

Abstract
This study was designed to observe the treatment effects of flavokawain B (FKB) on gastric cancer both in SGC-7901 cells and nude mice. When SGC-7901 cells were exposed to 10 μg/mL FKB, cellular proliferative and apoptotic capacities and cell cycle were detected utilizing CCK-8 and flow cytometry assays. The results showed that FKB treatment induced cell apoptosis and G2/M arrest and suppressed cell proliferation for SGC-7901 cells. Western blot results showed that FKB upregulated proapoptotic proteins as well as downregulated antiapoptotic and cell cycle-related proteins in SGC-7901 cells. SMAD4, TGF-β1, and TSPAN12 proteins were tested in FKB-induced SGC-7901 cells. Following exposure to FKB, SMAD4, TGF-β1, and TSPAN12 expression was augmented in SGC-7901 cells. si-SMAD4 transfection weakened cell apoptosis and accelerated cell proliferation. Furthermore, FKB reversed the change in apoptotic and cell cycle-related proteins induced by si-SMAD4. A nude mouse tumorigenesis model was constructed, which was treated by FKB. In the nude mouse tumorigenesis model, FKB activated the TSPAN12 expression and TGF-β1/SMAD4 pathway. Also, FKB treatment prolonged the survival time of nude mice and lowered tumor weight. iNOS and CD86 expression was significantly enhanced, and Arg-1 and CD206 expression was significantly decreased in THP-1 cells cultured in conditioned media from FKB-treated SGC-7901 cells. Additionally, FKB-treated SGC-7901 cells weakened macrophage migration. Collectively, this evidence suggested that FKB accelerated apoptosis and suppressed the proliferation of gastric cancer cells and attenuated M2 macrophage polarization, thereby exerting an anticancer effect on gastric cancer.
1. Introduction
Gastric cancer is a gastrointestinal malignant tumor with the highest incidence and fatality rate worldwide [1–3]. Surgical removal is the first choice for gastric cancer treatment [4, 5]. Most patients are already at an advanced stage and have distant metastases at the time of diagnosis [6]. The prognosis remains pessimistic, and 5-year survival rate is less than 20%. However, chemotherapy drugs usually cause severe side effects, such as liver toxicity [7], nephrotoxicity [8], and immunosuppression [9]. Therefore, the development and research of natural product anticancer drugs with low toxicity and high selectivity have become a research hotspot.
The occurrence of gastric cancer is in close relationship with the imbalance of apoptosis and proliferation [10, 11]. Previous research has found that Chinese medicine monomer can lead to cell cycle arrest at different stages, such as Cannabidiol [12], ethanolic extract of Cordyceps cicadae [13], and Phloretin [14]. Therefore, regulation of proliferation and apoptosis of cancer cells is a critical direction for gastric cancer therapy. Mammalian cell cycle progression can be mediated by various enzymes. It has been confirmed that cyclin-dependent protein kinase- (CDK-) cyclin complexes are activated in time of the cell cycle, which can be induced and mediated via environmental factors (ultraviolet rays, ionizing radiation, thermal damage, industrial chemicals, etc.) [15–17]. Apoptosis is the prime type of programmed cell death. During the process, cysteine-aspartic protease (caspase) family membranes are activated [18–20]. Therefore, the application of chemical and biological reagents to induce cell cycle arrest, reduce cell proliferation, and even induce cell apoptosis has been become a promising intervention for the therapy of gastric cancer.
Transforming growth factor- (TGF-) β is a polypeptide cytokine with multiple biological activities, which is one of the most important tumor suppressor pathways [21, 22]. Abnormal TGF-β/SMAD4 pathway contributes to tumorigenesis of gastric cancer [23]. Tetraspanin-12 belongs to the family of four transmembrane proteins, characterized by four transmembrane domains and two extracellular loops [24]. TSPAN12 has been found to participate to various cancers such as ovarian cancer [25], lung cancer [26], and colorectal cancer [27]. However, its expression and role in gastric cancer remain unclear. Flavokawain B (FKB) extracted from the rhizomes of Alpinia pricei Hayata, has a strong antiproliferative effect on a variety of cancer cells glioma [28], prostate cancer [29], breast cancer [30], thyroid cancer [31], etc. FKB can exert an antitumor biological activity through multiple mechanisms. For example, FKB could inhibit proliferative capacity and motivate cell cycle arrest in colon cancer cells [32]. Furthermore, FKB suppresses ROS-mediated apoptosis and autophagic cell death of lung adenocarcinoma cells [33]. However, the specific effect of FKB on gastric cancer remains unknown. Furthermore, whether FKB is related to the TGF-β/SMAD4 pathway is unclear.
Macrophages exert key roles in the initiation and progression of human cancers [34–36]. Tumor-associated macrophages exhibit enhanced plasticity as well as predominantly show as M2 phenotype, that is, in relation to cancer metastases and undesirable prognosis [37]. Thus, alleviating M2 macrophage polarization within the tumor microenvironment represents a promising therapeutic regimen against gastric cancer. Limited evidence indicates that both FKB and TGF-β1-SMAD4 signaling pathway mediate macrophage differentiation. For instance, FKB reduces LPS-triggered proinflammatory IL-6 secretion in macrophages [38]. Moreover, SMAD4 variant facilitates macrophage recruitment as well as inflammatory responses through activating TGF-β signaling in thoracic aortic aneurysm and dissection [39]. Herein, we hypothesized that FKB could expedite cell cycle arrest and apoptosis of gastric cancer cells via the TGF-β1-SMAD4 signaling pathway as well as weaken the induction capacity of SGC-7901 cells in macrophage polarization.
2. Materials and Methods
2.1. Cell Culture
Human gastric cancer cell line SGC-7901 was obtained from Chinese Academy of Sciences (Shanghai, China), which was grown in RPMI-1640 medium (Invitrogen, USA) plus 10% fetal bovine serum (FBS), penicillin (100 U/mL) and streptomycin (100 μg/mL). The cells were fostered in a humidified incubator containing 5% CO2 at 37°C.
2.2. Cell Transfection and Drug Treatment
SGC-7901 cells were seeded in a 6-well plate (2.5 × 105 cells/well) for 24 h. si-SMAD4 (sense: 5′-GCCACUGUAUCCAGCAGAUTT-3′; antisense: 5′-GCCACUGUAUCCAGCAGAUTT-3′) or negative control (NC) synthesized by RIBOBIO (Guangzhou, China) was transfected into cells via Lipofectamine 2000 (Applied Biosystems, Life technologies, USA). Following 48 h of transfection, the cells were harvested.
FKB solution (10 μg/mL; LKT Laboratories, Inc., St. Paul, MN, USA) was prepared in DMSO [33]. FKB was diluted to the concentration of 0.1% using DMSO. Then, SGC-7901 cells were exposed to FKB solution.
2.3. Cell Counting Kit-8 (CCK-8) Assay
24 h after treatment with FKB (10 μg/mL) or si-SMAD4, SGC-7901 cells (5.0 × 103) were plated into a 96-well plate. After culturing for 1, 2, 3, and 4 days, the original culture medium was removed, and serum-free medium containing 10 μL CCK-8 solution (Dojindo, Japan) was added to each well and incubated for 1 h at 37°C. Using an enzyme-linked immunoassay instrument, the corresponding absorbance (OD value) was monitored at a wavelength of 570 nm.
2.4. Annexin V-FITC/PI Assay
Cells in the logarithmic growth phase were taken and digested utilizing trypsin without EDTA. Then, they were centrifuged to the cell suspension at 2000 rpm for 5 min. The cell pellet was collected, resuspended in PBS buffer, and centrifuged at 2,000 rpm for 5 min. The cells were resuspended with 500 μL binding solution, which were then incubated with 10 μL Annexin V-FITC staining solution (Meilunbio, Dalian, China). Afterwards, 10 μL propidium iodide (PI; Meilunbio) staining solution was added and incubated for 15 min at room temperature in the dark. Within 1 h, apoptotic rate was assessed utilizing flow cytometry (BD Biosciences, USA).
2.5. Western Blot
Tissues or cells were fully lysed through RIPA lysis buffer (Beyotime, China). After standing on ice for 30 min, the samples were centrifuged at 13,000 g for 10 min. The supernatant was stored at -20°C. The Bradford method was utilized to determine the protein concentration. After being separated, the sample was transferred to the PVDF membrane (Millipore, USA), followed by being sealed in 5% skimmed milk powder for 2 h. Following being washed with TBST, the diluted primary antibodies were added to the membrane and shook it overnight at 4°C. After being washed again, corresponding HRP-labeled secondary antibody (Santa Cruz, USA) was added dropwise to the membrane and fostered at room temperature for 90 min. The membrane was incubated with ECL luminous liquid (Beyotime). The gel imaging analyzer was used to analyze the gray value. Primary antibodies included TGF-β1 (Abcam, USA, ab92486, 1 : 200), SMAD4 (Abcam, ab40759, 1 : 3000), TSPAN12 (Boster, A05472, 1 : 500), caspase-3 (Abcam, ab13847, 1 : 500), caspase-7 (Abcam, ab69540, 1 : 1000), caspase-8 (Abcam, ab25901, 1 : 1000), caspase-9 (Abcam, ab25758, 1 : 300), Bcl-2 (Abcam, ab692, 1 : 500), Bax (Abcam, ab32503, 1 : 5000), cyclin A (Boster, A03889-1, 1 : 1000), cyclin B1 (Boster, BA3168-2, 1 : 200), Cdc2 (Boster, PB0561, 1; 500), Cdc25C (Boster, A01343, 1 : 500), iNOS (Abcam, ab178945, 1 : 1000), CD86 (Abcam, ab239075, 1 : 1000), Arg-1 (Abcam, ab183333, 1 : 1000), CD206 (Abcam, ab64693), CCR2 (Abcam, ab203128, 1 : 1000), and GAPDH (Abcam, ab199553, 1 : 5000) antibodies.
