

Abstract
Massive recruitment of eosinophils into the dermis is a hallmark of bullous pemphigoid pathogenesis. Identifying the chemoattractant(s) guiding eosinophils into the skin in bullous pemphigoid is a prerequisite to therapeutic targeting of eosinophil recruitment. Galectin-9 is a potent chemoattractant for eosinophils, but its potential role in bullous pemphigoid is unknown. The aim of this study was to determine the expression levels of galectin-9 in serum and skin of patients with bullous pemphigoid. Galectin-9 levels were significantly elevated in serum of patients with bullous pemphigoid compared with age- and sex-matched controls, but did not correlate with disease activity assessed with the Bullous Pemphigoid Disease Area Index. Galectin-9 expression was also increased in lesional skin of patients with bullous pemphigoid, and was expressed predominantly in eosinophils, neutrophils and keratinocytes. In conclusion, these results support the notion that galectin-9 may play a role in the pathogenesis of bullous pemphigoid.
Key words: autoimmune disease, bullous pemphigoid, galectin-9, eosinophil
Bullous pemphigoid (BP) is the most common autoimmune bullous disease and belongs to the group of pemphigoid diseases (1). Pemphigoid diseases are defined by antibody-driven autoimmunity against different components of the dermal–epidermal adhesion complex. In BP, the hemidesmosomal protein BP180 (type XVII collagen) is targeted (1, 2). The deposition of autoantibodies at the dermal–epidermal junction alone does not initiate the eruption of skin inflammation, but requires recruitment of granulocytes into the dermis. The dermal inflammatory infiltrate is dominated by eosinophils (3, 4). Their massive recruitment into the dermis is, therefore, presumed to be crucial for the pathogenesis of BP (3), but the molecular networks regulating skin inflammation in BP are still only partially understood. This also pertains to the molecular cues orchestrating the recruitment of eosinophils. Their elucidation would be key to the development of therapeutic strategies suppressing skin inflammation by selectively disrupting the recruitment of eosinophils. On the molecular level, a predominance of TH2 cytokines and IL-17A in the peripheral blood and skin are established hallmarks of BP, which probably contribute significantly to its pathogenesis (5–7).
SIGNIFICANCE
Eosinophils are major culprit cells in bullous pemphigoid, but the mechanism of their recruitment into the skin is unknown. This study identified galectin-9 as a potential driver of eosinophil recruitment in bullous pemphigoid. The results also show that galectin-9 is a potential serum biomarker in bullous pemphigoid. This finding could lead to the development of composite biomarkers for the diagnosis of bullous pemphigoid from serum samples.
Galectin-9 (Gal-9) is a β-galactoside-binding lectin exhibiting diverse immunoregulatory activities through binding to T-cell-immunoglobulin- and mucin-domain-containing molecule-3 (TIM-3), CD44, and IgE (8). Binding of galectin-9 to Tim-3 on CD4 and CD8 T cells leads to induction of apoptosis. Furthermore, galectin-9 induces calcium entry and aggregation of TH1 cells. Via binding to CD44 it inhibits association with hyaluronan, and therefore prevents cells from rolling through CD44-dependent mechanisms (8). By binding to IgE, galectin-9 prevents formation of IgE immune complexes and reduces IgE-induced degranulation of mast cells (9).
Several immunoregulatory activities of galectin-9 may also play a role in shaping the inflammatory microenvironment in lesional skin of patients with BP. More specifically, galectin-9 potently chemoattracts eosinophils, induces their aggregation and superoxide production and can prolong their survival (8, 10–12). It also establishes a dominance of TH2 over TH1 and TH17 cytokines (13–15). In line with the activity profile, galectin-9 has been elevated and implicated as an important regulatory factor of chronic diseases exhibiting eosinophil and/or TH2 cell driven tissue inflammation, such as scleroderma, atopic dermatitis, rheumatoid arthritis, eosinophilic pneumonia, and allergic lung inflammation (15–19). However, the role of galectin-9 in BP has not been addressed.
