

Abstract
Shigella flexneri is a gram-negative bacterium that causes diarrhea and dysentery by invasion and spread through the colonic epithelium. Bacteria spread by assembling actin and other cytoskeletal proteins of the host into “actin tails” at the bacterial pole; actin tail assembly provides the force required to move bacteria through the cell cytoplasm and into adjacent cells. The 120-kDa S. flexneri outer membrane protein IcsA is essential for actin assembly. IcsA is anchored in the outer membrane by a carboxy-terminal domain (the β domain), such that the amino-terminal 706 amino acid residues (the α domain) are exposed on the exterior of the bacillus. The α domain is therefore likely to contain the domains that are important to interactions with host factors. We identify and characterize a domain of IcsA within the α domain that bears significant sequence similarity to two repeated domains of rickettsial OmpA, which has been implicated in rickettsial actin tail formation. Strains of S. flexneri and Escherichia coli that carry derivatives of IcsA containing deletions within this domain display loss of actin recruitment and increased accessibility to IcsA-specific antibody on the surface of intracytoplasmic bacteria. However, site-directed mutagenesis of charged residues within this domain results in actin assembly that is indistinguishable from that of the wild type, and in vitro competition of a polypeptide of this domain fused to glutathione S-transferase did not alter the motility of the wild-type construct. Taken together, our data suggest that the rickettsial homology domain of IcsA is required for the proper conformation of IcsA and that its disruption leads to loss of interactions of other IcsA domains within the amino terminus with host cytoskeletal proteins.
The gram-negative bacterium Shigella flexneri causes diarrhea and dysentery by directly invading the colonic epithelium and eliciting a severe local inflammatory response. After inducing its entry into the cell, S. flexneri rapidly lyses the phagocytic vacuole and is released into the cytoplasm (6, 21, 27, 40). Within the cytoplasm, bacteria assemble actin on one pole of the bacterial body into tails that consist of bundled actin filaments. The continuous polymerization of actin at the junction of the bacterial body with the tail and the incorporation of the resulting actin filaments into the tail provide the force to propel the microorganism through the cytoplasm at speeds of 12 μm min−1. Then, in a process that also requires actin assembly, bacteria induce cell surface protrusions, which enable bacterial spread into adjacent cells (23, 35, 37, 41).
IcsA, a 120-kDa S. flexneri outer membrane protein, has been shown to be essential and sufficient for actin tail formation (4, 17, 26, 30). A strain with a disruption of icsA invades cells and lyses the phagocytic vacuole but fails to induce polymerization of actin and remains confined to the cytoplasm (9). In addition, introduction of icsA into Escherichia coli confers upon this organism the ability to form actin tails in cytoplasmic extracts (17, 26).
IcsA is an 1,102-amino-acid protein that is unusual in that it is localized at one pole of S. flexneri (16). It is anchored in the outer membrane via a domain within the carboxy-terminal 344 amino acid residues (the β domain), thereby exposing its amino-terminal 706 amino acid residues (the α domain) on the exterior of the bacillus (45). Once translocated across the outer membrane, IcsA is slowly cleaved by IcsP (SopA) between Arg758 and Arg759 to release the 95-kDa α domain into the growth medium or the cytosol (12, 13, 42). The specific domain(s) of IcsA that is required for actin assembly has not been identified, although deletion of a large region encompassing amino acids 320 to 507 leads to loss of actin recruitment on the surface of the bacillus (46).
Several other intracellular microorganisms have been shown to assemble actin tails that are similar to those assembled by Shigella: the gram-positive bacterium Listeria monocytogenes (8, 32, 48), certain members of the Rickettsiaceae family of small gram-negative bacteria (19), and vaccinia virus (7). The specific protein that mediates actin assembly on L. monocytogenes, ActA, has been extensively characterized, while those that mediate actin assembly on rickettsiae and vaccinia virus have not yet been definitively identified. ActA has been shown to be the sole Listeria factor required for this process (11, 25) and bears no significant sequence similarity to IcsA. ActA has two domains that appear to have distinct roles in actin tail formation: an amino-terminal domain (amino acid residues 128 to 151) that is required for actin recruitment and a domain containing a series of four proline-rich repeats (amino acid residues 263 to 390) that directly binds the vasodilator-stimulated phosphoprotein (VASP), a focal adhesion protein, which in turn recruits profilin-actin to the surface of the bacterium (5, 24, 29, 36, 43). Deletion of the actin recruitment domain leads to loss of actin association with ActA (29, 36), whereas deletion of the proline-rich repeats leads to slowing of the rate of actin assembly by approximately two-thirds (28, 33, 43). In addition, mounting evidence indicates that the Arp2/3 complex of seven proteins is also required for actin assembly on L. monocytogenes (49, 50).
Among the Rickettsiaceae, spotted-fever group rickettsiae (Rickettsia rickettsii, R. conorii, and R. akari) form actin tails in the cytoplasm of infected cells, whereas typhus-group rickettsiae do not. Since spotted-fever group rickettsiae express the 190-kDa outer membrane protein OmpA and typhus-group rickettsiae lack OmpA, it has been suggested that OmpA may have a role in rickettsial actin tail formation (19, 20).
In this context, we were interested in defining the IcsA domain(s) that is essential to its function in actin assembly. We identified a domain of IcsA that bears significant sequence similarity to a repeated domain of rickettsial OmpA. We therefore analyzed the role of this domain in translocation of IcsA to the bacterial surface and in actin assembly.
MATERIALS AND METHODS
Bacterial strains, plasmids, and cell lines.
The bacterial strains and plasmids used in this study are listed in Table 1. All strains carrying icsA alleles are derived from S. flexneri serotype 5 wild-type strain M90T (39) or E. coli MC1061 (31). Shigella strains were grown in tryptic soy broth, and E. coli strains were grown in Luria-Bertaini broth. Where appropriate, antibiotics were used at the following concentrations: ampicillin, 100 μg/ml; spectinomycin, 100 μg/ml; kanamycin, 40 μg/ml. HeLa and L2 cells used for plaque assays and Shigella intracellular motility assays were maintained in minimal essential medium supplemented with 10% fetal bovine serum and 1% nonessential amino acids.
TABLE 1.
