

ABSTRACT
Swiss‐born embryologist Wilhelm His, Sr. (1831–1904) was the first scientist to study embryos using paraffin histology, serial sectioning and three‐dimensional modelling. With these techniques, His made many important discoveries in vertebrate embryology and developmental neurobiology, earning him two Nobel Prize nominations. He also developed several theories of mechanical and evolutionary developmental biology. His argued that adult form is determined by the differential growth of developmental primordia. Furthermore, he suggested that changes in the growth parameters of those primordia are responsible for generating new phenotypes during evolution. His developed these theories in his book ‘Our Bodily Form’ (Unsere Körperform). Here, we review His's work with special emphasis on its potential importance to the disciplines of evolutionary developmental biology (evo‐devo) and mechanobiology.
Keywords: Wilhelm His, mechanobiology, evolution and development, evo‐devo, gastrulation, germ layer, morphogenesis, allometry, Entwicklungsmechanik
I. INTRODUCTION
With the recent death of Leipzig anatomist Wilhelm His [Sr.], we have seen the burial of one of the most idiosyncratic of research personalities. Certainly not a trail‐blazer like Carl Gegenbaur, or a fighter like his great opponent Ernst Haeckel, but a quiet scholar who worked with real German thoroughness… and immersed himself with real German sensibility in the details of phenomena … nonetheless, none of the other outstanding anatomists of the nineteenth century was treated with such hostility – almost hatred – as Wilhelm His. (Rawitz, 1904, p. 308)
Wilhelm His, Sr. (1831–1904) (Figs 1 and 2; Supplementary Note S1) was a Swiss‐born physician who pursued a career in embryological and anatomical research in Basel and then Leipzig. Landmarks in his career are given as online Supporting Information in Table S1. His published on a wide range of scientific topics (Fig. S2). For example, he showed that the lymphatic system is a closed system (see Table S2 for a glossary of terms); and he produced an early example of forensic craniofacial reconstruction, using the supposed skeletal remains of Johan Sebastian Bach (Supplementary Note S2). He also made important contributions to vertebrate embryology and developmental neurobiology, including neuron theory (Supplementary Note S3). His's success in anatomical research was due in part to his ability to understand the complex three‐dimensional (3‐D) relationships of structures, and how they change during development (Fig. 3).
Fig. 1.

Portraits of Wilhelm His. (A) 1849, portrait, aged 18, on the bowl of a pipe, from a watercolour by Hans Burckhardt; Anatomisches Museum, Basel. Photograph M. K. Richardson. (B) Photograph, 1872; courtesy of Roger and Hugo Kurz. (C) Photograph; courtesy of Fotosammlung 11, Universitätsbibliothek, Leipzig. An identical image in the Alan Mason Chesney Medical Archives is inscribed: ‘G. Brokesch 1882–83 Leipzig’. (D) Painting (oil on canvas), 62 × 49 cm, signed Albert Winther, dated 189?; Kunstbesitz der Universität Leipzig, inventory no. 1951:004, photographed by Karin Kranich; image rights: Kustodie der Universität Leipzig (for more information, see Supplementary Note S1). (E) Undated photograph, courtesy of the History of Medicine Collections, David M. Rubenstein Rare Book and Manuscript Library, Duke University. (F) Photograph, circa 1900 (source as B). B and F are figs 3 and 6 in Kurz (1992) from where the information provided here was taken.
Fig. 2.

Wilhelm His and other delegates at meetings (1895 and 1897) of the Anatomische Gesellschaft. (A) Basel (17–20 April, 1895), the meeting from which the ‘Basel’ Nomina Anatomica took its name (His, 1895b). Wilhelm His: seated, front row, far right. (B) Tübingen, 21–24 May, 1899. Wilhelm His: middle of second row, with round‐framed glasses. Images courtesy the Archive of the Anatomische Gesellschaft. For key to sitters see Fig. S1.
Fig. 3.

Scientific illustrations by Wilhelm His. (A) Fig. B1 from Plate VII in His (1880). Reconstruction of a human embryo from serial sections. The corresponding section numbers are shown by the scale (right). Note the semi‐transparent rendering of the liver (Lb) to reveal the intrahepatic course of the inferior vena cava. A note printed at the bottom of the plate attributes the drawings to His. (B) Figs 78 and 79, p. 130 in His (1885). Note the use of cut‐away layers to expose the endocardial tube. (C) Pen‐sketch by His, in a letter dated 20th March, 1889 to his former student Franklin Mall. His refers to a discussion between them over the developmental origin of the thymus. From the Mall Papers (Wilhelm His), image courtesy of the Alan Mason Chesney Medical Archives of the Johns Hopkins Medical Institutions. (D) Blackboard drawing made by His during his lecture to a meeting of the Anatomical Society of Great Britain and Ireland at the Royal Dublin Society on June 11th, 1897. The drawing was photographed and reproduced as fig. 1 in His (1897a). Image courtesy the Wellcome Collection.
In recent years, several historians and biologists have discussed His's research (e.g. Gould, 1977; Maienschein, 1991; Nyhart, 1995; Hopwood, 1999, 2000, 2002, 2005, 2007, 2012, 2013; Richardson & Keuck, 2001; Hoßfeld & Olsson, 2003; Brauckmann, 2006; Laubichler, Aird & Maienschein, 2007; Richards, 2008; Dupont, 2017; Glover et al., 2018). His is well‐known for his ‘mechanical’ theories of development. He wrote: ‘Embryology and morphology cannot proceed independently of all reference to the general laws of matter, – to the laws of physics and of mechanics’ (His, 1888a, p. 293). Our aims in this review are to explore these mechanical theories; to examine how His applied these theories to evolutionary questions; and to consider the potential relevance of His's theories to the modern disciplines of evolutionary developmental biology (evo‐devo) and mechanobiology.
We start by examining His's early research and show how it formed the basis of theories that he would advocate for most of his career. Then, we consider his (1875) book ‘Our Bodily Form’, which contained new hypotheses about vertebrate developmental mechanisms and their relationship to evolution. Finally, we examine how His's theories have been received by scientists and examine them in the light of current scientific knowledge. We should note that His's son, Wilhelm His Jr. was also a physician and scientist; he described the atrioventricular bundle in the heart (Roguin, 2006; Anderson & Mori, 2016).
II. WILHELM HIS'S EARLY RESEARCH
As a medical student, His spent three semesters at Berlin University. During that time, he and other students were invited by Robert Remak, one of the lecturers, to his home on the Unter den Linden. There, Remak had created a makeshift embryology laboratory, and he showed the students how to prepare chicken embryos for microscopical analysis (His, 1868b, p. 50; His, 1903, p. 26). These interactions with Remak stimulated in His an enduring interest in early chicken development. Remak stressed the importance of understanding the germ layers in relation to histogenesis: the development of mature tissues from precursor cells (Table S2). His took up these issues in a paper entitled ‘The membranes and cavities of the body’ (His, 1865b).
(1). Membranes and cavities
During the nineteenth century we see three great steps in anatomy: general anatomy, associated with the name of Bichat, the cell doctrine with that of Schwann, and histogenesis with that of His. (Mall, 1905, pp. 144–145).
In his paper ‘Membranes and cavities’, His coined the name ‘endothelia’ for epithelia that arise from the mesoderm, and which come to line body cavities, and the blood and lymphatic vessels (His, 1865b, p. 18). This paper contains highly original speculations on developmental mechanisms. Those speculations appear to be informed by his unpublished histological studies of chick embryos. His noted that embryonic cavities develop from splits in the mesoderm, and suggested that this splitting might be mediated by mechanical forces. When a split develops, he argued, it be becomes filled with fluid. This leads to swelling of the extracellular matrix, and the resulting pressure is modulated by elastic fibres in the matrix. Further, His argued that mechanical forces in the matrix might actually influence cell behaviour:
The remarks above concern an influence of mechanical moments on the development of connective tissue, which we can call directive [richtenden]; in this case, an indifferent tissue mass is assumed, and the special way of developing that it experiences under given external influences is followed. Now, mechanical influences, along with everything else that comes under the broad concept of stimuli, can act in a determinant [bestimmend] way upon cell proliferation and thereby on the formation of tissues. (His, 1865b, p. 30, his italics; the words in square brackets are those used in the German original)
His also speculated that ectoderm and endoderm might provide a chemical signal to the mesoderm which stimulates growth and blood vessel development (His, 1865b, p. 33).
His challenged Remak's view that the peripheral nervous system develops from the mesoderm, arguing (in agreement with data available today) that it develops from the ectoderm (Fig. 4; Table S3; Remak, 1855, p. 44; His, 1865b; pp. 7–9). His also disputed Remak's view that the nephric duct arises from the mesoderm, arguing for an ectodermal origin (the data available today are consistent with Remak's view). His later changed his mind when he realised that what he had taken to be the nephric precursor was actually the primordium of the peripheral nervous system. With this primordium, His had discovered the neural crest (a term later coined by Marshall; Fig. 4; Tables S2 and S3). His (1865b, p. 8) suggested that such disagreements about the origins of tissues arose because of the limited resolution of the histological techniques of the time.
Fig. 4.

Conflicting hypotheses of the origin of the neural crest and kidney duct. (A) Plate V, fig. 69 in Remak (1855). Transverse slice (possibly from a fresh embryo) through the trunk of a chicken embryo viewed with transmitted light. Individual cells are not shown; the tissue is simply block‐shaded, with selected mesodermal derivatives shown in a paler shade. (B) Fig. IVa, p. 32 in Hensen (1903) showing a transverse section with what Hensen believed to be the precursor of the nephric duct arising from a cell mass in the ectoderm; the mass grows ventrad towards the mesoderm and then becomes detached (our boxes a and b). (C) Plate XI (III) in His (1865a) A–D (from bottom to top) are sections from the same chicken embryo showing what His thought was the precursor of the nephric duct (Urnierenfalte, Unf.). (D) Plate VI, fig. III (4) in His (1868b). Transvere section through a chicken embryo showing the intermediate groove (Zr), the supposed precursor of the peripheral ganglia. (E) Plate VII, box I, figs 2, 4 in (His, 1868b). Our box a shows detail of intermediate cord (Z); our box b shows precursors of the ganglia as an inverted pyramid of tissue (arrowhead). Note that His's figures show individual cells, a reflection of his improved histological techniques.
(2). Improved histological techniques
Like my predecessors, I had little success in achieving a uniform view of the genesis of the nervous system by way of observation, and I could reach only indirect conclusions about the developmental history of ganglia. The obstacle to decisive observations seemed to lie in deficient technology, and this has kept me working to improve the technique of making sections as fine as possible. (His, 1868b, p. v)
A lack of adequate histological techniques was recognised as an obstacle to progress in embryology (Kölliker, 1876, pp. 102–103; Bischoff, 1877, pp. 2–6; Balfour, 1881, pp. 124–125; Hertwig, 1906, p. 2). The main difficulty lay in obtaining sufficiently thin sections. For most purposes, histologists cut tissue slices by hand with a razor (e.g. His, 1856, p. 3), but young embryos are too delicate to be sectioned in this way. Instead, they were dissected, while living, in a dish of water or saline (Remak, 1855, p. xxxvi; Dursy, 1866, pp. 11–14), essentially applying the dissection techniques of gross anatomy to embryos (Hertwig, 1906, p. 2). With such crude techniques it was impossible to resolve the details of early development, or to adequately employ the 3‐D reconstruction techniques that His had invented (His, 1887, pp. 383–384; Hopwood, 1999, 2000, 2002, 2012); those techniques required an unbroken series of sections of consistent thickness.
His addressed these problems by developing a protocol for the infiltration‐embedding of embryos with paraffin wax (van der Lem et al., 2021). Edwin Klebs had introduced paraffin wax to histology (Klebs, 1867) but soon abandoned it (Klebs, 1869, pp. 164–165) because wax does not penetrate watery tissue. His overcame this problem by dehydrating the embryos in alcohol and clearing them in oil of lavender, which is miscible with both alcohol and molten wax, and therefore acts as an intermediate reagent.
The sectioning of paraffin‐embedded embryos by hand was very time consuming (His, 1870a, p. 229). There were several mechanical aids to sectioning then available such as Valentin's ‘double‐knife’ (Fig. 5A, B), and Hensen's ‘section‐cutter’ (Fig. 5C, D; Hensen, 1863, p. 82; His, 1887, p. 383). His designed his own microtome (Fig. 6) for the free‐hand sectioning of paraffin‐embedded embryos. This microtome allowed sections to be cut individually (rather than in a ribbon; His, 1868b, p. 181) at His's preferred section thickness of 50 μm (His, 1887, p. 383). His's new workflow of infiltration embedding, sectioning and 3‐D modelling formed the basis of His's studies on the early embryology of vertebrates.
Fig. 5.

Early microtechniques before the introduction of His's microtome. (A, B) Valentin double‐knife made by Weiss, London, c. 1860. Photograph: Science and Society Picture Library, Science Museum, London (object number A135073), Wellcome Trust loan. (B) Detail of blades. Oschatz (1844, p. 129) stated that Valentin used a design of Purkyně's, but Valentin denied this (Valentin, 1845, p. 55 n.). (C, D) Hensen's ‘section‐cutter’; Key: b, blade; h, handle of chopper; l, lens; m, sub‐stage mirror; s, stage. The device was made by the instrument maker Beckmann (Hensen, 1863, p. 82 n.1). Anatomisches Museum, Basel; Photograph: M. K. Richardson. (E) Thick tissue slice from a chicken embryo, possibly cut underwater from a fresh embryo. From fig. 1 on p. 5 of Reichert (1861), cropped. Image courtesy of the University of Leiden, Special Collections Library.
Fig. 6.

The microtome of Wilhelm His. (A) Front view; (B) side view of stage; (C) engraved name ‘His’ on the strut. Background gridline spacing: 1 cm. The blade, held in the blade‐holder h (fastened with wire for display purposes) is drawn downwards, the holder steadied against the flat face of g. Between cuts, the block clamped in b is advanced under the arch of g using the micrometer screw (m). Anatomisches Museum, Basel; photograph: M. K. Richardson.
(3). Studies of early chicken development
In the 1860s and early 1870s, His published research on the early embryology of sharks, teleosts and the chicken (see bibliography in Fick, 1904). Among these works was a monograph on the early development of the chicken (His, 1868b) that established His's name as an embryologist (Marchand, 1905, pp. 329–330).
(a). The growth law and organ‐forming regions
The entire development of the organism can be derived from a growth law which originates as a relatively simple function of space and time. (His, 1868b, p. 220)
His's chicken embryology monograph (His, 1868b) was partly descriptive, partly theoretical. The theoretical part was a model of morphogenesis based on the differential growth of embryonic tissues. He wrote: ‘The first step in any attempt to deduce a physical form must therefore be the search for [a] basic law of growth’ (His, 1868b, p. 184). The values of the embryonic growth parameters, he says, are unique to each species and change during evolution. In our view, His's growth law was the first comprehensive model of morphogenesis and morphological evolution with a testable mechanism.
His measured the dimensions of chicken embryos at different stages of development. Using tissue thickness as a proxy for growth rate, he found gradients of growth rate in the blastoderm along all three anatomical axes (His, 1868b, pp. 188–190). His mapped the gradients onto the embryo using a system of 3‐D Cartesian coordinates (His, 1867a, pp. 620–621).
The causes of tissue growth, according to His, are cell proliferation, increase in cell size, cell movements where cells slide past one another (His, 1867a, p. 618; His, 1868b, p. 53), and changes in cell shape (His, 1868b, p. 53). The resultant increase in the volume of the tissues creates mechanical pressures which deform the tissues in accordance with their inherent elasticity (His, 1868b, pp. 52–53). His further argued that differential growth causes some regions of the blastoderm to become thicker — and therefore less elastic — than others. As a result, tissue elasticity becomes unequally distributed. Ultimately, the pressure of differential growth on an unevenly elastic embryo creates tensile and compressive forces that cause the blastoderm to fold.
His used the growth law to explain phenomena such as gastrulation and other morphogenetic movements (His, 1867a, p. 621). It also provided a possible mechanism for regional specification. His argued that the folding of the blastoderm divides it into territories with different developmental fates (His, 1868b, p. 197). These territories or ‘organ‐forming embryonic regions’ (His, 1875, p. 19) are similar to von Baer's ‘primitive organs’ (epithelial tubes, etc.; von Baer, 1837, p. 65), except that they are completely undifferentiated.
(b). The modelling of mechanical forces in development
His used clay, metal, rubber and other materials to model development (Table S4; His, 1867a, p. 623; His, 1868b, p. 182; His, 1894; Hopwood, 1999, 2002). He was also interested in the geological literature on tangential pressures formed in the Earth's crust as it cooled, and how these might cause the land to rise up as mountain ranges. His thought, by analogy, that pressure established by differential growth in the embryo might cause the folding and splitting of tissue layers [Fig. 7; His, 1894; Hopwood, 1999, p. 470 note (n.) 16]. A similar analogy was later drawn by (Roux, 1895a, pp. 6–7).
Fig. 7.

