

Abstract
Polygenic risk scores (PRSs) can boost risk prediction in late-onset Alzheimer’s disease (LOAD) beyond apolipoprotein E (APOE) but have not been leveraged to identify genetic resilience factors. Here, we sought to identify resilience-conferring common genetic variants in (1) unaffected individuals having high PRSs for LOAD, and (2) unaffected APOE-ε4 carriers also having high PRSs for LOAD. We used genome-wide association study (GWAS) to contrast “resilient” unaffected individuals at the highest genetic risk for LOAD with LOAD cases at comparable risk. From GWAS results, we constructed polygenic resilience scores to aggregate the addictive contributions of risk-orthogonal common variants that promote resilience to LOAD. Replication of resilience scores was undertaken in eight independent studies. We successfully replicated two polygenic resilience scores that reduce genetic risk penetrance for LOAD. We also showed that polygenic resilience scores positively correlate with polygenic risk scores in unaffected individuals, perhaps aiding in staving off disease. Our findings align with the hypothesis that a combination of risk-independent common variants mediates resilience to LOAD by moderating genetic disease risk.
Subject terms: Comparative genomics, Long-term memory, Clinical genetics
Introduction
Alzheimer’s disease (AD) is the leading cause of dementia [1]. AD exists as two genetically distinct forms: early-onset AD, which is caused by autosomal dominant mutations in one of several genes (PSEN1, PSEN2, APP, SORL1) and typically has an onset of symptoms between the ages of 40 and 60 years [2], and the more common late-onset AD (LOAD), which is sporadic, polygenic, and typically has an onset of symptoms in the mid-60s [3]. Elevated risk of LOAD is associated with a host of lifestyle factors and medical conditions, such as a high-fat diet, heavy drinking and smoking, cardiovascular disease, type-2 diabetes, and traumatic brain injury [4]. More importantly, the heritability of LOAD from twin studies was estimated at 58–79% [5], and its estimates from single-nucleotide polymorphisms (SNPs) range from 13 to 33% [6–9]. The goal of this study is to determine whether genes also play a role in resilience to LOAD. We used an innovative approach first introduced and applied in schizophrenia as a general framework for resilience research [10], focusing on individuals at the highest levels of genetic risk.
To date, genome-wide association studies (GWASs) have discovered close to 50 genome-wide significant loci (P < 5e-08) associated with LOAD risk [9, 11–20]. The ε4 allele of apolipoprotein E (APOE) is the polymorphism with the strongest effect on LOAD susceptibility [21]. Beyond APOE-ε4, there may be thousands of additional genetic polymorphisms that make small individual contributions to the overall risk for LOAD [22–25]. A polygenic risk score (PRS) [26] can be derived by summing the weighted effect of SNPs to identify a single genetic risk variable that reflects one’s relative susceptibility to LOAD. Recent LOAD PRSs capture most of the SNP heritability for LOAD [9, 24, 27]. Extensive research shows that PRSs boost the accuracy of LOAD diagnosis beyond the performance of APOE [22–25], and capture LOAD phenotypic variability not explained by APOE status [28, 29].
Revealing the genetic architecture of LOAD is vital for understanding its etiology and identifying molecular targets for innovative therapeutic interventions. Yet, knowledge of risk factors might be fruitfully complemented by an understanding of resilience-associated or -promoting mechanisms as well. As such, some AD research has shifted focus from symptomatic cases to healthy aging individuals or asymptomatic individuals at elevated risk [30]. This was motivated by the premise that high-risk asymptomatic individuals, yet unaffected, may provide clues that protect them against AD. Here, we employ the term “resilience” to indicate individuals who show better than expected outcomes in the face of high genetic risk for disease [30–35].
Increasing evidence suggests that several factors—including education, literacy, physical activity, and mental activity—can moderate the risk for LOAD [31, 32, 36], and it is estimated that one-third to 40% of dementia cases might be preventable [36, 37]. These moderation effects may be explained by reverse causation [38], but genetic influences—which are not subject to reverse causation—also underlie these factors. Educational attainment [39, 40] and, particularly, general cognitive ability [40, 41] are heritable. Thus, some of these factors may also confer resilience-enhancing genetic effects. Notably, some genetic variants, such as APOE-ε2 [42] and the APP A673T variant [43], have been identified as protective for LOAD. However, the biological mechanisms that drive the protective effects remain largely unknown. Importantly, we consider such protective effects to be fundamentally different from the “resilience” effects we sought in our study, in that protective factors are generally operative across the full range of risk, whereas resilience factors are only operative in those at the highest risk for disease. Very little work has been aimed at identifying additional genetic resilience factors that potentially moderate the genetic risk established by the cumulative effects of risk-associated alleles and their corresponding protective alleles. Genetic resilience against risk for LOAD has been investigated through diverse approaches based on varying conceptualizations and measurements used to identify individuals at high risk. As aggregation of beta-amyloid plaques and tau tangles in the brain are two of the neuropathological hallmarks of LOAD [44], a principal focus of resilience has been on asymptomatic individuals who have cognition levels that are better than predicted based on these pathologies [45–47]. Other studies have leveraged known genetic risk factors to study resilience. For example, in APOE-ε4 carriers, over a dozen SNPs have been reported to potentially facilitate resilience, such as rs10553596 in CASP7 [48] and the rs4934 nonsynonymous variant in SERPINA3 [49, 50]. However, a substantial part of the genetic risk for LOAD is neglected without incorporating the effects of genes other than APOE. Thus, although composite genetic risk indices (such as the PRS) are growing in popularity and utility, they have not been employed in the service of identifying genetic resilience for LOAD. Now, with very large numbers of LOAD samples and a more comprehensive profile of the genetic factors that confer LOAD risk, we are entering a period in which it is possible to study the interplay of genetic risk factors and genetic modifiers that reduce their penetrance.
Here, we posit the existence of common genetic variants, which have not been identified by GWAS as associated with AD as either risk or protective factors, that can help older adults remain LOAD-free despite a high genetic risk burden. We hypothesize that there exist resilience-associated variants that lower LOAD susceptibility in a manner that is statistically independent of the effects of risk-associated alleles (or their alternative protective alleles). We tested this hypothesis by capitalizing on the most comprehensive known PRS for LOAD [18] and APOE allelic status to develop two designs identifying unaffected individuals with the highest genetic likelihood of developing LOAD. Design 1 defined “resilient” individuals as normal controls with the highest PRSs for LOAD. Design 2 defined “resilient” individuals as normal controls with at least one APOE-ε4 allele and the highest LOAD PRSs (excluding the APOE region). We aimed to discover residual common genetic variants that confer resilience to unaffected individuals in the highest genetic risk tiers for LOAD. We then leveraged this profile of resilience-promoting genetic variants to build a polygenic resilience score for LOAD. We hypothesized that polygenic resilience scores would account for significant variation in affection status for LOAD among individuals with high genetic risk, and would show a significant positive correlation with PRSs in unaffected controls.
Methods
Research design
Our workflow is shown in Fig. 1. In stage 1, a recent GWAS meta-analysis for LOAD [18] was leveraged for identifying risk variants and polygenic risk scoring. In stage 2, we compared two analytic designs to identify high-risk “resilient” normal controls and “risk-matched” LOAD cases. In stage 3, a resilience GWAS was conducted for each design using the identified high-risk individuals. Then the polygenic resilience score weights were derived from resilience GWAS meta-analysis summary statistics. Finally, polygenic resilience scores were replicated in independent external studies for evaluating the performance in distinguishing high-risk “resilient” normal controls from “risk-matched” LOAD cases. The parameters of each analysis step are summarized in Supplementary Table 3.
Fig. 1. An illustration of the workflow of deriving polygenic resilience scores for late-onset Alzheimer’s disease (LOAD) for design 1 and design 2.

Stage 1: Using prior LOAD genome-wide association study (GWAS) results to calculate polygenic risk scores (PRSs). Stage 2: Identifying resilient individuals. In stage 2, we deployed two analysis designs differing in the definition of “resilient” individuals. In design 1, normal controls with LOAD PRSs ≥90th percentile were defined as “resilient” participants. In design 2, within the subset of normal controls who had at least one apolipoprotein E (APOE)-ε4 allele, a threshold of ≥80th percentile of PRSs (excluding SNPs in the APOE region) was used to define high-risk controls as “resilient”. Stage 3: Resilience GWAS and replication of polygenic resilience scores. GWAS was performed using “resilient” individuals and risk-matched affected cases from each of the two designs. For each design, polygenic resilience scores were derived and evaluated in external replication datasets. LD linkage disequilibrium, OR odds ratio, SNPs single-nucleotide polymorphisms.
