

Abstract
The heat shock proteins (HSPs) are ubiquitous and conserved protein families in both prokaryotic and eukaryotic organisms, and they maintain cellular proteostasis and protect cells from stresses. HSP protein families are classified based on their molecular weights, mainly including large HSPs, HSP90, HSP70, HSP60, HSP40, and small HSPs. They function as molecular chaperons in cells and work as an integrated network, participating in the folding of newly synthesized polypeptides, refolding metastable proteins, protein complex assembly, dissociating protein aggregate dissociation, and the degradation of misfolded proteins. In addition to their chaperone functions, they also play important roles in cell signaling transduction, cell cycle, and apoptosis regulation. Therefore, malfunction of HSPs is related with many diseases, including cancers, neurodegeneration, and other diseases. In this review, we describe the current understandings about the molecular mechanisms of the major HSP families including HSP90/HSP70/HSP60/HSP110 and small HSPs, how the HSPs keep the protein proteostasis and response to stresses, and we also discuss their roles in diseases and the recent exploration of HSP related therapy and diagnosis to modulate diseases. These research advances offer new prospects of HSPs as potential targets for therapeutic intervention.
Keywords: cancers, heat shock proteins, molecular chaperone, proteostasis, target therapy
Heat shock proteins (HSPs) are in charge of proteostasis and buffer the stresses. Malfunction of HSPs lead to many diseases. In this review, we discuss the current understanding about the functions and structures of the major HSP families, the HSP network in proteostasis and their roles in diseases, and the prospects of these proteins as potential targets for therapeutic intervention.
1. INTRODUCTION
In living organisms, cells are under constantly changing conditions and maintenance of cellular protein homeostasis is crucial for cell survival and integrity, 1 , 2 as protein misfolding and aggregation have been found to induce malfunction of proteins and lead to various diseases. 3 , 4 The ubiquitous and conserved heat shock proteins (HSPs), which exist in both prokaryote and eukaryote organisms, belong to a large family of proteins that are in charge of proteostasis. 5 They function in a wide range of cellular housekeeping processes, including the folding of newly synthesized polypeptide, refolding metastable proteins, protein complex assembly, the degradation of misfolded proteins, and dissociating protein aggregates. 5 , 6 Under normal conditions, HSPs account for 5–10% of the total cellular protein content and they work as an integrated network to maintain proteostasis. 5 Although under extreme conditions, the heat shock transcription factors (HSFs) are activated in response to stresses, leading the transcription of abundant HSPs to buffer the stresses. 7 , 8 And based on the primary molecular chaperon function, they also participate in multiple processes in eukaryotic cells, and malfunction of HSPs has reported to related with many diseases. In this review, we discuss the current understanding about the structures features and the chaperon functions of the major HSPs, and the regulation of HSPs by their cochaperons, describe the HSP network in proteostasis and how they response to the stresses. We also discuss their roles in cancers, neurodegeneration, autoimmune, inflammation, infection and cardiovascular diseases (CVDs), and summarize the prospects of these proteins as potential targets and biomarkers for therapeutic intervention, the development, and shortcomings of HSP inhibitors.
2. THE DISCOVERY OF HSPs
The protective ability of HSPs to cope with stress was discovered long before the function was understood. The chromosomal puffing induced by temperature shock was first reported on fruit fly in 1962, 9 and in the following years, similar chromosomal puffs were observed in different organisms including both prokaryotes and eukaryotes, and the puffing happened not only after heat shock, but also in response to other survival pressures, such as hypoxia, ischemia, reactive oxygen species (ROS), or endotoxins. 10 It indicates that this phenomenon executes protective function for cell survival under diverse stresses. 10 , 11 Moreover, the newly transcribed messenger RNA at the puff site induces the synthesis of HSPs in vivo. 12 , 13 Since then, identification and functional research of HSPs became the focus in this area.
Among the many HSPs, HSP60/70/90 families are the most studied HSPs. GroEL, the bacterial homolog of HSP60, was first reported in 1973 and found to participate in bacteriophage assembly. 14 , 15 Later, HSP60 homologs in plant chloroplast and in yeast mitochondria were also identified, and they facilitate the folding of peptides with adenosine triphosphate (ATP) and cofactor chaperonin‐10, and mediate protein importation into yeast mitochondria, indicating a role of HSP60 in protein folding and assembly. 16 , 17 In the few years later, the mechanism of GroEL‐GroES (as homologues of HSP60‐HSP10 in prokaryotes) folding machine was identified as a heptameric rings that encapsulate the substrates inside the complex and perform protein folding by ATP hydrolysis. 18 , 19 For HSP70 family, the first heat shock protein HSP70 was discovered in drosophila, then the Escherichia coli homolog of HSP70‐DnaK was found in the 1980s. In the later 1980s, evidence emerged that HSP70 can interact with the hydrophobic peptides and associate with nascent polypeptide in translation, 20 , 21 which was followed by the discovery of another two factors regulating the ATPase activity of HSP70, DnaJ and GrpE. 22 By reconstitution, the protein folding pathway in vitro, it was then found that protein folding is a highly organized cellular process that needs the cooperation of HSP70 and HSP60‐HSP10. 23 At this time, the molecular chaperon function of HSPs has been defined as to maintain the correct folding of polypeptide chains and assembly of protein complex. 21 , 24 In 1983, the first HSP90 gene was isolated from yeast, and then HSP90 protein was reported to associate with steroid receptors and Src kinase. This association is required for the functions of these proteins, as reduced HSP90 compromise the activity of steroid receptors. 25 , 26 Then in 1990s, it was revealed that HSP90 functions as molecular chaperones and participate in the folding of client proteins.
Later, it was observed that the HSPs are high evolutionarily conserved from bacteria to eukaryote species. 27 , 28 , 29 , 30 Since then, increasing understanding about the mechanism of HSPs were obtained by structural biology analysis, and the functions of HSPs expanded to almost every aspect in the life of proteins from de novo synthesis to degradation. 31 , 32
3. HSPs CLASSIFICATION
Based on molecular weights, HSPs are classified into large HSPs, HSP90, HSP70, HSP60, HSP40, and small HSPs families, the functions of which are involved in the entire metabolic process of proteins (Table 1). They function as holdase, foldase, sequestrase, aggregase, or disaggregase, and work as a network to keep the proteostasis in cells and also an efficient first line of defense in response to stress. 5 , 33
TABLE 1.
Brief summary of heat shock protein families’ members, cellular locations, and functions
| Family | Major Members and location | Characteristics | function | Reference | |
|---|---|---|---|---|---|
| Small HSPs | HSPB1‐HSPB10 |
Cytosol Mitochondria Nucleus |
ATP‐independent large heterogeneous oligomers |
Work as holdase to prevent aggregation; sequestrate misfolded proteins; |
72 , 80 , 81 |
| HSP40/DNAJ |
DNAJA DNAJB DNAJC |
Cytosol mitochondria nucleus |
J domain containing proteins Interact with HSP70 |
Serve as cochaperon of HSP70, regulating HSP70 ATPase activity; Hold misfolding substrates and recruit HSP70; |
22 , 40 , 43 |
| HSP60 | HSP60 | Mitochondria |
Double ring structure contains Two heptameric rings of HSP60, cooperates with HSP10; ATP‐dependent |
Protein foldase Prevents aggregation |
18 , 62 , 82 , 83 |
| TRiC | Cytosol |
Double ring structure contains two octameric rings of TRiC; ATP‐dependent. |
|||
| HSP70 |
HSPA1A/1B HSPA1L HSPA2 HSPA6 HSPA7 HSPA8 HSPA12A/12B HSPA13 HSPA14 |
Cytosol Nucleus |
Conserved structure containing NTD‐SBD domains; Chaperon function based on the allosteric conformation change cycle in ATP‐dependent way |
Multiple functions in proteostasis Work as holdase, foldase, prevent aggregation and triage the protein fates |
34 , 35 , 84 |
| HSPA5 | ER | ||||
| HSPA9 | Mitochondria | ||||
| HSP90 |
HSP90AA HSP90AB GRP9 TRAP1 |
Cytosol Cytosol Cytosol/ER Mitochondria |
Form homodimer and undergoes allosteric open‐closed conformation change in the folding process; ATP dependent |
Foldase for proteins de novo synthesized and refold misfolded proteins; major substrates include kinases, steroid receptors | 4 , 54 , 85 |
| Large HSPs |
HSP110 GRP170 |
Cytosol ER |
Belong to HSP70 superfamily |
Holdase; keep proteins from aggregation; Cochaperon of HSP70 |
47 , 48 , 77 |
3.1. HSP70
HSP70 /DnaK are conserved molecular chaperon proteins ubiquitously expressed in prokaryote and eukaryote organisms. 34 DnaK is expressed in the prokaryotes, and HSP70 is the eukaryote counterpart. In human genome, there are about 17 genes encoding multiple HSP70 proteins. 35 The expression of some of them is stress induced, called HSP70s, which activate to deal with the environment changes, the others are housekeeping HSC70 (heat shock cognate) that are constitutively expressed. 34 , 35 , 36 HSP70s are expressed in different cellular locations, including cytosol, nucleus, endoplasmic reticulum (ER), mitochondria, and secreted, and they keep the dynamic balance of the synthesis, folding, degradation, and translocation of proteins. 34 , 35 , 36
HSP70 proteins shares conserved structure features. The N‐terminal nucleotide binding domain (NTD) ligate to the C‐terminal substrate binding domain (SBD) by a flexible linker. SBD domain includes two subdomains, SBDα and SBDβ, followed by a C‐terminal tail with EEVD motif (conserved Glu‐Glu‐Val‐Asp, EEVD) that recognizes several cochaperones (Figure 1A). 34 The chaperon activity of HSP70 is based on the allosteric conformation change of HSP70, which is ATP‐dependent. In this process, ADP‐bound HSP70 recognizes the substrate at the SBD domain with high affinity. For DnaK in E. coli, it prefers peptides with a hydrophobic core of five residues, 37 and for HSP70, it seems to be more promiscuous for the substrate recognition. 38 Then nuclear exchange factors (NEFs) mediate the exchange from ADP to ATP, and ATP induces the rotation of HSP70 at the NBD lobes and the SBDα helix lid covers on the NBD. This ATP‐bound conformation of HSP70 has low affinity for substrates, causing the release of substrates. 34 , 39 Upon the interaction with the cochaperone J‐domain protein (JDP/HSP40) with HSP70, HSP70 undergoes ATP hydrolysis and returns to the ADP‐bound mode, triggering another round of cycle. 40 , 41 The released substrate could rebind HSP70 and undergoes further folding cycles until it reaches its native state (Figure 1B).
FIGURE 1.

(A) Schematic diagram of the HSP70 domains. (B) The conformational cycles of HSP70. (C) Schematic diagram of the HSP90 domains. (D) The ATP‐dependent chaperone cycles of HSP90. (E) The structures of half football (PDB ID: 6MRD) 63 and full football assembly (PDB ID: 6MRC) 63 of HSP60‐HSP10, TRiC (PDB ID: 4A0O), 64 images in Figure 1E adapted from Refs. 63 and 64. (F) The model of HSP60‐HSP10 chaperone cycles
The functions of HSP70 are regulated by the cochaperones binding to HSP70 at different domains. 42 The major cochaperones coupling with HSP70 NBD are J domain protein (JDP)/HSP40 and nucleotide exchange factor (NEF) families. The JDP/HSP40 families are not only chaperones by themselves, but also act as cochaperone of HSP70. 43 JDP/HSP40 recognize the hydrophobic features on the unfolded or nascent proteins and hand over the substrates to HSP70. 43 In addition, HSP70‐HSP40 complex elevates the HSP70 ATPase and folding activity, facilitating the binding with substrates, and preventing aggregates. 40 Another family of ubiquitous cochaperones that bind to HSP70 are NEFs, which in eukaryotes include three major classes, HSP110, BAG, and HSPBP1. NEFs interact with HSP70 at the NBD domain and mediate the dissociation of ADP and the rebinding of ATP. Different NEFs coupled with HSP70 serve diverse cellular functions. In the BAG family, BAG1 has been reported to mediate HSP70 with the proteasomal degradation pathway, whereas BAG3‐HSP70 complex is a hub for controlling protein aggregation. 44 , 45 , 46 HSP110 not only functions as a chaperon protein, but also is the most abundant NEFs of HSP70, promoting protein disaggregation. 47 , 48 Besides HIP protein that not belongs to the JDP and NEFs family also interacts with HSP70 at the NBD domain. HIP competes with BAG1 for HSP70 binding, and prevents the nucleotide exchange, which delays the release of substrate, indicating that it is important to avoid the premature substrate release. 42 Meanwhile, the C‐terminal EEVD motif of HSP70 interacts with E3 ligase CHIP, mediating the substrate degradation through ubiquitin‐proteasome system (UPS). 49
3.2. HSP90
HSP90s are highly conserved HSPs with molecular weight of 90 kDa. They are ubiquitously expressed in all species except for archaea. In human genome, there are six genes encoding HSP90, which are either expressed in cytosol (induced expressed HSP90AA and constitutive expressed HSP90AB), 50 , 51 ER (GRP94), or mitochondrial (TRAP) (Table 1). 52 , 53 , 54
All the HSP90 homologues share common structure elements. The HSP90 monomer contains three domains, including nucleotide‐binding domain (NTD) at N‐terminal, a middle domain (MD), and a C‐terminal domain (CTD) driving the homodimer of two subunits of HSP90 and contains an Met‐Glu‐Glu‐Val‐Asp (MEEVD) motif for cochaperone coupling (Figure 1C). 55 HSP90 folding activity is based on ATP‐dependent dynamic conformation changes and regulated by the multiple cochaperones. Without nucleotide bound, HSP90 stays in an NTD‐open conformation, forming dimers through the CTD domain. The unfolded client proteins are loaded to HSP90 in open conformation at the MD domain, and ATP binding induces the dimerization of NTD. The following ATP hydrolysis leads to a more twisted and closed conformation of HSP90, and ADP is released from HSP90, turning the conformation back to open mode. By repeating this cycle, the client proteins are folded and released in native conformations (Figure 1D). 56 In human proteome, about 10% proteins are HSP90 clients and their maturation is dependent on HSP90. By analyzing the clients’ categories by mass spectrum, Barrios‐Rodiles et al. found that HSP90 interacts with the relatively abundant proteins in human proteome, including transcription factors, ubiquitin proteins, and about 60% kinase proteins in human kinome. 57
In eukaryotes, many cochaperones of HSP90 have been found to regulate the function of HSP90. Some cochaperones adjust the ATPase activity of HSP90. For example, AHA1 is a cochaperone that stimulates the ATPase activity; meanwhile, p23 suppresses HSP90 ATPase activity by allosterically switching the conformation of the catalytic loop in HSP90. 58 Several tetratricopeptide repeat (TPR) domain‐containing cochaperones interact with the MEEVD peptide of HSP90, the most studied TPR protein is HSC70/HSP90‐organizing protein HOP, which stabilizes HSP90 in open conformation and behaves as a linker for substrate handover from HSP70 to HSP90. 59 The recognition for substrates also depends on general cochaperons, such as cochaperone CDC37, which is a universal cochaperone for kinase‐HSP90 interaction. Cochaperones also take part in the protein triage, as CHIP and Sgt1 are required for efficient proteasomal degradation of misfolded proteins.
HSP90 proteins function not only as protein foldases, but also participate the protein conformational maturation. For example, the structure stabilization of oncogenic kinases with activation mutations depends on HSP90 chaperon. Exposure to HSP90 inhibitor leads to the instability and degradation of kinase clients through proteasome. Due to its extensive involvement of the protein stability and activity, HSP90 plays many roles in cell survival derived from the chaperone function, such as cell signaling and cell cycle regulation.
3.3. HSP60
HSP60 is one of the most conserved proteins expressed in all species, and is also named as chaperonin or Cpn60. Two groups of chaperonins are found to be expressed in different locations in cells. Group I HSP60s are expressed in prokaryotes (known as GroEL) and in the eukaryotic mitochondria and chloroplasts, and they work with cofactor HSP10 (GroES in prokaryote). Group II HSP60s are distributed in archaea and the eukaryotic cytosol (TRiC or CCT). 60
The two types of chaperonins adopt similar conformation and substrate folding mechanism, both of which assist the protein folding through the ATP‐dependent cylinder folding cage. The group I GroEL‐GroES system is composed of three domains, an equatorial domain including ATP binding pocket, an apical domain, and an intermediate hinge domain in between. GroEL assembles into a back‐to‐back double ring structure with seven subunits in each ring. 61 Compared to the GroEL‐GroES complex, the eukaryotic HSP60‐HSP10 system shows some different features. The double ring structure of HSP60 in eukaryotes depends on the cofactor and nucleotide interaction. HSP60 exists as a single heptamer ring without HSP10 and ATP. In an ATP‐dependent manner, the double‐ring structures are formed between single rings and HSP60/HSP10 forms football‐type complexes (Figure 1E). 62 , 63 For group II chaperonins, TRiC works independent of cochaperone HSP10(Figure 1E). An extra domain in the apical domain of TRiC forms an iris‐like structure that functions as the lid of TRiC. TRiC forms octamer complex and utilizes a mechanism for protein folding similar to that of HSP60‐HSP10. 64 Upon ATP binding, heptamer HSP60 complex with HSP10 on the head region and the substrates are captured into the lumen of GroEL, after which the substrate is folded in the nanocage of HSP60 through ATP hydrolysis. Afterward, ATP binding with GroEL on the other ring induces the release of ADP and GroES, which allows the release of folded substrate (Figure 1F). Meanwhile, inadequately folded proteins can be encapsulated into GroEL again for further folding. 61 , 65 Chaperonin plays multiple roles in cells. For example, TRiC participates in the folding of about 10% new synthesized proteins. 66 Chaperonin also prevents the aggregation and refolding the denatured protein under stress. 67 Mitochondrial HSP60 is essential for maintaining the mitochondrial respiratory‐chain integrity. Meanwhile, HSP60s are dual‐directional regulators of apoptosis in response to extracellular or intracellular stresses. 68 , 69 , 70
3.4. Small HSPs
sHSP/HSPB are ATP‐independent molecular chaperones defined by their low molecular weights (12–43 Kda). Until now, about 10 small HSPs have been found in the cytosol and nucleus of mammalian cells with diverse functions as holdase, sequestrase, or aggregase. 71 Small HSPs are constitutively expressed to avoid protein aggregation, and they are upregulated rapidly under stress to buffer the protein unfolding pressure. 72
The common structural feature of small HSPs is the conserved α‐crystalline domain, 73 which is a β‐sandwich structure composed of seven or eight antiparallel β sheets. 72 On the N‐ and C‐side of ACD is the N‐ and C‐terminal regions (NTEs and CTEs), which are variable in sequence and length, and contribute to the flexibility of the small HSP structures. 71 In vivo, the HSPs form transient and plastic large oligomers as hollow spheres. During the formation of oligomers, sHSP monomers interact with each other through the ACD to form sHSP dimers. 74 Then using sHSP dimer as basic construction material, the oligomers assemble in diverse modes. In human HSPB2/B3, the NTE recognizes the groove formed in the ACD dimer. 75 While in other HSPs, such as Triticum aestivum HSP16.9, the dimers form higher oligomers through the NTE interactions between dimers. 76 In brief, the oligomers are dynamic and exchangeable depending on the stress or temperature. Unlike the foldase activity of HSP90/70/60 chaperone, sHSPs have a distinct holdase activity that is ATP‐independent. sHSPs dimers interact with the early‐unfolded intermediate substrates and sequestrate them in the core of the sHSP/substrate complexes to avoid aggregation. Because sHSPs do not have foldase activity, the sequestrated substrates can be triaged for refolding by HSP70/90 chaperon system. Meanwhile, some other sHSPs have aggregase activity and promote ubiquitin‐dependent aggregation of substrates under diverse stresses.
3.5. Large HSPs
The large HSPs, including HSP110 and Grp170, are HSPs with large molecular weights found in eukaryotes. HSP110 is universally distributed in both cytosol and nucleus, whereas Grp170 is specially located at ER. They are evolutionarily conserved and homologous to the HSP70 family, thus often categorized as members of the so‐called “HSP70 superfamily.” Both of them are expressed constitutively in cells, and under stresses, they can also be induced rapidly to buffer the proteostasis. 77 They work as holdases to prevent the aggregation of substrates. The yeast HSP110 (sse1p) also mediates both the ubiquitin‐dependent and ubiquitin‐independent degradation by interacting with the 19S subunit of the 29S proteasome. 78 Meanwhile, HSP110 is a cochaperon of HSP70, acting as NEF for the ADP‐ATP exchange. Cochaperone HSP‐110 is not only important for the cellular protein folding capability of HSP70, but also essential for the HSP70 disaggregation machinery and participates in the HSP70‐mediated degradation pathway. 79
4. DIVERGENT FUNCTIONS OF HSPSs
HSPs are widely distributed in cells and participate in a variety of cellular processes to keep cell integrity, maintain protein homeostasis, and response to stresses. They also participate in regulation of cell cycle, apoptosis, signal transduction, and other physiological processes. In fact, the wide roles of HSPs are based on the chaperon functions of stabilization multiple substrates that participate in almost every cell process.
4.1. Proteostasis network of HSPs
Protein synthesis and elimination are a dynamic cellular equilibrium process. In the proteome of mammalian cells, there are over10,000 proteins expressed, making it critical for proteostasis and cell survival to maintain protein stability and remove the aberrant proteins in the crowded cell (protein concentration > 300 g/L). 2 The folding of nascent proteins to their native conformation requires the assistance of HSP chaperon network. In this process, nascent proteins are synthesized on ribosome as linear chains, which must then be folded into correct conformations to execute their functions. For proteins with less than 100 amino acids, they can be folded to native state spontaneously and rapidly. 86 But the folding of larger proteins (>100 aa) or multidomain proteins are inefficient in cells and hampered by the increased number of folding intermediates or partially folded proteins 87 as well as tendency for protein aggregation upon exposure to solvent in the hydrophobic region of partially folded proteins. 88 , 89
Both prokaryotic and eukaryotic organisms share conserved molecular chaperons to keep the orchestrate of the proteome stability. HSP70, HSP90, and HSP60 families act as foldases and participate in the de novo folding of nascent proteins, and as holdases—the large and small HSPs, preventing the proteins from aggregation. During translation on ribosome, ribosome‐binding chaperons interact with the nascent peptides. Trigger factor 90 in bacteria first integrates with nascent peptide chain to assist the folding of 70% proteins, 90 , 91 and then the unfolded proteins are passed to the hub of folding network‐DnaK, which is the abundant DnaK‐DnaJ complex and fold about 20% proteins in bacteria proteome. The folding of the remaining 10% unfolded proteins to native conformation is accomplished on the GrpE fold machine. In eukaryotes, rather than the prokaryotic TF, nascent chain‐associated complex (NAC) and ribosome‐associated complex (RAC) associate with nascent polypeptides once they are synthesized on ribosomes. 92 , 93 The unfolded proteins are then delivered to HSP70‐HSP40, which work as an intersection before downstream folding machineries. On the one hand, HSP70 assists the folding of a fraction of proteins to native state. On the other hand, HSP70 connects the downstream folding chaperons and delivers the partially folded intermedia to HSP90 or TRiC chaperons. Some proteins in eukaryotes are transferred directly from HSP70 to HSP90 system by cofactor HOP for further folding, 94 including kinases important for cell signaling transduction. 57 By interacting with HSP90, the kinases are structurally stabilized and active in functions. 95 The complicate chaperone mechanism is lack of structure basis due to the transient and dynamic nature of client proteins folding process and the client–chaperon complex structure is hard to be solved. Recently, using GR as client model, the Agard group reported the chaperon complex structure with client protein loaded by cryoelectron microscopy. The structure of Hsp90–Hsp70–Hop–GR complex describes the molecular basis of HSP90 chaperon cycles in coordination with HSP70, 96 and based on the Hsp90–p23–GR structures, the complete remodeling cycle of client by chaperons was described for the first time. 97
The protein folding pathways in bacteria and eukaryotes suggest that the coordination of protein folding relies on the different recognition modes of HSPs. HSP70 shows plasticity and promiscuity in substrate binding by recognizing the hydrophobic core motif. 38 Therefore, HSP70 can participate in the folding of most of the intracellular proteins once they are synthesized and their hydrophobic residues exposed. TRiC recognizes a subset of domain‐specific folding intermediates folded by HSP70. Meanwhile, HSP90 recognizes substrates based on thermal and conformational stability. 57
Besides the folding of de novo proteins, the denatured proteins induced by stress or misfolded proteins are recognized by the HSP70 system for the protein triage decision, and some proteins can then be refolded to native state by HSP70 with assistance of HSP90. 5 , 6 However, the irreversibly misfolded proteins are transferred to downstream degradation machineries via either ubiquitin proteasome pathway or autophagy by coupling with multiple cochaperones. During proteasome degradation, CHIP, an E3 ligase, induces ubiquitylation of misfolded proteins and initiates proteolysis through proteasome by interact with EEVD domain of HSP70/90 and the a‐helical lid subdomain of HSP70 through the TPR domain. 49 CHIP preferentially ubiquitinates the substrates that stay with Hsp70 longer than others. Besides CHIP‐mediated degradation, BAG1, the NEF of HSP70, also induces protein degradation through TRC8 E3 ligase. 98 Meanwhile, BAG3 mediates degradation by autophagy and lysosome pathways (Figure 2). 44 , 45 , 46
FIGURE 2.

