

Abstract
Fumarylacetoacetate hydrolase (FAH) catalyzes the final step of Tyrosine (Tyr) degradation pathway essential to animals and the deficiency of FAH causes an inborn lethal disease. In plants, a role of this pathway was unknown until we found that mutation of Short-day Sensitive Cell Death1 (SSCD1), encoding Arabidopsis FAH, results in cell death under short day. Phenylalanine (Phe) could be converted to Tyr and then degraded in both animals and plants. Phe ingestion in animals worsens the disease caused by FAH defect. However, in this study we found that Phe represses cell death caused by FAH defect in plants. Phe treatment promoted chlorophyll biosynthesis and suppressed the up-regulation of reactive oxygen species marker genes in the sscd1 mutant. Furthermore, the repression of sscd1 cell death by Phe could be reduced by α-aminooxi-β-phenylpropionic acid but increased by methyl jasmonate, which inhibits or activates Phe ammonia-lyase catalyzing the first step of phenylpropanoid pathway, respectively. In addition, we found that jasmonate signaling up-regulates Phe ammonia-lyase 1 and mediates the methyl jasmonate enhanced repression of sscd1 cell death by Phe. These results uncovered the relation between chlorophyll biosynthesis, phenylpropanoid pathway and jasmonate signaling in regulating the cell death resulting from loss of FAH in plants.
Subject terms: Jasmonic acid, Plant molecular biology, Plant physiology
Introduction
Tyrosine (Tyr) degradation pathway includes the five-step enzymatic reactions, in which Tyr is first converted to 4-hydroxyphenylpyruvate by Tyr aminotransferase, then transformed into homogentisate by 4-hydroxyphenylpyruvate dioxygenase. Next, homogentisate dioxygenase catalyzes homogentisate to form maleylacetoacetate (MAA) that is isomerized by MAA isomerase to fumarylacetoacetate (FAA). Last, FAA is hydrolyzed by fumarylacetoacetate hydrolase (FAH) to fumarate and acetoacetate1,2. Tyr degradation is essential to animals, blockage of this pathway results in metabolic disorder diseases, in which the most severe disorder in humans is hereditary tyrosinemia type I (HT1), an inborn lethal disease caused by deficiency of FAH3,4. Loss of FAH in HT1 patients results in the accumulation of Tyr degradation intermediates including FAA and MAA, and then both would undergo spontaneous reduction to succinylacetoacetate followed by spontaneous nonenzymatic decarboxylation to succinylacetone (SUAC) toxic to cells and tissues3. Although the homologous genes putatively encoding homogentisate dioxygenase, MAA isomerase, and FAH were demonstrated to exist in plants2,5, the role of the Tyr degradation pathway in plants had been unclear until we cloned the short-day sensitive cell death 1 (SSCD1) gene encoding an Arabidopsis putative FAH6. Loss of FAH in the sscd1 mutant leads to spontaneous cell death under short-day conditions (SD)6 and the accumulation of SUAC7. SUAC inhibits the activity of δ-aminolevulinic acid dehydratase involved in Chlorophyll (Chl) biosynthesis, resulting in high production of the Chl biosynthesis intermediate protochlorophyllide (Pchlide) in the dark under SD8. The excessive accumulation of Pchlide induces the production of reactive oxygen species (ROS) upon light irradiation and thereby causes cell death8.
Phenylalanine (Phe) is catalyzed by Phe hydroxylase to Tyr and then degraded in animals, and the dietary restriction of Tyr and Phe can improve the condition of HT1 patients4,9. In plants, Phe could also be converted into Tyr and then via homogentisate, to plastoquinones and tocopherols, or to degradation of the aromatic ring10. However, Phe can carry through phenylpropanoid metabolism to produce secondary metabolites10. Phenylpropanoids are precursors to flavonoids, isoflavonoids, cumarins, and stilbenes, which have important functions in plant defense against pathogens and other predators, as UV light protectants, and as regulatory molecules in signal transduction and communication with other organisms11. In phenylpropanoid pathway, the first step is that Phe ammonia-lyase (PAL) catalyzes the deamination of Phe to give cinnamic acid12,13. The α-aminooxi-β-phenylpropionic acid (AOPP) is an inhibitor of PAL, treatment with AOPP could reduce the activity of PAL14–16. Jasmonates (JAs) including jasmonic acid, methyl jasmonate (MeJA), and other derivatives, are a basic class of plant hormones involved in plant growth, development, and responses to biotic and abiotic stresses17–20. Treatment with MeJA could increase the activity of PAL in plants such as Chinese bayberry21, wheat22, and tuberose16. The PAL gene expression is responsive to a variety of environmental stimuli including pathogen infection, wounding, nutrient depletion, UV irradiation, extreme temperatures, and other stress conditions23–26.
To investigate whether the uptake of Phe in plants increases the cell death caused by loss of FAH as it does in animals, in this study the sscd1 mutant was treated with Phe and it was found that the death of sscd1 seedlings is not increased but suppressed by Phe treatment. With further investigation, we found that Phe treatment promotes Chl biosynthesis and represses the up-regulation of ROS marker genes in the sscd1 mutant. Furthermore, the repression of sscd1 cell death by Phe is reduced by AOPP whereas enhanced by MeJA. In addition, MeJA up-regulates the PAL1 gene and enhances the repression of sscd1 cell death by Phe through JA signaling. Our study shows that Phe has different effects on the cell death caused by FAH loss in plants and animals and uncovers the relation between Chl biosynthesis, phenylpropanoid pathway, and JA signaling in regulating the cell death resulting from loss of FAH, which will help to further investigate the regulation of Tyr degradation, phenylpropanoid biosynthesis and the cell death in plants.
