

We hypothesized that female mice had differences (vs males) in their microbiota after sepsis. Females demonstrated different microbiota 14 days after abdominal sepsis. This difference in recovery may play a role in outcome differences between sexes after sepsis.
KEY WORDS: Fecal bacteria, resilience, 16S rRNA, persistent inflammation immunosuppression and catabolism syndrome, mice
BACKGROUND
Sepsis-induced gut microbiome alterations contribute to sepsis-related morbidity and mortality. Given evidence for improved postsepsis outcomes in females compared with males, we hypothesized that female mice maintain microbiota resilience versus males.
METHODS
Mixed-sex C57BL/6 mice underwent cecal ligation and puncture (CLP) with antibiotics, saline resuscitation, and daily chronic stress and were compared with naive (nonsepsis/no antibiotics) controls. For this work, the results of young (3–5 months) and old (18–22 months) adult mice were analyzed by sex, independent and dependent of age. Mice were sacrificed at days 7 and 14, and 16S rRNA gene sequencing was performed on fecal bacterial DNA. α and β diversity were determined by Shannon index and Bray-Curtis with principal coordinate analysis, respectively. False discovery rate (FDR) correction was implemented to account for potential housing effect.
RESULTS
In control mice, there was no difference in α or β diversity between male and female mice (FDR, 0.76 and 0.99, respectively). However, male mice that underwent CLP with daily chronic stress had a decrease in microbiota α diversity at 7 days post-CLP (Shannon FDR, 0.005), which was sustained at 14 days post-CLP (Shannon FDR, 0.001), compared with baseline. In addition, male mice maintained differences in β diversity even at day 14 compared with controls (FDR, <0.0001). In contrast, female mice had a decreased microbiota α diversity (Shannon FDR, 0.03) and β diversity (FDR, 0.02) 7 days post-CLP but recovered their α and β diversity by post-CLP day 14 (Shannon FDR, 0.5, and FDR, 0.02, respectively). Further analysis of females revealed that only young female mice were not different (β diversity) post-CLP day 14 to controls.
CONCLUSION
Although sepsis-induced perturbations of the intestinal microbiota occur initially in both male and female C57BL/6 mice, females demonstrate different microbiota by day 14. This may be seen primarily in younger females. This difference in recovery may play a role in outcome differences between sexes after sepsis.
Globally, sepsis is a leading cause of death,1 and the World Health Organization has made sepsis a health priority.2 In addition, sepsis accounts for >$20 billion of total hospital costs in the United States.3 Although early sepsis survival has improved over the past decade, this decrease in mortality has yielded a rapidly expanding population of sepsis survivors who develop chronic critical illness (CCI).4 Chronic critical illness is characterized by persistent organ dysfunction requiring prolonged intensive care unit (ICU) treatment. Subsequently, CCI frequently manifests thereafter in the host as low-grade systemic inflammation, global immunosuppression, and cachexia/muscle wasting,5 known as the persistent inflammation, immunosuppression, and catabolism syndrome (PICS) endotype.4,5 Sepsis survivors with CCI/PICS are often discharged to long-term acute care facilities, where they experience repeat infections and ~40% 1-year postdischarge mortality.6,7 Specific treatments for these sepsis survivors are lacking, because, in part, of (1) inadequate knowledge of its pathobiology and (2) a paucity of studies that are able to address sepsis in a manner that befits precision medicine, including but not limited to sex.8,9 In fact, it is now recognized that “[a] better understanding of sex-and gender-dependent differences may serve to increase translational research success.”8
Although there is still significant debate as to whether females have improved outcomes after sepsis,8,9 studies indicate that females are less likely to have postsepsis morbidity and mortality, even after menopause.10,11 In fact, one study demonstrated that, in patients younger than 90 years, women had significantly decreased 1-year mortality compared with males.10 Regardless, it is clear that females and males respond differently to infection.8–10 Future successful interventions for sepsis will need to use personalized/precision medical therapies, which will require a better understanding of the differences in host response due to severe infection.
One key aspect to septic outcomes is the host microbiome.12,13 The microbiota is the collection of trillions of microorganisms that form a symbiont and pathobiont relationship with its host. Its role in various human diseases, including posttrauma recovery, has gained much traction recently given its interaction with the immune system and host metabolic processes.14–16 The ability to study this ecosystem is predicated on taking advantage of the unique 16S ribosomal subunit gene in prokaryotic organisms. Because of the slow evolutionary change imparted on this gene, sequencing allows reconstruction of bacterial phylogeny present in a host microbiota. Subsequently, the abundance (total number), richness (the assortment of bacterial species in an ecosystem, e.g., your gut microbiome), and diversity (the amount of individual bacteria from each of the bacterial species present) can be determined. Microbial community diversity can be measured within a cohort (α diversity) or between cohorts from different environments (β diversity).17 Through bioinformatic analysis, any noted changes based on host, treatment, and so on, can be determined to give greater insight into how the microbiome may play a role in human disease.
Critical illness has been shown to disturb the host microbiome,12 and evidence exists demonstrating that a host dysbiome worsens outcomes after sepsis.13 Importantly, it is not simply the changes in host microbiome that contributes to its pathology but the ability (or inability) of the microbiome to recover from perturbations, known as microbiota resilience, that can determine host morbidity and mortality.18 Furthermore, sex is considered one the most important factors affecting the gut microbiota.11 For example, investigators have conducted sex-specific fecal microbiota transplantation animal studies from C57BL/6 J conventional mice in germ-free mice of the opposite sex. Their results illustrated that female recipients lost significantly more weight after receiving male microbiota compared with receiving female microbiota, suggesting a sexually dimorphic impact of the microbiome on metabolic regulation.19
Using a murine model of surgical/abdominal sepsis that combines cecal ligation and puncture with daily chronic stress (CLP-DCS) and is associated with PICS and CCI, our goal was to determine differences in the diversity of the murine microbiome at post–cecal ligation and puncture (CLP) days 7 and 14 between female and male mice. Furthermore, we specifically sought to determine if differences existed in the gut microbiota in female versus male adult mice in postsepsis recovery, as differences in microbiota resilience are potentially targets of therapy,16 and may help to explain sex-based differences in postsepsis recovery.
MATERIALS AND METHODS
Animals
All animal experiments were approved by the University of Florida Institutional Animal Care and Use Committee and followed Animal Research Reporting of In Vivo Experiments guidelines (https://www.nc3rs.org.uk/arrive-guidelines) (Supplemental Digital Content, Supplementary Data 1, http://links.lww.com/TA/C417). The animals were cared for and used according to the Guide for the Care and Use of Laboratory Animals.20 C57BL/6 J (B6) mice of both sexes were purchased from Jackson Laboratory (Bar Harbor, ME) and the National Institute on Aging (Baltimore, MD). Mice were cared for by the University of Florida Animal Care Services and housed in transparent cages (three to four animals of same sex/age per cage) under specific pathogen-free conditions in a single room. Animals were provided standard irradiated pelleted diet and water ad libitum for the duration of the study. Before initiation of the experiment, mice were acclimated to a 12-hour light-dark cycle for a minimum of 14 days, and National Institute on Aging and Jackson Laboratory old adult mice were housed together. Because of the coprophagic nature of mice, this assured that mice in the same cage would have similar microbiota composition and structure.21 Only animals of the same sex, age, and treatment group were housed together. For this work, young (3–5 months) and old (18–22 months) adult murine microbiome results were combined and analyzed by the variable of sex.