Tensile and compressive forces in development. (A) Fig. 44 in His (1875) showing the formation of a split in the ventral germ layer (Dd) of a bilaminar chicken embryo, resulting from the lateral tension produced by the faster growth of the upper layer (pp. 58–59). The loose cells torn from the lower layer will become the lower leaf of the mesoderm. oG, epiblast; Z, cell stretched across the gap. (B) Fig. 10 in His (1894). Sheet of clay with a sudden transition (arrow) from thick (8 mm) to thin (4 mm). In the lower illustration there is a span of 15 cm, but in the upper illustration the sheet has been pushed from the left to decrease the span to 12 cm. The result is an asymmetric fold resembling the head fold in C. (C) Fig. 26 in His (1894), ray (Torpedo sp.) embryo showing the head fold. Original rotated 90° anticlockwise and horizontally reflected by us. (D) Fig. 17 in His (1894; also reproduced as fig. 3 in Hopwood, 1999). A strip of leather, forced into a curve and fixed along one side. The resultant forces throw the leather into folds resembling intersomitic clefts.
In addition to modelling with physical materials, His asked Eduard Hagenbach to model development mathematically. A theoretical, circular elastic plate, representing the blastodisc, was transformed by specified growth parameters (His, 1868b, pp. 191–194). According to Hagenbach's calculations, the circular plate would be transformed into an ellipse (Brauckmann, 2006, p. 401; Hopwood, 2015, pp. 97–98).
(4). The growth law in conflict with the biogenetic law
His did not incorporate Darwinian theory into his models of development and evolution. He appreciated the explanatory potential of Darwin's ideas, but felt that they were unproven (His, 1868b, pp. 223–224). Ultimately, His thought that Darwinian theory was irrelevant: all that was needed to explain morphological evolution were the mechanics of tissue growth, and species‐differences in the growth law (His, 1868b, pp. 211–213). His's agnosticism towards Darwinian theory is in contrast with the pro‐Darwinian stance of Ernst Haeckel.
Haeckel shared His's desire to explain development on ‘physiological’ or physico‐chemical grounds [Haeckel, 1866, volume (vol.) 1, p. 107; Haeckel, 1866, vol. 2, p. 295]. But while His believed that the embryo contains all causal explanations for its own development, Haeckel believed that the causal mechanisms of development – adaptation and heredity – lay outside the egg, in its phylogenetic history (Haeckel, 1866, pp. 50–60; Richards, 2008, pp. 282–286; Abzhanov, 2013; Olsson, Levit & Hossfeld, 2017).
His and Haeckel's scientific differences quickly became acrimonious (Supplementary Note S4; Spitzer, 1886; Rádl, 1909; Maienschein, 1991; Nyhart, 1995, pp. 188–190; Richards, 2008, pp. 280–291; Hopwood, 2015, pp. 123–126; see Supplementary Note S5 for His's clashes with other scientists). It is likely that his angry exchanges with Haeckel prompted His to write his book, ‘Our Bodily Form’, as a riposte (Richards, 2008, p. 298). Indeed, one advertisement for ‘Our Bodily Form’ specifically recommends it to owners of Haeckel's ‘Anthropogeny’ (Fig. 8A). Furthermore, His wrote his book in just a few weeks (His, 1931, p. 25), suggesting perhaps that he was anxious to give a quick reply to Haeckel's most recent attacks (Supplementary Note S4). ‘Our Bodily Form’ contains scientific arguments against Haeckel's phylogenetic approach, and personal attacks on Haeckel's scientific credibility. It also contains important new ideas about development and evolution.
Fig. 8.

(A) Advertisement for ‘Our Bodily Form’ in Gegenwart, Issue 13, March 27th, 1875, p. 208. (B) Figure on p. 45 of His (1868b) showing lines of folding that subdivide the embryo into organ‐forming regions. (C) Fig. 24 from His (1875). Fate map showing organ‐forming regions demarcated by folding of the embryo. Key: Bauch, abdomen; Brust, ventral thorax; Damm, perineum; Hals, ventral region of neck; Hinterkopf, occipital region; Kreuz, sacral spine; Mundbucht, stomodaeum; Nacken, cervical spine; Oberkiefer, maxilla; Rücken, thoracic spine; Scheitel, vertex; Schwanz, coccygeal region; Stirn, forehead; Unterkiefer, mandible.
III. ‘OUR BODILY FORM’ (UNSERE KÖRPERFORM)
Finally, prompted by my ‘Natural History of Creation’ and ‘Anthropogeny’, His gives a general exposition of the same in his 1875 publication ‘Our Bodily Form’, the most important part of which is particularly directed against me.” (Haeckel, 1875, p. 13)
His's semi‐popular book ‘Our Bodily Form’ (His, 1875; often mis‐cited as 1874; see Table S5) is of particular interest because of its many new theories about evolution and development. Its full title can be translated as: ‘Our Bodily Form and the Physiological Problem of its Origin: Letters to a Natural Scientist Friend’. A translation of the Table of Contents is provided in Table S5. For other discussions of ‘Our Bodily Form’ see Maienschein (1978, pp. 132–134), Maienschein (1981, pp. 95–96), Gould (1997), Hopwood (2015), Picken (1956) and Richards (2008). The ‘natural scientist friend’ alluded to in the title is His's nephew Friedrich Miescher (His, 1897b, p. 18), who incidentally discovered DNA (in pus that he collected from used surgical dressings; Miescher, 1871, pp. 9, 441; His, 1897b; Dahm, 2005).
One of His's stated goals in publishing ‘Our Bodily Form’ was to transform embryology into a new discipline of ‘physiological embryology’: a quantitative science with a reductionist, mechanistic approach (His, 1903, p. 35; Mall, 1905, p. 142). He was inspired by Carl Ludwig's transformation of physiology into a ‘new physiology’ that aimed to explain biological phenome in terms of the underlying chemistry and physics, and to do so using the research methodology of the exact sciences: rigorously planned experiments amenable to mathematical analysis (Ludwig, 1852, 1856; His, 1895c; His, 1898; Rádl, 1909, pp. 549–554; Drischel, 1965; Frank & Weiss, 1966; Rosen, 1973; Fye, 1986; Zimmer, 1996).
(1). Folding and elasticity of the chicken embryo
According to His, the early chicken embryo becomes demarcated by longitudinal and transverse folds in predictable locations (Fig. 8B, C). One of these folds is the Wolffian ridge, named by His in honour of its discoverer Caspar Wolff (His, 1868b, p. 88; Stephens, 1982). His argued that limb buds arise at the points where the Wolffian ridge intersects a transverse fold. To help his reader understand these folds, His suggested that they try folding a piece of paper into the form of a postal envelope, each corner representing the site of a limb bud (Table S4).
The folding is influenced by the elasticity that, according to His, is an intrinsic property of the chicken germinal disc (and the teleost germinal vesicle; His, 1873b, p. 36). His's evidence for that elasticity is the histological appearance of an elastic extracellular matrix in the embryo (His, 1865b, p. 33). In his ‘Monograph on Calcareous Sponges’, Haeckel (1872, p. 472 n.) denied this elasticity: ‘the germinal disc … is not elastic!’ and criticised His for his mechanical approach to evolution and development. In ‘Our Bodily Form’, His responded by noting that Haeckel provided no evidence for his denial of elasticity, and suggested resolving the issue by taking a germinal disc from an incubated chicken egg and placing it in a dish of serum. The disc, when prodded with a probe, will show its elasticity by springing back into its original shape (His, 1875, pp. 48–49). In any case, Haeckel considered his own phylogenetic approach (as embodied in the biogenetic law), and His's mechanical approach, to be irreconcilable.
(2). Organ‐forming regions and a fate map
His illustrated a series of ‘organ‐forming regions’ using what we might now call a fate map (Fig. 8B, C). Today, fate maps are constructed using cell marking and other techniques to track the behaviour of cells and their progeny during development (Le Douarin & Kalcheim, 1999). By contrast, His's fate map is a thought‐experiment in which the definitive body regions are projected, hypothetically, onto the early embryo. His suggested imagining the body of a bird or mammal opened along the ventral midline and flattened out:
If you have carried out in your mind this flattening of the body, it will be clear that on the one hand every point on the embryonic region of the blastoderm must correspond to a later organ, or part of an organ, and that on the other hand any organ that develops from the blastoderm must have a preformed primordium in some spatially defined region of the blastoderm. (His, 1875, p. 19)
His later published a theoretical fate map of the neural primordium (fig. 8 in His, 1893a).
(3). Differential growth in morphogenesis and evolution
His presented measurements from the embryos of various vertebrate species (His, 1875, pp. 208–210) showing that the early embryo is similar in dimensions in the species examined. But, as the embryos become folded into organ‐forming regions, different quantities of tissue are allocated to each region, and the quantity allocated varies among species. His argued that each primordium has its own ‘partial growth law’, and so some regions grow faster than others (His, 1875, pp. 82–83, 152). The brain primordium, for example, has more tissue allocated to it than other regions and grows faster.
The configuration which the organ finally assumes is therefore dependent on the law of its own growth, on its spatial relations with neighbouring parts, and on the growth of the latter. The principle of unequal growth, according to what has been said, also retains its significance as a form‐determining [morphogenetic] principle in the further course of development. (His, 1875, p. 83)
As an example, he modelled the role of unequal growth in the formation of intersomitic clefts using a strip of leather fixed along one side. When deformed into a curve, the leather assumes a series of segment‐like folds (Fig. 7D; His, 1875, pp. 64–65).
(4). Germ layers and the parablast theory
In 1875, the germ layers were a battleground for diverse opinions (His, 1875, p. 38; Hertwig & Hertwig, 1881, p. 117), mainly because the available histological techniques made it difficult to resolve the germ layers in detail. His's improved histological techniques led him to a new model of germ layer development: the parablast theory (His, 1866a,b, 1868b; His, 1875, pp. 41–44). This theory envisaged a dual genetic origin for the embryo, with the mesoderm largely arising from maternal ‘parablast’ cells that migrate from the ovary into the embryo via the white yolk (the pale‐coloured yolk surrounding the embryo). The parablast theory has some similarities with Reichert's idea that the embryo develops from yolk cells (the ‘formative yolk’; Reichert, 1840, pp. 88–96; Reichert, 1843, pp. 23–27; Stricker, 1872, p. 1201).
(5). Concrescence theory
Thus the primitive rudiment of the body is a flat ring, whose width and thickness is maximal at the future head end, and minimal at the opposite end, the tail. The two lateral halves of the ring come successively into apposition and unite as symmetrical body halves. (His, 1875, p. 198)
Another of His's new ideas – concrescence – was inspired by his studies of teleost development (His, 1874; His, 1875, pp. 188–189; His, 1877b). His suggested that the trunk was formed from left and right embryonic halves that fuse in the midline (Fig. 9). This process of fusion supposedly takes place during epiboly, and became known as ‘concrescence’ (Fig. 9; reviewed by Kopsch, 1904). In one thought experiment, His imagined concrescence as if a ring of rubber tubing were pushed inwards at one point; the resulting two‐layered indentation represents the nascent embryo and the converging halves of the germ ring (His, 1877b, p. 109, n.). A similar idea was perhaps touched on by Lereboullet when he described malformed pike (Esox sp.) embryos with a partial split along the longitudinal axis (Lereboullet, 1863; Morgan, 1895, p. 419; Kopsch, 1904, p. 6); it was possible that the split represented the fusion line of two halves.
Fig. 9.

Concrescence in salmon and cat shark embryos. (A) Figs 127–130 in His (1875). This sophisticated schematic uses semi‐transparent rendering to show the marginal ring progressing over the yolk mass during epiboly. The sequence of stages is: top left, top right, bottom left and bottom right. (B) Fig. 126 in His (1875). Schematic illustration of concrescence in bony fish development. Cells from the inner curvature of the germ ring (upper two arrows) contribute to lateral parts of the body; cells from the outer curvature (lower four arrows) contribute to structures closer to the midline. According to the concrescence model, the trunk region is formed by fusion of right and left halves, while the head and tail each consist of one mass from the outset.
(6). Quantitative comparative embryology and a developmental hourglass
(a). His's ‘physiognomy’ of embryos
An identity in the external form of animal embryos, as so often asserted, does not exist. Even at an early stage of development, the embryos have their class and their order characters, just as we can scarcely doubt their species and gender, even their individual characters. (His, 1875, p. 201)
As part of his attack on the biogenetic law, His tried to disprove Haeckel's claim that embryos of different vertebrate species are identical in morphology during organogenetic stages (reviewed by Richardson & Keuck, 2002). His compared six amniote embryos at limb‐bud stages (Figs 10, 11, 12) and quantified their differences using a simple morphometric analysis: he traced the outline of each embryo onto paper, cut round the outlines, then weighed the cut‐outs to estimate their surface area. His analysis showed, for example, that the head of the human embryo is massive, and its body small, compared to the pig embryo. His noted the massive development of the eye in the chicken embryo, suggesting that this compresses the surrounding tissue, causing the beak to develop (Fig. 13).
Fig. 10.

Embryos of six amniotes. From His (1875), limb‐bud stages, left lateral aspect. See a similar composite in Hopwood (2015; fig. 7.14); as Hopwood notes, the embryos were originally on different pages. (A) Fig. 132, human; (B) Fig. 133, pig, arrow indicates the nascent snout described by His; (C) Fig. 134, deer; (D) Fig. 135, rabbit, 14 days (d) post‐fertilisation; (E) Fig. 136, guinea‐pig; (F) Fig. 137, chicken, 5 d post‐fertilisation.
Fig. 11.

Schematic analysis of some of the six amniote embryos shown in Fig. 10, showing our interpretation of His's (1875, pp. 192–206) text. Double arrowheads indicate the images to be compared in each example. (A, B) Pairwise comparison illustrating that human (A) and pig (B) embryos share many common characters (highlighted in green): (cf, cervical flexure; fl, forelimb; hl, hindlimb; hy, hyoid hyomandibular groove; na, nasal pit; nl, nasolachrymal groove; op, optic Anlage; ot, otocyst; so, somites; tb, tailbud. (C, D) Comparison of human (C) and pig (D) showing that common characters present different character states (highlighted in purple). Thus the simple first pharangeal arch (I) in D is divided into mandibular (md) and maxillary (mx) processes in C. His (1875, p. 196) suggested that the modest development of the human tailbud (tb) is a foreshadowing of the vestigial state of the adult coccyx. (E, F) Comparison of human (E) and pig (F) embryos showing relatively massive development of the head, rostral to the eye (shaded blue) in the human; and the relatively massive size of the trunk overlying the pleuro‐peritoneal cavity in the pig (grey). (G, H) The human embryo (G) has a larger telencephalon (te, shaded blue) than the pig (H). The nasal processes (red outline) give the nasal margin (nm) the appearance of a rudimentary snout. The hyoid arch (hy, brown) of the pig embryo is ‘plumper’ (His, 1875, p. 195) than that of the human. (I, J, K) The eye (op) and midbrain (me) together occupy two‐thirds of the area of the head in the chick (K) but less in the human (I) and pig (J). Yellow shading in the two mammals is mainly the lens, whereas in the chicken a large part of the optic cup in addition to the lens is also exposed and shaded. The optic Anlage is much larger in the chicken than in the mammals.
Fig. 12.

Chart summarising His's morphometric analysis of six amniote embryos. Measurements taken from table in His (1875, p. 201). His notes that the chicken and human embryos have relatively large heads (blue), while the deer and pig both have relatively large pleuroperitoneal regions (pink). The dorsal trunk and limbs combined (green) are similar in relative size in all six species.
Fig. 13.

Allometric growth and the development of the beak. Our interpretation of figures of the rostral aspect of the face and text in His (1875, pp. 204–206). (A, B) Fig. 139, 14 day (d) rabbit; (C, D) Fig. 139, 5 d chicken; (E, F) Fig. 140, 6 d chicken. According to His, the eye (yellow) of the chicken embryo grows enormously in size compared with that of the rabbit. As a consequence, the stomodeum (blue) and internasal region in the chicken are laterally compressed (arrows in D), causing the nasal pits (red) to converge (arrows in F). His argued that these changes contribute to the formation of a narrow, protruding beak. Key: nl, nasolachrymal duct.
(b). A developmental hourglass
His compared embryos of different species stage‐by‐stage and showed that early embryos exhibit species differences in the size and distribution of the yolk and other factors:
… the most pressing question is how, from such different developmental starting points, do the similarities in subsequent [stages], and in the adult body, arise? (His, 1875, pp. 190–191)
His explained that, after these divergent early stages, the embryo undergoes folding into organ‐forming regions; the folding releases mechanical tensions in the tissues and so the embryo reaches an equilibrium state (His, 1875, p. 208). The embryos of different species become more similar in appearance, but nonetheless show the unique ‘physiognomy’ of their species. As development progresses further, the morphology of the embryos becomes increasingly divergent as they develop class characters (for example, feathers and beak in the chicken; His, 1875, p. 126). With this analysis, he described a pattern known today as the ‘developmental hourglass’ (Duboule, 1994), although His did not use that term.
(7). Early brain development
Differential growth had been suggested as a causal factor in brain development by Reichert (1861) and later by His (1873a). In ‘Our Bodily Form’, His pursued this topic by analysing neural tube development in different vertebrates (His, 1875, pp. 93–118). His showed that rubber tubes, variously manipulated, can adopt shapes resembling the neural tube at different stages of development (Figs 14 and 15). The forms produced by the tubes bear a striking resemblance to the developing optic primordia, the rhomboid sinus, the infundibulum and the pontine flexure, implying that mechanical forces might play a part in neural tube morphogenesis.
Fig. 14.

