

Abstract
The two most urgent and interlinked environmental challenges humanity faces are climate change and biodiversity loss. We are entering a pivotal decade for both the international biodiversity and climate change agendas with the sharpening of ambitious strategies and targets by the Convention on Biological Diversity and the United Nations Framework Convention on Climate Change. Within their respective Conventions, the biodiversity and climate interlinked challenges have largely been addressed separately. There is evidence that conservation actions that halt, slow or reverse biodiversity loss can simultaneously slow anthropogenic mediated climate change significantly. This review highlights conservation actions which have the largest potential for mitigation of climate change. We note that conservation actions have mainly synergistic benefits and few antagonistic trade‐offs with climate change mitigation. Specifically, we identify direct co‐benefits in 14 out of the 21 action targets of the draft post‐2020 global biodiversity framework of the Convention on Biological Diversity, notwithstanding the many indirect links that can also support both biodiversity conservation and climate change mitigation. These relationships are context and scale‐dependent; therefore, we showcase examples of local biodiversity conservation actions that can be incentivized, guided and prioritized by global objectives and targets. The close interlinkages between biodiversity, climate change mitigation, other nature's contributions to people and good quality of life are seldom as integrated as they should be in management and policy. This review aims to re‐emphasize the vital relationships between biodiversity conservation actions and climate change mitigation in a timely manner, in support to major Conferences of Parties that are about to negotiate strategic frameworks and international goals for the decades to come.
Keywords: biodiversity conservation, carbon sequestration, climate change mitigation, convention on biological diversity, nature‐based solutions, restoration
This review provides evidence that biodiversity conservation actions have mostly synergistic benefits and few antagonistic trade‐offs with climate change mitigation. Specifically, we identify co‐benefits in 14 out of the 21 action targets of the draft post‐2020 global biodiversity framework of the Convention on Biological Diversity. These relationships are context‐ and scale‐dependent; therefore, based on 12 case studies in different parts of the world, we showcase local biodiversity conservation actions that can be incentivized, guided and prioritized by global objectives and targets. We highlight the close interlinkages between biodiversity, climate change mitigation, other nature's contributions to people and good quality of life which need to be better integrated in management and policy.

1. INTRODUCTION
Increasing lines of evidence show the important contribution of nature to climate change mitigation. More than 30% of anthropogenic CO2 emissions are estimated to be re‐absorbed annually into the land surface (12.5 ± 3.3 GtCO2e year−1 (2010–2019), Friedlingstein et al., 2020) through forest regrowth, enhanced photosynthetic CO2 uptake and sequestration (Pugh et al., 2019; Ahlström et al., 2015; Schimel et al., 2015). A further ca. 25% of anthropogenic CO2 emissions is estimated to be absorbed by the ocean (9.2 ± 2.2 GtCO2e year−1 (2010–2019), Friedlingstein et al., 2020; IPCC, 2019), due to both CO2 dissolution in the ocean and the organic carbon cycle driven largely by photosynthesis, carbon sequestration in coastal vegetated habitats and the biological pump that moves carbon from the upper ocean layers to the deep ocean waters and sediments. Uncertainties in these estimates are large and reflect multiple challenges such as uncertain hindcasts of land‐use change, diverging process representations in models that contribute to these estimates, different sensitivities of these models to inter‐annual variation in weather and climate. The overall presence of a large natural sink is well constrained, however, by the measured increase of atmospheric CO2 concentrations and the relatively well‐quantified fossil fuel and industrial emissions. These powerful land and ocean sinks are currently by far the leading natural climate mitigation processes globally. Their carbon sequestration potential can be protected, and even enhanced, through ecosystem management on land and in the oceans. In the United Nations Framework Convention on Climate Change (UNFCCC) and Convention on Biological Diversity (CBD), the concept of nature‐based solutions has been proposed as a way to harness natural processes to help solve the climate challenge and reduce the loss of biodiversity, while providing other co‐benefits for nature's contributions to people.
Implementing nature‐based solutions, therefore, takes advantage of the strong connections between the climate system, the oceans, the land, and nature within these realms. Crucially, this needs to be managed without compromising the many nature's contributions to people (NCP) (Girardin et al., 2021). In this paper, we provide evidence of the potential effects of biodiversity conservation on the climate system including the greater storage of CO2 emissions by land and marine ecosystems, lowering greenhouse gas (GHG) emissions, for example, by altered wildfire management and land use practices, and changing the reflection of solar energy from the land surface (albedo change). The central question we address is the extent to which actions taken to halt or reverse biodiversity loss have consequences for these climate change mitigation processes, and how, when and where the form and strength of such links vary.
The last decade has seen increased concerns about biodiversity loss, with multiple lines of evidence that nature and its contributions to people are declining globally at unprecedented rates (Diaz et al., 2019; IPBES, 2019; WWF, 2020). National level responses have not been at the level of required actions, partially achieving only a handful of the Aichi Biodiversity Targets of the CBD Strategic Plan for Biodiversity 2011–2020 (Butchart et al., 2019; Secretariat of the Convention on Biological Diversity, 2020). Thus, there are high expectations for the upcoming CBD fifteenth Conference of the Parties (CBD COP 15) which will be held in 2022 in Kunming (China) to finalize a new set of well‐defined goals and targets that would incentivize strong and ambitious actions to reverse the loss of biodiversity. In the first draft of the post‐2020 global biodiversity framework of the CBD released in July 2021 (CBD 2021), there is a dedicated target on mitigation and adaptation to climate change—‘Target 8: Minimize the impact of climate change on biodiversity, contribute to mitigation and adaptation through ecosystem‐based approaches, contributing at least 10 GtCO2e per year to global mitigation efforts, and ensure that all mitigation and adaptation efforts avoid negative impacts on biodiversity’. The UNFCCC Paris Agreement, under Decision 1/CP.21, made a singular reference to biodiversity where Parties noted in the preamble ‘the importance of ensuring the integrity of all ecosystems, including oceans, and the protection of biodiversity, recognized by some cultures as Mother Earth, and noting the importance for some of the concept of “climate justice”, when taking action to address climate change’. Although we see a step forward in the recognition that biodiversity and climate change are interconnected by decision‐ and policy‐makers in these two separate Conventions, the two issues are still largely addressed separately and in an unbalanced manner.
Based on the work conducted for section 5 of the scientific outcome of the IPBES‐IPCC co‐sponsored workshop on biodiversity and climate change (Pörtner et al., 2021), we review recent scientific evidence relevant to assessing potential synergy between slowing and halting biodiversity loss and avoiding dangerous climate change. To what extent are these most urgent and important challenges facing humanity today interlinked by mechanistic links and feedbacks? Here, we focus on links between biodiversity conservation actions and climate change mitigation. Such actions and interventions are context and scale‐dependent; therefore, we showcase examples of local biodiversity conservation actions that can be incentivized, guided and prioritized by global objectives and targets given all actions matter.
2. POST‐2020 BIODIVERSITY GOALS HAVE STRONG POTENTIAL CO‐BENEFITS FOR CLIMATE CHANGE MITIGATION
Many policy measures designed to address biodiversity loss and degradation of NCP have potential co‐benefits with climate change mitigation (Girardin et al., 2021; Pörtner et al., 2021). The level of such co‐benefits largely depends on which processes and nature components are targeted by management actions. Just as it is important to distinguish between carbon capture (e.g. by photosynthesis), storage (e.g. in the bodies of organisms) and sequestration (e.g. protected from microbial activity in soils and sediments for periods of centuries to millennia) (Bax et al., 2021; Siegel et al., 2021), understanding differences between carbon sinks and feedbacks also greatly aids understanding of climate and biodiversity interactions. Albedo feedbacks (and other feedbacks arising from biophysical processes at the land surface) on climate may be an important component of climate change, but they are currently ignored by UNFCCC guidelines in accounting for the climate benefits of actions taken in support of climate change mitigation (Duveiller et al., 2020; Jia et al., 2019; Perugini et al., 2017).
Carbon sinks result from net carbon capture and storage and can be mediated by physicochemical (e.g. direct oceanic uptake of CO2 via the solubility pump, which leads to ocean acidification) or biological processes (photosynthesis and subsequent storage of the assimilated carbon). Sinks can be either local (the carbon is captured and stored in, e.g. forests or peatlands) or act by exporting the carbon in remote sites (e.g. kelp forests exporting to deep seas, or the marine vertical biological pump). Many natural carbon sinks and the capacity of processes driving those sinks are reduced by climate change, thereby exacerbating climate change further (positive feedback; Arneth et al., 2010). In contrast, some carbon sinks, such as polar continental shelves and boreal forests (taiga) increase with climate change, so they strengthen mitigation (negative feedback; Piao et al., 2006; Zhu et al., 2016). Biodiversity conservation measures and nature‐based climate solutions can be powerful in regulating climate when they concern natural carbon sinks that are large and have negative feedbacks on climate change.
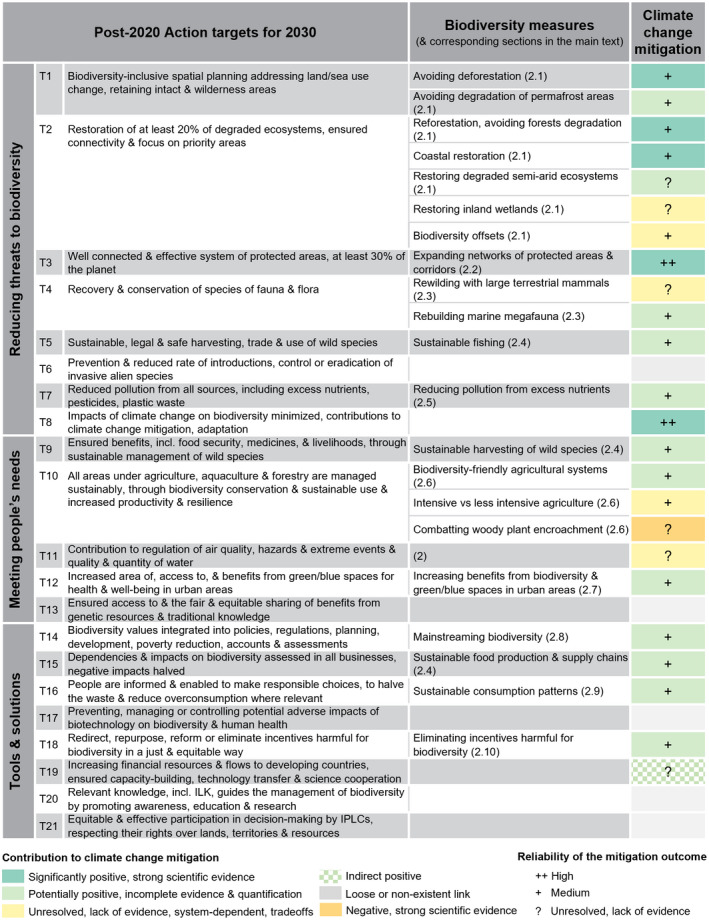
The first draft of the post‐2020 Global Biodiversity Framework provides 21 action‐oriented targets for 2030 which aim to contribute to the 2050 Vision for Biodiversity. Most of the framework targets have direct or indirect impacts on climate change mitigation (Table 1), even though they were not primarily designed with this goal. Here, we highlight a subset of biodiversity measures that are shown to have impacts on the climate system, based on potential contributions to carbon capture, storage and sequestration, the albedo effect and non‐CO2 GHG fluxes.
TABLE 1.
Action targets for 2030, from the first draft of the post‐2020 global biodiversity framework of the CBD (refer to CBD (2021) for the full and exact wording of the targets), and examples of biodiversity measures with contributions to climate change mitigation, and the reliability associated with achieving mitigation outcomes. The colour coding reflects expert judgement based on scientific literature (see Table S1 and main text). Target 8 is the outcome of all targets having significant and reliable positive contributions to climate change mitigation. (T: Target, ILK: Indigenous and Local Knowledge, IPLC: Indigenous Peoples and Local Communities)
2.1. Restoring degraded natural areas and retaining existing intact wilderness areas (Targets 1 and 2)
Restoring degraded natural areas is a flagship target of the post‐2020 global biodiversity framework of the CBD and put in the spotlight by the UN Decade on Ecosystem Restoration (2021–2030). Restoration is particularly critical where natural systems are so damaged that spontaneous recovery is unlikely or too slow compared with their degradation rate. Initially designed for protecting nature and its contributions to people, restoration programs provide opportunities for climate change mitigation, if selected ecosystems are both rich in species and potentially large carbon sinks.
2.1.1. Reforestation, avoided deforestation and degradation of forests
Large‐scale degradation of tropical and subtropical forests and woodlands is mainly driven by agricultural expansion and biofuel production and adversely affects both biodiversity and carbon stocks (Curtis et al., 2018; FAO, 2020; IPBES, 2019; Laurance et al., 2014; Mackey et al., 2020). Tropical deforestation contributed to almost one fifth of global anthropogenic GHG emissions during the 1990s (~5.5 GtCO2e year−1; Gullison et al., 2007). Recent remote sensing studies highlight that the increasing area over which tropical forest is being degraded may already match or exceed the area of tropical deforestation (Bullock et al., 2020; Matricardi et al., 2020). Above‐ground carbon losses due to degradation could increase estimates of gross deforestation losses by between 25% and more than 600% (Baccini et al., 2017; Maxwell et al., 2019; Pearson et al., 2017). Additional losses from tropical forest soils are unknown. Dryland forest and savannah have been deforested and degraded for many decades, in South America (e.g. Chaco and Cerrado systems; Mustin et al., 2017), Australia (e.g. Eucalypt woodlands; Queensland Department of Science, Information Technology and Innovation, 2017) and Asia (Tölle et al., 2017), with African woodlands having some of the highest deforestation rates in the world (e.g. 2,500–3,000 km2 year−1 in Zambia; Vinya et al., 2011).
The adoption of the REDD+mechanism (reducing emissions from deforestation and forest degradation in developing countries) by the UNFCCC in 2007, has provided a significant opportunity to align national climate change mitigation and biodiversity goals and has strengthened international efforts to slow and ultimately avoid deforestation (Johnson et al., 2019). Recent evidence shows that REDD+projects have been effective in some regions, for example, leading to the avoidance of 1.5 (±0.4) GtCO2e emissions from tropical forest in Brazil alone, between 2006 and 2017 (West et al., 2019) but efforts are not always sustained over the long term and a range of barriers exists in some other tropical regions such as in Indonesia (Ekawati et al., 2019) and in Africa (Gizachew et al., 2017).
Reforestation or restoration of degraded forests and woodlands with indigenous species plays a role in addressing losses of biodiversity and NCP, including through recovering the soil carbon stocks of these ecosystems (e.g. Edwards et al., 2021; Sileshi, 2016), and by targeting spatial spots that allow to re‐establish forest habitat continuity with additional positive impacts (e.g. Atlantic Forest; Newmark et al., 2017; Strassburg et al., 2018). It has been estimated that reforesting up to 3.7 million km2 of degraded tropical forest (less than half the potentially reforestable area) could support a carbon uptake rate of 5.5 GtCO2e year−1 by 2030, while contributing to conservation of forest‐dependent vertebrate species (Kemppinen et al., 2020). Reforestation using monoculture plantations of non‐indigenous species (e.g. Lewis et al., 2019), as well as some large scale sylviculture programs (e.g. Brazil; Mustin et al., 2017; Ethiopia; Pistorius et al., 2017) pose significant risks for nature and its contributions to people (Reisman‐Berman et al., 2019) but these practices are currently being incentivized financially.
2.1.2. Coastal restoration
Coastal habitats and ecosystems (e.g. mangroves, seagrass, salt marshes, coral reefs) are highly productive areas, harbouring large amounts of biological diversity, and providing valuable ecosystem services (e.g. water quality, carbon sequestration, food, livelihoods, cultural services and coastal protection; Mcleod et al., 2011). Coastal ecosystems are exposed to increase in temperature, acidification, sea level rise, salinification, and exposure to intensified storms (IPCC, 2014; Hoegh‐Guldberg et al., 2018). Urbanisation and coastal hardening further exert a strong pressure with increasing clustering of cities and other forms of development along the coasts (Barragán & de Andrés 2015; Liu et al., 2018; Loke et al., 2019). All these pressures have considerably shrunk the extent of many coastal ecosystems such as mangroves (Babcock et al., 2019), coral reefs (Oppenheimer et al., 2019) or seagrass (Waycott et al., 2009). Critically, the destruction and the degradation of these habitats result in reduced ‘blue carbon’ stocks by slowing biomass accumulation and exposing soils to increased oxidation of organic deposits (Mcleod et al., 2011). Compared to terrestrial forests, the global carbon sequestration is much lower in coastal systems due to their smaller extent, but the amount of carbon sequestration per unit of coastal vegetated area is typically much higher (Donato et al., 2011).
The success and costs of restoration options has varied between coastal ecosystems. Bayraktarov et al., (2016) reviewed restoration costs across a range of coastal ecosystems and found that coral reefs and seagrass beds were among the most expensive ecosystems to meaningfully restore, whereas mangrove restoration projects were the least expensive per unit area. It has also been shown that mangrove forests are capable of storing and sequestering a substantial proportion of carbon in both their biomass and soil substrates (Sanderman et al., 2018) even when fringing dense urban development areas (e.g. in Singapore; Friess et al., 2015).
2.1.3. Avoiding degradation of permafrost areas
The perennially frozen ground, known as permafrost, stores large amounts of organic carbon. The permafrost found in the Arctic and high mountain regions contains twice as much carbon as the atmosphere and about four times as much as all the carbon emitted by human activity from 1960 to 2019 (Canadell et al., 2021; Friedlingstein et al., 2020; Schädel et al., 2014). Permafrost wetlands degrade due to climate warming and commercial minerals extraction (CAFF, 2021; Opekunova et al., 2018; Peterson, 2001). Because of global warming, permafrost thaw, more frequent wildfires and shifts in hydrological flows in the permafrost region are anticipated (Canadell et al., 2021; Mishra et al., 2021). The upshot is the release of the carbon stored in the soil, biogenic CO2 and CH4 emissions and water quality reduction (Bruhwiler et al., 2021). These alterations impact biodiversity negatively because of soil moisture change and habitat loss, and increase the risk of extinction of wetland endemic and dependent species (Shin et al., 2019). Better management of permafrost wetlands, stopping destructive activities (drainage or excavation), preserving undamaged peatlands, rewetting artificially drained areas and restoring degraded areas will help maintaining their biodiversity and keeping carbon locked in the ground (Anisha et al., 2020; Avagyan, et al., 2017). Such management actions have been successfully implemented by the plan on the Long‐Term Gravel Pad Reclamation in Alaska (Peterson, 2001) and the Strategic Plan for peatland conservation and wise use in Mongolia (Ariunbaatar et al., 2017). In northern high‐latitude ecosystems, introducing large herbivores compacts snow and decreases its depth due to winter grazing and animal movements. This substantially reduces the thermal insulation efficiency of snow during wintertime and exposes permafrost to colder temperatures, thereby preventing or decreasing CH4 release from permafrost thawing. In addition, the selective grazing by large herbivores changes vegetation and soil properties, by decreasing shading and surface roughness, which may result in an increase of summer albedo (Cahoon et al., 2012; Falk et al., 2015; Schmitz et al., 2018; te Beest et al., 2016). Such an ecosystem‐based management experience could be scaled up to the entire Arctic permafrost region as a strategy to support mitigation of the global climate (Table 2 and Case study 11 in Table S3).
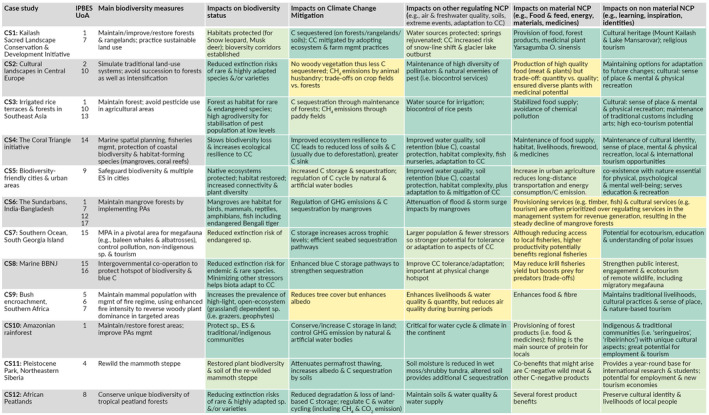
TABLE 2.
Impacts of biodiversity conservation measures on biodiversity, climate change mitigation and other NCP in twelve case studies (supporting references in Table S2). For the color coding, refer to Figure 1 legend. BBNJ: Biodiversity Beyond National Jurisdiction, C: carbon, CC: climate change, CH4: methane, CS: case study, ES: ecosystem service, GHG: greenhouse gas, mgmt: management, MPA: marine protected area, NCP: nature's contributions to people, PA: protected area, sp.: species, UoA: unit of analysis
2.1.4. Restoring degraded semi‐arid ecosystems
Degradation of semi‐arid ecosystems leads to significant carbon emissions via soil erosion and degradation (e.g. Chappell et al., 2016, 2019). Reversal of soil degradation is a longstanding focus of the UN Convention to Combat Desertification. Rebuilding soil (especially) and plant carbon stocks in semi‐arid regions is seen as a potentially significant contribution to mitigation of CO2 emissions because these regions are vast, and they appear to affect both the interannual variability and trend in the land carbon sink (Ahlström et al., 2015). But this view is not fully supported by the evidence (e.g. Yusuf et al., 2015), and the efficacy of restoring degraded semi‐arid systems is, thus, somewhat contested (Gosnell et al., 2020). Many semi‐arid systems have been observed as having ‘greening’ trends (Fensholt et al., 2012; Leroux et al., 2017; Stevens et al., 2017), tentatively linked to plant fertilization by rising atmospheric CO2 (Deng et al., 2021; Donohue et al., 2013; Zhu et al., 2016), with one of the outcomes being an increase in the competitive advantage of woody plants over grasses and increasing woody cover in these ecosystems. Global analysis suggests that this greening is associated with soil drying as a result of higher plant cover (Deng et al., 2020) and that associated shrub encroachment reduces grazing potential (Anadón et al., 2014).
2.1.5. Restoring inland wetlands
Inland wetland ecosystems provide vital services, such as food and freshwater, water purification and flood prevention. Humans use inland wetlands intensively for agriculture, aquaculture and urban development causing widespread degradation (IPBES, 2018). While important for global carbon sequestration, the disturbance of wetlands could result in increases of GHGs (Adhikari et al., 2009). Conversion, drainage and degradation of tropical wetlands and peatlands are important drivers of current increases in the atmospheric concentration of CH4 (Shukla et al., 2019). Notably, many irrigated rice areas are Ramsar sites for the protection of endangered species (Xi et al., 2020), but these are also important emitters of CH4 in the atmosphere (Shukla et al., 2019).
The protection and restoration of wetlands and peatlands is expected to reduce net carbon loss to the atmosphere between 0.15 and 0.81 GtCO2e year−1 up to 2050 (Couwenberg et al., 2009; Griscom et al., 2017; IPCC, 2019) and provide continued or restored natural CO2 removal (IPCC, 2019). There has been significant knowledge gained over the last decade on wetland drainage and rewetting practices (IPCC, 2013), while the carbon storage and flux rates, in particular the balance between CH4 sources and CO2 sinks are still hard to quantify (IPCC, 2019; Spencer et al., 2016). Recent evidence shows that tropical wetland CH4 emissions are underestimated, perhaps by a factor of two (IPCC, 2019). This could be due to the lack of inclusion of release by tree stems (Pangala et al., 2017). However, consistent with inventory data, agriculture may be a more probable source of increased emissions, for example, from wetland rice and livestock production systems in the tropics (Patra et al., 2016; Schaefer et al., 2016; Wolf et al., 2017). For peatlands models show mixed results for their role as future sink (Chaudhary et al., 2017; Ise et al., 2008; Spahni et al., 2013). Extensive historical data sets suggest that the currently global peatland sink could increase slightly until 2100 and decline thereafter, under scenarios of future warming (Gallego‐Sala et al., 2018).