Samples and genotypes
We acquired the largest available collection of genome-wide SNP data for clinically diagnosed or autopsy-confirmed LOAD to ensure adequate power. Table 1 shows the number of normal controls, LOAD cases, high-risk “resilient” normal controls, and “risk-matched” LOAD cases in each study. Summary statistics of age-at-onset (AAO) for LOAD cases and age-at-last-examination (AAE) for normal controls are presented in Supplementary Table 1. In design 1 and design 2, the mean AAE of high-risk “resilient” normal controls and the mean AAO of “risk-matched” LOAD cases ranged from 70.3 to 80.9, and there were no significant age differences between groups. A common lower bound for AAO of LOAD is 65; however, the age cutoff has no specific biological significance [3], and many genetic studies of LOAD have included cases with AAO as low as 60 (and the same AAE for unaffected comparison subjects). Therefore, we included participants in our analysis having AAO/AAE ≥ 60 years old. The full name and accessibility of each study can be found in Supplementary Table 2. All 26 studies in the discovery stage came from the stage-1 AD GWAS meta-analysis of Kunkle et al. [18]. The eight studies in the replication stage are fully independent of the discovery studies. Full descriptions of the discovery and replication samples were published previously [9, 17, 18, 51, 52]. Genotypes for all studies were imputed using the Haplotype Reference Consortium (HRC) r1.1 2016 reference panel [53]. Detailed quality control (QC) steps for samples and genotypes are described in Supplementary Methods.
Table 1.
The number of LOAD cases and normal controls, high-risk normal controls (“resilient” individuals), and risk-matched LOAD cases identified in each of the discovery and replication studies.
| Design 1 | Design 2 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Normal controls | LOAD cases | Normal controls | LOAD cases | |||||||||||
| Study | Sub-study | N High-risk | N Total | % Retained | N Risk-matched | N Total | % Retained | N High-risk | N Total | % Retained | N Risk-matched | N Total | % Retained | |
| Discovery | ADGC | ADC1 | 52 | 512 | 10.2 | 842 | 1524 | 55.2 | 30 | 150 | 20.0 | 773 | 1021 | 75.7 |
| ADC2 | 16 | 155 | 10.3 | 386 | 620 | 62.3 | 9 | 41 | 22.0 | 212 | 353 | 60.1 | ||
| ADC3 | 57 | 566 | 10.1 | 379 | 666 | 56.9 | 27 | 132 | 20.5 | 357 | 391 | 91.3 | ||
| ADC4 | 38 | 376 | 10.1 | 222 | 304 | 73.0 | 20 | 100 | 20.0 | 115 | 160 | 71.9 | ||
| ADC5 | 51 | 503 | 10.1 | 187 | 285 | 65.6 | 24 | 119 | 20.2 | 147 | 178 | 82.6 | ||
| ADC6 | 34 | 337 | 10.1 | 68 | 213 | 31.9 | 19 | 93 | 20.4 | 94 | 121 | 77.7 | ||
| MAYO | 112 | 1117 | 10.0 | 626 | 754 | 83.0 | 63 | 313 | 20.1 | 436 | 492 | 88.6 | ||
| UMVUMSSM | 113 | 1108 | 10.2 | 753 | 1123 | 67.1 | 51 | 251 | 20.3 | 430 | 661 | 65.1 | ||
| ACT + GenDiff | 157 | 1556 | 10.1 | 425 | 524 | 81.1 | 63 | 314 | 20.1 | 198 | 234 | 84.6 | ||
| MIRAGE | 51 | 509 | 10.0 | 101 | 137 | 73.7 | 38 | 190 | 20.0 | 68 | 78 | 87.2 | ||
| WASHU1 | 18 | 176 | 10.2 | 137 | 253 | 54.2 | 10 | 48 | 20.8 | 65 | 141 | 46.1 | ||
| ROSMAP | 72 | 717 | 10.0 | 192 | 237 | 81.0 | 23 | 114 | 20.2 | 18 | 87 | 20.7 | ||
| UPITT | 82 | 811 | 10.1 | 794 | 1152 | 68.9 | 32 | 158 | 20.3 | 545 | 664 | 82.1 | ||
| OHSU | 18 | 180 | 10.0 | 119 | 174 | 68.4 | 5 | 25 | 20.0 | 49 | 73 | 67.1 | ||
| Tgen II | 36 | 360 | 10.0 | 364 | 613 | 59.4 | 16 | 76 | 21.1 | 251 | 396 | 63.4 | ||
| NIA-LOAD | 102 | 1007 | 10.1 | 582 | 760 | 76.6 | 58 | 289 | 20.1 | 463 | 571 | 81.1 | ||
| ROSMAP2 | 22 | 214 | 10.3 | 50 | 59 | 84.7 | 3 | 15 | 20.0 | 0 | 18 | 0.0 | ||
| WASHU2 | 8 | 71 | 11.3 | 29 | 36 | 80.6 | 4 | 16 | 25.0 | 16 | 19 | 84.2 | ||
| MTC | 19 | 188 | 10.1 | 220 | 252 | 87.3 | 5 | 22 | 22.7 | 48 | 143 | 33.6 | ||
| TARCC | 18 | 176 | 10.2 | 241 | 306 | 78.8 | 10 | 47 | 21.3 | 122 | 184 | 66.3 | ||
| WHICAP | 56 | 553 | 10.1 | 17 | 72 | 23.6 | 23 | 113 | 20.4 | 8 | 16 | 50.0 | ||
| ADNI | ADNI-1 | 13 | 127 | 10.2 | 174 | 350 | 49.7 | 6 | 28 | 21.4 | 97 | 230 | 42.2 | |
| CHARGE | CHS | 168 | 1661 | 10.1 | 274 | 451 | 60.8 | 66 | 330 | 20.0 | 99 | 148 | 66.9 | |
| FHS | 191 | 1901 | 10.0 | 137 | 288 | 47.6 | 74 | 370 | 20.0 | 54 | 94 | 57.4 | ||
| EADI | 620 | 6172 | 10.0 | 1636 | 2167 | 75.5 | 247 | 1235 | 20.0 | 870 | 1072 | 81.2 | ||
| GERAD | 139 | 1388 | 10.0 | 2354 | 2992 | 78.7 | 62 | 310 | 20.0 | 1006 | 1623 | 62.0 | ||
| N Total | 2263 | 22441 | 10.1 | 11309 | 16312 | 69.3 | 988 | 4899 | 20.2 | 6541 | 9168 | 71.3 | ||
| Replication | AddNeuroMed | 19 | 183 | 10.4 | 38 | 223 | 17.0 | 9 | 44 | 20.5 | 19 | 121 | 15.7 | |
| ADGC | ADC7 | 79 | 784 | 10.1 | 45 | 513 | 8.8 | 51 | 252 | 20.2 | 57 | 320 | 17.8 | |
| ADNI | ADNI-GO/2/3 | 38 | 374 | 10.2 | 37 | 211 | 17.5 | 23 | 113 | 20.4 | 38 | 142 | 26.8 | |
| PGC-ALZ | Norwegian DemGene Network | 75 | 741 | 10.1 | 163 | 1085 | 15.0 | 34 | 168 | 20.2 | 130 | 671 | 19.4 | |
| GENDER/SATSA/HARMONY | 77 | 764 | 10.1 | 39 | 306 | 12.7 | 40 | 196 | 20.4 | 34 | 148 | 23.0 | ||
| TwinGene | 608 | 6080 | 10.0 | 30 | 287 | 10.5 | 353 | 1765 | 20.0 | 31 | 156 | 19.9 | ||
| AIBL | 77 | 762 | 10.1 | 13 | 111 | 11.7 | 36 | 180 | 20.0 | 12 | 71 | 16.9 | ||
| Sydney MAS | 83 | 830 | 10.0 | 16 | 95 | 16.8 | 37 | 182 | 20.3 | 10 | 31 | 32.3 | ||
| N Total | 1056 | 10518 | 10.0 | 381 | 2831 | 13.5 | 583 | 2900 | 20.1 | 331 | 1660 | 19.9 | ||
LOAD late-onset Alzheimer’s disease.