Rubber tubes used to model the morphogenesis of the cranial neural tube. Figures from His (1875). (A) Fig. 85. Flexion produces a transverse groove (gr) and two ‘ears’ (actually the optic Anlagen, arrowheads). (B) Fig. 86. When the tube is anchored at the white circle, then pushed in the direction of the black arrows, two ‘ears’, representing the optic Anlagen (arrowheads) and an oblique groove (white arrow) are formed. The anchorage point is the infundibulum attached to Rathke's pouch. (C) Fig. 82 (detail). Chicken embryo, 2 d incubation, ventral aspect of head end. The anchorage point in B is analogous to the infundibulum (white circle) in this embryo. (D) Fig. 84. Brain of chicken embryo, 3 d incubation, right lateral aspect. The infundibulum (black circle and Tr) represents the anchor point (white circle in B, C) which causes the bulging (curved arrow). (E) Fig. 87. A spindle‐shaped hole cut in the tube gapes to form a diamond‐shaped cavity when the tube undergoes convex flexure. (F) Fig. 88. With convex flexure of the slit tube, the floor of the aperture forms a transverse, saddle‐shaped ridge (r). (G) Fig. 14; chicken embryo, end of first day; dorsal aspect. The rubber model in E (shown in detail, left) resembles the shape of the neural Anlage where it passes from the head to neck (dashed lines). (H) Fig. 90. Chicken embryo, day 2, dorsal aspect. The open neural plate in the lumbar region resembles the slit tube in F (shown in detail, right).
Fig. 15.

