

Abstract
The recent emergence of multiple SARS‐CoV‐2 variants has caused considerable concern due to both reduced vaccine efficacy and escape from neutralizing antibody therapeutics. It is, therefore, paramount to develop therapeutic strategies that inhibit all known and future SARS‐CoV‐2 variants. Here, we report that all SARS‐CoV‐2 variants analyzed, including variants of concern (VOC) Alpha, Beta, Gamma, Delta, and Omicron, exhibit enhanced binding affinity to clinical grade and phase 2 tested recombinant human soluble ACE2 (APN01). Importantly, soluble ACE2 neutralized infection of VeroE6 cells and human lung epithelial cells by all current VOC strains with markedly enhanced potency when compared to reference SARS‐CoV‐2 isolates. Effective inhibition of infections with SARS‐CoV‐2 variants was validated and confirmed in two independent laboratories. These data show that SARS‐CoV‐2 variants that have emerged around the world, including current VOC and several variants of interest, can be inhibited by soluble ACE2, providing proof of principle of a pan‐SARS‐CoV‐2 therapeutic.
Keywords: COVID‐19, treatment, clinical trial, vaccine
Subject Categories: Microbiology, Virology & Host Pathogen Interaction
Recombinant human ACE2 is reported as a “universal” therapeutic approach, exhibiting strong potency and efficacy for the inhibition of SARS‐CoV‐2 infection, especially towards current variants of concern.
The paper explained.
Problem
SARS‐CoV‐2 infections caused an unprecedented global pandemic with millions of fatalities and morbidities. While great progress has been made in vaccinations, drugs for the treatment of COVID‐19, including antibodies, convalescent sera, and even small molecule‐based therapeutics, are inherently vulnerable to viral escape mutations. Consequently, there is a large unmet medical need for therapeutics that can act on all current and future SARS‐CoV‐2 variants to limit the progression of COVID‐19.
Results
In this study, we provide evidence for the therapeutic potential of soluble recombinant human ACE2 as a pan‐SARS‐CoV‐2 molecular decoy therapeutic approach. We systematically tested the binding of SARS‐CoV‐2 variants with a focus on variants of concern to recombinant soluble human ACE2 and provide evidence for an increase in binding affinity and avidity for all tested variants of concern. This increased binding affinity correlates with a substantially enhanced potency of the soluble ACE2 decoy toward all variants of concern, including Delta and Omicron, when compared to the original viral isolate of SARS‐CoV‐2. Data from an independent laboratory confirm these findings.
Impact
The efficacy of soluble ACE2 toward all tested variants of concern, including the delta and omicron sub‐lineages, highlights the therapeutic potential of ACE2 a universal strategy to treat potentially all current and future SARS‐CoV‐2 variants and additional coronaviruses that use ACE2 as a cell entry receptor. Clinical studies on the safety and tolerability of soluble ACE2 inhalation are currently ongoing to provide the basis to test its efficacy to prevent and treat SARS‐CoV‐2 infections, an approach that should be feasible for a large population.
Introduction
The emergence of SARS‐CoV‐2 has resulted in an unprecedented COVID‐19 pandemic with dire economic, social, and health consequences for hundreds of millions of people. The initial step of SARS‐CoV‐2 infection is binding of the viral Spike protein to Angiotensin‐converting enzyme 2 (ACE2) (Shang et al, 2020; Wang et al, 2020; Zhou et al, 2020), followed by proteolytic processing of the trimeric Spike (Benton et al, 2020; Hoffmann et al, 2020) and subsequent infection of target cells (Hu et al, 2021). Inhibition of Spike/ACE2 interaction is the fundamental principle for the activity of neutralizing antibodies induced by all current vaccines (Kyriakidis et al, 2021). Similarly, approved monoclonal antibodies act by blocking the interaction of the cell entry receptor ACE2 and the viral Spike protein (see https://www.covid19treatmentguidelines.nih.gov/ for further information). Thus, blocking Spike/ACE2 binding has become a central strategy of both vaccine design and multiple therapeutic approaches including ACE2‐based therapeutics (Linsky et al, 2020; preprint: Svilenov et al, 2020; preprint: Hassler et al, 2021; Higuchi et al, 2021; Tanaka et al, 2021). This has created an intense research focus on the molecular details of these processes, thereby making the Spike/ACE2 interaction one of the best validated drug targets in medicine.
Both vaccines and antibody therapeutics have had an enormous impact and are a remarkable testament to the rapid translatability of basic research. However, although coronaviruses mutate less frequently, as compared to viruses like influenza, many variants of SARS‐CoV‐2 have emerged throughout the pandemic (Banerjee et al, 2021; Harvey et al, 2021) (see also WHO and CDC resources online). Some of these variants have been designated as variants of concern (VOCs) by the WHO because of their increased infectivity and transmissibility. Mutations in the viral Spike protein seem to be of particular relevance in this respect. These mutations do not only affect the infectivity and transmissibility of SARS‐CoV‐2, but also reduce the potency of vaccines, convalescent sera, and monoclonal antibody therapeutics (Harvey et al, 2021; Planas et al, 2021; Garcia‐Beltran et al, 2021; Cele et al, 2021; Greaney et al, 2021; Lopez Bernal et al, 2021; Jangra et al, 2021; Takashita et al, 2022; Zhou et al, 2022; Peng et al, 2022). The recent emergence of the Delta and Omicron variants and symptomatic and sometimes even severe infections of doubly vaccinated people (Bergwerk et al, 2021; Farinholt et al, 2021; Christensen et al, 2022; Kuhlmann et al, 2022) is one such example, and more such variants will possibly develop. It remains to be determined whether increased natural immunity, mass vaccinations, and the increased application of therapeutic monoclonal antibodies and antiviral therapeutics will affect or drive further viral evolution. To prevent further severe disruptions to life and economies due to SARS‐CoV‐2, it is therefore paramount to design universal strategies for the prevention and treatment of current VOCs and possibly even to variants that will emerge in future.
Here, we report that soluble ACE2 (APN01), already being tested in clinical trials (NCT04335136), binds receptor‐binding domain (RBD) and full‐length Spike proteins of SARS‐CoV‐2 variants and especially VOCs with increased affinity when compared to the SARS‐CoV‐2 reference strain Spike and effectively neutralizes infections of all tested variants. Clinical Phase I testing of an inhalation approach, ultimately aiming at directly neutralizing SARS‐CoV‐2 at its site of entry, is currently underway (NCT05065645). Since Spike/ACE2 interaction is the crucial first step of viral infection, the viral Spike cannot mutate to escape ACE2 binding, without a loss in infectivity and tissue tropism. Our data provide the blueprint for a universal anti‐COVID‐19 agent with the potential to treat or even prevent infections against all current and potentially also future SARS‐CoV‐2 variants, as well as novel emerging coronaviruses using ACE2 as cell entry receptor.
Results
Enhanced affinity of clinical grade ACE2 to the Spike RBDs of emerging SARS‐CoV‐2 variants
Viral evolution of SARS‐CoV‐2 has been demonstrated to be focused on the Spike protein, which is instrumental for the early steps of viral infection (Pereson et al, 2021; Rochman et al, 2021). Many single or compound mutations, especially in the RBD of the viral Spike, have been described and either hypothesized or demonstrated to affect binding to the cell entry receptor ACE2 (see Harvey et al, 2021 for a review). To systematically test whether these emerged variants affect Spike/ACE2 interactions, we selected viral variants that have been described in the literature and in various databases. The RBD variants analyzed in this study and the location(s) of the respective mutations are depicted in a 3D model of the viral Spike RBD (Fig 1A). In an initial set of experiments, we performed ELISA analyses with plate‐bound ACE2/APN01 to evaluate the impact of the indicated single and compound mutations on Spike RBD/ACE2 interactions. Intriguingly, almost all tested variant RBDs exhibited increased binding to APN01 (Appendix Fig 1A). These results prompted us to extend the number of analyzed variants to include current VOCs and to biophysically characterize RBD/ACE2 interactions using Biacore surface plasmon resonance (SPR) analysis.