Intra-abdominal Sepsis and Daily Chronic Stress Model
To recapitulate the human condition of CCI and PICS, a murine model of sepsis and persistent inflammation, previously described by our laboratory, was used.22 General anesthesia was induced with inhaled isoflurane, and CLP was performed via a midline laparotomy with exteriorization of the cecum to induce a model of sepsis. The cecum was ligated with 2-0 silk suture 1 cm from its tip, a 25-gauge needle was used to puncture the cecum, and the laparotomy was closed in one layer with surgical clips. Buprenorphine analgesia was provided for 48 hours postsurgery. Imipenem monohydrate (25 mg/kg in 1 mL 0.9% normal saline) was administered subcutaneously 2 hours post-CLP and then continued twice daily for 72 hours. Subsequently, we added a component of daily chronic stress (DCS) to mimic the ICU stay of human patients who develop CCI.19 Daily chronic stress was conducted by placing mice in weighted plexiglass animal restraint holders (Kent Scientific, Torrington, CT) for 2 hours daily commencing the day after CLP. The CLP-DCS mice, along with mixed-sex naive mice (no CLP, no DCS, no antibiotics, and no fluid resuscitation) (old, n = 13; young, n = 13), were euthanized on day 7 or 14 post–CLP-DCS. Mice were euthanized via cervical dislocation following isoflurane inhalation, after which stool from the descending colon (during exploratory laparotomy, after euthanasia, the distal portion of the colon was incised and feces were pushed directly into a cryogenic vial) was collected under sterile conditions in a laminar flow hood into sterile Eppendorf tubes (Fisher Scientific, Waltham, WA) and snap frozen in liquid nitrogen immediately. Subsequently, these samples were stored in a −80°C freezer.
Bacterial DNA Isolation and 16S rRNA Gene Sequencing
Whole genome bacterial DNA was isolated from individual animal stool using the QIAamp Fast DNA Stool Mini Kit (Qiagen, Germantown, MD) per manufacturer guidelines. Library preparation was performed using the Quick-16S NGS (Next Generation Sequencing) Library Prep Kit (Zymo Research, Irvine, CA) according to manufacturer protocol. The V3-V4 hypervariable region of the 16S rRNA gene was amplified, and paired-end adapter sequences with unique eight nucleotide barcodes were used during library preparation to allow multiplexing. The final polymerase chain reaction products were quantified with quantitative polymerase chain reaction and pooled based on equal molarity. The final pooled library was purified using the Mag-Bind TotalPure NGS kit (Omega Bio-tek, Inc., Norcross, GA); quality control and quantification were performed with TapeStation (Agilent Technologies, Santa Clara, CA) and Qubit (Thermo Fisher Scientific, Waltham, WA). Sequencing was subsequently completed using the Illumina MiSeq sequencer (Illumina, Inc., San Diego, CA), producing paired end reads, 300 bases long, which produced 11,834,108 total reads (each end) for the 48 samples.
Plasma Testosterone and Estrogen Assay
Blood was collected in dipotassium ethylenediaminetetraacetic acid tubes. Plasma samples were collected after centrifugation of blood samples at 1,800g for 10 minutes and stored at −80°C until used for quantitative determination of testosterone and estradiol using the testosterone and estradiol assay kits (Bio-Techne, Minneapolis, MN). Standard curves were created using a four-parameter logistic curve fit (Softmax Pro 3.1 software; Molecular Devices Corporation, Sunnyvale, CA).
Statistical Analyses
Paired end reads were joined using fastq-join from ea-utils (https://expressionanalysis.github.io/ea-utils/) with a minimum overlap of 6 bp and up to 8% maximum difference. The joined reads were quality filtered using Quantitative Insights into Microbial Ecology (QIIME) version 1.9.123 with a score of Q20 or higher in at least 75% of the input read length. The first and last 10 bases of each read were then trimmed, and any read <100 bases long were removed via Trimmomatic (http://www.usadellab.org/cms/?page=trimmomatic). An additional filtering step was applied to remove PhiX and mouse/host sequences using KneadData (http://huttenhower.sph.harvard.edu/kneaddata). Preindexed bowtie2 files for PhiX and Mus musculus Ensembl GRCm38 from Illumina's iGenomes collection were used as references. The surviving reads were fed to QIIME to pick operational taxonomic units at 97% similarity level using an open-reference picking strategy with the Greengenes reference data set (release 13.8). Chimeric sequences were detected and removed using ChimeraSlayer (http://microbiomeutil.sourceforge.net/#A_CS). Operational taxonomic units that had ≤0.005% of the total number of sequences were excluded according to Bokulich and colleagues,23 and the final set contained 13,455,787 reads. Taxonomic assignment was carried out using the Ribosomal Database Project classifier version 2.224 through QIIME with confidence set to 50%. Counts were then normalized and log10 transformed according to the following formula25:
where RC is the read count for a particular operational taxonomic unit in a particular sample, n is the total number of reads in that sample, the sum of x is the total number of reads in all samples, and N is the total number of samples.
Principal coordinate analysis (PCoA) was performed on Bray-Curtis distance of the normalized and log10-transformed counts using the phyloseq R package.26α Diversity was assessed using Chao1, and Shannon indexes were assessed using rarefied counts (set to 89,087 reads representing the minimum number of reads among all samples). All statistical analyses were performed in R version 4.0.3.27 A linear mixed-effects model (lme function) in the R lme4 package (version 1.1–25), with the residual maximum likelihood method, was used to fit a generalized mixed linear model of the following form: var ~ state +1|cage, where var is PCoA axis, Chao1 index, Shannon index, taxa normalized, and log10-transformed count (considering only taxa present in at least 25% of the samples) or log10 read count in each sample. The latter was performed to ensure that the sparsity of 16S rRNA count data did not contribute to sample clustering. State is defined as male/female and post–CLP-DCS (7 days or 14 days)/naive and one |cage indicates that we used the cage as a random effect to account for cohousing effects.25p Values were obtained from the analysis of variance on the previous model and were false discovery rate (FDR) corrected to control for potential cage effect using the Benjamini and Hochberg approach.28 A one-way analysis of variance with Tukey's multiple comparisons test was performed to compare plasma testosterone and estradiol levels between indicated cohorts using Prism 8.0 (GraphPad Software, San Diego, CA.) A p value of ≤0.05 was considered statistically significant.
RESULTS
Female B6 Mice Recover From Sepsis and Stress-Induced Microbiota Diversity Changes Over Time Compared With Nonrecovering Male Mice
We have previously demonstrated the impact of age on the stability of the murine microbiota after CLP-DCS.29 However, it is unknown what impact sex has in relation to microbiome changes after sepsis over the adult age range. Given the clinical data demonstrating survival differences after sepsis between male and female patients, we sought to determine whether the changes in the host microbiota in a murine model of sepsis/PICS are based on the sex of the host. For this work, we analyzed the results of our young and old adult mice, combined looking at the results by sex only and, subsequently, looking at the differences in sex in old and young mice separately. Because of the small sample sizes, this latter analysis must be considered preliminary.