2.1.6. Biodiversity offsets
Biodiversity offsetting is the practice of mitigating the negative impacts of developments on biodiversity (e.g. urban development, mining, agricultural expansion) by setting aside areas for restoring or protecting biodiversity elsewhere. Biodiversity offsets are meant to compensate for the services of carbon storage in biomass and soils through the development of newly restored or created habitats either in public or private lands. There are 12,983 listed biodiversity offsets under no net loss (NNL) principles implemented across 37 countries, predominantly forest ecosystems, covering about 153,679 km2 (Bull & Strange, 2018). The true benefits of biodiversity offsetting are currently being questioned. Although a large number of offset projects are located in less industrialized and emerging economies, existing studies on biodiversity offsets focused on North America, Western Europe and Australasia (Bull & Strange, 2018). In addition, although biodiversity offset programs advocate NNL principles, a recent review revealed that only one third of biodiversity offsets met the NNL principle with varied performance across different ecosystems (zu Ermgassen et al., 2019), suggesting the limited capacity of existing biodiversity offsetting projects to regulate climate and compensate for biodiversity loss. It should also be noted that, even when the offsetting program meets the NNL principle and is successful in maintaining carbon storage for climate change mitigation, biodiversity offsets can limit local people's access to, or cause loss of benefits from, the biodiversity and NCP on which their livelihoods depend, and so impacting their adaptation to climate change (Jones et al., 2019). However, existing studies on biodiversity offsets rarely assess potential trade‐offs between carbon storage and other ecosystem services (Sonter et al., 2020). Applying the NNL principle to offsetting programs will not necessarily minimize the trade‐offs and disconnects between the loss of local benefits from biodiversity, with gains in remote or global benefits. To avoid such trade‐offs, the type and distributions of NCP should be taken into consideration in the offsetting process along with the NNL principle, through more spatially explicit evaluation of NCP.
2.2. Implementing a well‐connected and effective system of protected areas (Target 3)
2.2.1. Expanding the network of protected areas
As of July 2021, protected areas cover 15.7% of terrestrial habitats and 7.7% of marine habitats (UNEP‐WCMC, 2021). There is increasing evidence that creating new protected areas and maintaining existing ones can help mitigating climate change through carbon sequestration and storage on land (Dinerstein et al., 2020; Soares‐Filho et al., 2010; UNEP, 2019) and at sea (O’Leary et al., 2016; Roberts et al., 2017; Sala et al., 2021). At sea, ecological representation and connectivity between marine protected areas would require at least 30% of sea protected, with a focus on areas most affected by human activities (Roberts et al., 2020). Most known, and nearly all measured examples of linking marine protection to climate change mitigation are in coastal wetlands, yet the vast majority of ocean (and protected ocean) is deep water. Although little studied, deep water ecosystems can hold considerable and important seabed carbon, such as around remote islands and seamounts (Barnes et al., 2019), Arctic and Antarctic continental shelves (see Souster et al., 2020 and Bax et al., 2021, respectively). On land, it is estimated that current protected areas store between 12% and 16% of land carbon stocks (Dinerstein et al., 2020; Melillo et al., 2016). To both reverse biodiversity loss and stabilize the climate, Dinerstein et al., (2020) suggest that protected areas should cover 50.4% of the terrestrial realm, storing a total of 1420 GtC. There is a substantial overlap of 92% between areas that require reversing biodiversity loss and the areas needing protection for enhancing carbon storage and drawdown. It is argued that by limiting global warming to 2°C and conserving 30% of the terrestrial surface, aggregate extinction risk could be reduced by more than half compared with business as usual scenario of unmitigated climate change and no increase in conserved areas (Hannah et al., 2020). These studies, while needing to be consolidated, suggest a stronger interlinkage between biodiversity conservation and climate change mitigation.
2.2.2. Establishing ecological corridors
Enhancing efficiency and effectiveness of protected areas in fragmented land‐ and sea‐scapes require establishment of ecological corridors (Dinerstein et al., 2017; Keeley et al., 2018; Littlefield et al., 2019). The carbon densities found in most of the ecological corridors are similar to those found in the protected areas they connect (Jantz et al., 2014). The ‘Global Safety Net’ plan—that aims to reverse biodiversity loss and increase carbon storage and drawdown by connecting all protected areas—indicates the need of only 4.3% of additional areas (but based on 2.5 km corridor width) (Dinerstein et al., 2020). Hallmarks of successful connectivity conservation includes community involvement, habitat priority setting, restoration actions and environmental services payments that satisfy tenets of climate‐smart conservation and improve the resilience of human and ecological communities (Littlefield et al., 2019; Townsend & Masters, 2015). Progress in protecting and restoring habitat connectivity has been slow (Keeley et al., 2018), and their climate benefits have not been fully explored.
2.3. Recovering and conserving wild species (Target 4)
Gaining increasing attention and supported by the 2021–2030 UN’s decade of ecosystem restoration (https://www.decadeonrestoration.org/), ‘rewilding’ conceives the restoration and protection of natural ecosystem processes, with no or little human interference following initial restoration. On land, vegetation and soils in most natural ecosystems store more carbon than systems managed for agriculture, forestry or grasslands. Rewilding is, therefore, considered a potentially important contribution to climate change mitigation, since the regrowing plants remove carbon dioxide from the atmosphere, storing this carbon in biomass and soils (Arneth et al., 2021; Smith et al., 2020; Strassburg et al., 2020). Rewilding, however, often also means trophic rewilding such as the reintroduction of large mammal herbivores or carnivores into ecosystems, supporting overall restoration efforts by rebuilding trophic cascades and promoting system self‐regulation (Bakker & Svenning, 2018; Sandom et al., 2020). Presence or absence of animals and the relative abundance of different animal groups affect ecosystem functioning, for example, by altering the amount of above‐ground biomass, changing light transfer into the canopy, albedo and evapotranspiration, altering plant species composition, affecting wildfire and mediating soil and ecosystem carbon and nitrogen turnover rates (Perino et al., 2019; Schmitz et al., 2018). It is widely accepted that the reintroduction of animals as part of rewilding will not only gain—often charismatic—species but also bring their ecosystem function. The impact of ecosystem processes relevant for climate change mitigation may well be considerable, as inferred from experimental plots, satellite remote sensing analyses, as well as assessment of paleo‐data (Cromsigt et al., 2018; Perino et al., 2019; Sandom et al., 2020; Schmitz et al., 2018). Whether or not trophic rewilding could be part of targeted mitigation strategies is unclear, however, and discussed controversially (Bakker & Svenning, 2018; Cromsigt et al., 2018; Sandom et al., 2020; Schmitz et al., 2018). Trophic rewilding could trigger processes that support mitigation as well as opposing it (Bakker & Svenning, 2018; Cromsigt et al., 2018; Sandom et al., 2020; Schmitz et al., 2018). Likely, the net climate impacts will differ strongly between regions and ecosystem types, and also how climate change will impact trophic interactions and species communities (Bakker & Svenning, 2018; Cromsigt et al., 2018; Sandom et al., 2020; Schmitz et al., 2018).
At sea, marine mammals, sharks and big predatory fish have been severely overexploited for decades (Myers & Worm, 2003; Roman, 2003) and are now the focus of many conservation programs around the world. As for terrestrial mammals, the contribution of these emblematic species in the global carbon cycle has been neglected until recent studies show the role of these predators either as carbon sinks or mediators of carbon sequestration in the deep ocean (Atwood et al., 2015; Heithaus et al., 2014; Lavery et al., 2010; Mariani et al., 2020; Passow & Carlson, 2012; Roman & McCarthy, 2010).
The role of animals has been particularly scrutinized in marine vegetated coastal habitats, identified as carbon‐rich ecosystems, where predators are essential to control the abundance of herbivores and bioturbators, which in turn impact the canopy height, root and shoot densities of the macrophytes, all characteristics playing a role in carbon capture and storage in plants, sequestration in sediments, and particle trapping (Atwood et al., 2015). Trophic downgrading triggered by the loss of predators can lead to the complete loss of salt marshes and seagrass habitats (Atwood et al., 2015), or severe reduction in the density of kelp forests (Wilmers et al., 2012). The case of the green turtle, a vulnerable and emblematic species, poses an interesting conservation challenge, as this seagrass grazer, when at high densities as a result of intense rewilding programs, and in the absence of predators (overexploited sharks), can overgraze and deplete seagrass beds (Heithaus et al., 2014).
In offshore waters, whales contribute to the biological pump, that is, the removal of carbon from the euphotic zone to the deep sea and sea bottom where it can be sequestered for several centuries or more (Passow & Carlson, 2012). While the sinking of whales’ carcasses is negligible compared with other drivers of the biological pump, it serves as a synergistic positive outcome of rebuilding programs (Pershing et al., 2010). Possibly more important is the role played by whales’ faecal plumes in fertilizing surface waters in allochthonous limiting nutrients, iron in particular, boosting primary production and thereby capturing atmospheric carbon down to deeper waters via the ocean biological pump (Lavery et al., 2010; Roman & McCarthy, 2010).
2.4. Ensuring sustainable harvesting of wild species, food production and supply chains (Targets 5, 9 and 15)
With the global human population projected to reach over 9 billion by 2050 (Adam, 2021), it is likely that we will need to produce more food, from land and the oceans, as well as to substantially reduce food loss and waste. Agriculture is one of the main causes of biodiversity loss on land (Green et al., 2005; IPBES, 2019a, 2019b; Newbold et al., 2015, 2016), due to a wide range of impacts including agriculture expansion into natural ecosystems, conversion for livestock farming, pollution from pesticides and fertilizers and its contribution to climate change (Crist et al., 2017; IPBES, 2019a). The biodiversity status of agricultural land and food supply chains can be improved by interventions such as the following: (a) sustainable intensification of production (Pretty et al., 2018), which allows land to be freed for nature conservation (Balmford et al., 2018; and see Section 2.6), (b) less intensive farming practices, for example, by adopting agroecological techniques (Albrecht et al., 2020; Tittonell et al., 2020; and see Section 2.6)—although this could exacerbate the clearance of natural ecosystems for agriculture if it resulted in lower productivity (Phalan et al., 2011), and (c) demand‐side changes in the food supply chain, such as dietary shifts toward more plant‐based diets containing less meat and dairy (Alexander et al., 2016; Bajželj et al., 2014; Xu et al., 2021), and reducing food loss and waste (Alexander et al., 2017; Gustavsson et al., 2011), which reduces demand for products with a large land footprint (Hayek et al., 2021). These interventions to improve the biodiversity status of agricultural land also have significant climate change mitigation and adaptation benefits with mitigation potentials ranging from 0.1 to 8 GtCO2e year−1, and adaptation benefits accruing to up to 2.3 billion people (Smith et al., 2020a).
In the ocean, fishing wild species as the main source of seafood production is a major driver of biodiversity loss as a result of overexploitation, bycatch and destruction of habitats (IPBES, 2019a, 2019b; Rogers et al., 2020). Fishing can also impact carbon fluxes, by exporting ocean carbon to land and ultimately to the atmosphere that would otherwise be sequestered in the deep sea (Mariani et al., 2020; Sala et al., 2021). Downward passive transport of carbon from the surface to the deep ocean occurs through sinking of dead carcasses, faecal pellets of fish and invertebrates, and this has been shown to be a significant contribution to the biological pump. By preventing these natural processes to happen, large pelagic fisheries have released an estimated minimum of 0.73 GtCO2e since 1950 (Mariani et al., 2020). In addition, fishing impacts the biological pump by extracting organisms that realize active diurnal vertical migration (DVM), feeding at the surface at night, and then joining the deeper mesopelagic domain during daytime where they produce faecal pellets. The flux of carbon driven by DVM is estimated to be 3.85 ± 0.5 GtCO2 year−1, about 18% of the passive flux of carbon (Aumont et al., 2018). In the Southern Ocean, fishing krill (Euphausia superba) has the potential to impact the biological pump significantly as krill is estimated to be responsible for about 35% of the current export of carbon to the ocean floor in the marginal ice zone (Belcher et al. 2019). An additional effect of fishing comes from the disruption and resuspension of sediments by bottom trawling, enhancing remineralization of organic matter and releasing CO2 in the water column (Atwood et al., 2020). The release of carbon into the atmosphere is massive during the first years of bottom trawling. For the surface currently trawled each year (1.3% of the global ocean), in a fictitious scenario where this surface would be free from previous disturbance, carbon emissions after 1 year of trawling are estimated at 1.47 Gt aqueous CO2, equivalent to about 15–20% of the atmospheric CO2 absorbed by the ocean each year (Sala et al., 2021).
Concerns regarding unsustainable fish production have driven a number of efforts to minimize environmental impacts, including developing sustainable aquaculture practices. These efforts first focused on replacement of fish‐derived protein and oil in aquaculture feeds with plant products resulting in a reduction of the trophic level of aquaculture species (Cottrell et al., 2021). This has a direct impact on fishing wild fish species for feed, with indirect consequences on the biological pump of carbon. There has also been a focus on development of integrated multitrophic aquaculture (IMTA) and cultivation of low trophic level species that do not require inputs from fisheries. IMTA relies on raising species from different trophic levels in close proximity to one another so that waste materials from one species cultivation serve as input food and nutrients for others (Knowler et al., 2020). Examples of IMTA include the cultivation of salmon with mussels and kelp or the growth of sea cucumbers with seaweeds and mussels (e.g. Knowler et al., 2020; Stenton‐Dozey et al., 2020). Cultivation of seaweeds has been concentrated in south east Asia but is now expanding globally in areas suitable for growth (Cai et al., 2021). Seaweeds can be used as a healthy food source, as food additives (e.g. phycocolloids), as animal feeds (reducing methane production from ruminants) and a range of other products such as bioplastics (Ditchburn & Carballeira, 2019; Kim et al., 2019). Seaweed cultivation can also have significant environmental benefits including removal of excess macronutrients such as N and P from coastal waters (Xiao et al., 2017), CO2 capture (e.g. Sondak et al., 2016) and can form habitat for natural populations of marine animals such as fish (also for bivalve cultivation; Theuerkauf et al., 2021). Research is currently underway to determine the scope of expanding IMTA and low trophic level aquaculture geographically as well as the environmental carrying capacity of these forms of food production if it is to be undertaken sustainably (e.g. Cai et al., 2021; Froehlich et al., 2019; Stenton‐Dozey et al., 2020).
2.5. Reducing pollution from excess nutrients (Target 7)
As the human population grows, so have the inputs of nutrients and organic matter to inland and coastal waters. Excess nitrogen, and in some cases phosphorus, originating from agricultural fertilizer runoff on land, industrial, wastewater and stormwater discharges, fossil‐fuel burning or aquaculture facilities lead to algal blooms and in some cases hypoxia in fresh, estuarine and coastal waters (Deininger & Frigstad, 2019; Jeppesen et al., 2010; Nazari‐Sharabian et al., 2018; Rabalais et al., 2014). This phenomenon, termed eutrophication, can modify the biogeochemical cycles of carbon, nitrogen, phosphorus, sulphide and silica as well as food webs and other ecosystem processes (Jeppesen et al., 2010; Li et al., 2021; Rabalais et al., 2014). There are more than 500 coastal locations and hundreds of freshwater lakes where oxygen loss occurs, accompanied by rising carbon dioxide levels due to microbial decomposition of excess primary and secondary production stimulated by eutrophication (Breitburg et al., 2018; Jane et al., 2021). Warming of fresh and ocean waters increases respiration rates and may tip eutrophic areas into hypoxia or anoxia; thus, it can be difficult to attribute observed oxygen and pH declines solely to eutrophication versus climate change (e.g. Kessouri et al., 2021).
Rising eutrophication combined with warming may increase GHG emissions in freshwater bodies, creating a positive feedback loop that accelerates both climate change and eutrophication, but with some complex, counteracting effects. This loop involves enhanced methane (CH4) release (Davidson et al., 2018); phytoplankton blooms that release CO2 but also dimethyl sulphide (DMS) that reduces solar radiation; deposition of acid nitrogen and sulphur compounds that promote ammonium oxidation releasing nitrous oxide (N2O) and warming‐enhanced stratification that might limit CH4 release and facilitate its storage (Li et al., 2021). Under eutrophication and anoxia in freshwater, the coupling of methanotrophy and denitrification may ameliorate N2O release (Naqvi et al., 2018). Also, eutrophic freshwater lakes (with >30 μgTP L−1) bury 5 times more organic carbon than non‐eutrophic lakes (Anderson et al., 2014). Biogeochemical feedbacks to climate from expanded coastal hypoxia may include increased denitrification and ammonium oxidation in coastal waters and release of N20 (Naqvi et al., 2010). Release of inorganic phosphate and iron from sediments under anoxic conditions stimulates further primary production and oxygen consumption as is the case in several oxygen minimum zones (Linsy et al., 2018; Lomnitz et al., 2016). Under some circumstances hydrogen sulphide, which is highly toxic, may be generated in anoxic water or sediments.
Control of nutrient pollution (oligotrophication) may lead to a significant decrease in coastal deoxygenation and the climate feedbacks associated with CH4 and N2O emissions or phosphorus and iron release. Effective tools to decrease coastal deoxygenation and associated GHG emissions include altered agricultural practices, various eco‐engineering approaches such as river diversions through wetlands to employ natural processes that reduce nitrogen loads (Engle, 2011) or new wetland construction (Duarte & Krause‐Jensen, 2018; Jahangir et al., 2016). Both eutrophication and the incidence of red tides (phytoplankton blooms) and green tides (macroalgal blooms) are predicted to increase under future warming scenarios (Gao et al., 2017; Gilbert, 2020; Xiao et al., 2019). The reduction of harmful algal blooms, which act as co‐stressors by releasing toxins and consuming oxygen, is a co‐benefit of oligotrophication (Griffith & Gobler, 2019; Pitcher & Jacinto, 2019).
By limiting nutrient inputs to both freshwater bodies and the ocean, it is possible to address eutrophication and climate change simultaneously, in part by preventing the two‐way feedbacks between eutrophication and climate. Societal choices about land and ocean management need to ensure that regionally rising precipitation (e.g. in the US or Asia) does not negate the nutrient removal benefits of these choices (Sinha et al., 2019).
2.6. Supporting the productivity, sustainability and resilience of biodiversity in agricultural and other managed ecosystems (Target 10).
2.6.1. Biodiversity‐based and biodiversity‐friendly agricultural systems
Reducing biodiversity loss and enhancing biodiversity in agricultural systems can help mitigate climate change and enhance a wide range of NCP (Leippert et al., 2020; Vanbergen et al., 2020; Wanger et al., 2020). Biodiversity can be promoted in agricultural systems directly—for example, through greater crop diversity, agroforestry or integration of crop production with livestock raising or aquaculture; or indirectly through practices that are biodiversity friendly—for example, through organic amendments to soils, reduced tillage or reduced pesticide use (Smith et al., 2020a; Tamburini et al., 2020). In general, these practices do not compromise agricultural yields, and in addition to enhancing biodiversity, they reduce nutrient losses, reduce soil erosion and improve soil fertility (Tamburini et al., 2020). Biodiversity‐based and biodiversity friendly agricultural practices also tend to increase carbon sequestration, but have highly variable effects on total GHG emissions, so identifying and implementing win‐win practices for biodiversity and climate change mitigation need to be done with this in mind (Smith et al., 2020a; Tamburini et al., 2020). Practices that promote biodiversity in agricultural systems include agroecology (which relies in part on the use of ecological processes to substitute for chemical inputs), regenerative agriculture (which focuses on restoring soil health and reversing biodiversity loss) and organic agriculture, as well as certain aspects of climate‐smart agriculture, conservation agriculture and sustainable intensification (Doré et al., 2011; FAO, 2019a; Giller et al., 2021; Pretty et al., 2018).
In situ conservation and restoration of biodiversity is one of a suite of practices falling within agroecological principles. Agroecology can also include promoting local and national food production, small‐scale farming and local innovations and resource use (Altieri et al., 2012). Mbow et al., (2014) provide an example of African smallholder farmers using agroecological practices (agroforestry) such as diversification of trees on‐farm and within the landscape to increase carbon content, prepare for climate extremes at the same time reduce and/or avoid crop failures. In dryland agriculture, soil and water conservation measures potentially improve ground cover and soil carbon content (Vanbergen et al., 2020; Wanger et al., 2020) and albedo (Creed et al., 2018).
The three main objectives of climate‐smart agriculture are to sustainably increase agricultural productivity and incomes; adapt to and build resilience to climate change and reduce GHG emissions (FAO, 2019a). Many of the practices promoted for climate‐smart agriculture are also good for biodiversity. For example, the Government of India and its Indian Council of Agricultural Research identified the districts most vulnerable to climate change and implemented climate‐smart agricultural interventions such as appropriate use of nitrogen fertilizers, which also reduces negative effects of nitrogen losses on non‐agricultural ecosystems, and conservation tillage for increased soil carbon content, which also enhances soil biodiversity. In addition, these measures helped farming groups protect their agricultural systems for local food security and increase adaptive capacity (Rao et al., 2020; Vanbergen et al., 2020) in climate‐smart villages (Aggarwal et al., 2018).
2.6.2. Intensive vs less intensive agriculture and the land sharing‐land sparing debate
GHG emissions will continue to increase with continued agricultural expansion and continued conventional intensification (Vanbergen et al., 2020). Scenarios that achieve climate change targets generally require substantial changes in agricultural intensification and demand for agricultural products (IPCC, 2019). One approach to conserving biodiversity could be to boost yields per unit area, through sustainable intensification on existing farmland that could in principle spare land for remaining natural habitats (Balmford et al., 2018; Smith et al., 2020a). However, intensive high‐yield farming raises other concerns because it can generate high levels of GHG emissions and nutrient losses. For example, excessive fertilization of crops results in N2O emissions, which is a potent GHG, and also results in other gaseous nitrogen losses that contribute to dry and wet deposition of nitrogen into terrestrial ecosystems that can reduce species richness (Galloway et al., 2003; Gerber et al., 2016; Tian et al., 2020). Moreover, NO x emissions can result in increased tropospheric ozone which can reduce productivity of natural ecosystems (Galloway et al., 2003). In addition, intensive high‐yield systems may move the provision of non‐material benefits (aesthetics, sense of place etc.) to larger distances from people's centres of livelihood, in contrast to less intensive and often more biodiverse agriculture. Others have argued that the most beneficial approach to conserving biodiversity in agricultural landscapes is to ‘share’ land more effectively with biodiversity, often by reducing agricultural intensity (Kremen, 2015). However, this approach runs the risk of increasing land conversion elsewhere to compensate for reduced agricultural yields per unit area, resulting in an overall negative impact on biodiversity and climate change mitigation (Balmford et al., 2018; Kremen, 2015). A growing consensus is that the benefits and drawbacks of these approaches are highly context dependent, not mutually exclusive and require careful spatial planning (Egli et al., 2018; Kremen, 2015; Salles et al., 2017).
In terms of demand for agricultural products, Van Meijl et al., (2017) indicate that demand is more influenced by population growth and changes in dietary preferences than for instance by GDP growth. This implies that in the end, agricultural pathway choices are about quality versus quantity and that high yield agriculture based on high inputs of energy, fertilizers and pesticides may not be necessary if demand shifts to reduce overconsumption, reduce food waste and loss and increase the fraction of plant‐based foods (Clark et al., 2019).