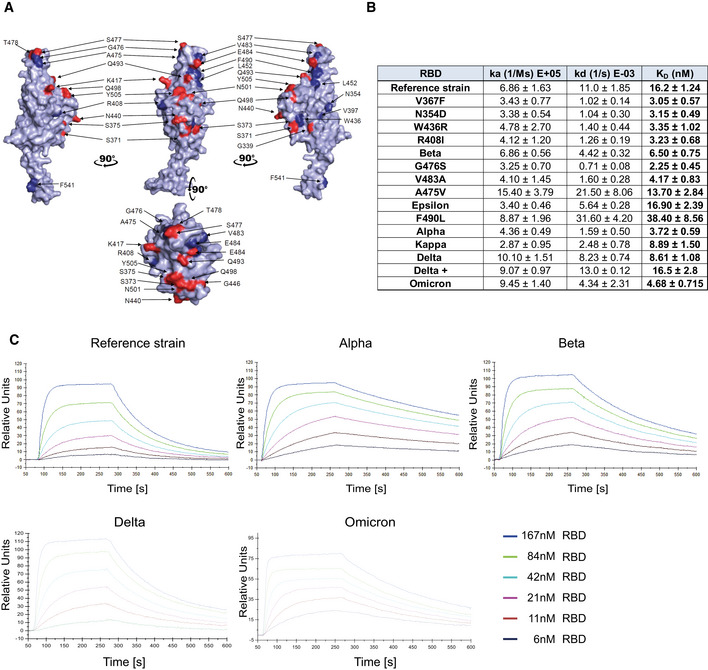
Figure 1. Increased affinity of APN01 interactions with SARS‐CoV‐2‐RBD variants.
-
APyMOL rendered visualization of the SARS‐CoV‐2 RBD. Rendering depicts the SARS‐CoV‐2 RBD from the Wuhan reference isolate with mutation sites observed in SARS‐CoV‐2 variants shown in red. Indicated in blue are positions mutated in the various strains of SARS‐CoV‐2 used in experiments in this study.
-
BSurface Plasmon Resonance analysis to derive kinetic constants (ka, kd) and affinity values (KD) of SARS‐CoV‐2 RBD/APN01 interaction. The constants represent mean values and standard deviations obtained from sensorgram fittings performed in quadruplicate. The table lists both the tested variants and the introduced amino acid substitution as well as the designation of the respective Variant of Concern (VOC) or Variant of Interest (VOI) mutations tested in this study. Reference strain RBD sequence corresponds to the Wuhan SARS‐CoV‐2 isolate.
-
CRepresentative SPR sensorgram images for the SARS‐CoV‐2 RBD/APN01 interaction of current VOCs.
For comparative kinetic binding analysis of SARS‐CoV‐2 RBD variants, dimeric APN01 was covalently coupled as ligand to optical sensor chips. Commercially available or in‐house purified RBDs containing the amino acid changes described in Fig 1A were passed as analytes over the immobilized APN01 ligand in twofold serial dilutions. These proteins contained previously identified RBD mutations of current VOCs, as well as amino acid substitutions identified in variants of interest (VOI). Binding on‐rates (association constants; ka), off‐rates (dissociation constants; kd), and binding affinities (KD reported as nM) of SARS‐CoV‐2 RBD/ACE2 interactions were determined by mathematical sensorgram fitting, applying a monomeric Langmuir 1:1 interaction model (A + B = AB) using BiaEvaluation 4.1 software. The results are summarized in Fig 1B, listing both the tested variants and the introduced amino acid substitution as well as the designation of the respective VOI and VOC that were tested with SPR analysis. Of note, the reference RBD sequence corresponds to the original Wuhan SARS‐CoV‐2 isolate. Importantly, our SPR analyses showed that affinities of all tested VOC RBDs to APN01 are substantially increased, with the Alpha variant RBD showing the highest affinity (Fig 1B and C); see Appendix Fig 1B for SPR sensorgrams of variants Kappa and Delta+). Of note, among these VOC RBDs, we observed a significantly lower off‐rate for the Alpha variant and a significantly faster on‐rate for the Delta variant. Binding of the Omicron variant was characterized by a high association rate, comparable to the Delta variant, but an off‐rate reduction, as observed for the early variants of concern.
Combined with the increased binding affinity, changes in these kinetic parameters might contribute along with conformational changes of the whole Spike and the impact of mutations affecting proteolytic processing to the enhanced infectivity of VOC. These data show that, as expected and also in part reported by others (Cai et al, 2021; Gobeil et al, 2021; Kim et al, 2021; Ramanathan et al, 2021; Tchesnokova et al, 2021), viral evolution of the Spike protein led to an increase in Spike RBD/ACE2 binding affinity as well as altered kinetic constants of RBD/APN01 interaction, especially observed for VOC.
Enhanced binding of SARS‐CoV‐2 VOC Spike trimers to clinical grade ACE2/APN01
SARS‐CoV‐2 RBD binding to ACE2 occurs in the context of trimeric Spike proteins in the pre‐fusion conformation. Spike trimer structures have been solved in receptor bound or unbound forms (Walls et al, 2020; Wrapp et al, 2020; Cerutti et al, 2022; Mannar et al, 2022; Zhou et al, 2022), and it has been demonstrated that the conformation of the Spike (open or closed) is of critical relevance for receptor interactions (Cai et al, 2020; Xu et al, 2021). Importantly, mutations outside the RBD that alter the Spike conformation have been reported to also affect viral infectivity and receptor binding (Mansbach et al, 2021; Yang et al, 2021). To test whether our observations on the increased affinity of VOC RBD/APN01 interaction is also observed in the context of the full‐length Spike, we assessed APN01 binding to recombinant pre‐fusion trimeric SARS‐CoV‐2 Spike proteins. Since APN01 is a dimeric molecule thus allowing for bivalent target interaction, the VOC trimeric pre‐fusion Spike variant proteins were immobilized to an optical sensor chip surface by covalent amine coupling. APN01 was passed over the immobilized Spike proteins in serial dilution in single binding cycles. Using BiaEvaluation 4.1 software, subsequent kinetic analysis was carried out by sensorgram fitting applying a Langmuir binding and a bivalent analyte binding model. Kinetic binding constants derived from the Langmuir model represent the apparent affinity. The bivalent analyte model calculates separate kinetic constants for the affinity determining first step A + B = AB and the avidity determining second step AB + B = AB2 of the binding process.
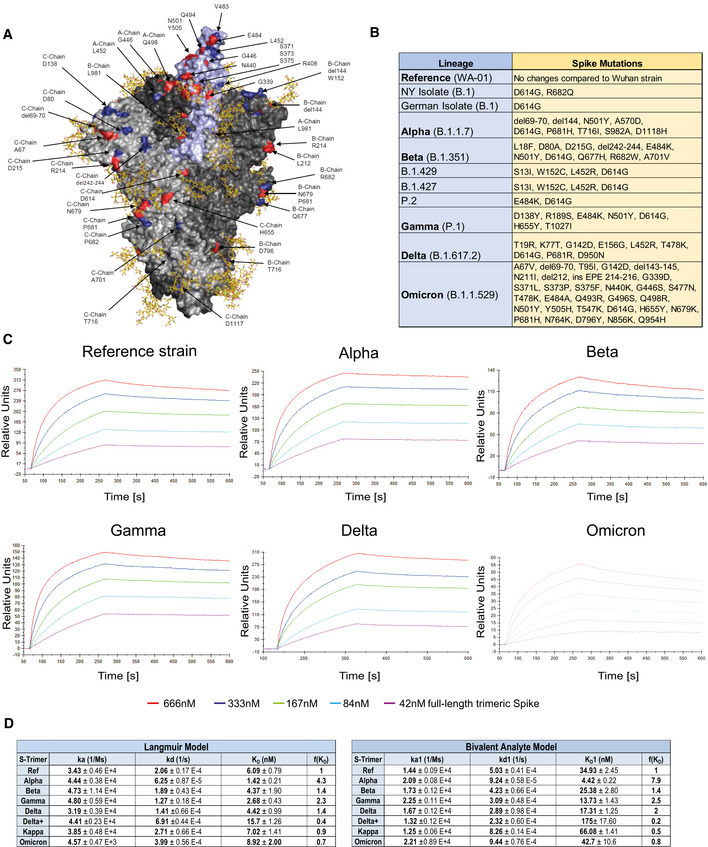
Structural rendering of the trimeric full‐length SARS‐CoV‐2 Spike protein and the positions mutated in the various strains of SARS‐CoV‐2 used in this study are shown in Fig 2A. Lineages and corresponding mutations of the SARS‐CoV‐2 isolates including VOCs and VOIs used in these biophysical analyses and cellular assays are listed in Fig 2B. SPR analysis showed strong binding of APN01 to all tested variants of the pre‐fusion Spike trimers including the original Wuhan viral isolate trimer as well as the Alpha, Beta, Gamma, Delta, and Kappa, and Omicron variants (Fig 2C; Appendix Fig 2A). Sensorgram fitting showed enhanced apparent affinity (avidity; Langmuir model) and calculated first step affinity (KD1, bivalent analyte model) of Alpha, Beta, Gamma, and Delta trimeric Spike proteins compared with Spike trimers of the reference strain (Fig 2C and D). For Omicron, no affinity enhancement was observed in this particular experimental setup. These results confirm that dimeric recombinant soluble human ACE2 (APN01) binds to the pre‐fusion Spike trimers of all current VOCs with strong affinity/avidity.
