

Abstract
SARS-CoV2 responsible for the severe acute respiratory syndrome known as COVID-19 has rapidly spread in almost every country and devastated the global economy and health care system. Lung injury is an early disease manifestation believed to be a major contributor to short and long-term pathological consequences of COVID-19, thus drug discovery aiming to ameliorate lung injury could be a potential strategy to treat COVID-19 patients. By inducing SARS-like pulmonary disease model through infecting A/J mice with murine hepatitis virus strain 1 (MHV-1), we show that intravenously administration of pazopanib ameliorates acute lung injuries without affecting the MHV-1 replication. Pazopanib reduces cell apoptosis in MHV-1 infected lungs. Furthermore, we also identified that pazopanib has to be given no later than 48 hours after the virus infection without compromising the therapeutic effect. Our study provides a potential treatment for coronavirus-induced lung injuries and support for further evaluation of pazopanib in COVID-19 patients.
Keywords: Pazoapnib, Coronavirus, COVID-19, Acute lung injury
INTRODUCTION
The current COVID-19 pandemic is caused by a novel coronavirus, designated as severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). The disease causes the death of about 1.2% of cases probably due to massive alveolar damage and progressive respiratory failure [1]. Published pulmonary pathology and radiological reports of COVID-19 and previous SARS-CoV or Middle Eastern respiratory syndrome (MERS) provide a clear association of the disease with acute respiratory distress syndrome (ARDS) [2-5].
ARDS is a severe form of acute lung injury (ALI), which is caused by direct or indirect insults to the lung [6, 7]. In the case of COVID-19, lung injury is likely caused by both direct and indirect means. SARS-CoVs have been shown to cause necrosis of lung epithelial cells (direct insult) [2, 8]. In addition, strong inflammatory responses elicited by the viral infection should also cause lung injury (indirect insult). Besides viral infection, there are many other causes for ALI/ARDS, including bacterial infection, gastric acid aspiration, and trauma. The incidence of ALI/ARDS is reported to be around 200,000 per year in the US (excluding COVID-19) with a mortality rate of around 40% [6]. Currently, there is no pharmacological intervention for the disease. Care of these conditions is largely dependent on supportive measures [9, 10]. This probably contributes to the high fatality rate of COVID-19 and/or many of COVID-19 associated short-term and long-term morbidities as well as mortality.
Pazopanib is a multi-kinase inhibitor that acts as an angiogenesis inhibitor targeting VEGF, PDGF, and c-kit receptors among others. By binding to its receptors, pazopanib crosses the cell membrane to interact with the intracellular domain of the tyrosine kinase receptors and later on to compete with adenosine triphosphate (ATP), where it inhibits downstream signaling pathways involved in tumor angiogenesis, growth, and metastasis [11, 12]. Pazopanib was approved by the FDA in 2012 for use in soft tissue sarcomas (STS). We recently showed that pazopanib was also a high affinity, substrate-specific inhibitor of serine/threonine protein kinases MAP3K2 and 3 for their phosphorylation of Nox2 subunit p47phox at Ser208 [13]. Pazopanib treatment ameliorated lung injuries in both LPS (inflammation)-induced and acid (direct insult)-induced acute lung injury mouse models. Mechanistically, pazopanib acted through moderate increases in reactive oxygen species (ROS) generation from neutrophils, which enhances pulmonary vasculature integrity and promotes lung epithelial cell survival and proliferation, leading to the increased pulmonary barrier function and resistance to ALI [13]. Because there is an abundant presence of lung neutrophils in SARS-CoV-induced ARDS [2-5], a hallmark of ALI/ARDS [14], we postulate that pazopanib may be effective in curbing lung injury associated with the viral infection including COVID-19.
Intranasal infection of A/J mice with the murine coronavirus, MHV-1, produced pulmonary pathological features of SARS. All MHV-1-infected A/J mice developed progressive interstitial edema, neutrophil/macrophage infiltrates, and hyaline membranes [15]. In this study, using MHV-1 infected A/J mice as an animal model, we found that pazopanib, an FDA-approved drug, can reduce lung injuries caused by coronavirus infection, which provides a potential treatment for COVID-19.