2.6. Cell Cycle Analysis
Cells were harvested as well as rinsed by cold PBS twice. Thereafter, they were fixed with cold 70% ethanol at 4°C overnight, followed by treatment with 10 μg/mL RNase at 37°C for 30 min. Afterwards, they were dyed utilizing 50 μg/mL PI for 5 min away from light. Cell cycle was analyzed with flow cytometry (BD Biosciences, USA).
2.7. Tissue Specimens
Totally, 20 cases of tumor tissues and normal tissues were collected from 20 gastric cancer patients at the General Hospital of Ningxia Medical University between January 2017 and July 2019. The selection criteria of the samples were as follows: patients with primary gastric cancer who had been pathologically diagnosed as gastric adenocarcinoma; no history of chronic diseases such as cardiovascular and cerebrovascular diseases; no history of other malignant tumors; patients who did not receive any treatment such as radiotherapy or chemotherapy; and patients who had the complete clinical data. All of them provided written informed consent, and the study was approved by the Ethics Committee of General Hospital of Ningxia Medical University (2017-030).
2.8. Animals
Female BALB/c nude mice aged 4-6 weeks were purchased from Beijing Laboratory Animal Research Center (China). All animals were reared in an environment of 23 ± 2°C temperature, 55 ± 5% humidity, and 12 : 12 h light/dark cycle. This assay was conducted in line with the guidelines of the General Hospital of Ningxia Medical University Animal Ethics Research Board (2017-030).
2.9. Tumor Cell Inoculation
SGC-7901 cells (5 × 106) that were mixed in 200 μL matrix gel were subcutaneously injected into the armpit of the nude mice. After 7 days, these animals were randomly divided into control group (0.1% DMSO) and FKB group. In the FKB group, mice were intraperitoneally injected by 1.5 mg/kg FKB every second day. Meanwhile, control mice were intraperitoneally injected by equal amount of saline. All animals were allowed to survive until they died of natural causes. The tumor tissue was stored at -80°C for protein extraction.
2.10. Immunohistochemistry
The tumor tissues from nude mice were formalin-fixed and paraffin-embedded. The sections were treated with xylene and a series of ethanol. Following antigen retrieval, the sections were blocked and stained with primary antibodies against TGF-β1 (Abcam, USA, ab92486, 1 : 3000), SMAD4 (Abcam, ab40759, 1 : 3000), and TSPAN12 (Boster, A05472, 1 : 3000), followed by being incubated with secondary antibodies. Then, the sections were counterstained by hematoxylin. The images were acquired under a microscope (Nikon, Japan).
2.11. Preparation of Conditioned Media from SGC-7901 Cells
SGC-7901 cells were seeded with 2 × 104 cells/cm2 lasting 72 h. After reaching 90% confluence, the cells were administrated with or without 10 μg/mL FKB for 6 h. The medium was exchanged with serum-free fresh medium lasting 24 h. Thereafter, the cells were centrifuged at 2,000 g at 4°C lasting 10 min and were filtered with 0.22 mm filters. The conditioned media were stored at –80°C.
2.12. Transwell Assay
THP-1 cells were planted into 6-well plates (2 × 105 cells/well), followed by incubation with conditioned medium. At 48 h, THP-1 cells were harvested for transwell assays. THP-1 cells were maintained in the upper Transwell chamber with FBS-free RPMI-1640 media, and 600 μL RPMI-1640 media plus 20% FBS was added to the lower chamber. After incubation for 24 h, the cells in the lower chamber were fixed with 4% paraformaldehyde as well as stained crystal violet staining solution. Three random fields were chosen.
2.13. Statistical Analysis
The GraphPad Prism 7.0 software was utilized to present statistical analyses. Measurement data were denoted as the mean ± standard deviation. Comparisons of two groups were presented via Student's t test. More than 2 independent groups were compared by use of one-way analysis of variance. P < 0.05 was set as the cutoff value.
3. Results
3.1. FKB Suppresses Growth and Induces Apoptosis of Gastric Cancer Cells
The CCK-8 was utilized to detect cell proliferative activity. The results suggested that, compared with controls, FKB treatment slowed down the growth of gastric cancer SGC-7901 cells, which started from day two (Figure 1(a)). Annexin V-FITC/PI test was presented to examine cell apoptosis. In comparison with controls, the apoptotic rate of SGC-7901 cells in the FKB treatment group was significantly increased (Figures 1(b) and 1(c)). Proapoptotic and antiapoptotic proteins were tested in SGC-7901 cells under FKB exposure by western blot (Figure 1(d)). FKB treatment increased Bax expression (Figure 1(e)) and decreased Bcl-2 expression (Figure 1(f)) in SGC-7901 cells. Furthermore, the Bax/Bcl − 2 ratio was distinctly increased following FKB treatment (Figure 1(g)). We also examined Caspase family members in the two groups. FKB treatment increased expression of caspase-3, caspase-7, caspase-8, and caspase-9 (Figures 1(h)–1(k)). Collectively, FKB could activate both extrinsic and intrinsic apoptotic pathways, thereby exhibiting apoptotic effects in gastric cancer cells.
Figure 1.
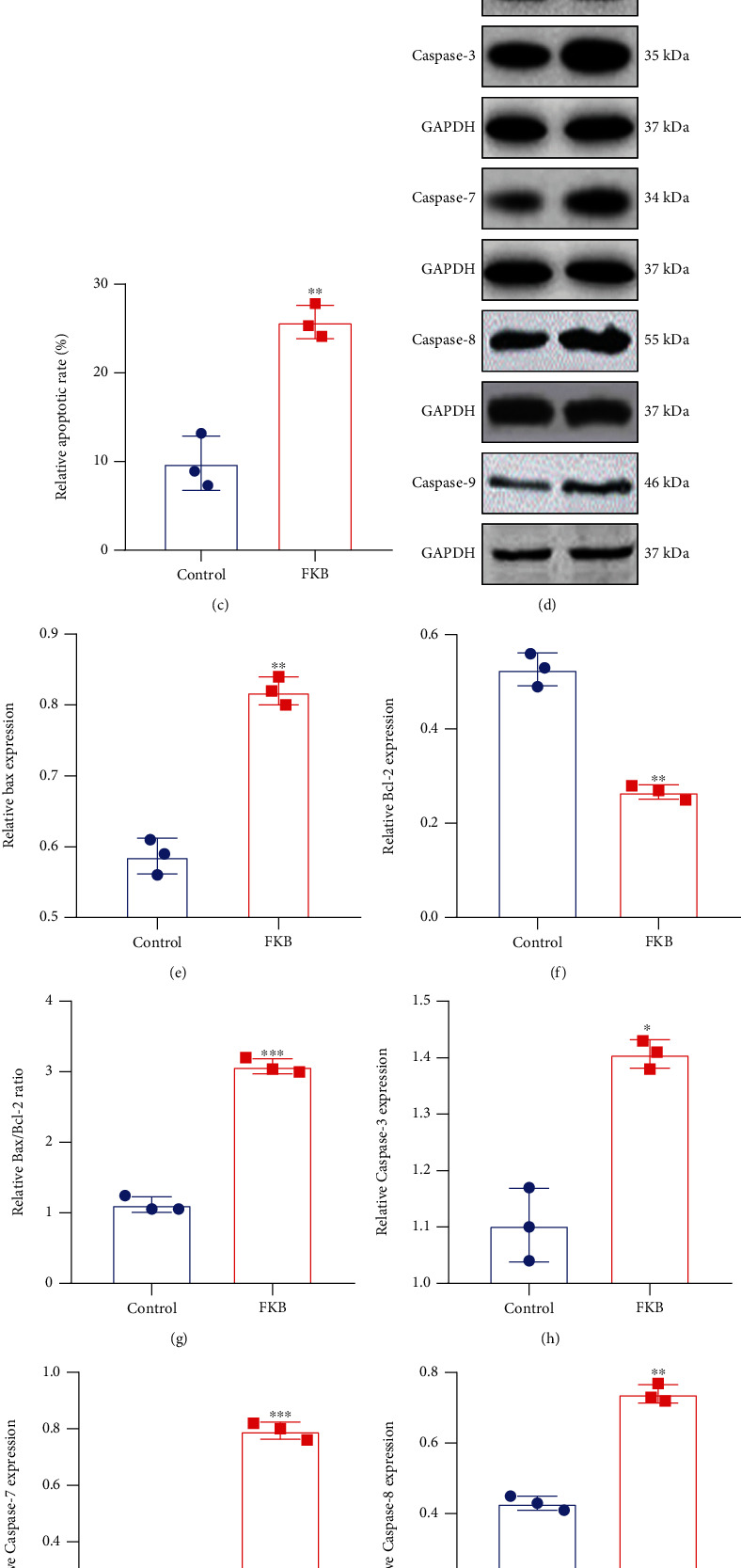
FKB suppresses growth and induces apoptosis of gastric cancer cells. (a) Cell growth curve for SGC-7901 cells after treatment with FKB. (b, c) Apoptosis of SGC-7901 cells was investigated following FKB treatment utilizing Annexin V-FITC/PI test. (d) Representative images of western blots. (e) Bax, (f) Bcl-2, (g) Bax/Bcl-2, (h) caspase-3, (i) caspase-7, (j) caspase-8, and (k) caspase-9 expression was quantified in SGC-7901 cells of the two groups. ∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001; ∗∗∗∗p < 0.0001.