Figure 2. Increased binding affinity of APN01 to full‐length pre‐fusion trimeric Spike proteins from SARS‐CoV‐2 variants of concern.
-
APyMOL rendering of the trimeric full‐length SARS‐CoV‐2 Spike protein. One RBD is shown in violet. Indicated in blue are positions mutated in the various strains of SARS‐CoV‐2 used in experiments in this study. Mutations described for the Omicron VOC are depicted in red. Shown in yellow are the glycan modifications of the spike protein (Capraz et al, 2021).
-
BTable lists the SARS‐CoV‐2 strains and their respective mutations within the Spike protein that were used in this study.
-
CRepresentative sensorgram images for the SPR analysis conducted with full‐length trimeric spike proteins in pre‐fusion state with APN01. Reference strain corresponds to original Wuhan viral isolate spike sequence. Indicated are VOC Alpha, Beta, Gamma, and Delta, and Omicron.
-
DTables listing ka, kd, as well as KD values for the interaction of APN01 and full‐length trimeric spike proteins. The constants represent mean values and standard deviations obtained from sensorgram fittings performed in quadruplicate. Values are derived from calculations based upon the Langmuir (left table) or Bivalent Analyte sensorgram fitting (right table).
ACE2/APN01 effectively neutralizes SARS‐CoV‐2 VOCs
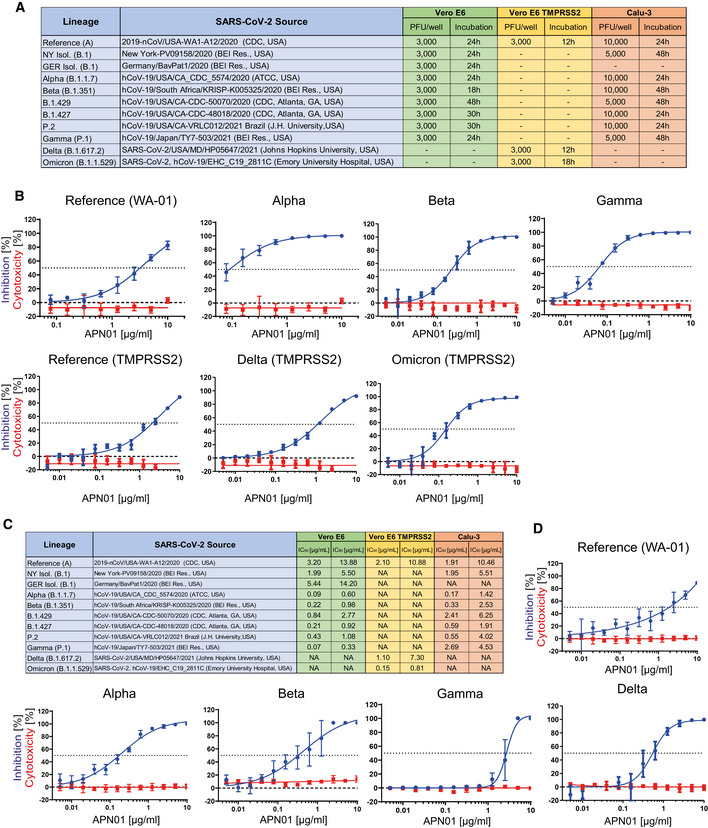
We have previously reported that clinical grade APN01 can effectively reduce the SARS‐CoV‐2 viral load in VeroE6 cells and 3D organoids in a dose‐dependent manner, using a reference virus isolated early during the pandemic (Monteil et al, 2020). This virus carries the same Spike sequence as the originally reported virus isolated in Wuhan. To test whether APN01 can also neutralize variant clinical SARS‐CoV‐2 isolates including VOC strains (see Fig 3A for a list of tested strains), we performed neutralization assays in VeroE6 cells and VeroE6 cells overexpressing TMPRSS2 (VeroE6‐TMPRSS2) and compared the inhibitory potency side by side to our reference strain. APN01 potently neutralized all the SARS‐CoV‐2 isolates we tested in VeroE6 cells (Fig 3B; Appendix Fig 3A). Intriguingly, this inhibition was markedly enhanced against all the VOCs tested with sometimes up to 20 times lower IC50 and IC90 values (Fig 3C).
Figure 3. Increased neutralization potency of APN01 toward SARS‐CoV‐2 variants.
-
ATable depicts source of the tested viral isolates, as well plaque‐forming units (PFU) and the infection time used in these experiments for both VeroE6/VeroE6‐TMPRSS2 and Calu‐3 cells.
-
BPanels depict both neutralization of the indicated SARS‐CoV‐2 isolates (blue line) and cytotoxicity of APN01 (red line) in VeroE6 or VeroE6‐TMPRSS2 cells. Analysis was done in quadruplicate with mean and standard deviations shown. Y‐axis depicts the percentage of neutralization and cytotoxicity, respectively.
-
CTable depicts IC50 and IC90 values for APN01‐mediated neutralization of viral infection in VeroE6/VeroE6‐TMPRSS2 and Calu‐3 cells.
-
DSame experimental setup as in (a) but conducted with the epithelial lung cancer cell line Calu‐3. Analysis was done in quadruplicate with mean and standard deviations shown. Y‐axis depicts the percentage of neutralization and cytotoxicity, respectively.
To extend our results to a physiologically more relevant cell system, we infected Calu‐3 human lung epithelial cells with the reference SARS‐CoV‐2 isolate and the indicated SARS‐CoV‐2 variants. We again observed that clinical grade soluble APN01 potently reduced viral load of all tested variants in a dose‐dependent manner (Fig 3C and D; Appendix Fig 3B). Importantly, the observed neutralization potency closely correlated with the Spike/APN01 binding affinity as assessed by SPR. In some cases, we detected 10 to 20 times lower IC50 and IC90 values when compared to values obtained with the SARS‐CoV‐2 reference strain. These data show that ACE2/APN01 not only binds significantly stronger to RBD or full‐length Spike proteins of the tested variants, but also more potently inhibits viral infection by these strains in both VeroE6 and human lung epithelial cells.
Independent validation studies
Experimental setups and model systems as well as culture conditions sometimes have a dramatic impact on experimental results. To ensure the reproducibility of our results, we conducted confirmatory experiments in a different and independent laboratory. These validation experiments were performed at the Karolinska Institutet, Stockholm, where VeroE6 cells were infected with reference virus isolated from the first Swedish patient. This virus was previously reported (Monteil et al, 2020, 2021) and carries the same Spike amino acid sequence as reported for the first Wuhan virus isolate. Experiments were performed at different Multiplicities of Infection (MOIs) to test the inhibitory potency of APN01. As reported before (Monteil et al, 2020), APN01 markedly reduced viral replication of the SARS‐CoV‐2 reference strain in a dose‐dependent manner (Fig 4A and B). Importantly, the inhibitory potency of APN01 was again significantly increased toward the VOC Alpha and Beta, providing an independent validation of the neutralization results. This increase in neutralization potency of APN01, when the reference virus was compared with the VOC strains Alpha and Beta, could be observed at all APN01 concentrations and MOIs tested (Fig 4A–C).
Figure 4. Increased potency of APN01 against SARS‐CoV‐2 VOC.
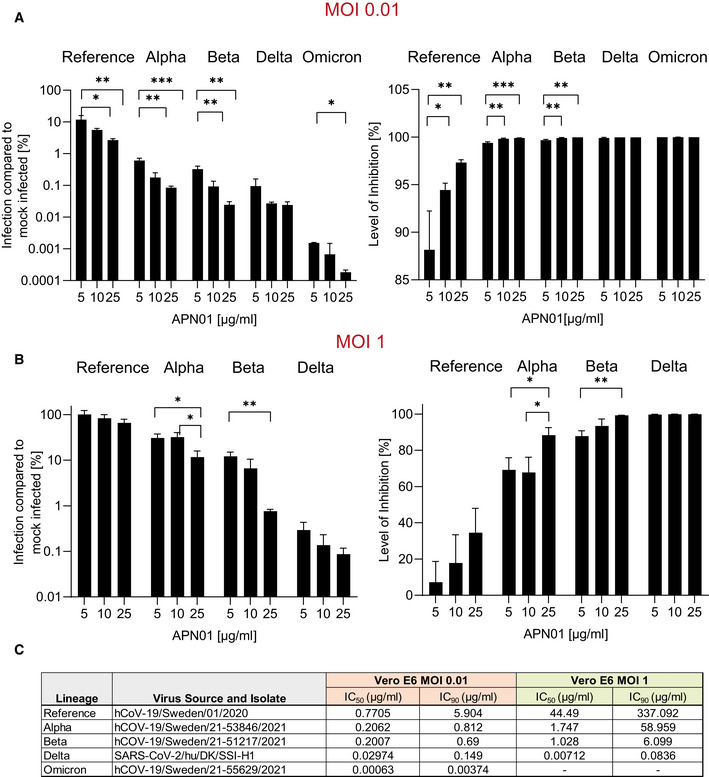
-
A, BDiagrams depict the level of infection with the indicated SARS‐CoV‐2 isolates at MOI 0.01 (A) and MOI 1 (B) of VeroE6 cells after pre‐incubation with increasing concentrations of APN01 as compared to infections in the absence of APN01 pre‐incubation. MOIs as depicted reflect multiplicities of infection before the pre‐incubation with APN01 that was followed by the viral infection of cells. Shown are means of biological replicates (n = 3) analyses with standard deviations. Statistical significance is indicated by asterisks (*P‐value < 0.05; **P‐value < 0.01, ***P‐value < 0.001 as calculated with one‐way ANOVA).