Comparison of brain development in three different vertebrates: (A) pike; (B) chicken (top) and frog (bottom). Figs 92, 101 and 102, respectively, in His (1875). (C, D, E) Our interpretation of His's text (His, 1875, pp. 107–108) for the pike, chicken and frog, respectively. The midbrain (mb) is relatively long in the pike. The pontine flexure (pf) is massively developed in the pike, weakly developed in the chicken and scarcely present in the frog. The massive development of the pike pontine flexure causes part of the hindbrain (hb) to appear subducted along the ventral aspect of the midbrain. hf, hook flexure or ‘Hakenkrümung’. (F) Model formed by pushing two ends of a slit rubber tube towards each other; subduction occurs (arrow) of part of the tube at the point of the slit (asterisk). This configuration resembles the pontine flexure in A, and the apparent subduction of part of the hindbrain on the ventral aspect of the midbrain (A, C; it is only ‘apparent’ because the midbrain is always rostral to the hindbrain).
(a). Flexures in the early neural tube influence the form of the adult brain
His noted that the midbrain in avian and osteichthyan embryos is relatively larger than in mammals (Fig. 15). He argued that these size differences influence the subsequent development of the brain and its adult form (he had made these arguments before: His, 1873a, pp. 331–332). He explained the relatively massive expanse of the human hemispheres in terms of the strong development of the pontine flexure, which in turn causes the hindbrain to be subducted under the ventral aspect of the midbrain. The brain is therefore shortened and more readily overgrown by the hemispheres (His, 1875, pp. 114–116).
(b). Each brain region ‘inherits’ a unique growth rate from its early primordium
According to His's earlier research (His, 1868b) the forebrain lies at the zenith of three growth gradients. Its relatively rapid growth continues in later stages, as though an autonomous growth rate is ‘inherited’ from its antecedent in the neurula. The forebrain grows so fast that it comes to bulge beyond the rostral tip of the notochord.
(c). Evolutionary differences in the relative size of the brain regions are established by heterochrony
His presented evidence that teleost embryos have a long midbrain compared to other vertebrates — and a correspondingly small forebrain. This size difference persists in the adult. One mechanism suggested by His to explain this phenomenon is an evolutionary change in developmental timing (His, 1873a, p. 331), which we would now call ‘heterochrony’ (Table S2). For example, the pontine flexure and ‘hook flexure’ (His's fourth primary flexure, see Fig. 15) are prominent in the developing salmon, and develop relatively early. By contrast, in the chicken at a similar stage, neither flexure is well developed.
(8). After ‘Our Bodily Form’
In the years following the publication of ‘Our Bodily Form’ , His's major publications included a landmark study of human developmental anatomy (His, 1880, 1882a, 1885). He also made important contributions to developmental neurobiology, proposing a scheme of neural tube organisation in which it is divided into ‘longitudinal zones’: the roof plate, floor plate, and side walls. The latter were subdivided into what he called alar and basal plates (His, 1888b, p. 350; Table S2). Orr (1887, p. 329) had also alluded to the longitudinal organisation of the neural tube (in Anolis sagrei and other lizards) but did not provide a zonation scheme.
IV. THE INFLUENCE AND FATE OF HIS'S WORK
Prof. His has been led by his researches to adopt peculiar views concerning the causation of animal forms. These he has explained at some considerable length in his great work on the “Development of the Chick,” and elsewhere, but they have not met with very general acceptance (M.F., 1875)
(1). An outstanding legacy in developmental neurobiology and descriptive embryology
‘In almost every branch of anatomy [His] has left his mark, and during the last thirty years no one has done so much to advance our knowledge of that subject. … there is no one who has exercised a more powerful influence in moulding anatomical thought in almost all branches of that subject.’ (Cunningham, 1906, pp. 1235–1236)
The contributions of Wilhelm His, Sr to neurobiology (Supplementary Note S3) and descriptive embryology were of great importance and he coined many anatomical terms still in current use (Table S2; Swanson, 2015, p. 17). His's contributions to neuron theory earned him two Nobel Prize nominations (Table S1). Minot (1890, p. 891) referred to His's ‘great’ monograph on chicken development (His, 1868b) and one reviewer described it as the finest work of its kind since von Baer (Anon., 1869). His's ‘Anatomy of Human Embryos’ (His, 1880, 1882a, 1885) is a foundational work in that field (O'Rahilly, 1979; Dupont, 2018, p. 2).
His's ideas influenced Weiss's ‘contact guidance’ theory of neuronal target‐finding (Weiss, 1955, p. 356), and his discovery of the neural crest is a landmark in the history of developmental neurobiology (Hall, 2008; Bronner & Simoes‐Costa, 2016; Dupont, 2018; Glover et al., 2018). On the other hand, his epi‐spinal spaces in the brain may be artefacts (Woollam & Millen, 1954), and his facial reconstruction of J. S. Bach is questionable (Supplementary Note S2).
… [His's] work on the development of the human hindbrain published in 1890 provided novel ideas that more than 100 years later form the basis for penetrating molecular investigations of the regionalization of the hindbrain neural tube … (Glover et al., 2018, p. S14)
His's longitudinal scheme of the nervous system was also highly influential, although Kingsbury (1920) argued that it may apply only to the truncal neural tube. Recent studies (reviewed by Puelles, Domenech‐Ratio & Martinez‐De‐La‐Torre, 1987; Glover et al., 2018; Puelles, 2018) suggest that His correctly identified the rostral extent of the alar‐basal boundary, but not that of the floor and roof plates (Luis Puelles, personal communication). The ‘updated prosomeric model’ is a synthesis of His's concept with current data; according to that model, the brain of all vertebrates can be divided into His's longitudinal zones, and several transverse segments or neuromeres (Puelles, 2018) (Fig. 16A, B).
Fig. 16.
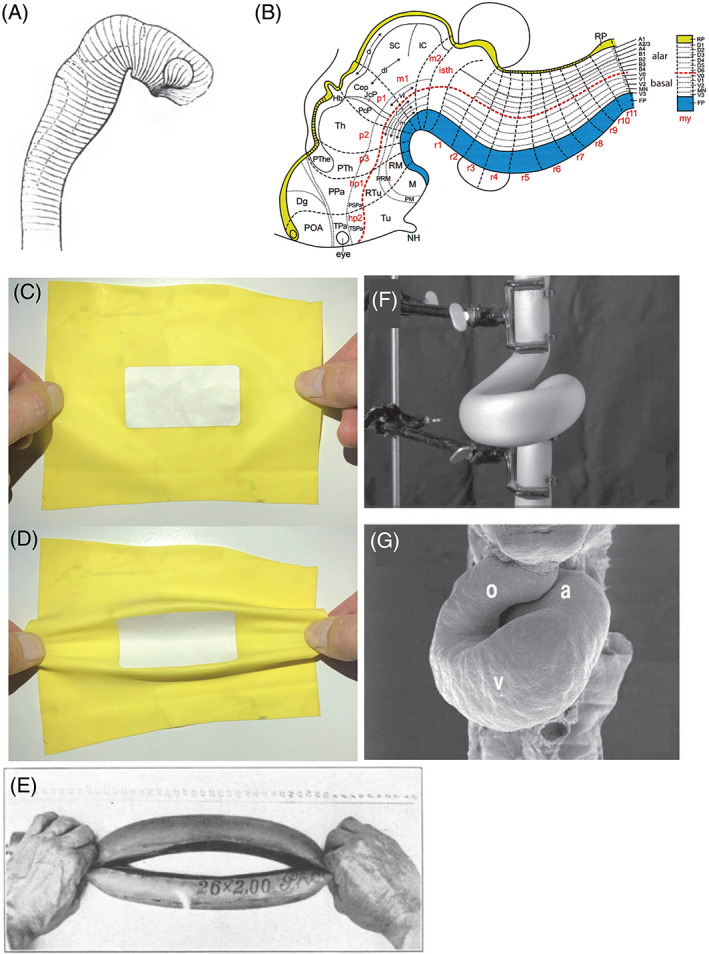
(A) His's model of longitudinal zones in the neural tube. Fig. 13 from His (1888b). (B) The ‘updated prosomeric’ model of brain organisation. Yellow, roof plate; blue, floor plate. Fig. 8 from Puelles (2018). (C, D) Folding along elasticity boundaries as a mechanism of early vertebrate morphogenesis (reconstructed by Dr. Fleury, based on fig. 16 in Fleury et al., 2015). A paper sticker is placed on a rubber sheet, thereby stiffening the central area. The stretched sheet buckles around the sticker, along boundaries between elastic and stiff regions. This demonstration is reminiscent of His' suggestion that the early embryo has variable elasticity and undergoes folding at unstable boundaries. (E) Vaihinger modelled the opening and closing of plant stomata by pushing and pulling a rubber tube (fig. 8 from Vaihinger, 1941). Courtesy of Universiteitsbibliotheek Utrecht. (F, G) Manipulation of a rubber tube (F) can create shapes resembling a scanning electron micrograph (G) of the developing cardiac tube of the chicken (fig. 5B, D from Männer, 2004).
In contrast to these impressive discoveries, His's theories of development and evolution did not have an enduring influence. This is partly because, as Mall explains, His ‘was never inclined to develop a school nor was he anxious to have pupils.’ (Mall, 1905, p. 150).
(2). Failure of the parablast and concrescence theories
… after the fall of the parablast theory, the concrescence theory became [His's] problem child … he demonstrated concrescence using rubber and wax models very beautifully and clearly; however, rubber and wax are not the materials from which embryos are composed. (Rabl, 1897, pp. xiii–xiv)
The parablast and concrescence theories were, in the early days, highly influential. Van Beneden (1878, p. 51) referred to the ‘famous’ parablast theory as did Fick (1904, p. 187). Edwin Klebs developed a ‘parablastome’ theory to classify cancers according to their supposed parablastic origin (Klebs, 1889, pp. 573–574). As for concrescence, Rabl was sceptical but nonetheless described it as ‘ … one of the most important developmental theories about the structure of the vertebrate body…’ (Rabl, 1897, p. xii). Kopsch, also a sceptic, argued that: ‘the epoch‐making significance of His's concept consists in the extraordinarily simple way in which the embryonic development of vertebrates and invertebrates can now be explained by the same processes’ (Kopsch, 1904, p. 4). Concrescence was still cited in textbooks well into the 20th century.
Both theories, however, were ultimately shown to be false (Brauckmann, 2006). Cell‐labelling experiments were particularly important in ruling out concrescence (Keller et al., 1991). For details of the scientific evidence against these theories, see Supplementary Note S6. Picken (1956) argued that the failure of the parablast and concrescence theories may have tarnished His's scientific reputation.
(3). His's mechanical developmental biology
Wilhelm His's approach to development is often called ‘mechanical’, a word that describes: (i) causal mechanisms; and (ii) the role of physical forces in development (Supplementary Note S7).
My attempts to introduce some elementary mechanical or physiological conceptions into embryology have not generally been agreed to by morphologists. (His, 1888a, p. 294)
His's mechanical developmental biology was important because it stimulated the study of developmental mechanisms, and because it provided a challenge to Haeckel's phylogenetic embryology (Supplementary Note S7). However, many scientists were critical because His inferred developmental mechanisms on the basis of histological studies and modelling, and not from in vivo experiments. Oscar Hertwig wrote: ‘ … I think more physiologically than His, who just wants to be the representative of a physiologically‐minded histology …’ (Hertwig, 1883, p. 124).
His's suggestion that differential growth is a major driver of gastrulation movements was contradicted by Rhumbler's studies suggesting that changes in cell shape and position, rather than growth, were important (see Table S4; Rhumbler, 1902, pp. 422–423). Subsequent studies have confirmed Rhumbler's view (Keller, 2002; Heisenberg & Bellaiche, 2013).
His suggested that neurulation was driven by pressure from tissues surrounding the neural primordium (Alexander Goette made similar suggestions; Goette, 1875, p. 186). However, Wilhelm Roux argued that the slightest variations in the parameters of the system would then cause the embryo to develop abnormally (Roux, 1895a, p. 242). Roux seems to be arguing that His's mechanism lacks what we now call developmental precision and robustness (see Kerszberg & Wolpert, 2007, pp. 205–206). Roux reported that surgically‐isolated neural primordia of chicken embryos in culture could still undergo normal neurulation movements, despite having been freed from any surrounding tissues (Roux, 1895a, pp. 247–248).
(a). His and developmental mechanics ( Entwicklungsmechanik)
… [to His] we owe a whole series of causal deductions. However, we cannot deny that the applicability of this method to making causal deductions is very limited, and in many cases the conclusions thus obtained do not offer the certainty which is so desirable in the case of such fundamental questions. (Roux, 1895a, p. 30)
The developmental mechanics or Entwicklungsmechanik of Wilhelm Roux (Maienschein, 1991) was ‘mechanics’ not in the sense of physical forces, but in the sense of causal mechanisms (Roux, 1895a, p. 11; Rádl, 1909, pp. 517–522; Sander, 1991; Counce, 1994). Roux praised His for establishing a causal approach to development (Roux, 1895a, p. 244), but complained that His did not test his hypotheses experimentally (Roux, 1895b, p. 174). So, while His and Roux shared a common interest in mechanisms, Roux was an experimentalist, His was not. Because of this, Roux felt that His's explanations were insufficient. For example, he argued that His's model of differential growth in the embryo does not explain what causes the differential growth in the first place (Roux, 1895a, p. 147); similar criticisms were voiced by Spitzer (1886, p. 211).
(b). His and the modern discipline of mechanobiology
In ‘Membranes and cavities’, His speculated about the effects of physical forces and cell–matrix interactions in determining cell behaviour. The modern discipline of mechanobiology addresses such phenomena (Carter et al., 1998; Moore, Roca‐Cusachs & Sheetz, 2010; Iskratsch, Wolfenson & Sheetz, 2014; De, Hwang & Kuhl, 2015, p. v.; Jansen et al., 2015; Paluch et al., 2015). Mechanobiology is a recent discipline, one of the earliest mentions of which is in van der Meulen, Beaupre & Carter (1993).
Cells can sense forces applied to the extracellular matrix, and the transduction of these forces can influence stem cell behaviour (Engler et al., 2006; Discher, Mooney & Zandstra, 2009). There is increasing evidence that these phenomena are important in development — as His had suggested. The application of biomechanics to development was called ‘embryo mechanics’ by Davidson, who cites ‘Our Bodily Form’ as an early work in the field (Davidson, 2011, 2017).
Physical forces are important in modulating cell behaviour in the developing nervous system (Van Essen, 1997; Hilgetag & Barbas, 2005). Some recent authors have found merit in His's rubber tube models of neural tube development (Sheesley et al., 2014; Edelman et al., 2016). Others have presented evidence that mechanical forces play an important role in the formation of gyri and sulci on the surface of the cortex (Tallinen et al., 2016). Physical forces established by blood flow patterns (haemodynamics) are important in the development of the cardiovascular system (reviewed by Stekelenburg‐de Vos et al., 2003; Groenendijk et al., 2005; Andres‐Delgado & Mercader, 2016; Poelmann & Gittenberger‐de Groot, 2018). Fusion of the cardiac tubes at the midline may be influenced by tensile forces in the endoderm (Varner & Taber, 2012).
At least one recent paper on mechanobiology cites ‘Our Bodily Form’ as an early work relevant to the field (Keller, Shook & Skoglund, 2008). But other reviews of the history of mechanobiology make no mention of His (Iskratsch et al., 2014; Wall et al., 2018), suggesting that their authors do not regard him as a major influence on the field.
(c). The viscoelasticity of embryos
The dispute between His and Haeckel over the elasticity of the embryo appears to have been resolved in His's favour. Elasticity has been demonstrated in early chicken (Agero, Glazier & Hosek, 2010) and Xenopus laevis (Zhou, Kim & Davidson, 2009) embryos. One current model sees embryos as viscoelastic, i.e. having both fluid and elastic properties. Viscoelasticity varies at different locations in early chicken embryos (Marrese et al., 2019). Fleury et al. (2015) modelled the viscoelastic properties of early embryos, and how they are deformed by physical forces. In one experiment, the authors applied forces to a sheet of rubber with a paper sticker fastened to it (Fig. 16C, D) and showed that boundaries between regions differing in elasticity were crucial hinge‐points (Fleury et al., 2015).
(4). His and evo‐devo
“… comparative developmental mechanics” is a science that lies in the future (Roux, 1895a, p. 441)
Evo‐devo (evolutionary developmental biology) is a discipline which came to prominence following the publication of Ontogeny and Phylogeny by Stephen Gould (Gould, 1977) and the cloning of homeobox genes in Drosophila and mammals in 1984 (see Richardson, 2009). One of the aims of evo devo is to explain phenotypic evolution in terms of evolutionary changes in developmental mechanisms (Raff, 2000; Swalla, 2002; Gilbert, 2003; Hall, 2003; Tickle & Urrutia, 2017). His's work has had little influence on the modern discipline of evo‐devo; Gould is the only reviewer among those cited above who mentions His's work. Nevertheless, we argue that ‘Our Bodily Form’ contains many ideas relevant to this field.
(a). Ontogenetic allometry
Mehnert, inspired by His's growth law and organ‐forming regions, suggested that organ primordia might function as independent growth centres (Mehnert, 1898; pp. 76–77). Today, such phenomena are considered under the rubric of ontogenetic allometry: changes in the relative dimensions of an individual or its parts during development (Gould, 1966; pp. 587, 601–602). Eduard Hagenbach's calculations, published in His (1868b), are an early example of the mathematical modelling of ontogenetic allometry; His's morphometric analysis of six amniote embryos is another (see Figs 10, 11, 12).
His's suggestion that the massive growth of the eye in the chick embryo causes the beak to develop, is an example of ontogenetic allometry (Fig. 13; His, 1875, pp. 204–206). Haeckel (1875, pp. 27–28) argued that it could just as easily be the case that the beak drives development of the eye. His's idea, however, is more plausible, given that the eye develops much earlier than the beak (a point suggested to us by Robert Poelmann). Discussions of beak morphogenesis are interesting in the light of a recent study of the variations in beak shape among Darwin's finches (Geospiza spp.; Abzhanov et al., 2004) which found that the timing of Bmp4 expression in the upper beak primordium might explain the ontogenic allometry of the beak.
(b). The problem with phenetic comparisons
His's comparison of six amniote embryos (Figs 10, 11, 12) is problematic for many reasons. The stages are not equivalent (Fig. 10; Spitzer, 1886, p. 152), and only external characters are considered. Additionally, His's drawing (see Fig. 10) of the deer embryo is problematic because of its tendentious depiction of the acropodia (Richardson & Keuck, 2001). Another major objection to His's comparison is that it is a phenetic comparison — a pre‐cladistic approach to grouping organisms based on their overall similarity (Cain & Harrison, 1958, 1960; Sneath & Sokal, 1962; Vernon, 1988; Richardson et al., 2001; Rossello‐Mora & Amann, 2001). As Spitzer (1886, p. 155) put it, His's comparisons were based on the ‘impression of the aesthetic similarity’ of embryos.
(c). The ‘developmental hourglass’