3.2. FKB Downregulates Cell Cycle-Related Proteins in Gastric Cancer Cells
We further observed the effects of FKB treatment on cyclins and CDKs during cell cycle progression in SGC-7901 cells (Figure 2(a)). After treatment of 10 μg/mL FKB for 24 h, Cdc2 (Figure 2(b)), Cdc25C (Figure 2(c)), cyclin A (Figure 2(d)), and cyclin B1 (Figure 2(e)) expression was remarkedly reduced in SGC-7901 cells. The flow cytometry analysis of the cell cycle showed that the percentage of G2/M was higher in 10 μg/mL FKB-treated SGC-7901 cells than controls (Figures 2(f) and 2(g)), indicating that FKB triggered G2/M cell cycle arrest in gastric cancer cells.
Figure 2.
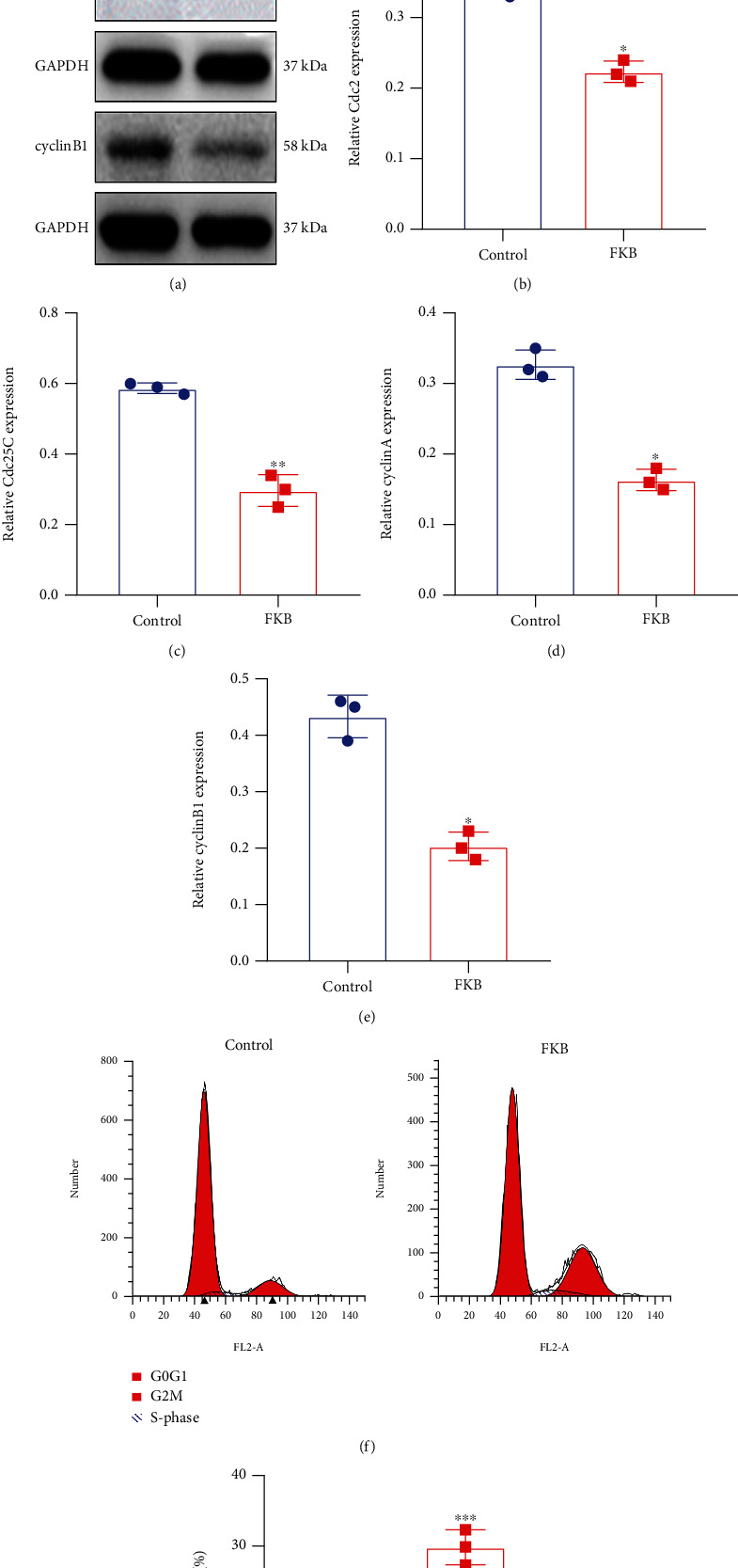
FKB downregulates the expression of cell cycle-related proteins in gastric cancer cells. (a) Representative images of western blots. (b) Cdc2, (c) Cdc25C, (d) cyclin A, and (e) cyclin B1 expression was quantified in SGC-7901 cells of the two groups. (f, g) The flow cytometry analysis of the cell cycle in SGC-7901 cells of the two groups. ∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001.
3.3. Inactivation of the TSPAN12 Expression and TGF-β1/SMAD4 Pathway in Gastric Cancer
Western blot was utilized to detect TSPAN12, TGF-β1, and SMAD4 expression in gastric cancer tissues and normal tissues. In Figure 3(a), TSPAN12 expression was distinctly weakened in gastric cancer tissues than normal tissues. Furthermore, low expression of TGF-β1 (Figure 3(b)) and SMAD4 (Figures 3(c) and 3(d)) was found in gastric cancer tissues. The above results were confirmed by immunohistochemistry (Figure 3(e)).
Figure 3.
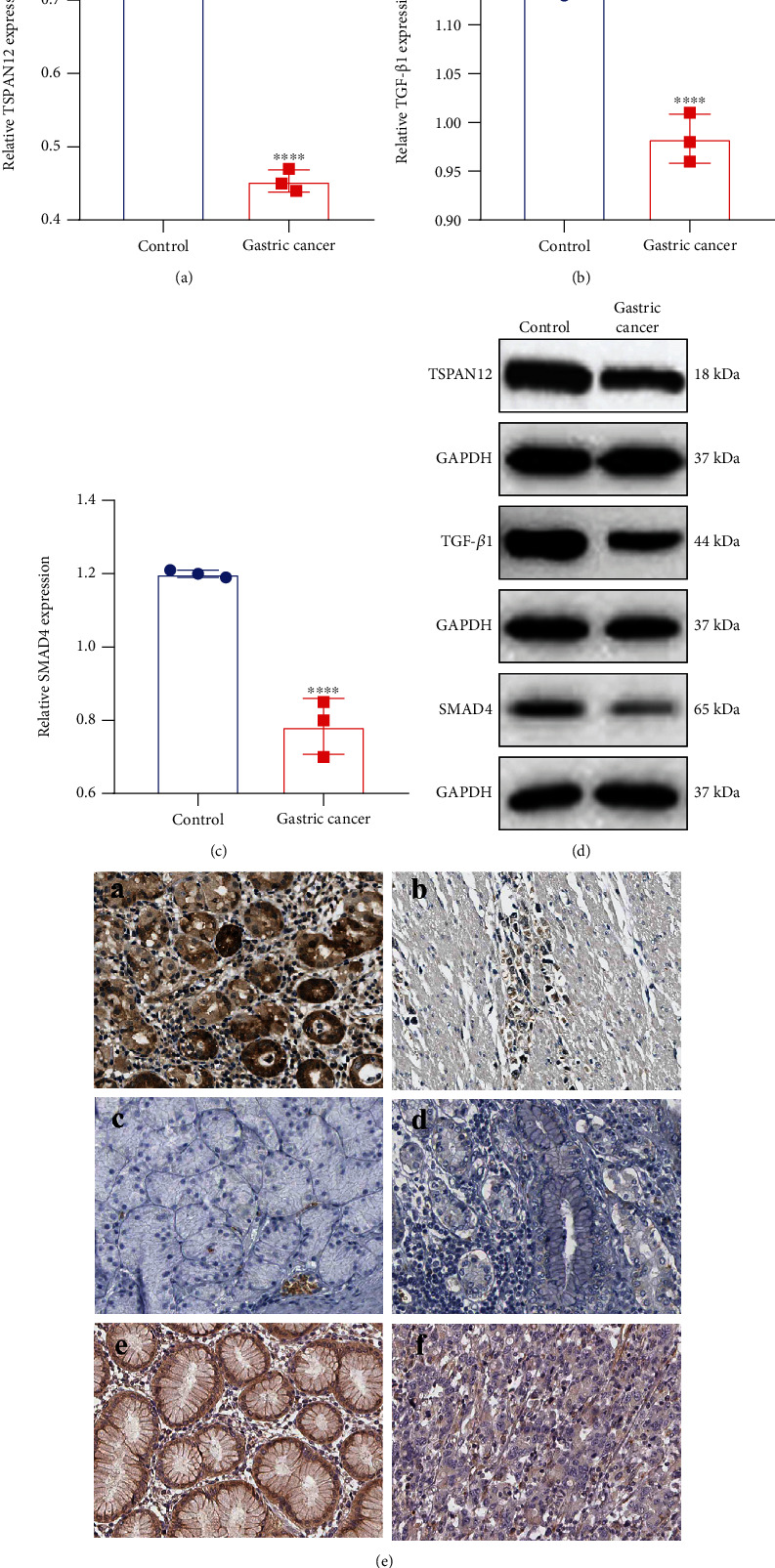
Inactivation of the TSPAN12 expression and TGF-β1/SMAD4 pathway in gastric cancer. (a–c) TSPAN12, TGF-β1, and SMAD4 expression was tested in gastric cancer and normal tissues utilizing western blot. (d) Representative images of western blots. (e) Immunohistochemistry results of (a, b) TSPAN12, (c, d) TGF-β1, and (e, f) SMAD4 in gastric cancer and normal tissues. ∗∗∗∗p < 0.0001.