Results
Phe treatment suppresses the death of sscd1 seedlings
Since Phe could be converted into Tyr and then degraded in plants10, we wondered whether Phe treatment promotes the death of sscd1 seedlings. To this end, the seeds of wild type and the sscd1 mutant were plated on MS medium without or with 0.1, 0.5, 1 and 2 mM Phe and grown under SD. Unexpectedly, the death of sscd1 seedlings treated with Phe was not increased, on the contrary, it was reduced (Fig. 1). When the medium was supplemented with 0.1 mM Phe, the rate of death seedlings was slightly reduced compared to that without Phe, however, it was significantly reduced as concentrations of Phe were increased (Fig. 1a). For example, on the 7th day, more than 80% of sscd1 seedlings in the medium without Phe were dead, however, when the medium was supplemented with 0.5, 1, and, 2 mM Phe, the rates of death seedlings were only about 70%, 30%, and 5%, respectively (Fig. 1a). The phenotype of seedlings shown in Fig. 1b clearly displayed that Phe treatment reduced the death of sscd1 seedlings. The sscd1 seedlings are normal under long-day conditions (LD), however, once transferred to SD and grown for several days, some leaves of sscd1 seedlings are dead and show bleaching, but the seedlings would not die6. To test whether the death of sscd1 seedling leaves is also weakened by Phe, the seedlings growing under LD were treated with Phe and then transferred to SD. As shown in Fig. 1c, when sscd1 seedlings growing in the medium with Phe under LD were transferred to SD, the extent of seedling leaf death was obviously attenuated. All these results indicated that Phe treatment suppresses the death of sscd1 seedlings.
Figure 1.

The death of sscd1 seedlings was suppressed by treatment with Phe. (a) The death rate of sscd1 seedlings which were grown in medium added without (control) or with 0.1, 0.5, 1, or 2 mM Phe under SD for 5–9 days. Error bars represent standard deviations (n > 30). The experiment was performed in three independent biological repeats. The single asterisk and double asterisks represent the significance of differences compared to the control (two-tailed Student’s t-test) at the level of P < 0.05 and P < 0.01, respectively. (b) The phenotype of WT and sscd1 seedlings which were grown in medium added without (control) or with 0.5, 1 or 2 mM Phe under SD for 7 days. (c) The phenotype of WT and sscd1 seedlings which were first grown in MS medium under LD for 7 days and transplanted to MS medium added without (control) or with 2 mM Phe for an additional 7-d growth under LD, and then transferred to SD for a 7-day growth. Phe phenylalanine; SD short day; WT wild type, Col-0; LD long day.
The up-regulation of ROS marker genes in sscd1 could be repressed by Phe treatment
Previously, we have speculated that the ROS resulting from excessive accumulation of Pchlide causes the sscd1 cell death8. Because ROS marker genes such as ascorbate peroxidase 2 (APX2)27, oxidative signal inducible 1 (OXI1)28,29, bonzai1-associated protein 1 (BAP1), and transcription factor (ZP)30 were up-regulated in the sscd1 mutant8,31, we next investigated whether the repression of sscd1 cell death by Phe is correlated with the expression level of these genes. As shown in Fig. 2, the expression levels of APX2, OXI1, BAP1, and ZP in the sscd1 mutant were significantly increased compared to that in wild type, however, they were clearly reduced after Phe treatment. Therefore, the up-regulation of ROS marker genes in sscd1 could be repressed by Phe treatment. Since the expression level of ROS marker genes is positively correlated to the content of ROS27,29,30, the repression of the up-regulation of ROS marker genes in sscd1 by Phe treatment (Fig. 2) indicated the reduction of ROS after Phe treatment.
Figure 2.

Expression of ROS marker genes in sscd1 was down-regulated by treatment with Phe. Relative expression levels of APX2, OXI1, BAP1 and ZP in WT and sscd1 seedlings which were first grown in MS medium under LD for 7 days and transplanted to MS medium added without (−) or with (+) 2 mM Phe under LD for an additional 7-day growth, and then transferred to SD for a 3-d growth. ACTIN2 expression was used as the internal control. Each value is the mean of three independent biological replicates ± standard deviation. An asterisk represents the significance of differences (two-tailed Student’s t-test) at the level of P < 0.05. WT wild type, Col-0; Phe phenylalanine; LD long day; SD short day.
Chl biosynthesis could be promoted by Phe treatment
Since the sscd1 cell death is mediated by Chl biosynthetic pathway8, we next investigated whether the Phe treatment influences Chl biosynthesis. We first determined the content of Chl and found that it was increased after Phe treatment (Fig. 3a). In the Chl biosynthesis pathway, there are two pivotal control points, one is the formation of the initial precursor, δ-aminolevulinic acid, by glutamyl-tRNA reductase, and another is the metal-ion insertion step by Mg-chelatase32–34. In Arabidopsis, the HEMA1 gene encodes the glutamyl-tRNA reductase and the CHLH gene encodes the H subunit of Mg-chelatase32,34. The transcriptional regulation of both HEMA1 and CHLH could affect Chl biosynthesis35. Thus, we next tested whether the transcription of these genes changes after Phe treatment by RT-qPCR. As we expected, the expression levels of HEMA1 and CHLH in both wild type and sscd1 were also increased after Phe treatment (Fig. 3b). Therefore, Phe treatment promotes Chl biosynthesis.
Figure 3.

Chlorophyll synthesis was increased after Phe treatment. (a) The content of chlorophyll in WT and sscd1 seedlings which were first grown in MS medium under LD for 7 days and transplanted to MS medium added without (−) or with (+) 2 mM Phe under LD for an additional 7-day growth, and then transferred to SD for a 3-day growth. (b) The relative expression levels of CHLH and HEMA1 in WT and sscd1 seedlings which were first grown in MS medium under LD for 7 days and transplanted to MS medium added without (−) or with (+) 2 mM Phe under LD for additional 7-d growth, and then transferred to SD for a 3-d growth. ACTIN2 expression was used as the internal control. Each value is the mean of three independent biological replicates ± standard deviation. An asterisk represents the significance of differences (two-tailed Student’s t-test) at the levels of P < 0.05. WT wild type, Col-0; LD long day; Phe phenylalanine; SD short day.
Repression of the sscd1 cell death by Phe could be reduced by AOPP and enhanced by MeJA
Phe is a precursor of phenylpropanoid biosynthesis and PAL catalyzes the first step of this pathway36. If the repression of sscd1 cell death by Phe is related to phenylpropanoid pathway, it would be changed by inhibition or activation of PAL. To confirm that, firstly, the sscd1 seedlings were treated with AOPP, a potent inhibitor of PAL15, on the basis of Phe treatment. As we expected, treated with Phe and AOPP, the death sscd1 seedlings were clearly increased compared to that treated with Phe alone (Fig. 4a). For example, the death rate of 7-d-old sscd1 seedlings treated with 1 mM Phe was approximately 30%, however, when seedlings were treated with 1 mM Phe and 100 μM AOPP, it was increased to approximately 72% (Fig. 4b). These results indicated that the inhibition of PAL activity by AOPP could reduce repression of the sscd1 cell death by Phe.