-
CList and source of strains used at the Karolinska Institutet and IC50 and IC90 values obtained for the indicated MOIs. See Materials and Methods section for a detailed list of viral mutations for the strains used.
In addition to altering the interaction with the cell entry receptor human ACE2, mutations in the SARS‐CoV‐2 Spike have been reported to affect potential transmission from humans to domesticated and wild animals, as has been reported for laboratory strains of mice (Hobbs & Reid, 2021; Kuchipudi et al, 2022) and https://www.cdc.gov/coronavirus/2019‐ncov/daily‐life‐coping/animals.html). While this might have a positive impact on animal modeling of SARS‐CoV‐2 infection, this is also of potentially high concern due to the possibility of additional viral reservoirs, as well as the accumulation of mutations that might affect viral fitness upon transmission back to human hosts. We indeed observed that SARS‐CoV‐2 reference strain infection of Calu‐3 cells could not be inhibited by recombinant mouse ACE2 but the Omicron infection was potently suppressed by both human and mouse recombinant ACE2 for the indicated MOIs, albeit at lower potency when using soluble mouse ACE2 (Appendix Fig 4). These data are in line with reported findings on the effect of some VOC Spike mutations on transmissibility to non‐human hosts.
While the origin of the Omicron VOC is still not fully clarified, the high number of mutations in Omicron has raised concerns in terms of immune evasion and the protective effect of currently available vaccines and therapeutic antibodies. To test the potential impact of these mutations on the protection status of vaccinated individuals, we compared the potency of vaccinees' sera to neutralize both reference (Wuhan) and the Omicron SARS‐CoV‐2 virus. In support of previous observations, vaccinees' sera exhibited a significantly decreased potency in viral neutralization when reference and Omicron infections of VeroE6 cells were compared (Appendix Fig 5). Our data are in line with recently published observations on the waning efficacy of current vaccines, as well as a (partial) loss of neutralization potency of many currently marketed or developed monoclonal antibodies (Cele et al, 2021; Garcia‐Beltran et al, 2021; Greaney et al, 2021; Harvey et al, 2021; Jangra et al, 2021; Lopez Bernal et al, 2021; Planas et al, 2021; Peng et al, 2022; Takashita et al, 2022; Zhou et al, 2022). The results described in this study, using different viral isolates as well as different experimental procedures, independently confirm that clinical grade soluble human APN01 strongly blocks SARS‐CoV‐2 infections of recently emerged VOC and indicate that this therapeutic approach provides a potentially universally efficacious therapeutic approach to treat all current and future variants of SARS‐CoV‐2.
Discussion
Since emerging from Wuhan, China, in December of 2019, SARS‐CoV‐2 has been causing devastating severe respiratory infections in humans worldwide. Multiple variants of SARS‐CoV‐2 have emerged and circulated around the world throughout the COVID‐19 pandemic with some strains displaying even greater infectivity and transmissibility (see https://www.cdc.gov/coronavirus/2019‐ncov/variants/variant‐info.html and https://www.who.int/en/activities/tracking‐SARS‐CoV‐2‐variants/ for further information). Some of these variants have been classified by the WHO as Variants of Concern (VOC), defined as “a variant for which there is evidence of an increase in transmissibility, more severe disease, significant reduction in neutralization by antibodies generated during previous infection or vaccination, reduced effectiveness of treatments or vaccines, or diagnostic detection failures” or Variants of Interest (VOI), defined as “a variant with specific genetic markers that have been associated with changes to receptor binding, reduced neutralization by antibodies generated against previous infection or vaccination, reduced efficacy of treatments, potential diagnostic impact, or predicted increase in transmissibility or disease severity.” The current VOCs are B.1.1.7 (Alpha), B.1.351 (Beta), P.1 (Gamma), B.1.617.2 (Delta), and in particular Omicron (B.1.529), whereas multiple VOI are currently circulating as well. Both large‐scale vaccination programs, emerging treatments as well as protracted infections in immune‐compromised hosts or animal reservoirs, could potentially drive further molecular evolution of SARS‐CoV‐2. It is, therefore, to be expected that novel variants will arise, some of which will rapidly spread and even re‐infect fully vaccinated people, sometimes leading to severe breakthrough COVID‐19, as has been observed for the Delta VOC and is currently observed for the Omicron variant (Farinholt et al, 2021; Christensen et al, 2022). In this arena of global vaccination efforts and antibody therapeutics in multiple permutations, it therefore remains critical to identify and design universal strategies that might help prevent and treat infections with all current and potential future variants.
The SARS‐CoV‐2 Spike protein interacts with high affinity with its main entry receptor ACE2, followed by a subsequent membrane fusion step. Although alternative ACE2‐independent modes of viral uptake and infection have been reported, genetic modeling in ACE2 deficient mice has provided unambiguous evidence that ACE2 is the essential receptor for SARS‐CoV‐2 infections and subsequent development of COVID‐19 in mice (Gawish et al, 2022). Indeed, most neutralizing antibodies from vaccinations, convalescent plasma therapies, and monoclonal antibodies or nanobodies interfere with the Spike/ACE2 interaction (Harvey et al, 2021) and have, due to their therapeutic or prophylactic efficacy, provided further evidence for the essential role of ACE2 as the relevant in vivo receptor for SARS‐CoV‐2 and COVID‐19 in humans. Conceptually, all SARS‐CoV‐2 variants and “escape mutants” still bind to ACE2 (Cai et al, 2021; Gobeil et al, 2021; Motozono et al, 2021; Ou et al, 2021; Tchesnokova et al, 2021; Tian et al, 2021; Yuan et al, 2021; Zhou et al, 2021), in part already validated in various studies (Cai et al, 2021; Yuan et al, 2021; Zhang et al, 2021). ACE2 is a carboxypeptidase that controls angiotensin II peptide levels and thereby is involved in critical aspects of physiology such as blood pressure control or sodium retention as well as disease processes including heart failure, blood vessel and kidney pathologies in diabetes, tissue fibrosis, or regulation of inflammatory cytokines (Kuba et al, 2010). Our group was the first to show that soluble dimeric ACE2 protects mice from acute lung injury and acute respiratory distress syndrome (ARDS) (Imai et al, 2005), which triggered preclinical and clinical development of recombinant human soluble ACE2 (Treml et al, 2010; Haschke et al, 2013), termed APN01, for lung disease. It was, therefore, critical to systematically assess whether APN01 could indeed bind to the RBD and more importantly full‐length Spike of emerged SARS‐CoV‐2 variants and neutralize infections with these variants—especially recently emerged VOC. Our data show that APN01 associates with the RBD and pre‐fusion trimeric Spike of all variants tested, often at markedly increased affinity and avidity, which is in line with the enhanced infectivity of these variants. Importantly, using different cell types including lung epithelial cells, APN01 showed markedly improved efficacy to block infection of all VOC, demonstrating that the prediction holds true—clinical grade APN01 can effectively block all tested SARS‐CoV‐2 variants and this inhibition is markedly improved against all tested VOCs and VOIs.