3.4. FKB Activates the TSPAN12 Expression and TGF-β1/SMAD4 Pathway in Gastric Cancer Cells
TSPAN12, TGF-β1, and SMAD4 proteins were examined in SGC-7901 cells treated with FKB via western blot. FKB treatment prominently increased SMAD4 (Figure 4(a)), TGF-β1 (Figure 4(b)), and TSPAN12 (Figure 4(c)) expression in SGC-7901 cells. The results indicated that FKB could exert an anticancer effect via activation of TSPAN12 and the TGF-β1/SMAD4 pathway.
Figure 4.
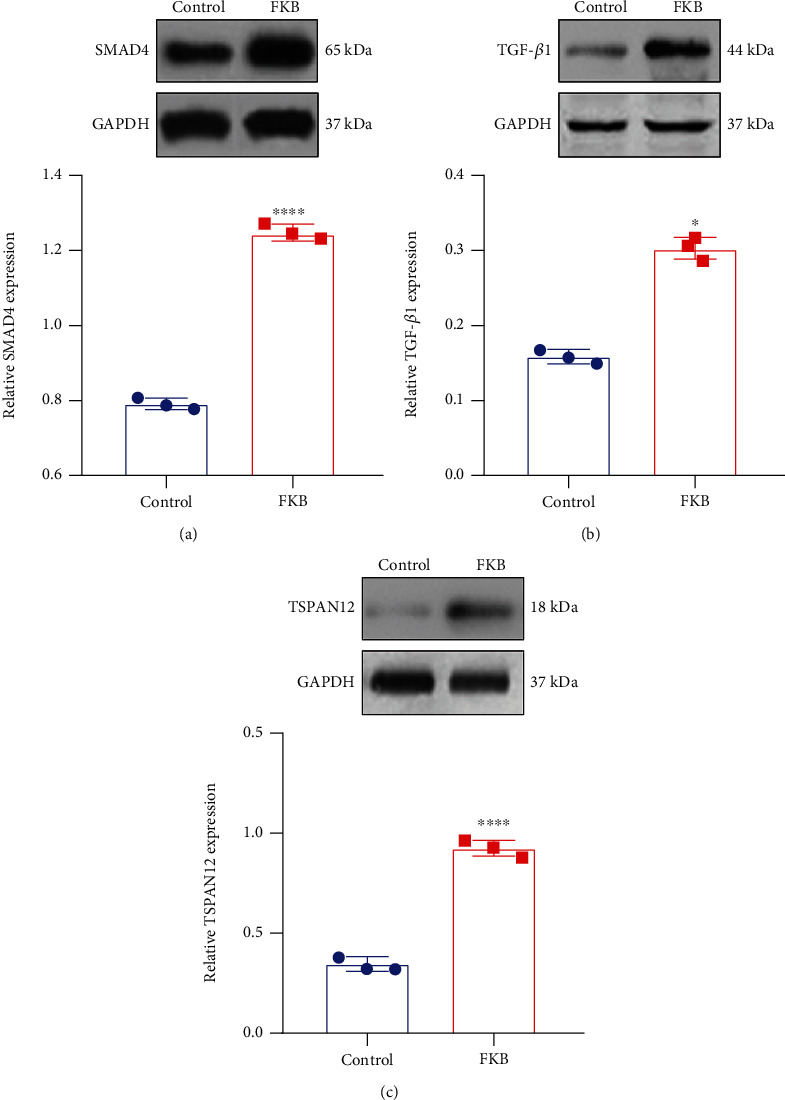
FKB activates the TSPAN12 expression and TGF-β1/SMAD4 pathway in gastric cancer cells. (a) SMAD4, (b) TGF-β1, and (c) TSPAN12 expression was quantified in SGC-7901 cells treated with FKB. ∗p < 0.05; ∗∗∗∗p < 0.0001.
3.5. FKB Suppresses Proliferation and Accelerates Apoptosis of Gastric Cancer Cells Related to SMAD4
We next probed into the mechanisms of FKB treatment on proliferative and apoptotic capacities of gastric cancer cells. si-SMAD4 was utilized to silence SMAD4 in SGC-7901 cells. CCK-8 results demonstrated that the cellular proliferative capacities were boosted after transfection with si-SMAD4 (Figure 5(a)). Following cotransfection with FKB and si-SMAD4, the cellular proliferative capacities were suppressed in comparison with the si-SMAD4 group (Figure 5(a)). Compared to controls, the apoptotic rate of SGC-7901 cells was accelerated by FKB treatment, which was inhibited by si-SMAD4 (Figures 5(b) and 5(c)). In comparison with the si-SMAD4 group, the apoptosis rate of SGC-7901 cells was improved when cotransfection with FKB and si-SMAD4. These findings revealed that FKB could suppress growth and accelerate apoptosis of gastric cancer cells, which was related to SMAD4. We also found that SMAD4 knockdown inhibited Bax expression (Figures 5(d) and 5(e)) and promoted Bcl-2 expression (Figure 5(f)) in SGC-7901 cells in comparison to controls. Moreover, the Bax/Bcl − 2 ratio was decreased when transfection with si-SMAD4 (Figure 5(g)). si-SMAD4 transfection decreased the expression of caspase-3, caspase-7, caspase-8, and caspase-9 in SGC-7901 cells (Figures 5(h)–5(k)). However, FKB treatment obviously reversed the expression of above proteins induced by si-SMAD4. Taken together, FKB could increase proapoptotic proteins and decrease antiapoptotic proteins in gastric cancer cells, which was related to SMAD4.
Figure 5.
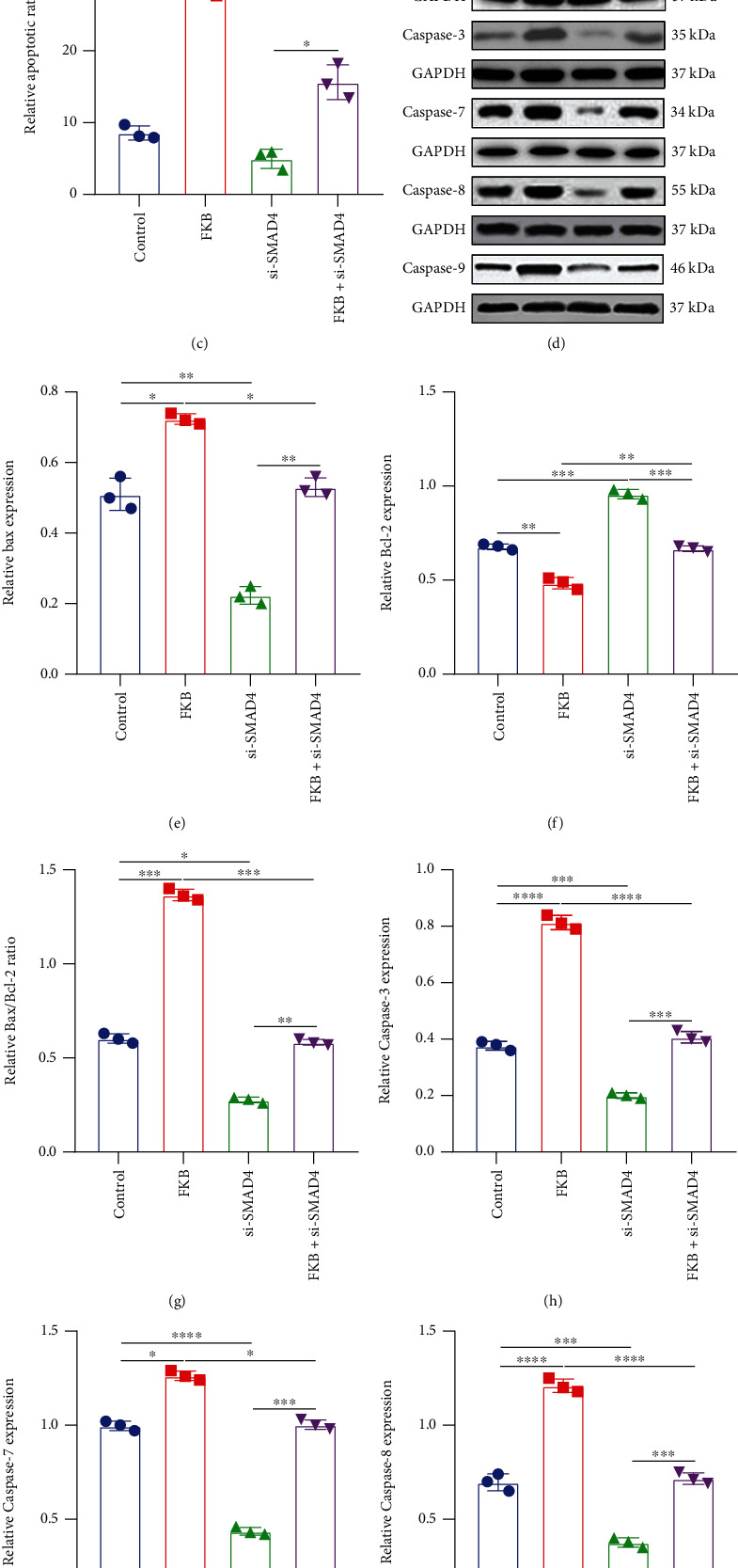
FKB suppresses growth and accelerates apoptosis of gastric cancer cells partly through SMAD4. (a) Cell growth curve for SGC-7901 cells after treatment with FKB and/or si-SMAD4. (b, c) Apoptosis of SGC-7901 cells was investigated when transfection with FKB and/or si-SMAD4 utilizing Annexin V-FITC/PI test. (d) Representative images of western blot. (e) Bax, (f) Bcl-2, (g) Bax/Bcl-2, (h) caspase-3, (i) caspase-7, (j) caspase-8, and (k) caspase-9 expression was quantified in SGC-7901 cells transfected with FKB and/or si-SMAD4. ∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001; ∗∗∗∗p < 0.0001.