To determine whether galectin-9 might play a role in the pathogenesis of BP, this study assessed the expression of galectin-9 in the serum of patients with BP compared with age- and sex-matched healthy controls. The study examined the correlation of galectin-9 levels with BPDAI disease activity score, age, autoantibody levels and various leucocyte subsets. In addition, the study tested the expression of galectin-9 in keratinocytes, eosinophils, and neutrophils in lesional tissue samples of patients and in healthy control skin.
METHODS
Patients
A total of 40 patients with BP and 40 age- and sex-matched controls were recruited at the University of Lübeck, Lübeck, Germany. Based on prior power calculations, 40 samples per group allowed a statistical power of >70% to be achieved with a type I error probability of 5% for a range of potential effect sizes (effect size 0.6/empirical power 72.7%; 0.7/0.851%; 0.8/93.3%). The minimum requirements for diagnosis of BP were a typical clinical presentation, the detection of linear depositions of IgG at the dermal–epidermal junction in the direct immunofluorescence and deposition of circulating IgG at the blister roof of salt-split skin in indirect immunofluorescence testing or binding to BP180-NC16A or BP230 detected by enzyme-linked immunoassay (ELISA). All patients included in the study were newly diagnosed with BP and had not received systemic immunosuppressive treatment prior to inclusion in the study. Collected serum samples were stored at –80 °C until usage. Skin biopsies were fixed in formalin, paraffin embedded, and stored at room temperature (RT) in darkness. Demographic, clinical, and serological characteristics of the patients are shown in Table I.
Table I.
Patient characteristics
| Characteristic | Patients with BP | Controls | p-value |
|---|---|---|---|
| n = 40 (50.0%) | n = 40 (50.0%) | ||
| Female, n (%) | 16 (40.0) | 16 (40.0) | |
| Male, n (%) | 24 (60.0) | 24 (60.0) | |
| Age, years, median (IQR) | 79.5 (73.5–82.0) | 79.0 (74.0–83.3) | 0.639 |
| Range | 66–89 | 66–93 | |
| BPDAI score, median (IQR) | 20 (10.5–39.5) | NA | |
| Range | 3–84 | NA | |
| BP180–NC16A (U/ml) | |||
| Tested, n (%) | 40 (100.0) | NA | |
| Positive, n (%) | 37 (92.5) | NA | |
| Median (IQR) | 167 (100–330) | NA | |
| Range | 30–3,412 | NA | |
| BP230–CF (U/ml) | |||
| Tested, n (%) | 21 (52.5) | NA | |
| Positive, n (%) | 13 (32.5) | NA | |
| Median (IQR) | 125 (42–157) | NA | |
| Range | 3–1,549 | NA | |
IQR: interquartile range; BP: bullous pemphigoid; NA: not applicable; BPDAI score: Bullous Pemphigoid Disease Area Index score.
Enzyme-linked immunosorbent assay
Serum galectin-9 levels were quantified using the Human Galectin-9 DuoSet ELISA kit (R&D Systems, Minneapolis, MI, USA, DY2045) according to the manufacturers’ instructions. Antibodies against BP180 were detected with Anti-BP180-NC16A-4XELISA (Euroimmun, EA 1502-1 G) and against BP230 with Anti-BP230-CF-ELISA (Euroimmun, Lübeck, Germany, EA 1502 G), according to the manufacturer’s instructions. Optical densities were measured at 450 nm using the Infinite M200Pro microplate reader (Tecan, Männedorf, Switzerland). Concentrations were calculated from the standard curve generated by the Megellan software version 7.2 (Tecan).