APN01 has now undergone phase 2 testing in severe (WHO scores 4, 5, and 6) COVID‐19 patients using intravenous infusions. Additionally, a new formulation of APN01 that can be inhaled as an aerosol to directly interfere with the earliest steps of viral infection and prevent lung damage and spreading to other organ systems has been developed (preprint: Shoemaker et al, 2021). Moreover, the preclinical efficacy of APN01 lung administration in a mouse‐adapted severe COVID‐19 model has been demonstrated, providing proof of concept for phase 1 clinical inhalation trials in humans that are currently ongoing. Of note, the FDA recently expanded emergency use authorization for a two‐antibody “cocktail” to allow its use in patients seeking protection from COVID‐19 following exposure to someone infected with SARS‐CoV‐2, providing a framework for preventive and post‐exposure prophylactic strategies. However, structural and functional analyses of the VOCs reveal decreased efficacy and viral escape from therapeutic antibodies or antibodies generated from natural infection and vaccinations (Zhou et al, 2022). Further viral evolution, especially under selective pressure by both widespread vaccination programs and monoclonal antibodies or antibody cocktails, might drive the emergence of resistant novel variants and strains. In particular, the Delta and Omicron VOC strains have already substantially contributed to a new global wave of infection and concerningly large numbers of re‐infections. Our data show that clinical grade soluble ACE2/APN01 blocks infectivity of all tested variants. As expected from the stoichiometry of these interactions, this neutralization was dependent upon the ratio of infectious particles to neutralizing APN01, that is, APN01 potency correlated with the number of infectious particles. These results support the notion that this therapeutic is inherently resistant to escape mutations that constitute a major problem for both vaccines and antibody‐based therapeutics. Data provided in this study and in (preprint: Shoemaker et al, 2021) as well as clinical data already generated for APN01 set the stage for the development of universal and pan‐variant SARS‐CoV‐2 prevention and therapy.
Limitations of this study
Our study used two different cell types and should be expanded using additional human cell types and also organoids. Additionally, different incubation times with viral nucleoprotein staining as experimental readout might affect the accuracy of the obtained experimental results. Moreover, to indeed make a claim on universality, additional variants should (and can) be tested using affinity/avidity measurements and neutralization assays.
Materials and Methods
Surface plasmon resonance analysis
Recombinant SARS‐CoV‐2 spike proteins (His‐tagged) were purchased from Acro Biosystems Inc. (Newark, USA) or produced in‐house by overexpression of respective constructs in HEK293T cells followed by purification via nickel NTA chromatography and size exclusion chromatography. The purity of recombinant proteins was documented by SDS–PAGE analysis. SPR measurements were performed on a Biacore 3000 instrument (GE Healthcare). For comparative kinetic analysis of SARS‐CoV‐2 RBD variants, APN01, which does not contain a capture tag, was immobilized on optical sensor chip surfaces by covalent coupling at pH = 4.5 at ligand densities of 2,672 ± 145 RU following the Biacore amine coupling protocol. Amine‐activated flow cell 1 (FC1) was used as a reference to allow for the generation of background‐subtracted binding sensorgrams. SARS‐CoV‐2 RBD protein variants were passed over the immobilized APN01 ligand as analytes in twofold serial dilution (167, 84, 42, 21, 11, and 6 nM) in single binding cycles, at a flow rate of 30 μl/min in HBS‐EP buffer (0.1 M HEPES, 1.5 M NaCl, 0.03 M EDTA and 0.5% v/v Surfactant P20). Bound analyte was removed after each cycle by surface regeneration with 3 M MgCl2. Reference flow cell (FC1) subtracted sensorgram overlays with additional correction by subtracting buffer (c = 0) sensorgrams (double referencing) were generated and used for kinetic binding analysis. Subtraction spikes occurring at the injection start were removed in the sensorgrams shown in the figures. Kinetic binding constants (ka, kd, and KD) were generated by mathematical sensorgram fitting. Generally, a Langmuir 1:1 interaction model (A + B = AB) was applied, using BiaEvaluation 4.1 software. A series of 4 curve fittings was performed for each binding reaction, using a simultaneous single fitting algorithm for each of the sensorgram overlays. Mean values and standard deviations were obtained from fitting runs with Chi2 values ≤3% of Rmax. Binding affinities (reported as nM) were calculated from on‐ and off‐rate constants.
SPR analysis of APN01 binding to recombinant full‐length trimeric SARS‐CoV‐2 spike proteins was performed with the spike proteins immobilized to CM5 optical sensor chip surfaces by covalent amine coupling to reach a surface density of 900–1,000 RU. APN01 was passed over the immobilized spike proteins as dimeric bivalent analyte at five concentrations (42, 84, 167, 333 and 666 nM) in repetitive single binding cycles. Using BiaEvaluation 4.1 software, kinetic analysis was carried out by applying a Langmuir 1:1 (apparent affinity) and a bivalent analyte binding algorithm, which separates first step binding (A + B = AB; affinity) from the second step binding (AB + B = AB2; avidity).
Cell lines and cell culture
Infection and APN01‐mediated viral neutralization assays were conducted at the Integrated Research Facility at Fort Detrick (IRF‐Frederick) of the National Institute of Allergy and Infectious Diseases (NIAID) or at the Department of Laboratory Medicine (Unit of Clinical Microbiology) of the Karolinska Institutet and Karolinska University Hospital. Vero E6 cells (Vero C1008; American Type Culture Collection [ATCC], Manassas, VA, USA) were cultured in DMEM medium (Gibco, Gaithersburg, MD, USA or Thermo Fisher) containing 10% fetal bovine serum (FBS). Calu‐3 cells (HTB‐55; American Type Culture Collection [ATCC], Manassas, VA, USA) were cultured in DMEM F12 Medium (ATCC) with 20% FBS. Both cell lines were tested for mycoplasma.
Viral neutralization experiments conducted at the Karolinska Institutet
A 24 h after seeding of Vero E6 cells (5 × 104 per 48 well), APN01 was mixed with viral particles of the indicated strains at the given concentrations in DMEM Medium (Thermofisher) containing 5% FBS in 100 μl per well and incubated for 30 min at 37°C. After the incubation period medium was removed from Vero E6 cells, cells were washed once with PBS to remove any non‐attached cells and virus/APN01 mixtures were added to the cells. Cells were incubated with virus for 15 h, after which cells were washed three times with PBS and lysed the Trizol, subsequently. RNA was extracted using the direct‐zol RNA kit (Zymo Research) and assayed by qRT–PCR as previously described (Monteil et al, 2020). Half‐maximal inhibitory concentration (IC50) and inhibitory concentration 90 (IC90) were calculated using GraphPad Prism Software (La Jolla, CA). For serum neutralization assays, Vero E6 cells were seeded in 48‐well plates as described above. 24‐h post‐seeding, indicated dilutions of vaccinated subjects sera were mixed with SARS‐CoV‐2 Wuhan or Omicron strains at an MOI of 0.01 in a final volume of 100 ml per well in DMEM (5% FBS) at 37°C under shaking conditions for 30 min. The serum dilutions used in this experiment were determined after a neutralization assay against the Wuhan reference strain. After 30 min, Vero E6 cells were infected with Serum/301 SARS‐CoV‐2 for 15 h. A 15‐h post‐infection, supernatants were removed, and cells were washed 3 times with PBS and then lysed using Trizol (Thermofisher) before analysis by qRT–PCR for viral RNA detection as previously described (Monteil et al, 2020).
Viral neutralization experiments conducted at the IRF‐Fredrick
For infection experiments, Vero E6 cells were seeded at 6,000 cells in 30 μl in 384 wells 24 h prior to infection and Calu‐3 cells were plated at 10,000 cells per well in 30 μl 48 h prior to infection in their respective culture media. For APN01 neutralization experiments, APN01 was diluted twofold in an eight‐ or twelve‐point dose curve. Each condition was tested in quadruplicate (n = 4) with an efficacy plate and a mock‐infected cytotoxicity plate run in parallel. Strains for infection and MOIs are listed in a table in the respective figures. Suitable MOIs were optimized previously for each cell line and virus using serial dilutions and staining for SARS‐CoV‐2 nucleoprotein, as described below. The viral inoculum was diluted in the respective cell culture medium containing indicated doses of APN01 and pre‐incubated for 60 min. 20 μl of the virus/APN01 mixture was transferred directly to plates containing cells to reach a final volume of 50 μl. After infection, cells were incubated for 24 h or 48 h, in accordance with optimal virus and cell‐line infection conditions. Cells were then fixed in 10% formalin and stained with a SARS‐CoV‐1 nucleoprotein‐specific antibody (cross‐reactive to SARS‐CoV‐2; see antibody list for further information), followed by a secondary antibody conjugated with fluorophore and/or horseradish peroxidase (HRP). Fluorescence was quantitated using a PerkinElmer Operetta high‐content imaging system (PerkinElmer, Massachusetts, USA). Chemiluminescence was read on a Tecan M1000 plate reader (Tecan, Switzerland). Cytotoxicity on mock‐infected plates was determined using the Promega CellTiter‐Glow Luminescent Cell Viability Assay (Promega, Madison, Wisconsin, USA). Half‐maximal inhibitory concentration (IC50) and 50% cytotoxic concentration (CC50) were calculated as described by Covés‐Datson et al (2019), using GraphPad Prism Software (La Jolla, CA). Z' factor scores were assessed as quality‐control parameters for each plate of each run. All plates included in the report passed quality‐control criteria.