3.6. FKB Weakens the Expression of Cell Cycle-Related Proteins and TGF-β1 in Gastric Cancer Cells Related to SMAD4
Western blot was presented to determine the expression of cyclins, CDKs, and TGF-β1 in SGC-7901 cells transfected with FKB and/or si-SMAD4 (Figure 6(a)). Firstly, we found that silencing SMAD4 significantly elevated Cdc2 (Figure 6(b)), Cdc25C (Figure 6(c)), cyclin A (Figure 6(d)), and cyclin B1 (Figure 6(e)) expression in SGC-7901 cells compared to controls. However, after cotreatment with FKB and si-SMAD4, the expression of these proteins was distinctly reversed. Furthermore, our data show that TGF-β1 expression was markedly decreased by SMAD4 knockdown, which was ameliorated following cotreatment with FKB (Figure 6(f)). The above data suggested that FKB could downregulate cell cycle-related proteins and TGF-β1 in gastric cancer cells party through SMAD4.
Figure 6.
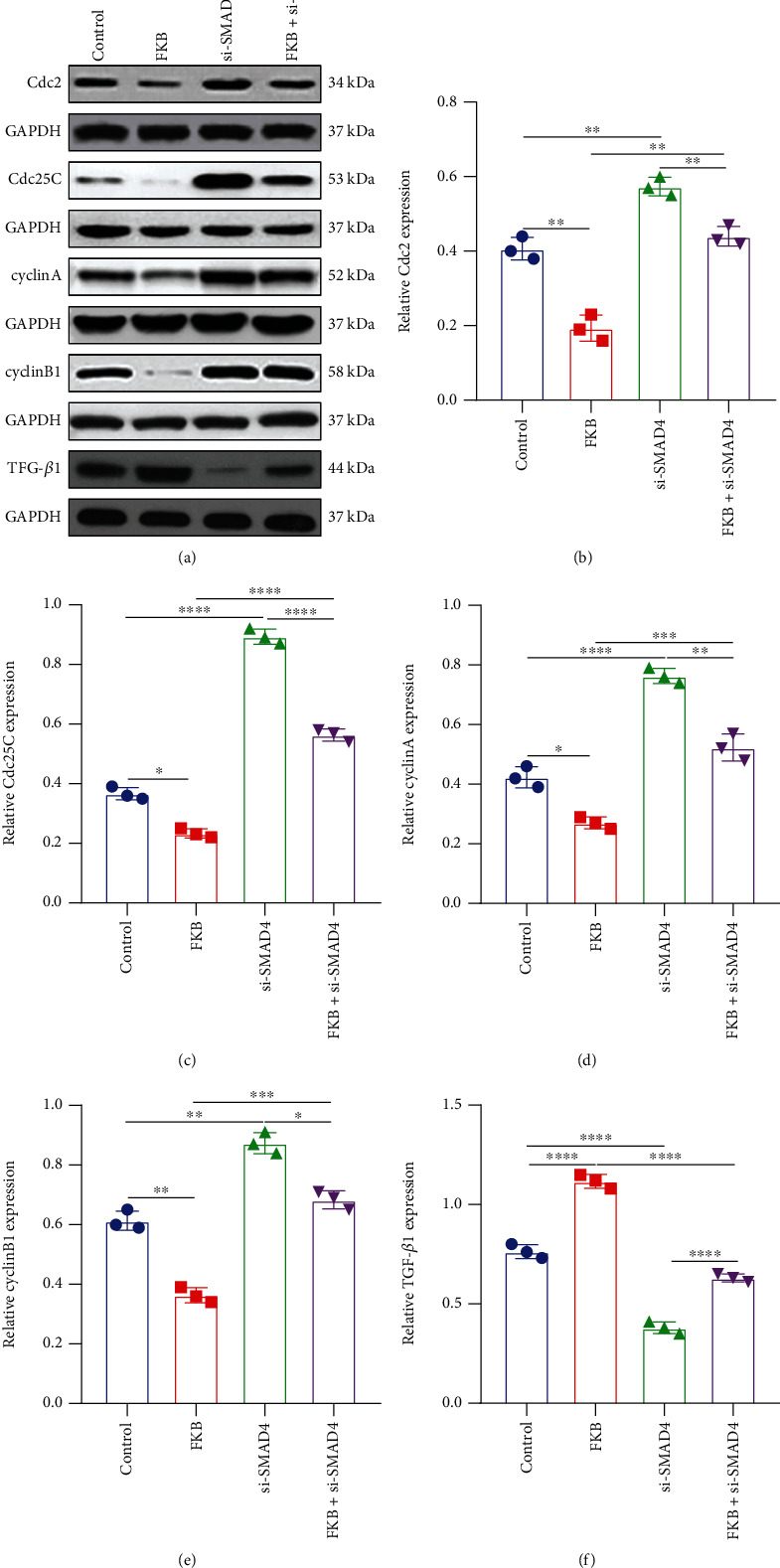
FKB weakens the expression of cell cycle-related proteins and TGF-β1 in gastric cancer cells related to SMAD4. (a) Representative images of western blots. (b) Cdc2, (c) Cdc25C, (d) cyclin A, (e) cyclin B1, and (f) TGF-β1 expression was quantified in SGC-7901 cells transfected with FKB and/or si-SMAD4. ∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001; ∗∗∗∗p < 0.0001.
3.7. FKB Activates the TSPAN12 Expression and TGF-β1/SMAD4 Pathway In Vivo
The anticancer effects of 4FKB were assessed using nude mice in vivo. SGC-7901 cells were xenografted into nude mice. There were no signs of toxicity in any of them. We examined TGF-β1, SMAD4, and TSPAN12 expression in gastric cancer tissues between the two groups. Both in immunohistochemistry and western blot, SMAD4, TGF-β1, and TSPAN12 expression was markedly higher in the FKB group than controls (Figures 7(a)–7(g)). The survival time of nude mice was distinctly prolonged after treatment with FKB in comparison to controls (Figure 7(h)). Additionally, the tumor weight of nude mice was lowered by FKB administration than controls (Figure 7(i)).
Figure 7.
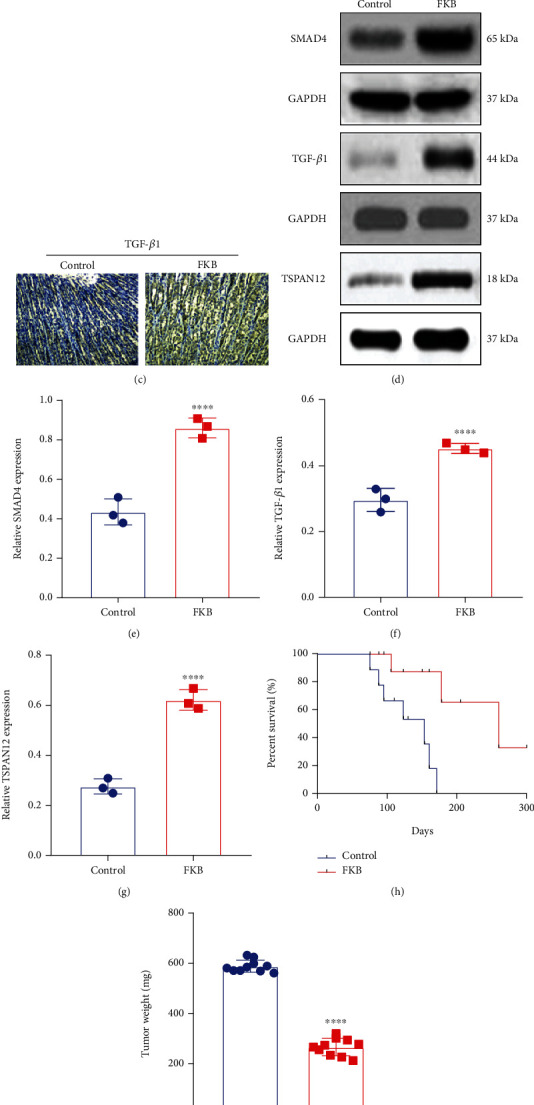
FKB activates TSPAN12 expression and TGF-β1/SMAD4 pathway in vivo. (a–c) Immunohistochemistry results of TGF-β1, SMAD4, and TSPAN12 proteins in gastric cancer tissues of FKB treatment and control groups. Bar = 20 μm. (d) Representative images of western blot. (e–g) TGF-β1, SMAD4, and TSPAN12 expression was examined in gastric cancer tissues of nude mice between FKB treatment and control groups using western blot. (h) FKB treatment prolonged the survival time of nude mice xenografted with SGC-7901 cells. (i) FKB treatment lowered the tumor weight of nude mice xenografted with SGC-7901 cells. ∗∗∗∗p < 0.0001.
3.8. FKB-Treated Gastric Cancer Cells Polarize Macrophages toward M1 Phenotype
Following treatment with 10 μg/mL FKB for 6 h, the cultured media of SGC-7901 cells were exchanged by fresh serum-free media. At 24 h, conditioned media were harvested for treating THP-1 cells. Alterations in THP-1 cell phenotype were evaluated through detecting the surface markers iNOS and CD86 (M1) as well as Arg-1 and CD206 (M2). In comparison to controls, iNOS and CD86 expression was significantly enhanced, and Arg-1 and CD206 expression was significantly decreased for THP-1 cells cultured in conditioned media from FKB-treated SGC-7901 cells (Figures 8(a)–8(e)), indicating that M2 induction capacities were reduced in FKB-treated SGC-7901cells.
Figure 8.