MATERIAL AND METHODS
Mice.
Female A/J mice 6 to 8 weeks of age (Jackson Laboratories) were maintained in specific pathogen-free facilities with free access to sterile water. The mice were housed under 12 hr light/dark cycles. All experiments and animal use were performed with the approval of the Institutional Animal Care and Use Committee at Yale University.
Virus.
Virus MHV-1 was originally obtained from the American Type Culture Collection. MHV-1 stocks used in these studies were grown and titered in L2 Percy cells by Dr. Susan Compton at Yale University. Mice were infected with 5,000 plaque-forming units (PFU) intranasally for all experiments.
MHV-1-induced lung injury.
All viral infection studies were performed in a viral isolation room. Mice were anesthetized with ketamine/xylazine (100 and 10 mg/kg). Immediately, mice received an intranasal inoculation of 5,000 PFU MHV-1 in 20 μl ice-cold Dulbecco's modified Eagle's medium. The virus (5,000 PFU) was instilled into the nares, and mice were observed until the virus was inhaled. Mice were treated with pazopanib (3mg/kg) or vehicle intravenously at the indicated time during MHV-1 infection. To measure pulmonary permeability, mice were injected with 100 μl of fluorescein isothiocyanate–labeled albumin (10 mg/ml) via retro-orbital vein two hours before sacrificing. Immediately after sacrifice, 1 ml of PBS was instilled into the lungs and retrieved via a tracheal catheter to obtain BAL. The FITC-albumin in the recovered fluid was measured by using a fluorescence plate reader.
Virus load determination.
Lung tissues with 1ml of Trizol snap-frozen at −80°C were thawed, and total RNAs were isolated from cells with Direct-zol RNA Kit (Zymo Research), as per the manufacturer's instructions. Complementary DNAs were synthesized from the RNAs with the iScript cDNA synthesis kit (Bio-Rad), according to the manufacturer's protocol. Quantitative PCR was done with iTaq Universal SYBR Green Supermix (Bio-Rad) [16]. The primer sequences for virus load detection are 5’-GTCATGAGGCTATTCCTACTA -3’ and 5’-ATACACATCTTTGGTGGG -3’.
Preparation of BAL leukocytes and lung-infiltrating leukocytes.
To prepare leukocytes in the BAL, 1 ml of PBS was instilled into the lungs and retrieved via a tracheal catheter to obtain BAL. Leukocytes in the BAL fluid were pelleted by centrifugation (500g for 5 min at 4°C). To prepare lung-infiltrating leukocytes, lungs were minced with scissors and incubated with digestion buffer (RPMI 1640 medium, 5% FBS, 1% penicillin-streptomycin, 25 mM HEPES, and 300 U collagenase (Sigma C0130) in a shaker for 1 hour at 37 °C. Dispersed cells were filtered through a 70-μm cell strainer to eliminate clumps and debris. After centrifugation for 5 min (500g) at 4 °C, cell pellets were resuspended in red blood cell lysis buffer (Sigma R7757) and incubated at room temperature for 5 min to remove erythrocytes. Cells were pelleted again and resuspended in PBS.
Histology.
H&E staining of unlavaged lung tissues was performed at the Comparative Pathology Research core at Yale School of Medicine. Briefly, mouse right ventricles were perfused with 10 ml PBS and the lungs were inflated with 4% paraformaldehyde at a constant pressure of 25 cm H2O, and then fixed in 4% paraformaldehyde for 24 hours at 4°C. The tissues were embedded in paraffin, and cut into 5-μm thick sections. Sections were stained with hematoxylin and eosin, and images were taken with a Keyence BZ-X800 Cell Imaging Microscope. ALI indices were quantified using H&E stained lung sections as previously described [17]. Five parameters (A. Neutrophils in the alveolar space, B. Neutrophils in the interstitial space, C. Hyaline membranes, D. Proteinaceous debris filling the airspaces, E. Alveolar septal thickening) were evaluated. The lung injury score was calculated as score = [(20 X A) + (14 X B) + (7 X C) + (7 X D) + (2 X E)] / (number of fields X 100). Each datum is the average score of the five criteria from five fields, which were uniformly and randomly picked from one lung section of one mouse. Five mice were included in each group. Images were evaluated by two investigators who were blinded to the identity of the sections.