2.6.3. Using fire and bush removal to combat woody plant encroachment
Woody plant encroachment has been observed on several continents, especially in tropical and subtropical latitudes, linked to a poorly understood mix of land management actions and climate change drivers, including CO2 fertilization of woody plants (e.g. Stevens et al., 2017). Woody plant encroachment and its reversal may have important implications for both biodiversity and carbon sequestration. In Namibia, for example, the extent of bush encroachment is sufficient to offset national fossil fuel emissions (Ministry of Environment and Tourism, 2011), and this may reduce incentives to combat this trend at the cost of iconic species that are dependent on open ecosystems.
Bush encroachment converts open ecosystems to a more densely tree or bush‐covered state that alters biodiversity patterns significantly. For the open savannah plains fauna of Africa, clear direct negative impacts of bush encroachment are already visible for vulture, cheetah, and a myriad of smaller grassland bird species. Wildfire and browsing pressure used to maintain these systems in an ‘open’ condition may no longer be effective (e.g. Bond & Midgley, 2012), threatening the biodiversity of grassland and savannah landscapes across tropical Africa, South America, and Australasia. Experimental use of extreme fires and mechanical removal to reverse or halt bush encroachment have been tested (e.g. Smit et al., 2016), but the drive to maintain open ecosystems using disturbance can be misinterpreted as counter to the need for carbon sequestration. Apart from biodiversity benefits of reducing encroachment, maintenance of open grasslands can be motivated by the fact that carbon stocks of semi‐arid grassland ecosystems may match that of alternative woody ecosystems (Wigley et al., 2020) when below‐ground carbon stocks are taken into account. Maintenance of open ecosystems also helps to maintain streamflow (e.g. Creed et al., 2019) and reduce the intensity of wildfire regimes. In addition, open ecosystems provide multiple material benefits for subsistence livelihoods, including extensive grazing and thatching, as well as the irreplaceable cultural elements associated with these lifestyles.
Recognition of the natural cooling effects of high albedo of grasslands, and the plethora of local and global benefits provided by tropical open ecosystems to people support the need for sustainably managing these systems. In South Africa, active removal of invasive non‐indigenous woody plants has created millions of job opportunities, with some demonstrable results with respect to slowing woody plant encroachment rates (van Wilgen et al., 2012).
2.7. Increasing benefits from biodiversity and green/blue spaces in urban areas (Target 12)
The United Nations estimated that 55.3% of the world's population lived in urban settlements in 2018 (UNDESA, 2019). It is projected that the urbanization trend will continue to accelerate, while the majority of GHG emissions are generated by urban dwellers (United Nations Economist Network, 2020). Contrary to common perception, it has been shown that cities can harbour rich biodiversity (Secretariat of the Convention on Biological Diversity, 2012; Chan, 2019). As highlighted in the Edinburgh Declaration that highlights the commitment of subnational governments, cities and local authorities to the delivery of the post‐2020 Global Biodiversity Framework, cities can contribute to solutions for both biodiversity loss and climate change and do so in an integrated way across public, private and business sectors to be more effective. This has been the approach taken by cities such as Berlin, Edinburgh, Melbourne, Portland, Singapore, Toronto and Washington DC where biodiversity‐friendly, green and sustainable practices have been adopted (Beatley, 2016; Plastrik & Cleveland, 2018), to make them a more liveable and desirable habitat for people and nature.
Many of the methods used to conserve biodiversity in cities result in the enhancement of sinks for GHGs (Epple et al., 2016). Instead of relying on energy to cool down buildings, designing biodiversity‐friendly (‘biophilic’) buildings and building green infrastructure have gained much traction due to the multiple benefits that have been observed (Enzi et al., 2017). Planting native plants that attract native fauna in vertical greenery and roof‐top gardens provide habitats for wildlife as well as reduce ambient temperatures, thereby resulting in decreased energy consumption (Alhashimi et al., 2018; Wong et al., 2003). Other forms of green infrastructures result in multiple benefits such as the emulation of tropical rainforest with multi‐tiered and multi‐native species planting of roadsides (Chan, 2019), park connectors, the creation of sponge cities (Yu, 2020), the naturalization of drainage channels or the coverage of coastal walls with a range of different materials and forms that increase the establishment of marine biodiversity.
All these measures are implemented to increase biodiversity, with multiple benefits including the reduction in adverse effects of climate change (reduction of urban heat island effect, etc.), the enhancement of regulating (water quality, air quality, soil retention, etc.), material (urban agriculture in roof‐top gardens) and non‐material ecosystem services connecting people to nature to ensure their physical, psychological and mental well‐being (World Health Organization, 2016). The extent to which greening cities also contribute to climate change mitigation has yet to be better quantified and its potential to be prospected globally.
2.8. Mainstreaming biodiversity (Target 14)
The Convention on Biological Diversity has put a strong emphasis on the importance of biodiversity mainstreaming which entails ‘embedding biodiversity considerations into policies, strategies and practices of key public and private actors that impact or rely on biodiversity, so that it is conserved, and sustainably used, both locally and globally’ (Huntley & Redford, 2014). As biodiversity conservation and climate change challenges are intricately linked, it follows that biodiversity and climate are most effectively mainstreamed together (Pörtner et al., 2021).
Several examples illustrate the variety of ways in which biodiversity issues can be mainstreamed, and how this mainstreaming can be beneficial for climate change mitigation. Biodiversity reporting and natural capital accounting can help mainstreaming in governments and policies by informing decision‐making. In the case of the System of Environmental Economic Accounting (SEEA) framework, which has been implemented by more than twenty countries, national accounts are used to inform decision‐making on biodiversity, climate change mitigation and other environmental issues across government agencies (Secretariat of the Convention on Biological Diversity, 2005). Mainstreaming biodiversity in financial instruments, such as fiscal reforms, taxation models and fiscal incentives may also contribute to climate change mitigation. One of the most important and urgent reforms requiring cooperation across many actors are the reduction or elimination of subsidies that are harmful to both biodiversity and climate (see Section 2.10 for examples). Better integration of biodiversity into business operations and practices might also benefit climate change mitigation. Businesses are increasingly using GHG emissions accounting to identify and reduce their contributions to climate change, but adoption of biodiversity accounting has lagged behind, in part due to low awareness of biodiversity as a major issue for businesses and to a lack of well‐established biodiversity metrics for business to assess and value their impacts and dependencies on biodiversity (Smith et al., 2020b). Climate and biodiversity footprints of businesses are, however, intimately related because reducing biodiversity footprints depends on reducing GHG emissions because climate change is one of the major factors impacting on biodiversity and also relies on reducing the impacts of businesses on drivers that are common to both biodiversity loss and climate change such as deforestation, mining and unstainable agricultural practices (IPBES, 2019). Mainstreaming biodiversity across society, for example through education can be beneficial for climate change especially when they are part of an overall strategy to raise environmental awareness. For example, an examination of educational curricula in 46 countries found that fewer than half of education policies and curricula mentioned climate change and only a fifth made reference to biodiversity, leading to a recommendation that ‘more emphasis should be given to environmental themes in education, with a particular need to expand integration of climate change and biodiversity’ (UNESCO, 2021).
With a growing number of programmes and projects adopting the mainstreaming approach, there are now more case studies documenting their success stories. The Working for Water programme (WfW) in South Africa (Redford et al., 2015) demonstrates that mainstreaming biodiversity resulted in controlling invasive alien species and speeded the rate of legal protection of areas of high biodiversity. In Costa Rica, the joint policies of several Ministries (Environment, Agriculture, Planning and Finance) resulted in a national sustainable development plan that led to the creation of the Forest Incentives Programme where landowners could benefit from income derived from the conservation of forests. This would contribute to climate change mitigation from biodiversity conservation actions. Under the circumstances where climate change mitigation measures could have negative impacts on biodiversity conservation or vice versa, trade‐offs should be considered and comprehensively analysed (cf. 3.3).
2.9. Eliminating unsustainable consumption patterns (Target 16)
Where sustainable consumption occurs, biodiversity and ecosystems have been frequently shown to benefit, with some further climate change mitigation benefits. The largest potential for reducing agriculture, forestry and other land use (AFOLU) emissions of GHG is through reduced deforestation and forest degradation (0.4–5.8 GtCO2e year−1), a shift towards plant‐based diets (0.7–8.0 GtCO2e year−1) and reduced food and agricultural waste (0.8–4.5 CO2e year−1) (Jia et al., 2019). Thus, there is a high potential that consumers’ choices can directly impact both biodiversity and the climate. For example, the market for wood products that are sustainably harvested and/or produced has shown clear benefits for forest cover and diversity—and this may result in improved measures in carbon sequestration and albedo (Di Sacco et al., 2021; Heilmayr et al., 2020). Furthermore, the trend to demanding and consuming other products that are harvested or produced in a more biodiversity‐ and climate‐friendly way is clear—some examples here include sustainably produced meat (including wildlife products and meat produced using improved rangeland management; D’Aurea et al., 2021; Conant et al., 2017), sustainable fashion, potatoes, tea and coffee (Alom et al., 2021; Ruggeri et al., 2020; Vogt, 2020; Zhao et al., 2021)—and work on the quantification of such benefits is a valuable and growing field.
On the food demand side, nearly 10% of the agricultural land area could be spared globally through halving consumer waste arising from over‐consumption in some sectors of society (Alexander et al., 2017). In high‐income countries, consumer behaviour significantly influences the amount of food wasted, so raising awareness of the consequences on biodiversity and climate change among consumers but also along the whole supply chains involving industries and retailers, is of critical importance (FAO, 2019b). Likewise, studies that explore dietary scenarios of reduced consumption of animal protein estimate that between 10% and 30% of today's area under agriculture could be freed for other purposes (Alexander et al., 2016; Shin et al., 2019). The aforementioned Conant et al., (2017) study shows, for example, how better grazing management can increase soil carbon stocks—showing rates from 10 to more than 1000 MgC km−2 year−1.
2.10. Eliminating incentives harmful for biodiversity (Target 18)
Subsidies are often inefficient, expensive, socially inequitable, and environmentally harmful (IPBES, 2019a, 2019b; OECD, 2005). Despite the commitments of the governments to phase out or reform biodiversity harmful subsidies by 2020, they are still continuing but the detailed information on potential impacts of such subsidies is mostly unavailable (Dempsey et al., 2020). The financial resource allocated to environmentally harmful subsidies in various sectors outweigh the resources allocated to biodiversity conservation by a factor of 10 to 1 (OECD, 2019), indicating the pervasiveness of such subsidies. For example, in the agriculture sector, OECD countries spent US$100 billion in 2015 in activities that are potentially harmful to nature (OECD, 2019). Similarly, it is estimated that the annual global fossil fuel subsidies (US$300 to US$600 billion) generate negative externalities of at least US$4 trillion (Coady et al., 2019; Franks et al., 2018).
In the forestry sector, Brazil spent $14 billion (88 times more) subsidizing activities linked to deforestation compared to $158 million to stop deforestation (McFarland et al., 2015). In the fisheries sector, subsidies promoting sustainable exploitation are about $10 billion compared with $22 billion spent in causing overfishing (Sumaila et al., 2019). These discrepancies in environmentally beneficial and harmful subsidies arise partly due to difficulty in tracking such subsidies, and ignorance of the complexity of institutions. It is also partly due to activities around politicking and interest‐group lobbying, for example, for palm oil in Indonesia (Maxton‐Lee, 2018) and petroleum lobbying in Canada (Blue et al., 2018).
Further, difficulties arise from the effectiveness of environment‐friendly subsidies. In Europe, the European Court of Auditors (ECA) found that the foreseen expenditure on ‘farmland biodiversity’ of the European Commission, amounting to €66 billion between 2014 and 2020, had little effect (https://www.eca.europa.eu/en/Pages/DocItem.aspx?did={B5A7E9DE‐C42E‐4C1D‐A5D2‐03CA1FADE6F8}). Over the same period, more than a quarter of the Commissions subsidies under the Common Agricultural Policy had aimed to target climate change mitigation and adaptation, but GHG emissions from European farms are not decreasing (https://www.eca.europa.eu/en/Pages/DocItem.aspx?did={D6EB02B9‐C74E‐4017‐912B‐EE46E75127B1}). The ECA raises numerous flaws in the ways the subsidies are oriented: the unreliable way the Commission tracks biodiversity expenditure, the low potential of the measures financed, the poor formulation of the agriculture targets and the poor quality of the indicators used to track progress among the main reasons.
Fast and bold actions are needed to eliminate harmful subsidies to halt biodiversity loss and to mitigate climate change simultaneously (IPBES, 2019a, 2019b). Such actions include enhancing a culture of subsidy accountability among individuals and businesses; reforming policies for better transparency, reporting and assessments and using policy tools to incentivize individuals, communities and governments to maintain biodiversity, for example, public procurement, taxes and fees (Barbier et al., 2018, 2020; Girardin et al., 2021; Lundberg & Marklund, 2018).
3. INTEGRATING SYNERGIES AND TRADE‐OFFS BETWEEN MULTIPLE GOALS AT THE LEVEL OF LANDSCAPES AND SEASCAPES
The success of environmental measures, whether for biodiversity conservation or climate change mitigation, strongly depends on their context in a landscape or seascape, with consideration of the degree of its transformation, its multiple uses, local socio‐economic conditions and the quality of life of local communities. Ecosystem management is challenged with achieving multiple goals simultaneously in multifunctional and multiple‐use land‐ and sea‐scapes (hereafter referred to as ‘scapes), within which synergies and trade‐offs between biodiversity conservation and climate change mitigation can be realized (see section 2 in Pörtner et al., 2021). The use and transformation of ecosystems by human society occur mainly at local scales, but these local effects accumulate at larger spatial scales, resulting in significant changes in regional and higher‐scale biodiversity and ecosystem functioning. We, therefore, make use of local case studies (CS) to better understand how human appropriation of nature has resulted in the spatial fragmentation of ‘scapes and biodiversity loss and to unpack the enabling conditions (including incentives and governance factors) that have been effective in fulfilling multiple ‘scape objectives simultaneously (Table S3; Figure 1; Table 2). Protection of biodiversity is only one of a range of management objectives for a multi‐functional and multi‐use ‘scape. A clear need going forward is to improve our ability to mainstream biodiversity objectives and measure multiple benefits in specific contexts (Figure 1), but preferably with scope for upscaling and generalizing across cases. We propose an integrative analysis based on a selection of case studies that cover a wide range of IPBES units of analyses and are located on different continents, oceans and latitudes. Case studies also cover a diversity of conservation measures, types of NCP, needs of local communities, socio‐economic contexts and governance situations.
FIGURE 1.
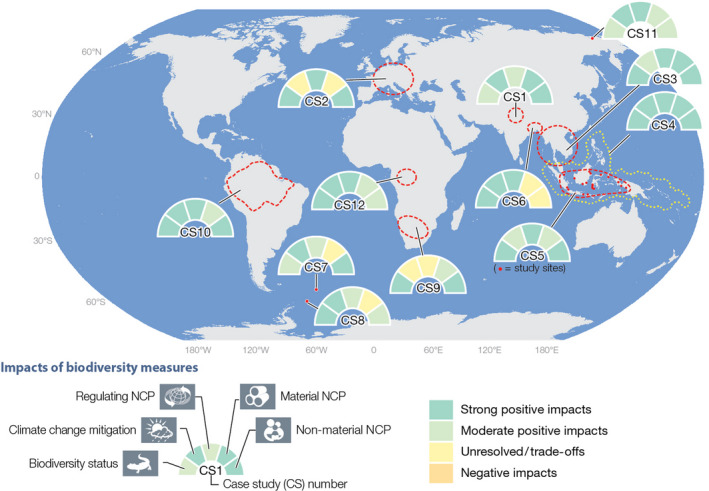
Example case studies (see Table 2, and Supplementary material for full description of the case studies and references) showing emerging synergies or trade‐offs between biodiversity conservation, climate change mitigation and nature's contributions to people (NCP). For each case study, five pieces of information are color‐coded in a pie chart regarding the impacts of biodiversity conservation measures on: biodiversity, climate change mitigation, regulating, material and non‐material NCP. None of the biodiversity measures implemented in the case studies resulted in negative impacts (indicated in orange), despite the fact that we had considered such negative impacts as possible in our assessment. CS: Case study, CS1: Kailash Sacred Landscape Conservation and Development Initiative, CS2: Cultural landscapes in Central Europe, CS3: Irrigated rice terraces and forests in Southeast Asia, CS4: The Coral Triangle initiative, CS5: Biodiversity‐friendly cities and urban areas, CS6: The Sundarbans, India‐Bangladesh, CS7: Southern Ocean, South Georgia Island, CS8: Marine BBNJ (Biodiversity Beyond National jurisdiction), South Orkney Islands, CS9: Bush encroachment, Southern Africa, CS10: Amazonian rainforest, CS11: Pleistocene Park, North‐eastern Siberia, CS12: African Peatlands
3.1. Local to regional actions and the critical role of scale and linkages
Ecosystems are used and transformed by human societies at local scales, but effects on biodiversity and ecosystem functioning accumulate and can be amplified at larger spatial scales. For example, nature‐based solutions in urban contexts can individually make a small contribution to global climate change mitigation and biodiversity protection, but given the high human densities in cities, all actions, collectively, have huge potential impact at the global scale, while improving the quality of life locally for a large share of the world population. On land, land use and land cover change result from increasing and changing human demands for nature's contributions, with the extent of change varying geographically, due to a complex interplay between biophysical, socioeconomic, and governance factors as illustrated by our set of case studies (Figure 1, Table 2 and Tables S2–S3). The configuration of anthropogenic landscapes offers opportunities to achieve various objectives in different locations relating to both human needs and sustainability objectives, including biodiversity and mitigation‐related regulating benefits like carbon storage and sequestration. Achieving specific objectives at local scales can together enable to reach multiple objectives at the ‘scape and global scale.
Land‐use and land cover change for increasing food provision or infrastructure expansion fragment and reduce the area of habitats and is currently the leading cause of terrestrial biodiversity loss (IPBES, 2018, 2019a, 2019b). While these processes also almost always result in net carbon release to the atmosphere (IPCC, 2019), they supply critical material benefits that maintain human society and contribute to good quality of life (Case studies CS 2, 3, 9, 10, 12, IPBES, 2019a, 2019b). Understanding how land cover can be allocated between competing uses is advancing and offers opportunities to optimize between multiple objectives (CS 2, 3). Such trade‐offs may include assessing the balance between biodiversity conservation, production of food and fibre (material NCP), carbon sequestration via reforestation (regulating NCP; CS 10) and restoration (regulating and cultural NCP; see CS 11 and 2.1 for the beneficial effects of rewilding mammoth steppe with large herbivores in Arctic permafrost areas).
Hannah et al., (2020) suggest that at the landscape to national scales, increasing conserved area from 20 to 30% significantly increases the resilience of the conserved area network to climate change (i.e. more species may be assured of persistence). The unequal distribution of biodiversity globally means that some regions have higher concentrations of rare species (Enquist et al., 2019) and prioritizing conservation objectives in these relatively small regions permits achievement of species conservation most efficiently (CS 1, 4). Spatial planning methodologies can be applied to maintain ecological functioning even in fragmented landscapes, through the consideration of zonation that takes into account landscape heterogeneity (Harlio et al., 2019; Moilanen et al., 2005). Many efforts are underway to green cities with multiple co‐benefits for human well‐being. Such efforts have the potential to connect cities to surrounding natural or managed areas and contribute to both biodiversity conservation and climate change mitigation regionally, as is the case in coastal cities for example (Beatley, 2014; 2.7 and CS 5).
In the ocean realm, governance differs greatly from that on land, with very little private ownership, and large amounts of global commons (CS 7, 8). Apart from coastal areas, marine ecosystem transformation occurs mainly via harvesting of consumer species for material benefits, with relatively low rates of plant use, and lower prevalence of high intensity food production systems. Important links between human use of the oceans and climate change mitigation have been identified, with local and regional harvesting scaling up to significantly alter the global food chain, with important impacts on processes like seabed sequestration of carbon and the biological pump of carbon (cf. 2.3, 2.4).
3.2. Realizing co‐benefits and synergies in land‐ and sea‐scapes
Species‐rich areas are often prioritized for biodiversity conservation measures at the ‘scape level, and in many cases, these same areas coincide with important carbon stores and sinks (CS 4, 6, 10, 12; Strassburg et al., 2010), making conservation actions doubly beneficial. The Amazon rainforest (CS 10) and mangrove forests (CS 4, 6), in particular, are two species‐rich iconic ecosystems that are typified by high rates of carbon sequestration (Donato et al., 2011; Guannel et al., 2016; Joly et al., 2018; Soares‐Filho et al., 2010). Mangroves are estimated to sequester on average between 600–800 MgCO2e km−2 year−1 in the sediments. This represents an annual carbon sequestration rate that is about four times more per unit area than some estimates for tropical forests (Donato et al., 2011), although estimates for climax forests, which are almost carbon‐neutral, should not be conflated with those for early succession forests, which are actively taking up carbon. By contrast, coral reefs that flourish in oligotrophic waters of tropical coastlines represent a counter example, where primary productivity and the build‐up of organic carbon over time are low, yet biodiversity is at least an order of magnitude higher than anywhere else in the ocean (Reaka‐Kudla, 1997).
There are also ecosystems with low species diversity but high carbon sequestration rates. In the Southern Ocean, for example, the sequestration of organic carbon is high while species richness is estimated to be lower than in non‐polar marine ecosystems (Bax et al., 2021), although precautions must be taken as these environments are not easily accessible for sampling. Protection of these ecosystems safeguards the trophic components of carbon pathways (e.g. krill, fish but also benthic communities), so that increased phytoplankton blooms (driven by sea ice losses and glacier retreat) are converted to higher seabed carbon storage, and possibly sequestration (CS 8) in oceans beyond national jurisdiction (Arrigo et al., 2008; Barnes et al., 2016).
In most of the case studies reported here (Figure 1; Table 2), conserving biodiversity in multi‐use and multi‐functional ‘scapes comes with a number of synergistic effects that help improve the quality of life of local people through the provisioning of context‐specific NCP. Such NCP could be materials (food, timber, fuelwood, fodder, medicinal plants) or regulating (water availability), or cultural/tourism related non‐material NCP (sense of place, cultural or sacred/religious heritage protection, ecotourism). In 2010, the Kailash Sacred Landscape Conservation and Development Initiative was launched covering parts of India, Nepal and China (CS 1) with the aim to contribute to local development and conservation—protecting threatened species (i.e. snow leopard, musk deer) and their habitats through a range of activities, such as reforestation, rangeland and farmland management. This initiative has great potential to generate climate change mitigation and adaptation co‐benefits through carbon sequestration and storage in natural systems—in forests, rangelands and soils (Aryal et al., 2018; Joshi et al., 2019; Liniger et al., 2020; Uddin et al., 2015). In addition, the initiative has benefited local and distant users through a range of NCP, such as timber, fodder, fuel wood, medicinal plants, water (Badola et al., 2017; Chaudhary et al., 2020; Liniger et al., 2020; Nepal et al., 2018; Tewari et al., 2020; Thapa et al., 2018), protection of Kailash Mountain and Mansarovar (cultural/religious sites), and the promotion of eco‐tourism (Adler et al., 2013; Pandey et al., 2016). Kailash Sacred Landscape also benefits distant downstream users through the (continued) provision of flowing waters for irrigation and other purposes (including hydro‐power generation) by protecting the sources.
Other such co‐benefits have been reported in various ‘scapes throughout the world. For example, about 50% of the poor people among the 7.2 million people of India and Bangladesh rely on Sundarbans (CS 6) for multiple benefits of nature (carbon sequestration, gas regulation, disturbance regulation) (IUCN, 2017, 2020). Similarly, conservation measures have generated co‐benefits to residents in cities (e.g. Beatley, 2016) which typically concentrate multi‐uses and multi‐functional spaces crossed by islands of biodiversity (cf. 2.7 and CS 5). The Coral Triangle Initiative (CS 4) is another example that generates multiple benefits from nature of local to regional significance (Foale et al., 2013; Table S3). Such co‐benefits are captured in the form of improvements to coastal water quality, nursery areas for fish, coastal protection, and maintenance of food, livelihoods, and cultural significance. In Africa, conservation of African peatlands yields high value water services to local people (CS 12). However, not all forms of benefits are equally prioritized due to the strong dependence of the livelihoods and income of poor people on material (fish, timber) and non‐material (tourism) contributions from nature (CS 1, 3, 4; Uddin et al., 2013).