Viral strains and isolates
Sources and strains used in the experiments at the NIAID and Karolinska Institutet are indicated in the respective figures, including a list of SARS‐CoV‐2 spike mutations identified by sequencing of viral isolates (NIAID). SARS‐CoV‐2 variant Delta was provided by Charlotta Polacek Strandh, Statens Serum Institut, Copenhagen, Denmark. Experiments dealing with SARS‐CoV‐2 were performed in a BSL‐3 laboratory under the approval of Public Health Agency of Sweden. Mutations of viral isolates used at Karolinska are listed in Appendix Table 1.
Preparation of recombinant human ACE2
Clinical grade recombinant human ACE2 (amino acids 18–740) was produced by the contract manufacturer Polymun Scientific (Klosterneuburg, Austria) from CHO cells according to GMP guidelines under serum‐free conditions and formulated as a physiologic aqueous solution, as described previously (Haschke et al, 2013; Zoufaly et al, 2020).
ELISA experiments
96‐well ELISA plates were coated with 100 μl of anti‐ACE2 coating antibody (2 μg/ml diluted in PBS pH 7.4) overnight at room temperature. Following coating, plates were washed 3 times with 300 μl of washing buffer (PBS + 0.05% Tween‐20) and blocked with 300 μl of blocking buffer (1% BSA in PBS, pH 7.4) for 1 h at room temperature. After blocking, plates were washed five times and 100 μl APN01, diluted in blocking buffer to a concentration of 2 μg/ml, was applied to the wells and incubated for 1 h at room temperature. Subsequently, plates were washed five times with washing buffer and 100 μl of SARS‐CoV‐2 Spike RBD variants were added to the plates in triplicates. As controls for non‐specific binding, wells were incubated in washing buffer without RBD proteins. Following incubation for 1 h at room temperature, plates were washed five times with washing buffer after which 100 μl of anti‐Histidine detection antibody (diluted 1/500 in blocking buffer) was added for another hour. After five washing steps, 100 μl secondary HRP conjugated antibody was added for 1 h at room temperature. Subsequently, the plates were washed seven times with washing buffer and 100 μl substrate (TMB Microwell Peroxidase Substrate, Seramun Diagnostika #S‐001‐2‐TMB, ready‐to‐use) was added to the wells. The colorimetric reaction was stopped by the addition of 50 μl of 1 M sulfuric acid and subsequently analyzed for absorption at 450 nm. To measure unspecific background binding, control values were generated by omitting the addition of RBD proteins. These control values were subtracted from RBD‐specific signals and plotted as OD values.
Visualizations of RBDs and full‐length Spike protein
Visualizations were rendered with pymol software (the PyMOL Molecular Graphics System, Version 2.4 Schrödinger, LLC), based on a model of the fully glycosylated Spike‐ACE2 complex described in Capraz et al (submitted for publication) and https://covid.molssi.org//models/#spike‐protein‐in‐complex‐with‐human‐ace2‐ace2‐spike‐binding.
Sera of vaccinees
Sera were taken 5–7 weeks after the second immunization with the mRNA vaccine Comirnaty (median dose interval 21 days (range 21–24) from four SARS‐CoV‐2‐naïve healthcare workers (75% female, median age 46 [IQR 37–59]) which took part in the COMMUNITY study. The COMMUNITY study was approved by the Swedish Ethical Review Authority (Dnr: 2020–01653). Informed consent was obtained from all subjects and experiments conformed to the principles set out in the WMA Declaration of Helsinki and the Department of Health and Human Services Belmont Report.
Primers and antibodies used in this study are listed in Appendix Tables 2 and 3.
Author contributions
Vanessa Monteil: Conceptualization; data curation; formal analysis; visualization; methodology. Brett Eaton: Data curation; formal analysis. Elena Postnikova: Data curation; formal analysis; investigation. Michael Murphy: Conceptualization; data curation. Benedict Braunsfeld: Data curation; software; visualization. Ian Crozier: Conceptualization. Franz Kricek: Conceptualization; data curation; formal analysis. Janine Niederhoefer: Data curation; formal analysis; methodology. Alice Schwarzboeck: Investigation. Helene Breid: Data curation. Stephanie Devignot: Formal analysis. Jonas Klingström: Formal analysis. Charlotte Thalin: Formal analysis. Max J Kellner: Formal analysis. Wanda Christ: Formal analysis. Sebastian Havervall: Formal analysis. Stefan Mereiter: Conceptualization. Sylvia Knapp: Conceptualization. Anna Sanchez Jimenez: Methodology. Agnes Bugajska‐Schretter: Conceptualization. Alexander Dohnal: Conceptualization. Christine Ruf: Formal analysis; investigation. Romana Gugenberger: Conceptualization. Astrid Hagelkruys: Conceptualization; funding acquisition. Nuria Montserrat: Conceptualization. Ivona Kozieradzki: Data curation. Omar Hasan Ali: Formal analysis. Johannes Stadlmann: Conceptualization. Michael R Holbrook: Conceptualization; data curation; formal analysis. Connie Schmaljohn: Conceptualization. Chris Oostenbrink: Conceptualization; software; formal analysis. Robert H Shoemaker: Conceptualization. Ali Mirazimi: Conceptualization; data curation; formal analysis. Gerald Wirnsberger: Conceptualization; data curation; formal analysis; supervision; funding acquisition; validation; investigation; visualization; methodology; project administration. Josef M Penninger: Conceptualization.
Disclosure and competing interests statement
J.M.P. declares a conflict of interest as a founder and shareholder of Apeiron Biologics. G.W., R.G., A.S., J.N, A.S.J, and H.B. are employees of invIOs. Apeiron holds a patent on the use of ACE2 for the treatment of various diseases and is currently testing APN01 (soluble recombinant human ACE2) for the treatment of COVID‐19. Josef Penninger is an EMBO Member. This has no bearing on the editorial consideration of this article for publication. The other authors declare that they have no conflict of interest.
Supporting information
Appendix
Acknowledgements
We like to thank employees of invIOs' R&D department for input and critical discussion during the course of this study. We would also like to acknowledge Mr. Gregory Kocher (IRF‐NIAID‐NIH) for growing SARS‐CoV‐2 virus variants. J.M.P. and A.H. and the research leading to these results has received funding from the T. von Zastrow foundation, the FWF Wittgenstein award (Z 271‐B19), the Austrian Academy of Sciences, the Innovative Medicines Initiative 2 Joint Undertaking (JU) under grant agreement No 101005026, and the Canada 150 Research Chairs Program F18‐01336 as well as the Canadian Institutes of Health Research COVID‐19 grants F20–02343 and F20‐02015. Additionally, this project has received funding from the Innovative Medicines Initiative 2 Joint Undertaking (JU) under grant agreement no. 101005026. The JU receives support from the European Union's Horizon 2020 research and innovation program and EFPIA. Parts of this project have been funded with federal funds from the National Cancer Institute, National Institutes of Health, under Contract No. 75N91019D00024, Task Order No. 75N91019F00130. O.H.A. has received a grant from the Swiss National Science Foundation (P400PM_194473). Parts of this project have been funded with federal funds from the National Cancer Institute, National Institutes of Health, under Contract No. 75N91019D00024, Task Order No. 75N91019F00130 and through Laulima Government Solutions, LLC prime contract with the US National Institute of Allergy and Infectious Diseases (NIAID) under Contract No. HHSN272201800013C (B.E., M.R.H.). M.M. and E.P. performed this work as employees of Tunnell Government Services (TGS), a subcontractor of Laulima Government Solutions, LLC. The content of this publication does not necessarily reflect the views or policies of the Department of Health and Human Services, nor does mention of trade names, commercial products, or organizations imply endorsement by the U.S. Government (IC).
EMBO Mol Med (2022) 14 e15230
Contributor Information
Gerald Wirnsberger, Email: gwi@invios.com.
Josef M Penninger, Email: josef.penninger@ubc.ca.
Data availability
This study includes no data deposited in external repositories.