The proteostasis network of heat shock proteins. The heat shock proteins participate in the folding of de novo synthesized peptides, also they hold the metastable proteins and folding intermediates, preventing them from aggregation. In addition, HSPs and their cochaperones also take charge of protein quality control and connect with protein degradation pathways
4.2. Stress response
Besides housekeeping functions, HSPs are also important for cells to cope with various stresses. In response to stresses, such as heat, hypoxia, oxidative stress, DNA damage, or accumulation of misfolded proteins, heat shock response (HSR) rapidly triggers the upregulation of multiple HSPs to buffer the disordered cellular environment and proteostasis, restoring cell structure and cellular metabolism. 99 The protein family of HSF acts as stress sensors in cells. 7 Under normal conditions, they are inactive in complexes with HSPs. Upon stress, they are released from the HSP‐HSF complexes, allowing HSF to oligomerize, translocate to nucleus, and bind to the promoter regions of multiple HSPs, triggering their rapid transcription. 99 , 100 However, not all the HSPs are regulated by HSF at the transcription level. Posttranscription and translational regulations are also key mechanisms for HSP expression. Under stressed conditions, 5′UTR methylation of stress‐induced transcripts are preserved by m6A “reader” YTHDF2, which, in turn, initiates the translation of HSPs in cap‐independent way, providing a novel regulation mechanism for stress induced mRNA translation. 101
In stress conditions, cell death may be executed through both the intrinsic and extrinsic apoptosis. HSPs play dual roles in the regulation cell apoptosis with complicated mechanisms. In the intrinsic pathway, HSP70 inhibits the cytochrome c release by forming Bax‐HSP70‐HSP40 complex attenuating Bax mitochondria translocation. Meanwhile, HSPs can also regulate apoptosis through their canonical chaperon activity. For example, HSP90 and HSP70 can attenuate apoptosis through stabilizing and activating AKT signal pathway, which facilitates cancer cell survival. 102 The inhibitor of apoptosis proteins (IAPs), such as c‐IAP1, XIAP, and cFLIPS/L (CFLAR), are also reported as obligate Hsp70 clients. 103 By stabilizing IAPs, RIP1‐dependent apoptotic and necroptotic cascades are suppressed by HSP70. 104 It has been recently reported that Bim act as HSP70 cochaperon and enhances the ATPase and oncogenic protein chaperon activities of HSP70 in order to prevent apoptosis in cancer cells. 105 In addition, BCR‐ABL fusion protein induces the interaction of Hsp70 with Bim to prevent apoptosis in BCR‐ABL positive cells. 106 Downstream of the mitochondria apoptosis pathway, direct HSP70‐Apaf1 interaction and HSP27‐cytochrome c interaction prevent apoptosome formation, thus inhibiting caspase 3/9 activations and terminating the intrinsic apoptosis pathway. 107 , 108 HSPs also participate in the regulation of extrinsic apoptosis pathway. For example, HSP70 interacts with DR4/5 to suppress the formation of TRAIL‐mediated death inducing signaling complex (DISC). 109 In some breast cancer cells, TRAIL‐ and TNFa‐induced extrinsic apoptosis is inhibited by HSPB2 through suppressing caspases‐8/10 activations, which then blocking downstream Bid cleavage and the activation of caspase‐3. 110
Although most HSPs are prosurvival, Hsp60 exhibits bidirectional apoptosis regulatory function through several mechanisms. They chaperon with anti‐apoptosis proteins Bcl‐2 and Bcl‐xl to attenuate the apoptosis, 111 meanwhile stabilizing the survivin and CypD, which increase the cryoprotection and maintain the mitochondrial permeability. 68 , 112 In addition, HSP60 also suppresses the P53‐dependent apoptosis. 68 In contrast, in the presence of certain pathological factors, HSP60 can also exhibit proapoptosis ability. 82 Exogenous HSP60 is found to induce apoptosis through TLR4‐NF‐KB‐Caspase signal pathway in certain disease states. 113 , 114 Additionally, some apoptosis inducers such as staurosporine, disrupt the procaspase3–HSP60 complex, making caspase3 easier to be activated by cytochrome c, and this process is coupled with the release of HSP60 into cytosol. 115
Due to the comprehensive functions of HSPs in protecting cell integrity and hemostasis, it has been found that multiple cancers show chaperon addiction and take advantages of HSPs to maintain the oncogenic protein's structure and activity, promote cancer cell survival, evade from the immune system, attenuate apoptosis, and develop drug resistance. 116 Therefore, HSPs emerge as potential therapeutic targets in a variety of diseases.
5. HEAT SHOCK PROTEINS IN DISEASES
5.1. The role of heat shock proteins in cancers
In many malignancies, HSPs are frequently overexpressed and linked with poor prognosis, including lung cancers, gastric cancers, breast cancers, glioblastoma, PCs, and so on. HSPs have been reported associated with cancer cell proliferation, metastasis, and invasion. Thus, HSPs are effective biomarkers for cancer therapy (Table 2).
TABLE 2.
Role of heat shock proteins in cancers
| Name | Cancer type | Effects | References |
|---|---|---|---|
| HSP27 | Prostate cancer | HSP27 is required for EGF‐mediated EMT via modulation of the β‐catenin/Slug signaling pathway in prostate cancer. | 121 |
| HSP27 plays an important role in prostate cancer cell motility and metastatic progression | 138 | ||
| HSP27 confers resistance to androgen ablation and chemotherapy in prostate cancer cells through eIF4E | 139 | ||
| Colorectal Cancer | HSP27 is a key mediator in the progression and metastasis of CRC by regulating the store‐operated calcium entry. | 140 | |
| The elevated expression of HSP27 protein is a frequent event during the progression of CRC. | 141 | ||
| Lung cancer | Increased HSP27 correlates with malignant biological behavior of nonsmall‐cell lung cancer and predicts patient's survival. | 142 | |
| Chemoresistance of lung cancer stem like cells depends on activation of HSP27 | 143 | ||
| Breast cancer | Phosphorylation of Ser78 of HSP27 is correlated with HER‐2/neu status and lymph node positivity in breast cancer | 144 | |
| HSP27 induces SUMOylation of HSPB8 to promote HSPB8 expression, thereby endorsing proliferation and metastasis of breast cancer cells. | 145 | ||
| HSP27 is associated with decreased survival in node‐negative breast cancer patients. | 146 | ||
| HSP47 | Colorectal cancer | HSP47 promotes tumor survival and therapy resistance by modulating AKT signaling via PHLPP1 in colorectal cancer. | 147 |
| HSP47 is a predictive marker for lymph node metastasis in patients with colorectal cancer. | 148 | ||
| Prostate cancer | HSP47 confers chemoresistance on pancreatic cancer cells by interacting with calreticulin and IRE1α. | 149 | |
| Breast cancer | HSP47 promotes metastasis of breast cancer by interacting with myosin IIA via the unfolded protein response transducer IRE1α. | 150 | |
| Lung cancer | HSP47 promotes cell migration and invasion through AKT signal in nonsmall‐cell lung cancer. | 151 | |
| Glioblastoma | HSP47 plays an important role in GBM tumor formation, invasion, and angiogenesis. | 126 | |
| Head and neck cancers | HSP47 is associated with the prognosis of laryngeal squamous cell carcinoma by inhibiting cell viability and invasion and promoting apoptosis. | 152 | |
| HSP60 | Hepatocellular carcinoma | HSP60 exerts a tumor suppressor function by inducing cell differentiation and inhibiting invasion in hepatocellular carcinoma. | 153 |
| Colorectal Cancer | High HSP60 expression is important for CRC progression. | 154 | |
| Gastric Cancer | HSP60 overexpression is associated with the progression and prognosis in gastric cancer. | 155 | |
| Prostate cancer | HSP60 expression is strongly associated with prostate cancer lymph node metastasis. | 122 | |
| HSP70 | Leukemia | HSP70 expression is increased in the blood cells of patients with ALL, and inhibiting HSP70 could suppress cell proliferation and induce apoptosis. | 156 |
| HSP70 is involved in the progression of FLT3‐ITD‐positive AML. | 119 | ||
| Breast cancer | High HSP70 expression is found to be related with breast cancer lymph node metastasis. | 127 | |
| HSP70 is overexpressed in breast cancer. | 129 | ||
| Lung cancer | HSP70 promotes SUMO of HIF‐1α and promotes lung cancer invasion and metastasis. | 157 | |
| Prostate cancer | HSP70 binds to the N‐terminal domain of androgen receptor and modulates the receptor function in prostate cancer cells. | 158 | |
| Colorectal Cancer | HSP70 overexpression can predict poor survival in colorectal cancer patients. | 159 | |
| Head and neck cancers | HSP70 is a potential biomarker for detecting tumors and for monitoring the clinical outcome of radiotherapy in SCCHN patients. | 160 | |
| Glioblastoma | HSP70 promotes survival of C6 and U87 glioma cells by inhibition of ATF5 degradation. | 161 | |
| HSP90 | Colorectal cancer | HSP90 plays an important role in promoting EMT transition, migration, and invasion in colorectal cancer. | 132 |
| HSP90 is needed to cooperate with CD24 to enhance STAT3‐mediated VEGF transcription to inducing colorectal cancer angiogenesis. | 135 | ||
| Leukemia | HSP90 acts as the molecular chaperone and is highly expressed in several therapy‐resistant leukemia subtypes, thereby ensuring correct protein folding of several oncogenic proteins such as BCR‐ABL1 and FLT3‐ITD. | 120 , 162 | |
| Prostate cancer | HSP90 plays an important role in advanced prostate cancer growth and survival. | 163 | |
| Lung cancer | High HSP90 expression correlates with poorer overall survival in lung cancer patients. | 164 | |
| Ovarian cancer | HSP90 interaction with Lamin‐A is required for DNA damage repair and chemoresistance of ovarian cancer cells. | 165 | |
| Breast cancer | Elevated HSP90 expression in breast ductal carcinomas contributes to the proliferative activity of breast cancer cells. | 166 |
Leukemia is a hematological neoplasm that affects blood and bone marrow. Both acute myeloid leukemia (AML) and acute lymphoblastic leukemia (ALL) cells were shown to express high levels of HSPs. Patients with lower HSP expression usually have a higher rate of complete remission (CR) and their overall survival (OS) was considerably longer. 117 For example, FLT3 is the most frequently mutated gene in human AML 118 and HSP70 plays an important role in the progression of FLT3‐ITD‐positive AML. Knocking down inducible HSP70 was enough to destabilized FLT3‐ITD protein and inhibit cell proliferation and tumor growth in FLT3‐ITD‐positive AML cells. Thus, for FLT3‐ITD‐positive AML, targeting HSP70 could be a feasible treatment strategy. 119 p210(BCR‐ABL) or p185(BCR‐ABL) fusion is an oncogenic driving force in chronic myelogenous leukemia (CML). ABL tyrosine kinase is one of the HSP90 clients implicated in the development of chronic myeloid leukemia (CML). When the interaction between BCR‐ABL and HSP90 is disrupted, the cellular level of BCR‐ABL protein is reduced and the BCR‐ABL1‐STAT5 signaling pathway is thus inhibited, 120 suggesting that HSP90 appears to play an essential role in BCR‐ABL mutant leukemia. PC is the world's second most frequent malignancy in men. Metastasis is a great challenge for PC and it can be not cured by chemotherapy and ionizing radiation. In PCs, HSP27 has been reported as a key regulator of Hippo pathway, and the upregulation of HSP27 activates oncogenic and metastatic pathways by increasing the nuclear localization of transcription factor YAP. Besides, HSP 27 is also essential for EGF‐mediated EMT via β‐catenin /slug signaling pathway. 121 In poorly differentiated PC cells, HSP 60 protein is often overexpressed, which is strongly associated with lymph node metastasis. 122 Increased level of HSP 60 expression is related to poor survival of PC patients. Castration‐resistant PC (CRPC) is dependent on HSP60 through moderate mitochondrial spare respiratory capacity. 123
Glioblastoma multiforme (GBM) is an aggressive and lethal brain tumor in adults. 124 In GBM, mTORC1 signaling is hyperactivated and promotes growth and proliferation, and thus could be a promising therapeutic target in GBM. Downregulation of HSP60 enhances the ROS generation, resulting in AMPK pathway activation. The activated AMPK reduces protein translation by suppressing mTORC1‐mediated S6K and 4EBP1 phosphorylation, inhibiting cancer cell proliferation. 125 HSP47 has been reported to be highly overexpressed in GBM and is associated with tumor grade. HSP47 is involved in tumor development, invasion, and angiogenesis. The high expression of HSP47 in GBM cell lines leads to an increase in CD44+ cells, promoting the survival of GBM stem like cells by modifying the extracellular matrix of the tumor microenvironment (TME) via the TGF pathway. 126
Breast cancer is the main cause of cancer death among women worldwide. With the rapid drug discovery in the past few decades, a variety of drugs with different action mechanisms have improved the prognosis for breast cancer patients. However, metastasis is still a great challenge for cancer therapy. In breast cancer models, high HSP70 expression is found to be related with lymph node metastasis. 127 The epithelial‐mesenchymal transition (EMT) is a developmental process that permits stationary epithelial cells to migrate and invade as single cells. EMT plays an important role in prompting cell metastasis. 128 Inhibiting HSP70 have been shown to reduce invasion and migration by downregulating EMT pathway through modulating the expression of proteins involved in EMT such as N‐Cadherin, P‐Cadherin, Vimentin, and so on. 129 Furthermore, HSP70, together with HSP90, activates matrix metalloproteinase 2 (MMP‐2), thereby increasing the migration and invasion of breast cancer cells. 130
The most common cancer‐related cause of mortality is colorectal cancer (CRC), and metastasis is the major cause of death in CRC patients. 131 The nuclear factor kappa B (NF‐kB) and hypoxia‐inducible factor‐1 (HIF‐1a) are crucial in modulating CRC metastasis, both of which are client proteins of HSP90. The protein HSP90 has been linked to the promotion of EMT transition, migration, and invasion in CRC s. 132 In addition, angiogenesis also plays a significant role in tumor progression, including CRC. 133 Overexpression of CD24 is common in colorectal malignancies, and HSP90 is required for enhancing STAT3‐mediated VEGF transcription and initiating CRC angiogenesis. 134 Inhibiting HSP90 shows great efficiency in blocking CD24‐mediated CRC angiogenesis. 135 Besides, HSP110 is highly expressed in CRC and plays a role in the proliferation of CRC through activating STAT3 and promoting its transcriptional function. 136 Also, HSP110 is related to nodal metastasis that is a predictive marker for CRC metastasis. 137
5.2. Roles in neurodegeneration diseases
Alzheimer's disease (AD) is the most prevalent neurodegenerative illness, characterized by aberrant accumulations of hyperphosphorylated tau protein in brain cells, resulting in neurofibrillary tangles (NFTs) and improper folding of amyloid‐peptides (Aβ). 167 According to research, HSP not only plays a vital function in tau protein accumulation/degradation, but it also prevents Aβ‐related toxicity. HSP90/HSP70 is important in maintaining the normal physiological state of Tau protein as well as blocking aberrant phosphorylation and accumulation of Tau protein, and involved in the pathologic process of the AD‐associated Tau protein and Aβ. 168 , 169 , 170 Tau protein serves as a client for HSP90/HSP70 that can regulate Tau protein metabolism. Many studies have shown that inhibiting HSP 90 could reduce phosphorylation of Tau through proteasomal degradation. 171 HSP70 maintains tau and Aβ homeostasis through three mechanisms: inhibiting tau and Aβ aggregation, mediating tau return to microtubules, and accelerating Aβ clearance. 172 Many neurodegenerative disorders, such as AD, Parkinson's disease, and Huntington's disease, have been linked to mitochondrial malfunction. 173 Multiple aspects of mitochondrial function are hampered by Aβ, including the creation of ROS, the disfunction in energy metabolism, and so on. 174 It has been reported that Aβ can decrease the activity of complex IV and HSP60 can eliminate this effect of Aβ. One of the reasons is that HSP60 can inhibit Aβ amyloid peptide aggregation by disrupting fibrillogenesis. Despite the fact that HSP60 has been shown to have a neuroprotective role in AD, many studies demonstrate that upregulation of HSP60 expression can worsen the disease through triggering the release of inflammatory cytokines and accelerating neuronal cell death by binding to TLR4. 175 Parkinson's disease is distinguished by the accumulation of cellular misfolded α‐Syn protein and the death of dopaminergic neurons in the brain. HSPs are the most effective and well‐preserved cellular defense system in neurons, maintaining α‐Syn protein homeostasis by correcting protein folding, refolding partially misfolded proteins, and destroying potentially harmful aggregates. 176 , 177 HSPs also play a role in the chaperone‐mediated autophagy apparatus and inhibiting α‐Syn aggregation by the UPS. 176 , 178 , 179
5.3. HSPs in cardiovascular diseases
CVDs are a group of complex pathophysiological processes that occur in the heart or blood vessels, such as hypertension, atherosclerosis, coronary artery disease, arrhythmias, heart failure, and idiopathic LV heart dysfunction. Despite a variety of advanced treatments available, CVDs remain a leading cause of morbidity and mortality worldwide and also a major cause of disease burden in the world. 180 , 181 And HSP involves in normal physiological pathological functions of cells, especially in CVDs.
HSPs are immunodominant molecules that stimulate both innate and disease‐related immune responses. Inflammation causes cardiovascular dysfunction by causing cellular stress events such as apoptosis, oxidative and shear stress, as well as cellular and humoral immune responses, all of which impair the system's structure and function 182 (Table 3).
TABLE 3.
Heat shock proteins involvement in cardiovascular diseases
| Superfamily | Protein names | Clinical Manifestations | References |
|---|---|---|---|
| sHSPs | HSPB1 | Coronary artery disease | 183 |
| HSPB2 | protective effects against heart diseases such as cardiac hypertrophy and ischemia | 184 | |
| HSPB3 | Deregulation of myoblast viability | 185 | |
| HSPB5 | Desmin‐related myopathy | 186 , 187 | |
| HSPB6 | Decreased contractile function | 188 | |
| HSPB7 | No protection against age‐related dysfunction | 189 | |
| HSPB8 | Lack of protection after myocardial ischemia | 190 , 191 | |
| HSP40 | DNAJA3 | Respiratory chain deficiency | 191 , 192 |
| HSP60 | HSP60 | Protection from atherosclerosis and proliferation of vascular smooth muscle cells | 193 |
| HSP70 | HSPA1A | during myocardial ischemia/reperfusion HSP70 had protective effects | 194 |
| HSPA8 | |||
| HSP90 | HSPC4 |
Increased in plaque stability Cardiomyocyte apoptosis |
195 |
In CVDs, HSPs are a double‐edged sword. 196 On the one hand, HSPs are initially induced by stress stimuli. HSPs are expressed pronouncedly in CVDs, which play a cardioprotective role by inhibiting apoptosis. 197 Numerous studies have shown that preinduction of HSPs by mild stress has a protective effect on a more severe stress, which is related to the amount of induced HSP. 198 In addition, overexpression of HSPs in cultured cardiomyocytes or transgenic animals or intact hearts using viral vectors also has a protective effect, which directly demonstrates the ability of HSPs to protect cardiac function. 198 On the other hand, HSPs and autoimmune responses directed against them may be involved in the pathogenesis of atherosclerosis. For example, the expression of host‐protected HSP60 (hHSP60) on vascular endothelial cells may be the target of cross‐reactive autoimmune responses. Thus, there may be a cross‐reaction to the immune response of bacterial HSPs during infection due to the high sequence consistency between human and microbial HSPs. GroEL has high immunogenicity and cross‐reacts with hHSP60 expressed on endothelial cells, leading to endothelial dysfunction and atherosclerosis. This triggers an inflammatory cascade and accelerates the progression of atherosclerosis. Early fatty streak lesions may progress to atherosclerotic plaque formation if infections and cardiovascular risk factors persist. Under normal physiologic conditions, HSPs play a protective role in the arterial wall. However, due to their highly conserved sequences, HSPs expressed on the vascular endothelial cell surface can act as targets for detrimental autoimmunity in disease states. 199
Variety of HSPBs including HSPB1, HSPB2, HSPB3, HSPB5, HSPB6, HSPB7, and HSPB8 express in cardiac and skeletal muscle cells. These family members are generally considered as the focus of CVDs researches and the latest evidence highlights their role in cardiovascular protection. HSPB1 is a widely expressed multifunctional protein chaperone. Compared to healthy subjects, HSPB1 secretion decreased in human atherosclerotic plaques and HSPB1 levels are reduced in plasma. High expression of HSPB2 in heart has protective roles on some heart diseases such as cardiac hypertrophy and ischemia. 200 HSP70 also plays a role in the development of hypertension. Due to the relationship between genetic polymorphisms of HSP70 and essential hypertension, HSP70 level is elevated in hypertensive patients’ circulation and kidney. 201
There is a modulation between chaperones and cochaperones during the development of CVDs. Several CVDs occur due to out of physiological balance between protein synthesis, folding, and degradation resulting in accumulation of misfolded proteins. It is worth mentioning that different HSP proteins have opposite effect in promotion or inhibition of CVDs as well as related symptoms. Therefore, it is challenged to develop therapeutic strategies for CVDs based on HSPs that need balance their physiologic and pathologic roles. Better understanding of the relationship between HSPs and CVDs may speed up the development of new therapeutic strategies. 202
5.4. HSPs in autoimmune diseases
As we known, immune responses could be enabled by danger‐associated molecular pattern (DAMP), which is released by damaged or stressed tissues. HSPs were originally incorrectly classified as DAMPs, because HSPs are a group of constitutive and/or stress‐induced proteins, including thermal stress, infectious agents, intracellular stress, and so on. 203 So far, the immune response to HSPs has widely been studied in various inflammatory and autoimmune diseases with either immunoregulatory or immunostimulatory responses (Figure 3). Their bimodal and sometimes paradoxical roles in autoimmune diseases are outlined as follows.
FIGURE 3.
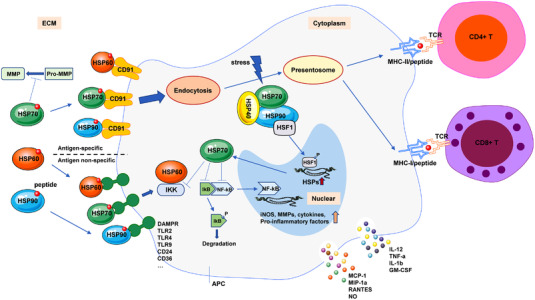
The HSP‐APC interaction modulates innate and adaptive immune responses
5.4.1. Small HSP in autoimmune diseases
Apart from chaperoning functions, the small HSPs also play an important role in cytoskeletal organization responding to cellular stress. 204 Among them, the most studied sHSP is HSP27. Aberrant phosphorylation of HSP27 was often increased in various autoimmune diseases, such as autoimmune skin diseases (pemphigus vulgaris and pemphigus foliaceus) 205 and myasthenia gravis (MG). 206 Anti‐HSP27 antibody levels were remarkably higher in broncho‐alveolar lavage (BAL) in patients with bronchiolitis obliterans (BOS) compared to lung transplant recipients without BOS. 207 HSP27 serum autoantibodies also correlate to Glaucoma (increased intraocular pressure) 208 and Guillain Barret syndromes. 209 NZBxW/F1 mice is a well‐established spontaneous lupus nephritis model. HSP27 could induce mesangial cell activation, which has a pivotal role in lupus phenotype development. 210 However, HSP27 can also exert anti‐inflammatory effect, as reduction of HSP27 leads to higher expression of pro‐IL‐1β and significantly more IL‐1β releasement in LPS‐treated monocytes. 211 In an ischemic acute kidney injury (AKI) mouse model, HSP27 was reported to play a protective role in kidney injury and neutrophil infiltration. 212
5.4.2. HSP40 in autoimmune diseases
HSP40 often exerts a pro‐inflammatory function in autoimmune diseases. HSP40 is reported as a biomarker of fibrillary glomerulonephritis, which is an autoimmune disease characterized by the extracellular deposition of nonamyloid fibrils. 213 DNAJB9, a member in HSP40 family, colocalizes with the fibrils, 214 indicating that HSP40 causes an immunostimulatory response in this context. In atherosclerosis, HDJ‐2, a homolog of human HSP40 found in E. coli, is highly expressed in atheromatous lesions, which may play an important role in T‐cell activation in the development of atherosclerosis. 215 HSP40/HSP70 expression is increased in stoke patients, which may lead to autoimmune responses against these HSPs. 216 Bullous pemphigoid, a bullous autoimmune disease, is characterized by the presence of autoantibodies against components of the dermal–epidermal junction. Compared to healthy control, circulating anti‐HSP40 autoantibodies were elevated in these patients. 217 Furthermore, HSP40 was found able to stimulate RAW264.7 cells to secrete IL‐6 by inducing PI3K/JNK signaling and inflammation. 218
However, HSP40 was responsible for the decline of pro‐inflammatory cytokine TNFα and the increase of anti‐inflammatory cytokine IL‐10 in the synovial fluid of juvenile idiopathic arthritis patients. Further study showed that the exposure of synovial fluid monocytes (SFMCs) to HSP40 peptide fragments induced Treg cells (CD4+, CD25high) with an increase of FoxP3, IL‐10, and CTLA‐4 mRNA, which may reverse the ongoing inflammation of the disease. 219
5.4.3. HSP60 in autoimmune diseases
HSP60 is mainly expressed in mammalian cells, whereas HSP65 is predominantly expressed in nonmammalian cells, like Mycobacterium. 220 HSP60/65 are HLA‐DR binders and able to present client peptides easily to antigen‐presenting cells (APCs). Self HSP60 undergoes a complete antigen processing within APCs and induces Th2 responses and tolerogenicity. On the other side, nonself HSP65 undergoes an incomplete antigen processing within APCs and induces Th1 responses and autoimmunity. 221 , 222 Molecular mimicry between human HSP60 and HSP65 could induce autoimmune phenomena. So, anti‐HSP60/65 autoantibodies were detected parallelly in autoimmune diseases, including SLE, Sjögren syndrome, undifferentiated connective tissue disease, 223 , 224 , 225 rheumatoid arthritis, 209 , 226 , 227 and autoimmune hepatitis. 228
Type I diabetes mellitus (T1D) is an autoimmune disease, and streptozotocin‐induced diabetes (STZ model) and nonobese diabetic 229 mice are two typical animal models for T1D. On the one hand, HSP60‐induced T‐cell activation and anti‐HSP60 antibody could be detected in experimental models of T1D, which contributes to diabetes aggravation. 230 On the other hand, HSP60 elicited a Th2 response and inhibited diabetes progression in an experimental STZ model. Immunization with mycobacterial HSP65 has also been found to prevent disease aggravation in NOD mice. 231 HSP60‐p277 peptide immunization in NOD mice induced a Th2 response with more IL‐10 and IL‐4 releasement, which was accompanied by Th1 response downregulation. 232
In a rat arthritis model, HSP60 administration showed increased number of Treg cells (CD4+, FoxP3+) in the joint‐draining lymph nodes and improved arthritis symptoms. HSP60 could induce Treg cell proliferation mediated by TLR9, leading to IL‐10 production. 233 Different domains of HSP65 exert different actions in autoimmune arthritis, and HSP65 P118‐388 causes Treg cells expansion and reduces autoimmune arthritis, whereas HSP65 P180‐188 does not. 234 Besides, immunization with HSP60/65 decreased autoantibodies against erythrocytes in mouse model of hemolytic anemia, 234 but increased inflammatory responses in mouse models of atherosclerosis 235 and intestinal autoimmune disease. 224
Interestingly, TLRs appear to play a key role in the immune regulation of HSP60/65. When HSP60/65 binds to TLR9/HLA‐DR, they induce a Th2 cytokine response in APCs. Otherwise, when HSP60/65 binds to TLR2/HLA‐DR, they induce a Th1 cytokine response in APCs. 221 , 236
5.4.4. HSP70 in autoimmune diseases
HSP70 sometimes induces an immunostimulatory response in autoimmune diseases by stimulating APCs to secrete more pro‐inflammatory cytokines, such as IL‐12, TNFα, and IL1β, and facilitate the maturation of immature dendritic cells. 237 , 238 , 239 HSP70/Ro52 and HSP70/Ro53 complexes were reported to induce more macrophages and cytotoxic T cells infiltration. 240 HSP70 serum autoantibodies were elevated in patients of thyroiditis, 241 inner ear disease, 242 and diabetic microangiopathy, 243 thereby involved in immune response modulation. In mouse model of salt‐sensitive hypertension, circulating anti‐HSP70 antibody was related to increased renal inflammatory infiltration. 244 Recombinant HSP70 immunization in RIP‐GP/P14 mice could induce the onset of diabetes, suggesting an in vivo pro‐inflammatory action of HSP70 in autoimmunity. 239
In most cases, HSP70s exert an immunoregulatory action in autoimmune diseases. Multiple HSP70 client peptides could bind to HLA‐DR and induce Treg cell expansion. In a mouse model of autoimmune arthritis, HSP70 treatment induced Treg cell expansion, elevated LAG3 expression in Treg cells, and increased IL‐10 production, resulting severity reduction of the disease. 245 Additionally, HSP70‐HINT1 (histidine triad nucleotide‐binding protein‐1) downregulated immune response through CD94 and NKG2D signaling in an experimental autoimmune encephalomyelitis (EAE) model, 246 suggesting that HSP70 expression was responsible for the reduced clinical inflammation scores in such models. In this context, inducible nitric oxide synthase (NOS) 247 production, RANTES, and NF‐κB mRNA transcription were downregulated, 248 indicating that HSP70 plays an anti‐inflammatory role in these autoimmune diseases.