The success of conservation measures is contingent on the extent of the operational and governance challenges encountered in implementing them. For example, in Amazonia (CS 10), the carbon sink function of rainforests is being negatively impacted by activities such as deforestation and the expansion of cattle and soybean production (Malhi et al., 2008), mining (Rosa et al., 2018), and the construction of big dams (Fearnside, 2016). In Pleistocene Park, the CH4 released by large re‐introduced animals could negatively affect the carbon cycle (Falk et al., 2015; Schmitz et al., 2018) (CS 11). Similarly, biodiversity and nature's contributions to people (fuelwood, fodder, water availability) have been adversely affected by the reforestation of dryland ecosystems (grasslands, savannah, forests) with exotic species (Acacia spp.) in Africa (CS 9). In all cases, it appears that without strong policy and operational coherences between countries, outcomes of the conservation measures would remain sub‐optimal (cf. 3.3).
Biodiversity conservation successes that generate climate change‐related co‐benefits depend on the consideration of the values held by the key stakeholders affected by such measures, primarily the indigenous and local people. Among the case studies examined, the differing values held by different groups of people are reflected in their actions in conservation or management of the ‘scapes. Some examples include the cultural values attached to sacred places in the Kailash Sacred Landscape (CS 1), the strong dependency of indigenous people on forest resources for identity and livelihoods in Amazon (CS 10), fishermen and their dependency on material benefits from fishing in the Coral Triangle (CS 4) and the Sundarbans (CS 6), and the strong and traditional livelihood linkages of local people with their surrounding ecosystems in Africa (CS 9).
3.3. Evaluating trade‐offs
Some biodiversity conservation measures and traditional land‐ and sea‐scape management have trade‐offs with climate change mitigation. In South Africa, wildfire management measures that limit bush encroachment and maintain open ecosystems contribute to surface cooling effects and biodiversity conservation by maintaining a high albedo surface, and provide a variety of NCP that support the livelihoods of local people (e.g. extensive grazing and thatching) (e.g. Creed et al., 2019). However, the carbon storage achieved through these measures are small compared to that provided by large‐scale afforestation (Wigley et al., 2020). Similarly, traditional grazing of livestock in Europe such as cattle, sheep and goats, whose primary purpose is to produce food (meat, milk, cheese, etc.), can also contribute to shaping and maintaining cultural landscapes as well as various ecosystem services such as water supply and flow regulation, carbon storage, erosion control, pollination (CS 2) (D’Ottavio et al., 2017). However, ruminant livestock produce large amounts of methane, but there is conflicting evidence about whether carbon storage achieved through grazing is sufficient to offset methane emission by livestock (Bengtsson et al., 2019). Trade‐offs also appear with the paddy rice cultivation in terraced fields in Southeast Asia that provides various NCP to local people, such as food production, water flow regulation, and sediment control, while the traditional cultural landscape shaped by paddy farming is also an important tourism resource for the livelihoods of locals (CS 3). However, rice paddies are known to be very large sources of methane emissions (Saunois et al., 2016; Zhang et al., 2020), which is reportedly concentrated during the monsoon season (Hayashida et al., 2013).
Trade‐offs in NCP have spatially differentiated consequences for their stakeholders with various types of flow of nature's contributions from providers to beneficiaries (Fisher et al., 2009; Serna‐Chavez et al., 2014; Syrbe & Walz, 2012). While beneficiaries of climate mitigation through improved carbon storage from, for instance, nature restoration, are spread across the globe, the restored nature can provide other NCP to locals such as water and soil regulation. Decisions on land‐ and sea‐scape uses are mostly determined locally for the benefit of locals, but it sometimes go counter to global benefits. For instance, Sundarbans mangroves are one of the largest mangrove forests in the world stretching across India and Bangladesh (CS 6). These and other mangrove forests contribute to global scale climate mitigation. Mangrove forests also provide other vital contributions such as firewood and timbers, fish and shrimps, water quality, sediment retention and disturbance regulation against extreme weathers such as cyclones and storm, most of which are benefited by locals to sustain their livelihood (Sannigrahi et al., 2020a; IUCN, 2020), whereas other contributions such as recreation and tourism are appreciated not only by locals but also by visitors. The use of mangroves for one of such contributions (e.g. aquaculture of fish and shrimps) will inevitably affect the state of the other contributions (e.g. carbon storage, water and soil regulation). Many existing studies on Sundarbans mangroves demonstrated that climate regulation is one of the vital NCPs along with habitat provision and disturbance regulation (Sannigrahi et al., 2020a, 2020b). However, these nature's contributions carry less weight in decision‐making by locals who often prioritize the production and use of NCP that bring direct benefits and revenues to local stakeholders and governments, and, in the end, mangrove forests are altered to aquaculture and tourism sites, diminishing mangrove's contribution to climate change mitigation (Uddin et al., 2013). The Sacred Landscape of Kailash is faced with a similar challenge due to the growing demand of tourism in the area which results in increased water and energy consumption, forest degradation, causing a trade‐off with climate change mitigation (Nepal et al., 2018; Pandey et al., 2016). This contrasts with consideration of climate change adaptation, which has direct implications for local communities.
Biodiversity conservation measures can have unintended consequences and challenges, which need to be recognized, rectified and addressed through proper planning and governance mechanisms. This could be done through a holistic, integrated, consultative and adaptive approach within and across nations. Transboundary cooperation by multiple countries can help manage trade‐offs among multiple NCP and simultaneously address biodiversity conservation, climate change mitigation and adaptation at regional level. For instance, the Coral Triangle Initiative (CS 4), which involves six participating countries (i.e. Indonesia, Malaysia, Papua New Guinea, Philippines, Solomon Islands and Timor‐Leste) in the Pacific and Indian Ocean, has developed marine protected area networks along with other joint efforts such as the identification of priority seascapes and spatial planning (Asaad et al., 2018; Weeks et al., 2014) that balance biodiversity measures, climate measures, and socio‐economic development at regional scale. In a similar manner, the Kailash Sacred Landscape Conservation and Development Initiative (CS 1) jointly established by China, India and Nepal, for the conservation of ecosystems, biodiversity, and quality of life along with cultural heritage of the pilgrimage to Mount Kailash, has contributed to establishing transboundary protected area networks and improving livelihood of locals (Zomer & Oli, 2011). The consensus building process among concerned countries is key to make the transboundary initiatives successful. This is especially true for the establishment of protected areas on the high seas, where agreement of member states is required under a multilateral environmental instrument to protect biodiversity, and is currently under negotiation by the UN (Tessnow‐von & Vadrot, 2020). The South Orkney Islands Southern Shelf Marine Protected Area (CS 8) is a good example of a successful transboundary conservation effort in high seas. Although the primary objective is the conservation of biodiversity in the region, its high carbon storage and sequestration capacity also contributes to climate change mitigation (Barnes et al., 2016; Trathan et al., 2014).
4. CONCLUSION
Species and their habitats contribute to regulate the climate system, by modifying the energy and water cycles, the consumption and production of radiatively active gases and aerosols. Actions for biodiversity conservation have not focused on this central role so far, but its recent recognition requires conservation actions to be better aligned with climate goals, and demands an assessment of where this alignment may be feasible, relevant and non‐conflictual.
Our review shows that many instances of conservation actions intended to slow, halt or reverse biodiversity loss can simultaneously slow anthropogenic climate change. Specifically, we identified direct co‐benefits in 14 out of the 21 action targets of the post‐2020 global biodiversity framework of the CBD, notwithstanding the indirect links that can as well support both biodiversity conservation and climate change mitigation. Avoiding deforestation and restoring ecosystems (especially high‐carbon ecosystems such as forests, mangroves or seagrass meadows) are among the conservation actions having the largest potential for mitigating climate change. Our analysis shows that conservation actions generally generate more mutually synergistic benefits than antagonistic trade‐offs with respect to climate change mitigation. Synergies between biodiversity conservation, climate change mitigation, other NCP and good quality of life are seldom quantified in an integrated way, and the evidence base for assessing these needs to be consolidated and collected routinely. The assessment of biodiversity and climate synergies would greatly benefit from the development of fully integrated indicators, models and scenarios, which would also facilitate decision‐making for mainstreaming and applying ecosystem‐based integrative approaches, while recognizing the multi‐use and multi‐function dimension of scapes.
Improving the linkages between the different scales of actions is essential for successfully implementing joint biodiversity and climate actions. Locally motivated biodiversity conservation actions can be incentivized, guided and prioritized by international objectives and targets, including climate mitigation and adaptation objectives. However, this should be done rigorously and based on evidence to avoid oversimplified objectives that assume positive synergies between biodiversity and climate too systematically, such as subsidizing large‐scale tree‐planting campaigns regardless of local needs and socio‐economic contexts. Choosing the right options locally is indeed crucial. Local initiatives matter since the benefits of many small and local biodiversity measures accumulate to make a large contribution to climate mitigation, while also providing multiple local benefits.
At the landscape or seascape level, many areas of high biodiversity have also high rates of carbon sequestration. However, there are exceptions to the generally positive synergy between biodiversity conservation and climate change mitigation. The realization of synergistic benefits is strongly dependent on which biomes, ecosystem uses, and sectoral interactions are under consideration. It may be impossible to achieve win‐win synergies, or even manage the trade‐offs between climate and biodiversity in every single small part of a landscape or seascape, but achieving synergies becomes progressively easier at the ‘scape level. Therefore, local to global policies and practices designed for biodiversity conservation and climate change mitigation should be considered in an integrated and consultative way in mixed‐use land‐ and sea‐scapes so that win‐win synergies and nature's contributions to people can be maximized.
CONFLICT OF INTEREST
The authors declare no competing interests.
AUTHOR CONTRIBUTION
Y.J.S., G.F.M., H.T.N., S.H., R.P., O.H.G., J.S., L.C., G.I., and E.A. conceived the initial ideas of the paper, and all authors reviewed the literature and wrote the manuscript.
Supporting information
Supplementary Material
ACKNOWLEDGEMENTS
We dedicate this article to Bob Scholes, whose rich knowledge, incredible dedication and leadership and love for nature contributed significantly to this article. He will be sorely missed as a friend and colleague. This paper is based on the work and findings from the IPBES‐IPCC co‐sponsored workshop on Biodiversity and Climate Change that was held in December 2020. The views expressed here, however, represent the individual views of the authors. We are grateful to the scientific steering committee of the IPBES‐IPCC workshop, review editors, the IPBES and IPCC Secretariat and Technical Support Units. We are especially grateful to Anne Larigauderie for her vision and continued support on all fronts, Yuka Estrada for having drawn Figure 1 and Table 2 of the paper, and Sally Archibald, David Cooper, Debora Ley, Valérie Masson‐Delmotte and Unai Pascual for their critical advice on previous versions of the manuscript. YJS acknowledges support from the Biodiversa and Belmont Forum project SOMBEE (BiodivScen ERA‐Net COFUND programme, ANR contract n°ANR‐18‐EBI4‐0003‐01), and the European Union’s Horizon 2020 research and innovation programme under grant agreement No 869300 (FutureMARES). GI was supported by the project No. AAAA‐A19‐119021990093‐8 (FMGE‐2019‐0007) of the Institute of Geography, Russian Academy of Sciences. ADR acknowledges the support of REVOcean during this work. SH was supported by the Research Institute for Humanity and Nature Project No. 14200103, the Environment Research and Technology Development Fund (S‐15) of the Ministry of the Environment, Japan, and CRRP2018‐03MY‐Hashimoto by the Asia‐Pacific Network for Global Change Research.
Shin, Y.‐J. , Midgley, G. F. , Archer, E. R. M. , Arneth, A. , Barnes, D. K. A. , Chan, L. , Hashimoto, S. , Hoegh‐Guldberg, O. , Insarov, G. , Leadley, P. , Levin, L. , Ngo, H. T. , Pandit, R. , Pires, A. P. F. , Pörtner, H.‐O. , Rogers, A. D. , Scholes, R. J. , Settele, J. , & Smith, P. (2022). Actions to halt biodiversity loss generally benefit the climate. Global Change Biology, 28, 2846–2874. 10.1111/gcb.16109
Contributor Information
Yunne‐Jai Shin, Email: yunne-jai.shin@ird.fr.
Alex D. Rogers, Email: alex.rogers@revocean.org.
DATA AVAILABILITY STATEMENT
The data that supports the findings of this study are available in the supplementary material of this article.
REFERENCES
- Adam, D. (2021). How far will global population rise? Researchers Can’t Agree. Nature, 597(462), 465. [DOI] [PubMed] [Google Scholar]
- Adhikari, S. , Bajracharaya, R. M. , & Sitaula, B. K. (2009). A review of carbon dynamics and sequestration in wetlands. Journal of Wetlands Ecology, 2, 2–46. 10.3126/jowe.v2i1.1855 [DOI] [Google Scholar]
- Adler, C. E. , McEvoy, D. , Chhetri, P. , & Kruk, E. (2013). The role of tourism in a changing climate for conservation and development. A problem‐oriented study in the Kailash Sacred Landscape. Nepal. Policy Sciences, 46(2), 161–178. 10.1007/s11077-012-9168-4 [DOI] [Google Scholar]
- Aggarwal, P. K. , Jarvis, A. , Campbell, B. M. , Zougmoré, R. B. , Khatri‐Chhetri, A. , Vermeulen, S. J. , Loboguerrero, A. M. , Sebastian, L. S. , Kinyangi, J. , Bonilla‐Findji, O. , Radeny, M. , Recha, J. , Martinez‐Baron, D. , Ramirez‐Villegas, J. , Huyer, S. , Thornton, P. , Wollenberg, E. , Hansen, J. , Alvarez‐Toro, P. , … Yen, B. T. (2018). The climate‐smart village approach: Framework of an integrative strategy for scaling up adaptation options in agriculture. Ecology and Society, 23(1), 14. https://www.ecologyandsociety.org/vol23/iss1/art14/ 10.5751/ES-09844-230114 [DOI] [Google Scholar]
- Ahlström, A. (2015). The dominant role of semi‐arid ecosystems in the trend and variability of the land CO2 sink. Science, 348(6237), 895–899. 10.1126/science.aaa1668 [DOI] [PubMed] [Google Scholar]
- Albrecht, M. , Kleijn, D. , Williams, N. M. , Tschumi, M. , Blaauw, B. R. , Bommarco, R. , Campbell, A. J. , Dainese, M. , Drummond, F. A. , Entling, M. H. , Ganser, D. , Arjen de Groot, G. , Goulson, D. , Grab, H. , Hamilton, H. , Herzog, F. , Isaacs, R. , Jacot, K. , Jeanneret, P. , … Sutter, L. (2020). The effectiveness of flower strips and hedgerows on pest control, pollination services and crop yield: A quantitative synthesis. Ecology Letters, 23(10), 1488–1498. 10.1111/ele.13576 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander, P. , Brown, C. , Arneth, A. , Finnigan, J. , Moran, D. , & Rounsevell, M. (2017). Losses, inefficiencies and waste in the global food system. Agricultural Systems, 153, 190–200. 10.1016/j.agsy.2017.01.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander, P. , Brown, C. , Arneth, A. , Finnigan, J. , & Rounsevell, M. D. A. (2016). Human appropriation of land for food: The role of diet. Global Environmental Change‐Human and Policy Dimensions, 41, 88–98. 10.1016/J.GLOENVCHA.2016.09.005 [DOI] [Google Scholar]
- Alhashimi, L. , Aljawi, L. , Gashgari, R. , & Almoudi, A. (2018). The effect of rooftop garden on reducing the internal temperature of the rooms in buildings. Proceedings of the 4th World Congress on Mechanical, Chemical, and Material Engineering (MCM’18) Madrid, Spain ‐ August 16–18, 2018. 10.11159/icmie18.114 [DOI] [Google Scholar]
- Altieri, M. A. , Funes‐Monzote, F. R. , & Petersen, P. (2012). Agroecologically efficient agricultural systems for smallholder farmers: Contributions to food sovereignty. Agronomy for Sustainable Development, 32, 1–13. 10.1007/s13593-011-0065-6 [DOI] [Google Scholar]
- Anadón, J. D. , Sala, O. E. , Turner, B. L. , & Bennett, E. M. (2014). Effect of woody‐plant encroachment on livestock production in North and South America. Proceedings of the National Academy of Sciences, 111(35), 12948–12953. 10.1073/pnas.1320585111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson, N. J. , Bennion, H. , & Lotter, A. F. (2014). Lake eutrophication and its implications for organic carbon sequestration in Europe. Global Change Biology, 20(9), 2741–2751. 10.1111/gcb.12584 [DOI] [PubMed] [Google Scholar]
- Anisha, N. F. , Mauroner, A. , Lovett, G. , Neher, A. , Servos, M. , Minayeva, T. , Schutten, H. , & Minelli, L. (2020). Locking carbon in wetlands: Enhancing climate action by including wetlands in NDCs. Alliance for Global Water Adaptation and Wetlands International. [Google Scholar]
- Ariunbaatar, A. , Batjargal, Z. , Banzragch, T. S. , Burenbaatar, G. , Dorjsuren, C. H. , ChuluunBaatar, I. , Dugarjav, C. H. , Enebish, D. , Enkh‐Amgalan, S. , Enkhbileg, S. , Heyde, J. , Jambaljav, Y. , Javzandulam, B. , Khosbayar, C. H. , Minayeva, T. , Nyambayar, B. , Saandari, M. , Saikhanjargal, D. , Sirin, A. , … Zoyo, D. (2017). Strategic planning for peatlands in Mongolia (Assessment Report TA‐8802; Technical Assistance, p. 273). Asian Development Bank.