Fluorescence-labelled immunohistochemistry
Galectin-9 was visualized on BP lesional skin samples in combination with myeloperoxidase (MPO) as a neutrophil marker (n = 8) or eosinophil peroxidase (EPX) as an eosinophil marker (n = 5) to identify the site of expression. In comparison, their expression was also tested in skin from corresponding anatomical sites of age- and sex-matched healthy controls (n = 5). Briefly, 6-μm paraffin sections were deparaffinized and rehydrated. After heat antigen retrieval for 10 min at 100°C alone (MPO staining) or together with subsequent Proteinase K digestion for 3 min at RT (EPX staining), samples were blocked with donkey anti-human Fab (Jackson Immuno Research Laboratories, West Grove, PA, USA, #709-006-149) for 45 min at RT and 10% (v/v) normal donkey serum (NDS) for 45 min at RT. Samples were incubated with rabbit anti-mouse/rat/human Galectin-9 antibody (Abcam, Cambridge, MA, USA, #ab69630) overnight at 4°C. Next day, secondary AF594 conjugated donkey anti-rabbit antibody (Jackson Immuno Research Laboratories, #711-585-152) was applied for 45 min at RT. After another blocking step with 10% (v/v) normal donkey serum (NDS), samples were either incubated with mouse anti-human MPO (R&D Systems, 392105) for 60 min at RT or with mouse anti-mouse/human EPX (Lee Laboratory, Mayo Clinic, Scottsdale, AZ, USA, #MM25-82.2) overnight at 4°C. Subsequently, all samples were labelled with secondary AF488 conjugated donkey anti-mouse antibody (Jackson Immuno Research Laboratories, #715-545-151) for 45 min at RT. All in-between washing steps were performed with 0.01M PBST, pH 7.2. In the end, slides were mounted with DAPI fluoromount G (SouthernBiotech, Birmingham, Alabama, USA). Images were acquired on a BZ-9000E series microscope (Keyence GmbH, Neu-Isenburg, Germany) and analysed using the BZ II Analyzer software (Keyence GmbH). To generate larger insets, original 200× magnification images were digitally magnified 3×.
Ethics approval
All patients and healthy controls provided written informed consent prior to their inclusion in the study. Ethics approval was obtained from the institutional review board of the University of Lübeck prior to the study (ethics board number 18-046).
Statistical analyses
Analyses were performed using R 3.6.0 (RCore Team 2017, Vienna, Austria). Associations between categorical and numerical variables were tested using a Wilcoxon rank-sum test (2 categorical groups). The χ2 test of independence was used to analyse contingency tables formed by 2 categorical variables. Correlations between 2 numeric variables were tested using Spearman’s rank-order correlation. Prior to the study a power calculation was performed utilizing the MKpower package to simulate different potential effect and samples sizes for Wilcoxon rank-sum test via Monte-Carlo simulation. p ≤ 0.05 were considered statistically significant unless otherwise specified.
RESULTS
Serum levels of galectin-9 were measured by ELISA in the serum of 40 patients with BP and 40 age-and sex-matched controls. Galectin-9 was detectable in significant amounts in both patients and healthy controls (Fig. 1), but levels were elevated in the serum of patients with BP (2.65 ± 1.02 ng/ml) compared with healthy controls (1.95 ± 1.18 ng/ml; pWilxoc = 0.0002). There was a weak positive association of galectin-9 with the age of healthy controls (Rho = 0.39, p = 0.014), but no correlation of galectin-9 with the age of patients with BP at time of diagnosis (Fig. S11).
Fig. 1.