Strains and plasmids
| Bacterial strain or plasmid | Relevant characteristics | Reference or source |
|---|---|---|
| S. flexneri strains | ||
| M90T | Wild-type serotype 5 | 39 |
| SC560 | M90T ΔicsA::Ω Spr | 9 |
| MAC1000 | SC560 carrying icsA on pMBG235, Spr Ampr | This study |
| MAC1003 | SC560 carrying icsA3 on pMAC103, Spr Ampr | This study |
| MAC1007 | SC560 carrying icsA7 on pMAC107, Spr Ampr | This study |
| MAC1008 | SC560 carrying icsA8 on pMAC108, Spr Ampr | This study |
| MAC1009 | SC560 carrying icsA9 on pMAC109, Spr Ampr | This study |
| MAC1010 | SC560 carrying icsA10 on pMAC210, Spr Ampr | This study |
| MAC1014 | SC560 carrying icsA14 on pMAC214, Spr Ampr | This study |
| MAC1015 | SC560 carrying icsA15 on pMAC215, Spr Ampr | This study |
| MAC1016 | SC560 carrying icsA16 on pMAC216, Spr Ampr | This study |
| MAC1017 | SC560 carrying icsA17 on pMAC217, Spr Ampr | This study |
| E. coli strains | ||
| BL21 | F−ompT [lon] hsdSB (rB−mB−) gal dcm | 44 |
| MC1061 | F−araD139 Δ(ara-leu)7696 galE15 galK16 Δ(lac)X74 rpsL (Strr) hsdR2 (rK− mK+) mcrA mcrB1 | 31 |
| MBG263 | MC1061 ompT::Km | 17 |
| MAC2000 | MBG263 carrying icsA on pMBG235, Kmr Ampr | This study |
| MAC2003 | MBG263 carrying icsA3 on pMAC103, Kmr Ampr | This study |
| MAC2009 | MBG263 carrying icsA9 on pMAC109, Kmr Ampr | This study |
| MAC2016 | MBG263 carrying icsA16 on pMAC216, Kmr Ampr | This study |
| MAC2017 | MBG263 carrying icsA17 on pMAC217, Kmr Ampr | This study |
| Plasmids | ||
| pMBG226 | pBR322 containing icsA from S. flexneri M90T, Ampr Cmr | This study |
| pMBG235 | pMBG226 with elimination of the BamHI site, Ampr Cmr | This study |
| pGEX-2T | gst gene fusion vector, Ampr | Pharmacia |
| pMBG282 | pGEX-2T with icsA 1912–2088a inserted at its BamHI and EcoRI sites | This study |
| pMAC103 | pMBG235 containing icsA3, which is icsA Δ(1909–2091) | This study |
| pMAC107 | pMBG235 containing icsA7, which is icsA Δ(1474–2175)::ompA 2671–2868b | This study |
| pMAC108 | pMBG235 containing icsA8, which is icsA Δ(1474–2175)::ompA 2128–2436 | This study |
| pMAC109 | pMBG235 containing icsA9, which is icsA Δ(1474–2175)::ompA 2320–2454 | This study |
| pMAC210 | pMBG235 containing icsA10, which is icsA 2032 GATACAGATGGCGAC 2047 changed to 2032 GCTACAGCTGGCGCT 2047 | This study |
| pMAC214 | pMBG235 containing icsA14, which is icsA 1972 GACTATACAAAA 1983 changed to 1972 GCCTATACAGCT 1983 | This study |
| pMAC215 | pMBG235 containing icsA15, which is icsA 1996 GACCAGAATAAA 2007 changed to 1996 GCCCAGAATGCG 2007 | This study |
| pMAC216 | pMBG235 containing icsA16, which is icsA Δ(1909–1992) | This study |
| pMAC217 | pMBG235 containing icsA17, which is icsA Δ(2020–2091) | This study |
Construction of in-frame deletions within icsA.
Plasmid pMBG235 (icsA WT [wild-type]) contains the entire coding sequence for icsA, as well as 523 bp of flanking DNA upstream of the translational start site and 56 bp of flanking DNA downstream of the translational stop codon. pMAC103 (icsA3), pMAC216 (icsA16), and pMAC217 (icsA17) were generated by reverse PCR on pMB6235 DNA, using oligonucleotide primer pairs 5′-CTGTGGATCCTTTATCTGCACTTAG-3′ and 5′-CTTACTGGATCCACTATTCTGGCA-3′, 5′-ACCTGTGGATCCTTTATCTGCACTTA-3′ and 5′-GCTATGGATCCGACCAGAATAAATTG-3′, and 5′-CCAGACTGGATCCGTAGATCAATTA-3′ and 5′-CTGAACTTACTGGATCCACTATTCT-3′, respectively. To construct the three site-directed mutants (icsA10, icsA14, and icsA15), reverse PCR was carried out with pMBG235 as a template and three different oligonucleotide primer pairs. pMAC210 (icsA10) was made with 5′-CTGGAATGATACAGATGGCGCTAGCCATG-3′ and 5′-TCCATGGCTAGCGCCAGCTGTAGCATTC-3′, thus creating NheI ends and resulting in the following three amino acid substitutions: Asp487 to Ala, Asp489 to Ala, and Asp491 to Ala. pMAC214 (icsA14) was made with 5′-GACTATACAGCTAGCTATATCAGTGAC-3′ and 5′-CTGATATAGCTAGCTGTATAGGCAATAG-3′ to create NheI ends and resulted in the following two amino acid substitutions: Asp467 to Ala and Lys470 to Ala. pMAC215 (icsA15) was generated with 5′-GTGACCAGAACGCGTTGATCTACGGTTT-3′ and 5′-CCGTAGATCAACGCGTTCTGGGCACTGA-3′ to generate MluI ends and resulted in the following two amino acid substitutions: Asp475 to Ala and Lys477 to Ala. The absence of the deleted segment or the presence of the point mutations in each case was confirmed by DNA sequencing.
To construct pMAC107 (icsA7), a region encompassing R. rickettsii ompA bp 2671 to 2868 was amplified by PCR with oligonucleotide primers 5′-GGAGCGGTGATTAGATCTACTACG-3′ and 5′-CTATATCCCCAGATCTTTGACTTAAC-3′, thereby generating BglII ends. This fragment was inserted into pMAC103 at the BamHI site in icsA3, thus resulting in insertion of rOmpA amino acids 868 to 933 after IcsA amino acid 507 with insertion of an Arg residue and a Ser residue. The presence of an insert was confirmed by restriction digestion and PCR analysis, and the orientation and nucleotide sequence were verified to be correct by DNA sequencing. pMAC108 (icsA8) and pMAC109 (icsA9) were made similarly, except with oligonucleotide primers 5′-CGACTAAAATAACGAGATCTGTGCA-3′ and 5′-GACTTAAAGATCTATTTAAATTTAACACA-3′. To construct pMBG282 (gst::icsA67), a fragment encompassing icsA bp 1897 to 2105 was amplified by PCR with oligonucleotide primers 5′-GCAGATAAAAGATCTACAGGTTTCAG-3′ and 5′-GCCAGAATAGTGAATTCAGTAAGTTC-3′, thus generating a BglII and an EcoRI end. This fragment was ligated into the cloning vector pGEX-2T (Pharmacia) at its BamHI and EcoRI sites. The resulting gene fusion codes for a 34-kDa protein (GST::IcsA67) consisting of glutathione S-transferase (GST) and IcsA amino acids 447 to 505 with a thrombin cleavage site (LVPRGS) between the two polypeptides and six additional amino acids at the carboxy terminus (EFIVTD). Stop codons in each of the three reading frames are located downstream of the coding sequence of the gene fusion. The fusion protein was expressed in E. coli BL21, and purification was carried out according to the manufacturer’s recommendations. DNA isolations, transformation, and cloning were performed by standard methods (38).
Protein preparation and analysis.
Whole-cell and supernatant extracts were prepared as described previously (1, 22). Protein extracts, resolved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis, were transferred to nitrocellulose and probed with the monoclonal antibody VIF8, which recognizes an epitope within the α domain of IcsA, or with affinity-purified IcsA antiserum (16). The VIF8 monoclonal antibody was synthesized according to standard procedures by using IcsA that had been purified as previously described (16). Visualization was performed by enhanced chemiluminescence (Amersham).
IcsA localization and cell infection assays.