- Arneth, A. , Harrison, S. P. , Zaehle, S. , Tsigaridis, K. , Menon, S. , Bartlein, P. J. , Feichter, J. , Korhola, A. , Kulmala, M. , O'Donnell, D. , Schurgers, G. , Sorvari, S. , & Vesala, T. (2010). Terrestrial biogeochemical feedbacks in the climate system. Nature Geoscience, 3(8), 525–532. 10.1038/ngeo905 [DOI] [Google Scholar]
- Arneth, A. , Olsson, L. , Cowie, A. , Erb, K. H. , Hurlbert, M. , Kurz, W. A. , Mirzabaev, A. , & Rounsevell, M. D. A. (2021). Restoring degraded land. Annual Review of Environment and Resources, 10.1146/annurev-environ-012320-054809 [DOI] [Google Scholar]
- Arrigo, K. R. , van Dijken, G. L. , & Bushinsky, S. (2008). Primary production in the Southern Ocean, 1997–2006. Journal of Geophysical Research: Oceans, 113(C8), C08004. 10.1029/2007JC004551 [DOI] [Google Scholar]
- Aryal, K. , Poudel, S. , Chaudary, R. P. , Chettri, N. , Ning, W. , Shaoliang, Y. , & Kotru, R. (2018). Conservation and management practices of traditional crop genetic diversity by the farmers: A case from Kailash Sacred Landscape. Nepal. Journal of Agriculture and Environment, 18, 15–28. 10.3126/aej.v18i0.19886 [DOI] [Google Scholar]
- Asaad, I. , Lundquist, C. J. , Erdmann, M. V. , Van Hooidonk, R. , & Costello, M. J. (2018). Designating spatial priorities for marine biodiversity conservation in the Coral Triangle. Frontiers in Marine Science, 5, 400. 10.3389/fmars.2018.00400 [DOI] [Google Scholar]
- Atwood, T. B. , Connolly, R. M. , Ritchie, E. G. , Lovelock, C. E. , Heithaus, M. R. , Hays, G. C. , Fourqurean, J. W. , & Macreadie, P. I. (2015). Predators help protect carbon stocks in blue carbon ecosystems. Nature Climate Change, 5(12), 1038–1045. 10.1038/nclimate2763 [DOI] [Google Scholar]
- Atwood, T. B. , Witt, A. , Mayorga, J. , Hammill, E. , & Sala, E. (2020). Global patterns in marine sediment carbon stocks. Frontiers in Marine Science, 7, 165. 10.3389/fmars.2020.00165 [DOI] [Google Scholar]
- Aumont, O. , Maury, O. , Lefort, S. , & Bopp, L. (2018). Evaluating the potential impacts of the diurnal vertical migration by marine organisms on marine biogeochemistry. Global Biogeochemical Cycles, 32(11), 1622–1643. 10.1029/2018GB005886 [DOI] [Google Scholar]
- Avagyan, A. , Baker, E. , Barthelmes, A. , Cisneros Velarde, H. , Dargie, G. , Guth, M. , Hergoualc’h, K. , Johnson, L. , Joosten, H. , Kieft, J. , Kopansky, D. , Miles, L. , Minayeva, T. , Montanarella, L. , Nuutinen, M. , van Paddenburg, A. , Peters, J. , Garcia Rangel, S. , Richards, J. , … Silvius, M. (2017). Smoke on water—Countering global threats from peatland loss and degradation. A UNEP Rapid Response Assessment (p. 70). [A RAPID RESPONSE ASSESSMENT]. United Nations Environment Programme and GRID‐Arendal. https://www.grida.no/publications/355
- Babcock, R. C. , Bustamante, R. H. , Fulton, E. A. , Fulton, D. J. , Haywood, M. D. E. , Hobday, A. J. , Kenyon, R. , Matear, R. J. , Plagányi, E. E. , Richardson, A. J. , & Vanderklift, M. A. (2019). Severe continental‐scale impacts of climate change are happening now: Extreme climate events impact marine habitat forming communities along 45% of Australia’s coast. Frontiers in Marine Science, 6, 411. 10.3389/fmars.2019.00411 [DOI] [Google Scholar]
- Baccini, A. , Walker, W. , Carvalho, L. , Farina, M. , Sulla‐Menashe, D. , & Houghton, R. A. (2017). Tropical forests are a net carbon source based on aboveground measurements of gain and loss. Science, 358, 230–233. 10.1126/science.aam5962 [DOI] [PubMed] [Google Scholar]
- Badola, E. , Rawal, R. , & Dhyani, P. (2017). Stories of Success: Narratives from a sacred land. GBPIHED. [Google Scholar]
- Bajželj, B. , Richards, K. S. , Allwood, J. M. , Smith, P. , Dennis, J. S. , Curmi, E. , & Gilligan, C. A. (2014). Importance of food‐demand management for climate mitigation. Nature Climate Change, 4(10), 924–929. 10.1038/nclimate2353 [DOI] [Google Scholar]
- Bakker, E. S. , & Svenning, J. C. (2018). Trophic rewilding: Impact on ecosystems under global change. Philosophical Transactions of the Royal Society B‐Biological Sciences, 10.1098/rstb.2017.0432 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balmford, A. , Amano, T. , Bartlett, H. , Chadwick, D. , Collins, A. , Edwards, D. , Field, R. , Garnsworthy, P. , Green, R. , Smith, P. , Waters, H. , Whitmore, A. , Broom, D. M. , Chara, J. , Finch, T. , Garnett, E. , Gathorne‐Hardy, A. , Hernandez‐Medrano, J. , Herrero, M. , … Eisner, R. (2018). The environmental costs and benefits of high‐yield farming. Nature Sustainability, 1(9), 477–485. 10.1038/s41893-018-0138-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barbier, E. , Burgess, J. , & Dean, T. (2018). How to pay for saving biodiversity. Science, 360, 486–488. 10.1126/science.aar3454 [DOI] [PubMed] [Google Scholar]
- Barbier, E. B. , Lozano, R. , Rodríguez, C. M. , & Troëng, S. (2020). Adopt a carbon tax to protect tropical forests. Nature, 578(7794), 213–216. 10.1038/d41586-020-00324-w [DOI] [PubMed] [Google Scholar]
- Barnes, D. K. A. , Ireland, L. , Hogg, O. T. , Morley, S. , Enderlein, P. , & Sands, C. J. (2016). Why is the South Orkney Island shelf (the world’s first high seas marine protected area) a carbon immobilization hotspot? Global Change Biology, 22(3), 1110–1120. 10.1111/gcb.13157 [DOI] [PubMed] [Google Scholar]
- Barnes, D. K. A. , Sands, C. J. , Richardson, A. , & Smith, N. (2019). Extremes in benthic ecosystem services; blue carbon natural capital shallower than 1000 m in isolated. Small, and Young Ascension Island’s EEZ, Frontiers in Marine Science. 10.3389/fmars.2019.00663 [DOI] [Google Scholar]
- Barragán, J. M. , & de Andrés, M. (2015). Analysis and trends of the world’s coastal cities and agglomerations. Ocean & Coastal Management, 114, 11–20. 10.1016/j.ocecoaman.2015.06.004 [DOI] [Google Scholar]
- Bax, N. , Sands, C. J. , Gogarty, B. , Downey, R. V. , Moreau, C. V. E. , Moreno, B. , Held, C. , Paulsen, M. L. , McGee, J. , Haward, M. , & Barnes, D. K. A. (2021). Perspective: Increasing blue carbon around Antarctica is an ecosystem service of considerable societal and economic value worth protecting. Global Change Biology, 27(1), 5–12. 10.1111/gcb.15392 [DOI] [PubMed] [Google Scholar]
- Bayraktarov, E. , Saunders, M. I. , Abdullah, S. , Mills, M. , Beher, J. , Possingham, H. P. , Mumby, P. J. , & Lovelock, C. E. (2016). The cost and feasibility of marine coastal restoration. Ecological Applications, 26(4), 1055–1074. 10.1890/15-1077 [DOI] [PubMed] [Google Scholar]
- Beatley, T. (2014). Blue urbanism: Exploring connections between cities and oceans. Island Press. [Google Scholar]
- Beatley, T. (2016). Handbook of biophilic city planning and design. Island Press. [Google Scholar]
- Belcher, A. , Henson, S. A. , Manno, C. , Hill, S. L. , Atkinson, A. , Thorpe, S. E. , Fretwell, P. , Ireland, L. , & Tarling, G. A. (2019). Krill faecal pellets drive hidden pulses of particulate organic carbon in the marginal ice zone. Nature Communications, 10(1), 889. 10.1038/s41467-019-08847-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bengtsson, J. , Bullock, J. M. , Egoh, B. , Everson, C. , Everson, T. , O’Connor, T. , O’Farrell, P. J. , Smith, H. G. , & Lindborg, R. (2019). Grasslands—More important for ecosystem services than you might think. Ecosphere, 10(2), e02582. [Google Scholar]
- Blue, G. , Daub, S. , Yunker, Z. , & Rajewicz, L. (2018). In the corporate interest: Fossil fuel industry input into Alberta and British Columbia’s climate leadership plans. Canadian Journal of Communication, 43(1). 10.22230/cjc.2018v43n1a3309 [DOI] [Google Scholar]
- Bond, W. J. , & Midgley, G. F. (2012). Carbon dioxide and the uneasy interactions of trees and savannah grasses. Philosophical Transactions of the Royal Society B: Biological Sciences, 367(1588), 601–612. 10.1098/rstb.2011.0182 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Breitburg, D. , Levin, L. A. , Oschlies, A. , Grégoire, M. , Chavez, F. P. , Conley, D. J. , Garçon, V. , Gilbert, D. , Gutiérrez, D. , Isensee, K. , Jacinto, G. S. , Limburg, K. E. , Montes, I. , Naqvi, S. W. A. , Pitcher, G. C. , Rabalais, N. N. , Roman, M. R. , Rose, K. A. , Seibel, B. A. , … Zhang, J. (2018). Declining oxygen in the global ocean and coastal waters. Science, 359(6371), eaam7240. 10.1126/science.aam7240 [DOI] [PubMed] [Google Scholar]
- Bruhwiler, L. , Parmentier, F.‐J. W. , Crill, P. , Leonard, M. , & Palmer, P. I. (2021). The Arctic carbon cycle and its response to changing climate. Current Climate Change Reports, 7(1), 14–34. 10.1007/s40641-020-00169-5 [DOI] [Google Scholar]
- Bull, J. W. , & Strange, N. (2018). The global extent of biodiversity offset implementation under no net loss policies. Nature Sustainability, 1(12), 790–798. 10.1038/s41893-018-0176-z [DOI] [Google Scholar]
- Bullock, E. L. , Woodcock, C. E. , Souza, C. , & Olofsson, P. (2020). Satellite‐based estimates reveal widespread forest degradation in the Amazon. Global Change Biology, 26(5), 2956–2969. 10.1111/gcb.15029 [DOI] [PubMed] [Google Scholar]
- Butchart, S. H. M. , Miloslavich, P. , Reyers, B. , Subramanian, S. M. , Adams, C. , Bennett, E. , Czucz, B. , Galetto, L. , Galvin, K. , Reyes‐Garcia, V. , Gerber, L. R. , Bekele, T. , Jetz, W. , Kosamu, I. B. M. , Palomo, M. G. , Panahi, M. , Selig, E. R. , Singh, G. S. , Tarkhnishvili, D. , Xu, H. , Lynch, A. J. , Mwampamba, T. H. , & Samakov, A. (2019). Chapter 3. Assessing progress towards meeting major international objectives related to nature and nature’s contributions to people. In Brondizio E. S., Settele J., Diaz S., & Ngo H. T. (Eds.), Global assessment report of the Intergovernmental Science‐Policy Platform on Biodiversity and Ecosystem Services. IPBES Secretariat, 214 pp. 10.5281/zenodo.3832053 [DOI] [Google Scholar]
- CAFF . (2021). Scoping for resilience and management of Arctic wetlands: Key findings and recommendations. Akureyri, Iceland. ISBN 978‐9935‐431‐97‐4. [Google Scholar]
- Cahoon, S. M. P. , Sullivan, P. F. , Post, E. , & Welker, J. M. (2012). Large herbivores limit CO2 uptake and suppress carbon cycle responses to warming in West Greenland. Global Change Biology, 18(2), 469–479. 10.1111/j.1365-2486.2011.02528.x [DOI] [Google Scholar]
- Cai, J. , Lovatelli, A. , Aguilar‐Manjarrez, J. , Cornish, L. , Dabbadie, L. , Desrochers, A. , Diffey, S. , Garrido, Gamarro E. , Geehan, J. , Hurtado, A. , Lucente, D. , Mair, G. , Miao, W. , Potin, P. , Przybyla, C. , Reantaso, M. , Roubach, R. , Tauati, M. , & Yuan, X. (2021). Seaweeds and microalgae: An overview for unlocking their potential in global aquaculture development. FAO. [Google Scholar]
- Canadell, J. G. , Monteiro, P. M. S. , Costa, M. H. , Cotrim da Cunha, L. , Cox, P. M. , Eliseev, A. V. , Henson, S. , Ishii, M. , Jaccard, S. , Koven, C. , Lohila, A. , Patra, P. K. , Piao, S. , Rogelj, J. , Syampungani, S. , Zaehle, S. , & Zickfeld, K. (2021). Global carbon and other biogeochemical cycles and feedbacks. In Masson‐Delmotte V., Zhai P., Pirani A., Connors S. L., Péan C., Berger S., Caud N., Chen Y., Goldfarb L., Gomis M. I., Huang M., Leitzell K., Lonnoy E., Matthews J. B. R., Maycock T. K., Waterfield T., Yelekçi O., Yu R., & Zhou B. (Eds.), Climate Change (2021). The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press. Available from https://www.ipcc.ch/report/ar6/wg1/downloads/report/IPCC_AR6_WGI_Chapter_05.pdf (last access 2021‐10‐17). [Google Scholar]
- CBD . (2021). First draft of the post‐2020 global biodiversity framework (CBD/WG2020/3/3). Convention on Biological Diversity. https://www.cbd.int/conferences/post2020/wg2020‐03/documents
- Chan, L. (2019). Chapter 5: Nature in the city. In Hamnet S., & Yuen B. (Eds.), Planning Singapore: The experimental city (pp. 109–129). London: Routledge. [Google Scholar]
- Chappell, A. , Baldock, J. , & Sanderman, J. (2016). The global significance of omitting soil erosion from soil organic carbon cycling schemes. Nature Climate Change, 6(2), 187–191. 10.1038/nclimate2829 [DOI] [Google Scholar]
- Chappell, A. , Webb, N. P. , Leys, J. F. , Waters, C. M. , Orgill, S. , & Eyres, M. J. (2019). Minimising soil organic carbon erosion by wind is critical for land degradation neutrality. Environmental Science & Policy, 93, 43–52. 10.1016/j.envsci.2018.12.020 [DOI] [Google Scholar]
- Chaudhary, A. , Sarkar, M. S. , Adhikari, B. S. , & Rawat, G. S. (2020). Ageratina adenophora and Lantana camara in Kailash Sacred Landscape, India: Current distribution and future climatic scenarios through modeling. BioRxiv. 10.1101/2020.09.14.295899 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaudhary, N. , Miller, P. A. , & Smith, B. (2017). Modelling Holocene peatland dynamics with an individual‐based dynamic vegetation model. Biogeosciences, 14(10), 2571–2596. 10.5194/bg-14-2571-2017 [DOI] [Google Scholar]
- Clark, M. A. , Springmann, M. , Hill,a J. , & Tilman, D. (2019). Multiple health and environmental impacts of foods. Proceedings of the National Academy of Sciences, 116, 23357–23362. 10.1073/pnas.1906908116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coady, D. , Parry, I. , Le, N.‐P. , & Shang, B. (2019). Global fossil fuel subsidies remain large: An update based on country‐level estimates. IMF Working Papers, 19(89), 39. 10.5089/9781484393178.001 [DOI] [Google Scholar]
- Conant, R. T. , Cerri, C. E. P. , Osborne, B. B. , & Paustian, K. (2017). Grassland management impacts on soil carbon stocks: A new synthesis. Ecological Applications, 27(2), 662–668. 10.1002/eap.1473 [DOI] [PubMed] [Google Scholar]
- Cottrell, R. S. , Metian, M. , Froehlich, H. E. , Blanchard, J. L. , Jacobsen, N. S. , McIntyre, P. B. , Nash, K. L. , Williams, D. R. , Bouwman, L. , Gephart, J. A. , Kuempel, C. D. , Moran, D. D. , Troell, M. , & Halpern, B. S. (2021). Time to rethink trophic levels in aquaculture policy. Reviews in Aquaculture, 10.1111/raq.12535 [DOI] [Google Scholar]
- Couwenberg, J. , Dommain, R. , & Joosten, H. (2009). Greenhouse gas fluxes from tropical peatlands in south‐east Asia: Greenhouse gas fluxes from tropical peatlands. Global Change Biology, 16(6), 1715–1732. 10.1111/j.1365-2486.2009.02016.x [DOI] [Google Scholar]
- Creed, I. F. , Jones, J. A. , Archer, E. , Claassen, M. , Ellison, D. , McNulty, S. G. , van Noordwijk, M. , Vira, B. , Wei, X. , Bishop, K. , Blanco, J. A. , Gush, M. , Gyawali, D. , Jobbágy, E. , Lara, A. , Little, C. , Martin‐Ortega, J. , Mukherji, A. , Murdiyarso, D. , … Xu, J. (2019). Managing forests for both downstream and downwind water. Frontiers in Forests and Global Change, 2, 10.3389/ffgc.2019.00064 [DOI] [Google Scholar]
- Creed, I. , van Noordwijk, M. , Archer, E. , Claassen, M. , Ellison, D. , Jones, J. A. , McNulty, S. G. , Vira, B. , & Wei, X. (2018). Forest, trees and water on a changing planet: How contemporary science can inform policy and practice. In Creed I. F., & van Noordwijk M. (Eds.), Forest and water on a changing planet: vulnerability, adaptation and governance opportunities: A global assessment report. IUFRO World Series, (Vol. 38, pp. 171–175). IUFRO. [Google Scholar]
- Crist, E. , Mora, C. , & Engelman, R. (2017). The interaction of human population, food production, and biodiversity protection. Science, 356(6335), 260–264. 10.1126/science.aal2011 [DOI] [PubMed] [Google Scholar]
- Cromsigt, J. , te Beest, M. , Kerley, G. I. H. , Landman, M. , le Roux, E. , & Smith, F. A. (2018). Trophic rewilding as a climate change mitigation strategy? Philosophical Transactions of the Royal Society B‐Biological Sciences, 10.1098/rstb.2017.0440 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Curtis, P. G. , Slay, C. M. , Harris, N. L. , Tyukavina, A. , & Hansen, M. C. (2018). Classifying drivers of global forest loss. Science, 361(6407), 1108–1111. [DOI] [PubMed] [Google Scholar]
- Davidson, T. A. , Audet, J. , Jeppesen, E. , Landkildehus, F. , Lauridsen, T. L. , Søndergaard, M. , & Syväranta, J. (2018). Synergy between nutrients and warming enhances methane ebullition from experimental lakes. Nat. Climate Change, 8, 156–160. 10.1038/s41558-017-0063-z [DOI] [Google Scholar]
- Deininger, A. , & Frigstad, H. (2019). Reevaluating the role of organic matter sources for coastal eutrophication, oligotrophication, and ecosystem health. Frontiers in Marine Science, 6, 210. 10.3389/fmars.2019.00210 [DOI] [Google Scholar]
- Dempsey, J. , Martin, T. G. , & Sumaila, U. R. (2020). Subsidizing extinction? Conservation Letters, 10.1111/conl.12705 [DOI] [Google Scholar]
- Deng, Y. , Li, X. , Shi, F. , Hu, X. , & Gillespie, T. (2021). Woody plant encroachment enhanced global vegetation greening and ecosystem water‐use efficiency. Global Ecology and Biogeography, 30(12), 2337–2353. 10.1111/geb.13386 [DOI] [Google Scholar]
- Deng, Y. , Wang, S. , Bai, X. , Luo, G. , Wu, L. , Chen, F. , Wang, J. , Li, C. , Yang, Y. , Hu, Z. , Tian, S. , & Lu, Q. (2020). Vegetation greening intensified soil drying in some semi‐arid and arid areas of the world. Agricultural and Forest Meteorology, 292–293, 10.1016/j.agrformet.2020.108103. 108103 [DOI] [Google Scholar]
- Di Sacco, A. , Hardwick, K. A. , Blakesley, D. , Brancalion, P. H. S. , Breman, E. , Cecilio Rebola, L. , Chomba, S. , Dixon, K. , Elliott, S. , Ruyonga, G. , Shaw, K. , Smith, P. , Smith, R. J. , & Antonelli, A. (2021). Ten golden rules for reforestation to optimize carbon sequestration, biodiversity recovery and livelihood benefits. Global Change Biology, 27, 1328–1348. 10.1111/gcb.15498 [DOI] [PubMed] [Google Scholar]
- Díaz, S. , Settele, J. , Brondízio, E. S. , Ngo, H. T. , Agard, J. , Arneth, A. , Balvanera, P. , Brauman, K. A. , Butchart, S. H. M. , Chan, K. M. A. , Garibaldi, L. A. , Ichii, K. , Liu, J. , Subramanian, S. M. , Midgley, G. F. , Miloslavich, P. , Molnár, Z. , Obura, D. , Pfaff, A. , … Zayas, C. N. (2019). Pervasive human‐driven decline of life on Earth points to the need for transformative change. Science (New York, N.Y.), 366. 10.1126/science.aax3100 [DOI] [PubMed] [Google Scholar]
- Dinerstein, E. , Joshi, A. R. , Vynne, C. , Lee, A. T. L. , Pharand‐Deschênes, F. , França, M. , Fernando, S. , Birch, T. , Burkart, K. , Asner, G. P. , & Olson, D. (2020). A “Global Safety Net” to reverse biodiversity loss and stabilize Earth’s climate. Science Advances, 6(36), eabb2824. 10.1126/sciadv.abb2824 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dinerstein, E. , Olson, D. , Joshi, A. , Vynne, C. , Burgess, N. D. , Wikramanayake, E. , Hahn, N. , Palminteri, S. , Hedao, P. , Noss, R. , Hansen, M. , Locke, H. , Ellis, E. C. , Jones, B. , Barber, C. V. , Hayes, R. , Kormos, C. , Martin, V. , Crist, E. , … Saleem, M. (2017). An ecoregion‐based approach to protecting half the terrestrial realm. BioScience, 67(6), 534–545. 10.1093/biosci/bix014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ditchburn, J.‐L. , & Carballeira, C. B. (2019). Versatility of the humble seaweed in Biomanufacturing. In 12th international conference interdisciplinarity in engineering, INTER‐ENG 2018, 4–5 October 2018, Tirgu Mures, Romania (Vol. 32, pp. 87–94). 10.1016/j.promfg.2019.02.187 [DOI] [Google Scholar]
- Donato, D. C. , Kauffman, J. B. , Murdiyarso, D. , Kurnianto, S. , Stidham, M. , & Kanninen, M. (2011). Mangroves among the most carbon‐rich forests in the tropics. Nature Geoscience, 4(5), 293–297. 10.1038/ngeo1123 [DOI] [Google Scholar]
- Donohue, R. J. , Roderick, M. L. , McVicar, T. R. , & Farquhar, G. D. (2013). Impact of CO2 fertilization on maximum foliage cover across the globe’s warm, arid environments. Geophysical Research Letters, 40(12), 3031–3035. 10.1002/grl.50563 [DOI] [Google Scholar]
- Doré, T. , Makowski, D. , Malézieux, E. , Munier‐Jolain, N. , Tchamitchian, M. , & Tittonell, P. (2011). Facing up to the paradigm of ecological intensification in agronomy: Revisiting methods, concepts and knowledge. European Journal of Agronomy, 34(4), 197–210. 10.1016/j.eja.2011.02.006 [DOI] [Google Scholar]
- D'Ottavio, P. , Francioni, M. , Trozzo, L. , Sedic, E. , Budimir, K. , Avanzolini, P. , Trombetta, M. F. , Porqueddu, C. , Santilocchi, R. , & Toderi, M. (2017). Trends and approaches in the analysis of ecosystem services provided by grazing systems: A review. Grass and Forage Science, 73(1), 15–24. [Google Scholar]
- Duarte, C. M. , & Krause‐Jensen, D. (2018). Intervention options to accelerate ecosystem recovery from coastal eutrophication. Front. Mar. Sci, 5, 470. 10.3389/fmars.2018.00470 [DOI] [Google Scholar]
- Duveiller, G. , Caporaso, L. , Abad‐Vinas, R. , Perugini, L. , Grassi, G. , Arneth, A. , & Cescatti, A. (2020). Local biophysical effects of land use and land cover change: Towards an assessment tool for policy makers. Land Use Policy, 91, 10.1016/j.landusepol.2019.104382 [DOI] [Google Scholar]
- Edwards, D. P. , Cerullo, G. R. , Chomba, S. , Worthington, T. A. , Balmford, A. P. , Chazdon, R. L. , & Harrison, R. D. (2021). Upscaling tropical restoration to deliver environmental benefits and socially equitable outcomes. Current Biology, 31(19), PR1326‐R1341. [DOI] [PubMed] [Google Scholar]
- Egli, L. , Meyer, C. , Scherber, C. , Kreft, H. , & Tscharntke, T. (2018). Winners and losers of national and global efforts to reconcile agricultural intensification and biodiversity conservation. Global Change Biology, 24, 2212–2228. 10.1111/gcb.14076 [DOI] [PubMed] [Google Scholar]