Note: “Retained” column indicates the percentage of high-risk normal controls of all normal controls retained for resilience genome-wide association analysis per study, or the percentage of risk-matched LOAD cases of all LOAD cases retained in analysis per study. ROSMAP2 study had no LOAD cases (in italic) whose risk matched with high-risk normal controls in design 2, and was not included in the analysis for design 2.
A list of study full names is in Supplementary Table 2.
Identifying individuals at high genetic risk
In design 1, a PRS was used to select individuals with high genetic risk. At the time of deploying our analyses, the Kunkle et al., 2019 study [18] was the largest publicly available GWAS using clinically diagnosed or autopsy-confirmed AD cases and CN controls, as opposed to proxy AD cases and controls that might lead to inaccurate risk estimation [27]. Therefore, we consider that this study will give the most accurate measure of AD risk and derived the PRS weights from its stage-1 AD GWAS meta-analysis summary statistics [18]. See Supplementary Methods for further details. The variance in AD explained by PRS maximizes at a P-value threshold of 0.5 in participants from GERAD (Genetic and Environmental Risk for Alzheimer’s disease) [23] and 22 locally available ADGC (Alzheimer’s Disease Genetics Consortium) studies (Supplementary Fig. 4). We, therefore, adopted this threshold to ensure that our risk measure captures as much of the genetic risk for AD as possible. This very conservative threshold will thus ensure that potential risk SNPs with even very small effect sizes will not be advanced for consideration as resilience SNPs. However, if studies other than Kunkle et al. 2019 were used to estimate genetic risk for AD, it may be the case that smaller P-value thresholds may be optimal (e.g., 5e−08, 1e−05, 0.1) [9, 22, 54, 55]. Within each study, LOAD cases and normal controls were ranked based on their PRSs. Note that as the true prevalence of resilience to AD in the population is unknown, we adopted the same high-risk percentile cutoff that proved effective in our original workflow [10], and classified the 10% of controls with the highest PRSs as “resilient”. The LOAD cases whose risk scores were between the 90th percentile and the maximum PRS in controls were retained as risk-matched LOAD cases for comparison.
In design 2, we restricted the analysis to APOE-ε4 carriers. APOE and its flanking region (chr19: 44,400 kb–46,500 kb) [23] were removed from the PRS. As this analysis was restricted to fewer individuals due to the APOE-ε4 stratification, we chose a more lenient high-PRS cutoff (80th percentile) for identifying “resilient” individuals to retain more participants and preserve power. In this design, “resilient” normal controls were identified as those with at least one APOE-ε4 allele, and a risk score ranked at ≥80th percentile. Risk-matched LOAD cases were defined as APOE-ε4 carriers whose PRSs fell within the high-PRS range of “resilient” normal controls.
Derivation, replication, and statistical analysis of polygenic resilience scores
GWASs of resilience were performed using logistic regression with Plink (version 1.9) [56]. Selected principal components, AAO/AAE, and sex were used as covariates. A GWAS meta-analysis was conducted in METAL [57] software using an inverse-variance random-effect model with genomic control. In accord with the pipeline described by Hess et al. [10], SNPs known to be associated with LOAD risk were excluded from the resilience-scoring algorithm; these were defined as those SNPs that showed an association with AD risk (P < 0.5) from the GWAS meta-analysis summary statistics [18], and variants that were in linkage disequilibrium (LD) (r2 ≥ 0.2 in a 1-Mb window) with those risk variants with associations of P < 0.5. This pruning step of excluding risk variants from consideration as resilience loci serves as a conservative measure to avoid re-discovering risk variants for resilience scoring. For both resilience designs, the polygenic resilience score weights were generated from the marginal SNPs of resilience GWAS meta-analysis summary statistics following the same series of QC steps (see Supplementary Methods).
Polygenic resilience scores were derived for 10 P-value thresholds, in a manner similar to the PRS algorithm, by summing up the weighted effective allele counts of SNPs [26]. Logistic regression was used to assess the likelihood of “resilient” group inclusion based on harboring a higher polygenic resilience score. Selected principal components, AAO/AAE and sex were used as covariates. For each polygenic resilience score, we meta-analyzed the natural logarithm of the odds ratio (OR) of being a high-risk “resilient” normal control versus a risk-matched LOAD case using a random-effects inverse-variance model using the R package metafor, and pooled variance explained in resilience across independent replication studies. All tests were two-tailed unless specified otherwise. See Supplementary Methods for further details.
Results
Resilience GWAS
Design 1 produced 2263 high-risk “resilient” normal controls and 11,309 risk-matched LOAD cases for the resilience GWAS meta-analysis. As expected, the sample size retained in design 2 was smaller, totalling 988 high-risk “resilient” normal controls and 6541 risk-matched LOAD cases (Table 1). Because our analytic approaches used only subsets of all available LOAD case–control GWAS data, we neither had nor anticipated having sufficient power to detect individual SNPs with genome-wide significant association with resilience (Supplementary Fig. 3). Instead, our focus was on deriving and evaluating polygenic resilience scores. As a necessary step to generate SNP-weights for summation in those scores, we performed individual-SNP association tests and briefly reported the results in Supplementary Results.
Replication and evaluation of polygenic resilience scores
After removing risk-associated SNPs (P < 0.5) and SNPs in LD with those risk-associated SNPs (r2 ≥ 0.2), clumping the remaining marginal SNPs, and applying QC steps, a profile of 18,723 SNPs was included in the resilience score for design 1, and 18,122 SNPs in design 2. Resilience scores for all 10 P-value thresholds were significantly associated with “resilient” group inclusion (“resilient” normal controls versus risk-matched LOAD cases) when tested in locally downloaded discovery datasets. Results of the association between “resilient” group inclusion and polygenic resilience scores from the replication datasets were meta-analyzed, yielding 1056 high-risk “resilient” normal controls and 381 risk-matched LOAD cases in design 1, and 583 high-risk “resilient” normal controls and 331 risk-matched LOAD cases in design 2 (Table 1).
In design 1, the meta-analysis found significant replication of the association between “resilient” group inclusion and polygenic resilience scores at two P-value thresholds (P < 0.1, P < 0.2) (Fig. 2A). The most significant association was found for the polygenic resilience score containing all independent marginal SNPs with resilience GWAS P < 0.1 (OR = 1.24, 95% confidence interval [CI] = 1.05–1.47, P = 0.010). Resilience scores for the 0.1 P-value threshold explained an average of 1.3% (standard deviation [58] = 5.3%) of the variance in “resilient” group inclusion or 1.2% (SD = 4.3%) (Fig. 2B) of the variance on the liability scale, i.e., SNP heritability of resilience. No significant (P < 0.05) replication of the association between “resilient” group inclusion and polygenic resilience scores was observed for any of the 10 polygenic resilience scores in design 2.
Fig. 2. The performance of polygenic resilience scores in capturing resilience variability in independent replication studies.

In design 1, normal controls with late-onset Alzheimer’s disease (LOAD) polygenic risk scores (PRSs) ≥90th percentile were defined as “resilient” participants. In design 2, a threshold of ≥80th percentile of PRSs (excluding SNPs in the apolipoprotein E [APOE] region) was used to define high-risk controls as “resilient” within the normal controls who have at least one APOE-ε4 allele. A, B design 1 (high-risk normal controls, n = 1,056; risk-matched LOAD cases, n = 381). C, D Design 2 (high-risk normal controls, n = 583; risk-matched LOAD cases, n = 331). The odds ratio (OR) and variance explained by polygenic resilience scores reflect meta-analytic results from independent replication samples. Nagelkerke’s pseudo-R2 values on the liability scale are weighted average using the weights from the meta-analysis of ORs. The dot-plots (A, C) show corresponding ORs for resilience scores across 10 P-value thresholds, wherein OR > 1.0 indicates higher resilience scores are associated with a higher likelihood of being a high-risk normal control (“resilient” individual) than being a risk-matched LOAD case. Error bars represent the 95% confidence intervals (CI) around each OR, which are the exponent of the 95% CI of β coefficients. The barplots (B, D) show the amount of variance in resilience (i.e., “resilient” high-risk normal controls versus risk-matched LOAD cases) on the liability scale that is explained by resilience scores. Asterisks (*) indicate P values <0.05 for ORs >1.0.