Figure 4.

Treatment with AOPP reduced the inhibitory effect of Phe on the sscd1 seedlings death. (a) The phenotype of 7-d-old wild-type WT and sscd1 seedlings which were first grown in the medium added without (control) or with 1 mM Phe under SD for 2 days and then the seedlings in 1 mM Phe were treated with ddH2O (1 mM Phe) or 100 μM AOPP (1 mM Phe + 100 μM AOPP) for an additional 5-d growth. (b) The death rate of 7-d-old sscd1 seedlings which were first grown in medium added without (control) or with 1 mM Phe under SD for 2 days and then the seedlings in 1 mM Phe were treated with ddH2O (1 mM Phe) or 100 μM AOPP (1 mM Phe + 100 μM AOPP) for an additional 5-d growth. Error bars represent standard deviations (n > 30). The experiment was performed in three independent biological repeats. An asterisk represents the significance of differences (two-tailed Student’s t-test) at the levels of p < 0.05. WT wild type, Col-0; Phe phenylalanine; SD short day.
Since the activity of PAL could be activated by MeJA16, we next investigated whether treatment with MeJA enhances the repression of sscd1 cell death by Phe. As shown in Fig. 5a, in the absence of Phe, treatment with 5 μM MeJA did not distinctly affect the phenotype of both wild type and sscd1 seedlings, however, after treatment with 5 μM MeJA and 0.5 mM Phe, the death seedlings of sscd1 obviously reduced compared to those only treated with 0.5 mM Phe. The death rate of 7-d-old sscd1 seedlings treated with 0.5 mM Phe was approximately 64% whereas it was less than 40% once treated with 5 μM MeJA and 0.5 mM Phe (Fig. 5b). Therefore, the activation of PAL activity by MeJA could enhance repression of the sscd1 cell death by Phe.
Figure 5.
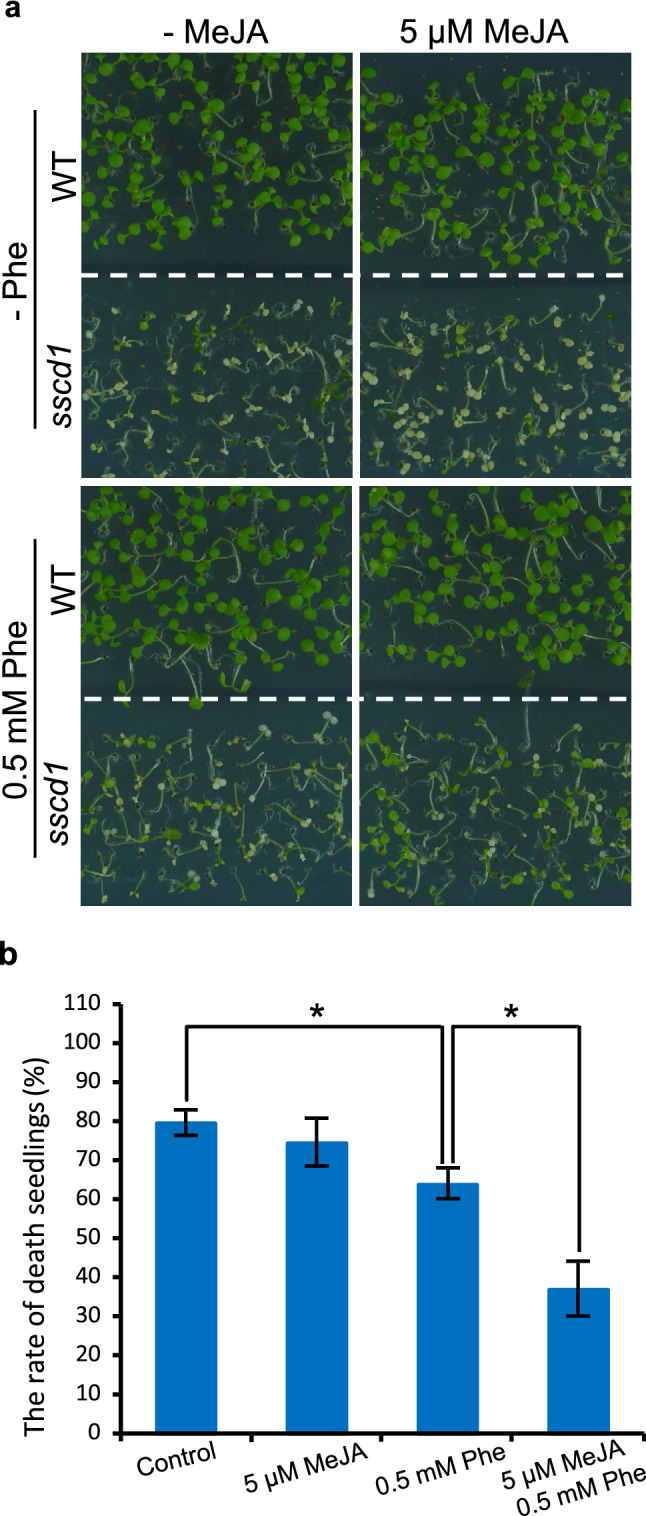
Treatment with MeJA enhanced the inhibitory effect of Phe on the sscd1 seedlings death. (a) The phenotype of WT and sscd1 seedlings which were grown in medium added without (−) or with 0.5 mM Phe or/and 5 μM MeJA under SD for 7 days. (b) The death rate of sscd1 seedlings which were grown in medium added without (control) or with 5 μM MeJA or/and 0.5 mM Phe under SD for 7 days. Error bars represent standard deviations (n > 30). The experiment was performed in three independent biological repeats. An asterisk represents the significance of differences (two-tailed Student’s t-test) at the level of P < 0.05. WT wild type, Col-0; Phe phenylalanine; MeJA methyl jasmonate; SD short day.
Treatment with MeJA causes the COI1-dependent up-regulation of PAL1
In Arabidopsis, PAL is encoded by a small gene family including PAL1, PAL2, PAL3, and PAL426,37. We next investigated whether treatment with MeJA affects some or all of these genes’ expression, and if so, is it dependent on COI117, a JA receptor of JA signaling38? To this end, the seedlings of wild type and the coi1-2 mutant, a coi1 leaky mutant39, were treated with MeJA and the expression levels of PAL1, PAL2, PAL3, and PAL4 were assessed by an analysis of RT-qPCR. As shown in Fig. 6, after treatment with MeJA, the expression level of PAL1 was significantly increased in wild type but not in coi1-2. However, the expression levels of PAL2, PAL3, and PAL4 were not significantly altered in both wild type and coi1-2 after being treated with MeJA (Fig. 6). These results suggested that treatment with MeJA up-regulates PAL1, and this up-regulation is dependent on COI1.