Mice of both sexes underwent the CLP-DCS model as indicated. To determine similarities of baseline murine microbiota based on sex, 16S rRNA gene sequencing was performed followed by a measure of α and β diversity between the cohorts. This demonstrated no difference in α diversity in naive (healthy, control) male versus female mice by the Chao1 index (FDR, 0.76; Fig. 1A), nor β diversity as measured by the Bray-Curtis dissimilarity index (FDR, 0.99; Fig. 1B). Given this, we next sought to determine temporal changes within each sex over time (0, 7, and 14 days post–CLP-DCS). This demonstrated that, compared with their naive (time 0) counterparts, male B6 mice had decreased α diversity at 7 days (Chao1 FDR, 0.003; Shannon FDR, 0.005; Fig. 1C) that persisted at 14 days post–CLP-DCS (Chao1 FDR, <0.001; Shannon FDR, 0.001; Fig. 1C). Interestingly, although the α diversity in female mice 7 days post–CLP-DCS was decreased (Chao1 FDR, 0.01; Shannon FDR, 0.03), this difference disappeared by day 14 (Chao1 FDR, 0.5; Shannon FDR, 0.5; Fig. 1D). Finally, PCoA on Bray-Curtis dissimilarity did not identify a significant difference in β diversity in male B6 mice at 7 days (FDR, 0.08; Fig. 2A) but was present at 14 days post–CLP-DCS (FDR, <0.001; Fig. 2B), demonstrating a delayed shift in the microbiota. However, as was the case with α diversity in the female cohort, B6 female mice demonstrated a significant difference in β diversity by PCoA at 7 days (FDR, 0.02; Fig. 2C), which generally recovered by 14 days (FDR, 0.07; Fig. 2D). These data demonstrate the association of sex with changes in the murine microbiota after sepsis and daily chronic stress.
Figure 1.
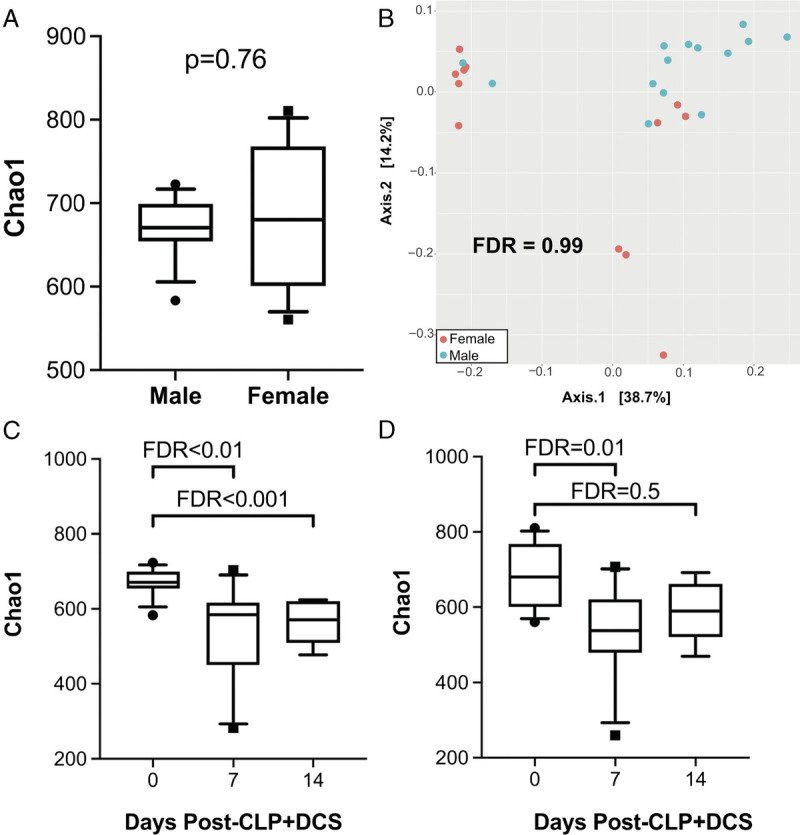
16S rRNA gene sequencing of the intestinal microbiota at baseline (“naive”) before CLP-DCS demonstrates no significant difference in α diversity by Chao1 (A) or β diversity by Bray-Curtis dissimilarity index (B). However, 7 and 14 days post–CLP-DCS demonstrated a significant difference in intestinal microbiota α diversity in male mice compared with baseline (C). While female mice also demonstrated a CLP-DCS–induced microbiota shift in diversity at 7 days, this change in diversity appears restored after 14 days (D).
Figure 2.
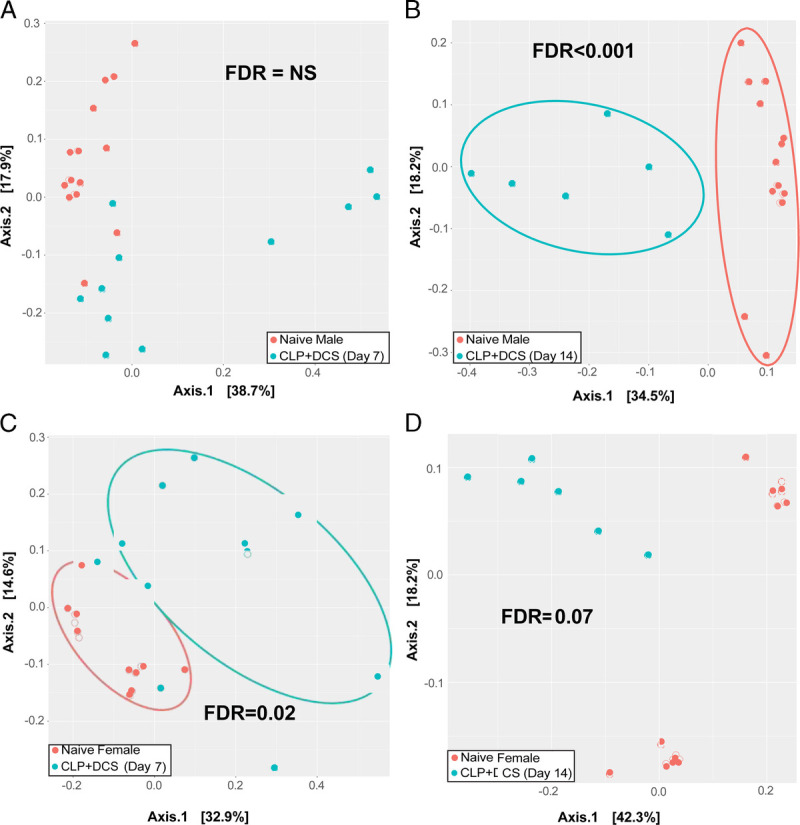
Principal coordinate analysis of Bray-Curtis dissimilarity index is presented, which demonstrates no difference in β diversity in male B6 mice post–CLP-DCS at 7 days (A) but at 14 days is significantly different (B). In contrast, female B6 mice had a shift in the β diversity at 7 days (C), which appeared restored by 14 days (D). NS, not significant.