Labeling for IcsA on the surface of bacteria grown in vitro was performed essentially as described previously (16). Labeling for IcsA and actin on S. flexneri-infected HeLa or L2 cell monolayers was performed essentially as previously described (16, 18). Plaque assays were performed essentially as described previously (42) with the exception that, where appropriate, ampicillin was added to the agarose overlay at a final concentration of 250 μg/ml. Monolayers were photographed 48 h after infection. Speeds of motile intracytoplasmic bacteria were determined as previously described (42).
Motility assays in Xenopus laevis egg cytoplasmic extracts.
Motility assays were performed as described previously (17). For competition studies, 8 μl of rhodamine-conjugated monomeric actin in G buffer (5 mM Tris [pH 8.0], 0.2 mM CaCl2, 0.2 mM ATP, 0.5 mM dithiothreitol [DTT]) was mixed with 3.2 μl of Kron buffer (0.05 mg of catalase per ml, 0.10 mg of glucose oxidase per ml, 2.5 mg of glucose per ml, 0.5 mM DTT). The following were then combined in a small Eppendorf tube: 5.5 μl of extracts; 0.7 μl of the rhodamine-conjugated actin mixture to a concentration of 40 μg/ml; 0.5 μl of phosphate-buffered saline (PBS), GST, or GST::IcsA67; and 0.5 μl of a suspension of formaldehyde-fixed bacteria in XB buffer (100 mM KCl, 0.1 mM CaCl2, 2 mM MgCl2, 5 mM EGTA, 10 mM K HEPES [pH 7.7], 50 mM sucrose). Following incubation on ice for 1 h, 0.5 μl of the assay mixture was spotted on a glass slide, covered with a 22- by 22-mm coverslip, sealed with nail polish, and examined by phase and fluorescence microscopy on a Nikon Diaphot 200 microscope with epifluorescence optics using standard rhodamine filters (Chroma, Brattleboro, Vt.). Between 250 and 350 individual bacteria were examined for each concentration of polypeptide per experiment and were evaluated for an association with polymerized actin in the form of clouds and tails.
RESULTS
Sequence similarity of IcsA amino acids 446 to 506 to rickettsial OmpA repeats.
IcsA has been shown to be anchored in the outer membrane by a domain within the carboxy-terminal 344 amino acids (β domain); as a result, the amino-terminal 706 amino acids of the mature protein (α domain) are exposed on the bacterial surface (45, 46) and thus constitute the region of the protein likely to be involved in interactions with host proteins. To aid in defining functional domains of IcsA, we compared the α domain of IcsA against the Swiss-Protein, EMBL, and GenBank databases. Sequence alignment reveals significant similarity of IcsA amino acids 446 to 506 to two discontinuous domains of the rickettsial outer membrane protein OmpA (Fig. 1B). Spotted-fever group Rickettsiae (R. rickettsii, R. conorii, and R. akari), like S. flexneri, form actin tails in the cytoplasm of cells, whereas typhus-group Rickettsiae do not. Since spotted-fever group Rickettsiae express OmpA and typhus-group Rickettsiae lack OmpA, it has been suggested that OmpA may have a role in rickettsial actin tail formation (19, 20). Significantly, the region that contains these two discontinuous domains is repeated 13 times in R. rickettsii OmpA, 14 times in R. conorii OmpA, and 11 times in R. akari OmpA (2, 14, 15). Within IcsA, the domain is not repeated. Of note, IcsA bears no sequence similarity to L. monocytogenes ActA.
FIG. 1.

Schematic of IcsA and sequence alignment of IcsA with the rickettsial homology domain. (A) Schematic of IcsA. SP and vertically striped bar, signal peptide; open bar, α domain; diagonally striped bar, β domain. (B) Sequence alignment of IcsA amino acids 450 to 498 with R. rickettsii OmpA amino acids 686 to 789. Dashes represent gaps introduced in the sequence to optimize alignment. (C) Sequence alignment of IcsA amino acids 446 to 506 with the three R. rickettsii OmpA domains exchanged for it in this study. Amino acids in boldface type are conserved between IcsA and the OmpA sequences. Amino acid designations for IcsA are those in GenBank accession no. M22802, and those for OmpA are those in GenBank accession no. M31227.
As shown in Fig. 1A, two discontinuous regions within the rickettsial OmpA repeat bear similarity to sequences within IcsA amino acids 446 to 506. One, OmpA amino acids 686 to 705, shares 50% sequence identity and 70% sequence similarity with IcsA amino acids 450 to 469; the other, OmpA amino acids 764 to 789, shares 34% sequence identity and 50% sequence similarity with IcsA amino acids 473 to 498.
We constructed a series of in-frame deletions that encompass all or portions of the coding region for the rickettsial homology domain of IcsA on plasmid pMBG235 (Fig. 2A). icsA3 contains a deletion that closely encompasses the rickettsial homology domain, while icsA16 and icsA17 contain deletions that roughly encompass the coding sequence for the amino-terminal and carboxy-terminal halves of the rickettsial homology domain, respectively. All icsA alleles constructed in this study contain the native icsA promoter, signal sequence, and coding region for the transmembrane domain.
FIG. 2.

Schematic of IcsA constructs made in this study. (A) Schema of internal in-frame deletions of IcsA. a, icsA WT, which encodes wild-type IcsA; b, icsA3, which encodes a deletion of amino acids 446 to 506, inclusive; c, icsA16, which encodes a deletion of amino acids 446 to 473, inclusive; d, icsA17, which encodes a deletion of amino acids 483 to 506, inclusive. Gaps represent deleted amino acids. Amino acid designations are those in GenBank accession no. M22802. SP and vertically striped bar, signal peptide; open bar, α domain; diagonally striped bar, β domain. (B) Alignment of IcsA amino acids 450 to 498 with the site-directed mutations within this domain. a, icsA WT, which encodes the wild-type amino acid sequence of IcsA (charged amino acids are underlined); b, icsA10, which encodes the amino acid substitutions Asp487 to Ala, Asp489 to Ala, and Asp491 to Ala; c, icsA14, which encodes the amino acid substitutions Asp467 to Ala and Lys470 to Ala; d, icsA15, which encodes the amino acid substitutions Asp475 to Ala and Lys477 to Ala. Alanine residues indicated in boldface type replace the charged residues underlined in line a.
Expression of icsA constructs in S. flexneri and E. coli grown in vitro.
IcsA is inserted into the outer membrane as a 120-kDa protein. Once in the outer membrane, it is slowly cleaved by IcsP (SopA) between Arg758 and Arg759, thereby releasing the 95-kDa α domain into the culture supernatant or host cell cytosol (10, 12, 13, 42). To determine whether the deletions within IcsA altered either the amount of IcsA present or cleavage by IcsP, protein samples prepared from approximately equivalent numbers of bacteria of each construct in both S. flexneri SC560 and E. coli MBG263 were analyzed by Western blotting.
As expected, S. flexneri strains M90T (wild type) and MAC1000 (icsA+ [i.e., the wild-type icsA allele on pMBG235]) exhibited strong bands corresponding to the full-length 120-kDa protein in whole-cell extracts (Fig. 3A, lanes 1 and 3) and the cleaved 95-kDa polypeptide in supernatant extracts (Fig. 3B, lanes 1 and 3). MAC1016 (icsA16) gave bands in both whole-cell extracts (Fig. 3A, lane 6) and supernatant extracts (Fig. 3B, lane 6) that were approximately as strong as those seen for M90T and MAC1000. Each of the other two constructs, MAC1003 (icsA3) and MAC1017 (icsA17), produced less protein in both whole-cell extracts (Fig. 3A, lanes 4 and 7) and supernatant extracts (Fig. 3B, lanes 4 and 7). For each construct, the apparent molecular masses of the observed bands correspond to the expected sizes of the deletion constructs. The relative faintness of the bands for MAC1003 and MAC1017 suggest that the mutant protein produced by alleles icsA3 and icsA17 might be slightly less stable than the native protein. To verify that the relative faintness of the signal was not due to partial loss of the epitope by the IcsA monoclonal antibody, Western blot analysis was also performed with a polyclonal antiserum and gave similar results (data not shown).