Figure 6.

An analysis of expression of the PAL genes upon MeJA treatment by RT-qPCR. The relative expression levels of PAL1, PAL2, PAL3 and PAL4 in WT and coi1-2 seedlings which were first grown in medium under LD for 12 days and removed to SD for a 3-d growth, and then treated with ddH2O (−) or 100 μM MeJA (+) for 1 day. ACTIN2 expression was used as the internal control. Each value is the mean of three independent biological replicates ± standard deviation. An asterisk represents the significance of differences (two-tailed Student’s t-test) at the levels of P < 0.05. WT wild type, Col-0; MeJA methyl jasmonate; LD long day; SD short day.
MeJA’s enhancement of Phe’s inhibition of sscd1 cell death depends on COI1
Since treatment with MeJA enhanced the repression of sscd1 cell death by Phe (Fig. 5) as well as up-regulated PAL1 in dependence of COI1 (Fig. 6), we next investigated whether the enhancement of MeJA on Phe in inhibiting the sscd1 cell death is also dependent on COI1. The seeds of the sscd1 single mutant and the sscd1coi1 double mutant were plated on medium added without or with 0.5 mM Phe or/and 5 μM MeJA and grown under SD, and then the death seedlings were counted. As shown in Fig. 7a, the death rate of sscd1coi1 seedlings was lower than that of sscd1, which is due to JA signaling positively regulating the sscd1 cell death31. Phe treatment also significantly reduced the death rate of sscd1coi1 seedlings (Fig. 7a, right), and that the reduction of seedling mortality was greater in sscd1coi1 (Fig. 7a, right) than in sscd1 (Fig. 7a, left), which is mainly resulted from that the blockage of JA signaling in sscd1coi1 suppresses the sscd1 cell death31 and Phe could also be degraded through the Tyr degradation pathway10. However, the death rate of sscd1coi1 seedlings treated with MeJA and Phe was not significantly reduced compared to that treated with Phe alone (Fig. 7a, right), which is unlike in sscd1 (Fig. 7a, left). From the Fig. 7b, we could see that MeJA treatment clearly enhances the repression of sscd1 seedlings death by Phe, however, this enhancement was not obvious in sscd1coi1 (Fig. 7b, right). Therefore, the MeJA’s enhancement of Phe’s inhibition of sscd1 cell death depends on COI1.
Figure 7.

MeJA-increased Phe inhibition of sscd1 seedlings death was dependent on COI1. (a) The death rate of sscd1 (left) and sscd1coi1 (right) seedlings which were grown in medium added without (Control) or with 5 μM MeJA or/and 0.5 mM Phe under SD for 6–8 days. Error bars represent standard deviations (n > 30). The experiment was performed in three independent biological repeats. An asterisk represents the significance of differences (two-tailed Student’s t-test) at the level of P < 0.05. (b) The phenotype of sscd1 and sscd1coi1 seedlings which were grown in medium added without (−) or with 0.5 mM Phe or/and 5 μM MeJA under SD for 8 days. MeJA methyl jasmonate; Phe Phenylalanine; SD short day.
Discussion
FAH catalyzes the final step of the Tyr degradation pathway and the deficiency of FAH in animals causes an inborn lethal disease, which was named HT1 in humans1,3. Phe could be converted to Tyr and then degraded in animals, and the dietary restriction of Tyr as well as Phe can improve the condition of HT1 patients4,9. In plants, the SSCD1 gene encodes the Arabidopsis FAH and the mutation of SSCD1 results in spontaneous cell death under SD6. Like as in animals, Phe could also be converted into Tyr in plants and then degraded10. However, in our study, the death of sscd1 seedlings was not increased but repressed by Phe treatment (Fig. 1). So, why would Phe treatment repress the cell death resulting from loss of FAH in plants?
Previously, we demonstrated that the sscd1 cell death is mediated by Chl biosynthesis8. The inhibition of the δ-aminolevulinic acid dehydratase activity by SUAC in the sscd1 mutant influences Chl biosynthesis resulting in impairment of feedback inhibition of Chl biosynthesis from the light–dark transition under SD, which activates Chl biosynthesis and accumulation of Pchlide in the dark, and then upon re-illumination the excessive accumulation of Pchlide induces the mass production of ROS and thereby causes cell death8. The main form of ROS induced by Pchlide is singlet oxygen, which is unstable and difficult to be quantitatively detected40, however, it could be assessed through the analysis of the expression levels of some genes including ZP and BAP1 activated specifically by singlet oxygen30. In this study, treatment of sscd1 seedlings with Phe distinctly repressed the up-regulation of ZP and BAP1 as well as other ROS marker genes such as APX2 and OXI1 (Fig. 2), In addition, we have tested hydrogen peroxide, another form of ROS, and found it was reduced in sscd1 once treated with Phe (Fig. S1). Therefore, ROS is reduced by Phe treatment. Treatment with Phe could promote Chl biosynthesis (Fig. 3). The increase in Chl biosynthesis would restore to some extent the feedback inhibition of Chl biosynthesis in the sscd1 mutant from the light–dark transition, as a result, reducing the accumulation of Pchlide in the dark and the production of ROS after subsequent exposure to light. Therefore, the increase of Chl biosynthesis should be one cause for the reduction of ROS and then the repression of cell death by Phe treatment.
In plants, Phe could be metabolized through the phenylpropanoid pathway to produce secondary metabolites, which plays an important role in plant against stress including UV-light, drought, and pathogen attack, due to their antioxidant function10,11,26,41–43. PAL catalyzes the first step of the phenylpropanoid pathway, which is a key step in phenylpropanoid biosynthesis13,34,44. The activity of PAL could be inhibited by AOPP and promoted by MeJA16. Treatment with AOPP prevents the increase in resistance to B. cinerea due to the application of external Phe43. In our study, the repression of sscd1 seedlings death by Phe was reduced by AOPP (Fig. 4), however, it was enhanced by MeJA (Fig. 5), which suggested that the suppression of sscd1 cell death by Phe is related to the phenylpropanoid pathway. Catechins, a class of flavonoids produced from Phe through phenylpropanoid pathway, have antioxidant activity45. In our study, treatment with catechins also suppressed the death of sscd1 seedlings (Fig. S2). Since the secondary metabolites produced by Phe metabolism through the phenylpropanoid pathway have antioxidant function42, ROS could also be reduced by the metabolism of Phe through the phenylpropanoid pathway, which should be another important cause for the repression of sscd1 cell death by Phe.