Alterations in α Diversity Associate With Age- and Sex-Based Differences in Mice Recovering From sepsis
To extend observations from our prior work that demonstrated stability of the murine microbiota in young mice compared with old after sepsis,29 but in the context of sex and temporal differences, changes in α diversity of the cohort were determined in subsets of the mice based on both age and sex (young male, old male, young female, and old female) pre–CLP-DCS as well as 7 and 14 days post–CLP-DCS. This demonstrated that, after sepsis, the α diversity of both young male (Fig. 3A) and young female mice remains stable compared with old, which maintains a persistent level of α diversity even at 14 days after CLP-DCS (Fig. 3). Specifically, old male mice had a significant decrease in Chao1 α diversity 14 days after CLP-DCS (FDR, <0.01; Fig. 3B), and old female mice had a significant decrease in α diversity 7 and 14 days post–CLP-DCS (FDR, <0.001; Fig. 3D).
Figure 3.
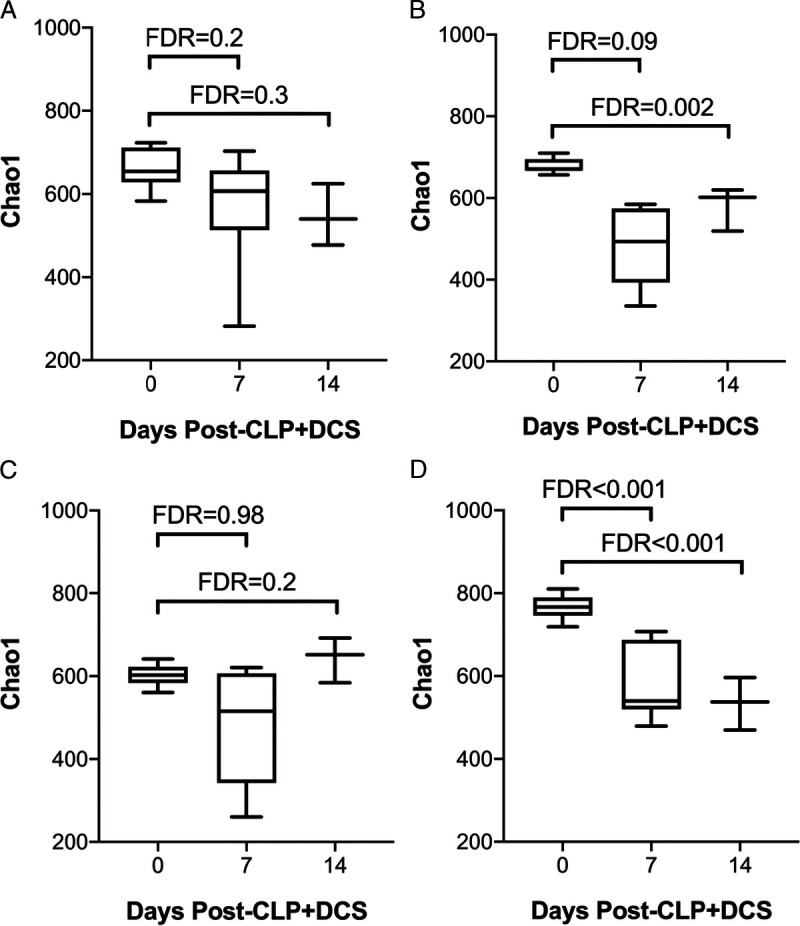
α Diversity by Chao1 of young and old mice that underwent CLP-DCS was analyzed for each sex and demonstrates that (A) young male mice do not have alterations in the α diversity of their intestinal microbiota after CLP-DCS but their old male counterparts do (B; FDR, 0.002). Similarly, young female mice that undergo CLP-DCS have stable α diversity after CLP-DCS (C), but old female mice were noted to have decreased α diversity over time after CLP-DCS (D; FDR, <0.001).
To determine if age drives the difference in β diversity seen in male and female mice (Fig. 2), Bray-Curtis dissimilarity index was next performed as previously described for male and female mice based on age with the PCoA plots illustrated in Figure 4. Given the difference in β diversity in male mice at 14 days post–CLP-DCS, subset analysis of this group based on age was performed to determine if this was a driver of the observation. Indeed, young male mice at 14 days post–CLP-DCS did not have a difference in their intestinal microbiota β diversity (FDR, 0.9; Fig. 4A); however, only old male mice had a significant difference in β diversity (FDR, <0.0001; Fig. 4B). Finally, given the differences in β diversity in female mice 7 days post–CLP-DCS, PCoA analysis was likewise performed on the cohort comparing young to old mice. This demonstrated that there was a difference in β diversity in young female mice at 7 days post–CLP-DCS (FDR, 0.04; Fig. 4C), but not old female mice (FDR, 0.99; Fig. 4D). However, at 14 days post–CLP-DCS, the β diversity of young female mice remained different compared with their baseline (FDR, 0.02; data not shown), while old female mice now had a significant difference in β diversity at 14 days post–CLP-DCS compared with pre–CLP-DCS (FDR, <0.0001; data not shown).
Figure 4.
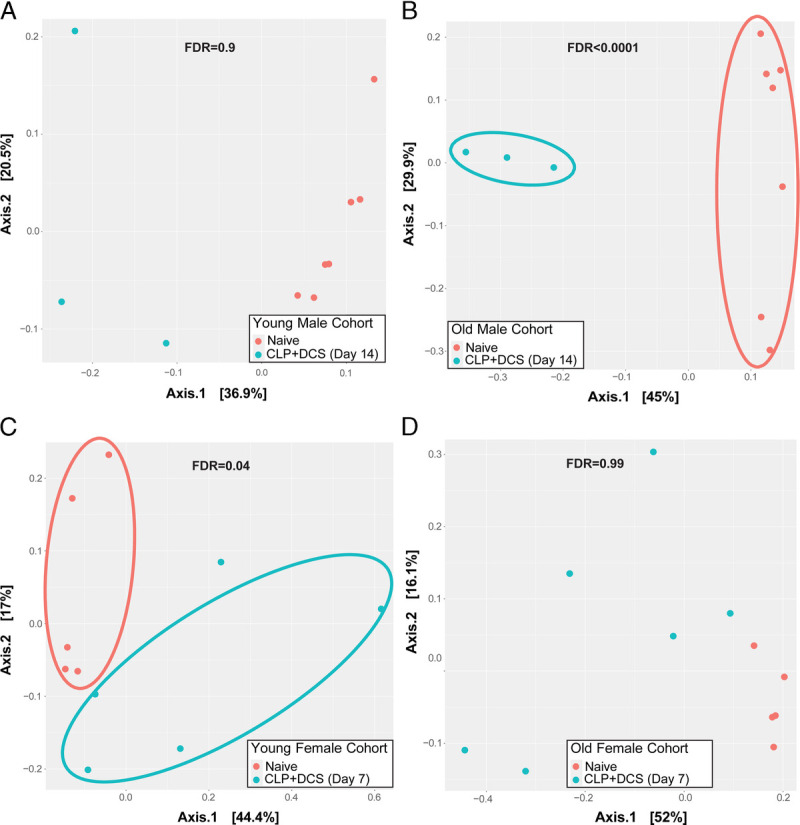
Principal coordinate analysis of Bray-Curtis dissimilarity index is presented for young and old male mice 14 days after CLP-DCS (A and B, respectively) as well as young and old female mice at 7 days post–CLP-DCS (C and D, respectively). This demonstrated a difference in the β diversity of the old male mice at 14 days post–CLP-DCS (FDR, <0.0001) and young female mice at 7 days post–CLP-DCS (FDR, 0.04).