FIG. 3.

Expression of IcsA constructs in S. flexneri SC560 (A and B) and E. coli MBG263 (C). Western blot analysis of whole-cell extracts (A and C) and supernatant extracts (B) was performed with monoclonal antibody to IcsA. For each panel, approximately equivalent amounts of protein preparations were loaded in each lane. Apparent molecular masses are indicated, in kilodaltons.
When expressed in E. coli, the level of protein expression from each of the mutant alleles was no different than that from the wild-type allele (Fig. 3C, lanes 3, 5, and 6 versus lane 1), suggesting that polypeptides expressed from icsA3 and icsA17 might be more stable in E. coli than in S. flexneri. The apparent molecular mass of each of the mutant constructs was appropriate. Since IcsA is not cleaved from the surface of E. coli MBG263, we did not examine supernatant protein preparations from the constructs in these strains.
Localization of IcsA on the surface of S. flexneri and E. coli is not altered by deletions within the rickettsial homology domain of IcsA.
IcsA is distributed unipolarly on the surface of S. flexneri (16) and circumferentially on E. coli MBG263 (17). To assess whether the deletions involving the rickettsial homology domain affected surface localization of IcsA, indirect immunofluorescence was performed on each strain. IcsA is present on the surface of S. flexneri SC560 carrying the alleles icsA3, icsA16, and icsA17 and is in the same unipolar distribution that it is on wild-type S. flexneri (Fig. 4 and data not shown). For the strains carrying the icsA3, icsA16, and icsA17 alleles, up to 60 to 75% of the bacteria did not have detectable IcsA on the surface. Individual bacteria that had IcsA on the surface appeared qualitatively to have less surface IcsA than did individual wild-type organisms, as indicated by a less intense signal on indirect immunofluorescence (data not shown).
FIG. 4.

Localization of IcsA on the surface of S. flexneri cells expressing IcsA constructs. Indirect immunofluorescence (A, C, and E) and phase-contrast (B, D, and F) micrographs are shown. (A and B) M90T (WT). (C and D) MAC1000 (icsA WT). (E and F) MAC1003 (icsA3). Arrows indicate unipolar localization of IcsA on bacteria. MAC1016 (icsA16) and MAC1017 (icsA17) each had a labeling pattern that was indistinguishable from that of MAC1003 (data not shown).
The expression of the icsA alleles on the surface of E. coli MBG263 is shown in Fig. 5. As for E. coli carrying the wild-type allele of icsA (MAC2000) (Fig. 5A and B), E. coli carrying each of the mutant alleles displayed IcsA distributed circumferentially on the bacterial surface (Fig. 5 C to F and data not shown). In E. coli MBG263, more than 90% of the bacteria observed for each strain displayed IcsA on the surface. In addition, the intensity of IcsA on the surface of MAC2003 (icsA3), MAC2016 (icsA16), and MAC2017 (icsA17) was approximately the same as that on MAC2000 (icsA WT).
FIG. 5.

Localization of IcsA on the surface of E. coli cells expressing IcsA alleles. Indirect immunofluorescence (A, C, and E) and phase-contrast (B, D, and F) micrographs are shown. (A and B) MAC2000 (icsA WT). (C and D) MAC2003 (icsA3). (E and F) MAC2009 (icsA9). MAC2016 (icsA16) and MAC2017 (icsA17) each had a labeling pattern that was indistinguishable from that of MAC2000 (data not shown).
Deletion of the rickettsial homology domain of IcsA results in loss of plaque formation in HeLa and L2 cell monolayers.
The ability of a Shigella strain to enter and spread between mammalian cells can be assessed by its ability to form plaques on cell monolayers (34). Plaques result when S. flexneri successfully enter cells, multiply within them, and spread by actin-based motility into cells that are immediately adjacent and, subsequently, into the next layer of adjacent cells. SC560 (icsA), while capable of entering cells and dividing within them, is unable to recruit actin and does not form plaques (10). To evaluate whether MAC1003 (icsA3) could form plaques, HeLa or L2 cell monolayers were infected with this strain. Unlike MAC1000 (icsA WT) (Fig. 6A, panel c, and 6B, panel c) and wild-type strain M90T (Fig. 6A, panel a, and 6B, panel a), MAC1003 (icsA3) (Fig. 6A, panel d, and 6B, panel d) did not form plaques on HeLa or L2 cells, indicating that it was either unable to enter cells or, once within the cell, unable to spread into adjacent cells. Of note, the plaques formed by wild-type strain M90T are larger than those formed by MAC1000 (icsA WT) (Fig. 6A and B, panels a versus panels c).
FIG. 6.

Plaque formation of S. flexneri cells expressing IcsA constructs. (A) HeLa cell monolayers. a, M90T (WT); b, SC560 (icsA); c, MAC1000 (icsA WT); d, MAC1003 (icsA3). (B) L2 cell monolayers. a to d, same as in panel A; e, MAC1010 (icsA10). Plaques formed by MAC1014 (icsA14) and MAC1015 (icsA15) were not significantly different from those formed by MAC1010 (icsA10). Petri dishes were photographed 48 h after infection. Bar, 2 mm.
Deletions within the rickettsial homology domain of IcsA abrogate actin tail formation and actin-based motility in S. flexneri.
To determine whether MAC1003 (icsA3) did not form plaques because of impairment in actin-based motility, HeLa cells were infected with this strain and the association of intracellular bacteria with actin was characterized. As expected, both the wild-type strain M90T (Fig. 7A and B) and MAC1000 (icsA WT) elaborated actin tails (Fig. 7C and D) and formed cell surface protrusions (data not shown). In contrast, MAC1003 (icsA3) gained access to the cytoplasm, but was not associated with actin and did not form tails (Fig. 7E and F) or cell surface protrusions (data not shown). In addition, while both MAC1016 (icsA16) and MAC1017 (icsA17) also gained access to the cytoplasm, neither was associated with actin or formed actin tails in the cytoplasm of HeLa cells (data not shown).
FIG. 7.

Actin tail formation of S. flexneri expressing IcsA constructs. Rhodamine-phalloidin signal (A, C, and E) and phase-contrast (B, D, and F) micrographs are shown. (A and B) S. flexneri M90T (WT). (C and D) MAC1000 (icsA WT). (E and F) MAC1003 (icsA3). Arrows indicate bacteria with tails; arrowheads indicate bacteria lacking tails.