Previously, we found that JA signaling is involved in the sscd1 cell death31. In the sscd1 mutant, the accumulation of SUAC results in the generation of ROS, which induces cell death as well as JA synthesis31. JA up-regulates the Tyr degradation pathway, producing more SUAC, which promotes cell death31. Once JA signaling is broken by mutation of COI1 encoding a JA receptor38, the up-regulation of Tyr degradation pathway by JA is eliminated, reducing the production of SUAC, as a result, the sscd1 cell death is repressed31. MeJA is an activator of PAL catalyzing the first step of phenylpropanoid pathway16. In this study, MeJA treatment markedly increased the expression level of PAL1 in wild type but not in the coi1-2 mutant (Fig. 6), indicating that JA signaling can up-regulates the phenylpropanoid pathway through activating PAL1. The repression of sscd1 cell death by Phe could be enhanced by MeJA treatment in the sscd1 mutant (Figs. 5 and 7) but not in the sscd1coi1 mutant (Fig. 7), which suggested that MeJA treatment enhances Phe inhibition of the sscd1 cell death through JA signaling. Therefore, JA has a dual regulatory effect on the sscd1 cell death. On the one hand, JA up-regulates the Tyr degradation pathway, promoting the sscd1 cell death, on the other hand, JA up-regulates the phenylpropanoid pathway, inhibiting the sscd1 cell death. For this reason, the death of sscd1 seedlings was not increased by MeJA treatment, it seemed to decrease slightly (Figs. 5 and 7), which suggested that the effect of MeJA treatment on the sscd1 cell death through the phenylpropanoid pathway might be greater than that through Tyr degradation pathway.
In addition, we observed that with Phe treatment alone the decrease of death seedlings in sscd1coi1 is greater than that in sscd1 at 8 days (Fig. 7). One main reason for that should be that the blockage of JA signaling in sscd1coi1 decreases the seedlings death because JA signaling positively regulates the sscd1 cell death31. Another main reason for that should be that Phe could also be degraded through the Tyr degradation pathway10, inducing production of more ROS and subsequent cell death, however, blockage of JA signaling in sscd1coi1 could suppress the sscd1 cell death31. As a result, with Phe treatment alone the suppression of the seedlings death in the double mutant sscd1coi1 should be more obvious than that in the single mutant sscd1.
In conclusion, although Phe can be degraded through the Tyr degradation pathway, unlike in animals, Phe treatment does not increase the cell death resulting from loss of FAH in plants, instead, it represses the cell death. A possible mechanism for the repression of sscd1 cell death by Phe treatment can be described as follows (Fig. 8). Loss of FAH in the sscd1 mutant results in a decline of Chl biosynthesis, which impairs the feedback inhibition of Chl biosynthesis from light–dark transition under SD, leading to the accumulation of ROS and then cell death. Phe treatment, on the one hand, promotes Chl biosynthesis, increasing the feedback inhibition of Chl biosynthesis from light–dark transition under SD, and on the other hand, activates the phenylpropanoid pathway, both of which reduce ROS and subsequent cell death. In addition, in the sscd1 mutant ROS induces cell death as well as JA synthesis. JA signaling up-regulates the Tyr degradation pathway, promoting the sscd1 cell death, however, it also up-regulates PAL1 which activates the phenylpropanoid pathway, repressing the sscd1 cell death. Since the effect of MeJA treatment on the sscd1 cell death through the phenylpropanoid pathway might be greater than that through the Tyr degradation pathway, the repression of sscd1 cell death by Phe could be enhanced by MeJA treatment.
Figure 8.

The possible mechanisms by which Phe treatment represses the sscd1 cell death. The SSCD1 gene encodes FAH catalyzing the last step of Tyr degradation pathway. Disruption of FAH in sscd1 reduces Chl biosynthesis, which impairs the feedback inhibition of Chl biosynthesis from light–dark transition under SD, leading to the accumulation of ROS and then cell death (red lines and arrows). Phe treatment promotes Chl biosynthesis, which increases the feedback inhibition of Chl biosynthesis from light–dark transition under SD, reducing ROS and subsequent cell death (green arrows and lines). Meanwhile, Phe treatment activates phenylpropanoid pathway, which reduces ROS and subsequent cell death (blue arrows and lines). In the sscd1 mutant, ROS induces cell death and the JA production. JA up-regulates Tyr degradation pathway through COI1, promoting cell death, however, it also up-regulates PAL1 through COI1, which activates phenylpropanoid pathway, repressing cell death. The effect of MeJA treatment on sscd1 cell death through phenylpropanoid pathway might be greater than that through Tyr degradation pathway, resulting in that MeJA treatment enhances the repression of sscd1 cell death by Phe. Arrows indicate induction or positive regulation, whereas lines indicate repression or negative regulation. Arrows with dashed lines indicate multiple steps. FAH fumarylacetoacetate hydrolase; Chl chlorophyll; SD short day; ROS reactive oxygen species; Phe phenylalanine; JA jasmonates; MeJA methyl jasmonate; HGA homogentisate; FAA fumarylacetoacetate; FA fumarate; AA acetoacetate.
Methods
Plant materials and growth conditions
Arabidopsis thaliana L. ecotype Columbia-0 (Col-0) was obtained from the Arabidopsis Biological Resource Center (ABRC; Ohio State University, Columbus, OH, USA) and the mutants used in this study are in Col-0 background. The sscd1 mutant6 was isolated by Han et al. in our laboratory. The coi1-2 mutant39 was kindly provided by Professor Xie (Tsinghua University, Beijing, China) and the sscd1coi1 double mutant31 was generated through a cross of sscd1 with coi1-2 by Zhou et al. in our laboratory. Experimental research on plants including the collection of plant material was performed in accordance with relevant institutional, national, and international guidelines and legislation.