Late Taxonomic Changes in Male Mice After Sepsis, As Well as Stress-Induced Microbiota Changes, Are Dominated by a Decrease in Clostridiales and Increased Lactobacillus
Subsequent analysis of the 16S sequencing from each sex cohort was performed to determine taxa associated with the diversity changes. The 7-day post–CLP-DCS changes in diversity in females compared with naive was dominated by a 65-log fold decrease in the order Clostridiales (FDR, <0.001), which comprised the top 10 altered taxa. Conversely, the 7-day post–CLP-DCS changes in diversity in males were dominated by a 278-log fold decrease in the order Bacteroidales (FDR, <0.001; Supplemental Digital Content, Supplementary Table 1, http://links.lww.com/TA/C418). Given the recovery of the female intestinal microbiota as evidenced by no difference in diversity compared with naive, we sought to determine the taxa responsible for the persistent shift in diversity among male mice at 14 days post–CLP-DCS. The change was notably driven by a 449-log fold decrease in the order Clostridiales (FDR, <0.001) and 474-log fold increase in genus Lactobacillus (Supplemental Digital Content, Supplementary Table 2, http://links.lww.com/TA/C418).
Plasma Estradiol and Testosterone Levels Are Unaffected by Sepsis and Stress-Induced Changes in the Intestinal Microbiota of B6 Mice
Given prior reports of the impact of sepsis on sex hormones and the possibility that the identified intestinal microbiota changes may be secondary to changes in these hormones induced by stress,8–11,30 plasma was isolated from male and female mice after CLP-DCS. This demonstrated no significant difference in estradiol levels between naive versus 7 days post–CLP-DCS and 14 days post–CLP-DCS (p = 0.2 and 0.99, respectively; Fig. 5A). Similarly, estradiol levels were similar in male B6 mice at 7 and 14 days post–CLP-DCS compared with their naive control (p = 0.99 and 0.91, respectively; Fig. 5A). Plasma testosterone levels were measured in female and male B6 mice after CLP-DCS. This demonstrated stable testosterone levels in female mice at 7 and 14 days post–CLP-DCS (p = 0.99 and 0.99, respectively; Fig. 5B) compared with their naive control as well as stability of testosterone levels in male mice 7 and 14 days post–CLP-DCS (p = 0.23 and 0.1, respectively; Fig. 5B). It should be noted that we were not able to determine the stage of the estrous cycle in the naive and CLP-DCS female mice,31 although this may not be necessary for all sex-based murine research.32 In addition, it has been demonstrated that, while there are clear increases in plasma estradiol in female mice during estrous (approximately 4 days), differences in plasma E2 levels between males and females during the nonestrous female days are not as clear cut.31,32 These data indicate that sepsis and daily chronic stress-induced changes in the murine intestinal microbiota may not be directly related to changes plasma sex hormone concentrations, at least in this wide age range of animals.
Figure 5.
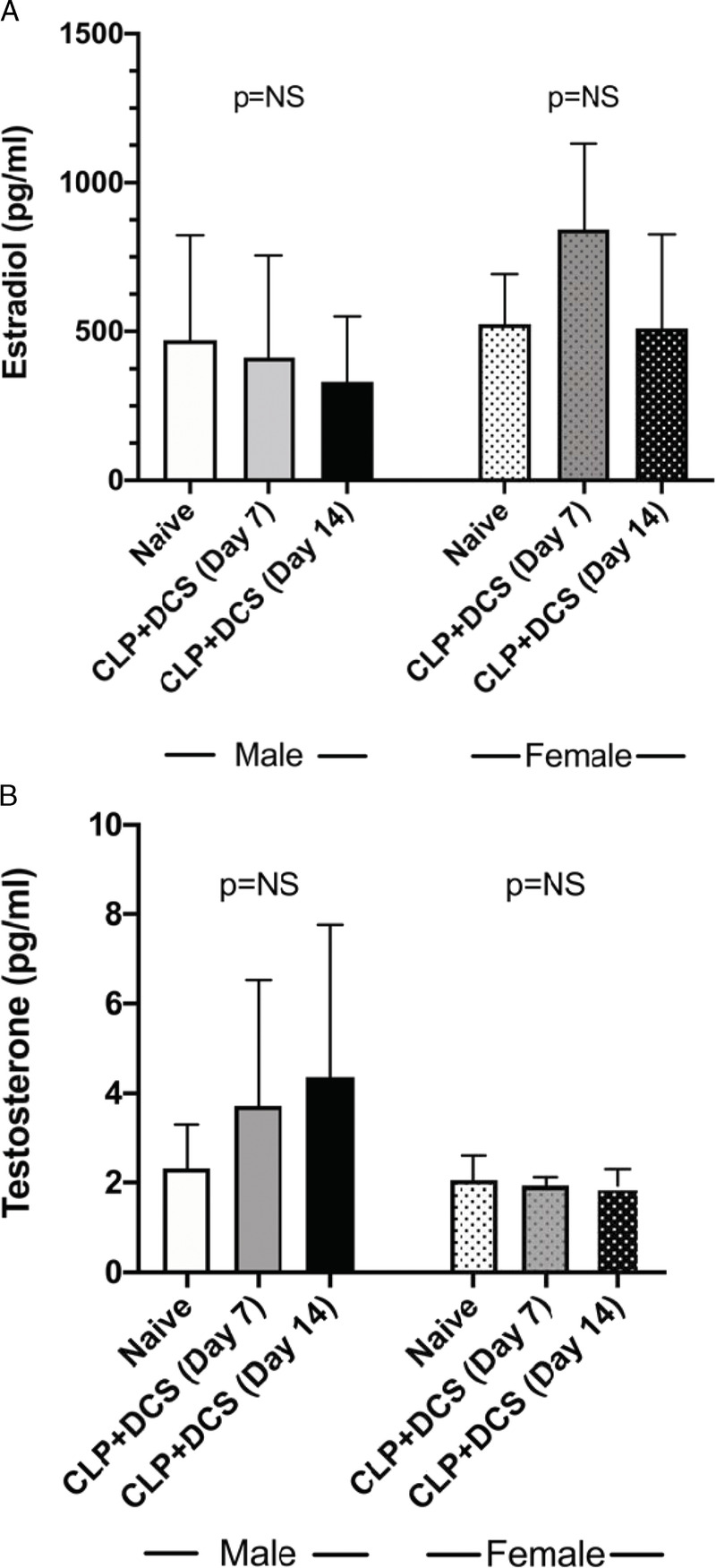
Plasma measurement of estradiol (A) and testosterone (B) in male and female B6 mice at baseline, 7 days, and 14 days post–CLP-DCS demonstrated no significant difference in plasma levels within sexes at the indicated time points. NS, not significant.