The influential developmental ‘hourglass’ (egg‐timer) model in evo‐devo is based on phenetic comparisons and on transcriptome analyses. It is a statement of the gross similarity of embryos at different stages of development. It envisages convergence of the external morphology of young stages towards a mid‐embryonic, conserved stage; and then divergence in morphology as embryos progress to postembryonic stages (reviewed by Duboule, 1994; Richardson & Keuck, 2002). The relatively high similarity of these mid‐embryonic stages among different species is suggested to reflect the conservation of gene regulatory networks active at those stages (Duboule, 1994; Prud'homme, Gompel & Carroll, 2007).
Von Baer had noted the similarity of early vertebrate embryos, and their subsequent divergence as they develop (von Baer, 1828, p. 221). He argued that all animals develop from a phenotypically similar early stage (von Baer, 1828, pp. 223–224): a vesicle, which Haeckel later equated with his ‘blastula’ (Haeckel, 1877, p. 153). Because von Baer's pattern of divergence represents only the upper part of the developmental hourglass, it has been called the developmental ‘funnel’ (Abzhanov, 2013).
It was His who first identified the hourglass pattern and Keibel (1906, p. 172) confirmed this, although neither used the term ‘developmental hourglass’. Recent transcriptomic studies have largely confirmed the existence of a middle stage in development where transcriptomes are relatively enriched with conserved developmental genes (Quint et al., 2012; Tena et al., 2014; Drost et al., 2015, 2016; Cridge, Dearden & Brownfield, 2016; Marlétaz et al., 2018; Yanai, 2018; discussed by Richardson, 2012).
(d). Embryonic characters in phylogenetic studies
His believed that ‘younger’ embryos show class, genus, species and individual traits; and he tried to group the species based on measurements of their bodily proportions (see Fig. 12; His, 1875, p. 201). Spitzer argued that this could potentially be used in a new classification of animals based on embryonic characters (Spitzer, 1886, pp. 154–156) but was sceptical about whether embryos could provide as much information as adult stages (Spitzer, 1886, p. 155). He also argued that using His's approach would mean that human and chick embryos, on the basis of their large heads, would be more closely related to one another than to the other species considered; a conclusion that would be inconsistent with known phylogenies.
(e). Quantitative methods in comparative embryology
It was, perhaps, in his views on the importance of measurement for the understanding of morphogenetic processes that His displayed the greatest originality. (Picken, 1956, p. 1163)
His's comparison of six amniote embryos, in which he applied morphometrics to comparative embryology (see Fig. 12) is a landmark in the history of evolution and development. It attempts to supplement phenetic statements about embryonic similarity with quantitative data on the relative dimensions of embryonic regions. However, His's data were limited to three regions in six embryos at a single stage with no replicates. Modern quantitative approaches to comparative embryology are found in the study of heterochrony (e.g. Jeffery et al., 2005; de Jong et al., 2009; reviewed by Klingenberg, 1998; Keyte & Smith, 2014), allometry (Klingenberg, 2016), and developmental transcriptomics.
(f). Coordinate systems, positional information and gradients
Some of the concepts that His explored have more recently become part of our understanding of developmental mechanisms. These include such concepts as coordinate systems, positional information gradients and cell–cell signalling. However, His's work on these concepts did not lead to lasting insights, and later interest in those same subjects appears to be independent of any influence or knowledge of His's work.
His used a coordinate system to map loci of differential growth, and growth gradients, in the embryo (His, 1875, pp. 121–122; His, 1867a, p. 619; His, 1868b, p. 185). Later, D'Arcy Thompson used x–y coordinates to map body plans onto rubber sheets; he then stretched the sheets to model the effects of differential growth (Thompson, 1945, p. 1087). Driesch also considered development in terms of coordinates: ‘what actually will happen in each of the blastula cells in any special case of development experimentally determined depends on the position of that cell in the whole, if the ‘the whole’ is put into relation with any fixed system of co‐ordinates … ’ (Driesch, 1908, p. 80; see also Driesch, 1891, pp. 28–29). Another example of the use of coordinate systems in anatomy is in its application to brain structure (Nieuwenhuys & Puelles, 2016).
His postulated that each organ primordium adopts its own growth law depending on its spatial relationships with its neighbours (His, 1875, p. 83). This is a statement of how a rapidly growing tissue might apply mechanical force to its neighbours. An analogous suggestion was made by Schwendener in his positional theory of phylotaxis based on mechanical influences (Schwendener, 1878).
His proposed that cells and tissues might influence each other via the secretion of chemicals (His, 1865b, p. 33). However, he considered gradients not in terms of concentration gradients in chemicals, but as growth gradients. The concept of growth gradients that His proposed does not appear to have influenced the development of gradient theory by Child (1928), Runnström (1914) and others (reviewed by Spemann, 1938, pp. 318–345). Spemann stated that the concept of developmental gradients ‘as far as I know, was first proposed and confirmed by Th. Boveri’ (Spemann, 1938, p. 318). Gradients of morphogens became a key concept in Wolpert's positional information model (Wolpert, 1969), although he later modified this view (Kerszberg & Wolpert, 2007; Wolpert, 2009).
(5). Modelling
His used a variety of physical materials in order to model developmental processes (Table S4), and to make 3‐D models for teaching (Hopwood, 1999, 2002, 2012). Haeckel poured scorn on His's ‘comical’ models that used rubber tubes and postal envelopes (Haeckel, 1875, pp. 26–27). However, many developmental biologists have used models of various kinds (e.g. Fig. 16; Table S4): Rhumbler (1902) made a model of the blastula out of corset stays, Vaihinger (1941) used rubber tubes to model the biomechanics of stomatal opening, and Männer (2004) applied forces to a rubber tube to model cardiac looping (Fig. 16C–G; see also Taber, Lin & Clark, 1995).
(6). The fate map and organ‐forming regions
This view was formerly introduced into embryology by His as the principle of organ‐forming germ regions; it shows affinity to the views of Weismann and Roux (Maas, 1903, p. 65). (The 'view' referred to was that different regions of the early embryo develop into distinct parts of the adult)
(a). The fate map
One of His's research goals was to establish an exact topography of the chicken embryonic disc (His, 1877a, p. 112). His's map of organ‐forming regions (Fig. 8) is the earliest fate map known to us. However, it was entirely speculative; true fate maps would only emerge from cell‐labelling experiments (Goodale, 1911; Smith, 1914; Vogt, 1929; Psychoyos & Stern, 1996). When Rawles compared His's fate map to her own experimental results, she concluded: ‘while a certain striking similarity exists between the theoretical concepts of His and the present experimental analysis with regard to some points, His's scheme is based on too simple a ground to justify detailed comparison.’ (Rawles, 1936, p. 312).
(b). Organ‐forming regions and mosaicism
His's theory of organ‐forming regions was influential in discussions of mosaic development (Supplementary Note S8). Roux wrote: ‘If a whole arises from several or many independently differentiating parts, it is assembled, like a mosaic, from individual parts that have been formed; I have called this kind of development a “mosaic work”’ (Roux, 1895a, p. 821). Roux noted that ‘His’ principle of organ‐forming germ‐regions … only has a causal meaning insofar as it designates the locations of the resultant formation of manifold interactions' (Roux, 1895a, p. 825).
Weismann Parker & Rönnfeldt (1893) state that His's theory of organ‐forming regions was the first, albeit imperfect, expression of the predestination of cells. They argue that His's theory correctly suggests that the ‘differentiating principle’ of development lies within the cell itself and not in external influences (Weismann et al., 1893, pp. 134–135). His's concept also influenced Wilson's ‘germinal localisation’ theory (reviewed by Kollmann, 1904; Wilson, 1904). For more recent opinions on regulation and mosaicism, see Boiani et al. (2019) and Stern (2006).
(c). Organ‐forming regions as fields
The organ‐forming regions of His are growth centres. But he did also talk in terms of ‘fields’ demarcated by ‘boundary markers’ in the early chick embryo (His, 1875; pp. 45–46) as, later, did Wolpert (1969; see also Jacobson, 1983, p. 1036). The concept of morphogenetic fields was elaborated first by Alexander Gurwitsch and then by Paul Weiss (Beloussov, Opitz & Gilbert, 1997). Today, developmental fields are considered to be regions undergoing morphogenesis or pattern formation (Wolpert, 1969; De Robertis, 2009).
(7). Technical innovations
The technic of investigation made little progress, and it is in this particular that His made a change. He had prepared himself by a study of the embryology of the chick and in connection with this study had worked out a successful technic; he fixed his embryos, he had constructed an apparatus for section cutting which, primitive as it now seems, led the way to our modem microtomes, and he had thought out a method of reconstruction which has been gradually improved, especially by Born, into a method that is now absolutely indispensable in embryological investigation. (Keibel & Mall, 1910, p. xv)
His's microtome was certainly not the first section cutter. In 1770, Hill described a ‘cutting engine’ designed by Cummings for the sectioning of woody plant tissue (Hill, 1770). Adams' microtome had a micrometer‐screw advance (Adams & Kanmacher, 1798, p. 128; Chvátal, 2017). Chevalier used the name ‘microtome’ for such devices (Chevalier, 1839, p. 192). A microtome for the sectioning of animal tissue was developed by Oschatz in Purkyně's lab, long before the invention of paraffin embedding (Oschatz, 1844; Chvátal, 2017).
The sections that His cut routinely were 50 μm in thickness — far thicker than the 5–7 μm sections common today. His was still using his microtome in his studies of human embryos in the 1880s. But it is likely, from reading his text and tables (His, 1880, pp. 118, 137; His, 1885, pp. 7–9) that he used even thicker sections (up to 200 μm) for particuarly large embryos and fetuses. Hermann Fol suggested that this was too thick, and that it may have led to errors in interpretation (Fol, 1884, p. 381; His, 1885, p. 11 n.). Hochstetter also questioned the reliability of His's interpretations and criticised the poor quality of his human embryos (Hochstetter, 1919; pp. 1, 9). On these grounds, Hochstetter argued that the fame and influence of His's publications on human embryology may not be entirely deserved. Eventually, His abandoned his own microtome in favour of the Altmann‐Schanze microtome which yielded sections of 20–25 μm (Anon., 1885, pp. 547–548; His, 1885, p. 7).
V. CONCLUSIONS
His was a developmental anatomist and technical innovator. Much of his descriptive work is now part of the fabric of embryology, anatomy and neurobiology.
His's publications were full of new ideas about evolution, development and mechanical developmental biology. Few of these ideas had an enduring influence, perhaps because His never cultivated a school of followers. Another reason is that these theories were speculations based solely on histological data and modelling. Experimental embryology later would provide more persuasive data and more robust theories.
Despite the criticism that they received, many of His's ideas have analogues in modern biology. They are only analogues – independent occurrences of the same idea. But the study of their origin and fate provides a fascinating case study in the history of scientific ideas.
Supporting information
Fig. S1. Keys to sitters in the group photographs shown in Fig. 2 of the main text.
Fig. S2. Bibliographic analysis of the research themes of Wilhelm His.
Table S1. Wilhelm His, Sr. – selected biographical landmarks.
Table S2. Glossary of terms and concepts relevant to this review.
Table S3. Opinions of earlier researchers on the origins of the peripheral ganglia and the nephric duct.
Table S4. Models used by His and others to explain and understand developmental processes.
Table S5. Table of Contents of ‘Our Bodily Form’ (His, 1875) with chapter summaries.
Supplementary Note S1. Additional information on the oil painting of Wilhelm His shown in Fig. 1D.
Supplementary Note S2. Contributions of Wilhelm His Sr. to anthropology and forensic craniofacial reconstruction.
Supplementary Note S3. Contributions of Wilhelm His Sr. to developmental neurobiology.
Supplementary Note S4. The conflict between Wilhelm His and Ernst Haeckel.
Supplementary Note S5. Other polemics and disputes.
Supplementary Note S6. Scientific opinion on the parablast and concrescence theories.
Supplementary Note S7. His's ‘mechanical’ developmental biology: its meaning and its reception.
Supplementary Note S8. Mosaic versus regulatory development.
VI. ACKNOWLEDGEMENTS
We are indebted to many people for valuable discussions and for sourcing documents. This review does not necessarily reflect their opinions, and any mistakes are ours. We thank: Thomas Bach, Peter Bowler, Frédéric Bussmann, Alexandr Chvátal, Lance Davidson, Christine Feja, Vincent Fleury, Michael Gerbl, Manfred Gessler, Andrew Harrison, Carl‐Phillip Heisenberg, Nick Hopwood, Phil Ingham, Cecile de Jong, Cornelia Junge, Ray Keller, Vanessa Klein, Hugo Kurz, Roger Kurz, Liz Lockett, Jane Maienschein, Jörg Männer, Friedrich Paulsen, Robert Richards, Martin Rücklin, Carl Stephan, and Richard Zegers. The suppliers of figures are acknowledged in the figure legends. The Universitäts‐ und Landesbibliothek Münster, provided a copy of Acten in Sachen (His, 1868a). Luis Puelles and Thomas Mueller were particularly helpful in helping us understand the significance of His's longitudinal model of the brain. Merijn de Bakker, Robert Poelmann, Luis Puelles and Colin Richardson kindly provided useful comments on the manuscript.
[Corrections added on 22 February 2022, after first online publication: Minor modifications have been applied in this version.]
REFERENCES
- Abzhanov, A. (2013). von Baer's law for the ages: lost and found principles of developmental evolution. Trends in Genetics 29, 712–722. [DOI] [PubMed] [Google Scholar]
- Abzhanov, A. , Protas, M. , Grant, B. R. , Grant, P. R. & Tabin, C. J. (2004). Bmp4 and morphological variation of beaks in Darwin's finches. Science 305, 1462–1465. [DOI] [PubMed] [Google Scholar]
- Adams, G. & Kanmacher, F. (1798). Essays on the Microscope, Second Edition. Jones, London. [Google Scholar]
- Agero, U. , Glazier, A. & Hosek, M. (2010). Bulk elastic properties of chicken embryos during somitogenesis. BioMedical Engineering OnLine 9, 19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson, R. H. & Mori, S. (2016). Wilhelm His Junior and his bundle. Journal of Electrocardiology 49, 637–643. [DOI] [PubMed] [Google Scholar]
- Andres‐Delgado, L. & Mercader, N. (2016). Interplay between cardiac function and heart development. Biochimica et Biophysica Acta 1863, 1707–1716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anon (1869). Untersuchungen über die erste Anlage des Wirbelthierleibes, von Wilhelm His. Leipzig: Vogel. Monthly Microscopical Journal 1, 246–247. [Google Scholar]
- Anon (1885). Schanze's microtome. Journal of the Royal Microscopical Society 5, 547–548. [Google Scholar]
- Anon (1904a). Obituary. Professor Wilhelm His. Boston Medical and Surgical Journal 150, 631. [Google Scholar]
- Anon (1904b). Obituary: Wilhelm His, Professor of Anatomy, University of Leipzig. British Medical Journal 21, 1225–1226. [Google Scholar]
- Assmann, F. G. (1847). Quellenkunde der vergleichenden Anatomie als Vorläufer einer pragmatischen Geschichte der Zootomi. Vieweg, Braunschweig. [Google Scholar]
- Balfour, F. M. (1875). On the development of the spinal nerves in elasmobranch fishes. Proceedings of the Royal Society of London 24, 134–136. [Google Scholar]
- Balfour, F. M. (1876). On the development of the spinal nerves in elasmobranch fishes. Philosphical Transactions of the Royal Society of London. Series B, Biological Sciences 166, 175–195. [Google Scholar]
- Balfour, F. M. (1878). A Monograph on the Development of Elasmobranch Fishes. Macmillan, London. [Google Scholar]
- Balfour, F. M. (1880). A Treatise on Comparative Embryology, Edition (Volume 1). Macmillan and Co, London. [Google Scholar]
- Balfour, F. M. (1881). A Treatise on Comparative Embryology, Edition (Volume 2). Macmillan and Co, London. [Google Scholar]
- Beard, J. (1888). Morphological studies II. The development of the peripheral nervous system of vertebrates. Part I. Elasmobranchii and Aves. Quarterly Journal of Microscopical Science 29(New Series, 153–227. [Google Scholar]
- Beloussov, L. V. , Opitz, J. M. & Gilbert, S. F. (1997). Life of Alexander G. Gurwitsch and his relevant contribution to the theory of morphogenetic fields. International Journal of Developmental Biology 41, 771–777. [PubMed] [Google Scholar]
- Bertolini, R. (1965). Wilhelm His (1831–1904). In Bedeutende Gelehrte in Leipzig (Volume 2, ed. Steinmetz M.), pp. 87–94. Karl‐Marx‐Universität, Leipzig. [Google Scholar]
- Bidder, F. & Kupffer, C. (1857). Untersuchungen über die Textur des Rückenmarks und die Entwicklung seiner Formelemente. Breitkopf und Haertl, Leipzig. [Google Scholar]
- Bischoff, T. L. W. (1877). Historisch‐kritische Bemerkungen zu den neuesten Mittheilungen über die erste Entwicklung der Säugethiereier. Literarisch‐artistische Anstalt, München. [Google Scholar]
- Boiani, M. , Casser, E. , Fuellen, G. & Christians, E. S. (2019). Totipotency continuity from zygote to early blastomeres ‐ a model under revision. Reproduction 158, R49–R65. [DOI] [PubMed] [Google Scholar]
- Braem, F. (1895). Was ist ein Keimblatt? Biologisches Centralblatt 15, 427–443; 466–476; 491–506. [Google Scholar]
- Brauckmann, S. (2006). August Rauber (1841–1917): from the primitive streak to Cellularmechanik. The International Journal of Developmental Biology 50, 439–449. [DOI] [PubMed] [Google Scholar]
- Bronn, H. G. (1860). Charles Darwin, über die Entstehung der Arten im Thier‐ und Pflanzen‐Reich durch natürliche Züchtung, oder Erhaltung der vervollkommneten Rassen im Kampfe um's Daseyn. Schweizerbart, Stuttgart. [Google Scholar]
- Bronner, M. E. & Simoes‐Costa, M. (2016). The neural crest migrating into the twenty‐first century. Current Topics in Developmental Biology 116, 115–134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cain, A. J. & Harrison, G. A. (1958). An analysis of the taxonomist's judgment of affinity. Proceedings of the Zoological Society of London 131, 85–98. [Google Scholar]