Seeds were surface sterilized with 20% (v/v) chlorine bleach containing 0.1% (v/v) Triton X-100 for 10 min and washed three to five times with sterile water, then plated on Murashige & Skoog medium supplemented with 1% (m/v) sucrose and 0.7% (w/v) agar (pH 5.8) (MS). The different concentrations of Phe (SIGMA) were added to the MS medium. Plates were chilled at 4 °C in darkness for 3 days and then transferred to a growth chamber with LD (16-h light/8-h dark) or SD (8-h light/16-h dark) under 150 μmol photons m−2 s−1, controlled temperature (22 ± 2 °C).
For RT-qPCR analysis (Figs. 2, 3b) and determination of Chl content (Fig. 3a), 1-week-old seedlings growing under LD were transplanted onto MS added without or with Phe and grown under LD for an additional 1 week’s growth, and then transferred to SD for three days growth. Since the cell death of sscd1 seedlings occurs on the 4th day after transferred from LD to SD6, the seedlings were harvested at the end of the third day's light for the determination of Chl content or harvested at 2 h light after three days for RT-qPCR analysis.
Determination of the dead seedlings
A dead seedling is one for which all leaves were completely bleached. The rate of seedling death was calculated as the percentage of dead seedlings. The number of seedlings counted was approximately 100, and the experiment was performed in three independent biological repeats.
RT-qPCR analysis
Total RNA was isolated using TRIZOL reagent (Life Technologies, https://www.thermofisher.com/us/en/home/brands/life-technologies.html). After incubation with DNase I (RNase Free, Thermo Fisher Scientific, https://www.thermofisher.com/) at 37 °C for 30 min and then at 65 °C for 10 min to remove genomic DNA, the RNA concentration and purity were measured spectrophotometrically using OD260/OD280 and OD260/OD230 ratios (ND-1000, NanoDrop, THERMO FISHER SCIENTIFIC). Complementary DNA was synthesized from the mixture of oligo-dT primers and random primers using a ReverTraAce qPCR RT kit (perfect real time) according to the manufacturer’s instructions (Toyobo, http://www.toyobo-global.com/).
RT-qPCR was performed in 96-well blocks using a SYBR qPCR mix (Roche, https://lifescience.roche.com/) with a Bio-Rad CFX Connect™ Real-Time PCR detection system (http://www.biorad.com/) following the manufacturer’s instructions. The RT-qPCR amplifications were performed under the following conditions: initial denaturation at 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 s and 60 °C for 60 s. The primers of genes tested by RT-qPCR are listed in Table 1, and ACTIN2 was used as an internal control. The gene expression for each sample was calculated on three analytical replicates, and the relative expression was quantified using the 2−△△Ct method. The experiment was performed in three independent biological repeats. The significance of differences between datasets was evaluated using the two-tailed Student’s t-test.
Table 1.
Primers of genes tested by real-time quantitative PCRs.
| Gene | Forward primer | Reverse primer |
|---|---|---|
| APX2 (AT3G09640) | 5'-ACAAAGTTGAGCCACCTCCT-3' | 5'-AAGGTGTGTCCACCAGACAA-3' |
| OXI1 (AT3G25250) | 5'-GTTGAGGAAATCAAGGGTCATG-3' | 5'-TGGACGATATTCTCCACATCC-3' |
| ZP (AT5G04340) | 5'-TACGAAGGAAAGAACGGAGGC-3' | 5'-GGTATCGGCGGTATGTTGAGG-3' |
| CHLH (AT5G13630) | 5'-CAGCCAACATCAGTCTTGCT-3' | 5'-ACCTGCTTCTTCTCAGCCAT-3' |
| BAP1 (AT3G61190) | 5'-ATCGGATCCCACCAGAGATTACGG-3' | 5'-AATCTCGGCCTCCACAAACCAG-3' |
| HEMA1 (AT1G58290) | 5'-GTTGCTGCCAACAAAGAAGA-3' | 5'-AATCCCTCCATGCTTCAAAC-3' |
| PAL1 (AT2G37040) | 5'-TTTTGGTGCTACTTCTCATCG-3' | 5'-CTTGTTTCTTTCGTGCTTCC-3' |
| PAL2 (AT2G37040) | 5'-GTGCTACTTCTCACCGGAGA-3' | 5'-TATTCCGGCGTTCAAAAATC-3' |
| PAL3 (AT5G04230) | 5'-CAACCAAACGCAACAGCA-3' | 5'-CTCCAGGTGGCTCCCTTTTA-3' |
| PAL4 (AT3G10340) | 5′-GGTGCACTTCAAAATGAGCT-3′ | 5′-CAACGTGTGTGACGTGTCC-3' |
| ACTIN2 (AT3G18780) | 5′-AGCACTTGCACCAAGCAGCATG-3′ | 5′-ACGATTCCTGGACCTGCCTCATC-3′ |
Determination of Chl content
The content of Chl was determined referring to the method described by Lichtenthaler46. Weighed segments of frozen crushed material (about 0.04 g) were homogenized in 1 mL 80% acetone and stood for 5–6 h, then centrifuged for 10 min at 5000 rpm at 4 °C and assayed spectrophotometrically at 663 nm and 645 nm. Result calculation: C(mg/g) = (17.32 A663 + 7.18 A645)/m(g). The experiment was performed in three independent biological repeats.
AOPP treatment
Seeds were germinated on MS added with 1 mM Phe (SIGMA) and grown under SD. On the third day seedlings were sprayed with 100 μM AOPP (Wako) or ddH2O (as a control) once a day for 5 days, then the rate of death seedlings was counted, and the seedlings were photographed. The experiment was performed in three independent biological repeats.
MeJA treatment
For determination of dead seedlings (Figs. 5 and 7), seeds were germinated on MS added without or with 0.5 mM Phe and/or 5 μM MeJA (SIGMA) and grown under SD for 6–8 days. For RT-qPCR analysis (Fig. 6), about 2-week-old seedlings growing under LD were transferred to SD and on the fourth day the seedlings were sprayed with 100 μM MeJA or ddH2O (as a control). After MeJA treatment for one day, the seedlings were harvested and used for RT-qPCR analysis. The experiment was performed in three independent biological repeats.
Supplementary Information
Acknowledgements
This work was supported by Grants from the Program for Key Basic Research of the Ministry of Science and Technology of China (2014CB160308) and the National Science Foundation of China (30671121 and 31571802).