5.4.5. HSP90 in autoimmune diseases
HSP90 could exert either immunoregulation or immunostimulation effects upon the assembly of different receptors and signaling pathways. When binding with certain receptors (CD36, CD91, and TLR2/4) on APCs, HSP90 blocks CTL (CD8+ T) expansion and downregulates T‐cell responses; when binding with other receptors (CD25 and P2 × 7R), HSP90 promotes autoimmunity. HSP90 induces IL‐1β production through binding with ATP‐gated P2X cation channel receptor (P2 × 7R) on APCs. HSP90 can also induce macrophage activation through binding with CD25, and propagate T‐cell responses, which is then followed by anti‐HSP90 antibody production. 236 HSP90 autoantibodies were detected in systemic lupus erythematosus and other autoimmune diseases. 227 In mouse models of autoimmune exocrinopathy 249 and anti‐collagen VII autoimmunity, 250 HSP90 could induce the infiltration of inflammatory cells and promote autoimmunity. But in other mouse models of autoimmune diseases, sucg as EAE, T1D, and skin diseases, immunization with HSP90 could reduce autoimmunity. 251 , 252 , 253 In a rat model of autoimmune arthritis, HSP90 reduced arthritis symptoms and induced immune tolerogenicity. 234
Taken altogether, immune responses could be finely tuned by the functions of different HSPs, which work like a double‐edged sword. Different HSPs, in cooperation with different receptors (MHC‐II, TLR, etc.), play different immunomodulatory functions in autoimmune diseases, which could be cell‐, tissue‐, organism‐, disease‐, or HSP‐specific (Figure 4).
FIGURE 4.
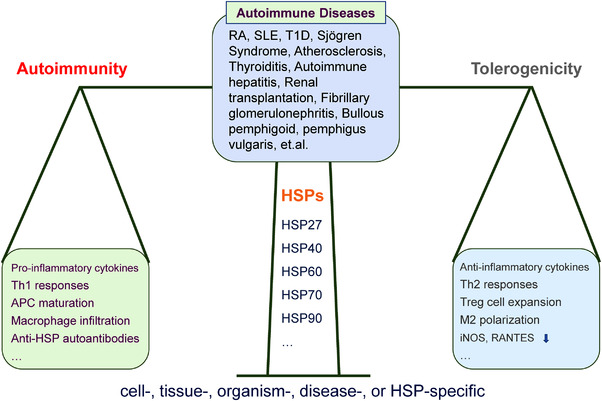
Immunomodulatory actions of HSPs in autoimmune diseases
6. HSPSs AS POTENTIAL THERAPEUTIC TARGETS
Studies of the roles of HSPs in the cause and progression of other severe diseases, such as neurodegeneration, CVD, autoimmune disease, and infection, have made tremendous efforts aiming to identify novel therapies directly targeting HSPs. 254 Therefore, HSPs, such as HSP90 and HSP70, have aroused great interest as potential therapeutic targets 255 , 256 (Table 4).
TABLE 4.
HSPs as potential therapeutic targets in different types of diseases
| Diseases | Therapeutic targets | Clients or mechanisms | References |
|---|---|---|---|
| Cancers | Oncogenic HSP90 |
Cell cycle regulation, immune responses Modulate aberrant protein: p53, HER2/ERBB2, AKT, BCR‐ABL, RAF1, and so on |
257 , 258 |
| HSP70 |
Affect variety apoptosis‐inducing pathways, DISC Tumor suppression pathways, lysosomal membranes, and so on |
259 | |
| Exosomal HSP60 | Increased in patient tumor‐tissue samples or EXs from blood | 260 , 261 , 262 , 263 | |
| HSPgp96 | Multiple clients: HER2, integrins, TLRs, LRP6, and IGF | 264 , 265 | |
| HSF1 | Supporting of cell migration, invasion, proliferation, and cancer cell metabolism | 266 , 267 , 268 | |
| HSP27 | Increased or low expression levels in different cancers or during chemotherapy | 269 , 270 , 271 , 272 | |
| HSP40 | Upregulated in different cancers and serum of cancer patients | 273 , 274 , 275 , 276 | |
| Neurodegeneration diseases | HSP90 | p‐tau, p35, LRRK2, and so on | 277 , 278 , 279 , 280 |
| HSP70 | α‐syn, Reducing loss of neurons | 281 , 282 | |
| HSP27 |
Antioxidant activity Inhibiting cell death pathways Modulating tau dynamics Amyloid β (Aβ) peptides, α‐syn aggregation |
283 , 284 , 285 , 286 , 287 | |
| Cardiovascular diseases | HSP22 | NOS, protects mitochondrial function, Rho GTPase pathway, glycogen synthesis | 288 , 289 , 290 , 291 |
| HSP27 | Angiotensin II, Nuclear factor‐B pathway, Cytokines | 292 , 293 , 294 , 295 , 296 , 297 , 298 , 299 | |
| HSP60 | Mechanism behind remains unclear | 300 , 301 , 302 , 303 , 304 | |
| HSP70 | NO generation, Ca2+ channel, ATP‐sensitive potassium channels | 305 , 306 , 307 | |
| Autoimmune diseases | HSP60 |
Stimulate macrophages, Reacted with various infectious microorganisms |
308 , 309 , 310 , 311 |
| Inflammation | HSP90, HSP70 | Production of proinflammatory cytokines, antigen presentation, and so on | 312 , 313 , 314 , 315 |
| Infection diseases | HSP90 | antifungal in morphogenesis | 316 |
| HSP70 | HIV in CD4+ T cells | 317 , 318 | |
| HSPgp96 | Hepatitis B virus | 319 | |
| HSP40 | Stimulates Th1 and Th17 against Streptococcus pneumoniae | 320 |
6.1. HSPs as targets in cancers
Since HSPs help proteins folding and function in cells, they are present at high levels in cancer cells where defective and mutant proteins are abundant. 321 Blocking the activity HSP90 is already being explored in the treatment of cancers. 322 Other HSPs, including HSP70 and gp96, are also being well studied in vaccines to treat cancers. 323 , 324
Among all the HSPs, HSP90 acquired the most interest and is the best understood during the drug discovery process. 325 HSP90 is a ubiquitously expressed gene that is one of the most abundant intracellular proteins in mammalian cells. HSP90 plays a crucial role in the conformational maturation, protein stability, and function of oncogenic signaling, such as mutated p53, AKT, HER2/ERBB2, BCR‐ABL, and RAF1, as well as many other members that are important in cell cycle progression and immune responses, and it has been recognized as a potential therapeutic target for a number of diseases correlated with aberrant protein signaling, especially in cancers. 257 , 258 The interest in anticancer ability of HSP90 was first discovered during a compound screening for v‐Src transformed cells, in which the antibiotic geldanamycin (GM) emerged as a hit. 326 The potential therapeutic benefit of the selective and potent anticancer activity of GM inspired researchers to identify its target. Later, GM was verified as an HSP90 inhibitor that targets the N‐terminal nucleotide binding pocket by different groups. 255 , 327 Subsequently, the mechanistic study uncovered the HSP90 substrate preference of mutant p53 over wild‐type p53, which explained the selectivity of GM and its derivative 17‐AAG through targeting a ubiquitous protein. Researchers further showed that, compared to normal cells, the HSP90 multichaperone complexes derived from tumor cells exhibited increased ATPase activity and showed higher affinity for HSP90 inhibitors, which provided a reasonable explanation for the selectivity observed with HSP90 inhibitors. 328
However, cancer cells only harbor about 20–30% HSP90 of total, consisting of stressed HSP90 chaperones that are involved in oncogenic partners (e.g., cochaperones) to maintain the malignant phenotype, thereby also named as oncogenic HSP90. Inhibitors have varying ability to recognize the oncogenic HSP90 fraction. 329 Moreover, studies have shown that posttranslational modifications (PTMs) may influence HSP90 chaperone activity and their therapeutic benefit in several ways, including affecting the affinity and hydrolysis of ATP, as well as the association with cochaperones and client proteins. The extents and types of PTMs in normal versus aberrant cells are also different and can impart drug selectivity towards cancer cells, providing a therapeutic rationale for HSP90 as anticancer targets. 330 The development of HSP90 inhibitors is by far the most advanced, and there have been over 20 inhibitors entering into clinical evaluation for the treatment of cancers, suggesting the great therapeutic advantage of HSP90. 325 , 331
HSP70 is one of the cochaperones of HSP90, and it also cooperates with other chaperone systems including HSP60 and small HSPs. Together, they compose a dynamic and functionally versatile network in a broad range of cellular housekeeping activities and stress‐related responses. It is known that some members of the HSP70 families are abundantly present in cancer cells and provide malignant cells with selective advantages by diminishing apoptosis signaling and promoting metastasis that is associated with poor prognosis, thus making them potential drug targets. 256 , 332 For instance, genetic studies strongly suggest that HSP70 plays critical roles in tumor progression and metastasis, which may involve the downregulation of tumor suppression pathways. 333 This direct involvement of HSP70 in cancer tightly associates tumor survival and growth with the HSP70 expression. HSP70s not only inhibit various apoptosis‐inducing pathways, but also block death‐inducing signaling complex (DISC) formation. 334 , 335 Moreover, studies have indicated that HSP70 stabilizes lysosomal membranes, allowing tumor cells to escape from cell death. 336 Further research revealed that inactivation of HSP70 reduced tumor invasiveness and metastatic potential of different cancer cell lines (e.g., breast cancer). 259 These explorations may give new clues for drug development in order to negatively modulate HSP70 activity for cancer treatment.
Blachere's group showed that HSP70‐specific peptide‐complex induced antigen‐specific CD8+ T cell response, suggesting that the immunogenic feature of HSP70 comes from its ability to bind antigenic peptides derived from tumors. 337 However, HSP70 or the 14‐mer HSP70 peptide (TKDNNLLGRFELSG, TKD) alone is not sufficient to stimulate NK cells, and the presence of IL‐2 or IL‐15 is also required. 338 Importantly, the types and dosage of HSP70s can also affect immune responses. The extracellular HSP70 (eHSP70) activates T regulatory cells (Treg) and interacts with APC, suggesting that HSP70s trigger both innate and adaptive immune responses. 339 In addition, HSP70‐positive exosomes were shown to activate myeloid‐derived suppressor cells (MDSCs), leading to IL‐6 production. 340 These experiments opened new perspectives for the utilizing of HSP70 as an adjuvant for tumor immunotherapy. HSP70 functions in different states through its catalytic cycle, suggesting that it can be multifunctional in malignant cells. In cancer patients, tumor cells release large amounts of HSP70 into the extracellular microenvironment, which can result in different outcomes for patient survival. 341 Given its clinical significance, many small‐molecule inhibitors targeting HSP70 were developed. 342 , 343 , 344 In addition, several HSP70‐based immunotherapy approaches have been evaluated in clinical trials. 345
HSP60 or HSPD1, located on the surface of exosomes secreted by tumor cells but not normal cells, was reported as one of the key players during the progress of cancer. Studies demonstrated that exosomal HSP60 has high potential for clinical use, such as a potential clinical biomarker for diagnosing, monitoring disease progression, and assessing prognosis of a variety of cancers. 260 , 261 , 262 Research showed that HSP60 is increased in patient tumor–tissue samples or EXs obtained from the blood of patients, and some groups are evaluating the application of potential HSP60 inhibitors for treatment of certain cancers, 262 induced a decrease in the HSP60 levels or its PTMs leading to cell death. 263 These findings suggest HSP60 has potential clinical applications of both diagnosis and therapy for cancers.
HSPgp96, or GRP94, is one of the HSP90 family members that promotes survival signaling of cancer cells through its multiple client proteins, including HER2, integrins, TLRs, LRP6, and IGF. 264 , 265 Recently, studies showed that HSP gp96 expression positive (>90%) was an independent poor prognostic predictor in gallbladder cancers. 346 Gp96 forms a complex with tumor antigens through acting as a molecular chaperone before the APCs incorporate the antigen, which is then incorporated into dendritic cells via gp96 receptor. 347 Some studies documented that gp96 purified from tumors was able to initiate efficient antitumor responses and protective immunity in both animal models and clinical patients, suggesting that gp96 worked in both preventive and therapeutic protocols. 264 , 323 , 348 Therefore, gp96 plays a critical role in antitumor immunity and is now being studied in vaccine applications to treat cancers.
Other HSPs also participate in cancer development in various ways. HSF1 is a chaperone that associates with both HSP70 and HSP90, and it has been shown to play a significant role in different cancers based on its functions in supporting cell migration, invasion, proliferation, and cancer cell metabolism. 266 Studies in a broad spectrum of cancers showed that active HSF1 is located in the cell nucleus and is associated with poor prognosis. 267 Moreover, genetic knockdown studies of HSF1 have validated the targeted effects in cancers, suggesting HSF1 as a potential therapeutic target. 268 In addition, HSF1 was investigated as a biomarker for patient outcomes in various cancers. 266
Small chaperone HSP27 acts as an ATP‐independent chaperone with potent antiapoptotic property and is involved in cell growth, differentiation, migration, and tumor progression. 269 However, the protective or counter‐protective capabilities of HSP27 in different diseases make it a controversial drug target. 270 Studies showed an aberrant high level of HSP27 in various types of cancers, such as prostate, brain, ovarian, and breast cancers. Recently, research on HSP27 is being conducted to elucidate its possible contributions in invasion and metastasis cascade that affect the overall patient survival, and the results indicated a possible association of HSP27 levels with poor prognosis. 269 , 270 , 332 There are also studies showing that HSP27 levels in serum and in TME were significantly higher in both PC and breast cancer patients, whereas low level of HSP27 was associated with nonresponsiveness to chemotherapy treatment, suggesting that HSP27 could be used as a biomarker for the diagnosis of certain human cancer. 271 , 272 In terms of drug development, there are now some preliminary trials targeting HSP27 for cancer therapy, primarily by downregulating HSP27. 349 However, unlike other HSPs, HSP27 does not bind ATP, making HSP27 difficult to target with small compounds.
HSP40 represents a large and poorly studied family of cochaperones. It was reported that the HSP40 family regulates HSP70 function and usually acts together with HSP90 and HSP70 to participate in different cell survival signal pathways. 273 , 274 Current studies showed that HSP40 upregulated in both brain tumor and lung cancer, and the levels of HSP40 in the serum of cancer patients can be used for tumor diagnosis. However, a study also uncovered a controversial role of HSP40 in human cancers that confers potential resistance to cytotoxic management of 5‐fluorouracil (5‐FU) and carboplatin. 275 , 276 Furthermore, knockdown or inhibition of HSP27 and HSP40 decreases survival of drugs resistant cells. 275 Further understanding of the role of HSP40 family members in cancer may lead to the discovery of new targets for possible cancer treatment.
6.2. HSPs as targets in neurodegeneration diseases
HSPs have recently become a focus of research in neurodegenerative diseases (such as AD, Parkinson's disease, polyglutamine (polyQ) disease, Amyotrophic lateral sclerosis, Huntington's, and many others) because the pathogenesis of these diseases is highlighted by the intracellular proteins aberrant folding and inclusion body formation. 282 HSPs serve as protein‐folding machinery and work together with UPS to assist in removing aberrant proteins, exert antiapoptotic effects, and maintain the dynamic homeostasis of dopaminergic neurons against stress conditions such as oxidative damage. Dysfunction of HSPs, including HSP90, HSP70, and HSP27, may contribute to the pathogenesis of Parkinson's disease as a study showed, suggesting that HSPs may come to be potential therapeutic targets for neurodegenerative disorders such as PD. 350
Overexpression of HSP90 has been reported to reduce the inclusion and accumulation of pathogenic proteins, and to improve phenotypes in neuronal cells and mouse models. 351 Studies have shown that HSP90/P23/Pin1 complex promotes the dephosphorylation and refolding of tau. 277 However, when the refolding process is not complete, tau is transferred to the HSP70/CHIP complex and further ubiquitination system mediates their degradation. 278 HSP90 inhibitors also exert treatment effects through selective mediate proteasomal degradation of their client proteins. In neurodegenerative diseases, HSP90 inhibitors significantly not only reduce the total amount of phosphorylated tau through inducing p35 degradation, but also reduce aggregated forms of tau. 277 , 279 Mutant leucine‐rich repeat kinase 2 (LRRK2) is another client protein of Hsp90, and HSP90 inhibitors rescued the axon growth retardation caused by the overexpression of G2019S mutant LRRK2 in neurons. 280 These studies indicated HSP90 as a therapeutic target of neurodegeneration diseases. Elucidation of its pathophysiological functions using animal models has led to the development of Hsp90 inhibitors and HSP inducers (e.g., Arimoclomol), which suppress the pathogenic process of neuronal degeneration and show clinical effects. 352
The elevation of HSP70 levels has a neuroprotective effect in some animal neurodegeneration models as indicated in several studies. 353 , 354 Recently, studies showed that during the exercise, the released extracellular HSPs 70 (eHSP70) can be internalized by the motoneurons, and then act as intracellular chaperons, protecting cells against protein denaturation, whereas reduced expression of inducible HSP 70 (iHSP70) is associated with neurodegenerative diseases, stressing the importance of understanding the physiological function of extracellular HSP70 in the treatment of neurodegenerative and other neuronal diseases. 355 Moreover, many studies shown that HSP70 may solubilize α‐syn and promote the degradation of its insoluble forms via chaperone‐mediated autophagy and the proteasome pathway in PD. 281 , 282 In addition, other studies suggested that HSP70 plays a role in neuroprotective in diseases that affect CNS and showed promising outcome in reducing the loss of neuron cells. 356 Therefore, future strategies for developing treatments for neurodegenerative diseases through targeting HSP70 can be proposed. Researchers have already showed that the pharmacological HSP70 activators exhibited beneficial effects in neuronal cells during neurodegenerative‐inducing diseases models by intranasal administration of full‐length recombinant human HSP70 protein, which improved the survival of injured neurons by inducing the production of endogenous HSP70. 357 , 358 However, several studies also showed that in the brains of AD patients, neuroprotective functions of molecules such as HSP70 partners are modest. It is clear that the expression and functionality of the inducible forms of HSP70, as well as other important HSPs, are reduced in aged tissues, which needs to be sorted out to ensure that these agents provide therapeutic benefit in neurodegeneration diseases in the future. 283
The expression level of HSP27 in neurons is low but can be increased by proteotoxic stress. In neurodegenerative diseases, elevated levels of HSP27 in glia and neurons correlate with pathogenic deposition of aberrant proteins and showed protective effects in neuronal cells. 283 HSP27 acts as a mediator in the survival response to central nervous system (CNS) injury with its antioxidant activity and ability to inhibit cell death pathways. One study demonstrated the role of HSP27 in AD through modulating tau dynamics and assisting in tau clearance from the brain. 284 Other studies uncovered the role of HSP27 in regulating Amyloid β (Aβ) peptides that is associated with senile plaques in AD brain tissue in in vitro study, and in reducing α‐syn aggregation in Lewy Bodies. 285 , 286 Moreover, several studies in animal models revealed the potential involvement of HSP27 in peripheral nerve injuries, which may yield benefit to selective neuronal loss in AD and PD. For example, one study reported that HSP27 increased axonal growth after peripheral nerve damage and initiated motor regenerative response. 287 These studies suggest that HSP27 could be a promising new therapeutic target for some neurodegenerative diseases. Therefore, it is feasible to develop agents that can stimulate HSP27 function as a potential therapeutic approach, providing insights for innovative treatments for neurological disorders.
6.3. HSPs as targets in cardiovascular diseases
The roles of HSPs in heart function have been extensively studied and recent studies highlighted their therapeutic effects in CVDs. There are numerous studies indicating that HSPs (e.g., HSP22, HSP27, HSP60, and HSP70) play critical roles in protecting cardiomyocytes against stress situations such as hypoxia, ischemia, and infection, suggesting their therapeutic potential for CVDs. 359 , 360 , 361
HSP22 adopts complex mechanisms in cardiomyocyte protection, in that it may activate a number of cell survival pathways, resist reversible damages, and have an antiapoptotic effect. HSP22 expression and phosphorylation increased during the compensatory period of heart failure after myocardial infarction and protect mitochondrial function. At the same time, by regulating proteostasis, HSP22 maintains mitochondrial function and integrity. 288 In vitro studies revealed that USP22 expression elevated when cardiomyocytes were exposed to hypoxia, thus promoting the expression of NOS in the cytoplasm and mitochondria. More studies showed that HSP22 protects mitochondrial function, inhibits mitochondrial apoptosis pathway, resists oxidative stress, and promotes myocardial cell survival and energy metabolism in animal myocardial infarction models, thereby improving myocardial ischemia and heart failure symptoms after myocardial infarction. 359 , 360 In addition, USP22 inhibits the Rho GTPase pathway, which is activated during the onset and progression of atherosclerosis, thus lowers the production of actin stress fibers and inhibits the lowering of peak calcium transients, which stabilizes cardiomyocyte structure and safeguard the atrium's electrophysiology and contractile function. 289 , 290 Furthermore, HSP22 promotes glycogen synthesis and provides the necessary resources for cardiomyocytes under stress. 291 Under normal conditions without stress, the heart structure and function in HSP22 knockout mice are unaffected. 360 , 362 , 363 Together, these findings imply that HSP22 may be a therapeutic target for reversing myocardial remodeling and may play a protective part in the progression of heart failure.
HSP27, a member of the small molecule HSP family, is found in abundance in a variety of malignancies and normal tissues, particularly cardiac tissue. The physiological functions of HSP27 linked to CVD are protecting cells from free radicals, heat, ischemia, and harmful chemicals. 361 HSP27 translocation from cytoplasm to myofibrils and hyperphosphorylated HSP27 in rat heart under ischemia were confirmed by immune‐hybridization analysis, suggesting a role of HSP27 in myocardial ischemia prevention. Studies showed that patients with atherosclerotic plaques have reduced or no HSP27 production compared to healthy people, according to studies. In patients, HSP27 expression and plasma levels were found to be considerably higher in the apparently normal vascular zone close to the plaque than in the plaque core region. 292 Other studies showed the elevated expression of HSP27 in patients with dilated cardiomyopathy and heart failure caused by ischemic cardiomyopathy. HSP27 reduced the extent of myocardial infarction inducing by closure of the left anterior descending coronary artery, therefore enhanced cardiac function in the Langendorff perfusion model of isolated mouse hearts, implying that HSP27 may be involved in cardiomyocyte protection during persistent myocardial ischemia. 293 , 294 , 295 , 296 Angiotensin II (Ang II) has pro‐inflammatory effects in thrombospondin vascular smooth muscle cells (VSMCs) by activating the nuclear factor B pathway, which is regulated by HSP27 and is dependent on the phosphorylation of p65. Extracellular HSP27 also works as a signaling molecule in macrophages, activating the nuclear factor‐B pathway. In addition, by acetylating low‐density lipoprotein, HSP27 inhibits the release of the proinflammatory cytokine IL‐1 from macrophages and enhances the secretion of IL‐10. These findings imply that HSP27 is involved in arterial protection and prevention of arterial wall inflammation. 297 , 298 , 299 Furthermore, acute ischemia can cause increased expression of myocardial HSP27. Under heat stress, the ability of aging rat hearts to synthesize HSP27 was dramatically reduced, suggesting that HSP27 synthesis may be a key element for the aging heart to adapt to stress. 364 , 365 Therefore, HSP27 is considered to be a potential therapeutic target for CVDs.