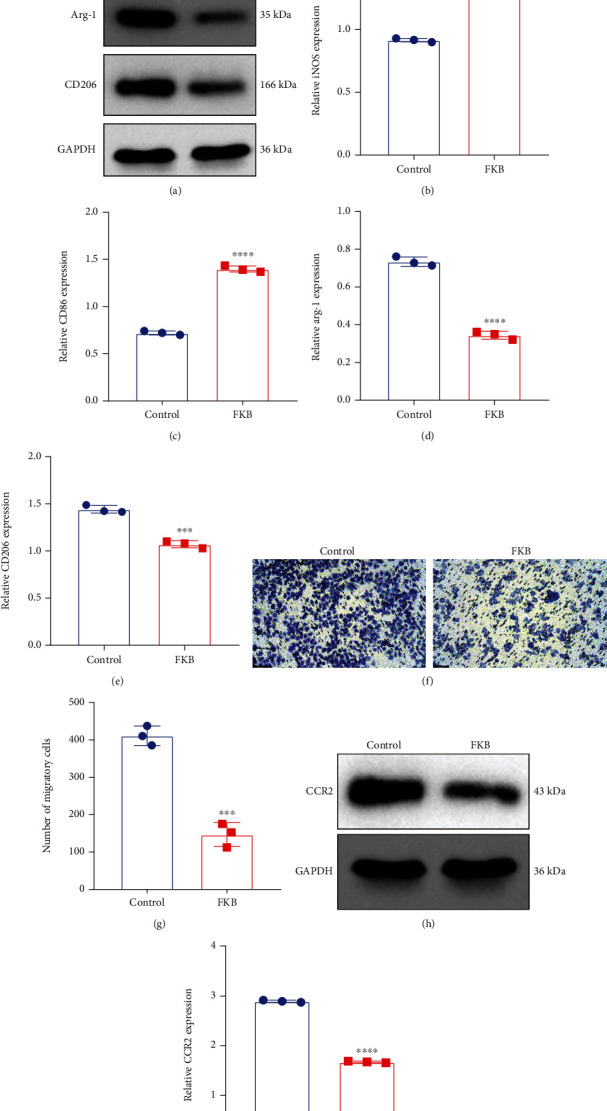
FKB-treated gastric cancer cells polarize macrophages toward M1 phenotype and weaken macrophage migration. (a–e) Western blot of the expression of iNOS and CD86 (M1) as well as Arg-1 and CD206 (M2) in THP-1 cells treated with conditioned media from normal or FKB-treated SGC-7901 cells. (f, g) Transwell of THP-1 cells treated with conditioned media from normal or FKB-treated SGC-7901 cells. Bar, 100 μm. (h, i) Western blot of the expression of CCR2 in THP-1 cells treated with conditioned media from normal or FKB-treated SGC-7901 cells. ∗∗∗p < 0.001; ∗∗∗∗p < 0.0001.
3.9. FKB-Treated Gastric Cancer Cells Weaken Macrophage Migration
Further analysis showed that the migratory capacities of THP-1 cells cultured in conditioned media from FKB-treated SGC-7901 cells were markedly decreased in comparison to that of controls (Figures 8(f) and 8(g)). Additionally, compared with controls, CCR2 expression was significantly reduced in THP-1 cells cultured in conditioned media from FKB-treated SGC-7901 cells (Figures 8(h) and 8(i)). Altogether, FKB-treated gastric cancer cells weakened macrophage migration. Figure 9 illustrates the flowchart of the study design.
Figure 9.
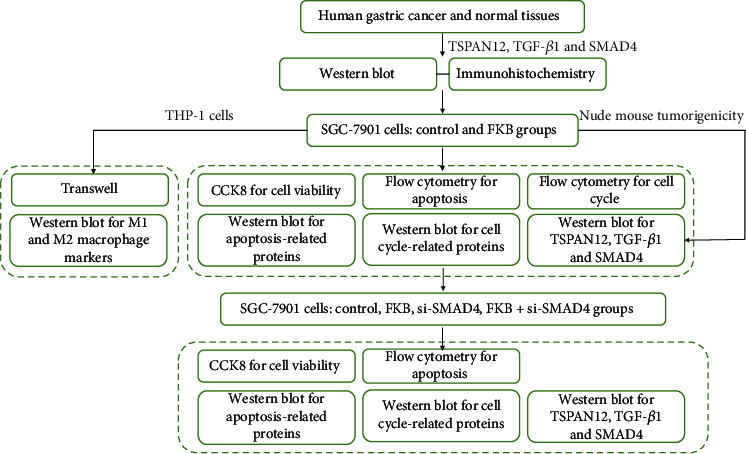
The flowchart of the study design.
4. Discussion
Conventional antitumor drugs for the treatment of gastric cancer, such as doxorubicin and methotrexate, are often limited due to their systemic toxicity and lack of specificity [40, 41]. Therefore, it is necessary to develop effective drugs. Many studies have shown that natural compounds derived from food and plants possess anticancer effects, including FKB [29]. FKB has been fully proven to have great potential as an anticancer agent [29]. Previous research has demonstrated that FKB has cytotoxic effects on human colon cancer (LoVo) [32], lung adenocarcinoma (A-549) [33], prostate cancer (PC3 and DU-145) [29], and squamous cell carcinoma (KB) [42] cells. FKB could induce apoptosis as well as G2/M cell cycle arrest of different cancer cells [32, 43]. In this study, we extracted the protein from gastric cancer tissues and found that the protein expression of TGF-β1, SMAD4, and TSPAN12 was prominently decreased in gastric cancer. Intriguingly, FKB treatment significantly increased their expression in gastric cancer cells. For the nude mouse gastric cancer model, the survival time of the FKB treatment group was significantly improved. Furthermore, FKB treatment significantly inhibited cell proliferation as well as accelerated cell apoptosis and G2/M arrest for SGC-7901 cells. These data suggest that FKB could play an antitumor effect related to the TGF-β1/SMAD4 pathway.
Apoptosis can balance cell division and cell death. As we all know, apoptosis is related to cancer, and apoptosis inducers have been explored as new ideas for cancer treatment. Herein, we evaluated the apoptosis regulatory mechanism of FKB in gastric cancer cells. Apoptosis is controlled by mitochondrial [44] and cell-surface death receptor pathways [45]. As a previous study, FKB-induced apoptosis depends largely on the damage of mitochondria [42]. The loss of mitochondrial membrane potential induces the release of cytochrome c from the mitochondria into the cytoplasm, which binds to the apoptotic protease activator 1 and caspase-9/7, thereby the activation of the downstream of caspase-3 [30]. In this study, we found that FKB promoted the expression of caspase-3, caspase-7, caspase-8, and caspase-9, suggesting that FKB may accelerate the apoptosis of gastric cancer cells partly through the mitochondrial pathway [46]. Apoptotic Bax and proapoptotic Bcl-2 are involved in the endogenous apoptotic pathway [47, 48]. The proapoptosis activity of Bax protein can be closely related to Bcl-2 protein. Both mediate cell apoptosis through homologous and heterologous complexes. Therefore, the Bax/Bcl − 2 ratio accelerates the execution of apoptosis [49]. In this study, the increase in FKB-induced apoptosis was in association with the decrease in the expression of Bcl-2 and Bax proteins. These results indicate that FKB can interfere with Bax/Bcl − 2, thereby causing apoptosis of gastric cancer cells. Data from current studies indicate that FKB induces apoptosis through the mitochondrial pathway and the cell membrane death receptor pathway.
The destruction of the cell cycle is one of the goals of the development of novel anticancer drugs. A few studies have shown that FKB induces G2/M blockage in leiomyosarcoma (LMS) cells, H460 cells, and ACC-2 cells [50–52]. This cell cycle arrest is related to the reduction of cyclin A, cyclin B1, Cdc2, and Cdc25C. Cdc2 kinase is mainly activated during the G2/M phase, which is in association with cyclin A and cyclin B1 [53]. At the beginning of mitosis, both Cdc2C and cyclin B1 residues are dephosphorylated by Cdc25C [54]. Inactivation of Cdc2 leads to G2/M stagnation after DNA damage [55]. Our data suggest that the inhibition effect of FKB on gastric cancer cell growth may be related to the regulation of cell cycle-related proteins including cyclin A, cyclin B1, Cdc2, and Cdc25C. In a previous study, it has been reported that FKB may induce cell apoptosis by p21-induced cell cycle arrest and activation of p38 in cervical cancer HeLa cells [56]. These findings confirm the antiapoptosis effect of FKB by regulating various cell cycle-related proteins in the treatment of cancers.
As we all know, SMAD4 is the main signal transduction pathway downstream of TGF-β. Evidence indicates that SMAD4 can act as a tumor suppressor gene in gastrointestinal cancer [57–59]. TGF-β has a potential tumor suppressor effect via inhibiting proliferative and apoptotic abilities of gastric cancer cells [21, 59]. As shown in this study, it was observed the decrease in the expression of TGF-β1 and SMAD4 proteins in gastric cancer tissues. This provided potential evidence that downregulated TGF-β1 and SMAD4 might participate in gastric cancer progression. TGF-β-induced cell cycle arrest is the induction of CDK inhibitor expression [60]. Moreover, TGF-β also limits the formation of gastric cancer by activating the apoptotic pathways [21]. From the data we obtained, FKB can induce the increase in the expression of TSPAN12 protein, suggesting that TSPAN12 may be a target for the treatment of FKB. Our data showed that after transfection with si-SMAD4, the therapeutic effect of FKB was significantly weakened, indicating that FKB may inhibit gastric cancer partly through the TGF-β1/SMAD4 pathway. Also, FKB treatment prolonged the survival time of nude mice. We also demonstrated iNOS and CD86 expression was significantly enhanced, and Arg-1 and CD206 expression was significantly decreased in THP-1 cells cultured in conditioned media from FKB-treated SGC-7901 cells. This demonstrated that FKB enabled to weaken the capacity of gastric cancer cells in inducing M2 macrophage polarization. Additionally, FKB-treated SGC-7901 cells weakened macrophage migration. Therefore, FKB may be a promising antigastric cancer drug, but its effect and mechanism deserve more in-depth study.