- Ekawati, S. , Subarudi, Xx., Budiningsih, K. , Sari, G. K. , & Muttaqin, M. Z. (2019). Policies affecting the implementation of REDD+ in Indonesia (cases in Papua, Riau and Central Kalimantan). Assessing Policies to Reduce Emissions from Land Use Change in Indonesia, 108, 101939. 10.1016/j.forpol.2019.05.025 [DOI] [Google Scholar]
- Engle, V. (2011). Estimating the provision of ecosystem services by Gulf of Mexico Coastal Wetlands. Wetlands, 31, 179–193. 10.1007/s13157-010-0132-9 [DOI] [Google Scholar]
- Enquist, B. J. , Feng, X. , Boyle, B. , Maitner, B. , Newman, E. A. , Jørgensen, P. M. , Roehrdanz, P. R. , Thiers, B. M. , Burger, J. R. , Corlett, R. T. , Couvreur, T. L. P. , Dauby, G. , Donoghue, J. C. , Foden, W. , Lovett, J. C. , Marquet, P. A. , Merow, C. , Midgley, G. , Morueta‐Holme, N. , … McGill, B. J. (2019). The commonness of rarity: Global and future distribution of rarity across land plants. Science. Advances, 5(11), eaaz0414. 10.1126/sciadv.aaz0414 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Enzi, V. , Cameron, B. , Dezsényi, P. , Gedge, D. , Mann, G. , & Pitha, U. (2017). Nature‐based solutions and buildings—The power of surfaces to help cities adapt to climate change and to deliver biodiversity (pp. 159–183). 10.1007/978-3-319-56091-5_10 [DOI] [Google Scholar]
- Epple, C. , Rangel, G. , Jenkins, M. , & Guth, M. (2016). Managing ecosystems in the context of climate change mitigation: A review of current knowledge and recommendations to support ecosystem‐based mitigation actions that look beyond terrestrial forests. Secretariat of the Convention on Biological Diversity. [Google Scholar]
- Falk, J. M. , Schmidt, N. M. , Christensen, T. R. , & Ström, L. (2015). Large herbivore grazing affects the vegetation structure and greenhouse gas balance in a high arctic mire. Environmental Research Letters, 10(4). 10.1088/1748-9326/10/4/045001 [DOI] [Google Scholar]
- FAO . (2019a). Climate‐smart agriculture and the Sustainable Development Goals: Mapping interlinkages, synergies and trade‐offs and guidelines for integrated implementation. FAO. [Google Scholar]
- FAO . (2019b). The State of the World’s biodiversity for food and agriculture. FAO Commission on Genetic Resources for Food and Agriculture Assessments. [Google Scholar]
- FAO . (2020). Global Forest Resources Assessment 2020: Main report. Rome, 10.4060/ca9825en [DOI] [Google Scholar]
- Fearnside, P. M. (2016). Greenhouse gas emissions from Brazil’s Amazonian hydroelectric dams. Environmental Research Letters, 11(1), 10.1088/1748-9326/11/1/011002. 011002. [DOI] [Google Scholar]
- Fensholt, R. , Langanke, T. , Rasmussen, K. , Reenberg, A. , Prince, S. D. , Tucker, C. , Scholes, R. J. , Le, Q. B. , Bondeau, A. , Eastman, R. , Epstein, H. , Gaughan, A. E. , Hellden, U. , Mbow, C. , Olsson, L. , Paruelo, J. , Schweitzer, C. , Seaquist, J. , & Wessels, K. (2012). Greenness in semi‐arid areas across the globe 1981–2007—An Earth Observing Satellite based analysis of trends and drivers. Remote Sensing of Environment, 121, 144–158. 10.1016/j.rse.2012.01.017 [DOI] [Google Scholar]
- Fisher, B. , Turner, R. K. , & Morling, P. (2009). Defining and classifying ecosystem services for decision making. Ecological Economics, 68(3), 643–653. 10.1016/j.ecolecon.2008.09.014 [DOI] [Google Scholar]
- Foale, S. , Adhuri, D. , Aliño, P. , Allison, E. H. , Andrew, N. , Cohen, P. , Evans, L. , Fabinyi, M. , Fidelman, P. , Gregory, C. , Stacey, N. , Tanzer, J. , & Weeratunge, N. (2013). Food security and the Coral Triangle Initiative. Marine Policy, 38, 174–183. 10.1016/j.marpol.2012.05.033 [DOI] [Google Scholar]
- Franks, M. , Lessmann, K. , Jakob, M. , Steckel, J. C. , & Edenhofer, O. (2018). Mobilizing domestic resources for the Agenda 2030 via carbon pricing. Nature Sustainability, 1(7), 350–357. 10.1038/s41893-018-0083-3 [DOI] [Google Scholar]
- Friedlingstein, P. , O'Sullivan, M. , Jones, M. W. , Andrew, R. M. , Hauck, J. , Olsen, A. , Peters, G. P. , Peters, W. , Pongratz, J. , Sitch, S. , Le Quéré, C. , Canadell, J. G. , Ciais, P. , Jackson, R. B. , Alin, S. , Aragão, L. E. O. C. , Arneth, A. , Arora, V. , Bates, N. R. , … Zaehle, S. (2020). Global Carbon Budget 2020. Earth Syst. Sci. Data, 12(3269–3340), 2020. 10.5194/essd-12-3269-2020 [DOI] [Google Scholar]
- Friess, D. A. , Richards, D. R. , & Phang, V. X. H. (2015). Mangrove forests store high densities of carbon across the tropical urban landscape of Singapore. Urban Ecosystem, 10.1007/s11252-015-0511-3 [DOI] [Google Scholar]
- Froehlich, H. E. , Afflerbach, J. C. , Frazier, M. , & Halpern, B. S. (2019). Blue growth potential to mitigate climate change through seaweed offsetting. Current Biology, 29(18), 3087–3093.e3. 10.1016/j.cub.2019.07.041 [DOI] [PubMed] [Google Scholar]
- Gallego‐Sala, A. V. , Charman, D. J. , Brewer, S. , Page, S. E. , Prentice, I. C. , Friedlingstein, P. , Moreton, S. , Amesbury, M. J. , Beilman, D. W. , Björck, S. , Blyakharchuk, T. , Bochicchio, C. , Booth, R. K. , Bunbury, J. , Camill, P. , Carless, D. , Chimner, R. A. , Clifford, M. , Cressey, E. , … Zhao, Y. (2018). Latitudinal limits to the predicted increase of the peatland carbon sink with warming. Nature Climate Change, 8(10), 907–913. 10.1038/s41558-018-0271-1 [DOI] [Google Scholar]
- Galloway, J. N. , Aber, J. D. , Erisman, J. W. , Seitzinger, S. P. , Howarth, R. W. , Cowling, E. B. , & Cosby, B. J. (2003). The nitrogen cascade. BioScience, 53(4), 341–356. 10.1641/0006-3568(2003)053[0341:TNC]2.0.CO;2 [DOI] [Google Scholar]
- Gao, G. G. , Clare, A. S. , Rose, C. , & Caldwell, G. S. (2017). Eutrophication and warming‐driven green tides (Ulva rigida) are predicted to increase under future climate change scenarios. Marine Pollution Bulletin, 114, 439–447. 10.1016/j.marpolbul.2016.10.003 [DOI] [PubMed] [Google Scholar]
- Gerber, J. S. , Carlson, K. M. , Garcia de Cortazar‐Atauri, I. , Herrero, M. , Launay, M. , Makowski, D. , Mueller, N. D. , O’Connell, C. S. , Smith, P. , & West, P. C. (2016). Spatially explicit N2O emissions estimates suggest climate mitigation opportunities from improved nitrogen management. Global Change Biology, 22, 3383–3394. 10.1111/gcb.13341 [DOI] [PubMed] [Google Scholar]
- Gilbert, P. (2020). (2020) Harmful algae at the complex nexus of eutrophication and climate change. Harmful Algae, 91, 101583. 10.1016/j.hal.2019.03.001 [DOI] [PubMed] [Google Scholar]
- Giller, K. E. , Hijbeek, R. , Andersson, J. A. , & Sumberg, J. (2021). Regenerative agriculture: An agronomic perspective. Outlook on Agriculture (in press), 10.1177/0030727021998063 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Girardin, C. A. J. , Jenkins, S. , Seddon, N. , Allen, M. , Lewis, S. L. , Wheeler, C. E. , Griscom, B. W. , & Malhi, Y. (2021). Nature‐based solutions can help cool the planet—If we act now. Nature, 593, 191–194. 10.1038/d41586-021-01241-2 [DOI] [PubMed] [Google Scholar]
- Gizachew, B. , Astrup, R. , Vedeld, P. , Zahabu, E. M. , & Duguma, L. A. (2017). REDD+ in Africa: Contexts and challenges. Natural Resources Forum, 41(2), 92–104. 10.1111/1477-8947.12119 [DOI] [Google Scholar]
- Gosnell, H. , Grimm, K. , & Goldstein, B. E. (2020). A half century of Holistic Management: What does the evidence reveal? Agriculture and Human Values, 37(3), 849–867. 10.1007/s10460-020-10016-w [DOI] [Google Scholar]
- Green, R. E. , Cornell, S. J. , Scharlemann, J. P. W. , & Balmford, A. (2005). Farming and the Fate of Wild Nature. Science, 307(5709), 550–555. 10.1126/science.1106049 [DOI] [PubMed] [Google Scholar]
- Griffith, A. , & Gobler, C. (2019). Harmful algal blooms: A climate change co‐stressor in marine and freshwater ecosystems. Harmful Algae, 91, 10.1016/j.hal.2019.03.008 [DOI] [PubMed] [Google Scholar]
- Griscom, B. W. , Adams, J. , Ellis, P. W. , Houghton, R. A. , Lomax, G. , Miteva, D. A. , Schlesinger, W. H. , Shoch, D. , Siikamäki, J. V. , Smith, P. , Woodbury, P. , Zganjar, C. , Blackman, A. , Campari, J. , Conant, R. T. , Delgado, C. , Elias, P. , Gopalakrishna, T. , Hamsik, M. R. , … Fargione, J. (2017). Natural climate solutions. Proceedings of the National Academy of Sciences, 114(44), 11645–11650. 10.1073/pnas.1710465114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guannel, G. , Arkema, K. , Ruggiero, P. , & Verutes, G. (2016). The power of three: Coral reefs, seagrasses and mangroves protect coastal regions and increase their resilience. PLoS One, 11(7), e0158094. 10.1371/journal.pone.0158094 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gullison, R. E. , Frumhoff, P. C. , Canadell, J. G. , Field, C. B. , Nepstad, D. C. , Hayhoe, K. , Avissar, R. , Curran, L. M. , Friedlingstein, P. , Jones, C. D. , & Nobre, C. (2007). Environment: Tropical forests and climate policy. Science, 316(5827), 985–986. 10.1126/science.1136163 [DOI] [PubMed] [Google Scholar]
- Gustavsson, J. , Cederberg, C. , & Sonesson, U. (2011). Global food losses and food waste: Extent, causes and prevention; study conducted for the International Congress Save Food! at Interpack 2011, [16 ‐ 17 May]. Food and Agriculture Organization of the United Nations. [Google Scholar]
- Hannah, L. , Roehrdanz, P. R. , Marquet, P. A. , Enquist, B. J. , Midgley, G. , Foden, W. , Lovett, J. C. , Corlett, R. T. , Corcoran, D. , Butchart, S. H. M. , Boyle, B. , Feng, X. , Maitner, B. , Fajardo, J. , McGill, B. J. , Merow, C. , Morueta‐Holme, N. , Newman, E. A. , Park, D. S. , … Svenning, J.‐C. (2020). 30% land conservation and climate action reduces tropical extinction risk by more than 50%. Ecography, 43(943–953), 10.1111/ecog.05166 [DOI] [Google Scholar]
- Harlio, A. , Kuussaari, M. , Heikkinen, R. K. , & Arponen, A. (2019). Incorporating landscape heterogeneity into multi‐objective spatial planning improves biodiversity conservation of semi‐natural grasslands. Journal for Nature Conservation, 49, 37–44. 10.1016/j.jnc.2019.01.003 [DOI] [Google Scholar]
- Hayashida, S. , Ono, A. , Yoshizaki, S. , Frankenberg, C. , Takeuchi, W. , & Yan, X. (2013). Methane concentrations over Monsoon Asia as observed by SCIAMACHY: Signals of methane emission from rice cultivation. Remote Sensing of Environment, 139, 246–256. [Google Scholar]
- Hayek, M. N. , Harwatt, H. , Ripple, W. J. , & Mueller, N. D. (2021). The carbon opportunity cost of animal‐sourced food production on land. Nature Sustainability, 4(1), 21–24. 10.1038/s41893-020-00603-4 [DOI] [Google Scholar]
- Heilmayr, R. , Echeverría, C. , & Lambin, E. F. (2020). Impacts of Chilean forest subsidies on forest cover, carbon and biodiversity. Nature Sustainability, 3(9), 701–709. 10.1038/s41893-020-0547-0 [DOI] [Google Scholar]
- Heithaus, M. R. , Alcoverro, T. , Arthur, R. , Burkholder, D. A. , Coates, K. A. , Christianen, M. J. A. , Kelkar, N. , Manuel, S. A. , Wirsing, A. J. , Kenworthy, W. J. , & Fourqurean, J. W. (2014). Seagrasses in the age of sea turtle conservation and shark overfishing. Frontiers in Marine Science, 1, 10.3389/fmars.2014.00028 [DOI] [Google Scholar]
- Hoegh‐Guldberg, O. , Jacob, D. , Taylor, M. , Bindi, M. , Brown, S. , Camilloni, I. , Diedhiou, A. , Djalante, R. , Ebi, K. L. , Engelbrecht, F. , Guiot, J. , Hijioka, Y. , Mehrotra, S. , Payne, A. , Seneviratne, S. I. , Thomas, A. , Warren, R. , & Zhou, G. (2018). Impacts of 1.5ºC global warming on natural and human systems. In Masson‐Delmotte V., Zhai P., Pörtner H.‐O., Roberts D., Skea J., Shukla P. R., Pirani A., Moufouma‐Okia W., Péan C., Pidcock R., Connors S., Matthews J. B. R., Chen Y., Zhou X., Gomis M. I., Lonnoy E., Maycock T., Tignor M., & Waterfield T. (Eds.), Global Warming of 1.5°C. An IPCC Special Report on the impacts of global warming of 1.5°C above pre‐industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty. IPCC. [Google Scholar]
- Huntley, B. J. , & Redford, K. H. (2014). Mainstreaming biodiversity in practice: A STAP advisory document. Global Environment Facility. [Google Scholar]
- IPBES . (2018). The IPBES assessment report on land degradation and restoration (p. 744). Secretariat of the Intergovernmental Science‐Policy Platform on Biodiversity and Ecosystem Services. [Google Scholar]
- IPBES . (2019a). Global assessment report on biodiversity and ecosystem services of the Intergovernmental Science‐ Policy Platform on Biodiversity and Ecosystem Services. Secretariat of the Intergovernmental Science‐Policy Platform on Biodiversity and Ecosystem services; [Google Scholar]
- IPBES . (2019b). Glossary. Secretariat of the Intergovernmental Science‐Policy Platform on Biodiversity and Ecosystem services. Retrieved from: https://www.ipbes.net/glossary [Google Scholar]
- IPCC . (2013). In Stocker T. F., Qin D., Plattner G.‐K., Tignor M., Allen S. K., Boschung J., Nauels A., Xia Y., Bex V., & Midgley P. M. (Eds.), Climate change 2013: The physical science basis. contribution of working group I to the fifth assessment report of the intergovernmental panel on climate change. Cambridge University Press, 1535 pp. [Google Scholar]
- IPCC . (2014). In C. B. Field, V.R. Barros, D. J. Dokken, K. J. Mach, M.D. Mastrandrea, T. E. Bilir, M. Chatterjee, K. L. Ebi, Y.O. Estrada, R. C. Genova, B. Girma, E. S. Kissel, A. N. Levy, S. MacCracken, P. R. Mastrandrea, & L. L. White (Eds.), Climate Change 2014: Impacts, adaptation, and vulnerability. Part A: Global and sectoral aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, 1132 pp. [Google Scholar]
- IPCC . (2019). In Shukla P. R., Skea J., Calvo Buendia E., Masson‐Delmotte V., Pörtner H.‐O., Roberts D. C., Zhai P., Slade R., Connors S., van Diemen R., Ferrat M., Haughey E., Luz S., Neogi S., Pathak M., Petzold J., Portugal Pereira J., Vyas P., Huntley E., Kissick K., Belkacemi M., & Malley J. (Eds.), Climate change and land: An IPCC special report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystem, in press. IPCC. [Google Scholar]
- Ise, T. , Dunn, A. L. , Wofsy, S. C. , & Moorcroft, P. R. (2008). High sensitivity of peat decomposition to climate change through water‐table feedback. Nature Geoscience, 1(11), 763–766. 10.1038/ngeo331 [DOI] [Google Scholar]
- IUCN . (2017). The Sundarbans 2017 conservation outlook assessment. IUCN. [Google Scholar]
- IUCN . (2020). Sundarbans National Park 2020 conservation outlook assessment (p. 24). IUCN. [Google Scholar]
- Jahangir, M. M. R. , Richards, K. , Healy, M. , Gill, L. , Müller, C. , Johnston, P. , & Fenton, O. (2016). Carbon and nitrogen dynamics and greenhouse gas emissions in constructed wetlands treating wastewater: A review. Hydrology and Earth System Sciences, 20, 109–123. 10.5194/hess-20-109-2016 [DOI] [Google Scholar]
- Jane, S. F. , Hansen, G. J. A. , Kraemer, B. M. , Leavitt, P. R. , Mincer, J. L. , North, R. L. , Pilla, R. M. , Stetler, J. T. , Williamson, C. E. , Iestyn Woolway, R. , Arvola, L. , Chandra, S. , DeGasperi, C. L. , Diemer, L. , Dunalska, J. , Erina, O. , Flaim, G. , Grossart, H.‐P. , Hambright, K. D. , … Rose, K. C. (2021). Widespread deoxygenation of temperate lakes. Nature, 594, 66–70. 10.1038/s41586-021-03550-y [DOI] [PubMed] [Google Scholar]
- Jantz, P. , Goetz, S. , & Laporte, N. (2014). Carbon stock corridors to mitigate climate change and promote biodiversity in the tropics. Nature Climate Change, 4(2), 138–142. 10.1038/nclimate2105 [DOI] [Google Scholar]
- Jeppesen, E. , Moss, B. , Bennion, H. , Carvalho, L. , DeMeester, L. , Feuchtmayr, H. , Friberg, N. , Gessner, M. O. , Hefting, M. , Lauridsen, T. L. , & Liboriussen, L. (2010). Interaction of climate change and eutrophication. Kernan/Climate Change Impacts on Freshwater Ecosystems, 119–51, 10.1002/9781444327397.CH6 [DOI] [Google Scholar]
- Jia, G. , Shevliakova, E. , Artaxo, P. , De Noblet‐Ducoudre, N. , Houghton, R. , House, J. , Kitajima, K. , Lennard, C. , Popp, A. , Sirin, A. , Sukumar, R. , & Verchot, L. (2019). Land–climate interactions. In Shukla P. R., Skea J., Calvo Buendia E., Masson‐Delmotte V., Pörtner H.‐O., Roberts D. C., Zhai P., Slade R., Connors S., R. van Diemen , Ferrat M., Haughey E., Luz S., Neogi S., Pathak M., Petzold J., Portugal Pereira J., Vyas P., Huntley E., … Malley J. (Eds.), Climate Change and Land: An IPCC special report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems (pp. 131–247). IPCC. [Google Scholar]
- Johnson, B. A. , Dasgupta, R. , Mader, A. D. , & Scheyvens, H. (2019). Understanding national biodiversity targets in a REDD+ context. Environmental Science & Policy, 92, 27–33. 10.1016/j.envsci.2018.11.007 [DOI] [Google Scholar]
- Joly, C. A. , Scarano, F. R. , Bustamante, M. , Gadda, T. , Metzger, J. P. , Seixas, C. S. , Ometto, J.‐P. , Pires, A. P. F. , Boesing, A. L. , Sousa, F. D. R. , Quintão, J. M. , Gonçalves, L. , Padgurschi, M. C. G. , de Aquino, M. F. S. , de Castro, P. D. , & de Santos, I. L. (2018). Sumário para tomadores de decisão: 1o diagnóstico brasileiro de biodiversidade e serviços ecossistêmicos. 10, 5935. 10.5935/978-85-5697-707-6 [DOI]
- Jones, J. P. G. , Bull, J. W. , Roe, D. , Baker, J. , Griffiths, V. F. , Starkey, M. , Sonter, L. J. , & Milner‐Gulland, E. J. (2019). Net gain: Seeking better outcomes for local people when mitigating biodiversity loss from development. One Earth, 1(2), 195–201. 10.1016/j.oneear.2019.09.007 [DOI] [Google Scholar]
- Joshi, S. , Liang, Y. , Wang, J. , Pasakhala, B. , Ismail, M. , Bisht, N. , Qamer, F. M. , Long, R. , Ning, W. , Foggin, M. , Weikang, Y. , Bari, F. , Shokirov, Q. , Ahmed, A. , Essa, M. , & Wenxuan, X. (2019). Rangeland resource and use assessment protocol.. International Centre for Integrated Mountain Development. ISBN 978 92 9115 687 0 (electronic). [Google Scholar]
- Keeley, A. T. H. , Basson, G. , Cameron, D. R. , Heller, N. E. , Huber, P. R. , Schloss, C. A. , Thorne, J. H. , & Merenlender, A. M. (2018). Making habitat connectivity a reality. Conservation Biology, 32(6), 1221–1232. 10.1111/cobi.13158 [DOI] [PubMed] [Google Scholar]
- Kemppinen, K. M. S. , Collins, P. M. , Hole, D. G. , Wolf, C. , Ripple, W. J. , & Gerber, L. R. (2020). Global reforestation and biodiversity conservation. Conservation Biology, 34(5), 1221–1228. 10.1111/cobi.13478 [DOI] [PubMed] [Google Scholar]
- Kessouri, F. , McWilliams, J. C. , Bianchi, D. , Sutula, M. , Renault, L. , Deutsch, C. , Feely, R. A. , McLaughlin, K. , Ho, M. , Howard, E. M. , Bednarsek, N. , Damien, P. , Molemaker, J. , & Weisberg, S. (2021). Coastal eutrophication drives acidification, oxygen loss, and ecosystem change in a major oceanic upwelling system. Coastal eutrophication drives acidification, oxygen loss, and ecosystem change in a major oceanic upwelling system. Proceedings of the National Academy of Sciences of the United States of America, 118 (21) e2018856118. 10.1073/pnas.2018856118 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim, J. , Stekoll, M. , & Yarish, C. (2019). Opportunities, challenges and future directions of open‐water seaweed aquaculture in the United States. Phycologia, 58, 446–461. [Google Scholar]
- Knowler, D. , Chopin, T. , Martínez‐Espiñeira, R. , Neori, A. , Nobre, A. , Noce, A. , & Reid, G. (2020). The economics of Integrated Multi‐Trophic Aquaculture: Where are we now and where do we need to go? Reviews in Aquaculture, 12, 1579–1594. [Google Scholar]
- Kremen, C. (2015). Reframing the land‐sparing/land‐sharing debate for biodiversity conservation. Annals of the New York Academy of Sciences, 1355, 52–76. 10.1111/nyas.12845 [DOI] [PubMed] [Google Scholar]
- Laurance, W. F. , Sayer, J. , & Cassman, K. G. (2014). Agricultural expansion and its impacts on tropical nature. Trends in Ecology & Evolution, 29(2), 107–116. 10.1016/j.tree.2013.12.001 [DOI] [PubMed] [Google Scholar]
- Lavery, T. J. , Roudnew, B. , Gill, P. , Seymour, J. , Seuront, L. , Johnson, G. , Mitchell, J. G. , & Smetacek, V. (2010). Iron defecation by sperm whales stimulates carbon export in the Southern Ocean. Proceedings of the Royal Society B: Biological Sciences, 277(1699), 3527–3531. 10.1098/rspb.2010.0863 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leippert, F. , Darmaun, M. , Bernoux, M. , & Mpheshea, M. (2020). The potential of agroecology to build climate‐resilient livelihoods and food systems. FAO and Biovision. 10.4060/cb0438en [DOI] [Google Scholar]
- Leroux, L. , Bégué, A. , Seen, D. L. , Jolivot, A. , & Kayitakire, F. (2017). Driving forces of recent vegetation changes in the Sahel: Lessons learned from regional and local level analyses. Remote Sensing of Environment, 191, 38–54. [Google Scholar]
- Lewis, S. L. , Wheeler, C. E. , Mitchard, E. T. A. , & Koch, A. (2019). Restoring natural forests is the best way to remove atmospheric carbon. Nature, 568(7750), 25–28. 10.1038/d41586-019-01026-8 [DOI] [PubMed] [Google Scholar]
- Li, Y. , Shang, J. , Zhang, C. , Zhang, W. , Niu, L. , Wang, L. , & Zhang, H. (2021). The role of freshwater eutrophication in greenhouse gas emissions: A review. Science of the Total Environment, 768(2021), 144582. [DOI] [PubMed] [Google Scholar]
- Liniger, H. P. , Bandy, J. , Bhuchar, S. , & Joshi, R. (2020). Spring revival through sustainable land management (SLM) in the Himalayan foothills: Uttarakhand, North India. WOCAT SLM Policy Brief, No. 1, WOCAT.