HSP60 is mostly found in the mitochondria, where it aids in the normal transport, folding, and assembly of cellular polypeptides or proteins. Overexpressed HSP60 can be recognized as a self‐antigen by the immune system upon stress to boost the body's immunological response, or plays a role in immune signal transduction as a signal molecule. Some groups showed that HSP60 expression is strongly correlated with the degree of atherosclerotic lesions and thickness of the intima, suggesting that HSP60 may play a role in the development of atherosclerosis. Other studies showed that during infection and inflammation, the pro‐atherosclerotic effects induce HSP60 expression, thus causing autoimmune responses and leading to vasculitis and atherosclerosis. 300 , 301 However, the mechanism behind remains unclear. 302 Moreover, HSP60 antibody levels in serum have been linked to the occurrence of coronary atherosclerotic heart disease, suggesting a correlation with the prognosis of severe atherosclerosis. 303 In addition, some studies in animal models revealed that HSP60 may play a role in dendritic cell antigen presentation and the production of costimulatory molecules in secondary atherosclerosis. 304
Recently, expression of myocardial HSP70 was detected in the myocardial infarction area in the rat heat shock model, thus promoting the recovery of myocardial contractile. This suggests that HSP70 has effects in myocardial protection and acts as a biomarker of stress or injury for cardiac cells. 366 The highly expressed HSP70 in rat cardiac tissue reduces NO generation by regulating the activation of inducible NOS, which can increase myocardial cell tolerance to ischemia, suggesting that HSP70 has high protective effects in both ischemia and reperfusion stages of myocardial ischemia. 367 Moreover, a study in 80 patients having cardiac surgery showed that HSP70 expression in cardiomyocytes was considerably elevated in patients without atrial fibrillation, imply that inducing HSP70 expression before surgery could be an effective way to prevent postsurgery atrial fibrillation and provide a novel strategy to myocardial protection. 368 , 369 Another group found that HSP70 can increase myocardial contractility and exert myocardial protection through enhancing the internal environment of ischemic myocardium Ca2+ channels and reducing cardiac Ca2+ overload. 305 There are also studies reporting the HSP70's cardioprotective function in controlling of ATP‐sensitive potassium channels. And HSP70 protects the heart against emergency‐induced harm through transporting between cytoplasm to nucleus, resulting in aberrant electrical activity of the heart in an injured state. 306 , 307 In addition, several studies investigated the pathogen load, HSP70 antibody, and serum HSP70 in patients with coronary heart disease, and demonstrated that decrease expression of HSP70 is a risk factor for coronary heart disease, whereas increasing HSP70 levels may help to prevent atherosclerosis, suggesting that HSP70 may be a therapeutic target for atherosclerosis. 370 , 371 Similarly, HSP70 antibody and HSP70 expression levels can be utilized as biomarkers for diagnosis of acute coronary syndrome. 372 , 373 , 374
6.4. HSPs as targets in autoimmune diseases, inflammation, and infection diseases
HSPs are endogenous adjuvants that induce strong tumor‐specific and pathogen‐specific immunity. Recent studies showed that some HSPs are secreted extracellularly during stress responses, prompting the immune system to respond to adverse cellular conditions. HSPs are associated with both pro‐ and anti‐inflammatory responses, and their effects on immune cells depend on the concentration of the respective HSPs secreted from cancer cells. 375 , 376
The correlation between HSPs and autoimmunity is complex although vaccination with HSPs could protect animals from autoimmune diseases. The high homology between human and microbial HSPs could cause autoimmune disorders through immune cross‐reactivity. Studies showed that HSPs could be involved in many autoimmune diseases, including Behcet's disease (BD), type 1 diabetes mellitus (T1DM), arthritis, and systemic lupus erythematosus. For example, human HSP60 is implicated in pro‐inflammation by stimulating macrophages to secrete IL‐6; TNF‐α, IL‐12, and IL‐15 in Type 2 diabetes. 308 HSP60 was also involved in the development of autoimmune encephalomyelitis in rats, but the mechanism is not clear. 309 Several studies discovered that antibodies reacting with HSP60 from various infectious agents (e.g., microorganisms) were found in patients suffering from spondylarthrosis, suggesting a role of HSP60 in autoimmune diseases involving infectious agents. 310 , 311 Therefore, some studies have reported the intervention strategies targeting HSP60 function in various human inflammatory diseases. For instance, the regulatory activities of HSP60 (the p277 peptide from human HSP60) were used to treat human type 1 diabetes (T1D) patients. 377
The molecular chaperones that play roles in signal transduction processes and immune regulation in general have also been reported. 378 For example, several HSPs, such as HSP90, HSP70, HSP60, HSP40, and HSP gp96, were shown to be involved in the production of pro‐inflammatory cytokines. 379 Studies also showed that some of the cytokines are secreted in response to the presence of HSPs, including IL‐1, IL‐6, IL‐12, TNF‐α, and anti‐inflammatory cytokines such as IL‐10. 379 Furthermore, some HSPs induce the release of chemokines from immune cells, and even modulate the maturation of DC cells. 378
Studies also indicate that HSPs facilitate the folding of receptors on immune cells, thus activating the innate immune response and eventually the adaptive immune system. For example, HSP90 serves as a signal to alert tumor‐immune response and is involved in antigen presentation. 312 The recognition of HSP70 by immune cells initiates signal transduction, resulting in the subsequent release of cytokines. 313 HSP70 was also shown to promote the production of anti‐inflammatory cytokines in chronic inflammatory diseases. 314 Moreover, HSP90 inhibition could lead to elevated expression of interferon response genes during tumor immunotherapy. 315 Together, these studies suggest that some HSPs could be potential therapeutic targets for both human autoimmune diseases and inflammations.
Microbial infections, such as pathogenic bacteria, viruses, parasites, and fungi, could put severe stress to cells, and the role of HSPs in infection is another active research area. 380 Organisms such as fungi and virus may express molecular chaperons such as HSPs to perform essential biological functions. 381 HSP90 was indicated as a potential antifungal target according to its role in morphogenesis. 316 Elevated HSP70 expression was observed in the lymphocytes of human immunodeficiency virus (HIV)‐positive patients. 317 Microbial HSP70 and the peptide epitope (aa 407–426) was capable of efficiently inhibiting type‐1 HIV infection of human CD4+ T cells. 318 One study discovered a significant correlation between the level of HSP gp96 and progression of diseases caused by hepatitis B virus. 319 Moreover, HSP40 was shown to stimulate Th1 and Th17 immune responses against Streptococcus pneumoniae infection in mice. 320 Therefore, HSPs inhibitors or vaccines were suggested as therapeutic agents for different virus infection diseases (e.g., HSP90 inhibitors for picornavirus infection, HSP70 vaccines for HSV and HIV‐1). 382 , 383 , 384 Together, these studies demonstrated that HSPs could be valuable therapeutic targets for infection diseases.
6.5. HSPs as disease biomarkers
In the development of targeted therapies and drugs, it is critical to select patients whose diseases depend on these targets prior to treatment so that they can benefit the most from the treatment. Hence, there is a pressing need to identify relevant biomarkers and develop diagnostic tools capable of identifying appropriate patients prior to clinical trials and giving the greatest chance for successful outcome. As mentioned above, some HSPs expression is higher in disease states. For example, HSP90, HSP70, HSP60, HSP27, and HSF1 have been investigated as potential biomarkers for patient outcomes in multiple cancers, including lung cancer, breast cancer, CRC, pancreatic carcinoma, hepatocellular carcinoma, ovarian cancer, PC, and leukemia. 112 , 266 , 332 , 335 , 385 The overexpression and crucial roles of HSPs in various cancers make them tumor markers for targeted drug delivery. Studies have shown that anti‐HSP therapy exhibits significant antitumor activity against human prostate and breast cancers in different clinical trials. For example, as a relevant biomarker for tumor metastasis, the synthetic heat shock protein HSP78 (GRP78)‐binding peptides conjugated with programmed cell death sequence can impair both prostate and breast cancer cell growth in different models. 386 The identified GRP78‐specific peptide, including WDLAWMFRLPVG and WIFPWIQL, were applied for targeted delivery of cytotoxic agents to tumor cells that overexpressed GRP78, such as PCs. 386 Moreover, the nanocarrier‐conjugated scFvs, such as quantum dot conjugated GRP78 scFv, exhibit biological antitumor activity in breast cancer models, suggesting the application as a therapeutic antibody for cancer treatment. 387 In addition, some studies identified the TME‐targeting nanoparticles (NPs), HSP‐NPs, which based on a natural Methanococcus jannaschii small HSPs (Mj‐sHSPs), and showed the ability in improving therapeutic efficacy and decreasing chemotherapy adverse effect in tumor therapy. 388
Besides the function of HSPs as target for drug delivery in cancers, studies also uncovered roles of targeting of HSPs in many other diseases. For example, targeted mitochondrial HSP90 with an inhibitor Gamitrinib‐triphenylphosphonium (G‐TPP) can chemically interfere mitochondrial protein folding, thus relieving Parkinson's disease by inducing mitophagy. 389 Some studies showed that the levels of host HSP65 and HSP71 can be used for the diagnosis of active tuberculosis (TB), whereas HSP16 was more specific for diseases in latency and was used as a diagnostic marker for latent tuberculosis infection (LTBI). 390 In multiple sclerosis (MS), HSP60 was reported as a useful biomarker for progression. 391 In addition, HSP70 antibody and the plasma concentrations of HSP70 were shown as potential screening biomarkers for early diagnosis of heart failure, including acute coronary syndrome. 372 , 373 , 374 , 392 Together, these studies demonstrated the important role of HSPs in disease diagnosis as clinical biomarkers.
However, unlike tyrosine kinase inhibitors targeting overexpressed or mutated kinases that could directly confer drug sensitivity in cells, the context in which these HSP drugs could be used is complicated. Previous studies uncovered that it is not only the levels of HSPs in different related diseases (e.g., cancer), but also the functional status as determined by complex formation as well as PTMs that determine sensitivity to inhibitors. 263 , 330 Therefore, a comprehensive understanding and discovery of other related clinical biomarkers may significantly improve patient treatment.
7. DRUG DISCOVERY FOR HSPSs
Oncogenic HSPs have been recognized as potential therapeutic targets and clinical biomarkers for diagnosis and prognosis for a lot of diseases associated with aberrant protein signaling. Until now, HSP90 and HSP70 have been extensively investigated in terms of drug discovery and they have received the most interest in the past two decades. Up to now, the development of HSP90 inhibitors is the most advanced among HSPs, and more than 20 inhibitors that have undergone clinical trials, some of which are limited by adverse toxicities during several clinical evaluation and none has been approved by the FDA yet. Other HSPs, including HSP70, gp96, and HSP27, are being studied in vaccines or antisense oligonucleotides to treat disease like cancers (Table 5). 323 , 324
TABLE 5.
Current status of HSP inhibitors and application in clinical trials a
| Drugs | Target | Conditions | Clinical trial phase |
|---|---|---|---|
| Geldanamycin | HSP90 |
Advanced solid tumors or non‐Hodgkin's lymphoma (NCT00019708, NCT00003969) |
Phase I terminated or completed |
| Tanespimycin | HSP90 |
Inoperable locoregionally advanced or metastatic thyroid cancer (NCT00118248) |
Phase II completed |
|
With hormone‐resistant prostate cancer (NCT00564928) |
Phase II completed | ||
| Alvespimycin | HSP90 |
Relapsed chronic lymphocytic leukemia, small lymphocytic lymphoma, or B‐cell prolymphocytic leukemia (NCT01126502) |
Phase I terminated or completed |
|
Metastatic or unresectable solid tumors or lymphomas (NCT00088868) |
Phase I completed | ||
|
Her2 positive breast cancer (NCT00803556) |
Phase II terminated | ||
| Retaspimycin | HSP90 | Nonsmall cell lung cancer (NCT01427946), hormone‐resistant prostate cancer (NCT00564928) | Phase I/II completed |
|
Inoperable locoregionally advanced or metastatic thyroid cancer (NCT00113204) |
Phase II completed | ||
| IPI‐493 | HSP90 |
Hormone‐resistant prostate cancer (NCT00564928) |
Phase II completed |
|
Hematologic malignancies (NCT01193491) |
Phase I terminated | ||
| Combination with everolimus in KRAS mutant non‐small cell lung cancer (NCT01427946) | Phase I/II completed | ||
| Trastuzumab pretreated, locally advanced or metastatic HER2 positive breast cancer (NCT00817362) | Phase II terminated | ||
|
Luminespib (NVP‐AUY922) |
HSP90 |
Advanced solid malignancies (NCT01602627) |
Phase I terminated |
| Metastatic pancreatic cancer who are resistant to first line chemotherapy (NCT01484860) | Phase II terminated | ||
| GIST (gastrointestinal stromal tumor) patients (NCT01389583) | Unknown | ||
|
Onalespib AT13387 |
HSP90 | Prostate cancer (NCT01685268) | Phase I/II completed |
| Relapsed/refractory ALK+ anaplastic large cell lymphoma (ALCL), mantle cell lymphoma (MCL), and BCL6+ diffuse large B cell lymphoma (DLBCL) (NCT02572453) | Phase II terminated | ||
|
BRAF V600E mutation present (NCT02097225), advanced triple negative breast cancer (NCT02474173) |
Phase I active, not recruiting | ||
| Ganetespib (STA‐9090) | HSP90 |
Advanced hepatocellular cancer (NCT01665937), metastatic breast cancer (NCT01273896), advanced esophagogastric cancer (NCT01167114), metastatic ocular melanoma (NCT01200238), relapsed or refractory small cell lung cancer (NCT01173523) |
Phase II completed |
| Combined with Crizotinib in ALK positive lung cancers (NCT01579994) | Phase I Completed | ||
|
Zelavespib (PU‐H71) |
HSP90 |
Metastatic breast cancer (NCT03166085), Myelofibrosis (NCT03373877) |
Phase I terminated |
|
Advanced malignancies (NCT01393509) |
Phase I active, not recruiting | ||
| Icapamespib (PU‐AD, PU‐HZ151) |
HSP90 |
Amyotrophic lateral sclerosis (ALS) (NCT04505358) |
Phase II withdrawn |
|
Recurrent malignant glioma (Glio) (NCT04782609) |
Phase I recruiting | ||
| MPC‐3100 | HSP90 |
Safety study in cancer patients who have failed other treatments (NCT00920205) |
Phase I completed |
| Debio0932 (CUDC‐305) | HSP90 |
Nonsmall cell lung cancer, advanced solid tumors or lymphoma (NCT01168752) |
Phase I completed |
|
BIIB021 CNF2024 |
HSP90 |
Advanced solid tumors (NCT01017198) |
Phase I completed |
| BIIB028 | HSP90 | Solid tumors (NCT00725933) | Phase I completed |
| SNX‐2112 | HSP90 | TP53 null cancers (NCT02612285) | Phase II terminated |
|
SNX‐5422 PF‐04929113 |
HSP90 | TP53 null cancers (NCT02612285) | Phase II terminated |
| Human epidermal growth factor receptor 2 (HER2) positive cancers (NCT01848756) | Phase I/II terminated | ||
| Solid tumor cancers and lymphomas, refractory hematological malignancies (NCT01635712), refractory solid tumor malignancies (NCT01892046) | Phase I Completed | ||
|
VER‐82576 NVP‐BEP800 |
HSP90 |
Acute lymphoblastique leukemia (NCT04437420) |
Recruiting |
| Autologous HSP70‐peptide complex in combination with imatinib mesylate | HSP70 |
Chronic myeloid leukemia (NCT00058747) |
Phase II |
| Autologous HSP70‐peptide complex (AG‐858) in combination with Gleevec | HSP70 |
Chronic myeloid leukemia (NCT00058747) |
Phase II |
The data were obtained from https://www.clinicaltrials.gov/.
7.1. HSP90 small molecule inhibitors
7.1.1. Geldanamycin analogues
GM is a benzoquinone‐derivative ansamycin antibiotic. GM was first reported in 1970 from a Streptomyces species, and inhibits the ATPase activity of Hsp90 with a nanomolar IC50, the binding mode was in the ATP‐pocket of HSP90 by mimicking the nucleotide. However, severe hepatotoxicity and poor solubility affected the conduct of clinical trials. 17‐AAG (tanespimycin) and 17‐DMAG (alvespimycin) are both the GM derivatives, which were obtained by replacement of the 17‐methoxy group with the different amine groups. In 1999, tanespimycin entered clinic as the first HSP90 inhibitor, and showed good therapeutic effect in various cancers either in single dose or in combinations, including melanoma, 393 ERBB2‐positive metastatic breast cancer, 394 refractory MM, 395 and so on. Unfortunately, due to the same shortcomings of GM such as the hepatotoxicity and poor solubility, the clinical trials of Alvespimycin had to be terminated. 396 By replacing the methoxy group with N, N‐dimethylpropan‐1‐amino moiety, the solubility, and oral bioavailability of Alvespimycin were improved. Subsequently, Alvespimycin was used in castration‐resistant PC, melanoma, renal cancer, chondrosarcoma, 396 and breast cancer. 397 The reduction of quinone to hydroquinone by reductase NQO1 (NADPH/quinone oxidoreductase I) causes the metabolic instability and hepatotoxicity. 398 By replacing quinone with hydroquinone, Retaspimycin was developed and the solubility can be improved in hydrochloride form, 399 , 400 and it was approved to enter clinic. In addition, IPI‐493 was a dealkylation compound of tanespimycin and was now being used in clinical practice (Figure S1A). 401
7.1.2. Resorcinol‐based HSP90 inhibitors
To overcome the limitations of GM analogs, such as hepatotoxicity and solubility, the resorcinol‐based HSP90 inhibitors were developed (Figure S1B). The resorcinol moiety of radicicol binds deeply of the ATP‐binding pocket of HSP90, and at the same time anchored by H2O‐mediated hydrogen bonds. Luminespib (NVP‐AUY922) was developed based on a previously discovered a resorcinol‐structural compound using structure‐based design approach, 402 and it showed high affinity to Hsp90. Luminespib can induce the proteasomal degradation of oncogenic client proteins 403 and was evaluated in multiple clinical trials. AT13387 was also a resorcinol‐based HSP90 inhibitor, which was optimized based on a potent lead compound using a fragment‐based design approach, 404 and it exhibited high affinity of HSP90 through its binding in the ATPase site of the N‐domain. AT13387 inhibited the growth, migration, and clone formation, and eventually showed significant antitumor effect in numerous in vitro and in vivo cancer models. 405 , 406 Recently, it was discovered that AT1387 can protect the Alveolo–Capillary Barrier and prevent HCl‐induced chronic lung injury and pulmonary fibrosis at nontoxic low doses. 407 Ganetespib (formerly known as STA‐9090) is a potent and selective Hsp90 inhibitor containing a resorcinol ring and a unique Triazolone, 408 and it is currently being evaluated in multiple clinical trials. 408 , 409
7.1.3. Purine‐based HSP90 inhibitors
Many HSP90 inhibitors were developed based on the purine ring of natural ligand ATP (Figure S1C). Substitution at C‐8 of the purine moiety was identified as a critical site for optimization, and a phenyl sulfide and amine in the alkyl chain were introduced at C‐8 and N‐9 of the purine ring. Zelavespib (PU‐H71) is also a purine‐based Hsp90 inhibitor, 410 which can specifically inhibit Hsp90 and chaperone’ function, promote the degradation of oncogenic signaling proteins, and eventually prevent tumor cell proliferation and survival. In 2011, Zelavespib (PU‐H71) was used for the treatment of lymphoma, solid tumors, metastatic solid tumor, myeloproliferative neoplasms (MPNs) in clinical trials, and recently was used in PML‐SYK fusion AML. 411 However, Zelavespib is unable to permeate the blood–brain barrier (BBB), which rendered it unfeasible for the treatment of diseases in CNS. Icapamespib (PU‐AD, PU‐HZ151) is developed based on the structure of Zelavespib (PU‐H71) aiming to overcome the limitation of BBB with the alkyl chain of the N9‐chain amine moiety modified from isopropyl to dimethylpropyl. 412 Zelavespib was used for the treatment of Alzheimer's disease (NCT03935568), PET imaging (NCT03371420), and recurrent GBM (NCT04782609). Based on Zelavespib, MPC‐3100 was later developed as a new clinical candidate with the N‐9 alkyl and the iodo of C‐8 arylsulfide modified to N‐substituted piperidine moiety and bromo, respectively. The potency and pharmacokinetic properties of MPC‐3100 were both improved, 413 and was used for the treatment of cancers (NCT00920205). Debio 0932 (CUDC‐305) was an Hsp90 inhibitor belonging to the purine analogs, the iodo group was changed to N, N‐dimethyl, and the N atom of the purine was removed. Debio 0932 (CUDC‐305) can overcome erlotinib resistance and was used for patients with nonsmall cell lung cancer (NCT01714037) and advanced solid tumors or lymphoma (NCT01168752). 414 In addition, BIIB021 was developed by replacing aryl substituent at N‐9 with a pyridyl motif and adding a chloro at C‐2 position. 415 BIIB021 entered clinical trials and showed partial responses in a Phase II trial of gastrointestinal stromal tumors (GISTs). 416 BIIB028 was further modified based on BIIB021 for improvement in potency, efficacy, and tolerability. In BIIB028, N7 was replaced with a carbon atom, the deeazapurine ring was designed, and a hydroxy alkynes group was substituted at this carbon atom. By adopting a prodrug form using phosphorylation, the solubility of BIIB028 was improved and it can be completely converted into its active metabolite in a short time. 417 BIIB028 was used as an orally available HSP90 inhibitor for the treatment of advanced solid tumors (NCT00725933).
7.1.4. Other structures HSP90 inhibitors
Although the natural product–derived GM analogs HSP90 inhibitors are efficacious, the hepatotoxicity and lack of oral availability limited the dosing frequency. To overcome this, a novel class Hsp90 inhibitor SNX‐2112 was developed (Figure S1D). SNX‐2112 is an ATP‐competitive Hsp90 inhibitor. SNX‐5422 is the prodrug of SNX‐2112 and highly orally bioavailable with potency against various cancers. Three phase 1 clinical studies of SNX‐5422 are currently recruiting including refractory hematologic and solid tumor malignancies. 418 NVP‐BEP800 is a novel orally bioavailable ATP‐competitive Hsp90 inhibitor. NVP‐BEP800 was developed through combination of hit identification strategies and the structure‐based medicinal chemistry program, and a new class of 2‐aminothieno[2,3‐d] pyrimidine Hsp90 inhibitors with oral efficacy in animal cancer models was identified. 419 NVP‐BEP800 showed favorable activity against numerous human tumor cell lines and primary human xenograft models at low concentrations. In clinical evaluation, NVP‐BEP800 was used on two different types of ALL (T‐ and B‐ALL) (NCT04437420). Recently, a novel HSP90 inhibitor (RGRN‐305) was reported for the treatment of plaque psoriasis, 420 but the structure was undisclosed.
7.2. Recombinant peptides or vaccines
7.2.1. Hsp70‐peptide
HSP70 was reported as a stress‐inducible protein with elevated expression in tumor cells but not normal cells. One of well‐developed recombinant peptide in clinical trial is the C‐terminal substrate‐binding domain of HSP70, comprising amino acids 450–461 (aa450–461), termed 14‐mer HSP70 peptide (TKD). TKD stimulated NK cells ex vivo with interleukin‐2 (IL‐2) and showed potential antitumor activity post reinfusion in different animal models or clinical patients. 421 , 422 TKD‐treated NK cells bind to tumor cells through structurally recognizing HSP70, thus mediating tumor cell lysis. Moreover, TKD alone is not sufficient to stimulate NK cells, and it also requires the presence of IL‐2 or IL‐15. 338
Another reported recombinant peptide is chemically identical to or similar to the endogenous HSP70. The autologous vaccination with tumor‐derived HSP70 stimulates the host mice immune system, permits antigen presenting cells (APCs) to present tumor antigens to their cell surface, and directly elicits a tumoricidal cytotoxic T lymphocyte (CTL) response, thereby increasing longevity and survival of tumor‐bearing hosts. 423
7.2.2. Vitespen, HSP gp96‐peptide complex
Vitespen is an autologous cancer vaccine derived from tumor‐specific HSPgp96 and is currently in different clinical trials (phase II and III) for cancers, including melanoma, CRC, lung cancer, glioblastoma, and renal cell carcinoma. HSPgp96 is a key regulator involved in dendritic cell maturation, migration, antigen processing, as well as T cell activation. Therefore, Vitespen acts as an inducer of major histocompatibility class I‐restricted immune responses in a range of tumor types in preclinical models and showed significant clinical responses in patients with early‐stage disease with few side effects. 424 , 425
7.2.3. Apatorsen, HSP27 antisense oligonucleotide OGX‐427
HSP27 has been found to be overexpressed in a variety of human cancers. Apatorsen is a second‐generation antisense oligonucleotide targeting HSP27 mRNA with potential antitumor and chemosensitizing activities. One recent study reported that Apatorsen induced tumor cell apoptosis and enhanced cell cytotoxicity through suppressing HSP27 expression in tumor cells. This potential drug is now in clinical trials for different cancers either alone or in combination treatment, so far it has shown improved outcomes in patients with metastatic NSCLC cancer and platinum‐resistant metastatic urothelial carcinoma. 426 , 427 Another study reported the evaluation of Apatorsen in phase II clinical in combination with prednisone for the treatment of castration‐resistant PCs (NCT01120470).
8. OUTLOOK
Since the discovery of the heat shock puff in 1962, deeper understanding is achieved about the HSPs structures, functions, and how they cooperate with each other, working as chaperones to orchestrate proteostasis and deal with stresses. However, at the same time, there are still unknown questions about the structures and mechanisms of HSPs. First, the structure mechanisms of how the HSPs work as a network in protein folding is not completely understood, mainly due to the transient and dynamic binding structures of HSPs complexed with clients which are difficult to resolve. Recently, David A. Agard group has resolved the high resolution cryoelectron microscopy structures of Hsp90–p23–GR and Hsp90–Hsp70–Hop–GR, which reveals that the basic mechanism of how the client is loaded to HSP90 from HSP70 and how the cochaperons work in protein folding cycle. 96 , 97 Meanwhile, the folding mechanism of large multidomain proteins remains unclear and need better understanding. 428 Second, the number of proteins in the proteome that rely on HSPs for structure stability still needs to be studied. Although some works has clarified the client proteins of HSP90 in proteomic range, 57 systematic proteomic studies for the client proteins of other HSPs are still needed. Recently, some principles for HSPs in the recognition of clients are summarized, which may be useful for discover new potential client proteins of HSPs. 37 , 38 , 57 , 429 As the protein homeostasis is important for cell integrity, survival and metabolism, impairment of chaperone‐assisted protein quality control leads to the onset and development of various diseases.
At the same time, although a lot of studies have proved that HSPs play an important role in the development of diseases, the current drug development for HSPs is not fulfilled. Many HSP90 inhibitors in clinical trials were terminated or delayed due to toxicity or lack of efficacy. 430 It has observed that the use of HSP90 inhibitors could lead to the activation of HSF1, which, in turn, induce the activation of other HSPs to overcome the lack of HSP90. Therefore, development of inhibitors against multiple HSPs or HSF1 may be a strategy to enhance the efficacy of HSP90 inhibitors and overcome the HSF1‐mediated feedback. In the meantime, we also need to pay attention to the safety of these inhibitors in use. Better understanding of how HSPs function in vivo and the collaboration between the HSPs in cancers will be crucial for reducing toxicity to normal cells and a more accurate indication selection.
CONFLICT OF INTERESTS
The authors declare no competing interests.
ETHICS APPROVAL
Not applicable.
CONTRIBUTIONS
C. Hu, J. Yang, Z.P. Qi, H. Wu, F.M. Zou, B.L. Wang, and H.S. Mei conceived the study, collected the literatures, and drafted the manuscript. Corresponding authors, including W.C. Wang, and Q.S. Liu, provided their corrective comments and tips. J.L revised the manuscript. All authors collaborated to write the article. All authors approved this manuscript for publication.