One possible explanation for the inability of MAC1003 (icsA3), MAC1016 (icsA16), and MAC1017 (icsA17) to associate with actin could be that the bacteria observed in the cytoplasm lacked surface IcsA. To ascertain whether the inability of MAC1003 (icsA3), MAC1016 (icsA16), and MAC1017 (icsA17) to recruit actin was attributable to reduced levels of surface IcsA, we evaluated the ability of the mutant constructs to recruit actin in the E. coli MBG263 background, since more than 90% of MBG263 cells carrying these alleles had levels of IcsA on their surfaces comparable to those on E. coli carrying the wild-type allele (icsA WT) (Fig. 5). We have previously shown that X. laevis egg cytoplasmic extracts support actin-based motility of IcsA-expressing E. coli MBG263 (15). Unlike MAC2000 (icsA WT), which moved at an average rate of 12.9 ± 7.8 μm/min in Xenopus egg cytoplasmic extracts (Fig. 8B), MAC2003 (icsA3), MAC1016 (icsA16), and MAC1017 (icsA17) did not associate with actin or move (Fig. 8A and data not shown).
FIG. 8.

Motility assay of E. coli MBG263 carrying IcsA constructs. Phase-contrast time-lapse images in Xenopus egg cytoplasmic extracts of MAC2003 (icsA3) (A) and MAC2000 (icsA WT) (B) are shown. The actin tail is seen as seen as a phase-lucent streak behind moving MAC2000. Sequential frames are separated by 30-s intervals. Images of MAC2016 (icsA16) and MAC2017 (icsA17) were indistinguishable from those of MAC2003.
Deletion of the rickettsial homology domain of IcsA leads to relative unmasking of IcsA within the host cell cytosol.
As mentioned above, a possible explanation for the inability of MAC1003 (icsA3), MAC1016 (icsA16), and MAC1017 (icsA17) to associate with actin in the cytosol would be a lack of IcsA on the surface of these strains in this setting. We therefore labeled MAC1003-infected cells for IcsA by using affinity-purified polyclonal antiserum. Approximately 70 to 75% of the observed intracellular bacteria displayed fluorescent label. Surprisingly, the signal was consistently more intense on MAC1003 (icsA3) (Fig. 9G and H) than on wild-type M90T (Fig. 9A and B) or MAC1000 (icsA WT) (Fig. 9E and F). This suggests that the mutant IcsA on MAC1003 was more accessible to the antibody than native IcsA on M90T or MAC1000 (icsA WT). Consistent with this, the signal from IcsA+ extracellular bacteria labeled with the same antiserum is significantly more intense than that from intracellular bacteria of any of the strains examined (data not shown).
FIG. 9.

Immunolabeling of IcsA on intracellular S. flexneri strains carrying IcsA alleles. Indirect immunofluorescence micrographs (A, C, E, and G), with affinity-purified polyclonal antiserum to IcsA, and phase-contrast micrographs (B, D, F, and H) are shown. (A and B) M90T (WT). (C and D) SC560 (icsA). (E and F) MAC100 (icsA WT). (G and H) MAC1003 (icsA3). The apparent output of the immunofluorescence signal was normalized for panels A, C, E, and G by setting the background nonspecific signals to equivalent levels. Arrows indicate intense signal from IcsA on MAC1003; arrowheads indicate significantly weaker signal from IcsA on M90T and MAC1000.
Replacement of IcsA amino acids 446 to 506 with the rickettsial repeat domain does not restore actin tail assembly.
To examine whether the rickettsial repeat domain that bears sequence similarity to IcsA amino acids 446 to 506 could functionally complement the similar sequence of IcsA, the coding sequence of each of three distinct repeat sequences from R. rickettsii OmpA was exchanged for that for amino acids 446 to 506 of IcsA, creating alleles icsA7, icsA8, and icsA9 (Fig. 1C).
The rickettsial repeats exchanged for IcsA amino acids 446 to 506 differ in length (Fig. 1C); icsA7, icsA8, and icsA9 are predicted to encode fusion proteins of 122, 126, and 120 kDa, respectively. On Western blot analysis, the S. flexneri strains carrying these alleles, strains MAC1007 (icsA7), MAC1008 (icsA8), and MAC1009 (icsA9), exhibited bands corresponding to the predicted masses of the fusion proteins that were fainter than the bands corresponding to wild-type IcsA (Fig. 3A and B, lane 5, and data not shown versus Fig. 3A and B, lanes 1 and 3). For each of these constructs in S. flexneri, a significantly lower percentage of bacteria expressed IcsA on the surface than for MAC1000 (icsA WT). None of these constructs in S. flexneri formed plaques on HeLa or L2 cell monolayers or associated with actin in the cytoplasm of infected HeLa cells (data not shown).
The E. coli MBG263 strains carrying these alleles, MAC2007 (icsA7), MAC2008 (icsA8), and MAC2009 (icsA9), exhibited bands corresponding to the predicted masses of the fusion proteins that were of the same approximate intensity as the bands corresponding to wild-type IcsA (Fig. 3C, lane 4, and data not shown versus Fig. 3C, lanes 1 and 3). Each of these strains expressed IcsA on the bacterial surface to the same extent and in the same circumferential distribution as did MAC2000 (icsA WT). However, none either associated with actin or moved in Xenopus egg cytoplasmic extracts (data not shown).
To evaluate potentially critical contact residues within IcsA amino acids 446 to 506, clusters of charged residues, which are likely to be exposed on the surface of the protein and therefore to be involved in protein-protein interactions, were identified and subjected to site-directed mutagenesis. This strategy has been successfully applied in a number of cases (3, 51). Three sets of mutations were introduced in the region (Fig. 2B): the first replaced amino acids Asp487, Asp489, and Asp491 with alanines (giving icsA10); the second replaced Asp467 and Lys470 with alanines (icsA14); and the third replaced Asp475 and Lys478 with alanines (icsA15). S. flexneri strains carrying the resultant icsA alleles were no different from S. flexneri carrying the wild-type allele (MAC1000) in any of the assays performed: they expressed protein at wild-type levels on Western blot analysis, expressed IcsA on the surface of approximately the same percentage of bacteria and at approximately the same intensity, formed plaques on HeLa and L2 cell monolayers that were not significantly different from MAC100 (Fig. 6B, panel c versus panel e), formed actin tails in HeLa cells, and moved in the cytoplasm of HeLa cells at speeds comparable to MAC1000 (icsA WT) (data not shown).
Competition of the rickettsial homology domain polypeptide in actin recruitment assays.
To further examine the role of IcsA amino acids 446 to 506 in actin-based motility, a polypeptide that consists of GST fused to amino acid residues 447 to 505 of IcsA (GST::IcsA67) was used in competition assays with MAC2000 (icsA WT) in the Xenopus egg cytoplasmic extracts. The polypeptide is greater than 90% pure (Fig. 10). As shown in Table 2, the fusion protein had no significant effect on either (i) the percentage of bacteria associated with actin clouds, which are accumulations of polymerized actin that is not organized into a bundle, or actin tails or (ii) the percentage of bacteria assembling actin tails. Further, the IcsA-derived peptide alone (after removal of GST) had no effect on actin polymerization in these assays (unpublished data).
FIG. 10.