Flow cytometry.
Flow cytometry was performed as previously described [18]. Cells in single-cell suspension were fixed with 2% PFA (Santa Cruz, sc-281692). After being washed with a flow cytometry staining buffer (eBioscience, 00-4222-26), cells were stained with antibodies for cell-surface markers for 1 hour on ice in the dark. The cells were then washed, pelleted and resuspended in the flow cytometry staining buffer for flow cytometry analysis. The absolute number of cells was counted by using CountBright™ Absolute Counting Beads (Invitrogen C36950), according to the manufacturer’s instruction. Antibodies for flow cytometry are as follows: mouse CD45-BUV395 (BD), mouse CD4-FITC (Biolegend), mouse CD8-Pacific blue (Biolegend), mouse CD11b-Pacific blue (Biolegend), mouse CD11b-APC (Biolegend), mouse Ly6G-Pacific blue (Biolegend), mouse Ly6C-FITC (Biolegend), mouse NK1.1-APC (Biolegend), mouse B220-PE (Biolegend), mouse F4/80-APC (Biolegend). Major immune cell populations were determined by CD45+CD11b+Ly6G+(Neutrophils), CD45+CD11b+Ly6G−Ly6C+(Monocytes), CD45+CD4−CD8−B220−NK1.1+(NK cells), CD45+CD4+(CD4 T cells), CD45+CD8+(CD8 T cells), CD45+CD11b+ F4/80+(Macrophages).
Immunohistochemistry.
Unlavaged lungs were inflated and then fixed with 4% PFA for 4–6 h on a shaker at 4 °C. They were then washed with PBS three times and perfused in 30% sucrose solution in PBS overnight at 4 °C. They were subsequently mounted in OCT embedding compound and frozen first at −20 °C and then at −80 °C. Tissue sections were prepared at 8-μm thickness with a cryostat and mounted onto gelatin-coated histological slides, which were stored at −80 °C. For immunostaining, slides were thawed to room temperature and fixed in pre-cold acetone for 10 min, then rehydrated in PBS for 10 min. The slides were incubated in a blocking buffer (1% horse serum and 0.02% Tween 20 in PBS) for 1 hour at room temperature, then incubated with anti-ABCA3 (ab24751, Abcam), anti-Cleaved caspase-3 antibodies (9661, Cell signaling technology), anti-PDPN (AF3244, R&D Systems) and anti-CD31 (AF3628, R&D Systems) which were diluted in the blocking buffer, overnight at 4°C. The slides were then washed three times with PBS and incubated with a secondary antibody in the incubation buffer for 1 hour at room temperature. After repeated washes, the slides were mounted with an anti-fade mounting media containing DAPI (Thermo Fisher, P36931) and visualized with a Keyence BZ-X800 Cell Imaging Microscope [19]. Apoptotic cells were quantified by using imageJ software. Lung apoptotic cells were assessed by normalizing the area of active caspase-3 positive signals to the area of DAPI signals. Lung apoptotic type I epithelial cells, type II epithelial cells and endothelial cells were assessed by normalizing the area of active caspase-3 and cell marker double positive signals to the area of cell marker positive signals. Each datum represents one field from one section of one mouse. Five fields were evaluated for each section and three mice were included in each group.
Serum pazopanib measurement.
Mice were anesthetized with ketamine/xylazine (100 and 10 mg/kg) for blood collection by retro-orbital bleeding. Serum was collected by centrifugation of the blood in BD Microtainer blood collection tubes (Catalog no.365985) at 10000g for one minute. Serum was then transferred to the Eppendorf tube for LC-MS measurement.
Statistical analysis.
Comparisons of means between two groups and multiple groups were tested by unpaired, two-tailed t test and One-way ANOVA-test using Prism 9.2.0 software (GraphPad). For Kaplan-Meier survival analysis, log-rank test was used. Statistical tests used biological replicates. *p < 0.05, **p < 0.01 and ***p < 0.001 were all considered statistically significant.