Supporting information
Supporting Information
ACKNOWLEDGMENTS
This work was supported by the National Natural Science Foundation of China (Grant Nos, 81903650, 82104198), the Natural Science Foundation of Anhui Province (Grant Nos, 2008085MH274, 2108085QH377), and the CASHIPS Director's Fund (Grant Nos. YZJJZX202011, YZJJ2021QN38). We are also grateful for the support of the Youth Innovation Promotion Association of CAS support for JY (No. 2022453) and HW (No. 2019437).
Hu C, Yang J, Qi Z, et al. Heat shock proteins: Biological functions, pathological roles, and therapeutic opportunities. MedComm. 2022;3:e161. 10.1002/mco2.161
Contributor Information
Wenchao Wang, Email: wwcbox@hmfl.ac.cn.
Qingsong Liu, Email: qsliu97@hmfl.ac.cn.
DATA AVAILABILITY STATEMENT
The data included in this study are available upon request from the corresponding authors.
REFERENCES
- 1. Richter K, Haslbeck M, Buchner J. The heat shock response: life on the verge of death. Mol Cell. 2010;40(2):253‐266. [DOI] [PubMed] [Google Scholar]
- 2. Ellis RJ. Protein misassembly: macromolecular crowding and molecular chaperones. Adv Exp Med Biol. 2007;594:1‐13. [DOI] [PubMed] [Google Scholar]
- 3. Balch WE, Morimoto RI, Dillin A, Kelly JW. Adapting proteostasis for disease intervention. Science. 2008;319(5865):916‐919. [DOI] [PubMed] [Google Scholar]
- 4. Hoter A, El‐Sabban ME, Naim HY. The HSP90 family: structure, regulation, function, and implications in health and disease. Int J Mol Sci. 2018;19(9):2560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Hartl FU, Bracher A, Hayer‐Hartl M. Molecular chaperones in protein folding and proteostasis. Nature. 2011;475(7356):324‐332. [DOI] [PubMed] [Google Scholar]
- 6. Saibil H. Chaperone machines for protein folding, unfolding and disaggregation. Nat Rev Mol Cell Biol. 2013;14(10):630‐642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Murshid A, Prince TL, Lang B, Calderwood SK. Role of heat shock factors in stress‐induced transcription. Methods Mol Biol. 2018;1709:23‐34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Lindquist S. The heat‐shock response. Annu Rev Biochem. 1986;55:1151‐1191. [DOI] [PubMed] [Google Scholar]
- 9. Ritossa F. A new puffing pattern induced by temperature shock and DNP in drosophila. Experientia. 1962;18(12):571‐573. [Google Scholar]
- 10. Ritossa FM. Experimental activation of specific Loci in polytene chromosomes of Drosophila. Exp Cell Res. 1964;35:601‐607. [DOI] [PubMed] [Google Scholar]
- 11. Lindquist S, Craig EA. The heat‐shock proteins. Annu Rev Genet. 1988;22:631‐677. [DOI] [PubMed] [Google Scholar]
- 12. Moran L, Mirault ME, Arrigo AP, Goldschmidt‐Clermont M, Tissieres A. Heat shock of Drosophila melanogaster induces the synthesis of new messenger RNAs and proteins. Philos Trans R Soc Lond B Biol Sci. 1978;283(997):391‐406. [DOI] [PubMed] [Google Scholar]
- 13. Mirault ME, Goldschmidt‐Clermont M, Moran L, Arrigo AP, Tissieres A. The effect of heat shock on gene expression in Drosophila melanogaster. Cold Spring Harb Symp Quant Biol. 1978;42(Pt 2):819‐827. [DOI] [PubMed] [Google Scholar]
- 14. Georgopoulos CP, Hendrix RW, Casjens SR, Kaiser AD. Host participation in bacteriophage lambda head assembly. J Mol Biol. 1973;76(1):45‐60. [DOI] [PubMed] [Google Scholar]
- 15. Hohn T, Hohn B, Engel A, Wurtz M, Smith PR. Isolation and characterization of the host protein groE involved in bacteriophage lambda assembly. J Mol Biol. 1979;129(3):359‐373. [DOI] [PubMed] [Google Scholar]
- 16. Cheng MY, Hartl FU, Martin J, et al. Mitochondrial heat‐shock protein hsp60 is essential for assembly of proteins imported into yeast mitochondria. Nature. 1989;337(6208):620‐625. [DOI] [PubMed] [Google Scholar]
- 17. Barraclough R, Ellis RJ. Protein synthesis in chloroplasts IX. Assembly of newly‐synthesized large subunits into ribulose bishopshate carboxylase in isolated intact pea chloroplasts. Biochim Biophys Acta (BBA) ‐ Nucl Acids Protein Synth. 1980;608(1):19‐31. [DOI] [PubMed] [Google Scholar]
- 18. Ostermann J, Horwich AL, Neupert W, Hartl FU. Protein folding in mitochondria requires complex formation with hsp60 and ATP hydrolysis. Nature. 1989;341(6238):125‐130. [DOI] [PubMed] [Google Scholar]
- 19. Langer T, Pfeifer G, Martin J, Baumeister W, Hartl FU. Chaperonin‐mediated protein folding: groES binds to one end of the GroEL cylinder, which accommodates the protein substrate within its central cavity. Embo J. 1992;11(13):4757‐4765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Flynn GC, Chappell TG, Rothman JE. Peptide binding and release by proteins implicated as catalysts of protein assembly. Science. 1989;245(4916):385‐390. [DOI] [PubMed] [Google Scholar]
- 21. Beckmann RP, Mizzen LE, Welch WJ. Interaction of Hsp 70 with newly synthesized proteins: implications for protein folding and assembly. Science. 1990;248(4957):850‐854. [DOI] [PubMed] [Google Scholar]
- 22. Liberek K, Marszalek J, Ang D, Georgopoulos C, Zylicz M. Escherichia coli DnaJ and GrpE heat shock proteins jointly stimulate ATPase activity of DnaK. Proc Natl Acad Sci U S A. 1991;88(7):2874‐2878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Langer T, Lu C, Echols H, Flanagan J, Hayer MK, Hartl FU. Successive action of DnaK, DnaJ and GroEL along the pathway of chaperone‐mediated protein folding. Nature. 1992;356(6371):683‐689. [DOI] [PubMed] [Google Scholar]
- 24. Ellis J. Proteins as molecular chaperones. Nature. 1987;328(6129):378‐379. [DOI] [PubMed] [Google Scholar]
- 25. Schuh S, Yonemoto W, Brugge J, et al. A 90,000‐dalton binding protein common to both steroid receptors and the Rous sarcoma virus transforming protein, pp60v‐src. J Biol Chem. 1985;260(26):14292‐14296. [PubMed] [Google Scholar]
- 26. Finkelstein DB, Strausberg S. Identification and expression of a cloned yeast heat shock gene. J Biol Chem. 1983;258(3):1908‐1913. [PubMed] [Google Scholar]
- 27. Bardwell JC, Craig EA. Major heat shock gene of Drosophila and the Escherichia coli heat‐inducible dnaK gene are homologous. Proc Natl Acad Sci U S A. 1984;81(3):848‐852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Seaton BL, Vickery LE. A gene encoding a DnaK/hsp70 homolog in Escherichia coli. Proc Natl Acad Sci U S A. 1994;91(6):2066‐2070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Lelivelt MJ, Kawula TH. Hsc66, an Hsp70 homolog in Escherichia coli, is induced by cold shock but not by heat shock. J Bacteriol. 1995;177(17):4900‐4907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Bardwell JC, Craig EA. Eukaryotic Mr 83,000 heat shock protein has a homologue in Escherichia coli. Proc Natl Acad Sci U S A. 1987;84(15):5177‐5181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Ellis RJ. Protein aggregation: opposing effects of chaperones and crowding. In: Wyttenbach A, O'Connor V, eds. Folding for the Synapse. Springer; 2011:9‐34. [Google Scholar]
- 32. Voos W, Röttgers K. Molecular chaperones as essential mediators of mitochondrial biogenesis. Biochim Biophys Acta. 2002;1592(1):51‐62. [DOI] [PubMed] [Google Scholar]
- 33. Yun CW, Kim HJ, Lim JH, Lee SH. Heat shock proteins: agents of cancer development and therapeutic targets in anti‐cancer therapy. Cells. 2019;9(1):60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Rosenzweig R, Nillegoda NB, Mayer MP, Bukau B. The Hsp70 chaperone network. Nat Rev Mol Cell Biol. 2019;20(11):665‐680. [DOI] [PubMed] [Google Scholar]
- 35. Brocchieri L, Conway de Macario E, Macario AJ. hsp70 genes in the human genome: conservation and differentiation patterns predict a wide array of overlapping and specialized functions. BMC Evol Biol. 2008;8:19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Daugaard M, Rohde M, Jaattela M. The heat shock protein 70 family: highly homologous proteins with overlapping and distinct functions. FEBS Lett. 2007;581(19):3702‐3710. [DOI] [PubMed] [Google Scholar]
- 37. Rudiger S, Germeroth L, Schneider‐Mergener J, Bukau B. Substrate specificity of the DnaK chaperone determined by screening cellulose‐bound peptide libraries. EMBO J. 1997;16(7):1501‐1507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Clerico EM, Tilitsky JM, Meng W, Gierasch LM. How hsp70 molecular machines interact with their substrates to mediate diverse physiological functions. J Mol Biol. 2015;427(7):1575‐1588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Brehmer D, Rüdiger S, Gässler CS, et al. Tuning of chaperone activity of Hsp70 proteins by modulation of nucleotide exchange. Nat Struct Biol. 2001;8(5):427‐432. [DOI] [PubMed] [Google Scholar]
- 40. Kityk R, Kopp J, Mayer MP. Molecular mechanism of J‐domain‐triggered ATP hydrolysis by Hsp70 Chaperones. Mol Cell. 2018;69(2):227‐237 e4. [DOI] [PubMed] [Google Scholar]
- 41. Kityk R, Kopp J, Sinning I, Mayer MP. Structure and dynamics of the ATP‐bound open conformation of Hsp70 chaperones. Mol Cell. 2012;48(6):863‐874. [DOI] [PubMed] [Google Scholar]
- 42. Tzankov S, Wong MJ, Shi K, Nassif C, Young JC. Functional divergence between co‐chaperones of Hsc70. J Biol Chem. 2008;283(40):27100‐27109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Misselwitz B, Staeck O, Rapoport TA. J proteins catalytically activate Hsp70 molecules to trap a wide range of peptide sequences. Mol Cell. 1998;2(5):593‐603. [DOI] [PubMed] [Google Scholar]
- 44. Minoia M, Boncoraglio A, Vinet J, et al. BAG3 induces the sequestration of proteasomal clients into cytoplasmic puncta: implications for a proteasome‐to‐autophagy switch. Autophagy. 2014;10(9):1603‐1621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Gamerdinger M, Hajieva P, Kaya AM, Wolfrum U, Hartl FU, Behl C. Protein quality control during aging involves recruitment of the macroautophagy pathway by BAG3. EMBO J. 2009;28(7):889‐901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Behl C. BAG3 and friends: co‐chaperones in selective autophagy during aging and disease. Autophagy. 2011;7(7):795‐798. [DOI] [PubMed] [Google Scholar]
- 47. Raviol H, Sadlish H, Rodriguez F, Mayer MP, Bukau B. Chaperone network in the yeast cytosol: hsp110 is revealed as an Hsp70 nucleotide exchange factor. EMBO J. 2006;25(11):2510‐2518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Schuermann JP, Jiang J, Cuellar J, et al. Structure of the Hsp110:hsc70 nucleotide exchange machine. Mol Cell. 2008;31(2):232‐243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Zhang H, Amick J, Chakravarti R, et al. A bipartite interaction between Hsp70 and CHIP regulates ubiquitination of chaperoned client proteins. Structure. 2015;23(3):472‐482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Shen Y, Liu J, Wang X, Cheng X, Wang Y, Wu N. Essential role of the first intron in the transcription of hsp90beta gene. FEBS Lett. 1997;413(1):92‐98. [DOI] [PubMed] [Google Scholar]
- 51. Zhang SL, Yu J, Cheng XK, et al. Regulation of human hsp90alpha gene expression. FEBS Lett. 1999;444(1):130‐135. [DOI] [PubMed] [Google Scholar]
- 52. Chen B, Zhong D, Monteiro A. Comparative genomics and evolution of the HSP90 family of genes across all kingdoms of organisms. BMC Genomics. 2006;7:156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Felts SJ, Owen BA, Nguyen P, Trepel J, Donner DB, Toft DO. The hsp90‐related protein TRAP1 is a mitochondrial protein with distinct functional properties. J Biol Chem. 2000;275(5):3305‐3312. [DOI] [PubMed] [Google Scholar]
- 54. Johnson JL. Evolution and function of diverse Hsp90 homologs and cochaperone proteins. Biochim Biophys Acta. 2012;1823(3):607‐613. [DOI] [PubMed] [Google Scholar]
- 55. Ali MM, Roe SM, Vaughan CK, et al. Crystal structure of an Hsp90‐nucleotide‐p23/Sba1 closed chaperone complex. Nature. 2006;440(7087):1013‐1017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Zierer BK, Rubbelke M, Tippel F, et al. Importance of cycle timing for the function of the molecular chaperone Hsp90. Nat Struct Mol Biol. 2016;23(11):1020‐1028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Taipale M, Krykbaeva I, Koeva M, et al. Quantitative analysis of HSP90‐client interactions reveals principles of substrate recognition. Cell. 2012;150(5):987‐1001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Biebl MM, Lopez A, Rehn A, et al. Structural elements in the flexible tail of the co‐chaperone p23 coordinate client binding and progression of the Hsp90 chaperone cycle. Nat Commun. 2021;12(1):828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Yamamoto S, Subedi GP, Hanashima S, et al. ATPase activity and ATP‐dependent conformational change in the co‐chaperone HSP70/HSP90‐organizing protein (HOP). J Biol Chem. 2014;289(14):9880‐9886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Balchin D, Milicic G, Strauss M, Hayer‐Hartl M, Hartl FU. Pathway of actin folding directed by the eukaryotic chaperonin TRiC. Cell. 2018;174(6):1507‐1521 e16. [DOI] [PubMed] [Google Scholar]
- 61. Saibil HR, Fenton WA, Clare DK, Horwich AL. Structure and allostery of the chaperonin GroEL. J Mol Biol. 2013;425(9):1476‐1487. [DOI] [PubMed] [Google Scholar]
- 62. Ishida R, Okamoto T, Motojima F, et al. Physicochemical properties of the mammalian molecular chaperone HSP60. Int J Mol Sci. 2018;19(2):489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Gomez‐Llorente Y, Jebara F, Patra M, et al. Structural basis for active single and double ring complexes in human mitochondrial Hsp60‐Hsp10 chaperonin. Nat Commun. 2020;11(1):1916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Cong Y, Schroder GF, Meyer AS, et al. Symmetry‐free cryo‐EM structures of the chaperonin TRiC along its ATPase‐driven conformational cycle. EMBO J. 2012;31(3):720‐730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Naqvi MM, Avellaneda MJ, Roth A, et al. Protein chain collapse modulation and folding stimulation by GroEL‐ES. Sci Adv. 2022;8(9):eabl6293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Khabirova E, Moloney A, Marciniak SJ, et al. The TRiC/CCT chaperone is implicated in Alzheimer's disease based on patient GWAS and an RNAi screen in Aβ‐expressing Caenorhabditis elegans. PLoS One. 2014;9(7):e102985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Kim YE, Hipp MS, Bracher A, Hayer‐Hartl M, Hartl FU. Molecular chaperone functions in protein folding and proteostasis. Annu Rev Biochem. 2013;82:323‐355. [DOI] [PubMed] [Google Scholar]
- 68. Ghosh JC, Dohi T, Kang BH, Altieri DC. Hsp60 regulation of tumor cell apoptosis. J Biol Chem. 2008;283(8):5188‐5194. [DOI] [PubMed] [Google Scholar]
- 69. Sarangi U, Singh MK, Abhijnya KV, et al. Hsp60 chaperonin acts as barrier to pharmacologically induced oxidative stress mediated apoptosis in tumor cells with differential stress response. Drug Target Insights. 2013;7:35‐51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Kumar S, O'Malley J, Chaudhary AK, et al. Hsp60 and IL‐8 axis promotes apoptosis resistance in cancer. Br J Cancer. 2019;121(11):934‐943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Haslbeck M, Franzmann T, Weinfurtner D, Buchner J. Some like it hot: the structure and function of small heat‐shock proteins. Nat Struct Mol Biol. 2005;12(10):842‐846. [DOI] [PubMed] [Google Scholar]
- 72. Basha E, O'Neill H, Vierling E. Small heat shock proteins and alpha‐crystallins: dynamic proteins with flexible functions. Trends Biochem Sci. 2012;37(3):106‐117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Pfoh R, Lacdao IK, Saridakis V. Deubiquitinases and the new therapeutic opportunities offered to cancer. Endocr Relat Cancer. 2015;22(1):T35‐T54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Hochberg GKA, Shepherd DA, Marklund EG, et al. Structural principles that enable oligomeric small heat‐shock protein paralogs to evolve distinct function. Science. 2018;359(6378):930‐935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Clark AR, Vree Egberts W, Kondrat FDL, et al. Terminal regions confer plasticity to the tetrameric assembly of human HspB2 and HspB3. J Mol Biol. 2018;430(18 Pt B):3297‐3310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. van Montfort RL, Basha E, Friedrich KL, Slingsby C, Vierling E. Crystal structure and assembly of a eukaryotic small heat shock protein. Nat Struct Biol. 2001;8(12):1025‐1030. [DOI] [PubMed] [Google Scholar]
- 77. Easton DP, Kaneko Y, Subjeck JR. The hsp110 and Grp1 70 stress proteins: newly recognized relatives of the Hsp70s. Cell Stress Chaperones. 2000;5(4):276‐290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Kandasamy G, Andreasson C. Hsp70‐Hsp110 chaperones deliver ubiquitin‐dependent and ‐independent substrates to the 26S proteasome for proteolysis in yeast. J Cell Sci. 2018;131(6):jcs210948. [DOI] [PubMed] [Google Scholar]
- 79. Tittelmeier J, Sandhof CA, Ries HM, et al. The HSP110/HSP70 disaggregation system generates spreading‐competent toxic alpha‐synuclein species. EMBO J. 2020;39(13):e103954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Bakthisaran R, Tangirala R, Rao CM. Small heat shock proteins: role in cellular functions and pathology. Biochim Biophys Acta (BBA) ‐ Proteins Proteom. 2015;1854(4):291‐319. [DOI] [PubMed] [Google Scholar]
- 81. Haslbeck M, Weinkauf S, Buchner J. Small heat shock proteins: simplicity meets complexity. J Biol Chem. 2019;294(6):2121‐2132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Cappello F, Conway de Macario E, Marasa L, Zummo G, Macario AJ. Hsp60 expression, new locations, functions and perspectives for cancer diagnosis and therapy. Cancer Biol Ther. 2008;7(6):801‐809. [DOI] [PubMed] [Google Scholar]
- 83. Weiss C, Jebara F, Nisemblat S, Azem A. Dynamic complexes in the Chaperonin‐Mediated protein folding cycle. Review. 2016;3:80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Fernandez‐Fernandez MR, Gragera M, Ochoa‐Ibarrola L, Quintana‐Gallardo L, Valpuesta JM. Hsp70 ‐ a master regulator in protein degradation. FEBS Lett. 2017;591(17):2648‐2660. [DOI] [PubMed] [Google Scholar]
- 85. Prodromou C. Mechanisms of Hsp90 regulation. Biochem J. 2016;473(16):2439‐2452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Brockwell DJ, Radford SE. Intermediates: ubiquitous species on folding energy landscapes?. Curr Opin Struct Biol. 2007;17(1):30‐37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Zou Z, Cao L, Zhou P, Su Y, Sun Y, Li W. Hyper‐acidic protein fusion partners improve solubility and assist correct folding of recombinant proteins expressed in Escherichia coli. J Biotechnol. 2008;135(4):333‐339. [DOI] [PubMed] [Google Scholar]
- 88. Schmidpeter PA, Schmid FX. Prolyl isomerization and its catalysis in protein folding and protein function. J Mol Biol. 2015;427(7):1609‐1631. [DOI] [PubMed] [Google Scholar]
- 89. Dinner AR, Sali A, Smith LJ, Dobson CM, Karplus M. Understanding protein folding via free‐energy surfaces from theory and experiment. Trends Biochem Sci. 2000;25(7):331‐339. [DOI] [PubMed] [Google Scholar]
- 90. Reikvam H, Nepstad I, Sulen A, Gjertsen BT, Hatfield KJ, Bruserud Ø. Increased antileukemic effects in human acute myeloid leukemia by combining HSP70 and HSP90 inhibitors. Expert Opin Investig Drugs. 2013;22(5):551‐563. [DOI] [PubMed] [Google Scholar]
- 91. Nilsson OB, Muller‐Lucks A, Kramer G, Bukau B, von Heijne G. Trigger factor reduces the force exerted on the nascent chain by a cotranslationally folding protein. J Mol Biol. 2016;428(6):1356‐1364. [DOI] [PubMed] [Google Scholar]
- 92. Kirstein‐Miles J, Morimoto RI. Ribosome‐associated chaperones act as proteostasis sentinels. Cell Cycle. 2013;12(15):2335‐2336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93. Preissler S, Deuerling E. Ribosome‐associated chaperones as key players in proteostasis. Trends Biochem Sci. 2012;37(7):274‐283. [DOI] [PubMed] [Google Scholar]
- 94. Baindur‐Hudson S, Edkins AL, Blatch GL. Hsp70/Hsp90 organising protein (hop): beyond interactions with chaperones and prion proteins. Subcell Biochem. 2015;78:69‐90. [DOI] [PubMed] [Google Scholar]
- 95. Taipale M, Krykbaeva I, Whitesell L, et al. Chaperones as thermodynamic sensors of drug‐target interactions reveal kinase inhibitor specificities in living cells. Nat Biotechnol. 2013;31(7):630‐637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96. Wang RY, Noddings CM, Kirschke E, Myasnikov AG, Johnson JL, Agard DA. Structure of Hsp90‐Hsp70‐Hop‐GR reveals the Hsp90 client‐loading mechanism. Nature. 2022;601(7893):460‐464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Noddings CM, Wang RY, Johnson JL, Agard DA. Structure of Hsp90‐p23‐GR reveals the Hsp90 client‐remodelling mechanism. Nature. 2022;601(7893):465‐469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98. Hantouche C, Williamson B, Valinsky WC, Solomon J, Shrier A, Young JC. Bag1 Co‐chaperone promotes TRC8 E3 ligase‐dependent degradation of misfolded human ether a go‐go‐related Gene (hERG) potassium channels. J Biol Chem. 2017;292(6):2287‐2300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99. Gomez‐Pastor R, Burchfiel ET, Thiele DJ. Regulation of heat shock transcription factors and their roles in physiology and disease. Nat Rev Mol Cell Biol. 2018;19(1):4‐19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100. Mahat DB, Salamanca HH, Duarte FM, Danko CG, Lis JT. Mammalian heat shock response and mechanisms underlying its genome‐wide transcriptional regulation. Mol Cell. 2016;62(1):63‐78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101. Zhou J, Wan J, Gao X, Zhang X, Jaffrey SR, Qian SB. Dynamic m(6)A mRNA methylation directs translational control of heat shock response. Nature. 2015;526(7574):591‐594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102. Chatterjee M, Andrulis M, Stuhmer T, et al. The PI3K/Akt signaling pathway regulates the expression of Hsp70, which critically contributes to Hsp90‐chaperone function and tumor cell survival in multiple myeloma. Haematologica. 2013;98(7):1132‐1141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103. Cesa LC, Shao H, Srinivasan SR, et al. X‐linked inhibitor of apoptosis protein (XIAP) is a client of heat shock protein 70 (Hsp70) and a biomarker of its inhibition. J Biol Chem. 2018;293(7):2370‐2380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104. Srinivasan SR, Cesa LC, Li X, et al. Heat shock protein 70 (Hsp70) suppresses RIP1‐dependent apoptotic and necroptotic cascades. Mol Cancer Res. 2018;16(1):58‐68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105. Guo Z, Song T, Wang Z, et al. The chaperone Hsp70 is a BH3 receptor activated by the pro‐apoptotic Bim to stabilize anti‐apoptotic clients. J Biol Chem. 2020;295(37):12900‐12909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106. Zhang H, Song T, Wang Z, et al. Bcr‐Abl drives the formation of Hsp70/Bim PPI to stabilize oncogenic clients and prevent cells from undergoing apoptosis. Biochem Pharmacol. 2022;198:114964. [DOI] [PubMed] [Google Scholar]
- 107. Saleh A, Srinivasula SM, Balkir L, Robbins PD, Alnemri ES. Negative regulation of the Apaf‐1 apoptosome by Hsp70. Nat Cell Biol. 2000;2(8):476‐483. [DOI] [PubMed] [Google Scholar]
- 108. Bruey JM, Ducasse C, Bonniaud P, et al. Hsp27 negatively regulates cell death by interacting with cytochrome c. Nat Cell Biol. 2000;2(9):645‐652. [DOI] [PubMed] [Google Scholar]
- 109. Guo F, Sigua C, Bali P, et al. Mechanistic role of heat shock protein 70 in Bcr‐Abl‐mediated resistance to apoptosis in human acute leukemia cells. Blood. 2005;105(3):1246‐1255. [DOI] [PubMed] [Google Scholar]
- 110. Oshita SE, Chen F, Kwan T, Yehiely F, Cryns VL. The small heat shock protein HspB2 is a novel anti‐apoptotic protein that inhibits apical caspase activation in the extrinsic apoptotic pathway. Breast Cancer Res Treat. 2010;124(2):307‐315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111. Shan YX, Liu TJ, Su HF, Samsamshariat A, Mestril R, Wang PH. Hsp10 and Hsp60 modulate Bcl‐2 family and mitochondria apoptosis signaling induced by doxorubicin in cardiac muscle cells. J Mol Cell Cardiol. 2003;35(9):1135‐1143. [DOI] [PubMed] [Google Scholar]
- 112. Sun B, Li G, Yu Q, Liu D, Tang X. HSP60 in cancer: a promising biomarker for diagnosis and a potentially useful target for treatment. J Drug Target. 2022;30(1):31‐45. [DOI] [PubMed] [Google Scholar]
- 113. Binder RJ, Vatner R, Srivastava P. The heat‐shock protein receptors: some answers and more questions. Tissue Antigens. 2004;64(4):442‐451. [DOI] [PubMed] [Google Scholar]