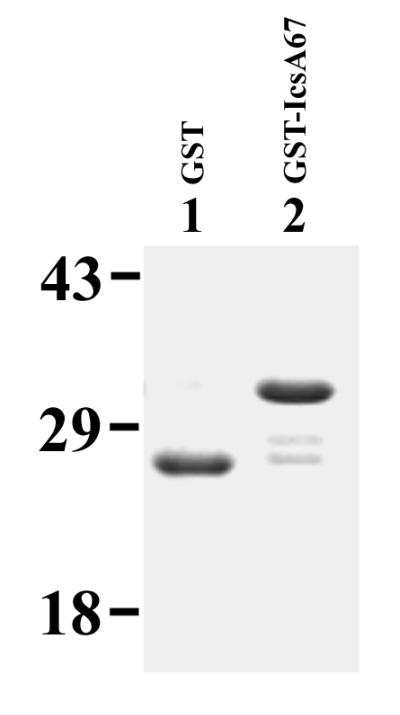
Purified IcsA rickettsial homology domain used in competition assays. A Coomassie-stained sodium dodecyl sulfate-polyacrylamide gel of GST (lane 1) and GST::IcsA67 (lane 2) is shown. Apparent molecular masses are indicated, in kilodaltons.
TABLE 2.
Competition assay with IcsA rickettsial homology domain polypeptide fused to GST
| Competitor | Concn (μM) | Accumulation of polymerized actina | Assembly of actin tailb |
|---|---|---|---|
| GST::IcsA67 | 35 | 55 ± 16 | 22 ± 8.3 |
| GST::IcsA67 | 0.35 | 66 ± 12 | 18 ± 8.4 |
| GST::IcsA67 | 0.035 | 71 ± 15 | 12 ± 8.6 |
| GST | 35 | 34 ± 23 | 17 ± 15 |
| PBS | 73 ± 7.6 | 24 ± 7.4 |
Values are percentages (mean ± standard deviation) of bacteria associated with actin clouds or actin tails in six separate experiments.
Values are percentages (mean ± standard deviation) of bacteria with actin tails in six separate experiments.
DISCUSSION
Shigella spp., like L. monocytogenes, spotted-fever group Rickettsiae, and vaccinia virus, are able to recruit host cell monomeric actin into a tail that consists of filamentous actin bundled in a parallel array, with the barbed end of the filaments uniformly directed towards the bacterium. Monomeric actin is incorporated into the filaments at the junction of the bacterium with the tail (47). The mechanism of actin tail formation can therefore be presumed to involve recruitment of monomeric actin to the bacterial pole, nucleation of this actin, orientation of the nuclei such that the barbed end faces the bacterium, polymerization from the barbed end while protecting it from barbed end capping proteins, and bundling of the filament into a parallel array. We have previously shown that the S. flexneri outer membrane protein IcsA is located unipolarly on the bacterium at the site of the forming tail (16), and we and others have shown that IcsA expressed in E. coli is sufficient to permit actin tail formation in cytoplasmic extracts (17, 26). Thus, IcsA is able to induce the specific series of steps outlined above.
In this work, we have identified a region of IcsA that is necessary for the interaction of IcsA with host cytoskeletal factors involved in actin assembly on S. flexneri. Deletion of the domain of amino acid residues 446 to 506 totally eliminates actin-based motility of either S. flexneri or E. coli strains that express the IcsA deletion construct. The mutant Shigella strain does not form actin tails or cell surface protrusions, does not recruit actin into a cloud at the bacterial surface, and does not form plaques on cell monolayers, indicating an inability to spread from cell to cell. Most striking, however, is the relative unmasking of the IcsA on the surface of S. flexneri strains expressing the mutant allele when in the cytoplasm of infected cells (Fig. 9). This suggests that native IcsA is made relatively inaccessible to antibody by its interactions with host cell factors, while the mutant allele is accessible by virtue of few or no such interactions.
These observations suggest that the deletion of amino acids 446 to 506 of IcsA either (i) leads to loss of a domain that directly interacts with key host cell factors or (ii) disrupts the native conformation of IcsA so as to impair the interaction of the IcsA domain that binds key host cell factors with those factors. On the basis of data presented here, we favor the latter. First, with the exception of the IcsA construct encoded by icsA16, all of the IcsA deletion constructs examined in this study are less stably expressed in S. flexneri and presented in lower amounts of the bacterial surface (Fig. 3 to 5). Second, alanine mutagenesis of clusters of charged residues within the region, which would be predicted to be on the surface of the protein in this region and therefore likely to interact with outer proteins, resulted in maintenance of the wild-type phenotype. Third, each of the small nonoverlapping deletions within amino acid residues 446 to 506 encoded by icsA16 and icsA17 has the phenotype of the larger deletion, which suggests that the intact rickettsial homology domain may be required for proper spacing or proximity of other domains. And finally, in vitro competition of a polypeptide of the region fused to GST did not alter the motility of the wild-type construct (Table 2). These data provide further refinement on the large region, encompassing amino acids 320 to 507, that was reported by Suzuki et al. to be required for actin recruitment (46).
IcsA is thought to cross the bacterial cytoplasmic membrane via the Sec secretion machinery, and subsequent translocation of the α domain of IcsA to the bacterial surface has been proposed to occur through a pore formed by 13 passages of the β domain through the outer membrane (45). We observed a moderate decrease in the surface presentation of most of the mutant deletion constructs expressed in S. flexneri. Notably, when expressed in E. coli, the amount of IcsA on the bacterial surface was not detectably decreased from those for native IcsA: IcsA was nevertheless unable to induce actin-based motility. The differences in levels of expression on the bacterial surface between the S. flexneri background and the E. coli background suggest that the mechanism of translocation of IcsA to the bacterial surface may differ in S. flexneri and E. coli.
Despite the sequence similarity between IcsA amino acids 446 to 506 and the repetitive domains of the rickettsial OmpA proteins, exchange of these domains did not yield a functional form of IcsA. One possible explanation for this is that the exchange of the rickettsial domain did not reconstitute the protein to its native conformation, as suggested by the observation that the levels of expression on the bacterial surface and the level of protein in whole-cell preparations did not return to those seen with wild-type IcsA. The presence of homologous domains in rickettsial OmpA suggests that the function served by these domains may be generalizable among a group of outer membrane proteins. How the function of the multiple copies of the domain that are present in rickettsial OmpA would differ from the function of the single copy that is present in IcsA is unclear.
ACKNOWLEDGMENTS
We thank T. Hackstadt for providing R. rickettsii genomic DNA, T. R. Evans for providing X. laevis eggs, J. Condeelis for helpful discussions and providing rhodamine-conjugated monomeric actin, S. Almo for helpful discussions and critical review of the manuscript, and the Analytical Imaging Facility and the DNA Sequencing Facility at the Albert Einstein College of Medicine for technical assistance.
This work was supported by NIH grants GM16654 (M.C.), AI35817 (M.B.G.), and AI36929 (J.A.T.); the Pew Scholars Program in the Biomedical Sciences (M.B.G.); and Established Investigator and Grant-in-Aid awards from the American Heart Association (M.B.G.).