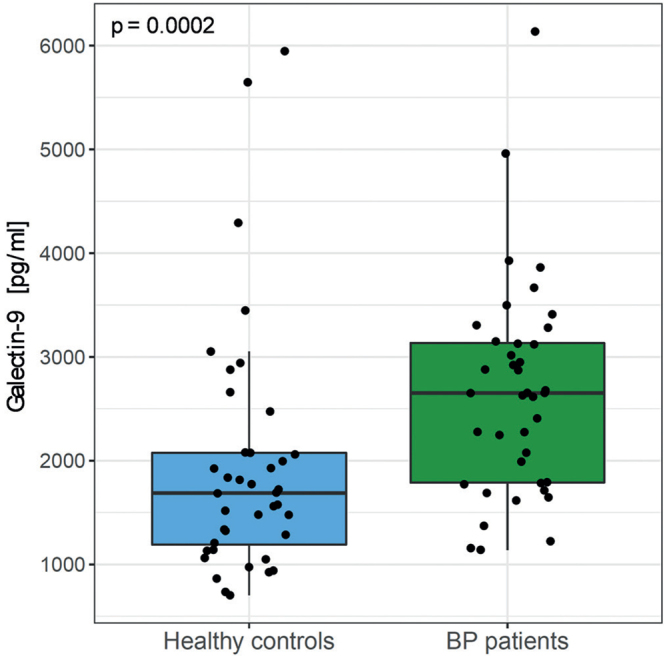
Patients with bullous pemphigoid (BP) have elevated galectin-9 serum levels. (a) Serum levels of galectin-9 measured by enzyme-linked immunoassay (ELISA) in patients with BP (n = 40) compared with healthy age- and sex-matched controls (n = 40). Bold lines of boxplots indicate the median; boxes illustrate lower and upper quartiles; whiskers show the lowest and highest data point still within 1.5× of interquartile range from lower or upper quartile, respectively; each dot represents an individual. Groups were compared by 2-sided Wilcoxon rank-sum test (p = 0.0002).
The patients with BP included in this study exhibited a wide range of disease activity as assessed by the BPDAI score (Table I). Serum levels of galectin-9 did not correlate with the BPDAI score (Rho = 0.045, p = 0.79, Fig. 2a). There was also no correlation of galectin-9 with the levels of anti-BP180 IgG (Rho = 0.27, p = 0.11, Fig. 2b) and anti-BP230 IgG (Rho =–0.039, p = 0.9, Fig. 2c) detectable.
Fig. 2.

Galectin-9 levels are not associated with disease activity and autoantibody levels. Galectin-9 levels do not correlate with (a) clinical disease activity, as measured with BPDAI score, (b) BP180-NC16A serum levels, or (c) BP230 serum levels at time of diagnosis. Line represents regression line, grey shading illustrates 95% confidence interval (95% CI), Spearman’s correlation test. Number in each plot based on patients with: (a) reported BPDAI score (n = 40 patients with bullous pemphigoid (BP)), (b) with positive anti-BP180-NC16A (n = 37 patients with BP), or (c) anti-BP230 antibody levels (n = 13 patients with BP).
This study also examined whether galectin-9 levels relate to the counts of different blood cell populations. While there was no significant correlation with eosinophils counts, which are typically increased in approximately 50% of patients with BP, there was a significant correlation between monocyte counts and galectin-9 levels (Rho = 0.36, p = 0.031, Fig. 3, Fig. S21).
Fig. 3.

Galectin-9 correlates weakly with the number of circulating monocytes. (a) Galectin-9 levels do not correlate with the number of circulating eosinophils. (b) Galectin-9 is weakly associated with the number of monocytes in the blood (rho = 0.36, p = 0.031). Line represents regression line, grey shading illustrates 95% confidence interval, Spearman’s correlation test; n = 40 patients with bullous pemphigoid.
The study subsequently assessed the expression of galectin-9 at the protein level in perilesional skin of patients with BP by fluorescence-labelled immunohistochemistry. Galectin-9 was pronouncedly more abundant in lesional as well as perilesional skin of patients with BP than in corresponding skin sections of age- and sex-matched healthy donors (Fig. 4). Keratinocytes of patients with BP, especially those of the basal and lower layers, seemed to express galectin-9 in their cytosol, while keratinocytes of healthy controls had mostly intranuclear galectin-9 expression. As galectin-9 was expressed in keratinocytes as well as in cells of the inflammatory dermal infiltrate, the skin sections were also stained for EPX and MPO as markers for eosinophils and neutrophils, respectively. As expected, both eosinophils and neutrophils were abundant in lesional skin (Fig. 5). Galectin-9 was expressed in the cytosol of more than 90% of eosinophils (Fig. 5a) as well as more than 90% of neutrophils neutrophils (Fig. 5b).
Fig. 4.