- Linsy, P. , Nagender, N. B. , Mascarenhas‐Pereira, M. B. L. , Vinitha, P. V. , Ray, D. , Prakash, B. C. , Ramalingeswara, R. B. , Kazip, A. , Sebastian, T. , Kocherla, M. , & Miriyala, P. (2018). Benthic cycling of phosphorus in the Eastern Arabian Sea: Evidence of present day phosphogenesis. Marine Chemistry, 199, 53–66. 10.1016/j.marchem.2018.01.007 [DOI] [Google Scholar]
- Littlefield, C. E. , Krosby, M. , Michalak, J. L. , & Lawler, J. J. (2019). Connectivity for species on the move: Supporting climate‐driven range shifts. Frontiers in Ecology and the Environment, 17(5), 270–278. 10.1002/fee.2043 [DOI] [Google Scholar]
- Liu, L. , Xu, W. , Yue, Q. , Teng, X. , & Hu, H. (2018). Problems and countermeasures of coastline protection and utilization in China. Ocean & Coastal Management, 153, 124–130. 10.1016/j.ocecoaman.2017.12.016 [DOI] [Google Scholar]
- Loke, L. H. L. , Heery, E. C. , & Todd, P. A. (2019). Shoreline defences. In Sheppard C. (Ed.), World seas: An environmental evaluation, vol. 3, ecological issues and environmental impacts (2nd ed., pp. 491–504). Academic Press. [Google Scholar]
- Lomnitz, U. , Sommer, S. , Dale, A. W. , Löscher, C. R. , Noffke, A. , Wallmann, K. , & Hensen, C. (2016). Benthic phosphorus cycling in the Peruvian oxygen minimum zone. Biogeosciences, 13(5), 1367–1386. 10.5194/bg-13-1367-2016 [DOI] [Google Scholar]
- Lundberg, S. , & Marklund, P.‐O. (2018). Green public procurement and multiple environmental objectives. Economia E Politica Industriale, 45(1), 37–53. 10.1007/s40812-017-0085-6 [DOI] [Google Scholar]
- Mackey, B. , Kormos, C. F. , Keith, H. , Moomaw, W. R. , Houghton, R. A. , Mittermeier, R. A. , Hole, D. , & Hugh, S. (2020). Understanding the importance of primary tropical forest protection as a mitigation strategy. Mitigation and Adaptation Strategies for Global Change, 25(5), 763–787. 10.1007/s11027-019-09891-4 [DOI] [Google Scholar]
- Malhi, Y. , Roberts, J. T. , Betts, R. A. , Killeen, T. J. , Li, W. , & Nobre, C. A. (2008). Climate change, deforestation, and the fate of the Amazon. Science, 319(5860), 169–172. 10.1126/science.1146961 [DOI] [PubMed] [Google Scholar]
- Mariani, G. , Cheung, W. W. L. , Lyet, A. , Sala, E. , Mayorga, J. , Velez, L. , Gaines, S. D. , Dejean, T. , Troussellier, M. , & Mouillot, D. (2020). Let more big fish sink: Fisheries prevent blue carbon sequestration—Half in unprofitable areas. Science. Advances, 6(44), eabb4848. 10.1126/sciadv.abb4848 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matricardi, E. A. T. , Skole, D. L. , Costa, O. B. , Pedlowski, M. A. , Samek, J. H. , & Miguel, E. P. (2020). Long‐term forest degradation surpasses deforestation in the Brazilian Amazon. Science, 369(6509), 1378–1382. 10.1126/science.abb3021 [DOI] [PubMed] [Google Scholar]
- Maxton‐Lee, B. (2018). Material realities: Why indonesian deforestation persists and conservation fails. Journal of Contemporary Asia, 48(3), 419–444. 10.1080/00472336.2017.1402204 [DOI] [Google Scholar]
- Maxwell, S. L. , Evans, T. , Watson, J. E. M. , Morel, A. , Grantham, H. , Duncan, A. , Harris, N. , Potapov, P. , Runting, R. K. , Venter, O. , Wang, S. , & Malhi, Y. (2019). Degradation and forgone removals increase the carbon impact of intact forest loss by 626%. Science Advances, 5(10), 10.1126/sciadv.aax2546 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mbow, C. , Van Noordwijk, M. , Luedeling, E. , Neufeldt, H. , Minang, P. A. , & Kowero, G. (2014). Agroforestry solutions to address food security and climate change challenges in Africa. Current Opinion in Environmental Sustainability, 6, 61–67. 10.1016/j.cosust.2013.10.014 [DOI] [Google Scholar]
- McFarland, W. , Whitley, S. , & Kissinger, G. (2015). Subsidies to key commodities driving forest loss. https://static1.squarespace.com/static/58d6cc1e17bffcffb801edde/t/5970e3c6e3df28c64461c095/1500570579966/Subsidies+driving+forest+loss.pdf
- Mcleod, E. , Chmura, G. L. , Bouillon, S. , Salm, R. , Björk, M. , Duarte, C. M. , Lovelock, C. E. , Schlesinger, W. H. , & Silliman, B. R. (2011). A blueprint for blue carbon: Toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2 . Frontiers in Ecology and the Environment, 9(10), 552–560. 10.1890/110004 [DOI] [Google Scholar]
- Melillo, J. M. , Lu, X. , Kicklighter, D. W. , Reilly, J. M. , Cai, Y. , & Sokolov, A. P. (2016). Protected areas’ role in climate‐change mitigation. Ambio, 45(2), 133–145. 10.1007/s13280-015-0693-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ministry of Environment and Tourism . (2011). Namibia second national communication to the united nations framework convention on climate change. Republic of Namibia, Ministry of Environment and Tourism. [Google Scholar]
- Mishra, U. , Hugelius, G. , Shelef, E. , Yang, Y. , Strauss, J. , Lupachev, A. , Harden, J. W. , Jastrow, J. D. , Ping, C.‐L. , Riley, W. J. , Schuur, E. A. G. , Matamala, R. , Siewert, M. , Nave, L. E. , Koven, C. D. , & Orr, A. (2021). Spatial heterogeneity and environmental predictors of permafrost region soil organic carbon stocks. Science. Advances, 7(9), eaaz5236, 10.1126/sciadv.aaz5236 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moilanen, A. , Franco, A. M. A. , Early, R. I. , Fox, R. , Wintle, B. , & Thomas, C. D. (2005). Prioritizing multiple‐use landscapes for conservation: Methods for large multi‐species planning problems. Proceedings of the Royal Society B: Biological Sciences, 272(1575), 1885–1891. 10.1098/rspb.2005.3164 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mustin, K. , Carvalho, W. D. , Hilário, R. R. , Costa‐Neto, S. V. , Silva, C. , Vasconcelos, I. M. , Castro, I. J. , Eilers, V. , Kauano, É. E. , Mendes‐Junior, R. N. G. , Funi, C. , Fearnside, P. M. , Silva, J. M. C. , Euler, A. M. C. , & Toledo, J. J. (2017). Biodiversity, threats and conservation challenges in the Cerrado of Amapá, an Amazonian savanna. Nature Conservation, 22, 107–127. 10.3897/natureconservation.22.13823 [DOI] [Google Scholar]
- Myers, R. A. , & Worm, B. (2003). Rapid worldwide depletion of predatory fish communities. Nature, 423(6937), 280–283. 10.1038/nature01610 [DOI] [PubMed] [Google Scholar]
- Naqvi, S. W. A. , Bange, H. , Farias, L. , Monteiro, P. , Scranton, M. , & Zhang, J. (2010). Marine hypoxia/anoxia as a source of CH4 and N2O. Biogeosciences, 7, 10.5194/bg-7-2159-2010 [DOI] [Google Scholar]
- Nazari‐Sharabian, M. , Ahmad, S. , & Karakouzian, M. (2018). Climate change and eutrophication: A short review. Engineering, Technology and Applied Science Research, 8(6), 3668–3672. [Google Scholar]
- Nepal, M. , Rai, R. , Saudamini, D. , Bhatta, L. , Kotru, R. , Khadayat, M. , Rawal, R. , & Gcs, N. (2018). Valuing cultural services of the kailash sacred landscape for sustainable management. Sustainability, 10, 10.3390/su10103638 [DOI] [Google Scholar]
- Newbold, T. , Hudson, L. N. , Arnell, A. P. , Contu, S. , Palma, A. D. , Ferrier, S. , Hill, S. L. L. , Hoskins, A. J. , Lysenko, I. , Phillips, H. R. P. , Burton, V. J. , Chng, C. W. T. , Emerson, S. , Gao, D. , Pask‐Hale, G. , Hutton, J. , Jung, M. , Sanchez‐Ortiz, K. , Simmons, B. I. , … Purvis, A. (2016). Has land use pushed terrestrial biodiversity beyond the planetary boundary? A Global Assessment. Science, 353(6296), 288–291. 10.1126/science.aaf2201 [DOI] [PubMed] [Google Scholar]
- Newbold, T. , Hudson, L. N. , Hill, S. L. L. , Contu, S. , Lysenko, I. , Senior, R. A. , Börger, L. , Bennett, D. J. , Choimes, A. , Collen, B. , Day, J. , Palma, A. D. , Díaz, S. , Echeverria‐Londoño, S. , Edgar, M. J. , Feldman, A. , Garon, M. , Harrison, M. L. K. , Alhusseini, T. , … Purvis, A. (2015). Global effects of land use on local terrestrial biodiversity. Nature, 520(7545), 45–50. 10.1038/nature14324 [DOI] [PubMed] [Google Scholar]
- Newmark, W. D. , Jenkins, C. N. , Pimm, S. L. , McNeally, P. B. , & Halley, J. M. (2017). Targeted habitat restoration can reduce extinction rates in fragmented forests. Proceedings of the National Academy of Sciences, 114(36), 9635–9640. 10.1073/pnas.1705834114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Leary, B. C. , Winther‐Janson, M. , Bainbridge, J. M. , Aitken, J. , Hawkins, J. P. , & Roberts, C. M. (2016). Effective coverage targets for ocean protection. Conservation Letters, 9(6), 398–404. 10.1111/conl.12247 [DOI] [Google Scholar]
- OECD . (2005). Environmentally harmful subsidies. 10.1787/9789264012059-en [DOI] [Google Scholar]
- OECD . (2019). Biodiversity: Finance and the Economic and Business Case for Action, report prepared for the G7 Environment Ministers’. OECD. [Google Scholar]
- Opekunova, M. G. , Opekunov, A. Y. , Kukushkin, S. Y. , & Arestova, I. Y. (2018). Evaluation of environmental transformation in areas of hydrocarbon deposits in the North of Western Siberia. Contemporary Problems of Ecology, 11(1), 99–110. 10.1134/S1995425518010109 [DOI] [Google Scholar]
- Oppenheimer, M. , Glavovic, B. C. , Hinkel, J. , van de Wal, R. , Magnan, A. K. , Abd‐Elgawad, A. , Cai, R. , Cifuentes‐Jara, M. , DeConto, R. M. , Ghosh, T. , Hay, J. , Isla, F. , Marzeion, B. , Meyssignac, B. , & Sebesvari, Z. (2019). Sea level rise and implications for low‐lying islands, coasts and communities. In Pörtner H.‐O., Roberts D. C., Masson‐Delmotte V., Zhai P., Tignor M., Poloczanska E., Mintenbeck K., Alegría A., Nicolai M., Okem A., Petzold J., Rama B., & Weyer N. M. (Eds.), IPCC special report on the ocean and cryosphere in a changing climate. IPCC. [Google Scholar]
- Pandey, A. , Kotru, R. , & Pradhan, N. (2016). Bridging cultural heritage, conservation and development through a transboundary landscape approach. In Asian sacred natural sites: Philosophy and practice in protected areas and conservation (pp. 145–158). Routledge. [Google Scholar]
- Pangala, S. R. , Enrich‐Prast, A. , Basso, L. S. , Peixoto, R. B. , Bastviken, D. , Hornibrook, E. R. C. , Gatti, L. V. , Marotta, H. , Calazans, L. S. B. , Sakuragui, C. M. , Bastos, W. R. , Malm, O. , Gloor, E. , Miller, J. B. , & Gauci, V. (2017). Large emissions from floodplain trees close the Amazon methane budget. Nature, 552(7684), 230–234. 10.1038/nature24639 [DOI] [PubMed] [Google Scholar]
- Passow, U. , & Carlson, C. (2012). The biological pump in a high CO2 world. Marine Ecology Progress Series, 470, 249–271. 10.3354/meps09985 [DOI] [Google Scholar]
- Patra, P. K. , Saeki, T. , Dlugokencky, E. J. , Ishijima, K. , Umezawa, T. , Ito, A. , Aoki, S. , Morimoto, S. , Kort, E. A. , Crotwell, A. , Ravi, Kumar K. , & Nakazawa, T. (2016). Regional methane emission estimation based on observed atmospheric concentrations (2002–2012). Journal of the Meteorological Society of Japan (Ser. II), 94(1), 91–113. 10.2151/jmsj.2016-006 [DOI]
- Pearson, T. R. H. , Brown, S. , Murray, L. , & Sidman, G. (2017). Greenhouse gas emissions from tropical forest degradation: an underestimated source. Carbon Balance and Management, 12, 10.1186/s13021-017-0072-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perino, A. , Pereira, H. M. , Navarro, L. M. , Fernandez, N. , Bullock, J. M. , Ceausu, S. , Cortes‐Avizanda, A. , van Klink, R. , Kuemmerle, T. , Lomba, A. , Pe'er, G. , Plieninger, T. , Benayas, J. M. R. , Sandom, C. J. , Svenning, J. C. , & Wheeler, H. C. (2019). Rewilding complex ecosystems. Science, 10.1126/science.aav5570 [DOI] [PubMed] [Google Scholar]
- Pershing, A. J. , Christensen, L. B. , Record, N. R. , Sherwood, G. D. , & Stetson, P. B. (2010). The impact of whaling on the ocean carbon cycle: Why bigger was better. PLoS One, 5(8), e12444. 10.1371/journal.pone.0012444 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perugini, L. , Caporaso, L. , Marconi, S. , Cescatti, A. , Quesada, B. , de Noblet‐Ducoudre, N. , House, J. , & Arneth, A. (2017). Biophysical effects on temperature and precipitation due to land cover change. Environmental Research Letters, 12, 10.1088/1748-9326/aa6b3f [DOI] [Google Scholar]
- Peterson, D. A. (2001). Long term gravel pad reclamation on Alaska’s north slope. University of Minnesota, Department of Horticultural Science. http://hdl.handle.net/11299/60127 [Google Scholar]
- Phalan, B. , Onial, M. , Balmford, A. , & Green, R. E. (2011). Reconciling food production and biodiversity conservation: Land sharing and land sparing compared. Science, 333(6047), 1289–1291. 10.1126/science.1208742 [DOI] [PubMed] [Google Scholar]
- Piao, S. (2006). Effect of climate and CO2 changes on the greening of the Northern Hemisphere over the past two decades. Geophysical Research Letters, 33(23), L23402. 10.1029/2006GL028205 [DOI] [Google Scholar]
- Pistorius, T. , Carodenuto, S. , & Wathum, G. (2017). Implementing Forest Landscape Restoration in Ethiopia. Forests, 8(3), 61. 10.3390/f8030061 [DOI] [Google Scholar]
- Pitcher, G. , & Jacinto, G. S. (2019). Ocean deoxygenation links to harmful algal blooms. Ocean deoxygenation: Everyone’s problem (pp. 153–170). IUCN International Union for Conservation of Nature. [Google Scholar]
- Plastrik, P. , & Cleveland, J. (2018). Life after carbon: The next Global Transformation of Cities. Island Press. [Google Scholar]
- Pörtner, H. O. , Scholes, R. J. , Agard, J. , Archer, E. , Arneth, A. , Bai, X. , Barnes, D. , Burrows, M. , Chan, L. , Cheung, W. L. , Diamond, S. , Donatti, C. , Duarte, C. , Eisenhauer, N. , Foden, W. , Gasalla, M. A. , Handa, C. , Hickler, T. , Hoegh‐Guldberg, O. , … Ngo, H. T. (2021). Scientific outcome of the IPBES‐IPCC co‐sponsored workshop on biodiversity and climate change. IPBES Secretariat, Bonn, Germany,, 10.5281/zenodo.4659158 [DOI] [Google Scholar]
- Pretty, J. , Benton, T. G. , Bharucha, Z. P. , Dicks, L. V. , Flora, C. B. , Godfray, H. C. J. , Goulson, D. , Hartley, S. , Lampkin, N. , Morris, C. , Pierzynski, G. , Prasad, P. V. V. , Reganold, J. , Rockström, J. , Smith, P. , Thorne, P. , & Wratten, S. (2018). Global assessment of agricultural system redesign for sustainable intensification. Nature Sustainability, 1(8), 441–446. 10.1038/s41893-018-0114-0 [DOI] [Google Scholar]
- Pugh, T. A. M. , Lindeskog, M. , Smith, B. , Poulter, B. , Arneth, A. , Haverd, V. , & Calle, L. (2019). Role of forest regrowth in global carbon sink dynamics. (p. 201810512). 10.1073/pnas.1810512116.Proceedings of the National Academy of Sciences [DOI] [PMC free article] [PubMed] [Google Scholar]
- Queensland Department of Science, Information Technology and Innovation . (2017). Land cover change in Queensland 2015–16: Statewide Landcover and Trees Study Report (p. 57).
- Rabalais, N. , Cai, W.‐J. , Carstensen, J. , Conley, D. , Fry, B. , Hu, X. , Quiñones‐Rivera, Z. , Rosenberg, R. , Slomp, C. P. , Turner, R. , Voss, M. , Wissel, B. , & Zhang, J. (2014). Eutrophication‐Driven Deoxygenation in the Coastal Ocean. Oceanography (Washington D.C.), 27, 172–183. 10.5670/oceanog.2014.21 [DOI] [Google Scholar]
- Rao, C. S. , Prasad, J. V. N. S. , Choudhari, S. K. , & Singh, A. K. (2020). Mainstreaming climate resilient villages in national programmes towards sustainability of agriculture and environment in India. Climate Change and Environmental Sustainability, 8(2), 116–133. 10.5958/2320-642X.2020.00013.7 [DOI] [Google Scholar]
- Reaka‐Kudla, M. L. (1997) The global biodiversity of coral reefs: a comparison with rain forests. In Reaka‐Kudla M. L., Wilson D. E., & Wilson E. O. (Eds.), Biodiversity II: Understanding and protecting our biological resources (pp. 83–108). Joseph Henry Press, 551 pp. [Google Scholar]
- Redford, K. H. , Huntley, B. J. , Roe, D. , Hammond, T. , Zimsky, M. , Lovejoy, T. E. , da Fonseca, G. A. B. , Rodriguez, C. M. , & Cowling, R. M. (2015). Mainstreaming biodiversity: Conservation for the twenty‐first century. Frontiers in Ecology and Evolution, 3(Article 137), 1–7. [Google Scholar]
- Reisman‐Berman, O. , Keasar, T. , & Tel‐Zur, N. (2019). Native and non‐native species for dryland afforestation: Bridging ecosystem integrity and livelihood support. Annals of Forest Science, 76(4), 114. 10.1007/s13595-019-0903-2 [DOI] [Google Scholar]
- Roberts, C. M. , O’Leary, B. C. , & Hawkins, J. P. (2020). Climate change mitigation and nature conservation both require higher protected area targets. Philosophical Transactions of the Royal Society B: Biological Sciences, 375(1794), 10.1098/rstb.2019.0121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts, C. M. , O’Leary, B. C. , McCauley, D. J. , Cury, P. M. , Duarte, C. M. , Lubchenco, J. , Pauly, D. , Sáenz‐Arroyo, A. , Sumaila, U. R. , Wilson, R. W. , Worm, B. , & Castilla, J. C. (2017). Marine reserves can mitigate and promote adaptation to climate change. Proceedings of the National Academy of Sciences of the United States of America, 114(24), 6167–6175. 10.1073/pnas.1701262114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogers, A. , Aburto‐Oropeza, O. , Appeltans, W. , Assis, J. , Balance, L. T. , Cury, P. , Duarte, C. , Favoretto, F. , Kumagai, J. , Lovelock, C. , Miloslavich, P. , Niamir, A. , Obura, D. , O’Leary, B. C. , Reygondeau, G. , Roberts, C. , Sadovy de Mitcheson, Y. , Sutton, T. , Tittensor, D. , & Velarde, E. (2020). Blue Paper 10: Critical habitats and biodiversity: Inventory, thresholds, and governance. Report prepared for the (Norwegian) Prime Minister’s high level panel on a sustainable ocean economy, 85 pp. https://www.oceanpanel.org/blue‐papers/critical‐habitats‐and‐biodiversity‐inventory‐thresholds‐and‐governance [Google Scholar]
- Roman, J. (2003). Whales before whaling in the North Atlantic. Science, 301(5632), 508–510. 10.1126/science.1084524 [DOI] [PubMed] [Google Scholar]
- Roman, J. , & McCarthy, J. J. (2010). The whale pump: Marine mammals enhance primary productivity in a coastal basin. PLoS One, 5(10), e13255. 10.1371/journal.pone.0013255 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosa, J. C. S. , Sánchez, L. E. , & Morrison‐Saunders, A. (2018). Getting to ‘agreed’ post‐mining land use—An ecosystem services approach. Impact Assessment and Project Appraisal, 36(3), 220–229. 10.1080/14615517.2018.1445175 [DOI] [Google Scholar]
- Ruggeri, G. , Mazzocchi, C. , & Corsi, S. (2020). Drinking biodiversity: A choice experiment on Franciacorta sparkling wines. British Food Journal, 122(8), 2531–2549. 10.1108/BFJ-06-2019-0451 [DOI] [Google Scholar]
- Sala, E. , Mayorga, J. , Bradley, D. , Cabral, R. B. , Atwood, T. B. , Auber, A. , Cheung, W. , Costello, C. , Ferretti, F. , Friedlander, A. M. , Gaines, S. D. , Garilao, C. , Goodell, W. , Halpern, B. S. , Hinson, A. , Kaschner, K. , Kesner‐Reyes, K. , Leprieur, F. , McGowan, J. , … Lubchenco, J. (2021). Protecting the global ocean for biodiversity, food and climate. Nature, 10.1038/s41586-021-03371-z [DOI] [PubMed] [Google Scholar]
- Salles, J. M. , Teillard, F. , & Tichit, M. (2017). Land sparing versus land sharing: an economist’s perspective. Regional Environmental Change, 17, 1455–1465. 10.1007/s10113-017-1142-4 [DOI] [Google Scholar]
- Sanderman, J. , Hengl, T. , Fiske, G. , Solvik, K. , Adame, M. F. , Benson, L. , Bukoski, J. J. , Carnell, P. , Cifuentes‐Jara, M. , Donato, D. , Duncan, C. , Eid, E. M. , Ermgassen, P. , Lewis, C. J. E. , Macreadie, P. I. , Glass, L. , Gress, S. , Jardine, S. L. , Jones, T. G. , … Landis, E. (2018). A global map of mangrove forest soil carbon at 30 m spatial resolution. Environmental Research Letters, 13(5), 55002. 10.1088/1748-9326/aabe1c [DOI] [Google Scholar]
- Sandom, C. J. , Middleton, O. , Lundgren, E. , Rowan, J. , Schowanek, S. D. , Svenning, J. C. , & Faurby, S. (2020). Trophic rewilding presents regionally specific opportunities for mitigating climate change. Philosophical Transactions of the Royal Society B‐Biological Sciences. 10.1098/rstb.2019.0125 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sannigrahi, S. , Pilla, F. , Bas, B. , Basu, A. S. , Zhang, Q. , Wang, Y. , Joshi, P. K. , Chakraborti, S. , Coscieme, L. , Keesstra, S. , Roy, P. S. , & Sutton, P. C. (2020). Identification of conservation priority zones using spatially explicit valued ecosystem services: A case from the Indian Sundarbans. Integrated Environmental Assessment and Management, 16(5), 773–787. 10.1002/ieam.4287 [DOI] [PubMed] [Google Scholar]
- Sannigrahi, S. , Zhang, Q. , Joshi, P. K. , Sutton, P. C. , Keesstra, S. , Roy, P. S. , Pilla, F. , Basu, B. , Wang, Y. , Jha, S. , Paul, S. K. , & Sen, S. (2020). Examining effects of climate change and land use dynamic on biophysical and economic values of ecosystem services of a natural reserve region. Journal of Cleaner Production, 257, 10.1016/j.jclepro.2020.120424. 120424. [DOI] [Google Scholar]
- Saunois, M. , Bousquet, P. , Poulter, B. , Peregon, A. , Ciais, P. , Canadell, J. G. , Dlugokencky, E. J. , Etiope, G. , Bastviken, D. , Houweling, S. , Janssens‐Maenhout, G. , Tubiello, F. N. , Castaldi, S. , Jackson, R. B. , Alexe, M. , Arora, V. K. , Beerling, D. J. , Bergamaschi, P. , Blake, D. R. , … Zhu, Q. (2016). The global methane budget 2000–2012. Earth System Science Data, 8(2), 697–751. 10.5194/essd-8-697-2016 [DOI] [Google Scholar]
- Schädel, C. , Schuu, E. A. G. , Bracho, R. , Elberling, B. , Knoblauch, C. , Lee, H. , Luok, Y. , Shaver, G. R. , & Turetsky, M. R. (2014). Circumpolar assessment of permafrost C quality and its vulnerability over time using long‐term 39 incubation data. Global Change Biology, 20(2), 641–652. 10.1111/gcb.12417 [DOI] [PubMed] [Google Scholar]
- Schaefer, H. , Fletcher, S. E. M. , Veidt, C. , Lassey, K. R. , Brailsford, G. W. , Bromley, T. M. , Dlugokencky, E. J. , Michel, S. E. , Miller, J. B. , Levin, I. , Lowe, D. C. , Martin, R. J. , Vaughn, B. H. , & White, J. W. C. (2016). A 21st‐century shift from fossil‐fuel to biogenic methane emissions indicated by 13CH4. Science, 352(6281), 80–84. 10.1126/science.aad2705 [DOI] [PubMed] [Google Scholar]
- Schimel, D. (2015). Effect of increasing CO2 on the terrestrial carbon cycle. Proceedings of the National Academy of Sciences of the United States of America, 112(2), 436–441. 10.1073/pnas.1407302112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmitz, O. J. , Wilmers, C. C. , Leroux, S. J. , Doughty, C. E. , Atwood, T. B. , Galetti, M. , Davies, A. B. , & Goetz, S. J. (2018). Animals and the zoogeochemistry of the carbon cycle. Science, 362(6419), eaar3213. 10.1126/science.aar3213 [DOI] [PubMed] [Google Scholar]
- Secretariat of the Convention on Biological Diversity . (2005). Connecting biodiversity and climate change mitigation and adaptation: Report of the second ad hoc technical expert group on biodiversity and climate change (Technical Series No. 41, p. 126).