- Cain, A. J. & Harrison, G. A. (1960). Phyletic weighting. Proceedings of the Zoological Society of London 135, 1–31. [Google Scholar]
- Čapek, D. & Müller, P. (2019). Positional information and tissue scaling during development and regeneration. Development 146, 1–16. [DOI] [PubMed] [Google Scholar]
- Carter, D. R. , Beaupre, G. S. , Giori, N. J. & Helms, J. A. (1998). Mechanobiology of skeletal regeneration. Clinical Orthopaedics and Related Research 355, S41–S55. [DOI] [PubMed] [Google Scholar]
- Chevalier, C. (1839). Des Microscopes et de Leur Usage. Crochard, Paris. [Google Scholar]
- Child, C. M. (1928). The physiological gradients. Protoplasma 5, 447–476. [Google Scholar]
- Chvátal, A. (2017). Jan Evangelista Purkyně (1787–1869) and his instruments for microscopic research in the field of neuroscience. Journal of the History of the Neurosciences 26, 1–19. [DOI] [PubMed] [Google Scholar]
- Counce, S. J. (1994). Archives for developmental mechanics. W. Roux, editor (1894–1924). Roux's Archives of Developmental Biology 204, 79–92. [DOI] [PubMed] [Google Scholar]
- Cridge, A. G. , Dearden, P. K. & Brownfield, L. R. (2016). Convergent occurrence of the developmental hourglass in plant and animal embryogenesis? Annals of Botany 117, 833–843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cunningham, D. J. (1906). Wilhelm His, K. S. Geheimer Rath, Professor of Anatomy, Leipzig, Honorary Fellow of the Royal Society of Edinburgh, 1900–1904. Proceedings of the Royal Society of Edinburgh 25, 1235–1240. [Google Scholar]
- Dahm, R. (2005). Friedrich Miescher and the discovery of DNA. Developmental Biology 278, 274–288. [DOI] [PubMed] [Google Scholar]
- Darwin, C. (1859). On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. Murray, London. [PMC free article] [PubMed] [Google Scholar]
- Davidson, L. A. (2011). Embryo mechanics: balancing force production with elastic resistance during morphogenesis. Current Topics in Developmental Biology 95, 215–241. [DOI] [PubMed] [Google Scholar]
- Davidson, L. A. (2017). Mechanical design in embryos: mechanical signalling, robustness and developmental defects. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 372, 1–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Jong, I. L. M. , Colbert, M. W. , Witte, F. & Richardson, M. K. (2009). Polymorphism in developmental timing: intraspecific heterochrony in a Lake Victoria cichlid. Evolution and Development 11, 625–635. [DOI] [PubMed] [Google Scholar]
- De Robertis, E. M. (2009). Spemann's organizer and the self‐regulation of embryonic fields. Mechanisms of Development 126, 925–941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De, S. , Hwang, W. & Kuhl, E. (2015). Preface. In Multiscale Modeling in Biomechanics and Mechanobiology (eds De S., Hwang W. and Kuhl E.), p. 286. Springer, London. [Google Scholar]
- DeFelipe, J. (2015). The dendritic spine story: an intriguing process of discovery. Frontiers in Neuroanatomy 9, 14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dejerine, J. (1895). Anatomie des Centres Nerveux. Rueff, Paris. [Google Scholar]
- Discher, D. E. , Mooney, D. J. & Zandstra, P. W. (2009). Growth factors, matrices, and forces combine and control stem cells. Science 324, 1673–1677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dixon, A. F. (1904). Professor Wilhelm His. Journal of Anatomy and Physiology 38, 503–505. [PMC free article] [PubMed] [Google Scholar]
- Driesch, H. (1891). Die mathematisch‐mechanische Betrachtung morphologischer Probleme der Biologie. Eine kritische Studie. Fischer, Jena. [Google Scholar]
- Driesch, H. (1892). Entwicklungsmechanische Studien. Zeitschrift für wissenschaftliche Zoologie 53, 160–184. [Google Scholar]
- Driesch, H. (1908). The Science and Philosophy of the Organism: Gifford Lectures Delivered at Aberdeen University, 1907–1908 by Hans Driesch. Printed for the University, Aberdeen. [Google Scholar]
- Drischel, H. (1965). Carl Friedrich Wilhelm Ludwig (1816–1895). In Bedeutende Gelehrte in Leipzig (Volume 2, ed. Steinmetz M.), pp. 73–86. Karl‐Marx‐Universität, Leipzig. [Google Scholar]
- Drost, H. G. , Bellstadt, J. , O'Maoileidigh, D. S. , Silva, A. T. , Gabel, A. , Weinholdt, C. , Ryan, P. T. , Dekkers, B. J. , Bentsink, L. , Hilhorst, H. W. , Ligterink, W. , Wellmer, F. , Grosse, I. & Quint, M. (2016). Post‐embryonic hourglass patterns mark ontogenetic transitions in plant development. Molecular Biology and Evolution 33, 1158–1163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drost, H. G. , Gabel, A. , Grosse, I. & Quint, M. (2015). Evidence for active maintenance of phylotranscriptomic hourglass patterns in animal and plant embryogenesis. Molecular Biology and Evolution 32, 1221–1231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duboule, D. (1994). Temporal colinearity and the phylotypic progression: a basis for the stability of a vertebrate Bauplan and the evolution of morphologies through heterochrony. Development (Suppl.) 1994, 135–142. [PubMed] [Google Scholar]
- Dupont, J.‐C. (2017). Wilhelm His and mechanistic approaches to development at the time of Entwicklungsmechanik. History and Philosophy of the Life Sciences 39, 211–219. [DOI] [PubMed] [Google Scholar]
- Dupont, J.‐C. (2018). Historical perspective on neuroembryology: Wilhelm His and his contemporaries. Genesis 56, e23218. [DOI] [PubMed] [Google Scholar]
- Dursy, E. (1866). Der Primitivstreif des Hühnchens. Schauenburg, Lahr. [Google Scholar]
- Dursy, E. (1869). Zur Entwicklungsgeschichte des Kopfes des Menschen und der höheren Wirbeltiere [Text]. Laupp, Tübingen. [Google Scholar]
- Edelman, D. B. , McMenamin, M. , Sheesley, P. & Pivar, S. (2016). Origin of the vertebrate body plan via mechanically biased conservation of regular geometrical patterns in the structure of the blastula. Progress in Biophysics and Molecular Biology 121, 212–244. [DOI] [PubMed] [Google Scholar]
- Engler, A. J. , Sen, S. , Sweeney, H. L. & Discher, D. E. (2006). Matrix elasticity directs stem cell lineage specification. Cell 126, 677–689. [DOI] [PubMed] [Google Scholar]
- Fick, A. (1895). Karl Ludwig. A. Nachruf von Adolf Fick. In Biographische Blätter. Jahrbuch für lebensgeschichtliche Kunst und Forschung (Volume 1), pp. 265–271. Hoffman, Berlin. [Google Scholar]
- Fick, R. (1904). Wilhelm His. Anatomischer Anzeiger 25, 161–208. [Google Scholar]
- Fleury, V. , Chevalier, N. R. , Furfaro, F. & Duband, J. L. (2015). Buckling along boundaries of elastic contrast as a mechanism for early vertebrate morphogenesis. The European Physical Journal E Soft Matter and Biological Physics 38, 92. [DOI] [PubMed] [Google Scholar]
- Fol, H. (1884). Description d'un embryon humain de cinq millimètres et six dixièmes. Recueil Zoologique Suisse 1, 357–401. [Google Scholar]
- Frank, M. H. & Weiss, J. J. (1966). The 'Introduction' to Carl Ludwig's textbook of human physiology. Medical History 10, 76–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freeman, R. B. (1977). The Works of Charles Darwin: An Annotated Bibliographical Handlist. Dawson/Archon, Folkestone/Hamden. [Google Scholar]
- Fye, W. B. (1986). Carl Ludwig and the Leipzig Physiological Institute: ‘a factory of new knowledge’. Circulation 74, 920–928. [DOI] [PubMed] [Google Scholar]
- Gilbert, S. F. (2003). The morphogenesis of evolutionary developmental biology. International Journal of Developmental Biology 47, 467–477. [PubMed] [Google Scholar]
- Glover, J. C. , Elliott, K. L. , Erives, A. , Chizhikov, V. V. & Fritzsch, B. (2018). Wilhelm His' lasting insights into hindbrain and cranial ganglia development and evolution. Developmental Biology 444(Suppl 1), S14–S24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goette, A. (1874). Beiträge zur Entwicklungsgeschichte der Wirbelthiere. Archiv für Mikroskopische Anatomie 10, 145–199. [Google Scholar]
- Goette, A. (1875). Die Entwicklungsgeschichte der Unke (Bombinator igneus) als Grundlage einer vergleichenden Morphologie der Wirbelthiere. Leopold Voss, Leipzig. [Google Scholar]
- Goodale, H. D. (1911). The early development of Spelerpes bilineatus (Green). American Journal of Anatomy 12, 173–247. [Google Scholar]
- Gould, S. J. (1966). Allometry and size in ontogeny and phylogeny. Biological Reviews 41, 587–640. [DOI] [PubMed] [Google Scholar]
- Gould, S. J. (1977). Ontogeny and Phylogeny. Belknap Press, Cambridge. [Google Scholar]
- Gould, S. J. (1997). The exaptive excellence of spandrels as a term and prototype. Proceedings of the National Academy of Sciences of the United States of America 94, 10750–10755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Groenendijk, B. C. , Hierck, B. P. , Vrolijk, J. , Baiker, M. , Pourquie, M. J. , Gittenberger‐de Groot, A. C. & Poelmann, R. E. (2005). Changes in shear stress‐related gene expression after experimentally altered venous return in the chicken embryo. Circulation Research 96, 1291–1298. [DOI] [PubMed] [Google Scholar]
- Haeckel, E. (1866). Generelle Morphologie der Organismen. Reimer, Berlin. [Google Scholar]
- Haeckel, E. (1868). Natürliche Schöpfungsgeschichte. Georg Reimer, Berlin. [Google Scholar]
- Haeckel, E. (1872). Die Kalkschwämme. Eine Monographie. Vol. 1 (Genereller Theil): Biologie der Kalkschwämme. Reimer, Berlin. [Google Scholar]
- Haeckel, E. (1874). Anthropogenie oder Entwickelungsgeschichte des Menschen: gemeinverständlich wissenschaftliche Vorträge über die Grundzüge der menschlichen Keimes‐ und Stammes‐Geschichte. Engelmann, Leipzig. [Google Scholar]
- Haeckel, E. H. P. A. (1875). Ziele und Wege der heutigen Entwicklungsgeschichte. Jenaische Zeitschrift für Naturwissenschaft 10(Suppl), 1–99. [Google Scholar]
- Haeckel, E. (1877). Biologische Studien. 2: Studien zur Gastraea‐Theorie. Dufft, Jena. [Google Scholar]
- Hall, B. K. (2003). Evo‐devo: evolutionary developmental mechanisms. International Journal of Developmental Biology 47, 491–495. [PubMed] [Google Scholar]
- Hall, B. K. (2008). The neural crest and neural crest cells: discovery and significance for theories of embryonic organization. Journal of Biosciences 33, 781–793. [DOI] [PubMed] [Google Scholar]
- Haymaker, W. & Schiller, F. (1970). The Founders of Neurology: One Hundred and Forty‐six Biographical Sketches by Eighty‐Eight Authors. Thomas, Springfield. [Google Scholar]
- Hehl, U. V. , Heydemann, G. , Fitschen, K. & König, F. (2010). Das zwanzigste Jahrhundert. Leipziger Universitätsverlag, Leipzig. [Google Scholar]
- Heisenberg, C. P. & Bellaiche, Y. (2013). Forces in tissue morphogenesis and patterning. Cell 153, 948–962. [DOI] [PubMed] [Google Scholar]
- Hensen, V. (1863). Studien Über das Gehörorgan der Decapoden. Engelmann, Leipzig. [Google Scholar]
- Hensen, V. (1864a). Ueber die Entwickelung des Gewebes und der Nerven im Schwanze der Froschlarve. Archiv für Pathologische Anatomie und Physiologie und für Klinische Medicin 31, 51–73. [Google Scholar]
- Hensen, V. (1864b). Zur Entwickelung des Nervensystems. Archiv für Pathologische Anatomie und Physiologie und für Klinische Medicin 30, 176–186. [Google Scholar]
- Hensen, V. (1867). Embryologische Mittheilungen. Archiv für Mikroskopische Anatomie 3, 500–503. [Google Scholar]
- Hensen, V. (1876). Beobachtungen über die Befruchtung und Entwicklung des Kaninchens und Meerschweinchens. Zeitschrift für Anatomie und Entwickelungsgeschichte 1, 213–273, 353–423. [Google Scholar]
- Hensen, V. (1903). Die Entwickelungsmechanik der Nervenbahnen im Embryo der Säugetiere: Ein Probeversuch. Lipsius & Tischer, Kiel & Leipzig. [Google Scholar]
- Hertwig, O. (1883). Studien zur Blättertheorie. 5: Die Entwicklung des mittleren Keimblattes der Wirbelthiere. Fischer, Jena. [Google Scholar]
- Hertwig, O. (1888). Lehrbuch der Entwicklungsgeschichte des Menschen und der Wirbelthiere, Second Edition. Fischer, Jena. [Google Scholar]
- Hertwig, O. (1892). Text‐Book of the Embryology of Man and Mammals (English transl.). Sonnenschein, London. [Google Scholar]
- Hertwig, O. (1906). Einleitung und allgemeine Litteraturübersicht. Fischer, Jena. [Google Scholar]
- Hertwig, O. & Hertwig, R. (1881). Studien zur Blättertheorie IV. Die Coelomtheorie. Versuch einer Erklärung des mittleren Keimblattes. Fischer, Jena. [Google Scholar]
- Hildebrand, R. (2005). “… that progress in anatomy is most likely to occur when its problems include the study of growth and function, as well as of structure”. Über den Anatomen und Physiologen Ernst Heinrich Weber (1795–1878) und über Wilhelm His (1831–1904), seinen Nachfolger auf dem Lehrstuhl für Anatomie an der Universität Leipzig. Annals of Anatomy 187, 439–459. [DOI] [PubMed] [Google Scholar]
- Hilgetag, C. C. & Barbas, H. (2005). Developmental mechanics of the primate cerebral cortex. Anatomy and Embryology 210, 411–417. [DOI] [PubMed] [Google Scholar]
- Hill, J. (1770). The Construction of Timber from Its Early Growth (etc.). Self‐published, London. [Google Scholar]
- His, W. (1854). Untersuchungen über den Bau der Hornhaut. Verhandlungen der Physikalisch‐medicinischen Gesellschaft in Würzburg 4, 90–96. [Google Scholar]
- His, W. (1856). Beiträge zur normalen und pathologischen Histologie der Cornea. Schweighauser, Basel. [Google Scholar]
- His, W. (1862). Untersuchungen über den Bau der Peyerschen Drüsen und der Darmschleimhaut. Zeitschrift für wissenschaftliche Zoologie 11, 416–442. [Google Scholar]
- His, W. (1863a). Ueber die Endigung der Gefässnerven. Archiv für pathologische Anatomie und Physiologie und für klinische Medicin 28, 427–428. [Google Scholar]
- His, W. (1863b). Ueber die Wurzeln der Lymphgefässe in den Häuten des Körpers. Zeitschrift für wissenschaftliche Zoologie 12, 223–254. [Google Scholar]
- His, W. (1865a). Beobachtungen über den Bau des Säugethier‐Eierstockes. Archiv für Mikroskopische Anatomie 1, 151–202. [Google Scholar]
- His, W. (1865b). Die Häute und Höhlen des Körpers: Academisches Programm. Schweighauser, Basel. [Google Scholar]
- His, W. (1865c). Ueber ein perivasculäres Kanalsystem in den nervösen Centralorganen und über dessen Beziehungen zum Lymphsystem. Zeitschrift für wissenschaftliche Zoologie 15, 127–140. [Google Scholar]
- His, W. (1866a). Ueber die erste Anlage des Wirbelthierleibes. Archiv für Mikroskopische Anatomie 2, 515–524. [Google Scholar]
- His, W. (1866b). Ueber die erste Anlage des Wirbelthierleibes. Verhandlungen der Naturforschenden Gesellschaft in Basel 4, 484–496. [Google Scholar]
- His, W. (1867a). Ueber die erste Anlage des Wirbelthierleibes (Fortsetzung). Verhandlungen der Naturforschenden Gesellschaft in Basel 4, 617–639. [Google Scholar]
- His, W. (1867b). Ueber Lymphgefässe in den nervösen Centralorganen. Verhandlungen der Naturforschenden Gesellschaft in Basel 4, 122–127. [Google Scholar]
- His, W. (1868a). Acten in Sachen der von Professor E. Dursy gegen W. His erhobenen Anklagen (Als Manuscript gedruckt). Vogel, Leipzig. [Google Scholar]
- His, W. (1868b). Untersuchungen über die erste Anlage des Wirbelthierleibes. Die erste Entwickelung des Hühnchens im Ei, Vogel, Leipzig. [Google Scholar]
- His, W. (1870a). Beschreibung eines Mikrotoms. Archiv für Mikroskopische Anatomie 6, 229–232. [Google Scholar]
- His, W. (1870b). Über die Bedeutung der Entwicklungsgeschichte für die Auffassung der organischen Natur. Rectoratsrede gehalten den 4. November 1869. Vogel, Leipzig. [Google Scholar]
- His, W. (1873a). Ueber die Gliederung des Gehirns. Verhandlungen der Naturforschenden Gesellschaft in Basel 5, 327–340. [Google Scholar]
- His, W. (1873b). Untersuchungen über die Erste Anlage des Wirbelthierleibes: Untersuchungen über das Ei und die Eientwickelung bei Knochenfischen. Vogel, Leipzig. [Google Scholar]
- His, W. (1874). Ueber die Bildung des Lachs‐Embryo. Sitzungsberichte der Naturforschenden Gesellschaft zu Leipzig 1874, 30. [Google Scholar]
- His, W. (1875). Unsere Körperform und das physiologische Problem ihrer Entstehung. Vogel, Leipzig. [Google Scholar]
- His, W. (1876). Die Entwicklungsgeschichte der Unke, als Grundlage einer vergleichenden Morphologie der Wirbelthiere. Von Dr. Alexander Goette. Referirt und besprochen von W. His. Zeitschrift für Anatomie und Entwickelungsgeschichte 1, 298–306; 465–472. [Google Scholar]
- His, W. (1877a). Neue Untersuchungen über die Bildung des Hühnerembryo. Archiv für Anatomie und Physiologie 1877, 112–187. [Google Scholar]
- His, W. (1877b). Ueber die Bildung der Haifischembryonen. Zeitschrift für Anatomie und Entwickelungsgeschichte 2, 108–124. [Google Scholar]
- His, W. (1879). Ueber die Anfänge des peripherischen Nervensystems. Archiv für Anatomie und Entwickelungsgeschichte 1879, 455–482. [Google Scholar]
- His, W. (1880). Anatomie menschlicher Embryonen I: Embryonen des ersten Monats (Text; with separate Atlas, plates I–VIII). Vogel, Leipzig. [Google Scholar]
- His, W. (1882a). Anatomie menschlicher Embryonen II: Gestalt‐ und Grössenentwicklung bis zum Schluss des 2. Monats [Text]. Vogel, Leipzig. [Google Scholar]
- His, W. (1882b). Die Lehre vom Bindesubstanzkeim (Parablast). Archiv für Anatomie und Entwickelungsgeschichte. 1882, 62–108. [Google Scholar]
- His, W. (1883). His, W. Über das Auftreten der weissen Substanz und der Wurzelfasern am Rückenmark menschlicher Embryonen. Archiv für Anatomie und Entwickelungsgeschichte 1883, 163–170. [Google Scholar]
- His, W. (1885). Anatomie menschlicher Embryonen III: Zur Geschichte der Organe. (Text; with separate Atlas, plates IX‐IV and I*). Vogel, Leipzig. [Google Scholar]
- His, W. (1886). Zur Geschichte des menschlichen Rückenmarkes und der Nervenwurzeln. Abhandlungen der Mathematisch‐Physischen Klasse der Königlich‐Sächsischen Gesellschaft der Wissenschaften 13, 479–543. [Google Scholar]
- His, W. (1887). Über die Methoden der plastischen Rekonstruktion und über deren Bedeutung für Anatomie und Entwickelungsgeschichte. Anatomischer Anzeiger 2, 382–394. [Google Scholar]
- His, W. (1888a). On the Principles of Animal Morphology. By Professor Wilhelm His of Leipzig. Letter to Mr John Murray, V.P.R.S.Ed. Proceedings of Royal Society of Edinburgh 15, 287–298. [Google Scholar]
- His, W. (1888b). Zur Geschichte des Gehirns sowie der centralen und peripherischen Nervenbahnen beim menschlichen Embryo. Abhandlungen der Königlich‐Sächsischen Gesellschaft der Wissenschaften 24, 341–392. [Google Scholar]
- His, W. (1889). Die Neuroblasten und deren Entstehung im embryonalen Mark. Abhandlungen der Mathematisch‐Physischen Klasse der Königlich‐Sächsischen Gesellschaft der Wissenschaften 15, 314–372. [Google Scholar]
- His, W. (1893a). Ueber das frontale Ende des Gehirnrohres. Archiv für Anatomie und Entwickelungsgeschichte 1893a, 157–171. [Google Scholar]
- His, W. (1893b). Vorschläge zur Eintheilung des Gehirns. Archiv für Anatomie und Entwickelungsgeschichte 1893b, 172–179. [Google Scholar]