Keratinocytes in both lesional and perilesional skin biopsies of patients with bullous pemphigoid (BP) express more galectin-9 than healthy controls. Representative immunofluorescence staining pictures of: (a) BP lesional skin (n = 8), (b) BP perilesional skin (n = 8) and (c) healthy control skin (n = 5). Arrows highlight examples of co-expression of galectin-9 and myeloperoxidase (MPO) in neutrophils; dashed lines indicate subepidermal split; scale bars: 100 µm.
Fig. 5.

Eosinophils and neutrophils in lesional bullous pemphigoid (BP) skin express galectin-9 in their cytosol. Histopathological analysis of galectin-9 expression in perilesional skin biopsies of patients with BP. Representative immunofluorescence (IF) staining pictures highlighting the expression of cell marker proteins (a) eosinophil peroxidase (EPX, eosinophils, n = 5) and (b) myeloperoxidase (MPO, neutrophils, n = 8) and their co-expression with galectin-9. Dashed lines indicate subepidermal split; scale bars: 100 µm.
DISCUSSION
This study found that galectin-9 serum levels are significantly elevated in patients with BP, and galectin-9 is expressed in lesional and perilesional skin of these patients. Collectively, these findings hint at a possible regulatory role of galectin-9 in the pathogenesis of BP.
Serum and tissue levels of galectin-9 have been reported previously to be increased in diverse chronic inflammatory conditions, including autoimmune as well as eosinophil- and TH2 cell-driven diseases. Thus, serum galectin-9 has been proposed as an easily accessible biomarker, for instance for systemic sclerosis, atopic dermatitis, systemic lupus erythematosus, and autoimmune hepatitis (15, 18, 20–22). However, the functional significance of galectin-9 has been addressed in only a few models of these diseases. These functional studies have delivered dichotomous results, arguing, depending on the model and experimental setup, either for a pro-inflammatory or protective net effect of galectin-9 on disease (13, 17, 19, 23–25).
For example, BALB/c mice sensitized and challenged with ovalbumin in a preclinical model of allergic lung inflammation presented with higher numbers of galectin-9 expressing eosinophils, elevated galectin-9 protein levels and increased TH2 cytokines in the bronchoalveolar lavage compared with control mice challenged with phosphate-buffered serum (PBS) (17). In in vitro models of rheumatoid arthritis (RA) galectin-9-treated granulocytes were shown to upregulate peptidyl arginine deiminase 4 (PAD-4), a key enzyme required for citrullination of intracellular granulocyte proteins that are known autoantigens to RA pathogenesis, i.e. MPO, alpha-enolase, MMP-9 (19). On the contrary, in a mouse collagen-induced arthritis model galectin-9 suppressed arthritis in a dose-dependent manner by reducing levels of pro-inflammatory cytokines (e.g. IL-17, IL-12, IFNγ), suppressing the generation of TH17 and promoting the induction of regulatory T cells (13). In spontaneous murine lupus models, i.e. in BXSB/MpJ and NZB/W F1 mice, galectin-9 interacted via CD44 with pDCs and B cells and abrogated their ability to mount cytokine responses to TLR7/TLR9 ligands.
Consequently, galectin-9 prevented TLR-mediated IFN secretion by pDCs and autoantibody generation by B cells via the mTOR/p70S6K pathway (23).