5. Conclusion
Taken together, our findings confirmed the antigastric cancer effect of FKB both in gastric cancer cells and nude mice with gastric cancer. Mechanically, FKB treatment could exert antiproliferation and proapoptosis effects, which was related to the TGF-β1/SMAD4 pathway. Additionally, FKB treatment weakened M2 macrophage polarization induction capacities of gastric cancer cells.
Acknowledgments
This work was funded by the Ningxia Natural Science Foundation (2020AAC03418) and the Foundation of General Hospital of Ningxia Medical University (2020/294).
Abbreviations
- FKB:
Flavokawain B
- CDKs:
Cyclin-dependent protein kinases
- TGF:
Transforming growth factor
Contributor Information
Shaoqi Yang, Email: shaoqiynh@nyfy.com.cn.
Fang He, Email: blue0708@nyfy.com.cn.
Data Availability
The datasets analyzed during the current study are available from the corresponding authors on reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
Authors' Contributions
Yongzhao Zhu and Weining Fan have contributed equally to this work and share first authorship.
References
- 1.Feng R. M., Zong Y. N., Cao S. M., Xu R. H. Current cancer situation in China: good or bad news from the 2018 Global Cancer Statistics? Cancer Commun (Lond) . 2019;39(1):p. 22. doi: 10.1186/s40880-019-0368-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Miller K. D., Nogueira L., Mariotto A. B., et al. Cancer treatment and survivorship statistics, 2019. CA: a Cancer Journal for Clinicians . 2019;69(5):363–385. doi: 10.3322/caac.21565. [DOI] [PubMed] [Google Scholar]
- 3.Siegel R. L., Miller K. D., Jemal A. Cancer statistics, 2019. CA: a Cancer Journal for Clinicians . 2019;69(1):7–34. doi: 10.3322/caac.21551. [DOI] [PubMed] [Google Scholar]
- 4.Qiu X. T., Song Y. C., Liu J., Wang Z. M., Niu X., He J. Identification of an immune-related gene-based signature to predict prognosis of patients with gastric cancer. World J Gastrointest Oncol . 2020;12(8):857–876. doi: 10.4251/wjgo.v12.i8.857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wang X., Cheng G., Miao Y., et al. Piezo type mechanosensitive ion channel component 1 facilitates gastric cancer omentum metastasis. Journal of Cellular and Molecular Medicine . 2021;25(4):2238–2253. doi: 10.1111/jcmm.16217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Chen T., Xu X. Y., Zhou P. H. Emerging molecular classifications and therapeutic implications for gastric cancer. Chinese Journal of Cancer . 2016;35(1):p. 49. doi: 10.1186/s40880-016-0111-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Xu J., Zhang Y., Jia R., et al. Anti-PD-1 antibody SHR-1210 combined with apatinib for advanced hepatocellular carcinoma, gastric, or esophagogastric junction cancer: an open-label, dose escalation and expansion study. Clinical Cancer Research . 2019;25(2):515–523. doi: 10.1158/1078-0432.CCR-18-2484. [DOI] [PubMed] [Google Scholar]
- 8.Sun L., Liu J., Yuan Y., Zhang X., Dong Z. Protective effect of the BET protein inhibitor JQ1 in cisplatin-induced nephrotoxicity. American Journal of Physiology. Renal Physiology . 2018;315(3):F469–f478. doi: 10.1152/ajprenal.00527.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Orditura M., Galizia G., Sforza V., et al. Treatment of gastric cancer. World Journal of Gastroenterology . 2014;20(7):1635–1649. doi: 10.3748/wjg.v20.i7.1635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ji C., Zhao Y., Kou Y. W., et al. Cathepsin F knockdown induces proliferation and inhibits apoptosis in gastric cancer cells. Oncology Research . 2018;26(1):83–93. doi: 10.3727/096504017X14928634401204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Xiong W., Deng Z., Tang Y., Deng Z., Li M. Downregulation of KMT2D suppresses proliferation and induces apoptosis of gastric cancer. Biochemical and Biophysical Research Communications . 2018;504(1):129–136. doi: 10.1016/j.bbrc.2018.08.143. [DOI] [PubMed] [Google Scholar]
- 12.Zhang X., Qin Y., Pan Z., et al. Cannabidiol induces cell cycle arrest and cell apoptosis in human gastric cancer SGC-7901 cells. Biomolecules . 2019;9(8):p. 302. doi: 10.3390/biom9080302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Xie H., Li X., Chen Y., Lang M., Shen Z., Shi L. Ethanolic extract of Cordyceps cicadae exerts antitumor effect on human gastric cancer SGC-7901 cells by inducing apoptosis, cell cycle arrest and endoplasmic reticulum stress. J Ethnopharmacol . 2019;231:230–240. doi: 10.1016/j.jep.2018.11.028. [DOI] [PubMed] [Google Scholar]
- 14.Xu M., Gu W., Shen Z., Wang F. Anticancer activity of phloretin against human gastric cancer cell lines involves apoptosis, cell cycle arrest, and inhibition of cell invasion and JNK signalling pathway. Medical Science Monitor . 2018;24:6551–6558. doi: 10.12659/MSM.910542. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 15.Palmer N., Kaldis P. Less-well known functions of cyclin/CDK complexes. Seminars in Cell & Developmental Biology . 2020;107:54–62. doi: 10.1016/j.semcdb.2020.04.003. [DOI] [PubMed] [Google Scholar]
- 16.Bernabò N., Ramal-Sanchez M., Valbonetti L., et al. Cyclin-CDK complexes are key controllers of capacitation-dependent actin dynamics in mammalian spermatozoa. International Journal of Molecular Sciences . 2019;20(17):p. 4236. doi: 10.3390/ijms20174236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Swaffer M. P., Jones A. W., Flynn H. R., Snijders A. P., Nurse P. CDK substrate phosphorylation and ordering the cell cycle. Cell . 2016;167(7):1750–1761.e16. doi: 10.1016/j.cell.2016.11.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Shalini S., Dorstyn L., Dawar S., Kumar S. Old, new and emerging functions of caspases. Cell Death and Differentiation . 2015;22(4):526–539. doi: 10.1038/cdd.2014.216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Brentnall M., Rodriguez-Menocal L., De Guevara R. L., Cepero E., Boise L. H. Caspase-9, caspase-3 and caspase-7 have distinct roles during intrinsic apoptosis. BMC Cell Biology . 2013;14(1):p. 32. doi: 10.1186/1471-2121-14-32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Larsen B. D., Sørensen C. S. The caspase-activated DNase: apoptosis and beyond. The FEBS Journal . 2017;284(8):1160–1170. doi: 10.1111/febs.13970. [DOI] [PubMed] [Google Scholar]
- 21.Xiong R., Gao J. L., Yin T. G3BP1 activates the TGF-β/Smad signaling pathway to promote gastric cancer. Oncotargets and Therapy . 2019;Volume 12:7149–7156. doi: 10.2147/OTT.S213728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Chen Z. L., Qin L., Peng X. B., Hu Y., Liu B. INHBA gene silencing inhibits gastric cancer cell migration and invasion by impeding activation of the TGF-β signaling pathway. Journal of Cellular Physiology . 2019;234(10):18065–18074. doi: 10.1002/jcp.28439. [DOI] [PubMed] [Google Scholar]
- 23.Zhang Y., Fan K. J., Sun Q., et al. Functional screening for miRNAs targeting Smad4 identified miR-199a as a negative regulator of TGF-β signalling pathway. Nucleic Acids Research . 2012;40(18):9286–9297. doi: 10.1093/nar/gks667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Zhang C., Lai M. B., Pedler M. G., et al. Endothelial cell-specific inactivation of TSPAN12 (Tetraspanin 12) reveals pathological consequences of barrier defects in an otherwise intact vasculature. Arteriosclerosis, Thrombosis, and Vascular Biology . 2018;38(11):2691–2705. doi: 10.1161/ATVBAHA.118.311689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ji G., Liang H., Wang F., Wang N., Fu S., Cui X. TSPAN12 precedes tumor proliferation by cell cycle control in ovarian cancer. Molecules and Cells . 2019;42:557–567. doi: 10.14348/molcells.2019.0015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Liang G., Meng W., Huang X., et al. miR-196b-5p-mediated downregulation of TSPAN12 and GATA6 promotes tumor progression in non-small cell lung cancer. Proceedings of the National Academy of Sciences of the United States of America . 2020;117(8):4347–4357. doi: 10.1073/pnas.1917531117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Liu J., Chen C., Li G., Chen D., Zhou Q. Upregulation of TSPAN12 is associated with the colorectal cancer growth and metastasis. American Journal of Translational Research . 2017;9(2):812–822. [PMC free article] [PubMed] [Google Scholar]