Note that the association between “resilient” group inclusion and polygenic resilience scores (P < 0.1, P < 0.2 of design 1) was not significant after multiple-testing correction for 10 P-value thresholds using the false discovery rate (FDR) or Bonferroni method. However, considering polygenic resilience scores were derived by aggregating SNPs within a series of escalating P-value thresholds, they were nested models and not independent. Therefore, a typical FDR or Bonferroni correction under the assumption of independence would be overly conservative.
Interaction of risk and resilience effects
In the full samples from three locally downloaded replication studies (Alzheimer Disease Centers Wave 7 [ADC7], AddNeuroMed, and Alzheimer’s Disease Neuroimaging Initiative stage GO/2/3 [ADNI-GO/2/3]; normal controls, n = 1321; LOAD cases, n = 943) (Table 1 and Supplementary Table 1), we tested for correlations between PRSs and polygenic resilience scores. As hypothesized, the standardized polygenic resilience scores of the optimal P < 0.1 threshold in design 1 exhibited a significant positive correlation with PRSs in normal controls (Pearson’s r = 0.102, 95% CI = 0.048–0.155, degree of freedom [df]=1319, P = 2.1e-04), and no significant correlation was observed in LOAD cases (Pearson’s r = 0.022, 95% CI = −0.042–0.085, df = 941, P = 0.51) (Fig. 3). As expected, the correlation coefficient between polygenic risk scores and polygenic resilience scores in normal controls was significantly larger than the one in LOAD cases (P = 0.03, one-tailed test).
Fig. 3. The correlation of standardized polygenic risk scores (PRSs) and polygenic resilience scores (design 1) in normal controls and late-onset Alzheimer’s disease (LOAD) cases.
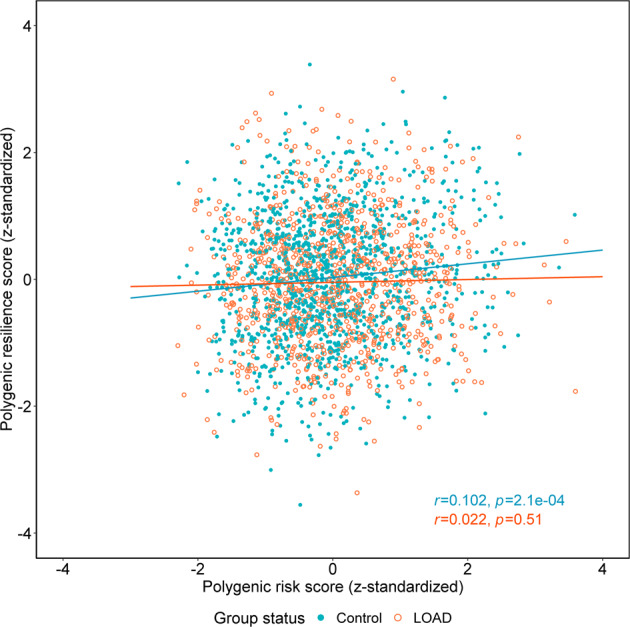
The analyses were performed in three independent replication studies not used in the resilience score derivation steps (i.e., ADC7, AddNeuroMed, and ADNI-GO/2/3; normal controls, n = 1321; LOAD cases, n = 943). The optimal P-value threshold for polygenic risk-scoring was 0.5, and the optimal P-value threshold for polygenic resilience scoring was 0.1 (see Fig. 2). The blue round dots indicate normal controls, and the orange circles indicate LOAD cases. The blue and orange lines represent the best fit for correlations between PRSs and resilience scores in normal controls and in LOAD cases, respectively. The blue and orange annotation text shows the Pearson correlation coefficient (r) and the P-value between PRSs and resilience scores in normal controls and LOAD cases, respectively. In this analysis, we excluded ultra-high-risk LOAD cases whose PRSs are higher than the maximum of all normal controls, and ultra-low-risk normal controls whose PRSs are lower than the minimum of all LOAD cases.
Discussion
We applied a validated analytic framework to detect common variants that, when combined into a polygenic resilience score, are associated with lower LOAD risk penetrance among older individuals with relatively high genetic risk of disease. We found reliable evidence to reinforce the notion that unaffected individuals with higher genetic risk loads may be protected from complex diseases, such as LOAD, by the collective effects of risk-independent common variants that reduce the penetrance of one’s overall genetic risk burden. Identifying genetic factors that moderate risk penetrance may prove valuable for explaining the missing heritability and etiologic heterogeneity of LOAD, which in turn could shed light on pathophysiological mechanisms and eventually lead to better interventions and preventive treatments.
Risk-countering effects of polygenic resilience scores
Individuals with higher polygenic resilience scores (P < 0.1 and P < 0.2 thresholds of design 1) had higher odds of being a “resilient” high-risk normal control than a risk-matched LOAD case. Polygenic resilience scores (design 1) significantly increased with higher PRSs in normal controls, but not in LOAD cases. Taken together, these results support the hypothesis that polygenic resilience scores capture risk-countering polygenic effects against the penetrance of high polygenic risk for LOAD, and that normal controls with higher PRSs are protected from LOAD by harboring correspondingly higher polygenic resilience scores. Although no polygenic resilience scores in design 2 demonstrated significant risk-buffering effects, we cannot rule out the possibility that common variants might reduce risk penetrance in normal controls with enriched risk from both APOE and PRSs. In fact, among APOE-ε4 carriers, higher resilience scores in design 1 at the P < 0.1 threshold (OR = 1.64, 95% CI = 1.08–2.50, P = 0.021) and the P < 0.2 threshold (OR = 1.98, 95% CI = 1.24–3.15, P = 3.9e-03) were associated with higher odds of being a “resilient” high-PRS normal control than a risk-matched LOAD case. Among “resilient” high-PRS controls, higher resilience scores in design 1 (P < 0.2 threshold) were significantly associated with increased odds of carrying at least one APOE-ε4 allele (OR = 1.57, 95% CI = 1.07–2.29, P = 0.021). A similar trend was observed when the P < 0.1 threshold was used, although this was not significant (OR = 1.30, 95% CI = 0.90–1.89, P = 0.16) (Supplementary Results). We, therefore, conclude that polygenic resilience scores may moderate the risk effects of the LOAD PRS generally, and the APOE-ε4 allele specifically. However, these analyses were carried out in relatively small studies (ADC7, AddNeuroMed, and ADNI-GO/2/3), and need to be repeated in larger, more powerful, replication samples.
Interplay of polygenic effects and APOE
In design 2, we hypothesized that a two-stage selection of individuals (with both higher PRSs and one or more APOE-ε4 alleles) would enrich for individuals with the absolute highest genetic risk for LOAD [59, 60]; yet, there was a substantial reduction in the performance of design 2 in contrast to design 1. The lack of significant replication of association with resilience in design 2 simply might be due to lower statistical power in both the resilience score development and replication stages, considering the total sample size of design 2 is approximately half that of design 1. Alternatively, resilience-promoting variants may be found among APOE-ε4 carriers through broader exploration of the model-parameter space (e.g., PRS threshold in particular), separate evaluation of APOE-ε4 homozygotes and various heterozygote combinations, and more accurate modeling of the genetic architecture of resilience (see limitations below). An important question future studies should address is to what extent common variants may influence the penetrance of genetic risk in larger samples of APOE-ε4 carriers, or whether the prevalence of risk-modifying common variants differs between APOE-ε4 carriers and noncarriers.
On the other hand, multiple studies [22–25, 28, 54, 61–63] have revealed that PRSs capture independent risk effects beyond APOE alone, while few studies have explored the risk-predictive performance of PRSs stratified by APOE status. Higher PRSs were found to be associated with increased susceptibility for LOAD in APOE-ε4 noncarriers [25, 29, 59]. Furthermore, the risk effects of PRS deciles across APOE status could be dependent on the ages of participants [29, 59, 62, 64]. Further mining of the complex relationship between the risk effects of PRSs and APOE is outside the scope of the current study; however, further investigations on the penetrance of high PRSs among APOE-ε4 carriers and noncarriers seem warranted.
