

Abstract
Mature granulated trophoblast binucleate cells (BNC) have been found in all ruminant placentas examined histologically so far. BNC are normally fairly evenly distributed throughout the fetal villus and all their granules contain a similar variety of hormones and pregnancy associated glycoproteins (PAGs). Only the Giraffe is reported to show a different BNC protein expression, this paper is designed to investigate that. Gold labelled Lectin histochemistry and protein immunocytochemistry were used on deplasticised 1 μm sections of a wide variety of ruminant placentomes with a wide range of antibodies and lectins. In the Giraffe placentomes, even though the lectin histochemistry shows an even distribution of BNC throughout the trophoblast of the placental villi, the protein expression in the BNC granules is limited to the BNC either in the apex or the base of the villi. Placental lactogens and Prolactin (PRL) are present only in basally situated BNC: PAGs only in the apical BNC. PRL is only found in the Giraffe BNC which react with many fewer of the wide range of antibodies used here to investigate the uniformity of protein expression in ruminant BNC. The possible relevance of these differences to ruminant function and evolution is considered to provide a further example of the versatility of the BNC system.
Keywords: Ruminant placenta, Trophoblast binucleate cell, Giraffe granule protein asymmetry
Introduction
Mature granulated trophoblast BNC have been found in all ruminant placentas examined histologically so far [1–3]. Recent investigations suggest that all mature BNC undergo the same characteristic migration out of the trophoblast followed by fusion with a uterine epithelial cell or derivative to form fetomaternal tissue throughout pregnancy [4]. The granules are released to the maternal compartment by exocytosis from this fetomaternal tissue. BNC are normally fairly evenly distributed throughout the fetal villus and all their granules contain a similar variety of hormones and pregnancy associated glycoproteins (PAGs) [1].
The one exception to this uniformity so far is the giraffe, (Giraffa camelopardalis) which is reported to show BNC whose granules react uniquely with a Prolactin antibody but are restricted to the base of the fetal villi [2].
The giraffoid clade now consists only of Giraffes, Okapis and Pronghorns [5] but they were much more widespread in the Miocene era [6]. They are now restricted to a few species in marked comparison with the enormous expansion of the other ruminant clades, in particular the Bovidae. Fortunately samples of Okapi and Pronghorn placentas were available, as well as a wide variety of other ruminant placentas [Supplementary Table S1].
Antibodies to Prolactin and purified Placental lactogens from three different species were used (Table 1) together with examples from the two phylogenetically distinct ancient and modern groups of PAGs [7, 14].
Table 1.
Antibodies and Lectins used
| Abbreviations | Antibody | Antibody origins | |
|---|---|---|---|
| “ancient”PAGs | |||
| RA | Anti native bovine-PAGs | Wooding et al 2005 [13] | |
| RD | Anti-boPAG 2 | ||
| BJ | Anti-boPAG 2 | Beckers et al 1994 [18] | R438 |
| “new”PAGs | |||
| RB | Anti native bo-PAGs | Wooding et al 2005 [13] | |
| RC | Anti –ovinePAG 1 | ||
| RU | |||
| FPIR | Anti bo PAG 1 | Touzard et al 2013 [14] | |
| BG | Anti-bo PAG 1 | Zoli et al 1991 [15] | R726 |
| Buffalo PAGs | |||
| BK | Anti-wbPAGs | Barbato et al 2013 [19] | R858 |
| BM | Anti-wbPAGs (different N terminal sequence) | R859 | |
| Goat PAGs | |||
| BB | Anti-capPAG55 + 59 kDa | Garbayo et al 1998 [16] | R708 |
| BE | Anti-capPAG55 + 62 kDa | R706 | |
| Ovine PAG | Anti-ovPAG57 | El-Amiri et al 2004 [20] | R780 |
| BN | Anti-bovine PAG 1 | Zoli et al 1991 [15] | |
| Placental Lactogens | |||
| Friesen oPL | Anti ovinePL | Chan et al 1978 [22] | |
| BF | Anti- bovine PL | Alvarez-Oxiley et al 2007 [17] | |
| PROLACTIN | Anti humanPRL | NIH | |
| SBU3 | Monoclonal ab, Homologous to ovine PAG | Gogolin-Ewens et al,1987 [21] | |
| LECTINS | Vector Laboratories, | Peterboro, UK | |
| DSA | |||
| DBA | |||
| ePHA | |||
| lPHA | |||
This paper reports a qualitative and quantitative study of ruminant BNC position and granule content in the species detailed above using the range of antibodies and lectins.
This variety of results should facilitate investigation of the possible relevance of any differences to ruminant function and evolution.