- 28.Wang J., Qi Q., Zhou W., et al. Inhibition of glioma growth by flavokawain B is mediated through endoplasmic reticulum stress induced autophagy. Autophagy . 2018;14(11):2007–2022. doi: 10.1080/15548627.2018.1501133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Li X., Pham V., Tippin M., et al. Flavokawain B targets protein neddylation for enhancing the anti-prostate cancer effect of Bortezomib via Skp2 degradation. Cell Communication and Signaling: CCS . 2019;17(1):p. 25. doi: 10.1186/s12964-019-0338-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Abu N., Akhtar M. N., Yeap S. K., et al. Flavokawain B induced cytotoxicity in two breast cancer cell lines, MCF-7 and MDA-MB231 and inhibited the metastatic potential of MDA-MB231 via the regulation of several tyrosine kinases in vitro. BMC Complementary and Alternative Medicine . 2016;16(1):p. 86. doi: 10.1186/s12906-016-1046-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.He Q., Liu W., Sha S., et al. Adenosine 5'-monophosphate-activated protein kinase-dependent mTOR pathway is involved in flavokawain B-induced autophagy in thyroid cancer cells. Cancer Science . 2018;109(8):2576–2589. doi: 10.1111/cas.13699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Palko-Łabuz A., Kostrzewa-Susłow E., Janeczko T., et al. Cyclization of flavokawain B reduces its activity against human colon cancer cells. Human & Experimental Toxicology . 2020;39(3):262–275. doi: 10.1177/0960327119882986. [DOI] [PubMed] [Google Scholar]
- 33.Hseu Y. C., Huang Y. C., Thiyagarajan V., et al. Anticancer activities of chalcone flavokawain B from Alpinia pricei Hayata in human lung adenocarcinoma (A549) cells via induction of reactive oxygen species-mediated apoptotic and autophagic cell death. Journal of Cellular Physiology . 2019;234(10):17514–17526. doi: 10.1002/jcp.28375. [DOI] [PubMed] [Google Scholar]
- 34.Chen L., Niu X., Qiao X., et al. Characterization of interplay between autophagy and ferroptosis and their synergistical roles on manipulating immunological tumor microenvironment in squamous cell carcinomas. Frontiers in Immunology . 2022;12, article 739039 doi: 10.3389/fimmu.2021.739039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Niu X., Chen L., Li Y., Hu Z., He F. Ferroptosis, necroptosis, and pyroptosis in the tumor microenvironment: perspectives for immunotherapy of SCLC. Seminars in Cancer Biology . 2022 doi: 10.1016/j.semcancer.2022.03.009. [DOI] [PubMed] [Google Scholar]
- 36.Liu X., Niu X., Qiu Z. A five-gene signature based on stromal/immune scores in the tumor microenvironment and its clinical implications for liver cancer. DNA and Cell Biology . 2020;39(9):1621–1638. doi: 10.1089/dna.2020.5512. [DOI] [PubMed] [Google Scholar]
- 37.Shi M., Gu Y., Jin K., et al. CD47 expression in gastric cancer clinical correlates and association with macrophage infiltration. Cancer Immunology, Immunotherapy . 2021;70(7):1831–1840. doi: 10.1007/s00262-020-02806-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Luo W., Yang L. B., Qian C. C., et al. Flavokawain B alleviates LPS-induced acute lung injury via targeting myeloid differentiation factor 2. Acta Pharmacologica Sinica . 2021 doi: 10.1038/s41401-021-00792-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Wang Y., Yin P., Chen Y. H., et al. A functional variant of SMAD4 enhances macrophage recruitment and inflammatory response via TGF-β signal activation in thoracic aortic aneurysm and dissection. Aging (Albany NY) . 2018;10(12):3683–3701. doi: 10.18632/aging.101662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Korga A., Ostrowska M., Iwan M., et al. Ethanol extracts of allium sp. regulate cyclooxygenase-2 and E-cadherin expression in gastric cancer MKN74 cell line and enhance doxorubicin toxicity. Food & Nutrition Research . 2019;63 doi: 10.29219/fnr.v63.3449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ter Veer E., Haj Mohammad N., van Valkenhoef G., et al. The efficacy and safety of first-line chemotherapy in advanced esophagogastric cancer: a network meta-analysis. Journal of the National Cancer Institute . 2016;108 doi: 10.1093/jnci/djw166. [DOI] [PubMed] [Google Scholar]
- 42.Lin E., Lin W. H., Wang S. Y., et al. Flavokawain B inhibits growth of human squamous carcinoma cells: involvement of apoptosis and cell cycle dysregulation in vitro and in vivo. J Nutr Biochem . 2012;23:368–378. doi: 10.1016/j.jnutbio.2011.01.002. [DOI] [PubMed] [Google Scholar]
- 43.Hua R., Pei Y., Gu H., Sun Y., He Y. Antitumor effects of flavokawain-B flavonoid in gemcitabine-resistant lung cancer cells are mediated via mitochondrial-mediated apoptosis, ROS production, cell migration and cell invasion inhibition and blocking of PI3K/AKT signaling pathway. Journal of BUON . 2020;25(1):262–267. [PubMed] [Google Scholar]
- 44.Brokatzky D., Dörflinger B., Haimovici A., et al. A non-death function of the mitochondrial apoptosis apparatus in immunity. The EMBO Journal . 2019;38(11) doi: 10.15252/embj.2018100907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Green D. R., Llambi F. Cell death signaling. Cold Spring Harbor Perspectives in Biology . 2015;7(12) doi: 10.1101/cshperspect.a006080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kalpage H. A., Bazylianska V., Recanati M. A., et al. Tissue-specific regulation of cytochrome c by post-translational modifications: respiration, the mitochondrial membrane potential, ROS, and apoptosis. The FASEB Journal . 2019;33(2):1540–1553. doi: 10.1096/fj.201801417R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Campbell K. J., Tait S. W. G. Targeting BCL-2 regulated apoptosis in cancer. Open Biology . 2018;8(5) doi: 10.1098/rsob.180002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Peña-Blanco A., García-Sáez A. J. Bax, Bak and beyond - mitochondrial performance in apoptosis. Febs j . 2018;285:416–431. doi: 10.1111/febs.14186. [DOI] [PubMed] [Google Scholar]
- 49.Liu S., Zhu Y., Yan S., et al. Phenethyl isothiocyanate induces IPEC-J2 cells cytotoxicity and apoptosis via S-G(2)/M phase arrest and mitochondria-mediated Bax/Bcl-2 pathway. Comparative Biochemistry and Physiology, Part C: Toxicology & Pharmacology . 2019;226, article 108574 doi: 10.1016/j.cbpc.2019.108574. [DOI] [PubMed] [Google Scholar]
- 50.Shao L. H., Liu S. P., Hou J. X., et al. Cathepsin B cleavable novel prodrug Ac-Phe-Lys-PABC-ADM enhances efficacy at reduced toxicity in treating gastric cancer peritoneal carcinomatosis: an experimental study. Cancer . 2012;118:2986–2996. doi: 10.1002/cncr.26596. [DOI] [PubMed] [Google Scholar]
- 51.Zhao X., Chao Y. L., Wan Q. B., et al. Flavokawain B induces apoptosis of human oral adenoid cystic cancer ACC-2 cells via up-regulation of Bim and down-regulation of Bcl-2 expression. Canadian Journal of Physiology and Pharmacology . 2011;89(12):875–883. doi: 10.1139/y11-088. [DOI] [PubMed] [Google Scholar]
- 52.An J., Gao Y., Wang J., et al. Flavokawain B induces apoptosis of non-small cell lung cancer H460 cells via Bax-initiated mitochondrial and JNK pathway. Biotechnology Letters . 2012;34(10):1781–1788. doi: 10.1007/s10529-012-0976-6. [DOI] [PubMed] [Google Scholar]
- 53.Choi Y. H. Isorhamnetin induces ROS-dependent cycle arrest at G2/M phase and apoptosis in human hepatocarcinoma Hep3B cells. General Physiology and Biophysics . 2019;38(6):473–484. doi: 10.4149/gpb_2019038. [DOI] [PubMed] [Google Scholar]
- 54.Jeon H. G., Yoon C. Y., Yu J. H., et al. Induction of caspase mediated apoptosis and down-regulation of nuclear factor-κB and Akt signaling are involved in the synergistic antitumor effect of gemcitabine and the histone deacetylase inhibitor trichostatin A in human bladder cancer cells. The Journal of Urology . 2011;186(5):2084–2093. doi: 10.1016/j.juro.2011.06.053. [DOI] [PubMed] [Google Scholar]
- 55.Sasaki M., Terabayashi T., Weiss S. M., Ferby I. The tumor suppressor MIG6 controls mitotic progression and the G2/M DNA damage checkpoint by stabilizing the WEE1 kinase. Cell Reports . 2018;24(5):1278–1289. doi: 10.1016/j.celrep.2018.06.064. [DOI] [PubMed] [Google Scholar]
- 56.Yeap S. K., Abu N., Akthar N., et al. Gene expression analysis reveals the concurrent activation of proapoptotic and antioxidant-defensive mechanisms in flavokawain B-treated cervical cancer HeLa cells. Integrative Cancer Therapies . 2017;16(3):373–384. doi: 10.1177/1534735416660383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Schiemer M., Schmitt-Graeff A., Brass V., Hasselblatt P. Gastric cancer and paraneoplastic dermatomyositis as complications of an unrecognized juvenile polyposis syndrome. Zeitschrift für Gastroenterologie . 2019;57(4):497–500. doi: 10.1055/a-0855-4404. [DOI] [PubMed] [Google Scholar]
- 58.Seidlitz T., Chen Y. T., Uhlemann H., et al. Mouse models of human gastric cancer subtypes with stomach-specific CreERT2-mediated pathway alterations. Gastroenterology . 2019;157:1599–1614.e 1592. doi: 10.1053/j.gastro.2019.09.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Yang T., Huang T., Zhang D., et al. TGF-β receptor inhibitor LY2109761 enhances the radiosensitivity of gastric cancer by inactivating the TGF-β/SMAD4 signaling pathway. Aging (Albany NY) . 2019;11(20):8892–8910. doi: 10.18632/aging.102329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Tiwari A., Swamynathan S., Alexander N., et al. KLF4 regulates corneal epithelial cell cycle progression by suppressing canonical TGF-β signaling and upregulating CDK inhibitors P16 and P27. Investigative Ophthalmology & Visual Science . 2019;60(2):731–740. doi: 10.1167/iovs.18-26423. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets analyzed during the current study are available from the corresponding authors on reasonable request.