References
- Banerjee A, Mossman K, Grandvaux N (2021) Molecular determinants of SARS‐CoV‐2 variants. Trends Microbiol 29: 1–3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benton DJ, Wrobel AG, Xu P, Roustan C, Martin SR, Rosenthal PB, Skehel JJ, Gamblin SJ (2020) Receptor binding and priming of the spike protein of SARS‐CoV‐2 for membrane fusion. Nature 588: 327–330 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bergwerk M, Gonen T, Lustig Y, Amit S, Lipsitch M, Cohen C, Mandelboim M, Gal Levin E, Rubin C, Indenbaum V et al (2021) Covid‐19 breakthrough infections in vaccinated health care workers. N Engl J Med 385: 1–11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cai Y, Zhang J, Xiao T, Lavine CL, Rawson S, Peng H, Zhu H, Anand K, Tong P, Gautam A et al (2021) Structural basis for enhanced infectivity and immune evasion of SARS‐CoV‐2 variants. Science 373: 642–648 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cai Y, Zhang J, Xiao T, Peng H, Sterling SM, Walsh RM, Rawson S, Rits‐Volloch S, Chen B (2020) Distinct conformational states of SARS‐CoV‐2 spike protein. Science 369: 1586–1592 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Capraz T, Kienzl NF, Laurent E, Perthold JW, Föderl‐Höbenreich E, Grünwald‐Gruber C, Maresch D, Monteil V, Niederhöfer J, Wirnsberger G et al (2021) Structure‐guided glyco‐engineering of ACE2 for improved potency as soluble SARS‐CoV‐2 decoy receptor. eLife 10 10.7554/elife.73641 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cele S, Gazy I, Jackson L, Hwa SH, Tegally H, Lustig G, Giandhari J, Pillay S, Wilkinson E, Naidoo Y et al (2021) Escape of SARS‐CoV‐2 501Y.V2 from neutralization by convalescent plasma. Nature 593: 142–146 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cerutti G, Guo Y, Liu L, Liu L, Zhang Z, Luo Y, Huang Y, Wang HH, Ho DD, Sheng Z et al (2022) Cryo‐EM structure of the SARS‐CoV‐2 omicron spike. Cell Rep 38: 110428 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christensen PA, Olsen RJ, Long SW, Subedi S, Davis JJ, Hodjat P, Walley DR, Kinskey JC, Ojeda Saavedra M, Pruitt L et al (2022) Delta variants of SARS‐CoV‐2 cause significantly increased vaccine breakthrough COVID‐19 cases in Houston, Texas. Am J Pathol 192: 320–331 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Covés‐Datson EM, Dyall J, DeWald LE, King SR, Dube D, Legendre M, Nelson E, Drews KC, Gross R, Gerhardt DM et al (2019) Inhibition of Ebola virus by a molecularly engineered banana lectin. PLOS Negl Trop Dis 13: e0007595 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farinholt T, Doddapaneni H, Qin X, Menon V, Meng Q, Metcalf G, Chao H, Gingras MC, Avadhanula V, Farinholt P et al (2021) Transmission event of SARS‐CoV‐2 Delta variant reveals multiple vaccine breakthrough infections. BMC Med 19 10.1186/s12916-021-02103-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia‐Beltran WF, Lam EC, St. Denis K, Nitido AD, Garcia ZH, Hauser BM, Feldman J, Pavlovic MN, Gregory DJ, Poznansky MC et al (2021) Multiple SARS‐CoV‐2 variants escape neutralization by vaccine‐induced humoral immunity. Cell 184: 2372–2383.e9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gawish R, Starkl P, Pimenov L, Hladik A, Lakovits K, Oberndorfer F, Cronin SJ, Ohradanova‐Repic A, Wirnsberger G, Agerer B et al (2022) ACE2 is the critical in vivo receptor for SARS‐CoV‐2 in a novel COVID‐19 mouse model with TNF‐ and IFNγ‐driven immunopathology. eLife 11: e74623 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gobeil SM‐C, Janowska K, McDowell S, Mansouri K, Parks R, Stalls V, Kopp MF, Manne K, Li D, Wiehe K et al (2021) Effect of natural mutations of SARS‐CoV‐2 on spike structure, conformation, and antigenicity. Science 373: eabi6226 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greaney AJ, Starr TN, Barnes CO, Weisblum Y, Schmidt F, Caskey M, Gaebler C, Cho A, Agudelo M, Finkin S et al (2021) Mapping mutations to the SARS‐CoV‐2 RBD that escape binding by different classes of antibodies. Nat Commun 12: 4196 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harvey WT, Carabelli AM, Jackson B, Gupta RK, Thomson EC, Harrison EM, Ludden C, Reeve R, Rambaut A, Peacock SJ et al (2021) SARS‐CoV‐2 variants, spike mutations and immune escape. Nat Rev Microbiol 19: 409–424 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haschke M, Schuster M, Poglitsch M, Loibner H, Salzberg M, Bruggisser M, Penninger J, Krähenbühl S (2013) Pharmacokinetics and pharmacodynamics of recombinant human angiotensin‐converting enzyme 2 in healthy human subjects. Clin Pharmacokinet 52: 783–792 [DOI] [PubMed] [Google Scholar]
- Hassler L, Wysocki J, Gelarden I, Tomatsidou A, Gula H, Nicoleascu V, Randall G, Henkin J, Yeldandi A, Batlle D (2021) A novel soluble ACE2 protein totally protects from lethal disease caused by SARS‐CoV‐2 infection short title: Novel soluble ACE2 to combat SARS‐CoV‐2. bioRxiv 10.1101/2021.03.12.435191 [PREPRINT] [DOI] [Google Scholar]
- Higuchi Y, Suzuki T, Arimori T, Ikemura N, Mihara E, Kirita Y, Ohgitani E, Mazda O, Motooka D, Nakamura S et al (2021) Engineered ACE2 receptor therapy overcomes mutational escape of SARS‐CoV‐2. Nat Commun 12: 3802 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hobbs EC, Reid TJ (2021) Animals and SARS‐CoV‐2: Species susceptibility and viral transmission in experimental and natural conditions, and the potential implications for community transmission. Transbound Emerg Dis 68: 1850–1867 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffmann M, Kleine‐Weber H, Schroeder S, Krüger N, Herrler T, Erichsen S, Schiergens TS, Herrler G, Wu NH, Nitsche A et al (2020) SARS‐CoV‐2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 181: 271–280.e8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu B, Guo H, Zhou P, Shi ZL (2021) Characteristics of SARS‐CoV‐2 and COVID‐19. Nat Rev Microbiol 19: 141–154 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imai Y, Kuba K, Rao S, Huan Y, Guo F, Guan B, Yang P, Sarao R, Wada T, Leong‐Poi H et al (2005) Angiotensin‐converting enzyme 2 protects from severe acute lung failure. Nature 436: 112–116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jangra S, Ye C, Rathnasinghe R, Stadlbauer D, Personalized Virology Initiative Study Group , Krammer F, Simon V, Martinez‐Sobrido L, García‐Sastre A, Schotsaert M (2021) SARS‐CoV‐2 spike E484K mutation reduces antibody neutralisation. Lancet Microbe 2: e283–e284 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim S, Liu Y, Lei Z, Dicker J, Cao Y, Zhang XF, Im W (2021) Differential interactions between human ACE2 and spike RBD of SARS‐CoV‐2 variants of concern. J Chem Theory Comput 17: 7972–7979 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuba K, Imai Y, Ohto‐Nakanishi T, Penninger JM (2010) Trilogy of ACE2: A peptidase in the renin‐angiotensin system, a SARS receptor, and a partner for amino acid transporters. Pharmacol Ther 128: 119–128 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuchipudi SV, Surendran‐Nair M, Ruden RM, Yon M, Nissly RH, Vandegrift KJ, Nelli RK, Li L, Jayarao BM, Maranas CD et al (2022) Multiple spillovers from humans and onward transmission of SARS‐CoV‐2 in white‐tailed deer. Proc Natl Acad Sci U S A 119, e2121644119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuhlmann C, Mayer CK, Claassen M, Maponga T, Burgers WA, Keeton R, Riou C, Sutherland AD, Suliman T, Shaw ML et al (2022) Breakthrough infections with SARS‐CoV‐2 omicron despite mRNA vaccine booster dose. Lancet 399: 625–626 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kyriakidis NC, López‐Cortés A, González EV, Grimaldos AB, Prado EO (2021) SARS‐CoV‐2 vaccines strategies: a comprehensive review of phase 3 candidates. NPJ Vaccines 6: 28 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Linsky TW, Vergara R, Codina N, Nelson JW, Walker MJ, Su W, Barnes CO, Hsiang T‐Y, Esser‐Nobis K, Yu K et al (2020) De novo design of potent and resilient hACE2 decoys to neutralize SARS‐CoV‐2. Science 370: 1208–1214 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez Bernal J, Andrews N, Gower C, Gallagher E, Simmons R, Thelwall