Methods and materials
Animals
Mid to late pregnant placental material was used from a wide variety of ruminants collected over many years (Supplemental Table S1 and see Wooding references in [1]). They were fixed by aldehyde immersion or perfusion.
Wild animals were shot as part of Wild life management or culling procedures and placentomes removed and immersion fixed within 20 minutes of the death of the animals. Fixatives used included Bouins, phosphate buffered Paraformaldehyde or Glutaraldehyde, and Surgipath (methyl alcohol and formaldehyde). The quality of fixation varied, but crucially, all produced comparable results with the antibodies and lectins (Table 1) used on the placentome sections.
Small pieces of Horse Anterior Pituitary were also fixed and processed as the placentome samples.
For details of the animals and origins see Supplemental Table S1.
At least two placentomes from a single animal were used in the case of the Wildebeest, at least two or more animals from each of the other species were used.
A central slice was cut from each fixed placentome. “Matchstick” samples from the central region of each slice from maternal to fetal edge were used. The samples were then embedded in epoxy resin with no osmium postfixation.
Semithin sections were cut, picked up on cover glass squares treated with APES, deresinated in sodium ethoxide and thoroughly washed in PBS.
Immunocytochemistry
The cover glass squares were then floated section side down on drops of antibody (see Table 1) followed by immunogold colloid (goat anti-rabbit G5, Jackson Immunoresearch Labs, USA) and then intensified with silver reagent (Aurion, Wagenigen, Netherlands). The coverslips were washed thoroughly between each incubation.
The antibodies used (Table 1) were to purified lactogens or Pregnancy Associated Glycoproteins (PAGs) from a variety of species (see Supplementary Table S1) used at a dilution of 1:1000.
Lectin histochemistry
The cover glass squares were floated section side down on drops of biotinylated lectins (see Table 1) followed by immunogold colloid.
(Goat antibiotin G5) and then intensified with silver reagent (Aurion, Wagenigen, Netherlands). The lectins used were chosen as the most BNC reactive although the silver enhancement did produce some background with the giraffe specimens.
The antibodies and lectins identified BNC granules in all species used. Controls with buffer substituted for antibody or lectin showed no significant labelling.
Assessment of Labeling
Visual estimate of antibody reaction on BNC granules, scale used: —, no reactivity, (+) sporadic/occasional: +, ++, +++ indicate increasing levels of reactivity.
Quantitation of BNC
The number of BNC visible in the X10 objective field of view was counted at random positions along the axis of the fetomaternal “matchstick” from the fetal to maternal side.
The area of the field of view was converted to mm 2 and the position along the “matchstick” estimated by eye.
The results for any particular lectin or antibody are consistent but the standard deviations are high mainly because of the variable areas of core villus connective tissue on the sections. To minimise this problem semiserial sections were used for any one comparative run.
Results
Our results show a remarkable degree of uniformity in the binucleate cell granule immunoreactivity in thirteen of the fourteen ruminant species we examined (Tables 2 and 4, Supplementary Fig. S1, Fig. 1).
Table 2.
Assessment of the immunoreactivity of non giraffoid Ruminant BNC
| ANTIBODY | COW | EWE | BISON | Tragulus | Springbok | Impala | Red deer | Chinese water deer | Wildebeest | White tail | Wapiti |
|---|---|---|---|---|---|---|---|---|---|---|---|
| “ancient”PAGs | |||||||||||
| RA | +++ | +++ | ++ | +++ | +++ | + | +++ | ++ | +++ | +++ | |
| RD | +++ | +++ | + | +++ | ++ | ++ | +++ | ||||
| BJ | ++ | ++ | + | ++ | + | +++ | |||||
| “new”PAGs | |||||||||||
| RB | +++ | +++ | +++ | +++ | ++ | +++ | + | +++ | +++ | +++ | |
| RC | + | +++ | +++ | +++ | ++ | ++ | + | +++ | +++ | +++ | |
| RU | ++ | +++ | +++ | ++ | ++ | +++ | +++ | +++ | +++ | +++ | |
| FPIR | +++ | ++ | +++ | ++ | ++ | +++ | |||||
| BC | ++ | ++ | ++ | +++ | ++ | + | + | +++ | +++ | ||
| BO | ++ | +++ | ++ | +++ | +++ | + | ++ | ++ | |||
| Buffalo PAGs | |||||||||||
| BK | ++ | ++ | +++ | +++ | + | + | ++ | ++ | ++ | +++ | +++ |
| BM | ++ | +++ | +++ | +++ | ++ | +++ | +++ | +++ | |||
| Goat PAGs | |||||||||||
| BB | +++ | +++ | +++ | +++ | +++ | + | +++ | ++ | +++ | +++ | +++ |
| BE | +++ | +++ | +++ | ++ | +++ | + | ++ | +++ | +++ | +++ | |
| Ovine PAG | |||||||||||
| BN | +++ | +++ | +++ | +++ | +++ | + | ++ | + | +++ | +++ | |
| SBU3 | ++ | ++ | ++ | ++ | + | ++ | ++ | ++ | |||
| Placental Lactogens | |||||||||||
| Friesen bPL | +++ | ++ | +++ | − | ++ | ++ | ++ | ++ | ++ | ||
| BF | ++ | ++ | +++ | − | − | + | ++ | + | +++ | + | +++ |
| PROLACTIN | − | − | − | − | − | − | − | − | − | − | − |
| SBU3 | ++ | ++ | ++ | ++ | + | ++ | ++ | ++ | |||
| LECTINS | |||||||||||
| DSA | ++ | ++ | +++ | ++ | |||||||
| DBA | +++ | +++ | +++ | +++ | +++ | +++ | |||||
| ePHA | +++ | ||||||||||
| lPHA | +++ |
Scale used: —, no reactivity, +, ++, +++ indicate increasing levels of reactivity. Empty oblong: assessment not done
Table 4.