Author contributions
C.R. conceived, designed and supervised the experiments; Y.J. and T.Z. performed the experiments; Q.Z. and H.Y. provided technical assistance to Y.J.; Y.J., Q.Z. and C.R. analyzed the data; Y.J. wrote the article with contributions from all authors; and C.R. supervised, modified and complemented the writing.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-022-17819-3.
References
- 1.Grompe M, et al. Loss of fumarylacetoacetate hydrolase is responsible for the neonatal hepatic dysfunction phenotype of lethal albino mice. Genes Dev. 1993;7:2298–2307. doi: 10.1101/gad.7.12a.2298. [DOI] [PubMed] [Google Scholar]
- 2.Dixon DP, Edwards R. Enzymes of tyrosine catabolism in Arabidopsis thaliana. Plant Sci. 2006;171:360–366. doi: 10.1016/j.plantsci.2006.04.008. [DOI] [PubMed] [Google Scholar]
- 3.Lindblad B, Steen SL. On the enzymic defects in hereditary tyrosinemia. Proc. Natl. Acad. Sci. USA. 1977;74:4641–4645. doi: 10.1073/pnas.74.10.4641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.St-Louis M, Tanguay RM. Mutations in the fumarylacetoacetate hydrolase gene causing hereditary tyrosinemia type I: Overview. Hum. Mutat. 1997;9:291–299. doi: 10.1002/(SICI)1098-1004(1997)9:4<291::AID-HUMU1>3.0.CO;2-9. [DOI] [PubMed] [Google Scholar]
- 5.Dixon DP, Cole DJ, Edwards R. Characterisation of a zeta class glutathione transferase from Arabidopsis thaliana with a putative role in tyrosine catabolism. Arch. Biochem. Biophys. 2000;384:407–412. doi: 10.1006/abbi.2000.2125. [DOI] [PubMed] [Google Scholar]
- 6.Han C, et al. Disruption of fumarylacetoacetate hydrolase causes spontaneous cell death under short-day conditions in Arabidopsis. Plant Physiol. 2013;162:1956–1964. doi: 10.1104/pp.113.216804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Zhou L, et al. A GC/MS method for determination of succinylacetone in Arabidopsis thaliana. Anal. Bioanal. Chem. 2016;408:4661–4667. doi: 10.1007/s00216-016-9551-7. [DOI] [PubMed] [Google Scholar]
- 8.Zhi T, et al. Loss of fumarylacetoacetate hydrolase causes light-dependent increases in protochlorophyllide and cell death in Arabidopsis. Plant J. 2019;98:622–638. doi: 10.1111/tpj.14235. [DOI] [PubMed] [Google Scholar]
- 9.Phaneuf D, et al. Cloning and expression of the cDNA encoding human fumarylacetoacetate hydrolase, the enzyme deficient in hereditary tyrosinemia: assignment of the gene to chromosome 15. Am. J. Hum. Genet. 1991;48:525–535. [PMC free article] [PubMed] [Google Scholar]
- 10.Pribat A, et al. Nonflowering plants possess a unique folate-dependent phenylalanine hydroxylase that is localized in chloroplasts. Plant Cell. 2010;22:3410–3422. doi: 10.1105/tpc.110.078824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ferrer JL, Austin MB, Stewart C, Jr, Noel JP. Structure and function of enzymes involved in the biosynthesis of phenylpropanoids. Plant Physiol. Biochem. 2008;46:356–370. doi: 10.1016/j.plaphy.2007.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Koukol J, Conn EE. The metabolism of aromatic compounds in higher plants. IV. Purification and properties of the phenylalanine deaminase of hordeum vulgare. J. Biol. Chem. 1961;236:2692–2698. doi: 10.1016/S0021-9258(19)61721-7. [DOI] [PubMed] [Google Scholar]
- 13.Bate NJ, et al. Quantitative relationship between phenylalanine ammonia-lyase levels and phenylpropanoid accumulation in transgenic tobacco identifies a rate-determining step in natural product synthesis. Proc. Natl. Acad. Sci. USA. 1994;91:7608–7612. doi: 10.1073/pnas.91.16.7608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ferrante A, Alberici A, Antonacci S, Serra G. Effect of promoter and inhibitors of phenylalanine ammonia-lyase enzyme on stem bending of cut gerbera flowers. Acta. Hortic. 2007;755:471–476. doi: 10.17660/ActaHortic.2007.755.64. [DOI] [Google Scholar]
- 15.Amrhein N, GöDeke KH. L-α-Aminooxy-β-phenyl-propionic acid–a potent inhibitor of L-phenylalanine ammonia-lyase in vitro and in vivo. Plant Sci. Lett. 1977;8:313–317. doi: 10.1016/0304-4211(77)90148-1. [DOI] [Google Scholar]
- 16.Kanani M, Nazarideljou MJ. Methyl jasmonate and α-aminooxi-β-phenyl propionic acid alter phenylalanine ammonia-lyase enzymatic activity to affect the longevity and floral scent of cut tuberose. Hortic. Environ. Biotechnol. 2017;58:136–143. doi: 10.1007/s13580-017-0055-y. [DOI] [Google Scholar]
- 17.Xie D, Feys BF, James S, Nieto-Rostro M, Turner JG. COI1: An Arabidopsis gene required for jasmonate-regulated defense and fertility. Science. 1998;280:1091–1094. doi: 10.1126/science.280.5366.1091. [DOI] [PubMed] [Google Scholar]
- 18.Wasternack C. Jasmonates: An update on biosynthesis, signal transduction and action in plant stress response, growth and development. Ann. Bot. 2007;100:681–697. doi: 10.1093/aob/mcm079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Browse J. Jasmonate passes muster: A receptor and targets for the defense hormone. Annu. Rev. Plant. Biol. 2009;60:183–205. doi: 10.1146/annurev.arplant.043008.092007. [DOI] [PubMed] [Google Scholar]
- 20.Avanci NC, Luche DD, Goldman GH, Goldman MH. Jasmonates are phytohormones with multiple functions, including plant defense and reproduction. Genet. Mol. Res. 2010;9:484–505. doi: 10.4238/vol9-1gmr754. [DOI] [PubMed] [Google Scholar]
- 21.Wang K, et al. Methyl jasmonate reduces decay and enhances antioxidant capacity in Chinese bayberries. J. Agric. Food Chem. 2009;57:5809–5815. doi: 10.1021/jf900914a. [DOI] [PubMed] [Google Scholar]
- 22.Motallebi P, Niknam V, Ebrahimzadeh H, Enferadi ST, Hashemim M. The effect of methyl jasmonate on enzyme activities in wheat genotypes infected by the crown and root rot pathogen Fusarium culmorum. Acta. Physiol. Plant. 2015;37:237. doi: 10.1007/s11738-015-1988-3. [DOI] [Google Scholar]