S, Stowe J, Tessier E, Groves N, Dabrera G et al (2021) Effectiveness of Covid‐19 vaccines against the B.1.617.2 (Delta) variant. N Engl J Med 385: 585–594 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mannar D, Saville JW, Zhu X, Srivastava SS, Berezuk AM, Tuttle KS, Marquez AC, Sekirov I, Subramaniam S (2022) SARS‐CoV‐2 omicron variant: Antibody evasion and cryo‐EM structure of spike protein‐ACE2 complex. Science 375: 760–764 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mansbach RA, Chakraborty S, Nguyen K, Montefiori DC, Korber B, Gnanakaran S (2021) The SARS‐CoV‐2 spike variant D614G favors an open conformational state. Sci Adv 7: 1–11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monteil V, Dyczynski M, Lauschke VM, Kwon H, Wirnsberger G, Youhanna S, Zhang H, Slutsky AS, Hurtado del Pozo C, Horn M et al (2021) Human soluble ACE2 improves the effect of remdesivir in SARS‐CoV‐2 infection. EMBO Mol Med 13: e13426 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monteil V, Kwon H, Prado P, Hagelkrüys A, Wimmer RA, Stahl M, Leopoldi A, Garreta E, Hurtado del Pozo C, Prosper F et al (2020) Inhibition of SARS‐CoV‐2 infections in engineered human tissues using clinical‐grade soluble human ACE2. Cell 181: 905–913.e7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Motozono C, Toyoda M, Zahradnik J, Saito A, Nasser H, Tan TS, Ngare I, Kimura I, Uriu K, Kosugi Y et al (2021) SARS‐CoV‐2 spike L452R variant evades cellular immunity and increases infectivity. Cell Host Microbe 29: 1124–1136.e11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ou J, Zhou Z, Dai R, Zhang J, Zhao S, Wu X, Lan W, Ren Y, Cui L, Lan Q et al (2021) V367F mutation in SARS‐CoV‐2 spike RBD emerging during the early transmission phase enhances viral infectivity through increased human ACE2 receptor binding affinity. J Virol 95: e0061721 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peng Q, Zhou R, Wang Y, Zhao M, Liu N, Li S, Huang H, Yang D, Au K‐K, Wang H et al (2022) Waning immune responses against SARS‐CoV‐2 variants of concern among vaccinees in Hong Kong. EBioMedicine 77: 103904 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pereson MIAJ, Flichman DM, Martinez AP, Baré P, Garcia GH, Di Lello FA (2021) Evolutionary analysis of SARS‐CoV‐2 spike protein for its different clades. J Med Virol 93: 3000–3006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Planas D, Veyer D, Baidaliuk A, Staropoli I, Guivel‐Benhassine F, Rajah MM, Planchais C, Porrot F, Robillard N, Puech J et al (2021) Reduced sensitivity of SARS‐CoV‐2 variant Delta to antibody neutralization. Nature 596: 276–280 [DOI] [PubMed] [Google Scholar]
- Ramanathan M, Ferguson ID, Miao W, Khavari PA (2021) SARS‐CoV‐2 B.1.1.7 and B.1.351 spike variants bind human ACE2 with increased affinity. Lancet Infect Dis 21: 1070 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rochman ND, Wolf YI, Faure G, Mutz P, Zhang F, Koonin EV (2021) Ongoing global and regional adaptive evolution of SARS‐CoV‐2. Proc Natl Acad Sci U S A 118: 1–10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shang J, Ye G, Shi K, Wan Y, Luo C, Aihara H, Geng Q, Auerbach A, Li F (2020) Structural basis of receptor recognition by SARS‐CoV‐2. Nature 581: 221–224 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shoemaker RH, Panettieri RA, Libutti SK, Hochster HS, Watts NR, Wingfield PT, Starkl P, Pimenov L, Gawish R, Hladik A et al (2021) Development of a novel, pan‐variant aerosol intervention for COVID‐19. bioRxiv 10.1101/2021.09.14.459961 [PREPRINT] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Svilenov HL, Sacherl J, Reiter A, Wolff L, Chen CC, Wachs FP, Pippig S, Wolschin F, Buchner J, Brockmeyer C et al (2020) Efficient inhibition of SARS‐CoV‐2 strains by a novel ACE2‐IgG4‐fc fusion protein with a stabilized hinge region. bioRxiv 10.1101/2020.12.06.413443 [PREPRINT] [DOI] [Google Scholar]
- Takashita E, Kinoshita N, Yamayoshi S, Sakai‐Tagawa Y, Fujisaki S, Ito M, Iwatsuki‐Horimoto K, Chiba S, Halfmann P, Nagai H et al (2022) Efficacy of antibodies and antiviral drugs against Covid‐19 omicron variant. N Engl J Med 386: 995–998 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanaka S, Nelson G, Olson CA, Buzko O, Higashide W, Shin A, Gonzalez M, Taft J, Patel R, Buta S et al (2021) An ACE2 triple decoy that neutralizes SARS‐CoV‐2 shows enhanced affinity for virus variants. Sci Rep 11: 1–12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tchesnokova V, Kulasekara H, Larson L, Bowers V, Rechkina E, Kisiela D, Sledneva Y, Choudhury D, Maslova I, Deng K et al (2021) Acquisition of the L452R mutation in the ACE2‐binding interface of spike protein triggers recent massive expansion of SARS‐CoV‐2 variants. J Clin Microbiol 59 10.1128/jcm.00921-21 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tian F, Tong B, Sun L, Shi S, Zheng B, Wang Z, Dong X, Zheng P (2021) N501Y mutation of spike protein in SARS‐CoV‐2 strengthens its binding to receptor ACE2. eLife 10 10.7554/elife.69091 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Treml B, Neu N, Kleinsasser A, Gritsch C, Finsterwalder T, Geiger R, Schuster M, Janzek E, Loibner H, Penninger J et al (2010) Recombinant angiotensin‐converting enzyme 2 improves pulmonary blood flow and oxygenation in lipopolysaccharide‐induced lung injury in piglets. Crit Care Med 38: 596–601 [DOI] [PubMed] [Google Scholar]
- Walls AC, Park YJ, Tortorici MA, Wall A, McGuire AT, Veesler D (2020) Structure, function, and antigenicity of the SARS‐CoV‐2 spike glycoprotein. Cell 181: 281–292.e6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Q, Zhang Y, Wu L, Niu S, Song C, Zhang Z, Lu G, Qiao C, Hu Y, Yuen KY et al (2020) Structural and functional basis of SARS‐CoV‐2 entry by using human ACE2. Cell 181: 894–904.e9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wrapp D, Wang N, Corbett KS, Goldsmith JA, Hsieh CL, Abiona O, Graham BS, McLellan JS (2020) Cryo‐EM structure of the 2019‐nCoV spike in the prefusion conformation. Science 367: 1260–1263 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu C, Wang Y, Liu C, Zhang C, Han W, Hong X, Wang Y, Hong Q, Wang S, Zhao Q et al (2021) Conformational dynamics of SARS‐CoV‐2 trimeric spike glycoprotein in complex with receptor ACE2 revealed by cryo‐EM. Sci Adv 7: 1–14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang T‐J, Yu P‐Y, Chang Y‐C, Liang K‐H, Tso H‐C, Ho M‐R, Chen W‐Y, Lin H‐T, Wu H‐C, Hsu S‐TD (2021) Effect of SARS‐CoV‐2 B.1.1.7 mutations on spike protein structure and function. Nat Struct Mol Biol 28: 731–739 [DOI] [PubMed] [Google Scholar]
- Yuan M, Huang D, Lee C‐CD, Wu NC, Jackson AM, Zhu X, Liu H, Peng L, van Gils MJ, Sanders RW et al (2021) Structural and functional ramifications of antigenic drift in recent SARS‐CoV‐2 variants. Science 373: 818–823 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang J, Cai Y, Xiao T, Lu J, Peng H, Sterling SM, Walsh RM, Rits‐Volloch S, Zhu H, Woosley AN et al (2021) Structural impact on SARS‐CoV‐2 spike protein by D614G substitution. Science 372: 525–530 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou D, Dejnirattisai W, Supasa P, Liu C, Mentzer AJ, Ginn HM, Zhao Y, Duyvesteyn HME, Tuekprakhon A, Nutalai R et al (2021) Evidence of escape of SARS‐CoV‐2 variant B.1.351 from natural and vaccine‐induced sera. Cell 184: 2348–2361.e6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou P, Yang X‐L, Wang X‐G, Hu B, Zhang L, Zhang W, Si H‐R, Zhu Y, Li B, Huang C‐L et al (2020) A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 588: E6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou T, Wang L, Misasi J, Pegu A, Zhang Y, Harris DR, Olia AS, Talana CA, Yang ES, Chen M et al (2022) Structural basis for potent antibody neutralization of SARS‐CoV‐2 variants including B.1.1.529. Science 376: eabn8897 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zoufaly A, Poglitsch M, Aberle JH, Hoepler W, Seitz T, Traugott M, Grieb A, Pawelka E, Laferl H, Wenisch C et al (2020) Human recombinant soluble ACE2 in severe COVID‐19. Lancet Respir Med 8: 1154–1158 [DOI] [PMC free article] [PubMed] [Google Scholar]