RESULTS
MHV-1 induces acute lung injury in A/J mice
To recapitulate pulmonary pathological manifestations of COVID-19 in mice, we infected A/J mice intranasally with 5,000 PFU of murine hepatitis virus strain 1 (MHV-1). Forty-eight hours after MHV-1 infection, we examined the lung vascular permeability, which indicated the severity of acute lung injury. Bronchoalveolar lavage (BAL) was collected from MHV-1 infected A/J mice two hours after injection of FITC-albumin intravenously. MHV-1 inoculation increased FITC-albumin extravasation into the BAL when compared with the control treatment, indicating that MHV-1 infection increased pulmonary permeability, a hall marker of ALI, in A/J mice (Fig. 1A). Neutrophils also infiltrated into BAL after MHV-1 infection, another hall marker of acute lung injury (Fig. 1B, S1A). Additionally, H&E (Haemotoxylin and Eosin) histological staining revealed that MHV-1 infection caused interstitial alveolar thickening, fluid accumulation in alveolar spaces (pulmonary edema) and heavy leukocyte infiltrates (Fig. 1C, 1D). Taken together, intranasal inoculation of MHV-1 induced lung injury 48 hours post challenge in A/J mice, with characteristics very similar to those observed in human COVID-19, including interstitial pulmonary infiltrates, congestion, and pulmonary edema.
Figure 1. Coronavirus MHV-1 induced lung injury in A/J mice.

(A-B) A/J mice (8 weeks old) were intranasally infected with 5,000 PFU of MHV-1. BAL was collected 48 hours after MHV-1 infection, followed by the measurements of pulmonary permeability (A) and the total number of neutrophils (B). Data were combined from two independent experiments with four or two mice per group per experiment, each datum represents one mouse. (C) Representative histology of injured lungs 48 hours after MHV-1 infection. (D) Quantification of lung injuries. Each datum is the average score of five fields, which were uniformly and randomly picked from one lung section of one mouse. Five mice were included in each group. The samples were collected from two independent experiments with three or two mice per group per experiment. The samples were then combined for the following processing, staining and evaluation. Data in (A), (B) and (D) are presented as means±SEM. ns, not significant, *p < 0.05, **p < 0.01 and ***p < 0.001, Student’s t test;
Pazopanib suppresses MHV-1-induced acute lung injury in A/J mice
Our previous study showed that pazopanib ameliorated acute lung injury by protecting pulmonary barrier functions in LPS and acid-induced mouse ALI models [13]. To test if pazopanib can protect MHV-1 induced lung injury, A/J mice were treated with pazopanib during MHV-1 infection. A pharmacokinetic study revealed a short half-life (5.9 hours) of pazopanib in the blood, suggesting that pazopanib was cleared quickly after intravenously delivery to mice (Fig. S1B). Based on this characteristic feature, pazopanib may need to be administrated to mice multiple times with a short interval. First, we tried a three-dose regimen of pazopanib treatment. The first dose was injected intravenously at 6 hours after MHV-1 inoculation. The second and third doses were delivered at 15 and 26 hours after the first dose. In this scenario, pazopanib significantly reduced pulmonary permeability versus vehicle control (Fig. 2A, 2B). Next, we examined whether two doses of pazopanib can also ameliorate lung injury. In this scenario, two doses of pazopanib were given at 24 and 33 hours after MHV-1 inoculation, and a significant reduction of pulmonary permeability was observed as well (Fig. 2C, 2D). Next, we proceeded to evaluate a one-dose regimen (24 hours after virus inoculation) of pazopanib treatment. Although the one-dose regimen had a lower concentration of pazopanib in the blood (Fig. S1C), it still showed a significant reduction in lung permeability (Fig. 2E, 2F). Histological assessment of the lung further demonstrated that treatment with pazopanib reduced MHV-1 induced lung pathology (Fig. 2G, 2H, S1D, S1E). Unlike the role of pazopanib in suppressing acute lung injury, liver and kidney injury induced by MHV-1 were not ameliorated by pazopanib treatment (Fig. S1F). To examine whether the effect of pazopanib on suppressing acute lung injury was due to alteration of virus replication, we determined the viral loads in the lungs from the MHV-1-infected mice with or without pazopanib treatment. Pazopanib did not affect viral loads regardless of the treatment modalities (Fig. S1G). Major immune cell populations in the BAL and lung were also examined after pazopanib treatment. Pazopanib didn’t affect neutrophil infiltration into BAL and lung (Fig. S2A, S2B). The infiltration of CD4 T cells, CD8 T cells or macrophages in the lung was not affected by pazopanib either (Fig. S2B, S2C). However, pazopanib treatment appeared to increase NK cell presence and CD8 expression in the lungs (Fig. S2D & S2E). These results together indicated that intravenously delivery of pazopanib suppressed MHV-1-induced acute lung injury in mice without affecting MHV-1 replication in the lungs.