REFERENCES
- 1.Allaoui A, Mounier J, Prevost M C, Sansonetti P J, Parsot C. icsB: a Shigella flexneri virulence gene necessary for the lysis of protrusions during intercellular spread. Mol Microbiol. 1992;6:1605–1616. doi: 10.1111/j.1365-2958.1992.tb00885.x. [DOI] [PubMed] [Google Scholar]
- 2.Anderson B E, McDonald G A, Jones D C, Regnery R L. A protective protein antigen of Rickettsia rickettsii has tandemly repeated, near-identical sequences. Infect Immun. 1990;58:2760–2769. doi: 10.1128/iai.58.9.2760-2769.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bass S H, Mulkerrin M G, Wells J A. A systematic mutational analysis of hormone-binding determinants in the human growth hormone receptor. Proc Natl Acad Sci USA. 1991;88:4498–4502. doi: 10.1073/pnas.88.10.4498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bernardini M L, Mounier J, d’Hauteville H, Coquis-Rondon M, Sansonetti P J. Identification of icsA, a plasmid locus of Shigella flexneri that governs bacterial intra- and intercellular spread through interaction with F-actin. Proc Natl Acad Sci USA. 1989;86:3867–3871. doi: 10.1073/pnas.86.10.3867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Chakraborty T, Ebel F, Domann E, Niebuhr K, Gerstel B, Pistor S, Temm-Grove C J, Jockusch B M, Reinhard M, Walter U, Wehland J. A focal adhesion factor directly linking intracellularly motile Listeria monocytogenes and Listeria ivanovii to the actin-based cytoskeleton of mammalian cells. EMBO J. 1995;14:1314–1321. doi: 10.1002/j.1460-2075.1995.tb07117.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Clerc P, Sansonetti P J. Entry of Shigella flexneri into HeLa cells: evidence for directed phagocytosis involving actin polymerization and myosin accumulation. Infect Immun. 1987;55:2681–2688. doi: 10.1128/iai.55.11.2681-2688.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Cudmore S, Cossart P, Griffiths G, Way M. Actin-based motility of vaccinia virus. Nature. 1995;378:636–638. doi: 10.1038/378636a0. [DOI] [PubMed] [Google Scholar]
- 8.Dabiri G A, Sanger J M, Portnoy D A, Southwick F S. Listeria monocytogenes moves rapidly through the host-cell cytoplasm by inducing directional actin assembly. Proc Natl Acad Sci USA. 1990;87:6068–6072. doi: 10.1073/pnas.87.16.6068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.d’Hauteville H, Sansonetti P J. Phosphorylation of IcsA by cAMP-dependent protein kinase and its effect on intracellular spread of Shigella flexneri. Mol Microbiol. 1992;6:833–841. doi: 10.1111/j.1365-2958.1992.tb01534.x. [DOI] [PubMed] [Google Scholar]
- 10.d’Hauteville H, Dufourcq Lagelouse R, Nato F, Sansonetti P J. Lack of cleavage of IcsA in Shigella flexneri causes aberrant movement and allows demonstration of a cross-reactive eukaryotic protein. Infect Immun. 1996;64:511–517. doi: 10.1128/iai.64.2.511-517.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Domann E, Wehland J, Rohde M, Pistor S, Hartl M, Goebel W, Leimeister-Wachter M, Wuenscher M, Chakraborty T. A novel bacterial virulence gene in Listeria monocytogenes required for host cell microfilament interaction with homology to the proline-rich region of vinculin. EMBO J. 1992;11:1981–1990. doi: 10.1002/j.1460-2075.1992.tb05252.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Egile C, d’Hauteville H, Parsot C, Sansonetti P J. SopA, the outer membrane protease responsible for polar localization of IcsA in Shigella flexneri. Mol Microbiol. 1997;23:1063–1073. doi: 10.1046/j.1365-2958.1997.2871652.x. [DOI] [PubMed] [Google Scholar]
- 13.Fukuda I, Suzuki T, Munakata H, Hayashi N, Katayama E, Yoshikawa M, Sasakawa C. Cleavage of Shigella surface protein VirG occurs at a specific site, but the secretion is not essential for intracellular spreading. J Bacteriol. 1995;177:1719–1726. doi: 10.1128/jb.177.7.1719-1726.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gilmore R D, Jr, Hackstadt T. DNA polymorphism in the conserved 190 kDa antigen gene repeat region among spotted fever group Rickettsiae. Biochim Biophys Acta. 1991;1097:77–80. doi: 10.1016/0925-4439(91)90027-7. [DOI] [PubMed] [Google Scholar]
- 15.Gilmore R D., Jr Comparison of the rompA gene repeat regions of Rickettsiae reveals species-specific arrangements of individual repeating units. Gene. 1993;125:97–102. doi: 10.1016/0378-1119(93)90752-o. [DOI] [PubMed] [Google Scholar]
- 16.Goldberg M B, Barzu O, Parsot C, Sansonetti P J. Unipolar localization and ATPase activity of IcsA, a Shigella flexneri protein involved in intracellular movement. J Bacteriol. 1993;175:2189–2196. doi: 10.1128/jb.175.8.2189-2196.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Goldberg M B, Theriot J A. Shigella flexneri surface protein IcsA is sufficient to direct actin-based motility. Proc Natl Acad Sci USA. 1995;92:6572–6576. doi: 10.1073/pnas.92.14.6572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Goldberg M B. Shigella actin-based motility in the absence of vinculin. Cell Motil Cytoskelet. 1997;37:44–53. doi: 10.1002/(SICI)1097-0169(1997)37:1<44::AID-CM5>3.0.CO;2-H. [DOI] [PubMed] [Google Scholar]
- 19.Hackstadt T. The biology of Rickettsiae. Infect Agents Dis. 1996;5:127–143. [PubMed] [Google Scholar]
- 20.Heinzen R A, Hayes S F, Peacock M G, Hackstadt T. Directional actin polymerization associated with spotted fever group Rickettsia infection of Vero cells. Infect Immun. 1993;61:1926–1935. doi: 10.1128/iai.61.5.1926-1935.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.High N, Mounier J, Prevost M C, Sansonetti P J. IpaB of Shigella flexneri causes entry into epithelial cells and escape from the phagocytic vacuole. EMBO J. 1992;11:1991–1999. doi: 10.1002/j.1460-2075.1992.tb05253.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hovde C J, Calderwood S B, Mekalanos J J, Collier R J. Evidence that glutamic acid 167 is an active-site residue of Shiga-like toxin I. Proc Natl Acad Sci USA. 1988;85:2568–2572. doi: 10.1073/pnas.85.8.2568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kadurugamuwa J L, Rohde M, Wehland J, Timmis K N. Intercellular spread of Shigella flexneri through a monolayer mediated by membranous protrusions and associated with reorganization of the cytoskeletal protein vinculin. Infect Immun. 1991;59:3463–3471. doi: 10.1128/iai.59.10.3463-3471.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kang F, Laine R O, Bubb M R, Southwick F S, Purich D L. Profilin interacts with the Gly-Pro-Pro-Pro-Pro-Pro sequences of vasodilator-stimulated phosphoprotein (VASP): implications for actin-based Listeria motility. Biochemistry. 1997;36:8384–8392. doi: 10.1021/bi970065n. [DOI] [PubMed] [Google Scholar]
- 25.Kocks C, Gouin E, Tabouret M, Berche P, Ohayon H, Cossart P. L. monocytogenes-induced actin assembly requires the actA gene product, a surface protein. Cell. 1992;68:521–531. doi: 10.1016/0092-8674(92)90188-i. [DOI] [PubMed] [Google Scholar]