Quantitation of the number of immunoreactive BNC per mm2 in four non giraffoid Ruminants
| VILLUS COUNT AREA | |||
|---|---|---|---|
| SPECIES | ANTIBODY OR LECTIN | FETAL END 0 -50% n = 5 | MATERNAL END 50 – 100% n = 5 |
| COW(Bos taurus) | BB | 80 ± 12 | 85 ± 11 |
| BC | 90 ± 6 | 95 ± 9 | |
| BF | 90 ± 12 | 85 ± 10 | |
| BG | 88 ± 10 | 85 ± 15 | |
| BK | 88 ± 15 | 80 ± 10 | |
| DBA | 45 ± 8 | 50 ± 11 | |
| EWE (Ovis aries) | RA | 68 ± 9 | 65 ± 10 |
| RB | 36 ± 8 | 40 ± 11 | |
| ovinePL | 28 ± 10 | 30 ± 8 | |
| bovinePL | 30 ± 8 | 32 ± 10 | |
| TRAGULUS spp | RU | 48 ± 14 | 50 ± 10 |
| PHA | 61 ± 9 | 69 ± 10 | |
| PRONGHORN (Antilocapra americana) | 97 ± 12 | 84 ± 15 | |
| RU | 78 ± 11 | 69 ± 12 | |
| DSA | 68 ± 7 | 70 ± 7 |
No significant differences were detected between fetal and maternal end counts
Figure 1.
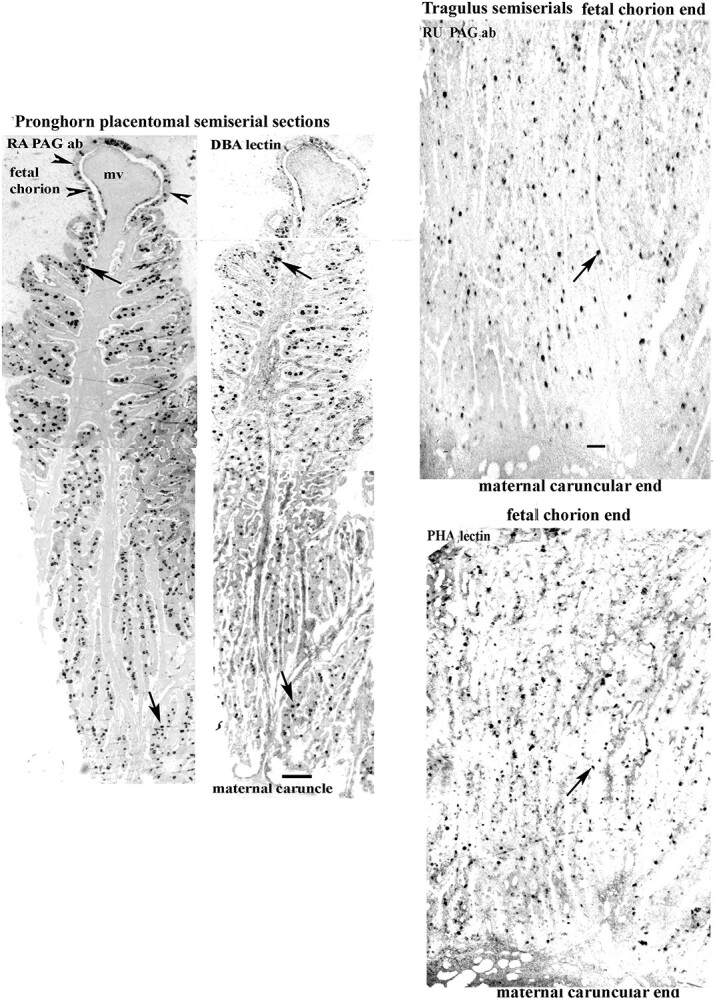
Pronghorn and Tragulus; placentomal semiserial sections showing similar localisations and even distributions of BNCs throughout the sections using PAG antibody or lectin localisations. Maternal villus, mv. Arrows indicate different levels of the same BNCs on the pairs of sections. Arrowheads identify the fetal basal chorionic layer on the Pronghorn. Scale bar = 250 μm. Tragulus; Scale bar = 80 μm.