Figure 2. Pazopanib treatment can reduce MHV-1 induced lung injury.

(A, C, E) Schematic diagram of the different regimens of pazopanib treatment. (B, D, F) BAL was collected 48 hours after MHV-1 infection and pulmonary permeability was examined. Data in (B) were combined from two independent experiments. The vehicle group included four and two mice per experiment. Pazo group included five or one mouse per experiment, each datum represents one mouse. Data in (D) were combined from two independent experiments with four or two mice per group per experiment, each datum represents one mouse. Data in (F) were combined from three independent experiments, in each independent experiment, each group contained two or three mice, and each datum represents one mouse. (G-H). A/J mice (8 weeks old) were intranasally infected with medium or 5,000 PFU of MHV-1. Pazopanib or vehicle was delivered 24 hours after MHV-1 inoculation. 48 hours after infection, mice were sacrificed and lungs were collected for histology analysis. Representative lung histology images were shown in (G). Quantification of lung injuries in (H). Each datum is the average score of five fields, which were uniformly and randomly picked from one lung section of one mouse. Five mice were included in each group. The samples were collected from two independent experiments with three or two mice per group per experiment. The samples were then combined for the following processing, staining and evaluation. Data in (B), (D) and (F) are presented as means±SEM, Student’s t test and data in (H) are presented as means±SEM, one-way ANOVA. ns, not significant, *p < 0.05, **p < 0.01 and ***p < 0.001;
The timing of pazopanib treatment is important.
The one-dose regimen of pazopanib treatment at 24 hours after virus inoculation provided significant protection from coronavirus-induced acute lung injury. In the real world, COVID-19 patients may not know they have been infected with the virus until the onset of the symptoms. To find out what is the best time for starting the pazopanib treatment, two more single-dose regimens, 48 hours or 72 hours after virus inoculation, were tested in the mouse model. Significant improvement was observed in the single-dose regimen at 48 hours post infection (Fig. 3A, 3B), but not in the regimen at 72 hours post infection (Fig. 3C, 3D). Since these mice had developed the severe progressive pulmonary disease at 48hr, the findings suggest that pazopanib treatment may be more effective if administrated early in the disease.
Figure 3. Pazopanib has to be given no later than 48 hours after MHV-1 infection.

(A, C) Schematic diagram of the different regimens of pazopanib treatment. (B, D) BAL was collected at the indicated time points after MHV-1 infection and pulmonary permeability was examined. Data in (B) were combined from two independent experiments, in the first independent experiment, each group (from left to right) contained two, four, and three mice, in the second independent experiment, each group (from left to right) contained two, two, and three mice, each datum represents one mouse. Data in (D) were combined from two independent experiments, in the first independent experiment, each group (from left to right) contained two, four, and three mice, in the second independent experiment, each group (from left to right) contained two mice, each datum represents one mouse. Data in (B) and (D) are presented as means±SEM, one-way ANOVA. ns, not significant, *p < 0.05, **p < 0.01 and ***p < 0.001;
Pazopanib promotes lung cell survival during MHV-1 infection
Our previous study showed that pazopanib could protect lung epithelial cells from apoptosis [13]. To determine whether pazopanib also has the same effect during MHV-1 infection, we examined the active caspase-3 expression in the lung tissues at 48 hours after MHV-1 inoculation. MHV-1 infection markedly induced the expression of active caspase-3 in the lungs, which was significantly reduced by pazopanib treatment (Fig. 4A, 4B). Next, we co-stained active caspase-3 with PDPN (type I epithelial cell marker), ABCA3 (type II epithelial cell marker) and CD31 (endothelial cell marker) respectively. Active caspase-3 was found in all three cell types, but was mainly co-localized with ABCA3. Pazopanib treatment reduced active caspase-3 in both epithelial cells and endothelial cells (Fig. 4C-H). These results suggested that pazopanib could inhibit lung cells apoptosis and prolong their survival.