- Secretariat of the Convention on Biological Diversity . (2012). Cities and Biodiversity Outlook. Secretariat of the Convention on Biological Diversity. Montreal, p. 64.
- Secretariat of the Convention on Biological Diversity . (2020). Global biodiversity outlook 5. Montreal.
- Serna‐Chavez, H. M. , Schulp, C. J. E. , van Bodegom, P. M. , Bouten, W. , Verburg, P. H. , & Davidson, M. D. (2014). A quantitative framework for assessing spatial flows of ecosystem services. Ecological Indicators, 39, 24–33. [Google Scholar]
- Shin, Y. J. , Arneth, A. , Chowdhury, R. , Midgley, G. F. , Bukvareva, E. , Heinimann, A. , Horcea‐Milcu, A. I. , Kolb, M. , Leadley, P. , Oberdorff, T. , Pichs Madruga, R. , Rondinini, C. , Saito, O. , Sathyapalan, J. , Boafo, Y. A. , Kindlmann, P. , Yue, T. , Krenova, Z. , & Osano, P. (2019). Plausible futures of nature, its contributions to people and their good quality of life. In IPBES, 2019. Global Assessment on Biodiversity and Ecosystem Services. (p. 264). https://www.ipbes.net/global‐assessment
- Shukla, P. R. , Skea, J. , Slade, R. , van Diemen, R. , Haughey, E. , Malley, J. , Pathak, M. , & Portugal Pereira, J. (2019). Technical summary. In Shukla P. R., Skea J., Calvo Buendia E., Masson‐Delmotte V., Pörtner H.‐O., Roberts D. C., Zhai P., Slade R., Connors S., R. van Diemen , Ferrat M., Haughey E., Luz S., Neogi S., Pathak M., Petzold J., Portugal Pereira J., Vyas P., Huntley E., Kissick K., Belkacemi M., & Malley J. (Eds.), Climate change and land: An IPCC special report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems (pp. 37–74). IPCC. [Google Scholar]
- Siegel, D. A. , DeVries, T. , Doney, S. C. , & Bell, T. (2021). Assessing the sequestration time scales of some ocean‐based carbon dioxide reduction strategies. Environmental Research Letters, in press. 10.1088/1748-9326/ac0be0 [DOI] [Google Scholar]
- Sileshi, G. W. (2016). The magnitude and spatial extent of influence of Faidherbia albida trees on soil properties and primary productivity in drylands. Journal of Arid Environments, 132, 1–14. 10.1016/j.jaridenv.2016.03.002 [DOI] [Google Scholar]
- Sinha, E. , Michalak, A. M. , Calvin, K. V. , & Lawrence, P. J. (2019). Societal decisions about climate mitigation will have dramatic impacts on eutrophication in the 21st century. Nature Communications, 10, 939. 10.1038/s41467-019-08884-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smit, I. P. , Asner, G. P. , Govender, N. , Vaughn, N. R. , & van Wilgen, B. W. (2016). An examination of the potential efficacy of high‐intensity fires for reversing woody encroachment in savannas. Journal of Applied Ecology, 53(5), 1623–1633. [Google Scholar]
- Smith, P. , Calvin, K. , Nkem, J. , Campbell, D. , Cherubini, F. , Grassi, G. , Korotkov, V. , Hoang, A. L. , Lwasa, S. , McElwee, P. , Nkonya, E. , Saigusa, N. , Soussana, J. F. , Taboada, M. A. , Manning, F. C. , Nampanzira, D. , Arias‐Navarro, C. , Vizzarri, M. , House, J. , … Arneth, A. (2020a). Which practices co‐deliver food security, climate change mitigation and adaptation, and combat land degradation and desertification? Global Change Biology, 26(3), 1532–1575. 10.1111/gcb.14878 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith, T. , Beagley, L. , Bull, J. W. , Milner‐Gulland, E. J. , Smith, M. , Vorhies, F. , & Addison, P. F. E. (2020b). Biodiversity means business: Reframing global biodiversity goalsfor the private sector. Conservation Letters, 13, e12690. 10.1111/conl.12690 [DOI] [Google Scholar]
- Soares‐Filho, B. , Moutinho, P. , Nepstad, D. , Anderson, A. , Rodrigues, H. , Garcia, R. , Dietzsch, L. , Merry, F. , Bowman, M. , Hissa, L. , Silvestrini, R. , & Maretti, C. (2010). Role of Brazilian Amazon protected areas in climate change mitigation. Proceedings of the National Academy of Sciences, 107(24), 10821–10826. 10.1073/pnas.0913048107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sondak, C. F. A. , Ang, P. O. Jr , Beardall, J. , Bellgrove, A. , Boo, S. M. , Gerung, G. S. , Hepburn, C. D. , Hong, D. D. , Hu, Z. , Kawai, H. , Largo, D. , Lee, J. A. , Phaik‐Eem Lim, P.‐E. , Mayakun, J. , Nelson, W. A. , Oak, J. H. , Phang, S.‐M. , Sahoo, D. , Peerapornpis, Y. , … Chung, I. K. (2016). Carbon dioxide mitigation potential of seaweed aquaculture beds (SABs). Journal of Applied Phycology, 10.1007/s10811-016-1022-1 [DOI] [Google Scholar]
- Sonter, L. J. , Gordon, A. , Archibald, C. , Simmonds, J. S. , Ward, M. , Metzger, J. P. , Rhodes, J. R. , & Maron, M. (2020). Offsetting impacts of development on biodiversity and ecosystem services. Ambio, 49(4), 892–902. 10.1007/s13280-019-01245-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Souster, T. A. , Barnes, D. K. A. , & Hopkins, J. (2020). Variation in zoobenthic blue carbon in the Arctic’s Barents Sea shelf sediments. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences, 378(20190362). 10.1098/rsta.2019.0362 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spahni, R. , Joos, F. , Stocker, B. D. , Steinacher, M. , & Yu, Z. C. (2013). Transient simulations of the carbon and nitrogen dynamics in northern peatlands: From the Last Glacial Maximum to the 21st century. Climate of the past, 9(3), 1287–1308. 10.5194/cp-9-1287-2013 [DOI] [Google Scholar]
- Spencer, T. , Schuerch, M. , Nicholls, R. J. , Hinkel, J. , Lincke, D. , Vafeidis, A. T. , Reef, R. , McFadden, L. , & Brown, S. (2016). Global coastal wetland change under sea‐level rise and related stresses: The DIVA Wetland Change Model. Global and Planetary Change, 139, 15–30. 10.1016/j.gloplacha.2015.12.018 [DOI] [Google Scholar]
- Stenton‐Dozey, J. M. E. , Heath, P. , Ren, J. S. , & Zamora, L. N. (2020). New Zealand aquaculture industry: Research, opportunities and constraints for integrative multitrophic farming. New Zealand Journal of Marine and Freshwater Research, 10.1080/00288330.2020.1752266 [DOI] [Google Scholar]
- Stevens, N. , Lehmann, C. E. R. , Murphy, B. P. , & Durigan, G. (2017). Savanna woody encroachment is widespread across three continents. Global Change Biology, 23(1), 235–244. 10.1111/gcb.13409 [DOI] [PubMed] [Google Scholar]
- Strassburg, B. B. N. (2010). Global congruence of carbon storage and biodiversity in terrestrial ecosystems. Conservation Letters, 3(2), 98–105. [Google Scholar]
- Strassburg, B. , Beyer, H. , Crouzeilles, R. , Iribarrem, A. , Barros, F. , de Siqueira, M. F. , Sánchez‐Tapia, A. , Balmford, A. , Sansevero, J. B. B. , Brancalion, P. H. S. , Broadbent, E. N. , Chazdon, R. L. , Filho, A. O. , Gardner, T. A. , Gordon, A. , Latawiec, A. , & Loyola, R. (2018). Strategic approaches to restoring ecosystems can triple conservation gains and halve costs. Nature Ecology & Evolution, 1, 64. 10.1038/s41559-018-0743-8 [DOI] [PubMed] [Google Scholar]
- Strassburg, B. B. N. , Iribarrem, A. , Beyer, H. L. , Cordeiro, C. L. , Crouzeilles, R. , Jakovac, C. C. , Braga, J. A. , Lacerda, E. , Latawiec, A. E. , Balmford, A. , Brooks, T. M. , Butchart, S. H. M. , Chazdon, R. L. , Erb, K.‐H. , Brancalion, P. , Buchanan, G. , Cooper, D. , Díaz, S. , Donald, P. F. , … Visconti, P. (2020). Global priority areas for ecosystem restoration. Nature, 10.1038/s41586-020-2784-9 [DOI] [PubMed] [Google Scholar]
- Sumaila, U. R. , Ebrahim, N. , Schuhbauer, A. , Skerritt, D. , Li, Y. , Kim, H. S. , Mallory, T. G. , Lam, V. W. L. , & Pauly, D. (2019). Updated estimates and analysis of global fisheries subsidies. Marine Policy, 109, 10.1016/j.marpol.2019.103695 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Syrbe, R. U. , & Walz, U. (2012). Spatial indicators for the assessment of ecosystem services: Providing, benefiting and connecting areas and landscape metrics. Ecological Indicators, 21, 80–88. [Google Scholar]
- Tamburini, G. , Bommarco, R. , Wanger, T. C. , Kremen, C. , van der Heijden, M. G. A. , Liebman, M. , & Hallin, S. (2020). Agricultural diversification promotes multiple ecosystem services without compromising yield. Science Advances, 6, eaba1715. 10.1126/sciadv.aba1715 [DOI] [PMC free article] [PubMed] [Google Scholar]
- te Beest, M. , Sitters, J. , Ménard, C. B. , & Olofsson, J. (2016). Reindeer grazing increases summer albedo by reducing shrub abundance in Arctic tundra. Environmental Research Letters, 11(12), 10.1088/1748-9326/aa5128. 125013 [DOI] [Google Scholar]
- Tessnow‐von, W. I. , & Vadrot, A. B. M. (2020). The Voice of Science on Marine Biodiversity Negotiations: A Systematic Literature Review. Frontiers in Marine Science, 7, 614282. 10.3389/fmars.2020.614282 [DOI] [Google Scholar]
- Tewari, P. , Singh, R. , Nagarkoti, P. , & Gumber, S. (2020). Temporal changes in livelihood and land usage patterns: Case study of a primitive tribe, Van Raji, from Uttarakhand, India (pp. 213–228). 10.1007/978-981-15-4712-6_13 [DOI] [Google Scholar]
- Thapa, S. , Chitale, V. , Rijal, S. J. , Bisht, N. , & Shrestha, B. B. (2018). Understanding the dynamics in distribution of invasive alien plant species under predicted climate change in Western Himalaya. PLoS One, 13(4), e0195752. 10.1371/journal.pone.0195752 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Theuerkauf, S. N. J. , Barrett, L. T. , Alleway, H. K. , Costa‐Pierce, B. A. , St. Gelais, A. , & Jones, R. C. (2021). Habitat value of bivalve shellfish and seaweed aquaculture for fish and invertebrates: Pathways, synthesis and next steps. Reviews in Aquaculture,. 10.1111/raq.12584 [DOI] [Google Scholar]
- Tian, H. Q. (2020). A comprehensive quantification of global nitrous oxide sources and sinks. Nature, 586(7828), 248. [DOI] [PubMed] [Google Scholar]
- Tittonell, P. , Piñeiro, G. , Garibaldi, L. A. , Dogliotti, S. , Olff, H. , & Jobbagy, E. G. (2020). Agroecology in large scale farming—A research agenda. Frontiers in Sustainable Food Systems, 4, 214. 10.3389/fsufs.2020.584605 [DOI] [Google Scholar]
- Tölle, M. H. , Engler, S. , & Panitz, H.‐J. (2017). Impact of abrupt land cover changes by tropical deforestation on southeast asian climate and agriculture. Journal of Climate, 30(7), 2587–2600. 10.1175/JCLI-D-16-0131.1 [DOI] [Google Scholar]
- Townsend, P. , & Masters, K. (2015). Lattice‐work corridors for climate change: A conceptual framework for biodiversity conservation and social‐ecological resilience in a tropical elevational gradient. Ecology and Society, 20. 10.5751/ES-07324-200201 [DOI] [Google Scholar]
- Trathan, P. N. , Collins, M. A. , Grant, S. M. , Belchier, M. , Barnes, D. K. A. , Brown, J. , & Staniland, I. (2014). The South Georgia and the South Sandwich Islands MPA: Protecting a biodiverse oceanic island chain situated in the flow of the antarctic circumpolar current. ADVANCES IN MARINE BIOLOGY, 69, 15–78. [DOI] [PubMed] [Google Scholar]
- Uddin, K. , Chaudhary, S. , Chettri, N. , Kotru, R. , Murthy, M. , Chaudhary, R. P. , Ning, W. , Shrestha, S. M. , & Gautam, S. K. (2015). The changing land cover and fragmenting forest on the Roof of the World: A case study in Nepal’s Kailash Sacred Landscape. Landscape and Urban Planning, 141, 1–10. 10.1016/j.landurbplan.2015.04.003 [DOI] [Google Scholar]
- Uddin, M. S. , de Ruyter van Steveninck, E. , Stuip, M. , & Shah, M. A. R. (2013). Economic valuation of provisioning and cultural services of a protected mangrove ecosystem: A case study on Sundarbans Reserve Forest, Bangladesh. Ecosystem Services, 5, 88–93. 10.1016/j.ecoser.2013.07.002 [DOI] [Google Scholar]
- UNEP . (2019). Emissions Gap Report 2019. UNEP ‐ UN Environment Programme. http://www.unep.org/resources/emissions‐gap‐report‐2019 [Google Scholar]
- UNEP‐WCMC . (2021). The world database on protected areas (WDPA). https://www.protectedplanet.net/en/thematic‐areas/wdpa?tab=WDPA [Google Scholar]
- UNESCO . (2021). Learn for our planet: A global review of how environmental issues are integrated in education. France. [Google Scholar]
- United Nations Economist Network . (2020). Report of the UN economist network for the UN 75th anniversary: Shaping the trends of our time. United Nations. [Google Scholar]
- United Nations, Department of Economic and Social Affairs, Population Division (UNDESA) (2019). World urbanization prospects: The 2018 revision (ST/ESA/SER.A/420). United Nations, Department of Economic and Social Affairs. https://population.un.org/wup/Publications/Files/WUP2018‐Report.pdf [Google Scholar]
- van Meijl, J. C. M. , Havlík, P. , Lotze‐Campen, H. , Stehfest, H. , Witzke, P. , Bodirsky, B. , van Dijk, M. , Doelman, M. , Humpenoeder, F. , Levin‐Koopman, J. , Mueller, C. , Popp, A. , Tabeau, A. A. , & Valin, H. (2017). In Pérez Domínguez I., & Fellmann T. (Eds.), Challenges of global agriculture in a climate change context by 2050: AgCLIM50. Publications Office. https://data.europa.eu/doi/ 10.2760/772445 [DOI] [Google Scholar]
- van Wilgen, B. W. , Forsyth, G. G. , Le Maitre, D. C. , Wannenburgh, A. , Kotzé, J. D. , van den Berg, E. , & Henderson, L. (2012). An assessment of the effectiveness of a large, national‐scale invasive alien plant control strategy in South Africa. Biological Conservation, 148(1), 28–38. [Google Scholar]
- Vanbergen, A. , Aizen, M. , Cordeau, S. , Garibaldi, L. , Garratt, M. , Kovács‐Hostyánszki, A. , Lecuyer, L. , Ngo, H. , Potts, S. , & Settele, J. (2020). Transformation of agricultural landscapes in the Anthropocene: nature’s contributions to people, agriculture and food security. Advances in Ecological Research, 10.1016/bs.aecr.2020.08.002 [DOI] [Google Scholar]
- Vinya, R. , Syampungani, S. , Kasumu, E. C. , Monde, C. , & Kasubika, R. (2011). Preliminary study on the drivers of deforestation and potential for REDD+ in Zambia. FAO/Zambian Ministry of Lands and Natural Resources. [Google Scholar]
- Vogt, M. A. B. (2020). Developing stronger association between market value of coffee and functional biodiversity. Journal of Environmental Management, 269, 10.1016/j.jenvman.2020.110777. 110777 [DOI] [PubMed] [Google Scholar]
- Wanger, T. C. , DeClerck, F. , Garibaldi, L. A. , Ghazoul, J. , Kleijn, D. , Klein, A.‐M. , Kremen, C. , Mooney, H. , Perfecto, I. , Powell, L. L. , Settele, J. , Solé, M. , Tscharntke, T. , & Weisser, W. (2020). Integrating agroecological production in a robust post‐2020 Global Biodiversity Framework. Nature Ecology & Evolution, 4(9), 1150–1152. 10.1038/s41559-020-1262-y [DOI] [PubMed] [Google Scholar]
- Waycott, M. , Duarte, C. M. , Carruthers, T. J. , Orth, R. J. , Dennison, W. C. , Olyarnik, S. , Calladine, A. , Fourqurean, J. W. , Heck, K. L. , & Hughes, A. R. (2009). Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proceedings of the National Academy of Sciences, 106(30), 12377–12381. 10.1073/pnas.0905620106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weeks, R. , Aliño, P. M. , Atkinson, S. , Beldia, P. , Binson, A. , Campos, W. L. , Djohani, R. , Green, A. L. , Hamilton, R. , & Horigue, V. (2014). Developing marine protected area networks in the Coral Triangle: Good practices for expanding the Coral Triangle Marine Protected Area System. Coastal Management, 42(2), 183–205. 10.1080/08920753.2014.877768 [DOI] [Google Scholar]
- West, T. A. , Börner, J. , & Fearnside, P. M. (2019). Climatic benefits from the 2006–2017 avoided deforestation in Amazonian Brazil. Frontiers in Forests and Global Change, 2, 52. 10.3389/ffgc.2019.00052 [DOI] [Google Scholar]
- Wigley, B. J. , Augustine, D. J. , Coetsee, C. , Ratnam, J. , & Sankaran, M. (2020). Grasses continue to trump trees at soil carbon sequestration following herbivore exclusion in a semiarid African savanna. Ecology, 101(5), 10.1002/ecy.3008 [DOI] [PubMed] [Google Scholar]
- Wilmers, C. C. , Estes, J. A. , Edwards, M. , Laidre, K. L. , & Konar, B. (2012). Do trophic cascades affect the storage and flux of atmospheric carbon? An analysis of sea otters and kelp forests. Frontiers in Ecology and the Environment, 10(8), 409–415. 10.1890/110176 [DOI] [Google Scholar]
- Wolf, J. , Asrar, G. R. , & West, T. O. (2017). Revised methane emissions factors and spatially distributed annual carbon fluxes for global livestock. Carbon Balance and Management, 12(1), 16. 10.1186/s13021-017-0084-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wong, N. H. , Cheong, D. K. W. , Yan, H. , Soh, J. , Ong, C. L. , & Sia, A. (2003). The effects of rooftop garden on energy consumption of a commercial building in Singapore. Energy and Buildings, 35(4), 353–364. 10.1016/S0378-7788(02)00108-1 [DOI] [Google Scholar]
- World Health Organization . (2016). Urban green spaces and health. WHO Regional Office for Europe. [Google Scholar]
- WWF . (2020). In Almond R. E. A., Grooten M. & Petersen T. (Eds.), Living Planet Report 2020—Bending the curve of biodiversity loss (1–159). WWF. [Google Scholar]
- Xi, Y. , Peng, S. , Ciais, P. , & Chen, Y. (2020). Future impacts of climate change on inland Ramsar wetlands. Nature Climate Change, 1–7, 10.1038/s41558-020-00942-2 [DOI] [Google Scholar]
- Xiao, X. , Agustí, S. , Lin, F. , Li, K. , Pan, Y. , Yu, Y. , Zheng, Y. , Wu, J. , & Duarte, C. (2017). Nutrient removal from Chinese coastal waters by large‐scale seaweed aquaculture. Scientific Reports, 7, 46613. 10.1038/srep46613 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiao, X. , Agustí, S. , Pan, Y. , Yu, Y. , Li, K. , Wu, J. , & Duarte, C. M. (2019). Warming amplifies the frequency of harmful algal blooms with eutrophication in Chinese Coastal Waters. Environmental Science and Technology, 53, 13031–13041. [DOI] [PubMed] [Google Scholar]
- Xu, X. M. , Sharma, P. , Shu, S. J. , Lin, T. S. , Ciais, P. , Tubiello, F. N. , Smith, P. , Campbell, N. , & Jain, A. K. (2021). Global greenhouse gas emissions from plant‐ and animal‐based food. Nature Food, 2, 724–732. 10.1038/s43016-021-00358-x [DOI] [PubMed] [Google Scholar]
- Yu, K. (2020). Sponge city: Leveraging nature as ecological infrastructure. CLC Webinar Series: Cities Adapting to a Disrupted World. https://www.clc.gov.sg/events/webinars/view/sponge‐cities
- Yusuf, H. M. , Treydte, A. C. , & Sauerborn, J. (2015). Managing semi‐arid rangelands for carbon storage: Grazing and woody encroachment effects on soil carbon and nitrogen. PLoS One, 10(10), e0109063. 10.1371/journal.pone.0109063 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, G. , Xiao, X. , Dong, J. , Xin, F. , Zhang, Y. , Qin, Y. , Doughty, R. , & Moore, B. (2020). Fingerprint of rice paddies in spatial–temporal dynamics of atmospheric methane concentration in monsoon Asia. Nature Communications, 11, 554. 10.1038/s41467-019-14155-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao, M. , Zhou, Y. , Meng, J. , Zheng, H. , Cai, Y. , Shan, Y. , Guan, D. , & Yang, Z. (2021). Virtual carbon and water flows embodied in global fashion trade: A case study of denim products. Journal of Cleaner Production, 303, 127080. [Google Scholar]
- Zhu, Z. , Piao, S. , Myneni, R. , Huang, M. , Zeng, Z. , Canadell, J. , Ciais, P. , Sitch, S. , Friedlingstein, P. , Arneth, A. , Cao, C. , Cheng, L. , Kato, E. , Koven, C. , Li, Y. , Lian, X. , Liu, Y. , Liu, R. , Mao, J. , & Zeng, N. (2016). Greening of the Earth and its drivers. Nature Climate Change, 6, 10.1038/nclimate3004 [DOI] [Google Scholar]
- Zomer, R. , & Oli, K. P. (2011). Kailash sacred landscape conservation initiative—Feasibility assessment report. ICIMOD. [Google Scholar]
- zu Ermgassen, S. O. S. E. , Baker, J. , Griffiths, R. A. , Strange, N. , Struebig, M. J. , & Bull, J. W. (2019). The ecological outcomes of biodiversity offsets under “no net loss” policies: A global review. Conservation Letters, 12(6), e12664. 10.1111/conl.12664 [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Material
Data Availability Statement
The data that supports the findings of this study are available in the supplementary material of this article.