- His, W. (1894). Über mechanische Grundvorgänge thierischer Formenbildung. Archiv für Anatomie und Entwickelungsgeschichte 1894, 1–80. [Google Scholar]
- His, W. (1895a). Anatomische Forschungen über Johann Sebastian Bach's Gebeine und Antlitz. Abhandlungen der Mathematisch‐Physischen Klasse der Königlich‐Sächsischen Gesellschaft der Wissenschaften 22, 381–420. [Google Scholar]
- His, W. (1895b). Die anatomische Nomenclatur. Nomina anatomica: Verzeichniss der von der anatomischen Gesellschaft auf ihrer IX. Versammlung in Basel angenommenen Namen. Veit, Leipzig. [Google Scholar]
- His, W. (1895c). Zum Gedächtnis an Carl Ludwig. Berichte über die Verhandlungen der Königlich‐Sächsischen Gesellschaft der Wissenschaften zu Leipzig . Mathematisch‐Physische Classe 47, 629–636. [Google Scholar]
- His, W. (1897a). Address upon the development of the brain. Transactions of the Royal Academy of Medicine in Ireland 1897, 3–21. [Google Scholar]
- His, W. (1897b). Friedrich Miescher. Die Histochemischen und Physiologischen Arbeiten. Vogel, Leipzig. [Google Scholar]
- His, W. (1898). Carl Ludwig and Carl Thiersch. Popular Science Monthly 52, 338–353. [Google Scholar]
- His, W. (1900). Lecithoblast und Angioblast der Wirbelthiere; Histogenetische Studien. Abhandlungen der Mathematisch‐Physischen Klasse der Königlich‐Sächsischen Gesellschaft der Wissenschaften 26, 172–326. [Google Scholar]
- His, W. (1903). Lebenserinnerungen. Als Manuskript gedruckt, Leipzig. [Google Scholar]
- His, W. (1904). Die Entwickelung des menschlichen Gehirns, während der ersten Monate: Untersuchungsergebnisse. Hirzel, Leipzig. [Google Scholar]
- His, W. & Rütimeyer, L. (1864). Crania Helvetica. Sammlung schweizerischer Schädelformen. Georg, Basel. [Google Scholar]
- His (Jr), W. (1931). Wilhelm His, der Anatom; ein Lebensbild. Urban & Schwarzenberg, Berlin. [Google Scholar]
- Hochstetter, F. (1919). Beiträge zur Entwicklungsgeschichte des menschlichen Gehirns. Deuticke, Vienna and Leipzig. [Google Scholar]
- Holtfreter, J. (1943). A study of the mechanics of gastrulation. Part I. Journal of Experimental Zoology 94, 261–318. [Google Scholar]
- Hopwood, N. (1999). “Giving body” to embryos. Modeling, mechanism, and the microtome in late nineteenth‐century anatomy. Isis 90, 462–496. [DOI] [PubMed] [Google Scholar]
- Hopwood, N. (2000). Producing development: the anatomy of human embryos and the norms of Wilhelm His. Bulletin of the History of Medicine 74, 29–79. [DOI] [PubMed] [Google Scholar]
- Hopwood, N. (2002). Embryos in Wax; Models from the Ziegler Studio. Whipple Museum of the History of Science, University of Cambridge; and The Institute of the History of Medicine, University of Bern, Cambridge and Bern. [Google Scholar]
- Hopwood, N. (2005). Visual standards and disciplinary change: normal plates, tables and stages in embryology. History of Science 43, 239–303. [Google Scholar]
- Hopwood, N. (2007). A history of normal plates, tables and stages in vertebrate embryology. International Journal of Developmental Biology 51, 1–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hopwood, N. (2012). A marble embryo: meanings of a portrait from 1900. History Workshop Journal 73, 5–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hopwood, N. (2013). Anatomist and embryo: a portrait sculpture. Lancet 381, 286–287. [DOI] [PubMed] [Google Scholar]
- Hopwood, N. (2015). Haeckel's Embryos: Images, Evolution, and Fraud. University of Chicago Press, Chicago. [Google Scholar]
- Hoßfeld, U. & Olsson, L. (2003). The road from Haeckel: the Jena tradition in evolutionary morphology and the origins of “evo‐devo”. Biology and Philosophy 18, 285–307. [Google Scholar]
- Huxley, J. & de Beer, G. (1963). The Elements of Experimental Embryology. Hafner Publishing Company, London. [Google Scholar]
- Iskratsch, T. , Wolfenson, H. & Sheetz, M. P. (2014). Appreciating force and shape‐the rise of mechanotransduction in cell biology. Nature Reviews Molecular Cell Biology 15, 825–833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacobson, M. (1983). Clonal organization of the central nervous system of the frog. III. Clones stemming from individual blastomeres of the 128‐, 256‐, and 512‐cell stages. Journal of Neuroscience 3, 1019–1038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacobson, M. (2005). Beginnings of the nervous system. In Developmental Neurobiology (Volume 4, eds Rao M. S. and Jacobson M.), pp. 365–413. Kluwer Academic/Plenum, New York. [Google Scholar]
- Jansen, K. A. , Donato, D. M. , Balcioglu, H. E. , Schmidt, T. , Danen, E. H. & Koenderink, G. H. (2015). A guide to mechanobiology:where biology and physics meet. Biochimica et Biophysica Acta 1853, 3043–3052. [DOI] [PubMed] [Google Scholar]
- Jeffery, J. E. , Bininda‐Emonds, O. R. , Coates, M. I. & Richardson, M. K. (2005). A new technique for identifying sequence heterochrony. Systematic Biology 54, 230–240. [DOI] [PubMed] [Google Scholar]
- Kästner, I. (2005). Der Anatom Wilhelm His. Ärzteblatt Sachsen 2005, 27–28. [Google Scholar]
- Katschenko, N. (1888). Zur Entwicklungsgeschichte des Selachierembryos. Anatomischer Anzeiger 3, 445–467. [Google Scholar]
- Keibel, F. (1897). In Normentafeln zur Entwicklungsgeschichte der Wirbelthiere (ed. Keibel F.). Verlag von Gustav Fischer, Jena. [Google Scholar]
- Keibel, F. (1899). Zu Mehnerts Bemerkungen über meine Kritiken und Referate. Anatomische Hefte 12, 567–573. [Google Scholar]
- Keibel, F. (1906). Die Entwickelung der äusseren Körperform der Wirbeltierembryonen, insbesondere der menschlichen Embryonen aus den ersten 2 Monaten. In Handbuch der vergleichenden und experimentellen Entwickelungslehre der Wirbelthiere, vol. 1 part 2 (ed. Hertwig O.), pp. 1–176. Gustav Fischer, Jena. [Google Scholar]
- Keibel, F. & Mall, F. P. (1910). Manual of Human Embryology. Lippincott, Philadelphia. [Google Scholar]
- Keller, R. E. (2002). Shaping the vertebrate body plan by polarized embryonic cell movements. Science 298, 19501954. [DOI] [PubMed] [Google Scholar]
- Keller, R. , Shih, J. , Wilson, P. & Sater, A. (1991). Pattern and function of cell motility and cell interactions during convergence and extension in Xenopus . In Cell‐Cell Interactions in Early Development (ed. Gerhart J.), pp. 31–62. Wiley‐Liss, New York. [Google Scholar]
- Keller, R. , Shook, D. & Skoglund, P. (2008). The forces that shape embryos: physical aspects of convergent extension by cell intercalation. Physical Biology 5, 015007. [DOI] [PubMed] [Google Scholar]
- Kerszberg, M. & Wolpert, L. (2007). Specifying positional information in the embryo: looking beyond morphogens. Cell 130, 205–209. [DOI] [PubMed] [Google Scholar]
- Kessel, R. G. (1960). The role of cell division in gastrulation of Fundulus heteroclitus . Experimental Cell Research 20, 277–282. [DOI] [PubMed] [Google Scholar]
- Keyte, A. L. & Smith, K. K. (2014). Heterochrony and developmental timing mechanisms: changing ontogenies in evolution. Seminars in Cell & Developmental Biology 34, 99–107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kingsbury, B. F. (1920). The extent of the floor‐plate of His and its significance. Journal of Comparative Neurology 32, 113–135. [Google Scholar]
- Klebs, E. (1867). Bemerkungen über Larynx‐Geschwülste. Archiv für pathologische Anatomie und Physiologie und für klinische Medicin 38, 202–220. [Google Scholar]
- Klebs, E. (1869). Die Einschmelzungs‐Methode, ein Beitrag zur mikroskopischen Technik. Archiv für Mikroskopische Anatomie 5, 164–166. [Google Scholar]
- Klebs, E. (1889). Die allgemeine Pathologie. Fischer, Jena. [Google Scholar]
- Klingenberg, C. P. (1998). Heterochrony and allometry: the analysis of evolutionary change in ontogeny. Biological Reviews 73, 79–123. [DOI] [PubMed] [Google Scholar]
- Klingenberg, C. P. (2016). Size, shape, and form: concepts of allometry in geometric morphometrics. Development Genes and Evolution 226, 113–137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kölliker, A. (1853). Manual of Human Histology by A. Kölliker. Translated and Edited by George Busk and Thomas Huxley. Sydenham Society, London. [Google Scholar]
- Kölliker, A. (1861). Entwicklungsgeschichte des Menschen und der höheren Thiere: Akademische Vorträge. Engelmann, Leipzig. [Google Scholar]
- Kölliker, A. (1876). Entwicklungsgeschichte des Menschen und der höheren Thiere, Second Edition. Engelmann, Leipzig. [Google Scholar]
- Kölliker, A. (1880). Grundriss der Entwickelungsgeschichte des Menschen und der höheren Tiere. Für Studierende und Ärzte. Engelmann, Leipzig. [Google Scholar]
- Kölliker, A. (1899). Erinnerungen aus meinem Leben. W. Engelmann, Leipzig. [Google Scholar]
- Kollmann, J. (1884). Der Randwulst und der Ursprung der Stützsubstanz. Archiv für Anatomie und Entwickelungsgeschichte 1884, 341–434. [Google Scholar]
- Kollmann, J. (1904). Wilhelm His. Worte der Erinnerung gesprochen am Begräbnistage den 4. Mai 1904. Verhandlungen der Naturforschenden Gesellschaft in Basel 15, 434–464. [Google Scholar]
- Kopsch, F. (1904). Untersuchungen über Gastrulation and Embryobildung bei den Chordaten I. Die morphologische Bedeutung des Keimhautrandes und die Embryobildung bei der Forelle. Thieme, Leipzig. [Google Scholar]
- Kunz, Y. W. (2004). Developmental Biology of Teleost Fishes. Springer, Dordrecht. [Google Scholar]
- Kurz, H. (1992). Wilhelm His (Basel und Leipzig): Seine Beiträge zur Weltgeltung der Anatomie im 19. Jahrhundert. Anatomisches Museum Basel, Basel. [Google Scholar]
- Laubichler, M. D. , Aird, W. C. & Maienschein, J. (2007). The endothelium in history. In Endothelial Biomedicine, pp. 5–19. Cambridge University Press, Cambridge. [Google Scholar]
- Lawson, K. A. , Meneses, J. J. & Pedersen, R. A. (1991). Clonal analysis of epiblast fate during germ layer formation in the mouse embryo. Development 113, 891–911. [DOI] [PubMed] [Google Scholar]
- Le Douarin, N. M. & Kalcheim, C. (1999). The Neural Crest. Cambridge University Press, Cambridge. [Google Scholar]
- Lehmstedt, M. (2003). Leipzig brennt: der Untergang des alten Leipzig am 4. Dezember 1943 in Fotografien und Berichten. Lehmstedt, Leipzig. [Google Scholar]
- Lereboullet, M. (1863). Recherches sur les monstruosités du brochet. Annales des Sciences Naturelles (Zoologie) 20, 177–271. [Google Scholar]
- Lewis, F. T. (1916). Charles Sedgwick Minot. Anatomical Record 10, 133–164. [Google Scholar]
- Lonie, I. M. (1981). The Hippocratic Treatises “On Generation”, On the Nature of the Child, “Diseases IV”. A Commentary. De Gruyter, Berlin. [Google Scholar]
- Lopez‐Munoz, F. , Boya, J. & Alamo, C. (2006). Neuron theory, the cornerstone of neuroscience, on the centenary of the Nobel Prize award to Santiago Ramon y Cajal. Brain Research Bulletin 70, 391–405. [DOI] [PubMed] [Google Scholar]
- Lotze, H. (1851). Von der Mechanik der Gestaltbildung. In Allgemeine Physiologie des koerperlichen Lebens. Book 2, Chapter 3, pp. 292–355. Weidmannsche Buchhandlung, Leipzig. [Google Scholar]
- Ludwig, C. (1852). Lehrbuch der Physiologie des Menschen, Edition (Volume 1), p. 458. Winter, Heidelberg. [Google Scholar]
- Ludwig, C. (1856). Lehrbuch der Physiologie des Menschen, Edition (Volume 2). Winter, Heidelberg. [Google Scholar]
- M.F (1875). His on morphological causation. Nature 12, 328. [Google Scholar]
- Maas, O. (1903). Einführung in die experimentelle Entwicklungsgeschichte (Entwicklungsmechanik). Bergmann, Wiesbaden. [Google Scholar]
- Maienschein, J. (1978). Cell lineage, ancestral reminiscence, and the biogenetic law. Journal of the History of Biology 11, 129–158. [Google Scholar]
- Maienschein, J. (1981). Shifting assumptions in american biology: embryology, 1890–1910. Journal of the History of Biology 14, 89–113. [DOI] [PubMed] [Google Scholar]
- Maienschein, J. (1991). The Origins of Entwicklungsmechanik. In A Conceptual History of Modern Embryology (ed. Gilbert S. F.), pp. 43–61. Springer, Boston. [Google Scholar]
- Mall, F. P. (1905). Wilhelm His. His relation to institutions of learning. American Journal of Anatomy 4, 139–161. [Google Scholar]
- Malpighi, M. (1673). Dissertatio Epistolica de Formatione Pulli in Ovo. Royal Society of London, London. [Google Scholar]
- Männer, J. (2004). On rotation, torsion, lateralization, and handedness of the embryonic heart loop: new insights from a simulation model for the heart loop of chick embryos. Anatomical Record Part A 278, 481–492. [DOI] [PubMed] [Google Scholar]
- Marchand, F. (1905). Wilhelm His: Nekrolog gesprochen in der öffentlichen Gesamtsitzung beider Klassen am 14. November 1904. Berichte über die Verhandlungen der Königlich Sächsischen Gesellschaft der Wissenschaften zu Leipzig. Mathematisch‐Physische Classe 56, 326–340. [Google Scholar]
- Marlétaz, F. , Firbas, P. N. , Maeso, I. , Tena, J. J. , Bogdanovic, O. , Perry, M. , Wyatt, C. D. R. , de la Calle‐Mustienes, E. , Bertrand, S. , Burguera, D. , Acemel, R. D. , van Heeringen, S. J. , Naranjo, S. , Herrera‐Ubeda, C. , Skvortsova, K. , et al. (2018). Amphioxus functional genomics and the origins of vertebrate gene regulation. Nature 564, 64–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marrese, M. , Antonovaite, N. , Nelemans, B. K. A. , Smit, T. H. & Iannuzzi, D. (2019). Micro‐indentation and optical coherence tomography for the mechanical characterization of embryos: experimental setup and measurements on chicken embryos. Acta Biomaterialia 97, 524–534. [DOI] [PubMed] [Google Scholar]
- Marshall, A. M. (1877). The early stages of development of the nerves in birds. Journal of Anatomy and Physiology 11, 491–515. [PMC free article] [PubMed] [Google Scholar]
- Marshall, A. M. (1878). The development of the cranial nerves in the chick. Quarterly Journal of Microscopical Science 18, 10–40. [Google Scholar]
- Marshall, A. M. (1879). Memoirs: the morphology of the vertebrate olfactory organ. Quarterly Journal of Microscopical Science 19(New Series, 300–340. [Google Scholar]
- Mehnert, E. (1898). Biomechanik: Erschlossen aus dem Principe der Organogenese. Fischer, Jena. [Google Scholar]
- Mehnert, E. (1899). Bemerkungen zu Keibels Kritiken und Referaten. Eine Replikation. Anatomische Hefte 12, 549–565. [Google Scholar]
- Miescher, F. (1871). Ueber die chemische Zusammensetzung der Eiterzellen. Medicinisch‐chemische Untersuchungen 4, 441–460. [Google Scholar]
- Minot, C. S. (1897). Human Embryology. Macmillan, New York. [Google Scholar]
- Minot, C. S. (1890). The mesoderm and the coelom of vertebrates. The American Naturalist 24, 877–898. [Google Scholar]
- Moore, S. W. , Roca‐Cusachs, P. & Sheetz, M. P. (2010). Stretchy proteins on stretchy substrates: the important elements of integrin‐mediated rigidity sensing. Developmental Cell 19, 194–206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morgan, T. H. (1895). The formation of the fish embryo. Journal of Morphology 10, 419–472. [Google Scholar]
- Morgan, T. H. (1897). The Development of the Frog's Egg. Macmillan, New York. [Google Scholar]
- Morgan, T. H. (1927). Experimental Embryology. Columbia University Press, New York. [Google Scholar]
- Müller, F. & O'Rahilly, R. (1986). Wilhelm His und 100 Jahre Embryologie des Menschen. Acta Anatomica 125, 73–75. [PubMed] [Google Scholar]
- Nieuwenhuys, R. & Puelles, L. (2016). Towards a new neuromorphology. In The Natural Coordinate System of the CNS (eds Nieuwenhuys R. and Puelles L.), pp. 132–140. Springer, Cham. [Google Scholar]
- Nissl, F. (1903). Die Neuronenlehre und ihre Anhänger. Fischer, Jena. [Google Scholar]
- Nyhart, L. K. (1995). Biology Takes Form: Animal Morphology and the German Universities, 1800–1900. University of Chicago Press, Chicago. [Google Scholar]
- Oken, L. & Kieser, D. G. (1806). Beiträge zur Vergleichenden Zoologie, Anatomie und Physiologie. Bamberg, Würzburg. [Google Scholar]
- Olsson, L. , Levit, G. S. & Hossfeld, U. (2017). The “Biogenetic Law” in zoology: from Ernst Haeckel's formulation to current approaches. Theory in Biosciences 136, 19–29. [DOI] [PubMed] [Google Scholar]
- Oppenheimer, J. M. (1955). Problems, concepts and their history. In Analysis of Development (eds Willier B. H., Weiss P. A. and Hamburger V.), pp. 1–24. Saunders, Philadelphia. [Google Scholar]
- O'Rahilly, R. & Müller, F. (1988). Der Vesal der Embryologie des Menschen. Anatomischer Anzeiger 166, 245–247. [PubMed] [Google Scholar]
- O'Rahilly, R. (1979). Early human development and the chief sources of information on staged human embryos. European Journal of Obstetrics, Gynecology, and Reproductive Biology 9, 273–280. [DOI] [PubMed] [Google Scholar]
- Orr, H. A. (1887). Contribution to the embryology of the lizard. Journal of Morphology 1, 311–372. [Google Scholar]
- Oschatz, A. F. (1844). Ueber Herstellung und Aufbewahrung mikroskopischer Präparate. In Beiträge zur physiologischen und pathologischen Chemie und Mikroskopie in ihrer Anwendung auf die praktische Medizin (Volume 1, ed. Simon F.), pp. 128–140. Hirschwald, Berlin. [Google Scholar]
- Oxford Dictionaries (2017). Oxford English Dictionary (OED online). Oxford University Press, Oxford. [Google Scholar]
- Paluch, E. K. , Nelson, C. M. , Biais, N. , Fabry, B. , Moeller, J. , Pruitt, B. L. , Wollnik, C. , Kudryasheva, G. , Rehfeldt, F. & Federle, W. (2015). Mechanotransduction: use the force(s). BioMed Central (BMC) Biology 13, 1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pander, C. H. (1817). Beiträge zur Entwickelungsgeschichte des Hühnchens im Eye. Würzburg. [Google Scholar]
- Picken, L. (1956). The fate of Wilhelm His. Nature 178, 1162–1165. [DOI] [PubMed] [Google Scholar]
- Poelmann, R. E. & Gittenberger‐de Groot, A. C. (2018). Hemodynamics in cardiac development. Journal of Cardiovascular Development and Disease 5, 1–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prévost, J.‐L. & Dumas, J.‐B. (1824). Deuxième mémoire sur la génération. Annales des Sciences Naturelles 2, 100–121; 129–149. [Google Scholar]
- Prud'homme, B. , Gompel, N. & Carroll, S. B. (2007). Emerging principles of regulatory evolution. Proceedings of the National Academy of Sciences of the United States of America 104, 8605–8612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Psychoyos, D. & Stern, C. D. (1996). Fates and migratory routes of primitive streak cells in the chick embryo. Development 122, 1523–1534. [DOI] [PubMed] [Google Scholar]
- Puelles, L. (2018). Developmental studies of avian brain organization. International Journal of Developmental Biology 62, 207–224. [DOI] [PubMed] [Google Scholar]
- Puelles, L. , Domenech‐Ratio, G. & Martinez‐De‐La‐Torre, M. (1987). Location of the rostral end of the longitudinal brain axis: review of an old topic in the light of marking experiments on the closing rostral neuropore. Journal of Morphology 194, 163–171. [DOI] [PubMed] [Google Scholar]