Figure 4. Pazopanib treatment reduced lung cells apoptosis.

A/J mice (8 weeks old) were intranasally infected with medium or 5,000 PFU of MHV-1. Pazopanib or vehicle was delivered 24 hours after MHV-1 inoculation. 48 hours after infection, mice were sacrificed and lung sections were stained with anti-active caspase-3 (Casp-3) and DAPI (A-B), anti-active caspase-3 (Casp-3), ABCA3 and DAPI (C-D), anti-active caspase-3 (Casp-3), PDPN and DAPI (E-F), anti-active caspase-3 (Casp-3), CD31 and DAPI (G-H) as indicated in the panels. Representative images are shown in (A, C, E, G). Quantifications are shown in (B, D, F, H). Each datum represents one field from one section of one mouse. Five fields were evaluated for each section and three mice were included in each group. The samples were collected from two independent experiments with two or one mouse per group per experiment. The samples were then combined for the following processing, staining and evaluation. Data in (B) are presented as means±SEM, one-way ANOVA and Data in (D), (F) and (H) are presented as means±SEM, student’s t test. ns, not significant, *p < 0.05, **p < 0.01 and ***p < 0.001, (B), (D), (F) and (H) n=15.
Pazopanib alone cannot extend the mice survival after MHV-1 inoculation
To test whether pazopanib can reduce mortality induced by MHV-1 infection, A/J mice were infected with MHV-1, and pazopanib was intravenously delivered to mice once daily beginning 24 hours after virus inoculation. The last dose was given at 120 hours after inoculation. Mice started to die on day 5, however, pazopanib treatment didn’t change the survival rate or bodyweight loss after MHV-1 inoculation (Fig. 5A, 5B). These results suggested that pazopanib alone cannot extend the survival of A/J mice after MHV-1 inoculation.
Figure 5. Pazopanib treatment does not affect mice survival in MHV-1 induced lung injury model.
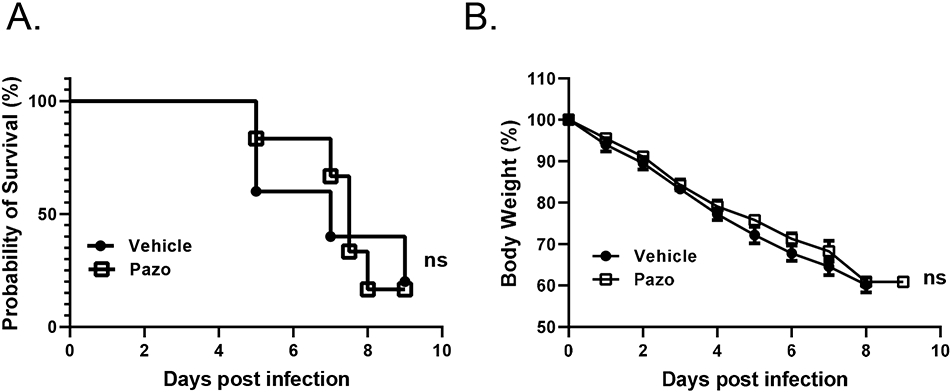
(A-B) The effect of pazopanib on mortality (A) and weight change (B) in mice was examined in MHV-1 induced ALI model. Experiments for (A) and (B) were conducted for two times with similar results (the first independent experiment contained 5 mice in the vehicle control group (Vehicle) and 6 mice in the pazopanib treatment group (Pazo), and the second independent experiment contained 7 mice per group). Data from the first independent experiment were shown in (A) and (B). Data in (A) are presented in Mantel-Cox log-rank test. ns, not significant;
DISCUSSION
In this study, we demonstrated that intravenous administration of pazopanib can significantly reduce lung injury in coronavirus MHV-1 infected A/J mice. Pazopanib is an FDA approved anti-cancer drug, which was identified as being inhibiting multiple tyrosine kinase receptors associated with angiogenesis and tumor cell proliferation. Our previous study showed that pazopanib was a potent inhibitor for MAP3K2/3-mediated phosphorylation of p47phox at Ser208 and acts largely through this MAP3K2/3-p47phox pathway to ameliorate lung injuries and extend survival in mouse LPS and HCl (Hydrogen Chloride) induced ALI models [13]. COVID-19, caused by SARS-CoV-2, has become a pandemic for almost two years. Most patients with COVID-19 exhibit mild to moderate symptoms, but approximately 20% develop ALI/ARDS [20]. Currently, there is no approved drug for treating this SARS-CoV-2 induced ALI/ARDS. Pazopanib has been approved by FDA in the clinic for years and is well tolerated even for long-term use [21]. Thus, our current study demonstrates that pazopanib has a promising potential for the treatment of ALI/ARDS in COVID patients.