- 114. Kol A, Lichtman AH, Finberg RW, Libby P, EA Kurt‐Jones. Cutting edge: heat shock protein (HSP) 60 activates the innate immune response: cD14 is an essential receptor for HSP60 activation of mononuclear cells. J Immunol. 2000;164(1):13‐17. [DOI] [PubMed] [Google Scholar]
- 115. Samali A, Cai J, Zhivotovsky B, Jones DP, Orrenius S. Presence of a pre‐apoptotic complex of pro‐caspase‐3, Hsp60 and Hsp10 in the mitochondrial fraction of jurkat cells. EMBO J. 1999;18(8):2040‐2048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116. Nagel R, Semenova EA, Berns A. Drugging the addict: non‐oncogene addiction as a target for cancer therapy. EMBO Rep. 2016;17(11):1516‐1531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117. Thomas X, Campos L, Le QH, Guyotat D. Heat shock proteins and acute leukemias. Hematology. 2005;10(3):225‐235. [DOI] [PubMed] [Google Scholar]
- 118. Smith CC. Disease diversity and FLT3 mutations. Proc Natl Acad Sci U S A. 2013;110(52):20860‐20861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119. Hu C, Zou F, Wang A, et al. Targeting chaperon protein HSP70 as a novel therapeutic strategy for FLT3‐ITD‐positive acute myeloid leukemia. Signal Transduct Target Ther. 2021;6(1):334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120. Tong WG, Estrov Z, Wang Y, et al. The synthetic heat shock protein 90 (Hsp90) inhibitor EC141 induces degradation of Bcr‐Abl p190 protein and apoptosis of Ph‐positive acute lymphoblastic leukemia cells. Invest New Drugs. 2011;29(6):1206‐1212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121. Vahid S, Thaper D, Gibson KF, Bishop JL, Zoubeidi A. Molecular chaperone Hsp27 regulates the Hippo tumor suppressor pathway in cancer. Sci Rep. 2016;6:31842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122. Castilla C, Congregado B, Conde JM, et al. Immunohistochemical expression of Hsp60 correlates with tumor progression and hormone resistance in prostate cancer. Urology. 2010;76(4):1017 e1‐1017 e6. [DOI] [PubMed] [Google Scholar]
- 123. Shkedi A, Taylor IR, Echtenkamp F, et al. Selective vulnerabilities in the proteostasis network of castration‐resistant prostate cancer. Cell Chem Biol. 2022;29(3):490‐501 e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124. Legler JM, Ries LA, Smith MA, et al. Cancer surveillance series [corrected]: brain and other central nervous system cancers: recent trends in incidence and mortality. J Natl Cancer Inst. 1999;91(16):1382‐1390. [DOI] [PubMed] [Google Scholar]
- 125. Tang H, Li J, Liu X, Wang G, Luo M, Deng H. Down‐regulation of HSP60 suppresses the proliferation of glioblastoma cells via the ROS/AMPK/mTOR pathway. Sci Rep. 2016;6:28388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126. Jiang X, Zhou T, Wang Z, Qi B, Xia H. HSP47 Promotes glioblastoma stemlike cell survival by modulating tumor microenvironment extracellular matrix through TGF‐beta pathway. ACS Chem Neurosci. 2017;8(1):128‐134. [DOI] [PubMed] [Google Scholar]
- 127. Kluger HM, Chelouche Lev D, Kluger Y, et al. Using a xenograft model of human breast cancer metastasis to find genes associated with clinically aggressive disease. Cancer Res. 2005;65(13):5578‐5587. [DOI] [PubMed] [Google Scholar]
- 128. Mittal V. Epithelial mesenchymal transition in tumor metastasis. Annu Rev Pathol. 2018;13:395‐412. [DOI] [PubMed] [Google Scholar]
- 129. Jagadish N, Agarwal S, Gupta N, et al. Heat shock protein 70–2 (HSP70‐2) overexpression in breast cancer. J Exp Clin Cancer Res. 2016;35(1):150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130. Li Y, Kang X, Wang Q. HSP70 decreases receptor‐dependent phosphorylation of Smad2 and blocks TGF‐beta‐induced epithelial‐mesenchymal transition. J Genet Genomics. 2011;38(3):111‐116. [DOI] [PubMed] [Google Scholar]
- 131. Hanahan D, Weinberg RA. The hallmarks of cancer. Cell. 2000;100(1):57‐70. [DOI] [PubMed] [Google Scholar]
- 132. Nagaraju GP, Long TE, Park W, et al. Heat shock protein 90 promotes epithelial to mesenchymal transition, invasion, and migration in colorectal cancer. Mol Carcinog. 2015;54(10):1147‐1158. [DOI] [PubMed] [Google Scholar]
- 133. Rajabi M, Mousa SA. The role of angiogenesis in cancer treatment. Biomedicines. 2017;5(2):34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134. Salnikov AV, Bretz NP, Perne C, et al. Antibody targeting of CD24 efficiently retards growth and influences cytokine milieu in experimental carcinomas. Br J Cancer. 2013;108(7):1449‐1459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135. Wang X, Zhang Y, Zhao Y, et al. CD24 promoted cancer cell angiogenesis via Hsp90‐mediated STAT3/VEGF signaling pathway in colorectal cancer. Oncotarget. 2016;7(34):55663‐55676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136. Berthenet K, Bokhari A, Lagrange A, et al. HSP110 promotes colorectal cancer growth through STAT3 activation. Oncogene. 2017;36(16):2328‐2336. [DOI] [PubMed] [Google Scholar]
- 137. Battaglin F, Naseem M, Lenz HJ, Salem ME. Microsatellite instability in colorectal cancer: overview of its clinical significance and novel perspectives. Clin Adv Hematol Oncol. 2018;16(11):735‐745. [PMC free article] [PubMed] [Google Scholar]
- 138. Voll EA, Ogden IM, Pavese JM, et al. Heat shock protein 27 regulates human prostate cancer cell motility and metastatic progression. Oncotarget. 2014;5(9):2648‐2663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139. Andrieu C, Taieb D, Baylot V, et al. Heat shock protein 27 confers resistance to androgen ablation and chemotherapy in prostate cancer cells through eIF4E. Oncogene. 2010;29(13):1883‐1896. [DOI] [PubMed] [Google Scholar]
- 140. Huang CY, Wei PL, Chen WY, Chang WC, Chang YJ. Silencing heat shock protein 27 inhibits the progression and metastasis of colorectal cancer (CRC) by maintaining the stability of stromal interaction molecule 1 (STIM1) Proteins. Cells. 2018;7(12):262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141. Yu Z, Zhi J, Peng X, Zhong X, Xu A. Clinical significance of HSP27 expression in colorectal cancer. Mol Med Reports. 2010;3(6):953‐958. [DOI] [PubMed] [Google Scholar]
- 142. Sheng B, Qi C, Liu B, Lin Y, Fu T, Zeng Q. Increased HSP27 correlates with malignant biological behavior of non‐small cell lung cancer and predicts patient's survival. Sci Rep. 2017;7(1):13807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143. Hsu HS, Lin JH, Huang WC, et al. Chemoresistance of lung cancer stemlike cells depends on activation of Hsp27. Cancer. 2011;117(7):1516‐1528. [DOI] [PubMed] [Google Scholar]
- 144. Zhang D, Wong LL, Koay ES. Phosphorylation of Ser78 of Hsp27 correlated with HER‐2/neu status and lymph node positivity in breast cancer. Mol Cancer. 2007;6:52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145. Wang S, Zhang X, Wang H, Wang Y, Chen P, Wang L. Heat shock protein 27 enhances SUMOylation of heat shock protein B8 to accelerate the progression of breast cancer. Am J Pathol. 2020;190(12):2464‐2477. [DOI] [PubMed] [Google Scholar]
- 146. Thanner F, Sutterlin MW, Kapp M, et al. Heat shock protein 27 is associated with decreased survival in node‐negative breast cancer patients. Anticancer Res. 2005;25(3A):1649‐1653. [PubMed] [Google Scholar]
- 147. Chern Y, Zhang P, Ju H, Tai IT. Heat shock protein 47 promotes tumor survival and therapy resistance by modulating AKT signaling via PHLPP1 in colorectal cancer. Cancer Biol Med. 2020;17(2):343‐356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148. Mori K, Toiyama Y, Otake K, et al. Proteomics analysis of differential protein expression identifies heat shock protein 47 as a predictive marker for lymph node metastasis in patients with colorectal cancer. Int J Cancer. 2017;140(6):1425‐1435. [DOI] [PubMed] [Google Scholar]
- 149. Yoneda A, Minomi K, Tamura Y. Heat shock protein 47 confers chemoresistance on pancreatic cancer cells by interacting with calreticulin and IRE1alpha. Cancer Sci. 2021;112(7):2803‐2820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150. Yoneda A, Minomi K, Tamura Y. HSP47 promotes metastasis of breast cancer by interacting with myosin IIA via the unfolded protein response transducer IRE1alpha. Oncogene. 2020;39(23):4519‐4537. [DOI] [PubMed] [Google Scholar]
- 151. Wu W, Hu Z, Xiong L, Zou J. Heat shock protein 47 promotes cell migration and invasion through AKT signal in non‐small cell lung cancer. Anticancer Drugs. 2022;33(3):268‐277. [DOI] [PubMed] [Google Scholar]
- 152. Song X, Liao Z, Zhou C, et al. HSP47 is associated with the prognosis of laryngeal squamous cell carcinoma by inhibiting cell viability and invasion and promoting apoptosis. Oncol Rep. 2017;38(4):2444‐2452. [DOI] [PubMed] [Google Scholar]
- 153. Zhang J, Zhou X, Chang H, et al. Hsp60 exerts a tumor suppressor function by inducing cell differentiation and inhibiting invasion in hepatocellular carcinoma. Oncotarget. 2016;7(42):68976‐68989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154. Guo J, Zhu S, Deng H, Xu R. HSP60‐knockdown suppresses proliferation in colorectal cancer cells via activating the adenine/AMPK/mTOR signaling pathway. Oncol Lett. 2021;22(2):630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155. Li XS, Xu Q, Fu XY, Luo WS. Heat shock protein 60 overexpression is associated with the progression and prognosis in gastric cancer. PLoS One. 2014;9(9):e107507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156. Guo D, Zhang A, Huang J, Suo M, Zhong Y, Liang Y. Suppression of HSP70 inhibits the development of acute lymphoblastic leukemia via TAK1/Egr‐1. Biomed Pharmacother. 2019;119:109399. [DOI] [PubMed] [Google Scholar]
- 157. Ling X, Wan J, Peng B, Chen J. Hsp70 promotes SUMO of HIF‐1alpha and promotes lung cancer invasion and metastasis. J Oncol. 2021;2021:7873085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158. Dong J, Wu Z, Wang D, et al. Hsp70 binds to the androgen receptor N‐terminal domain and modulates the receptor function in prostate cancer cells. Mol Cancer Ther. 2019;18(1):39‐50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159. Gao G, Liu S, Yao Z, Zhan Y, Chen W, Liu Y. The prognostic significance of Hsp70 in patients with colorectal cancer patients: a PRISMA‐compliant meta‐analysis. Biomed Res Int. 2021;2021:5526327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160. Gehrmann M, Specht HM, Bayer C, et al. Hsp70–a biomarker for tumor detection and monitoring of outcome of radiation therapy in patients with squamous cell carcinoma of the head and neck. Radiat Oncol. 2014;9:131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161. Li G, Xu Y, Guan D, Liu Z, Liu DX. HSP70 protein promotes survival of C6 and U87 glioma cells by inhibition of ATF5 degradation. J Biol Chem. 2011;286(23):20251‐20259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162. Katayama K, Noguchi K, Sugimoto Y. Heat shock protein 90 inhibitors overcome the resistance to Fms‐like tyrosine kinase 3 inhibitors in acute myeloid leukemia. Oncotarget. 2018;9(76):34240‐34258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163. Jansson KH, Tucker JB, Stahl LE, et al. High‐throughput screens identify HSP90 inhibitors as potent therapeutics that target inter‐related growth and survival pathways in advanced prostate cancer. Sci Rep. 2018;8(1):17239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164. Niu M, Zhang B, Li L, et al. Targeting HSP90 inhibits proliferation and induces apoptosis through AKT1/ERK pathway in lung cancer. Front Pharmacol. 2021;12:724192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165. Wang Y, Chen Q, Wu D, et al. Lamin‐A interacting protein Hsp90 is required for DNA damage repair and chemoresistance of ovarian cancer cells. Cell Death Dis. 2021;12(8):786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166. Zagouri F, Bournakis E, Koutsoukos K, Papadimitriou CA. Heat shock protein 90 (hsp90) expression and breast cancer. Pharmaceuticals (Basel). 2012;5(9):1008‐1020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167. Archibald JM, Cavalier‐Smith T, Maier U, Douglas S. Molecular chaperones encoded by a reduced nucleus: the cryptomonad nucleomorph. J Mol Evol. 2001;52(6):490‐501. [DOI] [PubMed] [Google Scholar]
- 168. Jinwal UK, O'Leary JC 3rd, Borysov SI, et al. Hsc70 rapidly engages tau after microtubule destabilization. J Biol Chem. 2010;285(22):16798‐16805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169. Miyata Y, Koren J, Kiray J, Dickey CA, Gestwicki JE. Molecular chaperones and regulation of tau quality control: strategies for drug discovery in tauopathies. Future Med Chem. 2011;3(12):1523‐1537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170. Salminen A, Ojala J, Kaarniranta K, Hiltunen M, Soininen H. Hsp90 regulates tau pathology through co‐chaperone complexes in Alzheimer's disease. Prog Neurobiol. 2011;93(1):99‐110. [DOI] [PubMed] [Google Scholar]
- 171. Koren J 3rd, Jinwal UK, Lee DC, et al. Chaperone signalling complexes in Alzheimer's disease. J Cell Mol Med. 2009;13(4):619‐630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172. Kundel F, De S, Flagmeier P, et al. Hsp70 inhibits the nucleation and elongation of tau and sequesters tau aggregates with high affinity. ACS Chem Biol. 2018;13(3):636‐646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173. Bross P, Magnoni R, Bie AS. Molecular chaperone disorders: defective Hsp60 in neurodegeneration. Curr Top Med Chem. 2012;12(22):2491‐2503. [DOI] [PubMed] [Google Scholar]
- 174. Chen JX, Yan SD. Amyloid‐beta‐induced mitochondrial dysfunction. J Alzheimers Dis. 2007;12(2):177‐184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175. Sun Y, Zheng J, Xu Y, Zhang X. Paraquat‐induced inflammatory response of microglia through HSP60/TLR4 signaling. Hum Exp Toxicol. 2018;37(11):1161‐1168. [DOI] [PubMed] [Google Scholar]
- 176. Maiti P, Manna J, Dunbar GL. Current understanding of the molecular mechanisms in Parkinson's disease: targets for potential treatments. Transl Neurodegener. 2017;6:28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177. Wyttenbach A. Role of heat shock proteins during polyglutamine neurodegeneration: mechanisms and hypothesis. J Mol Neurosci. 2004;23(1‐2):69‐96. [DOI] [PubMed] [Google Scholar]
- 178. Bruinsma IB, Bruggink KA, Kinast K, et al. Inhibition of alpha‐synuclein aggregation by small heat shock proteins. Proteins. 2011;79(10):2956‐2967. [DOI] [PubMed] [Google Scholar]
- 179. Aridon P, Geraci F, Turturici G, D'Amelio M, Savettieri G, Sconzo G. Protective role of heat shock proteins in Parkinson's disease. Neurodegener Dis. 2011;8(4):155‐168. [DOI] [PubMed] [Google Scholar]
- 180. Vasan RS, Benjamin EJ. The future of cardiovascular epidemiology. Circulation. 2016;133(25):2626‐2633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181. Roth GA. Global burden of cardiovascular diseases and risk factors, 1990–2019. J Am Coll Cardiol. 2020;76(25):2982‐3021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182. Krishnan‐Sivadoss I, Mijares‐Rojas IA, Villarreal‐Leal RA, Torre‐Amione G, Knowlton AA, Guerrero‐Beltrán CE. Heat shock protein 60 and cardiovascular diseases: an intricate love‐hate story. Med Res Rev. 2020;41(1):29‐71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183. Seibert TA, Hibbert B, Chen Y‐X, et al. Serum heat shock protein 27 levels represent a potential therapeutic target for atherosclerosis. J Am Coll Cardiol. 2013;62(16):1446‐1454. [DOI] [PubMed] [Google Scholar]
- 184. Ishiwata T, Orosz A, Wang X, et al. HSPB2 is dispensable for the cardiac hypertrophic response but reduces mitochondrial energetics following pressure overload in mice. PLoS One. 2012;7(8):e42118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185. Charmpilas N, Kyriakakis E, Tavernarakis N. Small heat shock proteins in ageing and age‐related diseases. Cell Stress Chaperones. 2017;22(4):481‐492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186. Selcen D, Engel AG. Myofibrillar myopathy caused by novel dominant negative alpha B‐crystallin mutations. Ann Neurol. 2003;54(6):804‐810. [DOI] [PubMed] [Google Scholar]
- 187. Fan GC, Ren X, Qian J, et al. Novel cardioprotective role of a small heat‐shock protein, Hsp20, against ischemia/reperfusion injury. Circulation. 2005;111(14):1792‐1799. [DOI] [PubMed] [Google Scholar]
- 188. Chen L, Lizano P, Zhao X, et al. Preemptive conditioning of the swine heart by H11 kinase/Hsp22 provides cardiac protection through inducible nitric oxide synthase. Am J Physiol Heart Circul Physiol. 2011;300(4):H1303‐H1310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189. Golenhofen N, Perng MD, Quinlan RA, Drenckhahn D. Comparison of the small heat shock proteins alphaB‐crystallin, MKBP, HSP25, HSP20, and cvHSP in heart and skeletal muscle. Histochem Cell Biol. 2004;122(5):415‐425. [DOI] [PubMed] [Google Scholar]
- 190. Depre C, Wang L, Sui X, et al. H11 kinase prevents myocardial infarction by preemptive preconditioning of the heart. Circ Res. 2006;98(2):280‐288. [DOI] [PubMed] [Google Scholar]
- 191. Henderson B, Pockley AG. Proteotoxic stress and circulating cell stress proteins in the cardiovascular diseases. Cell Stress Chaperones. 2012;17(3):303‐311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192. Hayashi M, Imanaka‐Yoshida K, Yoshida T, et al. A crucial role of mitochondrial Hsp40 in preventing dilated cardiomyopathy. Nat Med. 2006;12(1):128‐132. [DOI] [PubMed] [Google Scholar]
- 193. Xu Q. Role of heat shock proteins in atherosclerosis. Arterioscler Thromb Vasc Biol. 2002;22(10):1547‐1559. [DOI] [PubMed] [Google Scholar]
- 194. Vicencio JM, Yellon DM, Sivaraman V, et al. Plasma exosomes protect the myocardium from ischemia‐reperfusion injury. J Am Coll Cardiol. 2015;65(15):1525‐1536. [DOI] [PubMed] [Google Scholar]
- 195. Mu H, Wang L, Zhao L. HSP90 inhibition suppresses inflammatory response and reduces carotid atherosclerotic plaque formation in ApoE mice. Cardiovasc Ther. 2017;35(2). [DOI] [PubMed] [Google Scholar]
- 196. Leishman SJ, Ford PJ, West MJ. Heat shock proteins: a double‐edged sword linking periodontal and cardiovascular diseases. Future Cardiol. 2017;13(6):515‐519. [DOI] [PubMed] [Google Scholar]
- 197. Geraldine J, Mala S, Takeuchi S. Heat shock proteins in cardiovascular stress. Clin Med Cardiol. 2008;2:245‐256. [Google Scholar]
- 198. Latchman DS. Heat shock proteins and cardiac protection. Cardiovasc Res. 2001;51:637‐646. [DOI] [PubMed] [Google Scholar]
- 199. Kilic A, Mandal K. Heat shock proteins: pathogenic role in atherosclerosis and potential therapeutic implications. Autoimmune Dis. 2012;2012:1‐9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200. Zanconato F, Battilana G, Forcato M, et al. Transcriptional addiction in cancer cells is mediated by YAP/TAZ through BRD4. Nat Med. 2018;24(10):1599‐1610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201. Rodriguez‐Iturbe B, Lanaspa MA, Johnson RJ. The role of autoimmune reactivity induced by heat shock protein 70 in the pathogenesis of essential hypertension. Br J Pharmacol. 2019;176:1829‐1838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202. Mehta TA, Greenman J, Ettelaie C, Venkatasubramaniam A, Chetter IC, McCollum PT. Heat shock proteins in vascular disease—a review. Eur J Vasc Endovasc Surg. 2005;29(4):395‐402. [DOI] [PubMed] [Google Scholar]
- 203. Murshid A, Eguchi T, Calderwood SK. Stress proteins in aging and life span. Int J Hyperthermia. 2013;29(5):442‐447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204. Berkowitz P, Hu P, Liu Z, et al. Desmosome signaling. Inhibition of p38MAPK prevents pemphigus vulgaris IgG‐induced cytoskeleton reorganization. J Biol Chem. 2005;280(25):23778‐23784. [DOI] [PubMed] [Google Scholar]
- 205. Kostenko S, Johannessen M, Moens U. PKA‐induced F‐actin rearrangement requires phosphorylation of Hsp27 by the MAPKAP kinase MK5. Cell Signal. 2009;21(5):712‐718. [DOI] [PubMed] [Google Scholar]
- 206. Thanner J, Bekos C, Veraar C, et al. Heat shock protein 90alpha in thymic epithelial tumors and non‐thymomatous myasthenia gravis. Oncoimmunology. 2020;9(1):1756130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207. Verleden GM, Glanville AR, Lease ED, et al. Chronic lung allograft dysfunction: definition, diagnostic criteria, and approaches to treatment‐A consensus report from the Pulmonary Council of the ISHLT. J Heart Lung Transplant. 2019;38(5):493‐503. [DOI] [PubMed] [Google Scholar]
- 208. Yonekura K, Yokota S, Tanaka S, et al. Prevalence of anti‐heat shock protein antibodies in cerebrospinal fluids of patients with Guillain‐Barre syndrome. J Neuroimmunol. 2004;156(1‐2):204‐209. [DOI] [PubMed] [Google Scholar]
- 209. Agius MA, Kirvan CA, Schafer AL, Gudipati E, Zhu S. High prevalence of anti‐alpha‐crystallin antibodies in multiple sclerosis: correlation with severity and activity of disease. Acta Neurol Scand. 1999;100(3):139‐147. [DOI] [PubMed] [Google Scholar]
- 210. Ben‐Ami Shor D, Blank M, Reuter S, et al. Anti‐ribosomal‐P antibodies accelerate lupus glomerulonephritis and induce lupus nephritis in naive mice. J Autoimmun. 2014;54:118‐126. [DOI] [PubMed] [Google Scholar]
- 211. Hadadi E, Zhang B, Baidzajevas K, et al. Differential IL‐1beta secretion by monocyte subsets is regulated by Hsp27 through modulating mRNA stability. Sci Rep. 2016;6:39035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212. Skrzeczynska‐Moncznik J, Bzowska M, Nogiec A, et al. Rapid externalization of 27‐kDa heat shock protein (HSP27) and atypical cell death in neutrophils treated with the sphingolipid analog drug FTY720. J Leukoc Biol. 2015;98(4):591‐599. [DOI] [PubMed] [Google Scholar]
- 213. Nasr SH, Fogo AB. New developments in the diagnosis of fibrillary glomerulonephritis. Kidney Int. 2019;96(3):581‐592. [DOI] [PubMed] [Google Scholar]
- 214. Baker LW, Khan M, Cortese C, Aslam N. Fibrillary glomerulonephritis or complement 3 glomerulopathy: a rare case of diffuse necrotising crescentic glomerulonephritis with C3‐dominant glomerular deposition and positive DNAJB9. BMJ Case Rep. 2021;14(2):e239868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215. Nguyen TQ, Jaramillo A, Thompson RW, et al. Increased expression of HDJ‐2 (hsp40) in carotid artery atherosclerosis: a novel heat shock protein associated with luminal stenosis and plaque ulceration. J Vasc Surg. 2001;33(5):1065‐1071. [DOI] [PubMed] [Google Scholar]
- 216. Banecka‐Majkutewicz Z, Grabowski M, Kadzinski L, Papkov A, Wegrzyn A, Banecki B. Increased levels of antibodies against heat shock proteins in stroke patients. Acta Biochim Pol. 2014;61(2):379‐383. [PubMed] [Google Scholar]
- 217. Kasperkiewicz M, Tukaj S, Gembicki AJ, et al. Evidence for a role of autoantibodies to heat shock protein 60, 70, and 90 in patients with dermatitis herpetiformis. Cell Stress Chaperones. 2014;19(6):837‐843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218. Cui J, Ma C, Ye G, et al. DnaJ (hsp40) of Streptococcus pneumoniae is involved in bacterial virulence and elicits a strong natural immune reaction via PI3K/JNK. Mol Immunol. 2017;83:137‐146. [DOI] [PubMed] [Google Scholar]
- 219. Massa M, Passalia M, Manzoni SM, et al. Differential recognition of heat‐shock protein dnaJ‐derived epitopes by effector and Treg cells leads to modulation of inflammation in juvenile idiopathic arthritis. Arthritis Rheum. 2007;56(5):1648‐1657. [DOI] [PubMed] [Google Scholar]
- 220. Qamra R, Mande SC. Crystal structure of the 65‐kilodalton heat shock protein, chaperonin 60.2, of Mycobacterium tuberculosis. J Bacteriol. 2004;186(23):8105‐8113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221. Puga Yung GL, Fidler M, Albani E, et al. Heat shock protein‐derived T‐cell epitopes contribute to autoimmune inflammation in pediatric Crohn's disease. PLoS One. 2009;4(11):e7714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222. van der Zee R, Anderton SM, Prakken AB, Liesbeth Paul AG, van Eden W. T cell responses to conserved bacterial heat‐shock‐protein epitopes induce resistance in experimental autoimmunity. Semin Immunol. 1998;10(1):35‐41. [DOI] [PubMed] [Google Scholar]