BP is, in several respects, peculiar among the family of autoantibody-driven diseases. Its specific characteristics include a vast dominance of eosinophils and TH2 cytokines in the blood, and particularly in the inflamed tissues of patients with BP (3, 5). Both eosinophils and TH2 cytokines, specifically IL-4 and IL-5, are therefore considered important drivers of pathogenesis in BP (3). Therapeutic strategies directly targeting the recruitment and activation of eosinophils in the skin have been or are currently in clinical trials. These therapeutic strategies include the inhibition of IL-5 and CLL11/eotaxin-1 by neutralizing antibodies, as well as the dual inhibition of the lipid mediator leukotriene B4 and the cleavage of the anaphylatoxin C5a from the complement factor C5 by the biotherapeutic nomacopan (26–28). Like most immune cells, eosinophils simultaneously express several chemoattractant receptors and are therefore, in principle, responsive to many different chemoattractants (29, 30). Consequently, when several of these chemoattractants are expressed in the inflamed tissue in parallel, it must be considered that recruitment and activation is organized in a redundant manner (31, 32). As a chemoattractant for eosinophils, galectin-9 might contribute to the redundancy of eosinophil chemoattractant pathways in BP. The presence of galectin-9 in the skin of patients with BP should therefore be considered in the development of respective therapeutic strategies, as only inhibiting one select pathway of eosinophil attraction might be insufficient to ameliorate skin inflammation. The correlation of galectin-9 serum levels with monocytes blood counts in patients with BP indicates a potential interaction of these factors during the pathogenesis of BP. Indeed, monocytes differentiate to M2 macrophages through stimulation with galectin-9 (33). M2 macrophages express TH2 cytokines, such as IL-10, IL-4, TNF and IL-6, which are also found to be critical for pathogenesis of BP (1, 2, 34, 35). Furthermore, monocytes are among the first immune cells that infiltrate the skin after autoantibody binding to the dermal–epidermal junction and, after differentiation to macrophages, play an important role by supporting granulocyte recruitment and blister formation in BP (36). However, the spatiotemporal details of this presumed interplay between galectin-9 and monocytes remain to be clarified.
The current study is limited by its descriptive nature of an increased galectin-9 expression in the serum and lesional skin of patients with BP. Future functional studies in appropriate preclinical models are required to further elucidate the significance of galectin-9 in BP.
The increased levels of galectin-9 in the serum of patients with BP may also be considered in the future, in the development of composite biomarker panels to diagnose BP based on serum samples. Determination of galectin-9 in the serum of patients with BP may contribute to the proficiency of such panels.
In conclusion, these results highlight galectin-9 as another mediator possibly involved in the pathogenesis of BP. Through its eosinophil- and TH2 cytokine-promoting effects galectin-9 may facilitate the eruption of skin lesions in patients with BP. Further research is planned, focussing on the pathological significance of galectin-9 in BP, with functional studies in preclinical models.
The authors have no conflicts of interest to declare.
REFERENCES
- 1.Sadik CD, Schmidt E, Zillikens D, Hashimoto T. Recent progresses and perspectives in autoimmune bullous diseases. J Allergy Clin Immunol 2020; 145: 1145–1147. [DOI] [PubMed] [Google Scholar]