Strengths and limitations
Our approach has identified candidate resilience loci that may ultimately serve as targets for the promotion of resilience. We examined the performance of two polygenic resilience scores: design 1 selected participants with the highest polygenic risk regardless of APOE-ε4 status, while design 2 restricted analyses to APOE-ε4 carriers. To our knowledge, this is the first study to identify a polygenic resilience score for genetic LOAD risk, comprising thousands of risk-independent common variants that partially offset the genetic risk conferred by a relatively high PRS. An important distinction of the current study relative to prior work on genetic resilience to LOAD is that we accounted not only for the risk from APOE but also the aggregate effect of thousands of additional risk variants throughout the genome via the LOAD PRS.
A conservative variant-filtering strategy was applied, which resulted in the removal of common variants associated with LOAD risk variants (risk association P < 0.5) and those in liberal LD (r2 > 0.2) with LOAD risk variants. A strength of this approach is that we ensured the polygenic resilience scores derived in the current study are independent of the risk scores so that the SNPs comprising the polygenic resilience score are not sub-threshold risk SNPs. Our design of defining “resilient” groups from the same risk background instead of contrasting high-risk normal controls with low-risk LOAD cases also helped avoid re-discovering variants merely associated with risk. In addition, the resilience alleles of these risk-residual SNPs are not simply protective alleles defined in a risk framework, where each biallelic locus is defined by both a risk allele and a corresponding and opposing protective allele. Thus, this strategy helps identify resilience effects that are conditioned on net risk effects, owing to the combination of risk and protective alleles summed in polygenic risk scores. Yet, although our approach is conservative, it is limited in the identification of a better-performing resilience score because most of the genome has been discarded from the analysis. Biologically, it is plausible that variants nearby risk loci, such as those in the same LD block or in the same gene with risk SNPs, could exert modifying functions [65]. Our conservative strategy, discarding all SNPs with any semblance of risk association, and those in liberal LD threshold with such SNPs, consequently leads to lower power in uncovering variants with potentially higher biological functionality. This notion is borne out in the fact that no significant gene-ontology pathways were enriched by resilience-related common variants identified in this study (results not shown). With larger samples, resilience-conferring SNPs may be investigated using a stricter LD threshold (e.g., r2 > 0.1) to further restrain the “hitchhiking” of risk variants. More importantly, Mendelian randomization, conditional association testing, or simulation analysis may be better suited to evaluate the hypothesis that resilience signals are more likely to co-localize with risk loci or genes. In addition, filtering variants by LD with risk SNPs results in a low LD structure among the remaining SNPs as demonstrated previously [10], which diminishes our capacity to examine the genetic correlation of resilience to LOAD with other risk- or resilience-related phenotypes (e.g., via LD score regression). A high priority should be placed on the design of new methods that can detect resilience-associated SNPs that may reside in regions of strong LD with risk variants.
Resilience was defined by discrete groups in our analysis, which truncated effective sample sizes to the upper tail of the risk distribution. Choosing a lower percentile cutoff would increase the sample size available for the resilience analysis, while potentially diminishing the signal of resilience genes. In the future, when larger samples are available, higher risk thresholds may be applied and subgroups at more extreme risk could be leveraged to increase power. It is an important task for future studies to investigate which of these factors (i.e., sample size, signal: noise) would have the greater effect on power, and to better understand the prevalence of resilience to AD in the population. Theoretically, resilience may be a continuous measure; thus, our resilience approach might also be improved by leveraging all study samples and modeling the continuity of resilience using either linear or non-linear analysis. Despite the restricted sample sizes in the current study, two resilience scores in design 1 were sufficiently robust to replicate significantly in fully independent studies. Further replication would be key to testing the validity of these resilience scores. It is expected that the strength of our results (in terms of variance explained and the significance of associations) will only increase with the addition of more samples.
Several studies [9, 22, 54, 55] indicated that polygenic risk scores of P-value thresholds less than 0.5 (i.e., 5e−08, 1e−05, 0.1) might show better performance in predicting LOAD risk. Therefore, it may be valuable to compare the performance of resilience scores developed from risk scores at other p-value thresholds. In addition, it is likely that a subgroup of “resilient” normal controls identified in this work will eventually develop LOAD, but with later onset. Thus, all resilient participants demonstrate resilience against high levels of genetic risk for LOAD, but only those who never develop LOAD are additionally resistant against the disease itself. Lastly, the participants in our analysis were of European ancestry, so the degree of generalization of our results to non-European populations is presently unknown.
Future directions
Two analysis designs were deployed in the current study to select individuals with a high genetic risk burden from both PRSs and APOE, and other methods could be devised to expand the capabilities of our resilience approach in LOAD. It has been suggested, for example, that using a PRS with the APOE region removed and adding APOE alleles as a covariate may boost the performance of LOAD risk prediction [66], compared with incorporating APOE alleles as weighted SNPs in PRSs. In addition, it could be important to include the number of APOE-ε4 or ε2 alleles as covariates in resilience analysis models to better reflect the relative risk levels among individuals. In our study, we consider it important to utilize the most comprehensive risk profile of LOAD to identify resilient individuals, i.e., normal controls with the highest genetic risk from all sources. Future studies may be interested in examining the resilience effects that moderate a portion of the LOAD risk. For example, resilience to the risk effects of APOE-ε4 alone can be studied by defining all APOE-ε4 carrying (or homozygotic) normal controls as resilient. To examine whether the resilience scores remain predictive if the APOE region is excluded from the PRS, the resilience to residual polygenic risk effects excluding APOE can be investigated in normal controls without APOE-ε4, who land in the top percentiles of PRSs (excluding the APOE region). Previous studies [42, 67] demonstrated that women carrying APOE-ε4 alleles were at greater risk of developing AD than men with the same APOE-ε4 dosages, especially between the ages of 65 and 75. When larger sample sizes are available, limiting our analysis to females in design 2 may further enrich for high-risk individuals and increase resilience signals.
Potentially, polygenic resilience scores from the current study could be applied to other resilience-related questions. For example, it would be instrumental in discovering the extent to which polygenic resilience score is associated with other phenotypes that have been associated with resilience to LOAD risk (e.g., education, general cognitive ability in early life, and other indices of cognitive reserve, brain reserve, or brain maintenance) [31, 32, 68–72]. In follow-up studies, it might be illuminating to investigate whether these resilience-promoting genetic factors show protective effects for cognitive impairment or LOAD-related pathophysiological changes.
Conclusion
We found evidence to support the hypothesis that thousands of risk-independent common variants underlie resilience among unaffected individuals with higher genetic risk for LOAD. We conclude that common variants not in LD with known LOAD risk variants exert a protective effect on LOAD risk. Our findings provide a significant and novel contribution to the existing understanding of genetic resilience to LOAD risk. This novel approach highlights a window of opportunity for identifying risk-modifying biological mechanisms and potential pathways for intervention in populations at the highest risk for LOAD.
Supplementary information
Acknowledgements
We thank all the participants of this study for their contributions. Our effort on this project was supported by the U.S. National Institute on Aging [grant numbers R01AG064955 and R01AG054002]. Additional acknowledgements and detailed acknowledgements of funding sources for the study are provided in Supplementary acknowledgements.
Author contributions
JH, JLH, CCF, SVF, CFN, SJL, VEP, PH, SS, MTT, WSK, and SJG contributed to the conception and design of the study. The Alzheimer’s Disease Neuroimaging Initiative (ADNI), NA, JCB, BGB, IKK, GL, KN, EKO, AS, AT, QY, OAA, HB, MG, NAK, JCL, SML, CLM, KAM, NLP, DP, PSS, JW, PH, SS, and WSK were involved in the acquisition of data. JH performed the analyses of this study. JCB, BGB, IKK, GL, EKO, AS, AT, and QY performed the statistical analyses in non-local datasets and provided summary statistics. JH, JLH, SVF, VEP, PH, SS, WSK, and SJG were involved in the interpretation of data. JH wrote the first draft of the manuscript and SJG provided critical edits. JLH, BGB, IKK, EKO, AS, OAA, MG, SML, KAM, PSS, JW, SVF, CFN, VEP, PH, and WSK provided the critical review of the manuscript. All authors gave final approval of the version of the manuscript submitted for peer review.