The mechanism of action of pazopanib described in our previous study is through a paracrine mechanism. The moderate elevation of extracellular H2O2 released from neutrophils triggers the pulmonary microenvironment remodeling through crosstalk and interactions of different lung cell types, leading to the protection of lungs from acute injuries [13]. Neutrophils are also abundantly recruited to the lungs in SARS-CoV-2 induced ALI/ARDS [4, 5]. In autopsy samples from the lungs of COVID-19 patients, neutrophil infiltration was observed in pulmonary capillaries, and extravasation of neutrophils into the alveolar space was noted as well [22]. So we believe that pazopanib ameliorates coronavirus induced lung injuries through the same mechanism. In our study here, we found that neutrophils were accumulated in the lung of MHV-1 infected mice. Consistent with our previous observations in the other mouse ALI models, pazopanib treatment did not alter neutrophil infiltration into the lungs in this MHV-1 infection model. Our histology results showed that pazopanib treatment substantially inhibits the apoptosis of pulmonary cells. Our previous study has already demonstrated that pazopanib cannot affect pulmonary cells by itself [13], therefore these results suggested that pazopanib treated neutrophils impact pulmonary cells to favor enhancement of barrier functions via a paracrine mechanism in coronavirus induced ALI. As neutrophil presence is only dominant at the early stage of viral infection, it may help to explain why pazopanib is more efficacious at the early stage of the disease.
Pazopanib extends the survival of HCl or LPS induced ALI mice in our previous study, however, the survival is not significantly changed in this coronavirus induced ALI model. That’s probably because HCl or LPS only induces a transient insult to the lung, while coronavirus MHV-1 can proliferate in the lung and cause a continuous and less confined insult. Multiple organs including livers have been observed to be affected by MHV-1 infection [15] and our data indicated that pazopanib failed to reduce the liver and kidney injury induced by MHV-1 (Fig. S1F). Given that pazopanib cannot inhibit virus proliferation (Fig. S1G), the protective effect of pazopanib might be overridden as more and more viruses are amplifying. Future studies may be needed to investigate whether combining pazopanib with other anti-virus drugs could give us better results or even extend survival in coronavirus induced ALI model.
Supplementary Material
Key Points.
Pazopanib ameliorates MHV-1-induced lung injury in A/J mice.
Pazopanib reduces lung cell apoptosis induced by MHV-1 in A/J mice.
Acknowledgments
This work is supported by NIH grants to W.T. (R01HL145152), D.W. (R35HL135805).
Footnotes
COMPETING INTERESTS
D.W. is a co-founder and serves as a scientific advisory board member of Qx Therapeutics, which licensed the intellectual properties described in this study from Yale University. D.W., W.T., and Q.Y. are inventors of the patent application “Compounds, compositions and methods of treating or preventing acute lung injury” (no. 62/486,232).