- 2.Sadik CD, Schmidt E. Resolution in bullous pemphigoid. Semin Immunopathol 2019; 41: 645–654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Amber KT, Valdebran M, Kridin K, Grando SA. The role of eosinophils in bullous pemphigoid: a developing model of eosinophil pathogenicity in mucocutaneous disease. Front Med (Lausanne) 2018; 5: 201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Giusti D, Gatouillat G, Le Jan S, Plee J, Bernard P, Antonicelli F, et al. Eosinophil cationic protein (ECP), a predictive marker of bullous pemphigoid severity and outcome. Sci Rep 2017; 7: 4833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Feliciani C, Toto P, Mohammad Pour S, Coscione G, Amerio P, Amerio P. A Th2-like cytokine response is involved in bullous pemphigoid. the role of IL-4 and IL-5 in the pathogenesis of the disease. Int J Immunopathol Pharmacol 1999; 12: 55–61. [PubMed] [Google Scholar]
- 6.Gounni Abdelilah S, Wellemans V, Agouli M, Guenounou M, Hamid Q, Beck LA, et al. Increased expression of Th2 associated chemokines in bullous pemphigoid disease. Role of eosinophils in the production and release of these chemokines. Clin Immunol 2006; 120: 220–231. [DOI] [PubMed] [Google Scholar]
- 7.Chakievska L, Holtsche MM, Kunstner A, Goletz S, Petersen BS, Thaci D, et al. IL-17A is functionally relevant and a potential therapeutic target in bullous pemphigoid. J Autoimmun 2019; 96: 104–112. [DOI] [PubMed] [Google Scholar]
- 8.Norling LV, Perretti M, Cooper D. Endogenous galectins and the control of the host inflammatory response. J Endocrinol 2009; 201: 169–184. [DOI] [PubMed] [Google Scholar]
- 9.Niki T, Tsutsui S, Hirose S, Aradono S, Sugimoto Y, Takeshita K, et al. Galectin-9 is a high affinity IgE-binding lectin with anti-allergic effect by blocking IgE-antigen complex formation. J Biol Chem 2009; 284: 32344–32352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Matsumoto R, Hirashima M, Kita H, Gleich GJ. Biological activities of ecalectin: a novel eosinophil-activating factor. J Immunol 2002; 168: 1961–1967. [DOI] [PubMed] [Google Scholar]
- 11.Matsumoto R, Matsumoto H, Seki M, Hata M, Asano Y, Kanegasaki S, et al. Human ecalectin, a variant of human galectin-9, is a novel eosinophil chemoattractant produced by T lymphocytes. J Biol Chem 1998; 273: 16976–16984. [DOI] [PubMed] [Google Scholar]
- 12.Saita N, Goto E, Yamamoto T, Cho I, Tsumori K, Kohrogi H, et al. Association of galectin-9 with eosinophil apoptosis. Int Arch Allergy Immunol 2002; 128: 42–50. [DOI] [PubMed] [Google Scholar]
- 13.Seki M, Oomizu S, Sakata KM, Sakata A, Arikawa T, Watanabe K, et al. Galectin-9 suppresses the generation of Th17, promotes the induction of regulatory T cells, and regulates experimental autoimmune arthritis. Clin Immunol 2008; 127: 78–88. [DOI] [PubMed] [Google Scholar]
- 14.Fernandez-Santamaria R, Palomares F, Salas M, Dona I, Bogas G, Ariza A, et al. Expression of the Tim3-galectin-9 axis is altered in drug-induced maculopapular exanthema. Allergy 2019; 74: 1769–1779. [DOI] [PubMed] [Google Scholar]
- 15.Chihara M, Kurita M, Yoshihara Y, Asahina A, Yanaba K. Clinical significance of serum galectin-9 and soluble CD155 levels in patients with systemic sclerosis. J Immunol Res 2018; 2018: 9473243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Katoh S, Nobumoto A, Matsumoto N, Matsumoto K, Ehara N, Niki T, et al. Involvement of galectin-9 in lung eosinophilia in patients with eosinophilic pneumonia. Int Arch Allergy Immunol 2010; 153: 294–302. [DOI] [PubMed] [Google Scholar]
- 17.Sziksz E, Kozma GT, Pallinger E, Komlosi ZI, Adori C, Kovacs L, et al. Galectin-9 in allergic airway inflammation and hyper-responsiveness in mice. Int Arch Allergy Immunol 2010; 151: 308–317. [DOI] [PubMed] [Google Scholar]
- 18.Nakajima R, Miyagaki T, Oka T, Nakao M, Kawaguchi M, Suga H, et al. Elevated serum galectin-9 levels in patients with atopic dermatitis. J Dermatol 2015; 42: 723–726. [DOI] [PubMed] [Google Scholar]
- 19.Wiersma VR, Clarke A, Pouwels SD, Perry E, Abdullah TM, Kelly C, et al. Galectin-9 is a possible promoter of immunopathology in rheumatoid arthritis by activation of peptidyl arginine deiminase 4 (PAD-4) in granulocytes. Int J Mol Sci 2019; 20: 4046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wienke J, Bellutti Enders F, Lim J, Mertens JS, van den Hoogen LL, Wijngaarde CA, et al. Galectin-9 and CXCL10 as biomarkers for disease activity in juvenile dermatomyositis: a longitudinal cohort study and multicohort validation. Arthritis Rheumatol 2019; 71: 1377–1390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.van den Hoogen LL, van Roon JAG, Mertens JS, Wienke J, Lopes AP, de Jager W, et al. Galectin-9 is an easy to measure biomarker for the interferon signature in systemic lupus erythematosus and antiphospholipid syndrome. Ann Rheum Dis 2018; 77: 1810–1814. [DOI] [PubMed] [Google Scholar]