DISCUSSION
Using our mouse surgical-sepsis model (CLP-DCS) emulating the human condition of CCI and the PICS endotype,19 we determined that, although perturbations of the intestinal microbiota occur initially in both male and female C57BL/6 mice, the microbiota of males and females by day 14 postsepsis are not equivalent. This could potentially be due to improved microbial resilience by 2 weeks after severe infection in females. This resilience may be also driven by age, given the stability of the young female and male microbiota in these age groups. Regardless, our results may help to better explain sex differences in postsepsis ICU survival given that females may have better survival compared with males.8–11 These unique sex-related changes may be imparted on the host microbiota by the impact of sepsis and chronic stress. Such changes may lead to a chronic inflammatory state seen in PICS.4,5 The impact of CLP-DCS on microbiota diversity may not be directly reliant on plasma testosterone or estradiol concentration given the lack of differences of these hormones during our murine model of surgical sepsis.
Despite the lack of differences in circulating estradiol in our cohort of animals, the protective link between estrogen and the microbiome is well established.33,34 On one end, estrogen is positively correlated with gut microbiota species richness and α diversity in both males and females.35 This relationship is supported by a reduction of diversity under estrogen deprivation states such as menopause.36 Importantly, this relationship has been reported to be responsible for the sexually dimorphic protection against metabolic stress in cycling females and to mediate the preventive effect against chronic low-grade inflammation.37 Of note, the gut microbiome has also been reported to regulate production of estradiol, particularly in postmenopausal women, an aspect that is termed the “estrabolome” and defined as the aggregate of enteric bacterial genes whose products are capable of metabolizing estrogens, which has been linked to breast cancer.38 Thus, this all suggests a complex relationship and potential source of variability when trying to link estradiol levels to microbiome changes across different disease states and ages.
Our study is limited in that we were not able to monitor the estrogen cycle of the mice, ensuring that all cycling female were at the same stage of their estrus cycle when measurements were taken.39 Of note, mice in general are considered acyclic or have lower estrogen starting at 18 months of age.39 However, our primary goal was to determine if there were differences in the concentrations of testosterone or estradiol at days 7 and 14 after surgical sepsis that may explain changes in the microbiota. In addition, it has been stated that having mice all at the same stage of estrus may not be necessary for all sex-based murine research.32 Despite the inability to “normalize” females in their estrus cycle, the determination of plasma estradiol and testosterone levels supports that microbiota changes were primarily due to CLP-DCS. In addition, future studies addressing only DCS may be useful to differentiate changes secondary to sepsis from those induced by chronic stress. However, our model has been validated to replicate the clinical patient scenario and is thus advantageous in the current study.22,29,40 Finally, we were limited in the total number of elderly mice available for these studies, which may have an impact on the observed changes in α and β diversity. Future studies should focus on these older age groups with microbial reconstitution studies in an effort to “rescue” older mice from a state of PICS.
In conclusion, given the prior clinical studies whereby females, for unknown reasons, recover more expeditiously after sepsis and ICU stress than males, it is imperative that the factors explaining these differences between the sexes are determined. Herein we identify that changes in the intestinal microbiota occur after sepsis with chronic daily stress and, while changes in diversity are initially present in both male and female mice, female mice demonstrate a unique microbiota at 2 weeks after sepsis (vs. male mice), and this may be due to microbial resilience that is not present in the male mice. While further studies are required, our work may explain the persistent inflammatory state in males versus females that may make them more susceptible to CCI/PICS.
Supplementary Material
AUTHORSHIP
P.A.E., D.B.D., E.C.L., J.M., L.K., B.F., D.C.N., R.F.U., M.L.D., J.R., R.T.M., C.L., F.A.M., S.C.B., T.C.F., O.L., G.C., L.L.M., A.M.M., and R.M.T. contributed in the conception and design of the project, data acquisition, data analysis, and interpretation, as well as revision of article content and approval of its final form. All authors attest that they meet the current ICMJE criteria for authorship.
This work was supported, in part, by the following National Institutes of Health grants: R01 GM-113945, R01 GM-104481 (L.L.M.), R01 AG-037984 (T.C.F.), and P50 GM-111152 (L.L.M., P.A.E.), awarded by the National Institute of General Medical Sciences. In addition, this work was supported, in part, by a postgraduate training grant T32 GM-008721 (D.B.D., J.M., L.K., B.F.) in burns, trauma, and perioperative injury by National Institute of General Medical Sciences and the McKnight Brain Research Foundation. Finally, this work was supported by NIH R21 AG072011 (O.L.).
DISCLOSURE
The authors declare no conflicts of interest.
DISCUSSION
SUSANNAH E. NICHOLSON, M.D. (San Antonio, Texas): I congratulate Dr. Efron on his presentation of this impactful work. To date there is a knowledge gap regarding sex differences in post-sepsis morbidity and mortality and the relationship to the microbiota.
This study has the potential to contribute to the field of critical care medicine and to expand our collective knowledge on the human microbiome and disease.
This group utilized a murine abdominal sepsis model, cecal ligation and puncture, combined with a daily chronic stress to study sex differences in the microbiota over time to mimic an ICU stay.
They performed sequencing, targeting the V3-V4 region of the 16S rNA gene to evaluate the gut microbiota. The authors’ novel study found differences in diversity of the gut microbiota at Day 7, both males and females, compared to baseline.
However, the microbiota diversity in the female animals recovered at 14 days; whereas, the diversity in the males did not. This study has the potential to generate additional research in the field. I have several questions, though.
Number 1. What is the sample size of mice receiving cecal ligation and puncture, plus daily chronic stress? And how many were male versus female? How did you determine the group size? Was a power analysis done?
Number 2. When and how were the baseline samples obtained? Were the samples taken from stool from the animals prior to anesthetic expressed per rectum, or were they cecal samples obtained during the puncture procedure?
Number 3. Regarding the statistical analysis used, you assessed the beta diversity of the microbiota using the Bray Curtis Dissimilarity Index. However, a number of other indices are also routinely used in addition to Bray Curtis, and frequently a perma nova on a Bray Curtis Dissimilarity Index is employed to test for differences in strength in the statistical analysis. Was this considered during the analysis?
Number 4. Cohousing effects were addressed but the statistical modeling take into account variability and diversity resulting from factors such as batch effect and interspecies variability depending on shipment of animals, especially given that the animals were from two different facilities?
Number 5. You make the case that female animals demonstrate the resilience of their microbiota at 14 days, despite both males and females having altered microbiota at 7 days.
However, you also assert that the improved resiliency of the female microbiota may be secondary to changes imparted on the host microbiota by impact of sepsis and chronic stress. This does not really explain why there is still these sex differences.
Was there any consideration to doing a gonadectomy or employing hormone antagonists to see if the sex difference disappears? Are other hormones responsible or contributing such as progesterone?
Are there variations in hormonal levels occurring outside of the time points when levels were collected that could account for the microbiota differences observed?
I’d like to thank the AAST for the privilege of discussing this impactful work. It’s insightful, informative, and will contribute to the current literature on gut microbiota in the setting of sepsis and critical care medicine.
TIMOTHY A. PRITTS, M.D. (Cincinnati, Ohio): Dr. Efron, you showed the differences at 14 days between males and females. At what point do the males recover, if ever?
PHILIP A. EFRON, M.D., F.A.C.S., F.C.C.M. (Gainesville, Florida): Dr. Pritts, to answer your question, that’s a very important question.