We used a wide range of Pregnancy Associated Glycoproteins (PAGs) and Placental Lactogen antibodies (Table 1) raised against proteins purified from ruminant placentas of different species. The BNC granules were detected by gold labelled immunocytochemistry by most of the antibodies and lectins used on deplasticised araldite sections which included the full depth of the placentomal villi from maternal to fetal sides.
In the thirteen, BNC were evenly distributed throughout the fetal villi with no consistent indication of any zonation (Tables 2 and 4, Supplementary Fig. S1, Fig. 1).
There have been reports of differences in zonation [eg, 3], but there are no previous quantitative studies confirming them.
All the BNC granules on any one section showed similar levels of expression of PAGs or lactogens or lectin reactivity, although the levels of expression differed between the various species and the PAGs, lactogens and lectins (Tables 3 and 5, Figs 1, 2, 3).
Table 3.
The immunoreactivity of the BNC in the Giraffoid clade
| GIRAFFE | OKAPI | PRONGHORN | |||||
|---|---|---|---|---|---|---|---|
| ANTIBODY | Different Placentomes | Different animal | Different Placentomes | Different animal | |||
| “ancient”PAGs | G1A | G1B | G2 | P1 | P1G | P6 | |
| RA | (+) | (+) | (+) | +++ | ++++ | ++ | ++ |
| RD | − | − | − | +++ | +++ | ++ | +++ |
| BJ | − | +Mend | +Mend | ++ | ++ | +++ | ++ |
| “new”PAGs | |||||||
| RB | − | − | − | + | + | ++ | ++ |
| RC | − | +Mend | − | + | ++ | ++ | ++ |
| RU | ++Mend only | +++M end | ++Mend | + | + | + | + |
| FPIR | − | − | − | + | ++ | ++ | ++ |
| BC | − | − | − | − | +++ | ++ | ++ |
| BO | − | − | − | ++ | + | +++ | |
| Buffalo PAGs | − | ||||||
| BK | − | +Mend | − | + | ++ | ++ | ++ |
| BM | − | − | − | − | − | − | − |
| Goat PAGs | |||||||
| BB | ++Mend | ++Mend | − | +++ | ++ | ++ | ++ |
| BE | ++Mend | ++Mend | +++Mend | ++ | +++ | +++ | ++ |
| Ovine PAG | |||||||
| BN | ++Mend | ++Mend | ++Mend | − | ++ | + | +++ |
| SBU3 | − | − | − | − | − | − | − |
| Placental Lactogens | |||||||
| Friesen bPL | ++Fend | ++Fend | +++Fend | − | − | − | − |
| BF | ++Fend | ++Fend | ++Fend | + | − | − | − |
| PROLACTIN | ++Fend | ++Fend | ++Fend | − | − | − | − |
| LECTINS | |||||||
| DSA | +++ | +++ | +++ | ++ | +++ | +++ | |
| DBA | + | + | + | + | + | ||
| ePHA | ++ | + | ++ | +++ | +++ | ||
| lPHA | ++ | ++ | ++ | ++ | |||
Scale used: —, no reactivity, (+) sporadic/occasional: +, ++, +++ indicate increasing levels of reactivity. Empty oblong: assessment not done.
M/F ends: Maternal or Fetal ends of the Fetal villus
Table 5.