REFERENCES
- 1.Maitz T, et al. COVID-19 Cardiovascular Connection: A Review of Cardiac Manifestations in COVID-19 Infection and Treatment Modalities. Curr Probl Cardiol, 2022: p. 101186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ding Y, et al. The clinical pathology of severe acute respiratory syndrome (SARS): a report from China. J Pathol, 2003. 200(3): p. 282–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ng DL, et al. Clinicopathologic, Immunohistochemical, and Ultrastructural Findings of a Fatal Case of Middle East Respiratory Syndrome Coronavirus Infection in the United Arab Emirates, April 2014. Am J Pathol, 2016. 186(3): p. 652–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Tian S, et al. Pulmonary Pathology of Early-Phase 2019 Novel Coronavirus (COVID-19) Pneumonia in Two Patients With Lung Cancer. J Thorac Oncol, 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Xu Z, et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. The Lancet Respiratory Medicine, 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Johnson ER and Matthay MA, Acute Lung Injury: Epidemiology, Pathogenesis, and Treatment. Journal of Aerosol Medicine and Pulmonary Drug Delivery, 2010. 23(4): p. 243–252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Maca J, et al. Past and Present ARDS Mortality Rates: A Systematic Review. Respir Care, 2017. 62(1): p. 113–122. [DOI] [PubMed] [Google Scholar]
- 8.Li S, et al. SARS-CoV-2 triggers inflammatory responses and cell death through caspase-8 activation. Signal Transduct Target Ther, 2020. 5(1): p. 235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Brower RG, et al. Ventilation with lower tidal volumes as compared with traditional tidal volumes for acute lung injury and the acute respiratory distress syndrome. New England Journal of Medicine, 2000. 342(18): p. 1301–1308. [DOI] [PubMed] [Google Scholar]
- 10.Weinert CR, Gross CR, and Marinelli WA, Impact of randomized trial results on acute lung injury ventilator therapy in teaching hospitals. American Journal of Respiratory and Critical Care Medicine, 2003. 167(10): p. 1304–1309. [DOI] [PubMed] [Google Scholar]
- 11.Halim NA, et al. Safety and efficacy of pazopanib as a second-line treatment and beyond for soft tissue sarcomas: A real-life tertiary-center experience in the MENA region. Cancer Treat Res Commun, 2020. 26: p. 100275. [DOI] [PubMed] [Google Scholar]
- 12.Sloan B and Scheinfeld NS, Pazopanib, a VEGF receptor tyrosine kinase inhibitor for cancer therapy. Curr Opin Investig Drugs, 2008. 9(12): p. 1324–35. [PubMed] [Google Scholar]
- 13.Yuan Q, et al. Pazopanib ameliorates acute lung injuries via inhibition of MAP3K2 and MAP3K3. Sci Transl Med, 2021. 13(591). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Matthay MA and Zemans RL, The acute respiratory distress syndrome: pathogenesis and treatment. Annu Rev Pathol, 2011. 6: p. 147–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.De Albuquerque N, et al. Murine hepatitis virus strain 1 produces a clinically relevant model of severe acute respiratory syndrome in A/J mice. J Virol, 2006. 80(21): p. 10382–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chen Y, et al. p38 inhibition provides anti-DNA virus immunity by regulation of USP21 phosphorylation and STING activation. J Exp Med, 2017. 214(4): p. 991–1010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Matute-Bello G, et al. An official American Thoracic Society workshop report: features and measurements of experimental acute lung injury in animals. Am J Respir Cell Mol Biol, 2011. 44(5): p. 725–38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sun J, et al. The 11S Proteasome Subunit PSME3 Is a Positive Feedforward Regulator of NF-kappaB and Important for Host Defense against Bacterial Pathogens. Cell Rep, 2016. 14(4): p. 737–749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Qiu B, et al. Loss of FKBP5 Affects Neuron Synaptic Plasticity: An Electrophysiology Insight. Neuroscience, 2019. 402: p. 23–36. [DOI] [PubMed] [Google Scholar]
- 20.Cao X, COVID-19: immunopathology and its implications for therapy. Nat Rev Immunol, 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zivi A, et al. Safety and tolerability of pazopanib in the treatment of renal cell carcinoma. Expert Opin Drug Saf, 2012. 11(5): p. 851–9. [DOI] [PubMed] [Google Scholar]
- 22.Barnes BJ, et al. Targeting potential drivers of COVID-19: Neutrophil extracellular traps. J Exp Med, 2020. 217(6). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.