One thing I have to point out, first, about these aged mice, we get very limited numbers from the National Institute of Aging. They’re actually rationed. And we don’t get many of them for free so in order to do these experiments you have to buy them from a vendor like Jack’s Mice.
Each one of these mice is well over $300. So there are limitations, overall, as to how much we can do, the power analysis and how much analysis we can do, which is one of the reasons that this was sort of a we used male and females combined for both ages to try and reach appropriate statistical analysis.
So from that standpoint it is unclear yet when they resolve. Like we have grants that are moving forward to try and identify exactly how long and far out we can take these mice to sort of determine how long it does take for these individuals or, not individuals but these mice to recover. So it’s an important question. We’re just somewhat limited by the expense of the animals.
As for the first question about the sample size, it’s a very important point. We have sort of discussed in sort of our response to the reviewers that there are ways to actually, without doing a proper sample analysis, power analysis using FDR, to actually determine if, prevent the need to determine Type 1 or Type 2 errors have occurred.
So using our biostatistician, Dr. Li, who did this work with us, we were actually able to sort of avoid that which may have increased the number of mice we had to use.
I would say that for each group there were probably at least 30 mice which had to be sacrificed at different times. And overall, again, the statistical analysis was appropriate and sort of validated by several articles that say you can use these methods.
As for the stool samples, these actually, there may be some confusion. I have to explain in the paper a little better. We did not actually take stool samples from the animals at the time before they underwent CLP and DCS.
What we actually did was do the procedure, give them the antibiotics and volume resuscitation and then specifically the DCS and euthanize them. And at that point, under a laminar hood, we expressed the stool out from the left colon and used that into a sterile vial.
This being said, the naïve mice that I described get neither antibiotics, chronic stress, or infection, are actually used as our baseline. So we’re not actually comparing it to the mouse prior to but clonal mice that are specifically used for the time before.
Again, this is somewhat based on the fact that there is difficulties in being able to appropriately get the stool samples out of these animals from the same place sterile at each time, as well as trying to restrict overall total number of mice we have to use for these things.
The perma nova is actually a very good point. Many people actually do use this in their beta diversity and other sort of analysis points. And, again, talking to our statistician, Dr. Li, he says this is an outstanding point but that this isn’t particularly necessary to do the beta diversity that we were looking at.
We were trying to keep it similar to the initial analysis that we did with our older versus young mice at seven days, which we published and demonstrated was (Indistinguishable).
The batch effect, at this time we actually did save up the stool amounts and the – excuse me, the stool and the plasma so that there were not multiple batch effects when we did this and that they were analyzed at the same time.
I would also say that very few of the mice we used, less than 5 probably overall, were from the NIA as opposed to Jack’s. Again, we are somewhat not required but due to costs to use these mice when they are available.
I can say that although maybe not a perfect analogy because they’ve grown up in a different environment, we do have them spend at least four weeks together in the same cages so that through corpa fascia they obtain the same microbiome overall.
As for the gonadectomy and homoagonist, this is an outstanding point. We are actually specifically writing a grant at this point to do just that. And to take some of these animals out to these longer time points to do exactly what the reviewer asked which is determine what the effect is overall.
To be honest, with the hormones themselves, being female or being male isn’t necessarily7 the amount of hormone circulating at the time.
It is relevant to a mouse, at least a year’s worth of exposure to one or two of the other hormones that can have multiple effects in your body and different end organs, the epigenome, the transciptome and the microbiome.
And this is also true in males and females where you have a minimum of potentially 40 years of exposure to estrogen and testosterone which can have different effects on the body leading to these results that would be different when someone is potentially a little bit older and having sepsis. But these are, there is a key issue that needs to be addressed and experiments that need to be done.
Other types of hormone therapies, again, this is important to look at. There are multiple hormones that need to be looked at.
And, again, overall, I think there are even better analyses of estrogen and testosterone that could potentially be done, although the kits that we used are accepted throughout the country. But as many people who do hormone research realize, that testing these in mice can be extremely difficult and may require validation.
Other time points, again, are very important. It’s not feasible to necessarily euthanize all of these animals each day but a better understanding of the dynamics of what is occurring, it does absolutely need to be done so we can have a better idea of if or when an intervention and precision (Indistinguishable) needs to occur and how it tends to sort of go about.
So these are outstanding questions that we do need to address.
Footnotes
Published online: March 24, 2022.
This study was presented at the 80th Annual Meeting of the American Association for the Surgery of Trauma and Clinical Congress of Acute Care Surgery, September 29, 2021 to October 2, 2021, in Atlanta, Georgia.
Supplemental digital content is available for this article. Direct URL citations appear in the printed text, and links to the digital files are provided in the HTML text of this article on the journal’s Web site (www.jtrauma.com).
Contributor Information
Dijoia B. Darden, Email: Dijoia.Darden@surgery.ufl.edu.
Eric C. Li, Email: Chak.Li@medicine.ufl.edu.
Jennifer Munley, Email: Jennifer.Munley@surgery.ufl.edu.
Lauren Kelly, Email: Lauren.Kelly@surgery.ufl.edu.
Brittany Fenner, Email: Brittany.Fenner@surgery.ufl.edu.
Dina C. Nacionales, Email: Dina.Nacionales@surgery.ufl.edu.
Ricardo F. Ungaro, Email: Ricardo.Ungaro@surgery.ufl.edu.
Marvin L. Dirain, Email: mdirain@ufl.edu.
Jaimar Rincon, Email: Jaimar.Rincon@surgery.ufl.edu.
Robert T. Mankowski, Email: r.mankowski@ufl.edu.
Christiaan Leeuwenburgh, Email: cleeuwen@ufl.edu.
Fredrick A. Moore, Email: Frederick.Moore@surgery.ufl.edu.
Scott C. Brakenridge, Email: Scott.Brakenridge@surgery.ufl.edu.
Thomas C. Foster, Email: foster1@ufl.edu.
Orlando Laitano, Email: olaitano@fsu.edu.
Gemma Casadesus, Email: gcasadesus@ufl.edu.
Lyle L. Moldawer, Email: moldawer@surgery.ufl.edu.
Alicia M. Mohr, Email: Alicia.Mohr@surgery.ufl.edu.
Ryan M. Thomas, Email: Ryan.Thomas@surgery.ufl.edu.