Quantitative estimates of the number of immunoreactive BNC cells per mm2 in Giraffe
| Antibody or Lectin | Villus count position | ||||
|---|---|---|---|---|---|
| 0 – 25% | 25% -50% | 50% -75% | 75%- 100% | ||
| Animal | Fetal end | Maternal end | |||
| Giraffe A | DSA | 61 ± 13 | 60 ± 11 | 63 ± 12 | 62 ± 14 |
| bovinePL | 27 ± 6 | 1 ± 2 | 1 ± 1 | 0 | |
| PRL | 29 ± 6 | 1 ± 1 | 1 ± 1 | 0 | |
| BF | 67 ± 2 | 27 ± 8 | 11 ± 2 | 5 ± 2 | |
| BE | 0 | 0 | 0 | 65 ± 16 | |
| RU | 0 | 0 | 0 | 60 ± 9 | |
| Giraffe B | DSA | 56 ± 9 | 58 ± 11 | 55 ± 11 | 56 ± 12 |
| bovinePL | 28 ± 1 | 3 ± 1 | 2 ± 1 | 1 ± 1 | |
| ovinePL | 14 ± 2 | 0 | 0 | 0 | |
| PRL | 24 ± 1 | 0 | 0 | 0 | |
| BF | 53 ± 4 | 29 ± 9 | 24 ± 9 | 21 ± 6 | |
| BE | 0 | 0 | 0 | 26 ± 6 | |
| RU | 0 | 0 | 0 | 27 ± 5 | |
| BB | 0 | 0 | 0 | 20 ± 5 | |
| BJ | 0 | 0 | 0 | 14 ± 7 | |
| BN | 0 | 0 | 0 | 15 ± 5 | |
No significant differences were detected between fetal and maternal end counts of the Lectins
Figure 2.
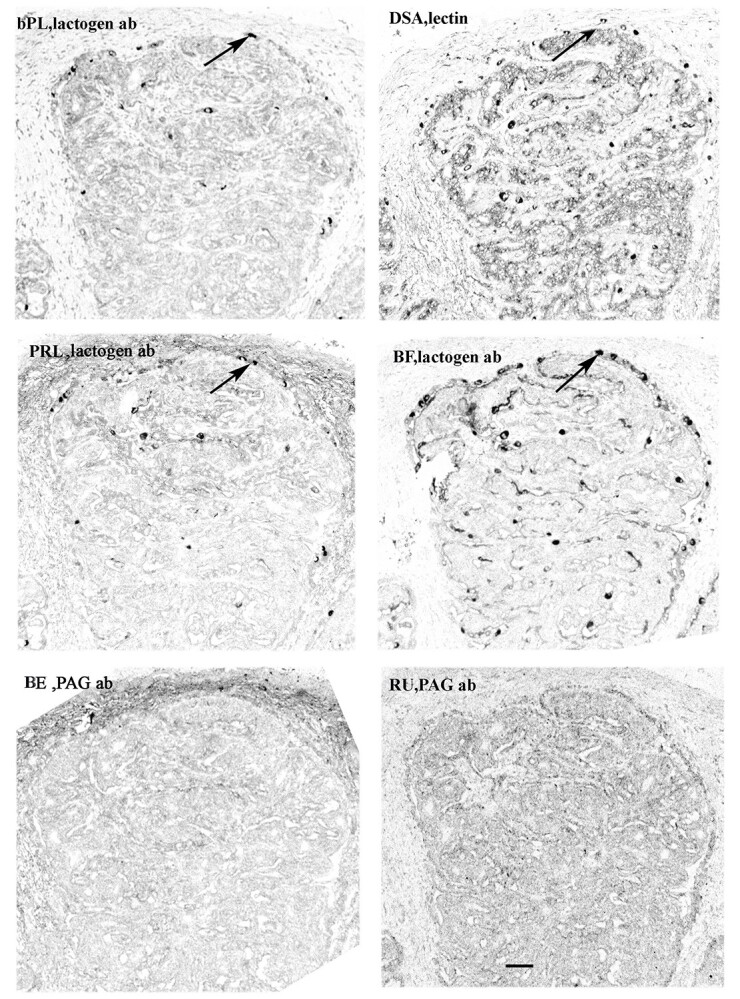
Giraffe placentomal fetal villus chorionic ends. Lactogen antibodies are present at the bases of the fetal chorionic villi (eg, at arrows); no BNCs can be identified using PAG antibody; BNCs are localised throughout the villi the using lectin. Arrows indicate different levels of probably the same BNCs on the sections using the lactogen antibody or lectin localisation. Scale bar =50 μm.
Figure 3.