Data availability
The Alzheimer’s Disease Genetics Consortium, the Alzheimer’s Disease Neuroimaging Initiative, and the AddNeuroMed data used in this study were provided under restricted access by NIAGADS (https://www.niagads.org), ADNI (http://adni.loni.usc.edu), and Synapse platform (https://www.synapse.org/#!Synapse:syn4907804), respectively. Only summary statistics were made available to us from the European Alzheimer’s Disease Initiative, the Genetic and Environmental Risk in Alzheimer’s Disease, the Cohorts for Heart and Aging Research in Genomic Epidemiology, the Psychiatric Genomic Consortium, the Australian Imaging, Biomarker & Lifestyle Study, and the Sydney Memory and Ageing Study. The resilience-scoring weights of design 1 are available from the corresponding author upon request.
Code availability
The code for data cleaning and analysis, and the resilience-scoring weights of design 1 are available from the corresponding author upon request.
Competing interests
OAA is a consultant to HealthLytix. SVF in the past year, received income, potential income, travel expenses continuing education support and/or research support from Aardvark, Akili, Genomind, Ironshore, KemPharm/Corium, Noven, Ondosis, Otsuka, Rhodes, Supernus, Takeda, Tris, and Vallon. With his institution, he has US patent US20130217707 A1 for the use of sodium-hydrogen exchange inhibitors in the treatment of ADHD. In previous years, he received support from: Alcobra, Arbor, Aveksham, CogCubed, Eli Lilly, Enzymotec, Impact, Janssen, Lundbeck/Takeda, McNeil, NeuroLifeSciences, Neurovance, Novartis, Pfizer, Shire, and Sunovion. He also receives royalties from books published by Guilford Press: Straight Talk about Your Child’s Mental Health; Oxford University Press: Schizophrenia: The Facts; and Elsevier: ADHD: Non-Pharmacologic Interventions. He is also Program Director of www.adhdinadults.com. He is supported by the European Union’s Horizon 2020 research and innovation programme under grant agreement No 965381; NIMH grants U01AR076092-01A1, 1R21MH1264940, R01MH116037; Oregon Health and Science University, Otsuka Pharmaceuticals, Noven Pharmaceuticals Incorporated, and Supernus Pharmaceutical Company. The remaining authors declare no competing interests.
Footnotes
Consortium author information: For the Alzheimer’s Disease Neuroimaging Initiative, data used in the preparation of this article were obtained from the Alzheimer’s Disease Neuroimaging Initiative (ADNI) database (adni.loni.usc.edu). As such, the investigators within the ADNI contributed to the design and implementation of ADNI and/or provided data but did not participate in the analysis or writing of this report. A complete listing of ADNI investigators can be found at: http://adni.loni.usc.edu/wp-content/uploads/how_to_apply/ADNI_Acknowledgement_List.pdf.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Members of the Alzheimer’s Disease Neuroimaging Initiative are listed in consortium author information.
Supplementary information
The online version contains supplementary material available at 10.1038/s41398-022-02055-0.
References
- 1.Alzheimer’s Association. Alzheimer’s disease facts and figures. Alzheimers Dement. 2021;17:327–406. [DOI] [PubMed]
- 2.Ryan NS, Nicholas JM, Weston PSJ, Liang Y, Lashley T, Guerreiro R, et al. Clinical phenotype and genetic associations in autosomal dominant familial Alzheimer’s disease: a case series. Lancet Neurol. 2016;15:1326–35. doi: 10.1016/S1474-4422(16)30193-4. [DOI] [PubMed] [Google Scholar]
- 3.Rossor MN, Fox NC, Mummery CJ, Schott JM, Warren JD. The diagnosis of young-onset dementia. Lancet Neurol. 2010;9:793–806. doi: 10.1016/S1474-4422(10)70159-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Edwards Iii GA, Gamez N, Escobedo G, Jr, Calderon O, Moreno-Gonzalez I. Modifiable risk factors for Alzheimer’s disease. Front Aging Neurosci. 2019;11:146. doi: 10.3389/fnagi.2019.00146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Gatz M, Reynolds CA, Fratiglioni L, Johansson B, Mortimer JA, Berg S, et al. Role of genes and environments for explaining Alzheimer disease. Arch Gen Psychiatry. 2006;63:168–74. doi: 10.1001/archpsyc.63.2.168. [DOI] [PubMed] [Google Scholar]
- 6.Lee SH, Harold D, Nyholt DR, Consortium AN, International Endogene C, Genetic. et al. Estimation and partitioning of polygenic variation captured by common SNPs for Alzheimer’s disease, multiple sclerosis and endometriosis. Hum Mol Genet. 2013;22:832–41. doi: 10.1093/hmg/dds491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ridge PG, Mukherjee S, Crane PK, Kauwe JS. Alzheimer’s disease genetics C. Alzheimer’s disease: analyzing the missing heritability. PLoS ONE. 2013;8:e79771. doi: 10.1371/journal.pone.0079771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Brainstorm C, Anttila V, Bulik-Sullivan B, Finucane HK, Walters RK, Bras J, et al. Analysis of shared heritability in common disorders of the brain. Science. 2018;360:eaap8757.. doi: 10.1126/science.aap8757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Zhang Q, Sidorenko J, Couvy-Duchesne B, Marioni RE, Wright MJ, Goate AM, et al. Risk prediction of late-onset Alzheimer’s disease implies an oligogenic architecture. Nat Commun. 2020;11:4799. doi: 10.1038/s41467-020-18534-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hess JL, Tylee DS, Mattheisen M, Schizophrenia Working Group of the Psychiatric Genomics C, Lundbeck Foundation Initiative for Integrative Psychiatric R. Borglum AD, et al. A polygenic resilience score moderates the genetic risk for schizophrenia. Mol Psychiatry. 2021;26:800–15. doi: 10.1038/s41380-019-0463-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Harold D, Abraham R, Hollingworth P, Sims R, Gerrish A, Hamshere ML, et al. Genome-wide association study identifies variants at CLU and PICALM associated with Alzheimer’s disease. Nat Genet. 2009;41:1088–93. doi: 10.1038/ng.440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lambert JC, Heath S, Even G, Campion D, Sleegers K, Hiltunen M, et al. Genome-wide association study identifies variants at CLU and CR1 associated with Alzheimer’s disease. Nat Genet. 2009;41:1094–9. doi: 10.1038/ng.439. [DOI] [PubMed] [Google Scholar]
- 13.Hollingworth P, Harold D, Sims R, Gerrish A, Lambert JC, Carrasquillo MM, et al. Common variants at ABCA7, MS4A6A/MS4A4E, EPHA1, CD33 and CD2AP are associated with Alzheimer’s disease. Nat Genet. 2011;43:429–35. doi: 10.1038/ng.803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Naj AC, Jun G, Beecham GW, Wang LS, Vardarajan BN, Buros J, et al. Common variants at MS4A4/MS4A6E, CD2AP, CD33 and EPHA1 are associated with late-onset Alzheimer’s disease. Nat Genet. 2011;43:436–41. doi: 10.1038/ng.801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lambert JC, Ibrahim-Verbaas CA, Harold D, Naj AC, Sims R, Bellenguez C, et al. Meta-analysis of 74,046 individuals identifies 11 new susceptibility loci for Alzheimer’s disease. Nat Genet. 2013;45:1452–8. doi: 10.1038/ng.2802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Marioni RE, Harris SE, Zhang Q, McRae AF, Hagenaars SP, Hill WD, et al. GWAS on family history of Alzheimer’s disease. Transl Psychiatry. 2018;8:99. doi: 10.1038/s41398-018-0150-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Jansen IE, Savage JE, Watanabe K, Bryois J, Williams DM, Steinberg S, et al. Genome-wide meta-analysis identifies new loci and functional pathways influencing Alzheimer’s disease risk. Nat Genet. 2019;51:404–13. doi: 10.1038/s41588-018-0311-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kunkle BW, Grenier-Boley B, Sims R, Bis JC, Damotte V, Naj AC, et al. Genetic meta-analysis of diagnosed Alzheimer’s disease identifies new risk loci and implicates Abeta, tau, immunity and lipid processing. Nat Genet. 2019;51:414–30. doi: 10.1038/s41588-019-0358-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wightman DP, Jansen IE, Savage JE, Shadrin AA, Bahrami S, Holland D, et al. A genome-wide association study with 1,126,563 individuals identifies new risk loci for Alzheimer’s disease. Nat Genet. 2021;53:1276–82. doi: 10.1038/s41588-021-00921-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Schwartzentruber J, Cooper S, Liu JZ, Barrio-Hernandez I, Bello E, Kumasaka N, et al. Genome-wide meta-analysis, fine-mapping and integrative prioritization implicate new Alzheimer’s disease risk genes. Nat Genet. 2021;53:392–402. doi: 10.1038/s41588-020-00776-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Corder EH, Saunders AM, Strittmatter WJ, Schmechel DE, Gaskell PC, Small GW, et al. Gene dose of apolipoprotein E type 4 allele and the risk of Alzheimer’s disease in late onset families. Science. 1993;261:921–3. doi: 10.1126/science.8346443. [DOI] [PubMed] [Google Scholar]