- 223. Yokota SI, Hirata D, Minota S, et al. Autoantibodies against chaperonin CCT in human sera with rheumatic autoimmune diseases: comparison with antibodies against other Hsp60 family proteins. Cell Stress Chaperones. 2000;5(4):337‐346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224. Horvath L, Czirjak L, Fekete B, et al. Levels of antibodies against C1q and 60 kDa family of heat shock proteins in the sera of patients with various autoimmune diseases. Immunol Lett. 2001;75(2):103‐109. [DOI] [PubMed] [Google Scholar]
- 225. Menge T, Rzepka R, Melchers I. Monoclonal autoantibodies from patients with autoimmune diseases: specificity, affinity and crossreactivity of MAbs binding to cytoskeletal and nucleolar epitopes, cartilage antigens and mycobacterial heat‐shock protein 60. Immunobiology. 2002;205(1):1‐16. [DOI] [PubMed] [Google Scholar]
- 226. Yoshida Y, Zhang XM, Wang H, et al. Elevated levels of autoantibodies against DNAJC2 in sera of patients with atherosclerotic diseases. Heliyon. 2020;6(8):e04661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227. Conroy SE, Faulds GB, Williams W, Latchman DS, Isenberg DA. Detection of autoantibodies to the 90 kDa heat shock protein in systemic lupus erythematosus and other autoimmune diseases. Br J Rheumatol. 1994;33(10):923‐926. [DOI] [PubMed] [Google Scholar]
- 228. Verma S, Goyal S, Jamal S, Singh A, Grover A. Hsp90: friends, clients and natural foes. Biochimie. 2016;127:227‐240. [DOI] [PubMed] [Google Scholar]
- 229. Lv K, Jiang J, Donaghy R, et al. CBL family E3 ubiquitin ligases control JAK2 ubiquitination and stability in hematopoietic stem cells and myeloid malignancies. Genes Dev. 2017;31(10):1007‐1023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230. Elias D, Prigozin H, Polak N, Rapoport M, Lohse AW, Cohen IR. Autoimmune diabetes induced by the beta‐cell toxin STZ. Immunity to the 60‐kDa heat shock protein and to insulin. Diabetes. 1994;43(8):992‐998. [DOI] [PubMed] [Google Scholar]
- 231. Bras A, Aguas AP. Diabetes‐prone NOD mice are resistant to Mycobacterium avium and the infection prevents autoimmune disease. Immunology. 1996;89(1):20‐25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232. Elias D, Meilin A, Ablamunits V, et al. Hsp60 peptide therapy of NOD mouse diabetes induces a Th2 cytokine burst and downregulates autoimmunity to various beta‐cell antigens. Diabetes. 1997;46(5):758‐764. [DOI] [PubMed] [Google Scholar]
- 233. Zonneveld‐Huijssoon E, van Wijk F, Roord S, et al. TLR9 agonist CpG enhances protective nasal HSP60 peptide vaccine efficacy in experimental autoimmune arthritis. Ann Rheum Dis. 2012;71(10):1706‐1715. [DOI] [PubMed] [Google Scholar]
- 234. Karopoulos C, Rowley MJ, Handley CJ, Strugnell RA. Antibody reactivity to mycobacterial 65 kDa heat shock protein: relevance to autoimmunity. J Autoimmun. 1995;8(2):235‐248. [DOI] [PubMed] [Google Scholar]
- 235. Zonneveld‐Huijssoon E, Albani S, Prakken BJ, van Wijk F. Heat shock protein bystander antigens for peptide immunotherapy in autoimmune disease. Clin Exp Immunol. 2013;171(1):20‐29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236. Androvitsanea A, Stylianou K, Drosataki E, Petrakis I. The pathophysiological role of heat shock response in autoimmunity: a literature review. Cells. 2021;10(10):2626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237. Niu L, Lou F, Sun Y, et al. A micropeptide encoded by lncRNA MIR155HG suppresses autoimmune inflammation via modulating antigen presentation. Sci Adv. 2020;6(21):eaaz2059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238. Elson CJ, Thompson SJ. Immunity, autoimmunity and immunotherapy: new frontiers in heat shock protein research. Clin Exp Immunol. 1994;98(2):175‐177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 239. Millar DG, Garza KM, Odermatt B, et al. Hsp70 promotes antigen‐presenting cell function and converts T‐cell tolerance to autoimmunity in vivo. Nat Med. 2003;9(12):1469‐1476. [DOI] [PubMed] [Google Scholar]
- 240. Purcell AW, Todd A, Kinoshita G, et al. Association of stress proteins with autoantigens: a possible mechanism for triggering autoimmunity?. Clin Exp Immunol. 2003;132(2):193‐200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241. Paggi A, Di Prima MA, Paparo BS, et al. Anti 70 kDa heat shock protein antibodies in sera of patients affected by autoimmune and non‐autoimmune thyroid diseases. Endocr Res. 1995;21(3):555‐567. [DOI] [PubMed] [Google Scholar]
- 242. Tebo AE, Szankasi P, Hillman TA, Litwin CM, Hill HR. Antibody reactivity to heat shock protein 70 and inner ear‐specific proteins in patients with idiopathic sensorineural hearing loss. Clin Exp Immunol. 2006;146(3):427‐432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243. Moin ASM, Nandakumar M, Diane A, Dehbi M, Butler AE. The role of heat shock proteins in type 1 diabetes. Front Immunol. 2020;11:612584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244. Petrakis I, Androvitsanea A, Stratakis S, Daphnis E, Stylianou K. Intense immunostaining of heat shock protein 70 within renal interstitium associates with long‐term renal survival in an ANCA‐associated vasculitis cohort. Cell Stress Chaperones. 2021;26(1):51‐65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245. van Herwijnen MJ, Wieten L, van der Zee R, et al. Regulatory T cells that recognize a ubiquitous stress‐inducible self‐antigen are long‐lived suppressors of autoimmune arthritis. Proc Natl Acad Sci U S A. 2012;109(35):14134‐14139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246. Galazka G, Jurewicz A, Domowicz M, Cannella B, Raine CS, Selmaj K. HINT1 peptide/Hsp70 complex induces NK‐cell‐dependent immunoregulation in a model of autoimmune demyelination. Eur J Immunol. 2014;44(10):3026‐3044. [DOI] [PubMed] [Google Scholar]
- 247. Knutson SK, Kawano S, Minoshima Y, et al. Selective inhibition of EZH2 by EPZ‐6438 leads to potent antitumor activity in EZH2‐mutant non‐Hodgkin lymphoma. Mol Cancer Ther. 2014;13(4):842‐854. [DOI] [PubMed] [Google Scholar]
- 248. Heneka MT, Sharp A, Murphy P, Lyons JA, Dumitrescu L, Feinstein DL. The heat shock response reduces myelin oligodendrocyte glycoprotein‐induced experimental autoimmune encephalomyelitis in mice. J Neurochem. 2001;77(2):568‐579. [DOI] [PubMed] [Google Scholar]
- 249. Khalafalla MG, Woods LT, Camden JM, et al. P2X7 receptor antagonism prevents IL‐1beta release from salivary epithelial cells and reduces inflammation in a mouse model of autoimmune exocrinopathy. J Biol Chem. 2017;292(40):16626‐16637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250. Kasperkiewicz M, Muller R, Manz R, et al. Heat‐shock protein 90 inhibition in autoimmunity to type VII collagen: evidence that nonmalignant plasma cells are not therapeutic targets. Blood. 2011;117(23):6135‐6142. [DOI] [PubMed] [Google Scholar]
- 251. Chandawarkar RY, Wagh MS, Kovalchin JT, Srivastava P. Immune modulation with high‐dose heat‐shock protein gp96: therapy of murine autoimmune diabetes and encephalomyelitis. Int Immunol. 2004;16(4):615‐624. [DOI] [PubMed] [Google Scholar]
- 252. Ye BX, Deng X, Shao LD, et al. Vibsanin B preferentially targets HSP90beta, inhibits interstitial leukocyte migration, and ameliorates experimental autoimmune encephalomyelitis. J Immunol. 2015;194(9):4489‐4497. [DOI] [PubMed] [Google Scholar]
- 253. Tukaj S, Zillikens D, Kasperkiewicz M. Heat shock protein 90: a pathophysiological factor and novel treatment target in autoimmune bullous skin diseases. Exp Dermatol. 2015;24(8):567‐571. [DOI] [PubMed] [Google Scholar]
- 254. Shrestha L, Bolaender A, Patel HJ, Taldone T. Heat shock protein (HSP) drug discovery and development: targeting heat shock proteins in disease. Curr Top Med Chem. 2016;16(25):2753‐2764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255. Stebbins CE, Russo AA, Schneider C, Rosen N, Hartl FU, Pavletich NP. Crystal structure of an Hsp90‐geldanamycin complex: targeting of a protein chaperone by an antitumor agent. Cell. 1997;89(2):239‐250. [DOI] [PubMed] [Google Scholar]
- 256. Kumar S, Stokes J, Singh UP, et al. Targeting Hsp70: a possible therapy for cancer. Cancer Lett. 2016;374(1):156‐166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 257. Miyata Y, Nakamoto H, Neckers L. The therapeutic target Hsp90 and cancer hallmarks. Curr Pharm Des. 2013;19(3):347‐365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258. Blagosklonny MV. Hsp‐90‐associated oncoproteins: multiple targets of geldanamycin and its analogs. Leukemia. 2002;16(4):455‐462. [DOI] [PubMed] [Google Scholar]
- 259. Kluger HM, Chelouche Lev D, Kluger Y, et al. Using a Xenograft model of human breast cancer metastasis to find genes associated with clinically aggressive disease. Cancer Res. 2005;65(13):5578‐5587. [DOI] [PubMed] [Google Scholar]
- 260. Merendino AM, Bucchieri F, Campanella C, et al. Hsp60 is actively secreted by human tumor cells. PLoS One. 2010;5(2):e9247‐e9247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 261. Fucarino A, Pitruzzella A. Role of HSP60/HSP10 in lung cancer: simple biomarkers or leading actors?. J Oncol. 2020;2020:4701868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262. Hoter A, Rizk S, Naim HY. Heat shock protein 60 in hepatocellular carcinoma: insights and perspectives. Mini Review. 2020;7:60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263. Meng Q, Li BX, Xiao X. Toward developing chemical modulators of Hsp60 as potential therapeutics. Front Mol Biosci. 2018;5:35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 264. Duan X, Iwanowycz S, Ngoi S, Hill M, Zhao Q, Liu B. Molecular Chaperone GRP94/GP96 in cancers: oncogenesis and therapeutic target. Front Oncol. 2021;11:629846‐629846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 265. Ansa‐Addo EA, Thaxton J, Hong F, et al. Clients and oncogenic roles of molecular Chaperone gp96/grp94. Curr Top Med Chem. 2016;16(25):2765‐2778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 266. Carpenter RL, Gökmen‐Polar Y. HSF1 as a cancer biomarker and therapeutic target. Curr Cancer Drug Targets. 2019;19(7):515‐524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 267. Santagata S, Hu R, Lin Nancy U, et al. High levels of nuclear heat‐shock factor 1 (HSF1) are associated with poor prognosis in breast cancer. Proc Natl Acad Sci. 2011;108(45):18378‐18383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 268. Dong B, Jaeger AM, Thiele DJ. Inhibiting heat shock factor 1 in cancer: a unique therapeutic opportunity. Trends Pharmacol Sci. 2019;40(12):986‐1005. [DOI] [PubMed] [Google Scholar]
- 269. Lianos GD, Alexiou GA, Mangano A, et al. The role of heat shock proteins in cancer. Cancer Lett. 2015;360(2):114‐118. [DOI] [PubMed] [Google Scholar]
- 270. Khalil AA, Kabapy NF, Deraz SF, Smith C. Heat shock proteins in oncology: diagnostic biomarkers or therapeutic targets?. Biochim Biophys Acta. 2011;1816(2):89‐104. [DOI] [PubMed] [Google Scholar]
- 271. Banerjee S, Lin CF, Skinner KA, et al. Heat shock protein 27 differentiates tolerogenic macrophages that may support human breast cancer progression. Cancer Res. 2011;71(2):318‐327. [DOI] [PubMed] [Google Scholar]
- 272. Mischak H, Allmaier G, Apweiler R, et al. Recommendations for biomarker identification and qualification in clinical proteomics. Sci Transl Med. 2010;2(46):46ps42. [DOI] [PubMed] [Google Scholar]
- 273. Mosser DD, Morimoto RI. Molecular chaperones and the stress of oncogenesis. Oncogene. 2004;23(16):2907‐2918. [DOI] [PubMed] [Google Scholar]
- 274. Qiu XB, Shao YM, Miao S, Wang L. The diversity of the DnaJ/Hsp40 family, the crucial partners for Hsp70 chaperones. Cell Mol Life Sci CMLS. 2006;63(22):2560‐2570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 275. Sharma A, Upadhyay AK, Bhat MK. Inhibition of Hsp27 and Hsp40 potentiates 5‐fluorouracil and carboplatin mediated cell killing in hepatoma cells. Cancer Biol Ther. 2009;8(22):2106‐2113. [DOI] [PubMed] [Google Scholar]
- 276. Langer R, Ott K, Specht K, et al. Protein expression profiling in esophageal adenocarcinoma patients indicates association of heat‐shock protein 27 expression and chemotherapy response. Clin Cancer Res. 2008;14(24):8279‐8287. [DOI] [PubMed] [Google Scholar]
- 277. Dickey CA, Kamal A, Lundgren K, et al. The high‐affinity HSP90‐CHIP complex recognizes and selectively degrades phosphorylated tau client proteins. J Clin Invest. 2007;117(3):648‐658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 278. Petrucelli L, Dickson D, Kehoe K, et al. CHIP and Hsp70 regulate tau ubiquitination, degradation and aggregation. Hum Mol Genet. 2004;13(7):703‐714. [DOI] [PubMed] [Google Scholar]
- 279. Dou F, Netzer WJ, Tanemura K, et al. Chaperones increase association of tau protein with microtubules. Proc Natl Acad Sci U S A. 2003;100(2):721‐726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 280. Wang L, Xie C, Greggio E, et al. The Chaperone activity of heat shock protein 90 is critical for maintaining the stability of leucine‐rich repeat kinase 2. J Neurosci. 2008;28(13):3384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 281. Witt SN. Molecular chaperones, α‐synuclein, and neurodegeneration. Mol Neurobiol. 2013;47(2):552‐560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 282. Beretta G, Shala AL. Impact of heat shock proteins in neurodegeneration: possible therapeutical targets. Ann Neurosci. 2022:09727531211070528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 283. Franklin TB, Krueger‐Naug AM, Clarke DB, Arrigo AP, Currie RW. The role of heat shock proteins Hsp70 and Hsp27 in cellular protection of the central nervous system. Int J Hyperther. 2005;21(5):379‐392. [DOI] [PubMed] [Google Scholar]
- 284. Abisambra JF, Blair LJ, Hill SE, et al. Phosphorylation dynamics regulate Hsp27‐mediated rescue of neuronal plasticity deficits in tau transgenic mice. J Neurosci. 2010;30(46):15374‐15382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 285. Wilhelmus MM, Boelens WC, Otte‐Höller I, et al. Small heat shock protein HspB8: its distribution in Alzheimer's disease brains and its inhibition of amyloid‐beta protein aggregation and cerebrovascular amyloid‐beta toxicity. Acta Neuropathol. 2006;111(2):139‐149. [DOI] [PubMed] [Google Scholar]
- 286. Cox D, Carver JA, Ecroyd H. Preventing α‐synuclein aggregation: the role of the small heat‐shock molecular chaperone proteins. Biochim Biophys Acta. 2014;1842(9):1830‐1843. [DOI] [PubMed] [Google Scholar]
- 287. Ma CH, Omura T, Cobos EJ, et al. Accelerating axonal growth promotes motor recovery after peripheral nerve injury in mice. J Clin Invest. 2011;121(11):4332‐4347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 288. Morrow G, Le Pécheur M, Tanguay RMJB. Drosophila melanogaster mitochondrial Hsp22: a role in resistance to oxidative stress, aging and the mitochondrial unfolding protein response. Biotechnology. 2016;17(1):61‐70. [DOI] [PubMed] [Google Scholar]
- 289. Reil J‐C, Hohl M, Oberhofer M, et al. Cardiac Rac1 overexpression in mice creates a substrate for atrial arrhythmias characterized by structural remodelling. Cardiovasc Res. 2010;87(3):485‐493. [DOI] [PubMed] [Google Scholar]
- 290. Ogata T, Ueyama T, Isodono K, et al. MURC, a muscle‐restricted coiled‐coil protein that modulates the Rho/ROCK pathway, induces cardiac dysfunction and conduction disturbance. Mol Cell Biol. 2008;28(10):3424‐3436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 291. Sanbe A, Daicho T, Mizutani R, et al. Protective effect of geranylgeranylacetone via enhancement of HSPB8 induction in desmin‐related cardiomyopathy. PLoS One. 2009;4(4):e5351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 292. Park HK, Park E‐C, Bae SW, et al. Expression of heat shock protein 27 in human atherosclerotic plaques and increased plasma level of heat shock protein 27 in patients with acute coronary syndrome. Circulation. 2006;114(9):886‐893. [DOI] [PubMed] [Google Scholar]
- 293. Clements RT, Sodha NR, Feng J, et al. Phosphorylation and translocation of heat shock protein 27 and αB‐crystallin in human myocardium after cardioplegia and cardiopulmonary bypass. J Thorac Cardiovasc Surg. 2007;134(6):1461‐1470. e3. [DOI] [PubMed] [Google Scholar]
- 294. Lu X‐Y, Chen L, Cai X‐L, Yang H‐T. Overexpression of heat shock protein 27 protects against ischaemia/reperfusion‐induced cardiac dysfunction via stabilization of troponin I and T. Cardiovasc Res. 2008;79(3):500‐508. [DOI] [PubMed] [Google Scholar]
- 295. Efthymiou CA, Mocanu MM, de Belleroche J, Wells DJ, Latchmann DS, DM Yellon. Heat shock protein 27 protects the heart against myocardial infarction. Basic Res Cardiol. 2004;99(6):392‐394. [DOI] [PubMed] [Google Scholar]
- 296. Hollander JM, Martin JL, Belke DD, et al. Overexpression of wild‐type heat shock protein 27 and a nonphosphorylatable heat shock protein 27 mutant protects against ischemia/reperfusion injury in a transgenic mouse model. Circulation. 2004;110(23):3544‐3552. [DOI] [PubMed] [Google Scholar]
- 297. Voegeli TS, Currie RW. siRNA knocks down Hsp27 and increases angiotensin II‐Induced phosphorylated NF‐κB p65 levels in aortic smooth muscle cells. Inflamm Res. 2009;58(6):336‐343. [DOI] [PubMed] [Google Scholar]
- 298. Salari S, Seibert T, Chen Y‐X, et al. Extracellular HSP27 acts as a signaling molecule to activate NF‐κB in macrophages. Cell Stress Chaperones. 2013;18(1):53‐63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 299. Rayner K, Chen Y‐X, McNulty M, et al. Extracellular release of the atheroprotective heat shock protein 27 is mediated by estrogen and competitively inhibits acLDL binding to scavenger receptor‐A. Circ Res. 2008;103(2):133‐141. [DOI] [PubMed] [Google Scholar]
- 300. Hammerer‐Lercher A, Mair J, Bonatti J, Watzka SB, Puschendorf B, Dirnhofer S. Hypoxia induces heat shock protein expression in human coronary artery bypass grafts. Cardiovasc Res. 2001;50(1):115‐124. [DOI] [PubMed] [Google Scholar]
- 301. Shor AJC. Mechanism of arterial infection by Chlamydia pneumoniae. Circulation. 2001;104(13):e75‐e75. [PubMed] [Google Scholar]
- 302. Xu Q, Schett G, Perschinka H, et al. Serum soluble heat shock protein 60 is elevated in subjects with atherosclerosis in a general population. Circulation. 2000;102(1):14‐20. [DOI] [PubMed] [Google Scholar]
- 303. Huittinen T, Leinonen M, Tenkanen L, et al. Autoimmunity to human heat shock protein 60, Chlamydia pneumoniae infection, and inflammation in predicting coronary risk. Arterioscler Thromb Vasc Biol. 2002;22(3):431‐437. [DOI] [PubMed] [Google Scholar]
- 304. Buono C, Pang H, Uchida Y, Libby P, Sharpe AH, Lichtman AH. B7‐1/B7‐2 costimulation regulates plaque antigen–specific T‐cell responses and atherogenesis in low‐density lipoprotein receptor–deficient mice. Circulation. 2004;109(16):2009‐2015. [DOI] [PubMed] [Google Scholar]
- 305. Liu J, Kam KW, Borchert GH, Kravtsov GM, Ballard HJ, TM Wong. Further study on the role of HSP70 on Ca2+ homeostasis in rat ventricular myocytes subjected to simulated ischemia. Am J Physiol Cell Physiol. 2006;290(2):C583‐C591. [DOI] [PubMed] [Google Scholar]
- 306. O'brien P, Li G, Locke M, Klabunde R, Ianuzo CD. Compensatory up‐regulation of cardiac SR Ca2+‐pump by heat‐shock counteracts SR Ca2+‐channel activation by ischemia/reperfusion. Mol Cell Biochem. 1997;173(1):135‐143. [DOI] [PubMed] [Google Scholar]
- 307. Zhang X, Chen S, Yoo S, et al. Mutation in nuclear pore component NUP155 leads to atrial fibrillation and early sudden cardiac death. Cell. 2008;135(6):1017‐1027. [DOI] [PubMed] [Google Scholar]
- 308. Juwono J, Martinus RD. Does Hsp60 provide a link between Mitochondrial stress and inflammation in diabetes mellitus?. J Diabetes Res. 2016;2016:8017571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 309. Quintana FJ, Cohen IR. The HSP60 immune system network. Trends Immunol. 2011;32(2):89‐95. [DOI] [PubMed] [Google Scholar]
- 310. Marino Gammazza A, Rizzo M, Citarrella R, et al. Elevated blood Hsp60, its structural similarities and cross‐reactivity with thyroid molecules, and its presence on the plasma membrane of oncocytes point to the chaperonin as an immunopathogenic factor in Hashimoto's thyroiditis. Cell Stress Chaperones. 2014;19(3):343‐353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 311. Gammazza AM, Bucchieri F, Grimaldi LM, et al. The molecular anatomy of human Hsp60 and its similarity with that of bacterial orthologs and acetylcholine receptor reveal a potential pathogenetic role of anti‐chaperonin immunity in myasthenia gravis. Cell Mol Neurobiol. 2012;32(6):943‐947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 312. Graner MW. HSP90 and immune modulation in cancer. Adv Cancer Res. 2016;129:191‐224. [DOI] [PubMed] [Google Scholar]
- 313. Vega VL, Rodríguez‐Silva M, Frey T, et al. Hsp70 translocates into the plasma membrane after stress and is released into the extracellular environment in a membrane‐associated form that activates macrophages. J Immunol. 2008;180(6):4299‐4307. [DOI] [PubMed] [Google Scholar]
- 314. Borges TJ, Wieten L, van Herwijnen MJ, et al. The anti‐inflammatory mechanisms of Hsp70. Front Immunol. 2012;3:95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 315. Mbofung RM, McKenzie JA, Malu S, et al. HSP90 inhibition enhances cancer immunotherapy by upregulating interferon response genes. Nat Commun. 2017;8(1):451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 316. Tiwari S, Thakur R, Shankar J. Role of heat‐shock proteins in cellular function and in the biology of fungi. Biotechnol Res Int. 2015;2015:132635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 317. Füst G, Beck Z, Bánhegyi D, Kocsis J, Bíró A, Prohászka Z. Antibodies against heat shock proteins and cholesterol in HIV infection. Mol Immunol. 2005;42(1):79‐85. [DOI] [PubMed] [Google Scholar]
- 318. Babaahmady K, Oehlmann W, Singh M, Lehner T. Inhibition of human immunodeficiency virus type 1 infection of human CD4+ T cells by microbial HSP70 and the peptide epitope 407–426. J Virol. 2007;81(7):3354‐3360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 319. Zhu XD, Li CL, Lang ZW, Gao GF, Tien P. Significant correlation between expression level of HSP gp96 and progression of hepatitis B virus induced diseases. World J Gastroenterol. 2004;10(8):1141‐1145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 320. Wu Y, Cui J, Zhang X, et al. Pneumococcal DnaJ modulates dendritic cell‐mediated Th1 and Th17 immune responses through Toll‐like receptor 4 signaling pathway. Immunobiology. 2017;222(2):384‐393. [DOI] [PubMed] [Google Scholar]
- 321. Lang BJ, Guerrero‐Giménez ME, Prince TL, Ackerman A, Bonorino C, Calderwood SK. Heat shock proteins are essential components in transformation and tumor progression: cancer cell intrinsic pathways and beyond. Int J Mol Sci. 2019;20(18):4507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 322. Pacey S, Wilson RH, Walton M, et al. A phase I study of the heat shock protein 90 inhibitor alvespimycin (17‐DMAG) given intravenously to patients with advanced solid tumors. Clin Cancer Res. 2011;17(6):1561‐1570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 323. Belli F, Testori A, Rivoltini L, et al. Vaccination of metastatic melanoma patients with autologous tumor‐derived heat shock protein gp96‐peptide complexes: clinical and immunologic findings. J Clin Oncol. 2002;20(20):4169‐4180. [DOI] [PubMed] [Google Scholar]
- 324. Enomoto Y, Bharti A, Khaleque AA, et al. Enhanced immunogenicity of heat shock protein 70 peptide complexes from dendritic cell‐tumor fusion cells. J Immunol. 2006;177(9):5946‐5955. [DOI] [PubMed] [Google Scholar]
- 325. Kim YS, Alarcon SV, Lee S, et al. Update on Hsp90 inhibitors in clinical trial. Curr Top Med Chem. 2009;9(15):1479‐1492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 326. Uehara Y, Hori M, Takeuchi T, Umezawa H. Phenotypic change from transformed to normal induced by benzoquinonoid ansamycins accompanies inactivation of p60src in rat kidney cells infected with Rous sarcoma virus. Mol Cell Biol. 1986;6(6):2198‐2206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 327. Whitesell L, Mimnaugh EG, De Costa B, Myers CE, Neckers LM. Inhibition of heat shock protein HSP90‐pp60v‐src heteroprotein complex formation by benzoquinone ansamycins: essential role for stress proteins in oncogenic transformation. Proc Natl Acad Sci U S A. 1994;91(18):8324‐8328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 328. Kamal A, Thao L, Sensintaffar J, et al. A high‐affinity conformation of Hsp90 confers tumour selectivity on Hsp90 inhibitors. Nature. 2003;425(6956):407‐410. [DOI] [PubMed] [Google Scholar]