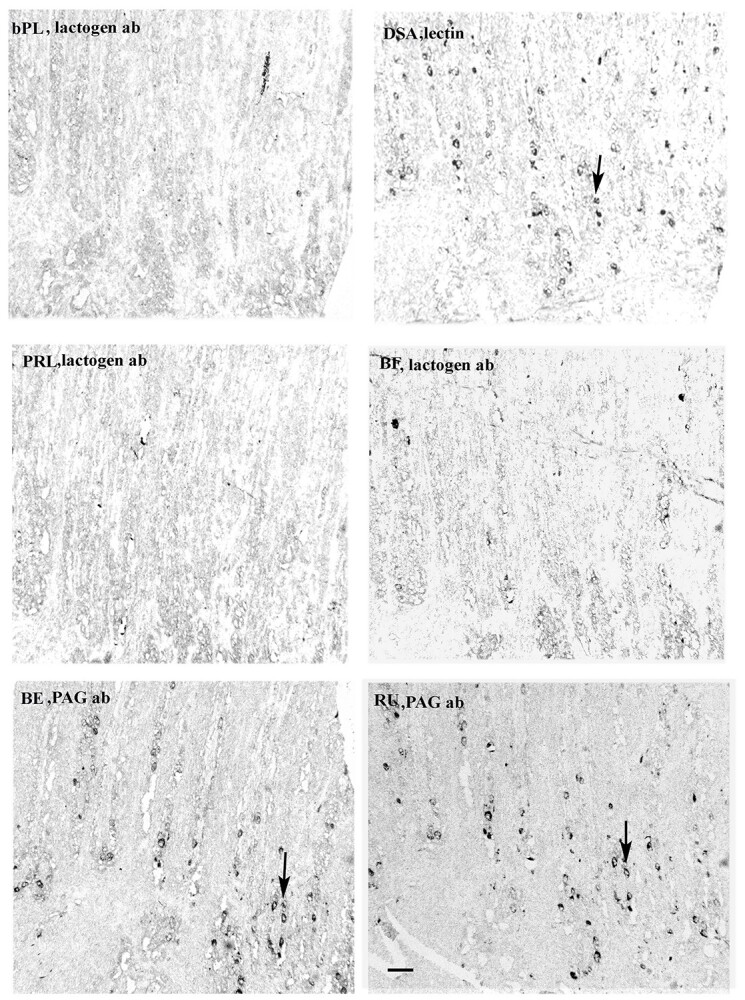
Giraffe placentomal fetal villus caruncular ends. PAG antibodies are present at the apices of the fetal chorionic villi (eg, at arrows); no BNCs can be identified using lactogen antibody; BNCs are localised throughout the villi using lectin. Arrows indicate BNCs in very similar positions using the PAG antibody or lectin localisation. Scale bar =50 μm.
In sharp contrast to the uniform immunoreactive BNC distribution in the 13 species described above, the Giraffe immunoreactive BNC distribution is unique. In this species the placental BNC granules react with many fewer of the antibodies used and when they do react it is in strictly localised areas, either the top or bottom 25% of the fetal villus length (Tables 3 and 5). This is more difficult to recognise when the entire length of the Giraffe sections are shown on Supplemental Fig. S2 and better appreciated at higher magnification on Figs. 2, 3).
All the antibodies expressed in the top 25% are against PAGs whereas all the basal group are anti PLs or Prolactin. The prolactin expression is unique to Giraffe, and it seems definite as the antibody can be absorbed against NIH prolactin on both placentome and Horse anterior pituitary sections (Supplementary Fig. S3).
One of the PLs (BF) does show a wider distribution than the top 25% of the villus but with a rapidly decreasing frequency towards the maternal end.
However localising giraffe BNC with the most strongly reacting lectin clearly demonstrates a uniform distribution throughout the fetal villi (Tables 3 and 4, Figs. 2, 3).
The two other members of the giraffoid clade, Okapi (Okapi johnstoni) and Pronghorn (Antilocapra americana) show no zonation of immunoreactive BNC positioning and their BNC react with many more of the antibodies than the giraffe. (Table 3) They also show individual differences compared to the giraffe – no Prolactin or placental lactogen expression for example.
Discussion
This paper establishes the asymmetry of the PAG and the PRL/lactogen distributions in the BNC granule content of the Giraffe placentomal villi compared with the uniformity of BNC granule content in the placentomal villi of all the other ruminant groups investigated so far. This asymmetry is also in contrast to the uniform distribution of the BNC in the Giraffe placentomal villi as clearly indicated by the lectin content of the BNC granules.
The Giraffe BNC granules react with many fewer of the antibodies used here but do show a unique content of PRL. This does not seem to be an artefact as shown by the ability to absorb the reactivity with pure PRL in both the giraffe placentome and the horse anterior pituitary sections (Supplementary Fig. S3).
There is also a clear difference in type at the two ends of the fetal villi with PRL and placental lactogens at the base and a range of PAGs at the top.
These peculiarities are not shared by the other members of the giraffoid clade, Okapi and Pronghorn, both of which show similar distribution of BNC granule immunoreactivity to the ruminant majority, although there are individual differences such as no reaction with the SBU3 or one of the Buffalo PAG antibodies.
It is perhaps of interest that the only other placental PRL trophoblast localisation reported is in the Elephant (Loxodonta africana) but the localisation is throughout the epithelium [8] and not restricted to any specialised cells.
The lectins used in this investigation were chosen for maximum reactivity. There are differences in lectin reactivity between Giraffoids and the other ruminants which are detailed in our previous papers [9–11].