- 26.Kocks C, Marchand J-B, Gouin E, d’Hauteville H, Sansonetti P J, Carlier M-F, Cossart P. The unrelated surface proteins ActA of Listeria monocytogenes and IcsA of Shigella flexneri are sufficient to confer actin-based motility on Listeria innocua and Escherichia coli respectively. Mol Microbiol. 1995;18:413–423. doi: 10.1111/j.1365-2958.1995.mmi_18030413.x. [DOI] [PubMed] [Google Scholar]
- 27.LaBrec E, Schneider H, Magani T, Formal S. Epithelial cell penetration as an essential step in the pathogenesis of bacillary dysentery. J Bacteriol. 1964;88:1503–1518. doi: 10.1128/jb.88.5.1503-1518.1964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lasa I, David V, Gouin E, Marchand J B, Cossart P. The amino-terminal part of ActA is critical for the actin-based motility of Listeria monocytogenes: the central proline-rich region acts as a stimulator. Mol Microbiol. 1995;18:425–436. doi: 10.1111/j.1365-2958.1995.mmi_18030425.x. [DOI] [PubMed] [Google Scholar]
- 29.Lasa I, Gouin E, Goethals M, Vancompernolle K, David V, Vandekerckhove J, Cossart P. Identification of two regions in the N-terminal domain of ActA involved in the actin comet tail formation by Listeria monocytogenes. EMBO J. 1997;16:1531–1540. doi: 10.1093/emboj/16.7.1531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Lett M C, Sasakawa C, Okada N, Sakai T, Makino S, Yamada M, Komatsu K, Yoshikawa M. virG, a plasmid-coded virulence gene of Shigella flexneri: identification of the VirG protein and determination of the complete coding sequence. J Bacteriol. 1989;171:353–359. doi: 10.1128/jb.171.1.353-359.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Meissner P S, Sisk W P, Berman M L. Bacteriophage lambda cloning system for the construction of directional cDNA libraries. Proc Natl Acad Sci USA. 1987;84:4171–4175. doi: 10.1073/pnas.84.12.4171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Mounier J, Ryter A, Coquis-Rondon M, Sansonetti P J. Intracellular and cell-to-cell spread of Listeria monocytogenes involves interaction with F-actin in the enterocytelike cell line Caco-2. Infect Immun. 1990;58:1048–1058. doi: 10.1128/iai.58.4.1048-1058.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Niebuhr K, Ebel F, Frank R, Reinhard M, Domann E, Carl U D, Walter U, Gertler F B, Wehland J, Chakraborty T. A novel proline-rich motif present in ActA of Listeria monocytogenes and cytoskeletal proteins is the ligand for the EVH1 domain, a protein module present in the Ena/VASP family. EMBO J. 1997;16:5433–5444. doi: 10.1093/emboj/16.17.5433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Oaks E V, Wingfield M E, Formal S B. Plaque formation by virulent Shigella flexneri. Infect Immun. 1985;48:124–129. doi: 10.1128/iai.48.1.124-129.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Pal T, Newland J W, Tall B D, Formal S B, Hale T L. Intracellular spread of Shigella flexneri associated with the kcpA locus and a 140-kilodalton protein. Infect Immun. 1989;57:477–486. doi: 10.1128/iai.57.2.477-486.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Pistor S, Chakraborty T, Walter U, Wehland J. The bacterial actin nucleator protein ActA of Listeria monocytogenes contains multiple binding sites for host microfilament proteins. Curr Biol. 1995;5:517–525. doi: 10.1016/s0960-9822(95)00104-7. [DOI] [PubMed] [Google Scholar]
- 37.Prévost M C, Lesourd M, Arpin M, Vernel F, Mounier J, Hellio R, Sansonetti P J. Unipolar reorganization of F-actin layer at bacterial division and bundling of actin filaments by plastin correlate with movement of Shigella flexneri within HeLa cells. Infect Immun. 1992;60:4088–4099. doi: 10.1128/iai.60.10.4088-4099.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sambrook J, Fritsch E F, Maniatis T. Molecular cloning: a laboratory manual. 2nd ed. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory Press; 1989. [Google Scholar]
- 39.Sansonetti P J, Kopecko D J, Formal S B. Involvement of a plasmid in the invasive ability of Shigella flexneri. Infect Immun. 1982;35:852–860. doi: 10.1128/iai.35.3.852-860.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Sansonetti P J, Ryter A, Clerc P, Maurelli A T, Mounier J. Multiplication of Shigella flexneri within HeLa cells: lysis of the phagocytic vacuole and plasmid-mediated contact hemolysis. Infect Immun. 1986;51:461–469. doi: 10.1128/iai.51.2.461-469.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Sansonetti P J, Mounier J, Prevost M C, Mege R M. Cadherin expression is required for the spread of Shigella flexneri between epithelial cells. Cell. 1994;76:829–839. doi: 10.1016/0092-8674(94)90358-1. [DOI] [PubMed] [Google Scholar]
- 42.Shere K D, Sallustio S, Manessis A, D’Aversa T G, Goldberg M B. Disruption of IcsP, the major Shigella protease that cleaves IcsA, accelerates actin-based motility. Mol Microbiol. 1997;25:451–462. doi: 10.1046/j.1365-2958.1997.4681827.x. [DOI] [PubMed] [Google Scholar]
- 43.Smith G A, Theriot J A, Portnoy D A. The tandem repeat domain in the Listeria monocytogenes ActA protein controls the rate of actin-based motility, the percentage of moving bacteria, and the localization of vasodilator-stimulated phosphoprotein and profilin. J Cell Biol. 1996;135:647–660. doi: 10.1083/jcb.135.3.647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Studier F W, Moffatt B A. Use of bacteriophage T7 RNA polymerase to direct selective high-level expression of cloned genes. J Mol Biol. 1986;189:113–130. doi: 10.1016/0022-2836(86)90385-2. [DOI] [PubMed] [Google Scholar]
- 45.Suzuki T, Lett M C, Sasakawa C. Extracellular transport of VirG protein in Shigella. J Biol Chem. 1995;270:30874–30880. doi: 10.1074/jbc.270.52.30874. [DOI] [PubMed] [Google Scholar]
- 46.Suzuki T, Saga S, Sasakawa C. Functional analysis of Shigella VirG domains essential for interaction with vinculin and actin-based motility. J Biol Chem. 1996;271:21878–21885. doi: 10.1074/jbc.271.36.21878. [DOI] [PubMed] [Google Scholar]
- 47.Theriot J A, Mitchison T J, Tilney L G, Portnoy D A. The rate of actin-based motility of intracellular Listeria monocytogenes equals the rate of actin polymerization. Nature. 1992;357:257–260. doi: 10.1038/357257a0. [DOI] [PubMed] [Google Scholar]
- 48.Tilney L G, Portnoy D A. Actin filaments and the growth, movement, and spread of the intracellular bacterial parasite Listeria monocytogenes. J Cell Biol. 1989;109:1597–1608. doi: 10.1083/jcb.109.4.1597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Welch M D, Iwamatsu A, Mitchison T J. Actin polymerization is induced by Arp2/3 protein complex at the surface of Listeria monocytogenes. Nature. 1997;385:265–269. doi: 10.1038/385265a0. [DOI] [PubMed] [Google Scholar]
- 50.Welch M D, Rosenblatt J, Skoble J, Portnoy D A, Mitchison T J. Interaction of human Arp2/3 complex and the Listeria monocytogenes ActA protein in actin filament elongation. Science. 1998;281:105–108. doi: 10.1126/science.281.5373.105. [DOI] [PubMed] [Google Scholar]
- 51.Wertman K F, Drubin D G, Botstein D. Systematic mutational analysis of the yeast ACT1 gene. Genetics. 1992;132:337–350. doi: 10.1093/genetics/132.2.337. [DOI] [PMC free article] [PubMed] [Google Scholar]