- 329. Moulick K, Ahn JH, Zong H, et al. Affinity‐based proteomics reveal cancer‐specific networks coordinated by Hsp90. Nat Chem Biol. 2011;7(11):818‐826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 330. Backe SJ, Sager RA, Woodford MR, Makedon AM, Mollapour M. Post‐translational modifications of Hsp90 and translating the chaperone code. J Biol Chem. 2020;295(32):11099‐11117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 331. Sidera K, Patsavoudi E. HSP90 inhibitors: current development and potential in cancer therapy. Recent Patents Anti‐Cancer Drug Discov. 2014;9(1):1‐20. [PubMed] [Google Scholar]
- 332. Ciocca DR, Calderwood SK. Heat shock proteins in cancer: diagnostic, prognostic, predictive, and treatment implications. Cell Stress Chaperones. 2005;10(2):86‐103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 333. Zylicz M, King FW, Wawrzynow A. Hsp70 interactions with the p53 tumour suppressor protein. EMBO J. 2001;20(17):4634‐4638. 10.1093/emboj/20.17.4634 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 334. Gong J, Weng D, Eguchi T, et al. Targeting the hsp70 gene delays mammary tumor initiation and inhibits tumor cell metastasis. Oncogene. 2015;34(43):5460‐5471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 335. Chatterjee S, Burns TF. Targeting heat shock proteins in cancer: a promising therapeutic approach. Int J Mol Sci. 2017;18(9):1978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 336. Nylandsted J, Gyrd‐Hansen M, Danielewicz A, et al. Heat shock protein 70 promotes cell survival by inhibiting lysosomal membrane permeabilization. J Exp Med. 2004;200(4):425‐435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 337. Blachere NE, Li Z, Chandawarkar RY, et al. Heat shock protein–peptide complexes, reconstituted in vitro, elicit peptide‐specific cytotoxic T lymphocyte response and tumor immunity. J Exp Med. 1997;186(8):1315‐1322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 338. Multhoff G, Pfister K, Gehrmann M, et al. A 14‐mer Hsp70 peptide stimulates natural killer (NK) cell activity. Cell Stress Chaperones. 2001;6(4):337‐344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 339. Wachstein J, Tischer S, Figueiredo C, et al. HSP70 enhances immunosuppressive function of CD4(+)CD25(+)FoxP3(+) T regulatory cells and cytotoxicity in CD4(+)CD25(‐) T cells. PLoS One. 2012;7(12):e51747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 340. Diao J, Yang X, Song X, et al. Exosomal Hsp70 mediates immunosuppressive activity of the myeloid‐derived suppressor cells via phosphorylation of Stat3. Med Oncol. 2015;32(2):1‐10. [DOI] [PubMed] [Google Scholar]
- 341. Pfister K, Radons J, Busch R, et al. Patient survival by Hsp70 membrane phenotype: association with different routes of metastasis. Cancer. 2007;110(4):926‐935. [DOI] [PubMed] [Google Scholar]
- 342. Leu JIJ, Pimkina J, Pandey P, Murphy ME, George DL. HSP70 inhibition by the small‐molecule 2‐phenylethynesulfonamide impairs protein clearance pathways in tumor cells. Mol Cancer Res. 2011;9(7):936‐947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 343. Yaglom JA, Wang Y, Li A, et al. Cancer cell responses to Hsp70 inhibitor JG‐98: comparison with Hsp90 inhibitors and finding synergistic drug combinations. Sci Rep. 2018;8(1):3010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 344. Wen W, Liu W, Shao Y, Chen L. VER‐155008, a small molecule inhibitor of HSP70 with potent anti‐cancer activity on lung cancer cell lines. Exp Biol Med. 2014;239(5):638‐645. [DOI] [PubMed] [Google Scholar]
- 345. Guzhova IV, Margulis BA. HSP70‐based anti‐cancer immunotherapy. Hum Vaccin Immunother. 2016;12(10):2529‐2535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 346. Chen Y, Chen C, Ma C, Sun S, Zhang J, Sun Y. Expression of heat‐shock protein gp96 in gallbladder cancer and its prognostic clinical significance. Int J Clin Exp Pathol. 2015;8(2):1946‐1953. [PMC free article] [PubMed] [Google Scholar]
- 347. Singh‐Jasuja H, Hilf N, Scherer HU, et al. The heat shock protein gp96: a receptor‐targeted cross‐priming carrier and activator of dendritic cells. Cell Stress Chaperones. 2000;5(5):462‐470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 348. Yedavelli SP, Guo L, Daou ME, Srivastava PK, Mittelman A, Tiwari RK. Preventive and therapeutic effect of tumor derived heat shock protein, gp96, in an experimental prostate cancer model. Int J Mol Med. 1999;4(3):243‐248. [DOI] [PubMed] [Google Scholar]
- 349. Choi S‐K, Kam H, Kim K‐Y, Park SI, Lee Y‐S. Targeting heat shock protein 27 in cancer: a druggable target for cancer treatment?. Cancers (Basel). 2019;11(8):1195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 350. Meriin AB, Sherman MY. Role of molecular chaperones in neurodegenerative disorders. Int J Hyperthermia. 2005;21(5):403‐419. [DOI] [PubMed] [Google Scholar]
- 351. Adachi H, Katsuno M, Waza M, Minamiyama M, Tanaka F, Sobue G. Heat shock proteins in neurodegenerative diseases: pathogenic roles and therapeutic implications. Int J Hyperthermia. 2009;25(8):647‐654. [DOI] [PubMed] [Google Scholar]
- 352. Kuta R, Larochelle N, Fernandez M, et al. Depending on the stress, histone deacetylase inhibitors act as heat shock protein co‐inducers in motor neurons and potentiate arimoclomol, exerting neuroprotection through multiple mechanisms in ALS models. Cell Stress Chaperones. 2020;25(1):173‐191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 353. Chaudhuri TK, Paul S. Protein‐misfolding diseases and chaperone‐based therapeutic approaches. FEBS J. 2006;273(7):1331‐1349. [DOI] [PubMed] [Google Scholar]
- 354. Sweeney P, Park H, Baumann M, et al. Protein misfolding in neurodegenerative diseases: implications and strategies. Transl Neurodegener. 2017;6:6‐6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 355. Krause M, da Costa Rodrigues‐Krause J. Extracellular heat shock proteins (eHSP70) in exercise: possible targets outside the immune system and their role for neurodegenerative disorders treatment. Med Hypotheses. 2011;76(2):286‐290. [DOI] [PubMed] [Google Scholar]
- 356. Tidwell JL, Houenou LJ, Tytell M. Administration of Hsp70 in vivo inhibits motor and sensory neuron degeneration. Cell Stress Chaperones. 2004;9(1):88‐98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 357. Bobkova N, Guzhova I, Margulis B, et al. Dynamics of endogenous Hsp70 synthesis in the brain of olfactory bulbectomized mice. Cell Stress Chaperones. 2013;18(1):109‐118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 358. Bobkova NV, Garbuz DG, Nesterova I, et al. Therapeutic effect of exogenous hsp70 in mouse models of Alzheimer's disease. J Alzheimer's Dis JAD. 2014;38(2):425‐435. [DOI] [PubMed] [Google Scholar]
- 359. Laure L, Long R, Lizano P, et al. Cardiac H11 kinase/Hsp22 stimulates oxidative phosphorylation and modulates mitochondrial reactive oxygen species production: involvement of a nitric oxide‐dependent mechanism. Free Radic Biol Med. 2012;52(11‐12):2168‐2176. [DOI] [PubMed] [Google Scholar]
- 360. Marunouchi T, Inomata S, Sanbe A, Takagi N, Tanonaka K. Protective effect of geranylgeranylacetone via enhanced induction of HSPB1 and HSPB8 in mitochondria of the failing heart following myocardial infarction in rats. Eur J Pharmacol. 2014;730:140‐147. [DOI] [PubMed] [Google Scholar]
- 361. Cao H, Xue L, Xu X, et al. Heat shock proteins in stabilization of spontaneously restored sinus rhythm in permanent atrial fibrillation patients after mitral valve surgery. Cell Stress Chaperones. 2011;16(5):517‐528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 362. Marunouchi T, Abe Y, Murata M, et al. Changes in small heat shock proteins HSPB1, HSPB5 and HSPB8 in mitochondria of the failing heart following myocardial infarction in rats. Biol Pharm Bull. 2013;36(4):529‐539. [DOI] [PubMed] [Google Scholar]
- 363. Qiu H, Lizano P, Laure L, et al. H11 kinase/heat shock protein 22 deletion impairs both nuclear and mitochondrial functions of STAT3 and accelerates the transition into heart failure on cardiac overload. Circulation. 2011;124(4):406‐415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 364. White MY, Hambly BD, Jeremy RW, Cordwell SJ. Ischemia‐specific phosphorylation and myofilament translocation of heat shock protein 27 precedes alpha B‐crystallin and occurs independently of reactive oxygen species in rabbit myocardium. J Mol Cell Cardiol. 2006;40(6):761‐774. [DOI] [PubMed] [Google Scholar]
- 365. Knowlton AA, Kapadia S, Torre‐Amione G, et al. Differential expression of heat shock proteins in normal and failing human hearts. J Mol Cell Cardiol. 1998;30(4):811‐818. [DOI] [PubMed] [Google Scholar]
- 366. Delogu G, Signore M, Mechelli A, Famularo G. Heat shock proteins and their role in heart injury. Curr Opin Crit Care. 2002;8(5):411‐416. [DOI] [PubMed] [Google Scholar]
- 367. Kim SH, Koh GY, Cho KW, Park W‐Y, Seo J‐S. Stretch‐activated atrial natriuretic peptide secretion in atria with heat shock protein 70 overexpression. Exp Biol Med (Maywood). 2003;228(2):200‐206. [DOI] [PubMed] [Google Scholar]
- 368. Elahi M, Hadjinikolaou L, Galiñanes M. Incidence and clinical consequences of atrial fibrillation within 1 year of first‐time isolated coronary bypass surgery. Circulation. 2003;108(suppl 1):II207‐II212. [DOI] [PubMed] [Google Scholar]
- 369. Mandal K, Torsney E, Poloniecki J, Camm AJ, Xu Q, Jahangiri M. Association of high intracellular, but not serum, heat shock protein 70 with postoperative atrial fibrillation. Ann Thorac Surg. 2005;79(3):865‐871. [DOI] [PubMed] [Google Scholar]
- 370. Hoag JB, Qian Y‐Z, Nayeem MA, D'Angelo M, Kukreja RC. ATP‐sensitive potassium channel mediates delayed ischemic protection by heat stress in rabbit heart. Am J Physiol. 1997;273(5):H2458‐H2464. [DOI] [PubMed] [Google Scholar]
- 371. Liu Y, Mayr S, Opitz‐Gress A, et al. Human skeletal muscle HSP70 response to training in highly trained rowers. J Appl Physiol. 1999;86(1):101‐104. [DOI] [PubMed] [Google Scholar]
- 372. Zhang X, Xu Z, Zhou L, et al. Plasma levels of Hsp70 and anti‐Hsp70 antibody predict risk of acute coronary syndrome. Cell Stress Chaperones. 2010;15(5):675‐686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 373. Kirchhoff S, Gupta S, Knowlton AA. Cytosolic heat shock protein 60, apoptosis, and myocardial injury. Circulation. 2002;105(24):2899‐2904. [DOI] [PubMed] [Google Scholar]
- 374. Rocnik E, Chow LH, Pickering JG. Heat shock protein 47 is expressed in fibrous regions of human atheroma and is regulated by growth factors and oxidized low‐density lipoprotein. Circulation. 2000;101(11):1229‐1233. [DOI] [PubMed] [Google Scholar]
- 375. Moudgil KD, Thompson SJ, Geraci F, De Paepe B, Shoenfeld Y. Heat‐shock proteins in autoimmunity. Autoimmune Dis. 2013;2013:621417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 376. Tukaj S, Kaminski M. Heat shock proteins in the therapy of autoimmune diseases: too simple to be true?. Cell Stress Chaperones. 2019;24(3):475‐479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 377. Huurman VA, Decochez K, Mathieu C, Cohen IR, Roep BO. Therapy with the hsp60 peptide DiaPep277 in C‐peptide positive type 1 diabetes patients. Diabetes Metab Res Rev. 2007;23(4):269‐275. [DOI] [PubMed] [Google Scholar]
- 378. Zininga T, Ramatsui L, Shonhai A. Heat shock proteins as immunomodulants. Molecules. 2018;23(11):2846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 379. Tsan M‐F, Gao B. Cytokine function of heat shock proteins. Am J Physiol‐Cell Physiol. 2004;286(4):C739‐C744. [DOI] [PubMed] [Google Scholar]
- 380. Bolhassani A, Agi E. Heat shock proteins in infection. Clin Chim Acta. 2019;498:90‐100. [DOI] [PubMed] [Google Scholar]
- 381. Binder RJ. Heat‐shock protein‐based vaccines for cancer and infectious disease. Expert Rev Vaccines. 2008;7(3):383‐393. [DOI] [PubMed] [Google Scholar]
- 382. Geller R, Vignuzzi M, Andino R, Frydman J. Evolutionary constraints on chaperone‐mediated folding provide an antiviral approach refractory to development of drug resistance. Genes Dev. 2007;21(2):195‐205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 383. Pack CD, Kumaraguru U, Suvas S, Rouse BT. Heat‐shock protein 70 acts as an effective adjuvant in neonatal mice and confers protection against challenge with herpes simplex virus. Vaccine. 2005;23(27):3526‐3534. [DOI] [PubMed] [Google Scholar]
- 384. Babaahmady K, Bergmeier LA, Lehner T. Combining human antisera to human leukocyte antigens, HIVgp120 and 70 kDa heat shock protein results in broadly neutralizing activity to HIV‐1. AIDS. 2008;22(11):1267‐1276. [DOI] [PubMed] [Google Scholar]
- 385. Gráf L, Barabás L, Madaras B, et al. High serum Hsp70 level predicts poor survival in colorectal cancer: results obtained in an independent validation cohort. Cancer Biomarkers: Sect A Dis Markers. 2018;23(4):539‐547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 386. Arap MA, Lahdenranta J, Mintz PJ, et al. Cell surface expression of the stress response chaperone GRP78 enables tumor targeting by circulating ligands. Cancer Cell. 2004;6(3):275‐284. [DOI] [PubMed] [Google Scholar]
- 387. Xu W, Liu L, Brown NJ, Christian S, Hornby D. Quantum dot‐conjugated anti‐GRP78 scFv inhibits cancer growth in mice. Molecules. 2012;17(1):796‐808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 388. Shi K, Wang Y, Zhou X, et al. Tumor microenvironment targeting with dual stimuli‐responsive nanoparticles based on small heat shock proteins for antitumor drug delivery. Acta Biomater. 2020;114:369‐383. [DOI] [PubMed] [Google Scholar]
- 389. Fiesel FC, James ED, Hudec R, Springer W. Mitochondrial targeted HSP90 inhibitor Gamitrinib‐TPP (G‐TPP) induces PINK1/Parkin‐dependent mitophagy. Oncotarget. 2017;8(63):106233‐106248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 390. Shekhawat SD, Purohit HJ, Taori GM, Daginawala HF, Kashyap RS. Evaluation of heat shock proteins for discriminating between latent tuberculosis infection and active tuberculosis: a preliminary report. J Infect Public Health. 2016;9(2):143‐152. [DOI] [PubMed] [Google Scholar]
- 391. Gerges SE, Alosh TK, Khalil SH, El Din MMW. Relevance of Helicobacter pylori infection in Egyptian multiple sclerosis patients. Egypt J Neurol Psychiatry Neurosur. 2018;54(1):41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 392. Li Z, Song Y, Xing R, et al. Heat shock protein 70 acts as a potential biomarker for early diagnosis of heart failure. PLoS One. 2013;8(7):e67964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 393. Pacey S, Gore M, Chao D, et al. A phase II trial of 17‐allylamino, 17‐demethoxygeldanamycin (17‐AAG, tanespimycin) in patients with metastatic melanoma. Invest New Drugs. 2012;30(1):341‐349. [DOI] [PubMed] [Google Scholar]
- 394. Modi S, Stopeck A, Linden H, et al. HSP90 inhibition is effective in breast cancer: a phase II trial of tanespimycin (17‐AAG) plus trastuzumab in patients with HER2‐positive metastatic breast cancer progressing on trastuzumab. Clin Cancer Res. 2011;17(15):5132‐5139. [DOI] [PubMed] [Google Scholar]
- 395. Richardson PG, Chanan‐Khan AA, Lonial S, et al. Tanespimycin and bortezomib combination treatment in patients with relapsed or relapsed and refractory multiple myeloma: results of a phase 1/2 study. Br J Haematol. 2011;153(6):729‐740. [DOI] [PubMed] [Google Scholar]
- 396. Arteaga CL. Why is this effective HSP90 inhibitor not being developed in HER2+ breast cancer?. Clin Cancer Res. 2011;17(15):4919‐4921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 397. Jhaveri K, Miller K, Rosen L, et al. A phase I dose‐escalation trial of trastuzumab and alvespimycin hydrochloride (KOS‐1022; 17 DMAG) in the treatment of advanced solid tumors. Clin Cancer Res. 2012;18(18):5090‐5098. [DOI] [PubMed] [Google Scholar]
- 398. Siegel D, Yan C, Ross D. NAD(P)H:quinone oxidoreductase 1 (NQO1) in the sensitivity and resistance to antitumor quinones. Biochem Pharmacol. 2012;83(8):1033‐1040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 399. Sydor JR, Normant E, Pien CS, et al. Development of 17‐allylamino‐17‐demethoxygeldanamycin hydroquinone hydrochloride (IPI‐504), an anti‐cancer agent directed against Hsp90. Proc Natl Acad Sci U S A. 2006;103(46):17408‐17413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 400. Ge J, Normant E, Porter JR, et al. Design, synthesis, and biological evaluation of hydroquinone derivatives of 17‐amino‐17‐demethoxygeldanamycin as potent, water‐soluble inhibitors of Hsp90. J Med Chem. 2006;49(15):4606‐4615. [DOI] [PubMed] [Google Scholar]
- 401. Porter JR, Ge J, Lee J, Normant E, West K. Ansamycin inhibitors of Hsp90: nature's prototype for anti‐chaperone therapy. Curr Top Med Chem. 2009;9(15):1386‐1418. [DOI] [PubMed] [Google Scholar]
- 402. Brough PA, Aherne W, Barril X, et al. 4,5‐Diarylisoxazole Hsp90 Chaperone inhibitors: potential therapeutic agents for the treatment of cancer. J Med Chem. 2008;51(2):196‐218. [DOI] [PubMed] [Google Scholar]
- 403. Eccles SA, Massey A, Raynaud FI, et al. NVP‐AUY922: a novel heat shock protein 90 inhibitor active against Xenograft tumor growth, angiogenesis, and metastasis. Cancer Res. 2008;68(8):2850‐2860. [DOI] [PubMed] [Google Scholar]
- 404. Woodhead AJ, Angove H, Carr MG, et al. Discovery of (2,4‐Dihydroxy‐5‐isopropylphenyl)‐[5‐(4‐methylpiperazin‐1‐ylmethyl)‐1,3‐dihydroisoindol‐2‐yl]methanone (AT13387), a novel inhibitor of the molecular Chaperone Hsp90 by fragment based drug design. J Med Chem. 2010;53(16):5956‐5969. [DOI] [PubMed] [Google Scholar]
- 405. Chan KC, Ting CM, Chan PS, et al. A novel Hsp90 inhibitor AT13387 induces senescence in EBV‐positive nasopharyngeal carcinoma cells and suppresses tumor formation. Mol Cancer. 2013;12(1):128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 406. Graham B, Curry J, Smyth T, et al. The heat shock protein 90 inhibitor, AT13387, displays a long duration of action in vitro and in vivo in non‐small cell lung cancer. Cancer Sci. 2012;103(3):522‐527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 407. Colunga Biancatelli RML, Solopov P, Dimitropoulou C, Gregory B, Day T, Catravas JD. The heat shock protein 90 inhibitor, AT13387, protects the Alveolo‐Capillary barrier and prevents HCl‐induced chronic lung injury and pulmonary fibrosis. Cells. 2022;11(6):1046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 408. Ying W, Du Z, Sun L, et al. Ganetespib, a unique triazolone‐containing Hsp90 inhibitor, exhibits potent antitumor activity and a superior safety profile for cancer therapy. Mol Cancer Ther. 2012;11(2):475‐484. [DOI] [PubMed] [Google Scholar]
- 409. Shimamura T, Perera SA, Foley KP, et al. Ganetespib (STA‐9090), a nongeldanamycin HSP90 inhibitor, has potent antitumor activity in in vitro and in vivo models of non‐small cell lung cancer. Clin Cancer Res. 2012;18(18):4973‐4985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 410. He H, Zatorska D, Kim J, et al. Identification of potent water soluble purine‐scaffold inhibitors of the heat shock protein 90. J Med Chem. 2006;49(1):381‐390. [DOI] [PubMed] [Google Scholar]
- 411. Sugita M, Wilkes DC, Bareja R, et al. Targeting the epichaperome as an effective precision medicine approach in a novel PML‐SYK fusion acute myeloid leukemia. npj Precision Oncol. 2021;5(1):44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 412. Bolaender A, Zatorska D, He H, et al. Chemical tools for epichaperome‐mediated interactome dysfunctions of the central nervous system. Nat Commun. 2021;12(1):4669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 413. Kim SH, Bajji A, Tangallapally R, et al. Discovery of (2S)‐1‐[4‐(2‐{6‐amino‐8‐[(6‐bromo‐1,3‐benzodioxol‐5‐yl)sulfanyl]‐9H‐purin‐9‐yl}ethyl)piperidin‐1‐yl]‐2‐hydroxypropan‐1‐one (MPC‐3100), a purine‐based Hsp90 inhibitor. J Med Chem. 2012;55(17):7480‐7501. [DOI] [PubMed] [Google Scholar]
- 414. Bao R, Lai CJ, Wang DG, et al. Targeting heat shock protein 90 with CUDC‐305 overcomes erlotinib resistance in non‐small cell lung cancer. Mol Cancer Ther. 2009;8(12):3296‐3306. [DOI] [PubMed] [Google Scholar]
- 415. Lundgren K, Zhang H, Brekken J, et al. BIIB021, an orally available, fully synthetic small‐molecule inhibitor of the heat shock protein Hsp90. Mol Cancer Ther. 2009;8(4):921‐929. [DOI] [PubMed] [Google Scholar]
- 416. Dickson MA, Okuno SH, Keohan ML, et al. Phase II study of the HSP90‐inhibitor BIIB021 in gastrointestinal stromal tumors. Ann Oncol. 2013;24(1):252‐257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 417. Lundgren K, Biamonte MA. CHAPTER 5 the discovery of BIIB021 and BIIB028. In: Machajewski TD, ed. Inhibitors of molecular chaperones as therapeutic agents. The Royal Society of Chemistry. 2014:158‐179. [Google Scholar]
- 418. Okawa Y, Hideshima T, Steed P, et al. SNX‐2112, a selective Hsp90 inhibitor, potently inhibits tumor cell growth, angiogenesis, and osteoclastogenesis in multiple myeloma and other hematologic tumors by abrogating signaling via Akt and ERK. Blood. 2009;113(4):846‐855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 419. Brough PA, Barril X, Borgognoni J, et al. Combining Hit identification strategies: fragment‐Based and in silico approaches to orally active 2‐aminothieno[2,3‐d]pyrimidine inhibitors of the Hsp90 molecular chaperone. J Med Chem. 2009;52(15):4794‐4809. [DOI] [PubMed] [Google Scholar]
- 420. Bregnhøj A, Thuesen KKH, Emmanuel T, et al. HSP90 inhibitor RGRN‐305 for oral treatment of plaque‐type psoriasis: efficacy, safety and biomarker results in an open‐label proof‐of‐concept study. Br J Dermatol. 2022;186:861‐874. [DOI] [PubMed] [Google Scholar]
- 421. Stangl S, Gehrmann M, Riegger J, et al. Targeting membrane heat‐shock protein 70 (Hsp70) on tumors by cmHsp70.1 antibody. Proc Natl Acad Sci. 2011;108(2):733‐738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 422. Krause SW, Gastpar R, Andreesen R, et al. Treatment of colon and lung cancer patients with ex vivo heat shock protein 70‐Peptide‐activated, autologous natural killer cells: a clinical phase I trial. Clin Cancer Res. 2004;10(11):3699‐3707. [DOI] [PubMed] [Google Scholar]
- 423. Kumar S, Deepak P, Acharya A. Autologous Hsp70 immunization induces anti‐tumor immunity and increases longevity and survival of tumor‐bearing mice. Neoplasma. 2009;56(3):259‐268. [DOI] [PubMed] [Google Scholar]
- 424. Testori A, Richards J, Whitman E, et al. Phase III comparison of Vitespen, an autologous tumor‐derived heat shock protein gp96 peptide complex vaccine, with physician's choice of treatment for stage IV Melanoma: the C‐100‐21 study group. J Clin Oncol. 2008;26(6):955‐962. [DOI] [PubMed] [Google Scholar]
- 425. Wood CG, Mulders P. Vitespen: a preclinical and clinical review. Future Oncol. 2009;5(6):763‐774. [DOI] [PubMed] [Google Scholar]
- 426. Spigel DR, Shipley DL, Waterhouse DM, et al. A randomized, double‐blinded, Phase II trial of Carboplatin and Pemetrexed with or without Apatorsen (OGX‐427) in patients with previously untreated stage IV non‐squamous‐non‐small‐cell lung cancer: the SPRUCE trial. Oncologist. 2019;24(12):e1409‐e1416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 427. Rosenberg JE, Hahn NM, Regan MM, et al. Apatorsen plus docetaxel versus docetaxel alone in platinum‐resistant metastatic urothelial carcinoma (Borealis‐2). Br J Cancer. 2018;118(11):1434‐1441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 428. Netzer WJ, Hartl FU. Recombination of protein domains facilitated by co‐translational folding in eukaryotes. Nature. 1997;388(6640):343‐349. [DOI] [PubMed] [Google Scholar]
- 429. Nakamoto H, Vígh L. The small heat shock proteins and their clients. Cel Mol Life Sci CMLS. 2007;64(3):294‐306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 430. Kim YS, Alarcon SV, Lee S, et al. Update on Hsp90 inhibitors in clinical trial. Curr Top Med Chem. 2009;9(15):1479‐1492. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supporting Information
Data Availability Statement
The data included in this study are available upon request from the corresponding authors.