The typical uniform BNC distribution with the unique formation of fetomaternal tissue by migration and fusion was established early in evolution as exemplified by the Tragulid line [12], and probably was a key factor in the success of the ruminant grouping. All of the mature ruminant placentas so far investigated show this characteristic pattern as do the BNC of Giraffe and Pronghorn [4].
There is no obvious or apparent functional reason for the PAG asymmetry unless it is necessary to establish a high concentration of PAGs in the caruncle. This could camouflage the area from invasive maternal lymphocytes to maintain the fetomaternal balance. This strategy has been suggested to be the case in the development of the bovine placental villi but no BNC asymmetry was necessary to produce that [13].
PRL and lactogens are considered to play important roles in the fetal metabolism but there is no reason to think that the asymmetric localisation would speed up the delivery to the fetus.
We have shown previously [2] that the stimulation of the protein expression is very precisely localised since the villus base (arcade) chorion presumably stimulates expression of BNC PRL and the lactogens but no significant PAGs whereas BNC in the adjacent intercotyledonary chorion express PAGs but no PRL or lactogens.
Both giraffe placentomal samples show a very similar unique BNC distribution and protein expression so it is unlikely to be a methodological artefact. Also the even distribution of the lectins here (and other antibodies used previously [2]) throughout the giraffe sections support this assumption.
Maybe it shows the versatility of the BNC system in solving the problems of maternofetal immunological balance in the extreme evolutionary niche occupied by the giraffe.
Supplementary Material
{kind=link}
{kind=link}
{kind=link}
Acknowledgements
The authors are very grateful to the many individuals (see Supplementary Table S1) who provided the wide range of suitable Ruminant samples.
Footnotes
† Grant Support: This research did not receive any specific grant from any funding agency in the public, commercial or not-for-profit sector.
Contributor Information
F B P Wooding, The Physiological Laboratory, University of Cambridge, Downing Site, Cambridge, CB2 3EG.
A J Forhead, The Physiological Laboratory, University of Cambridge, Downing Site, Cambridge, CB2 3EG.
S Wilsher, The Paul Mellon Laboratory of Equine Reproduction, Newmarket, Suffolk, CB8 9BJ.
W R Allen, The Paul Mellon Laboratory of Equine Reproduction, Newmarket, Suffolk, CB8 9BJ.
R M Roberts, Division of Animal Sciences, University of Missouri, Columbia, Missouri, USA.
J A Green, Division of Animal Sciences, University of Missouri, Columbia, Missouri, USA.
J F Beckers, Physiologie de la Reproduction, Faculte de Medecine Veterinaire, B-4000, Liege, Belgique.
de N Melo Sousa, Physiologie de la Reproduction, Faculte de Medecine Veterinaire, B-4000, Liege, Belgique.
G Charpigny, INRA, UMR1198, Biologie Devel Reprod, F-78532 Jouy et Josas, France.
Funding
This research did not receive any specific grant from any funding agency in the public, commercial or not-for- profit sector.
Conflict of interest
There is no potential Conflict of Interest with any public or commercial organisations.
Data availability
The data underlying this article are available in the article and in its online supplementary material.
References
- 1. Wooding FBP, Burton G. Comparative Placentation: structures functions and evolution. Springer-Verlag; 2008. [Google Scholar]
- 2. Wooding FBP, Wilsher S, Benirschke K, Jones CBP, Allen WR. Immunocytochemistry of the placentas of giraffe (Giraffa cameleopardalis giraffa) and okapi (okapi johnstoni): comparison with other ruminants. Placenta 2015; 36:77–87. [DOI] [PubMed] [Google Scholar]
- 3. Wilsher S, Greenwood RES, Mahon GD, Allen WR. Placentation and hormonal maintenance of pregnancy in the impala (Aepyceros melampus). Placenta 2020; 95:91–105. [DOI] [PubMed] [Google Scholar]
- 4. Wooding FBP, Osborn D, Killian GJ. Trinucleate uterine epithelial cells as evidence for white-tail deer trophoblast binucleate cell migration and as markers of placental binucleate cell dynamics in a variety of wild ruminants. Placenta 2018; 62:34–42. [DOI] [PubMed] [Google Scholar]
- 5. Price SA, Bininda-Emonds ORP, Gittleman JL. A complete phylogeny of the whales, dolphins and even-toed hoofed mammals (Cetartiodactyla). Biol Rev 2005; 80:445–473. [DOI] [PubMed] [Google Scholar]