REFERENCES
- 1.Gaieski DF, Edwards JM, Kallan MJ, Carr BG. Benchmarking the incidence and mortality of severe sepsis in the United States. Crit Care Med. 2013;41(5):1167–1174. [DOI] [PubMed] [Google Scholar]
- 2.World Health Organization . Editor improving the prevention, diagnosis and clinical management of sepsis (WHA 70.7). Geneva, Switzerland: Seventieth World Health Assembly; 2017. [Google Scholar]
- 3.Torio CM, Moore BJ. National Inpatient Hospital Costs: The Most Expensive Conditions by Payer, 2013: Statistical Brief #204. Healthcare Cost and Utilization Project (HCUP) Statistical Briefs [Internet]. Rockville, MD: Agency for Healthcare Research and Quality (US); 2016. [PubMed] [Google Scholar]
- 4.Gentile LF Cuenca AG Efron PA Ang D Bihorac A McKinley BA, et al. Persistent inflammation and immunosuppression: a common syndrome and new horizon for surgical intensive care. J Trauma Acute Care Surg. 2012;72(6):1491–1501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Stortz JA Murphy TJ Raymond SL Mira JC Ungaro R Dirain ML, et al. Evidence for persistent immune suppression in patients who develop chronic critical illness after sepsis. Shock. 2018;49(3):249–258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Stortz JA Mira JC Raymond SL Loftus TJ Ozrazgat-Baslanti T Wang Z, et al. Benchmarking clinical outcomes and the immunocatabolic phenotype of chronic critical illness after sepsis in surgical intensive care unit patients. J Trauma Acute Care Surg. 2018;84(2):342–349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Brakenridge SC Efron PA Cox MC Stortz JA Hawkins RB Ghita G, et al. Current epidemiology of surgical sepsis: discordance between inpatient mortality and 1-year outcomes. Ann Surg. 2019;270(3):502–510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zhang MQ Macala KF Fox-Robichaud A Mendelson AA Lalu MM, Sepsis Canada National Preclinical Sepsis Platform . Sex- and gender-dependent differences in clinical and preclinical Sepsis. Shock. 2021;56(2):178–187. [DOI] [PubMed] [Google Scholar]
- 9.Zhang M Montroy J Sharma R Fergusson DA Mendelson AA Macala KF, et al. The effects of biological sex on sepsis treatments in animal models: a systematic review and a narrative elaboration on sex- and gender-dependent differences in Sepsis. Crit Care Explor. 2021;3(6):e0433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Xu J Tong L Yao J Guo Z Lui KY Hu X, et al. Association of sex with clinical outcome in critically ill sepsis patients: a retrospective analysis of the large clinical database MIMIC-III. Shock. 2019;52(2):146–151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kim YS, Unno T, Kim BY, Park MS. Sex differences in gut microbiota. World J Mens Health. 2020;38(1):48–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Haak BW, Wiersinga WJ. The role of the gut microbiota in sepsis. Lancet Gastroenterol Hepatol. 2017;2(2):135–143. [DOI] [PubMed] [Google Scholar]
- 13.Kang H, Thomas RM. Bacteria and sepsis: microbiome to the rescue? J Clin Med. 2021;10(16):3578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Thomas RM, Jobin C. The microbiome and cancer: is the ‘oncobiome’ mirage real? Trends Cancer. 2015;1(1):24–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Rooks MG, Garrett WS. Gut microbiota, metabolites and host immunity. Nat Rev Immunol. 2016;16(6):341–352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kim SM DeFazio JR Hyoju SK Sangani K Keskey R Krezalek MA, et al. Fecal microbiota transplant rescues mice from human pathogen mediated sepsis by restoring systemic immunity. Nat Commun. 2020;11(1):2354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Knight R Vrbanac A Taylor BC Aksenov A Callewaert C Debelius J, et al. Best practices for analysing microbiomes. Nat Rev Microbiol. 2018;16(7):410–422. [DOI] [PubMed] [Google Scholar]
- 18.Dogra SK, Doré J, Damak S. Gut microbiota resilience: definition, link to health and strategies for intervention. Front Microbiol. 2020;11:572921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Fransen F van Beek AA Borghuis T Meijer B Hugenholtz F van der Gaast-de Jongh C, et al. The impact of gut microbiota on gender-specific differences in immunity. Front Immunol. 2017;8:754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.National Research Council (US) Committee for the Update of the Guide for the Care and Use of Laboratory Animals . Guide for the Care and Use of Laboratory Animals. 8th edition. Washington, DC: National Academies Press (US); 2011. Available at: https://www.ncbi.nlm.nih.gov/books/NBK54050/. Accessed April 11, 2022. [PubMed] [Google Scholar]
- 21.Franklin CL, Ericsson AC. Microbiota and reproducibility of rodent models. Lab Anim (NY). 2017;46(4):114–122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Stortz JA Hollen MK Nacionales DC Horiguchi H Ungaro R Dirain ML, et al. Old mice demonstrate organ dysfunction as well as prolonged inflammation, immunosuppression, and weight loss in a modified surgical sepsis model. Crit Care Med. 2019;47(11):e919–e929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Bokulich NA Subramanian S Faith JJ Gevers D Gordon JI Knight R, et al. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat Methods. 2013;10(1):57–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wang Q, Garrity GM, Tiedje JM, Cole JR. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl Environ Microbiol. 2007;73(16):5261–5267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.McCafferty J Muhlbauer M Gharaibeh RZ Arthur JC Perez-Chanona E Sha W, et al. Stochastic changes over time and not founder effects drive cage effects in microbial community assembly in a mouse model. ISME J. 2013;7(11):2116–2125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.McMurdie PJ, Holmes S. phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PloS One. 2013;8(4):e61217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Team RC . R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing; 2017. [Google Scholar]
- 28.Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc Series B Stat Methodol. 1995;57(1):289–300. [Google Scholar]
- 29.Mankowski RT Thomas RM Darden DB Gharaibeh RZ Hawkins RB Cox MC, et al. Septic stability? Gut microbiota in young adult mice maintains overall stability after sepsis compared to old adult mice. Shock. 2021;55(4):519–525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Valeri F, Endres K. How biological sex of the host shapes its gut microbiota. Front Neuroendocrinol. 2021;61:100912. [DOI] [PubMed] [Google Scholar]
- 31.Saito T, Ciobotaru A, Bopassa JC, Toro L, Stefani E, Eghbali M. Estrogen contributes to gender differences in mouse ventricular repolarization. Circ Res. 2009;105(4):343–352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Beery AK. Inclusion of females does not increase variability in rodent research studies. Curr Opin Behav Sci. 2018;23:143–149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Aguilera M, Galvez-Ontiveros Y, Rivas A. Endobolome, a new concept for determining the influence of microbiota disrupting chemicals (MDC) in relation to specific endocrine pathogenesis. Front Microbiol. 2020;11:578007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Yoon K, Kim N. Roles of sex hormones and gender in the gut microbiota. J Neurogastroenterol Motil. 2021;27(3):314–325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Flores R Shi J Fuhrman B Xu X Veenstra TD Gail MH, et al. Fecal microbial determinants of fecal and systemic estrogens and estrogen metabolites: a cross-sectional study. J Transl Med. 2012;10:253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Vieira AT, Castelo PM, Ribeiro DA, Ferreira CM. Influence of oral and gut microbiota in the health of menopausal women. Front Microbiol. 2017;8:1884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kaliannan K Robertson RC Murphy K Stanton C Kang C Wang B, et al. Estrogen-mediated gut microbiome alterations influence sexual dimorphism in metabolic syndrome in mice. Microbiome. 2018;6(1):205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Parida S, Sharma D. The microbiome-estrogen connection and breast cancer risk. Cells. 2019;8(12):1642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Brooks HL, Pollow DP, Hoyer PB. The VCD mouse model of menopause and perimenopause for the study of sex differences in cardiovascular disease and the metabolic syndrome. Physiology (Bethesda). 2016;31(4):250–257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Efron PA Darden DB Wang Z Nacionales DC Lopez MC Hawkins RB, et al. Transcriptomic responses from improved murine sepsis models can better mimic human surgical sepsis. FASEB J. 2021;35:e21156. [DOI] [PMC free article] [PubMed] [Google Scholar]