- 23.Edwards K, et al. Rapid transient induction of phenylalanine ammonia-lyase mRNA in elicitor-treated bean cells. Proc. Natl. Acad. Sci. USA. 1985;82:6731–6735. doi: 10.1073/pnas.82.20.6731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Liang X, Dron M, Cramer CL, Dixon RA, Lamb CJ. Differential regulation of phenylalanine ammonia-lyase genes during plant development and by environmental cues. J. Biol. Chem. 1989;264:14486–14492. doi: 10.1016/S0021-9258(18)71704-3. [DOI] [PubMed] [Google Scholar]
- 25.Liang X, Dron M, Schmid J, Dixon RA, Lamb CJ. Developmental and environmental regulation of a phenylalanine ammonia-lyase-β-glucuronidase gene fusion in transgenic tobacco plants. Proc. Natl. Acad. Sci. USA. 1989;86:9284–9288. doi: 10.1073/pnas.86.23.9284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Huang J, et al. Functional analysis of the Arabidopsis PAL gene family in plant growth, development, and response to environmental stress. Plant Physiol. 2010;153:1526–1538. doi: 10.1104/pp.110.157370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Karpinski S, et al. Systemic signaling and acclimation in response to excess excitation energy in Arabidopsis. Science. 1999;284:654–657. doi: 10.1126/science.284.5414.654. [DOI] [PubMed] [Google Scholar]
- 28.Rentel MC, et al. OXI1 kinase is necessary for oxidative burst-mediated signalling in Arabidopsis. Nature. 2004;427:858–861. doi: 10.1038/nature02353. [DOI] [PubMed] [Google Scholar]
- 29.Shumbe L, et al. Singlet oxygen-induced cell death in Arabidopsis under high-light stress is controlled by OXI1 kinase. Plant Physiol. 2016;170:1757–1771. doi: 10.1104/pp.15.01546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.op den Camp RGL, et al. Rapid induction of distinct stress responses after the release of singlet oxygen in Arabidopsis. Plant Cell. 2003;15:2320–2332. doi: 10.1105/tpc.014662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Zhou Z, et al. Cell death resulted from loss of fumarylacetoacetate hydrolase in Arabidopsis is related to phytohormone jasmonate but not salicylic acid. Sci. Rep. 2020;10:13714. doi: 10.1038/s41598-020-70567-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Cornah JE, Terry MJ, Smith AG. Green or red: What stops the traffic in the tetrapyrrole pathway? Trends Plant Sci. 2003;8:224–230. doi: 10.1016/S1360-1385(03)00064-5. [DOI] [PubMed] [Google Scholar]
- 33.McCormac AC, Fischer A, Kumar AM, Söll D, Terry MJ. Regulation of HEMA1 expression by phytochrome and a plastid signal during de-etiolation in Arabidopsis Thaliana. Plant J. 2001;25:549–561. doi: 10.1046/j.1365-313x.2001.00986.x. [DOI] [PubMed] [Google Scholar]
- 34.Matsumoto F, et al. Gene expression profiling of the tetrapyrrole metabolic pathway in Arabidopsis with a mini-array system. Plant Physiol. 2004;135:2379–2391. doi: 10.1104/pp.104.042408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Masuda T, Fujita Y. Regulation and evolution of chlorophyll metabolism. Photochem. Photobiol. Sci. 2008;7:1131–1149. doi: 10.1039/b807210h. [DOI] [PubMed] [Google Scholar]
- 36.Rohde A, et al. Molecular phenotyping of the pal1 and pal2 mutants of Arabidopsis thaliana reveals far-reaching consequences on phenylpropanoid, amino acid, and carbohydrate metabolism. Plant Cell. 2004;16:2749–2771. doi: 10.1105/tpc.104.023705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Raes J, Rohde A, Christensen JH, Van de Peer Y, Boerjan PW. Genome-wide characterization of the lignification toolbox in Arabidopsis. Plant Physiol. 2003;133:1051–1071. doi: 10.1104/pp.103.026484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Yan J, et al. The Arabidopsis CORONATINE INSENSITIVE1 protein is a jasmonate receptor. Plant Cell. 2009;21:2220–2236. doi: 10.1105/tpc.109.065730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Xu L, et al. The SCFcoi1 ubiquitin-ligase complexes are required for jasmonate response in Arabidopsis. Plant Cell. 2002;14:1919–1935. doi: 10.1105/tpc.003368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hideg E, et al. Detection of singlet oxygen and superoxide with fluorescent sensors in leaves under stress by photoinhibition or UV radiation. Plant Cell Physiol. 2002;43:1154–1164. doi: 10.1093/pcp/pcf145. [DOI] [PubMed] [Google Scholar]
- 41.Nakabayashi R, et al. Enhancement of oxidative and drought tolerance in Arabidopsis by overaccumulation of antioxidant flavonoids. Plant J. 2014;77:367–379. doi: 10.1111/tpj.12388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Zhang Y, et al. Different reactive oxygen species scavenging properties of flavonoids determine their abilities to extend the shelf life of tomato. Plant Physiol. 2015;169:1568–1583. doi: 10.1104/pp.15.00836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Oliva M, et al. Increased phenylalanine levels in plant leaves reduces susceptibility to Botrytis cinerea. Plant Sci. 2020;290:110289. doi: 10.1016/j.plantsci.2019.110289. [DOI] [PubMed] [Google Scholar]
- 44.Sewalt V, et al. Reduced lignin content and altered lignin composition in transgenic tobacco down-regulated in expression of L-phenylalanine ammonia-lyase or cinnamate 4-hydroxylase. Plant Physiol. 1997;115:41–50. doi: 10.1104/pp.115.1.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Engelhardt UH. Chemistry of Tea. Reference Module in Chemistry, Molecular Sciences and Chemical Engineering. Elsevier; 2013. pp. 4–5. [Google Scholar]
- 46.Lichtenthaler HK. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987;148:350–382. doi: 10.1016/0076-6879(87)48036-1. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.