- 22.Matsuoka N, Kozuru H, Koga T, Abiru S, Yamasaki K, Komori A, et al. Galectin-9 in autoimmune hepatitis: Correlation between serum levels of galectin-9 and M2BPGi in patients with autoimmune hepatitis. Medicine 2019; 98: e16924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Panda SK, Facchinetti V, Voynova E, Hanabuchi S, Karnell JL, Hanna RN, et al. Galectin-9 inhibits TLR7-mediated autoimmunity in murine lupus models. J Clin Invest 2018; 128: 1873–1887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Arikawa T, Watanabe K, Seki M, Matsukawa A, Oomizu S, Sakata KM, et al. Galectin-9 ameliorates immune complex-induced arthritis by regulating Fc gamma R expression on macrophages. Clin Immunol 2009; 133: 382–392. [DOI] [PubMed] [Google Scholar]
- 25.Zeggar S, Watanabe KS, Teshigawara S, Hiramatsu S, Katsuyama T, Katsuyama E, et al. Role of Lgals9 deficiency in attenuating nephritis and arthritis in BALB/c mice in a pristane-induced lupus model. Arthritis Rheumatol 2018; 70: 1089–1101. [DOI] [PubMed] [Google Scholar]
- 26.Sezin T, Murthy S, Attah C, Seutter M, Holtsche MM, Hammers CM, et al. Dual inhibition of complement factor 5 and leukotriene B4 synergistically suppresses murine pemphigoid disease. JCI Insight 2019; 4: e128239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Simon D, Yousefi S, Cazzaniga S, Burgler C, Radonjic S, Houriet C, et al. Mepolizumab failed to affect bullous pemphigoid: a randomized, placebo-controlled, double-blind phase 2 pilot study. Allergy 2020; 75: 669–672. [DOI] [PubMed] [Google Scholar]
- 28.Lee J, Werth VP, Hall RP, 3rd, Eming R, Fairley JA, Fajgenbaum DC, et al. Perspective from the 5th International Pemphigus and Pemphigoid Foundation Scientific Conference. Front Med (Lausanne) 2018; 5: 306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Diny NL, Rose NR, Cihakova D. Eosinophils in autoimmune diseases. Front Immunol 2017; 8: 484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Rosenberg HF, Dyer KD, Foster PS. Eosinophils: changing perspectives in health and disease. Nature Rev Immunol 2013; 13: 9–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sadik CD, Luster AD. Lipid-cytokine-chemokine cascades orchestrate leukocyte recruitment in inflammation. J Leukoc Biol 2012; 91: 207–215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sadik CD, Kim ND, Luster AD. Neutrophils cascading their way to inflammation. Trends Immunol 2011; 32: 452–460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Enninga EA, Nevala WK, Holtan SG, Leontovich AA, Markovic SN. Galectin-9 modulates immunity by promoting Th2/M2 differentiation and impacts survival in patients with metastatic melanoma. Melanoma Res 2016; 26: 429–441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Hirose M, Kasprick A, Beltsiou F, Dieckhoff Schulze K, Schulze FS, Samavedam UK, et al. Reduced skin blistering in experimental epidermolysis bullosa acquisita after anti-TNF treatment. Molec Med 2017; 22: 918–926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Schmidt E, Bastian B, Dummer R, Tony HP, Brocker EB, Zillikens D. Detection of elevated levels of IL-4, IL-6, and IL-10 in blister fluid of bullous pemphigoid. Arch Dermatol Res 1996; 288: 353–357. [DOI] [PubMed] [Google Scholar]
- 36.Chen R, Fairley JA, Zhao ML, Giudice GJ, Zillikens D, Diaz LA, et al. Macrophages, but not T and B lymphocytes, are critical for subepidermal blister formation in experimental bullous pemphigoid: macrophage-mediated neutrophil infiltration depends on mast cell activation. J Immunol 2002; 169: 3987–3992. [DOI] [PubMed] [Google Scholar]