- 22.Altmann A, Scelsi MA, Shoai M, de Silva E, Aksman LM, Cash DM, et al. A comprehensive analysis of methods for assessing polygenic burden on Alzheimer’s disease pathology and risk beyond APOE. Brain Commun. 2020;2:fcz047. doi: 10.1093/braincomms/fcz047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Escott-Price V, Sims R, Bannister C, Harold D, Vronskaya M, Majounie E, et al. Common polygenic variation enhances risk prediction for Alzheimer’s disease. Brain. 2015;138:3673–84. doi: 10.1093/brain/awv268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Escott-Price V, Shoai M, Pither R, Williams J, Hardy J. Polygenic score prediction captures nearly all common genetic risk for Alzheimer’s disease. Neurobiol Aging. 2017;49:e217–214 e211. doi: 10.1016/j.neurobiolaging.2016.07.018. [DOI] [PubMed] [Google Scholar]
- 25.Escott-Price V, Myers A, Huentelman M, Shoai M, Hardy J. Polygenic risk score analysis of Alzheimer’s disease in cases without APOE4 or APOE2 alleles. J Prev Alzheimers Dis. 2019;6:16–19. doi: 10.14283/jpad.2018.46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Schizophrenia Working Group of the Psychiatric Genomics C. Biological insights from 108 schizophrenia-associated genetic loci. Nature. 2014;511:421–7. doi: 10.1038/nature13595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Karlsson IK, Escott-Price V, Gatz M, Hardy J, Pedersen NL, Shoai M, et al. Measuring heritable contributions to Alzheimer’s disease: polygenic risk score analysis with twins. Brain Commun. 2022;4:fcab308. doi: 10.1093/braincomms/fcab308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ge T, Sabuncu MR, Smoller JW, Sperling RA, Mormino EC, Alzheimer’s Disease Neuroimaging I. Dissociable influences of APOE epsilon4 and polygenic risk of AD dementia on amyloid and cognition. Neurology. 2018;90:e1605–e1612. doi: 10.1212/WNL.0000000000005415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Najar J, van der Lee SJ, Joas E, Wetterberg H, Hardy J, Guerreiro R, et al. Polygenic risk scores for Alzheimer’s disease are related to dementia risk in APOE varepsilon4 negatives. Alzheimers Dement. 2021;13:e12142. doi: 10.1002/dad2.12142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Khachaturian ZS, Petersen RC, Snyder PJ, Khachaturian AS, Aisen P, de Leon M, et al. Developing a global strategy to prevent Alzheimer’s disease: Leon Thal Symposium 2010. Alzheimers Dement. 2011;7:127–32. doi: 10.1016/j.jalz.2011.02.005. [DOI] [PubMed] [Google Scholar]
- 31.Stern Y, Arenaza-Urquijo EM, Bartres-Faz D, Belleville S, Cantilon M, Chetelat G, et al. Whitepaper: defining and investigating cognitive reserve, brain reserve, and brain maintenance. Alzheimers Dement. 2020;16:1305–11. doi: 10.1016/j.jalz.2018.07.219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Perneczky R, Kempermann G, Korczyn AD, Matthews FE, Ikram MA, Scarmeas N, et al. Translational research on reserve against neurodegenerative disease: consensus report of the International Conference on Cognitive Reserve in the Dementias and the Alzheimer’s Association Reserve, Resilience and Protective Factors Professional Interest Area working groups. BMC Med. 2019;17:47. doi: 10.1186/s12916-019-1283-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Latimer CS, Burke BT, Liachko NF, Currey HN, Kilgore MD, Gibbons LE, et al. Resistance and resilience to Alzheimer’s disease pathology are associated with reduced cortical pTau and absence of limbic-predominant age-related TDP-43 encephalopathy in a community-based cohort. Acta Neuropathol Commun. 2019;7:91. doi: 10.1186/s40478-019-0743-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Arenaza-Urquijo EM, Vemuri P. Improving the resistance and resilience framework for aging and dementia studies. Alzheimers Res Ther. 2020;12:41. doi: 10.1186/s13195-020-00609-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Choi KW, Stein MB, Dunn EC, Koenen KC, Smoller JW. Genomics and psychological resilience: a research agenda. Mol Psychiatry. 2019;24:1770–8. doi: 10.1038/s41380-019-0457-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Livingston G, Huntley J, Sommerlad A, Ames D, Ballard C, Banerjee S, et al. Dementia prevention, intervention, and care: 2020 report of the Lancet Commission. Lancet. 2020;396:413–46. doi: 10.1016/S0140-6736(20)30367-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Livingston G, Sommerlad A, Orgeta V, Costafreda SG, Huntley J, Ames D, et al. Dementia prevention, intervention, and care. Lancet. 2017;390:2673–734. doi: 10.1016/S0140-6736(17)31363-6. [DOI] [PubMed] [Google Scholar]
- 38.Kremen WS, Beck A, Elman JA, Gustavson DE, Reynolds CA, Tu XM, et al. Influence of young adult cognitive ability and additional education on later-life cognition. Proc Natl Acad Sci USA. 2019;116:2021–6. doi: 10.1073/pnas.1811537116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lee JJ, Wedow R, Okbay A, Kong E, Maghzian O, Zacher M, et al. Gene discovery and polygenic prediction from a genome-wide association study of educational attainment in 1.1 million individuals. Nat Genet. 2018;50:1112–21. doi: 10.1038/s41588-018-0147-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Polderman TJ, Benyamin B, de Leeuw CA, Sullivan PF, van Bochoven A, Visscher PM, et al. Meta-analysis of the heritability of human traits based on fifty years of twin studies. Nat Genet. 2015;47:702–9. doi: 10.1038/ng.3285. [DOI] [PubMed] [Google Scholar]
- 41.Davies G, Lam M, Harris SE, Trampush JW, Luciano M, Hill WD, et al. Study of 300,486 individuals identifies 148 independent genetic loci influencing general cognitive function. Nat Commun. 2018;9:2098. doi: 10.1038/s41467-018-04362-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Farrer LA, Cupples LA, Haines JL, Hyman B, Kukull WA, Mayeux R, et al. Effects of age, sex, and ethnicity on the association between apolipoprotein E genotype and Alzheimer disease. A meta-analysis. APOE and Alzheimer Disease Meta Analysis Consortium. J Am Med Assoc. 1997;278:1349–56. doi: 10.1001/jama.1997.03550160069041. [DOI] [PubMed] [Google Scholar]
- 43.Jonsson T, Atwal JK, Steinberg S, Snaedal J, Jonsson PV, Bjornsson S, et al. A mutation in APP protects against Alzheimer’s disease and age-related cognitive decline. Nature. 2012;488:96–99. doi: 10.1038/nature11283. [DOI] [PubMed] [Google Scholar]
- 44.DeTure MA, Dickson DW. The neuropathological diagnosis of Alzheimer’s disease. Mol Neurodegener. 2019;14:32. doi: 10.1186/s13024-019-0333-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Reed BR, Mungas D, Farias ST, Harvey D, Beckett L, Widaman K, et al. Measuring cognitive reserve based on the decomposition of episodic memory variance. Brain. 2010;133:2196–209. doi: 10.1093/brain/awq154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Negash S, Bennett DA, Wilson RS, Schneider JA, Arnold SE. Cognition and neuropathology in aging: multidimensional perspectives from the Rush Religious Orders Study and Rush Memory And Aging Project. Curr Alzheimer Res. 2011;8:336–40. doi: 10.2174/156720511795745302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Hohman TJ, McLaren DG, Mormino EC, Gifford KA, Libon DJ, Jefferson AL, et al. Asymptomatic Alzheimer disease: defining resilience. Neurology. 2016;87:2443–50. doi: 10.1212/WNL.0000000000003397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ayers KL, Mirshahi UL, Wardeh AH, Murray MF, Hao K, Glicksberg BS, et al. A loss of function variant in CASP7 protects against Alzheimer’s disease in homozygous APOE epsilon4 allele carriers. BMC Genomics. 2016;17:445. doi: 10.1186/s12864-016-2725-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.DeKosky ST, Aston CE, Kamboh MI. Polygenic determinants of Alzheimer’s disease: modulation of the risk by alpha-1-antichymotrypsin. Ann N. Y Acad Sci. 1996;802:27–34. doi: 10.1111/j.1749-6632.1996.tb32595.x. [DOI] [PubMed] [Google Scholar]