- Quint, M. , Drost, H. G. , Gabel, A. , Ullrich, K. K. , Bonn, M. & Grosse, I. (2012). A transcriptomic hourglass in plant embryogenesis. Nature 490, 98–101. [DOI] [PubMed] [Google Scholar]
- Rabl, C. (1889). Theorie des Mesoderms. Morphologisches Jahrbuch 15, 113–252. [Google Scholar]
- Rabl, C. (1897). Theorie des Mesoderms, Edition (Volume 1). Engelmann, Leipzig. [Google Scholar]
- Rabl, C. (1909a). Das Anatomische Institut. Hirzel, Leipzig. [Google Scholar]
- Rabl, C. (1909b). Geschichte der Anatomie an der Universität Leipzig. Barth, Leipzig. [Google Scholar]
- Rabl, C. (1915). Édouard van Beneden und der gegenwärtige Stand der wichtigsten von ihm behandelten Probleme. Archiv für Mikroskopische Anatomie 88, 3–470. [Google Scholar]
- Rádl, E. (1909). Geschichte der biologischen Theorien in der Neuzeit. II. Teil: Geschichte der Entwicklungstheorien in der Biologie des XIX. Jahrhunderts. Engelmann, Leipzig. [Google Scholar]
- Raff, R. A. (2000). Evo‐devo: the evolution of a new discipline. Nature Reviews Genetics 1, 74–79. [DOI] [PubMed] [Google Scholar]
- Rasmussen, A. T. (1970). Wilhem His (1831–1904). In The Founders of Neurology: One Hundred and Forty‐six Biographical Sketches by Eighty‐Eight Authors (eds Haymaker W. and Schiller F.), pp. 48–51. Thomas, Springfield. [Google Scholar]
- Rathke, H. (1825). Kiemen bey Vogeln. Isis von Oken 1825, 1100–1101. [Google Scholar]
- Rauber, A. (1875). Embryonale Anlage des Hühnchens. III. Ursprung des Mesoderms. Centralblatt fur die medicinischen Wissenschaften 13, 257–259. [Google Scholar]
- Rauber, A. (1876). Über die Stellung des Hühnchens im Entwicklungsplan. Engelmann, Leipzig. [Google Scholar]
- Rauber, A. (1877). Primitivstreifen und Neurula der Wirbelthiere, in normaler und pathologischer Beziehung. Engelmann, Leipzig. [Google Scholar]
- Rawitz, B. (1904). Wilhelm His. Nachruf. Naturwissenschaftliche Rundschau 19, 308–310. [Google Scholar]
- Rawles, M. E. (1936). A study in the localization of organ‐forming areas in the chick blastoderm of the head‐process stage. Journal of Experimental Zoology 72, 271–315. [Google Scholar]
- Reichert, K. B. (1840). Das Entwickelungsleben im Wirbelthier‐Reich. Hirschwald, Berlin. [Google Scholar]
- Reichert, K. B. (1843). Beiträge zur Kenntniss des Zustandes der heutigen Entwickelungs‐Geschichte. Hirschwald, Berlin. [Google Scholar]
- Reichert, K. B. (1861). Der Bau des menschlichen Gehirns: Durch Abbildungen mit erläuterndem Texte dargestellt. Engelmann, Leipzig. [Google Scholar]
- Remak, R. (1855). Untersuchungen über die Entwicklung der Wirbelthiere. Reimer, Berlin. [Google Scholar]
- Rhumbler, L. (1902). Zur Mechanik des Gastrulationsvorganges insbesondere der Invagination. Eine entwickelungsmechanische Studie. Archiv für Entwicklungsmechanik der Organismen 14, 401–476. [Google Scholar]
- Richards, R. J. (2008). The Tragic Sense of Life: Ernst Haeckel and the Struggle over Evolutionary Thought. University of Chicago Press, Chicago. [Google Scholar]
- Richardson M. K. (2009). The Hox Complex ‐ an interview with Denis Duboule. The International Journal of Developmental Biology, 53(5‐6), 717–723. 10.1387/ijdb.072558mr [DOI] [PubMed] [Google Scholar]
- Richardson, M. K. (2012). A phylotypic stage for all animals? Developmental Cell 22, 903–904. [DOI] [PubMed] [Google Scholar]
- Richardson, M. K. , Jeffery, J. E. , Coates, M. I. & Bininda‐Emonds, O. R. (2001). Comparative methods in developmental biology. Zoology (Jena) 104, 278–283. [DOI] [PubMed] [Google Scholar]
- Richardson, M. K. & Keuck, G. (2001). A question of intent: when is a 'schematic' illustration a fraud? Nature 410, 144. [DOI] [PubMed] [Google Scholar]
- Richardson, M. K. & Keuck, G. (2002). Haeckel's ABC of evolution and development. Biological Reviews 77, 495–528. [DOI] [PubMed] [Google Scholar]
- Risau, W. & Flamme, I. (1995). Vasculogenesis. Annual Review of Cell and Developmental Biology 11, 73–91. [DOI] [PubMed] [Google Scholar]
- Roguin, A. (2006). Wilhelm His Jr. (1863–1934) ‐ the man behind the bundle. Heart Rhythm 3, 480–483. [DOI] [PubMed] [Google Scholar]
- Rosen, G. (1973). Ludwig, Carl Friedrich Wilhelm. In Dictionary of Scientific Biography (Volume 8, ed. Gillispie C. C.), pp. 540–542. Scribner's, New York. [Google Scholar]
- Rossello‐Mora, R. & Amann, R. (2001). The species concept for prokaryotes. Federation of European Microbiological Societies (FEMS) Microbiology Reviews 25, 39–67. [DOI] [PubMed] [Google Scholar]
- Roux, W. (1895a). Gesammelte Abhandlungen über Entwickelungsmechanik der Organismen. Abhandlung XIII–XXXIII: Über Entwickelungsmechanik des Embryo. Wilhelm Engelmann, Leipzig. [Google Scholar]
- Roux, W. (1895b). The problems, methods, and scope of developmental mechanics: an introduction to the “Archiv für Entwickelungsmechanik der Organismen”. In Biological Lectures Delivered at the Marine Biological Laboratory in Woods Hole, pp. 149–190. Ginn & Company, Boston. [Google Scholar]
- Roux, W. (1896). Über die Bedeutung geringer Verschiedenheiten der relativen Grösse der Furchungszellen für den Charakter des Furchungsschemas. Archiv für Entwicklungsmechanik der Organismen 4, 1–74. [Google Scholar]
- Runnström, J. (1914). Analytische Studien Über die Seeigelentwicklung. I. Archiv für Entwicklungsmechanik der Organismen 40, 526–564. [Google Scholar]
- Sabin, F. R. (1913). The origin and development of the lymphatic system. The Johns Hopkins Hospital Reports. Monographs. New Series No. 5, 1–94.
- Sabin, F. R. (1934). Biographical Memoir of Franklin Paine Mall. Biographical Memoirs. National Academy of Sciences 16, 65–122. [Google Scholar]
- Sander, K. (1991). Landmarks in developmental biology. Wilhelm Roux and his programme for developmental biology. Roux's Archives of Developmental Biology 200, 1–3. [DOI] [PubMed] [Google Scholar]
- Schwendener, S. (1878). Mechanische Theorie der Blattstellungen. Engelmann, Leipzig. [Google Scholar]
- Sheesley, P. , McMenamin, M. , Kusyk, J. & Pivar, S. (2014). The morphogenic mapping of the brain and the design of the nervous system. International Journal of Brain Science 2014, 7. [Google Scholar]
- Shindo, A. (2018). Models of convergent extension during morphogenesis. Wiley Interdisciplinary Reviews. Developmental biology 7, 1–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith, B. G. (1914). An experimental study of concrescence in the embryo of Cryptobranchus allegheniensis . Biological Bulletin 26, 245–261. [Google Scholar]
- Snarr, B. S. , Wirrig, E. E. , Phelps, A. L. , Trusk, T. C. & Wessels, A. (2007). A spatiotemporal evaluation of the contribution of the dorsal mesenchymal protrusion to cardiac development. Developmental Dynamics 236, 1287–1294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sneath, P. H. & Sokal, R. R. (1962). Numerical taxonomy. Nature 193, 855–860. [DOI] [PubMed] [Google Scholar]
- Spalteholz, W. (1906). His, Wilhelm. In Biographisches Jahrbuch und deutscher Nekrolog (1904) (Volume 9), pp. 231–242. Reimer, Berlin. [Google Scholar]
- Spemann, H. (1938). Embryonic Development and Induction (1967 reprint). Hafner, New York. [Google Scholar]
- Spitzer, H. (1886). Beiträge zur Descendenztheorie und zur Methodologie der Naturwissenschaft. Brockhaus, Leipzig. [Google Scholar]
- Stekelenburg‐de Vos, S. , Ursem, N. T. , Hop, W. C. , Wladimiroff, J. W. , Gittenberger‐de Groot, A. C. & Poelmann, R. E. (2003). Acutely altered hemodynamics following venous obstruction in the early chick embryo. Journal of Experimental Biology 206, 1051–1057. [DOI] [PubMed] [Google Scholar]
- Stephan, C. N. (2015a). Accuracies of facial soft tissue depth means for estimating ground truth skin surfaces in forensic craniofacial identification. International Journal of Legal Medicine 129, 877–888. [DOI] [PubMed] [Google Scholar]
- Stephan, C. N. (2015b). Facial approximation‐from facial reconstruction synonym to face prediction paradigm. Journal of Forensic Sciences 60, 566–571. [DOI] [PubMed] [Google Scholar]
- Stephens, T. D. (1982). The Wolffian ridge: history of a misconception. Isis 73, 254–259. [DOI] [PubMed] [Google Scholar]
- Stern, C. D. (2006). Evolution of the mechanisms that establish the embryonic axes. Current Opinion in Genetics & Development 16, 413–418. [DOI] [PubMed] [Google Scholar]
- Stern, C. D. (2004). Gastrulation: From Cells to Embryo. Cold Spring Harbor laboratory Press, Cold Spring Harbor. [Google Scholar]
- Stricker, S. (1872). Handbuch der Lehre von den Geweben des Menschen und der Thiere, Edition (Volume 2). Engelmann, Leipzig. [Google Scholar]
- Swalla, B. J. (2002). The evolution of developmental pathways. Science 296, 2147–2148. [Google Scholar]
- Swanson, L. W. (2015). Neuroanatomical Terminology. Oxford University Press, Oxford. [Google Scholar]
- Taber, L. A. , Lin, I. E. & Clark, E. B. (1995). Mechanics of cardiac looping. Developmental Dynamics 203, 42–50. [DOI] [PubMed] [Google Scholar]
- Tallinen, T. , Chung, J. Y. , Rousseau, F. , Girard, N. , Lefèvre, J. & Mahadevan, L. (2016). On the growth and form of cortical convolutions. Nature Physics 12, 588–593. [Google Scholar]
- Tena, J. J. , Gonzalez‐Aguilera, C. , Fernandez‐Minan, A. , Vazquez‐Marin, J. , Parra‐Acero, H. , Cross, J. W. , Rigby, P. W. , Carvajal, J. J. , Wittbrodt, J. , Gomez‐Skarmeta, J. L. & Martinez‐Morales, J. R. (2014). Comparative epigenomics in distantly related teleost species identifies conserved cis‐regulatory nodes active during the vertebrate phylotypic period. Genome Research 24, 1075–1085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson, D. W. (1910). In Historia Animalum. The Works of Aristotle (Volume 4, eds Smith J. A. and Ross W. D.), p. 633. The Clarendon Press, Oxford. [Google Scholar]
- Thompson, D. W. (1917). On Growth and Form, p. 793. Cambridge University Press, Cambridge. [Google Scholar]
- Thompson, D. W. (1945). On Growth and Form. Cambridge University Press, Cambridge. [Google Scholar]
- Tickle, C. & Urrutia, A. O. (2017). Perspectives on the history of evo‐devo and the contemporary research landscape in the genomics era. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 372, 1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torrey, T. W. (1967). Morphogenesis of the Vertebrates. Wiley, New York. [Google Scholar]
- Tubbs, R. S. , Loukas, M. , Shoja, M. M. & Cohen‐Gadol, A. A. (2009). Wilhelm His (1831–1904) and his contributions to neuroanatomy. Child's Nervous System 25, 1613–1615. [DOI] [PubMed] [Google Scholar]
- Vaihinger, K. (1941). Die Bewegungsmechanik der Spaltöffnungen. Protoplasma 36, 430–443. [Google Scholar]
- Valentin, G. (1845). Repertorium für Anatomie und Physiologie: Kritische Darstellung fremder und Ergebnisse eigener Forschung. Huber, Bern. [Google Scholar]
- Van Beneden, E. (1878). A contribution to the history of the embryonic development of the teleosteans. Quarterly Journal of Microscopical Science (New Series) 18, 41–57. [Google Scholar]
- Van der Lem, T. , de Bakker, M. , Keuck, G. & Richardson, M. K. (2021). Wilhelm His Senior und die Entwicklung der Paraffineinbettung. Der Pathologe 42, 424–430. An English translation is published online at:. 10.1007/s00292-021-00947-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Meulen, M. C. , Beaupre, G. S. & Carter, D. R. (1993). Mechanobiologic influences in long bone cross‐sectional growth. Bone 14, 635–642. [DOI] [PubMed] [Google Scholar]
- Van Essen, D. (1997). A tension‐based theory of morphogenesis and compact wiring in the central nervous system. Nature 385, 313–318. [DOI] [PubMed] [Google Scholar]
- Vanezis, M. & Vanezis, P. (2000). Cranio‐facial reconstruction in forensic identification‐historical development and a review of current practice. Medicine, Science, and the Law 40, 197–205. [DOI] [PubMed] [Google Scholar]
- Varner, V. D. & Taber, L. A. (2012). Not just inductive: a crucial mechanical role for the endoderm during heart tube assembly. Development 139, 1680–1690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vernon, K. (1988). The founding of numerical taxonomy. The British Journal for the History of Science 21, 143–159. [Google Scholar]
- Virchow, R. (1852). Die Identität von Knochen‐, Knorpel‐ und Bindegewebskörperchen, sowie über Schleimgewebe. Verhandlungen der Physikalisch‐Medizinischen Gesellschaft in Würzburg 2, 150–160. [Google Scholar]
- Virchow, R. (1856). Gesammelte Abhandlungen zur Wissenschaftlichen Medicin von Rudolph Virchow. Meidinger, Frankfurt. [Google Scholar]
- Virchow, R. (1859). Die Cellularpathologie in ihrer Begründung auf physiologische und pathologische Gewebelehre, Second Edition. Hirschwald, Berlin. [PubMed] [Google Scholar]
- Vogt, W. (1929). Gestaltungsanalyse am Amphibienkeim mit örtlicher Vitalfärbung. II. Teil. Gastrulation und Mesodermbildung bei Urodelen und Anuren. Wilhelm Roux' Archiv für Entwicklungsmechanik der Organismen 120, 384–706. [DOI] [PubMed] [Google Scholar]
- Voit, C. (1905). Wilhelm His. Sitzungsberichte der mathematisch‐physikalischen Klasse der Königlich Bayerischen Akademie der Wissenschaften 35, 328–337. [Google Scholar]
- Vollmer, H. (1980). Allgemeines Lexikon der bildenden Künstler von der Antike bis zur Gegenwart. Seemann, Lepzig. [Google Scholar]
- von Baer, K. E. (1828). Über Entwickelungsgeschichte der Thiere: Beobachtung und Reflexion. Erster Theil. Bornträger, Königsberg. [Google Scholar]
- von Baer, K. E. (1837). Über Entwickelungsgeschichte der Thiere: Beobachtung und Reflexion. Zweiter Theil. Bornträger, Königsberg. [Google Scholar]
- von Baer, K. E. & Oppenheimer, J. M. (1986). Autobiography of Karl Ernst von Baer. Science History Publications, Canton. [Google Scholar]
- Wagner, R. (1841). Elements of Physiology: For the Use of Students and with Especial Reference to the Wants of Practitioners (transl. Willis, R.A.). Sherwood, Gilbert, & Piper, London. [Google Scholar]
- Waldeyer, W. (1870). Eierstock und Ei: ein Beitrag zur Anatomie und Entwicklungsgeschichte der Sexualorgane. Engelmann, Leipzig. [Google Scholar]
- Waldeyer, W. (1883). Archiblast und Parablast. Archiv für Mikroskopische Anatomie 22, 1–77. [Google Scholar]
- Waldeyer, W. (1891). Ueber einige neuere Forschungen im Gebiete der Anatomie des Centralnervensystems. Thieme, Leipzig. [Google Scholar]
- Waldeyer, W. (1904). Wilhelm His. Sein Leben und Wirken. Deutsche Medizinische Wochenschrift 30, 1438–1439, 1469–1471, 1509–1511. [Google Scholar]
- Waldeyer‐Hartz, W. (1921). Lebenserinnerungen von Wilhelm von Waldeyer‐Hartz, Second Edition. Cohen, Bonn. [Google Scholar]
- Wall, M. , Butler, D. , El Haj, A. , Bodle, J. C. , Loboa, E. G. & Banes, A. J. (2018). Key developments that impacted the field of mechanobiology and mechanotransduction. Journal of Orthopaedic Research 36, 605–619. [DOI] [PubMed] [Google Scholar]
- Weismann, A. P. , Parker, W. N. & Rönnfeldt, H. (1893). The Germ‐Plasm: A Theory of Heredity (English Transl.). Scribner's, New York. [Google Scholar]
- Weiss, P. A. (1955). Nervous System (Neurogenesis). In Analysis of Development (eds Willier B. H., Weiss P. A. and Hamburger V.), pp. 346–401. Saunders, Philadelphia. [Google Scholar]
- Welcker, H. (1883). Schiller's Schädel und Todtenmaske, nebst Mittheilungen über Schädel und Todtenmaske Kant's. Vieweg und Sohn, Braunschweig. [Google Scholar]
- Wendler, D. & Rother, P. (1982). Wilhelm His Senior‐the life and work of the important Leipzig morphologist. Zeitschrift für die gesamte innere Medizin und ihre Grenzgebiete 37, 810–813. [PubMed] [Google Scholar]
- Wilson, E. B. (1893). Amphioxus, and the mosaic theory of development. Journal of Morphology 8, 579–638. [Google Scholar]
- Wilson, E. B. (1895). The embryological criterion of homology. Biological Lectures Delivered at the Marine Biological Laboratory of Wood's Holl 1894, 101–124. [Google Scholar]
- Wilson, E. B. (1904). Experimental studies on germinal localization. Journal of Experimental Zoology 1, 1–72. [Google Scholar]
- Wolff, C. F. (1768). De formatione intestinorum (parts 1 and 2). Novi commentari Academiae Scientiarum Imperialis Petropolitanae 12, 403–507. [Google Scholar]
- Wolff, C. F. (1812). Über die Bildung des Darmkanals im bebrüteten Hühnchen. Transl. J. F. Meckel. Rengersche Buchhandlung, Halle. [Google Scholar]
- Wolpert, L. (1994). Do we understand development? Science 266, 571–572. [DOI] [PubMed] [Google Scholar]
- Wolpert, L. (1969). Positional information and the spatial pattern of cellular differentiation. Journal of Theoretical Biology 25, 1–47. [DOI] [PubMed] [Google Scholar]
- Wolpert, L. (2009). Diffusible gradients are out ‐ an interview with Lewis Wolpert. Interviewed by Richardson, Michael K. International Journal of Developmental Biology 53, 659–662. [DOI] [PubMed] [Google Scholar]
- Woodger, J. H. (1929). Biological Principles: A Critical Study. Routledge and Kegan Paul, London. [Google Scholar]
- Woollam, D. H. M. & Millen, J. W. (1954). Perivascular spaces of the mammalian central nervous system. Biological Reviews 29, 251–283. [Google Scholar]
- Yanai, I. (2018). Development and evolution through the lens of global gene regulation. Trends in Genetics 34, 11–20. [DOI] [PubMed] [Google Scholar]
- Zegers, R. H. , Maas, M. , Koopman, A. T. & Maat, G. J. (2009). Are the alleged remains of Johann Sebastian Bach authentic? The Medical Journal of Australia 190, 213–216. [DOI] [PubMed] [Google Scholar]
- Zhou, J. , Kim, H. Y. & Davidson, L. A. (2009). Actomyosin stiffens the vertebrate embryo during crucial stages of elongation and neural tube closure. Development 136, 677–688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zimmer, H. G. (1996). Carl Ludwig: The man, his time, his influence. Pflügers Archiv 432, R9–R22. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Fig. S1. Keys to sitters in the group photographs shown in Fig. 2 of the main text.
Fig. S2. Bibliographic analysis of the research themes of Wilhelm His.
Table S1. Wilhelm His, Sr. – selected biographical landmarks.
Table S2. Glossary of terms and concepts relevant to this review.
Table S3. Opinions of earlier researchers on the origins of the peripheral ganglia and the nephric duct.
Table S4. Models used by His and others to explain and understand developmental processes.
Table S5. Table of Contents of ‘Our Bodily Form’ (His, 1875) with chapter summaries.
Supplementary Note S1. Additional information on the oil painting of Wilhelm His shown in Fig. 1D.
Supplementary Note S2. Contributions of Wilhelm His Sr. to anthropology and forensic craniofacial reconstruction.
Supplementary Note S3. Contributions of Wilhelm His Sr. to developmental neurobiology.
Supplementary Note S4. The conflict between Wilhelm His and Ernst Haeckel.
Supplementary Note S5. Other polemics and disputes.
Supplementary Note S6. Scientific opinion on the parablast and concrescence theories.
Supplementary Note S7. His's ‘mechanical’ developmental biology: its meaning and its reception.
Supplementary Note S8. Mosaic versus regulatory development.