- 6. Clauss SS, Rössner GE. Old world ruminant Morphophysiology, life history, and fossil record: exploring key innovations of a diversification sequence. Ann Zool Fennici 2014; 51:80–94. [Google Scholar]
- 7. Wallace RM, Pohler KG, Smith MF, Green JA. Placental PAGs: gene origins, expression patterns and use as markers of pregnancy. Reproduction 2015; 149:R115–R126. [DOI] [PubMed] [Google Scholar]
- 8. Yamamoto Y, Yamamoto T, Taya K, Watanabe GJ, Stansfield FJ, Allen WR. Placentation in the African elephant (Loxodonta africana). V. the trophoblast secretes placental lactogen. Placenta 2011; 32:506–510. [DOI] [PubMed] [Google Scholar]
- 9. Jones CJ, Wilsher SA, Wooding FB, Benirschke K, Allen WR. The binucleate cell of okapi and giraffe placenta shows distinctive glycosylation compared with other ruminants: a lectin histochemical study. Mol Phylogenet Evol 2015; 83:184–190. [DOI] [PubMed] [Google Scholar]
- 10. Jones CJP, Silvia WJ, Hamilton CJ, Geary TW, Zezeski AL, Wooding FBP. Glycosylation and immunocytochemistry of binucleate cells in pronghorn (Antilocapra americana, Antilocapridae) show features of both Giraffidae and Bovidae. Placenta 2017; 57:216–222. [DOI] [PubMed] [Google Scholar]
- 11. Klisch K, Wooding FBP, Jones CJP. The glycosylation pattern of secretory granules in binucleate trophoblast cells is highly conserved in ruminants. Placenta 2010; 31:11–17. [DOI] [PubMed] [Google Scholar]
- 12. Wooding FBP, Kimura J, Forhead AJ. Functional immunocytochemistry of Tragulus placenta: implications for ruminant evolution. Placenta 2014; 35:305–310. [DOI] [PubMed] [Google Scholar]
- 13. Wooding FBP, Roberts RM, Green JA. Light and electron microscope immunocytochemical studies of the distribution of pregnancy associated glycoproteins (PAGs) throughout pregnancy in the cow: possible functional implications. Placenta 2005; 26:807–827. [DOI] [PubMed] [Google Scholar]
- 14. Touzard E, Reinaud P, Dubois O, Guyader-Joly C, Humblot P, Ponsart C, Charpigny G. Specific expression patterns and cell distribution of ancient and modern PAG in bovine placenta during pregnancy. Reproduction 2013; 146:347–362. [DOI] [PubMed] [Google Scholar]
- 15. Pagnah Zoli A, Beckers JF, Wouters-Ballman P, Closset J, Falmagne P, Ectors F. Purification and characterization of a bovine pregnancy-associated Glycoprotein1. Biol Reprod 1991; 45:1–10. [DOI] [PubMed] [Google Scholar]
- 16. Garbayo JM, Remy B, Alabart JL, Folch J, Wattiez R, Falmagne P, Beckers JF. Isolation and partial characterization of a pregnancy-associated glycoprotein family from the goat Placenta1. Biol Reprod 1998; 58:109–115. [DOI] [PubMed] [Google Scholar]
- 17. Alvarez-Oxiley AV, Sousa NM, Hornick JL, Touati K, van der Weijden GC, Taverne MA, Szenci O, Sulon J, Debliquy P, Beckers JF. Radioimmunoassay of bovine placental lactogen using recombinant and native preparations: determination of fetal concentrations across gestation. Reprod Fertil Dev 2007; 19:877–885. [DOI] [PubMed] [Google Scholar]
- 18. Beckers JF, Roberts RM, Zoli AP, Ectors F, Derivaux J. Molécules de la famille des protéases aspartiques dans le placenta des ruminants: hormones ou protéines? Bull Mém Acad Royale Méd Belg 1994; 149:355–367. [PubMed] [Google Scholar]
- 19. Barbato O, Melo de Sousa N, Barile VL, Canali L, Beckers JF. Purification of pregnancy-associated glycoproteins from late-pregnancy Bubalus bubalis placentas and development of a radioimmunoassay for pregnancy diagnosis in water buffalo females. BMC Vet Res 2013; 9:89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. el Amiri B, Remy B, Melo de Sousa N, Beckers JF. Isolation and characterization of eight pregnancy-associated glycoproteins present at high levels in the ovine placenta between day 60 and day 100 of gestation. Reprod Nutr Dev 2004; 44:169–181. [DOI] [PubMed] [Google Scholar]
- 21. Gogolin-Ewens KJ, Lee CS, Mercer WR, Moseby AM, Brandon MR. Characterization of a sheep trophoblast-derived antigen first appearing at implantation. Placenta 1986; 7:243–255. [DOI] [PubMed] [Google Scholar]
- 22. Chan JSD, Robertson HA, Friesen HG. The purification and characterization of ovine placental Lactogen. Endocrinology 1976; 98:65–76. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The data underlying this article are available in the article and in its online supplementary material.