- 50.Kamboh MI, Sanghera DK, Ferrell RE, DeKosky ST. APOE*4-associated Alzheimer’s disease risk is modified by alpha 1-antichymotrypsin polymorphism. Nat Genet. 1995;10:486–8. doi: 10.1038/ng0895-486. [DOI] [PubMed] [Google Scholar]
- 51.Proitsi P, Lupton MK, Velayudhan L, Newhouse S, Fogh I, Tsolaki M, et al. Genetic predisposition to increased blood cholesterol and triglyceride lipid levels and risk of Alzheimer disease: a Mendelian randomization analysis. PLoS Med. 2014;11:e1001713. doi: 10.1371/journal.pmed.1001713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Lovestone S, Francis P, Kloszewska I, Mecocci P, Simmons A, Soininen H, et al. AddNeuroMed–the European collaboration for the discovery of novel biomarkers for Alzheimer’s disease. Ann N. Y Acad Sci. 2009;1180:36–46. doi: 10.1111/j.1749-6632.2009.05064.x. [DOI] [PubMed] [Google Scholar]
- 53.McCarthy S, Das S, Kretzschmar W, Delaneau O, Wood AR, Teumer A, et al. A reference panel of 64,976 haplotypes for genotype imputation. Nat Genet. 2016;48:1279–83. doi: 10.1038/ng.3643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Sleegers K, Bettens K, De Roeck A, Van Cauwenberghe C, Cuyvers E, Verheijen J, et al. A 22-single nucleotide polymorphism Alzheimer’s disease risk score correlates with family history, onset age, and cerebrospinal fluid Abeta42. Alzheimers Dement. 2015;11:1452–60. doi: 10.1016/j.jalz.2015.02.013. [DOI] [PubMed] [Google Scholar]
- 55.Leonenko G, Baker E, Stevenson-Hoare J, Sierksma A, Fiers M, Williams J, et al. Identifying individuals with high risk of Alzheimer’s disease using polygenic risk scores. Nat Commun. 2021;12:4506. doi: 10.1038/s41467-021-24082-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Chang CC, Chow CC, Tellier LC, Vattikuti S, Purcell SM, Lee JJ. Second-generation PLINK: rising to the challenge of larger and richer datasets. Gigascience. 2015;4:7. doi: 10.1186/s13742-015-0047-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Willer CJ, Li Y, Abecasis GR. METAL: fast and efficient meta-analysis of genomewide association scans. Bioinformatics. 2010;26:2190–1. doi: 10.1093/bioinformatics/btq340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Wan YW, Al-Ouran R, Mangleburg CG, Perumal TM, Lee TV, Allison K, et al. Meta-analysis of the Alzheimer’s disease human brain transcriptome and functional dissection in mouse models. Cell Rep. 2020;32:107908. doi: 10.1016/j.celrep.2020.107908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.van der Lee SJ, Wolters FJ, Ikram MK, Hofman A, Ikram MA, Amin N, et al. The effect of APOE and other common genetic variants on the onset of Alzheimer’s disease and dementia: a community-based cohort study. Lancet Neurol. 2018;17:434–44. doi: 10.1016/S1474-4422(18)30053-X. [DOI] [PubMed] [Google Scholar]
- 60.Longford NT. Classification in two-stage screening. Stat Med. 2015;34:3281–97. doi: 10.1002/sim.6554. [DOI] [PubMed] [Google Scholar]
- 61.Marioni RE, Campbell A, Hagenaars SP, Nagy R, Amador C, Hayward C, et al. Genetic stratification to identify risk groups for Alzheimer’s disease. J Alzheimers Dis. 2017;57:275–83. doi: 10.3233/JAD-161070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Bellou E, Baker E, Leonenko G, Bracher-Smith M, Daunt P, Menzies G, et al. Age-dependent effect of APOE and polygenic component on Alzheimer’s disease. Neurobiol Aging. 2020;93:69–77. doi: 10.1016/j.neurobiolaging.2020.04.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Stocker H, Perna L, Weigl K, Mollers T, Schottker B, Thomsen H, et al. Prediction of clinical diagnosis of Alzheimer’s disease, vascular, mixed, and all-cause dementia by a polygenic risk score and APOE status in a community-based cohort prospectively followed over 17 years. Mol Psychiatry. 2020;26:5812–22. doi: 10.1038/s41380-020-0764-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Fulton-Howard B, Goate AM, Adelson RP, Koppel J, Gordon ML, Alzheimer’s Disease Genetics C, et al. Greater effect of polygenic risk score for Alzheimer’s disease among younger cases who are apolipoprotein E-epsilon4 carriers. Neurobiol Aging. 2021;99:101 e101–101 e109. doi: 10.1016/j.neurobiolaging.2020.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Arboleda-Velasquez JF, Lopera F, O’Hare M, Delgado-Tirado S, Marino C, Chmielewska N, et al. Resistance to autosomal dominant Alzheimer’s disease in an APOE3 Christchurch homozygote: a case report. Nat Med. 2019;25:1680–3. doi: 10.1038/s41591-019-0611-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Ware EB, Faul JD, Mitchell CM, Bakulski KM. Considering the APOE locus in Alzheimer’s disease polygenic scores in the Health and Retirement Study: a longitudinal panel study. BMC Med Genomics. 2020;13:164. doi: 10.1186/s12920-020-00815-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Neu SC, Pa J, Kukull W, Beekly D, Kuzma A, Gangadharan P, et al. Apolipoprotein E genotype and sex risk factors for Alzheimer disease: a meta-analysis. JAMA Neurol. 2017;74:1178–89. doi: 10.1001/jamaneurol.2017.2188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Mukherjee S, Kim S, Gibbons LE, Nho K, Risacher SL, Glymour MM, et al. Genetic architecture of resilience of executive functioning. Brain Imaging Behav. 2012;6:621–33. doi: 10.1007/s11682-012-9184-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Mukherjee S, Kim S, Ramanan VK, Gibbons LE, Nho K, Glymour MM, et al. Gene-based GWAS and biological pathway analysis of the resilience of executive functioning. Brain Imaging Behav. 2014;8:110–8. doi: 10.1007/s11682-013-9259-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Hohman TJ, Dumitrescu L, Cox NJ, Jefferson AL, Alzheimer’s Neuroimaging I. Genetic resilience to amyloid related cognitive decline. Brain Imaging Behav. 2017;11:401–9. doi: 10.1007/s11682-016-9615-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Felsky D, Xu J, Chibnik LB, Schneider JA, Knight J, Kennedy JL, et al. Genetic epistasis regulates amyloid deposition in resilient aging. Alzheimers Dement. 2017;13:1107–16. doi: 10.1016/j.jalz.2017.01.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Ridge PG, Karch CM, Hsu S, Arano I, Teerlink CC, Ebbert MTW, et al. Linkage, whole genome sequence, and biological data implicate variants in RAB10 in Alzheimer’s disease resilience. Genome Med. 2017;9:100. doi: 10.1186/s13073-017-0486-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The Alzheimer’s Disease Genetics Consortium, the Alzheimer’s Disease Neuroimaging Initiative, and the AddNeuroMed data used in this study were provided under restricted access by NIAGADS (https://www.niagads.org), ADNI (http://adni.loni.usc.edu), and Synapse platform (https://www.synapse.org/#!Synapse:syn4907804), respectively. Only summary statistics were made available to us from the European Alzheimer’s Disease Initiative, the Genetic and Environmental Risk in Alzheimer’s Disease, the Cohorts for Heart and Aging Research in Genomic Epidemiology, the Psychiatric Genomic Consortium, the Australian Imaging, Biomarker & Lifestyle Study, and the Sydney Memory and Ageing Study. The resilience-scoring weights of design 1 are available from the corresponding author upon request.
The code for data cleaning and analysis, and the resilience-scoring weights of design 1 are available from the corresponding author upon request.