

Abstract
Youth behavior changes and their relationships to personality have generally been investigated using self-report studies, which are subject to reporting biases and confounding variables. Supplementing these with objective measures, like GPS location data, and twin-based research designs, which help control for confounding genetic and environmental influences, may allow for more rigorous, causally informative research on adolescent behavior patterns. To investigate this possibility, this study aimed to (1) investigate whether behavior changes during the transition from adolescence to emerging adulthood are evident in changing mobility patterns, (2) estimate the influence of adolescent personality on mobility patterns, and (3) estimate genetic and environmental influences on mobility, personality, and the relationship between them. Twins aged Fourteen to twenty-two (N=709, 55% female) provided a baseline personality measure, the Big Five Inventory, and multiple years of smartphone GPS data from June 2016 - December 2019. Mobility, as measured by daily locations visited and distance travelled, was found via mixed effects models to increase during adolescence before declining slightly in emerging adulthood. Mobility was positively associated with Extraversion and Conscientiousness (r of 0.17 – 0.25, r of 0.10 – 0.16) and negatively with Openness (r of −0.11 – −0.13). ACE models found large genetic (A = 0.56 – 0.81) and small-moderate environmental (C of 0.12 – 0.28, E of 0.07 – 0.15) influences on mobility. A and E influences were highly shared across mobility measures (rg = 0.70, re= 0.58). Associations between mobility and personality were partially explained by mutual genetic influences (rg of −0.27 – 0.53). Results show that as autonomy increases during adolescence and emerging adulthood, we see corresponding increases in youth mobility. Furthermore, the heritability of mobility patterns and their relationship to personality demonstrate that mobility patterns are informative, psychologically meaningful behaviors worthy of continued interest in psychology.
Keywords: GPS Location Data, Smartphone, Personality, Heritability, Environmental Influences, adolescent development, emerging adulthood
Introduction
In many cultures, late adolescence is the first period of substantial autonomy during the lifespan. Adolescents spend less time with their parents and more time with their peers and exert far greater control over their daily lives and activities than in childhood (Steinberg & Morris, 2001). In the United States and other western countries, developmental milestones like learning to drive, beginning to work, attending college, and leaving home all take place during late adolescence and further contribute to this expansion of autonomy (Remschmidt, 1994). As adolescents grow increasingly autonomous, adolescent personality plays a greater role in their daily experiences, behavior patterns, and life experiences (Johnson et al., 2013; McAdams et al., 2013). For example, adolescent personality is predictive of engagement in social activities, academic or career aspirations, artistic expression, and interest in recreational drug use (DeYoung et al., 2008; Wrzus et al., 2013). Additionally, life events under some degree of an adolescent’s control, like school suspensions, breaking up with a romantic partner, and starting or losing a job are also significantly associated with adolescent personality (Billig et al., 1996).
As behavior patterns which emerge during adolescence, such as eating habits, exercise, substance use, and sexual decision making, are highly predictive of important health outcomes, understanding how factors like personality contribute to their development carries significant scientific and public health implications (Alberga et al., 2012; Chambers et al., 2003; Sawyer et al., 2018). Understanding how adolescents move through and engage with their environments can help scientists, clinicians, and policy makers understand risk trajectories, identify at risk individuals, and design interventions to reduce the incidence of health problems like obesity or substance use.
Psychologists have historically relied on observational, self-report-based studies to understand developmental changes in adolescent behavior patterns. Self-report surveys are efficient to administer and adaptable to a wide variety of psychological constructs; they have helped us glean important insights into how adolescents’ daily activities change and how they are influenced by factors like personality (Csikszentmihalyi et al., 2014; Wrzus et al., 2013). However, while self-report based observational studies have proven useful, they come with methodological limitations that limit our ability to draw generalizable conclusions. For instance, they do not directly measure behavior, are subject to response biases, are limited by participant self-knowledge, and are often burdensome for participants to complete (Paulus & Vazire, 2007). Additionally, observational research is prone to confounding variables which can produce spurious correlations and render interpretation particularly difficult (Grimes & Schulz, 2002).
The limitations of self-report data can in part be mitigated through additional measures which are less prone to the biases associated with self-report. Smartphone GPS data, for example, can be used to unobtrusively observe and quantify aspects of participants’ daily activities (Harari et al., 2016; Miller, 2012). Smartphone data offer standardized, objective measures of participants’ locations and movement patterns which may be useful in corroborating the findings of existing research on adolescents’ daily lives. Previous research has demonstrated that human mobility patterns can be reliably measured using GPS data (Andrade et al., 2019) and that such patterns are meaningfully related to personality and daily activities in adolescence and young adulthood. Several studies have reported relationships between daily mobility patterns and personality traits in adolescence and young adulthood (Ai et al., 2019; Alessandretti et al., 2018; Stachl et al., 2020). Additionally, mobility based measures have been used to predict adolescent psychological and health outcomes like alcohol use, affect, anxiety and depression symptoms, and sleep patterns in adolescent and college aged samples (Jacobson & Bhattacharya, 2022; Ren et al., 2022; Santani et al., 2018; Sathyanarayana et al., 2016).
However, existing research has been conducted over short time spans in relatively small samples of adolescents, and research observing mobility patterns over the course of adolescence has yet to be conducted. Hence it remains an open question how mobility patterns change during this period of growing autonomy. Such information can help inform claims about how daily life changes during adolescence and help provide further information about whether daily mobility patterns contain useful information about human behavior over longer time spans.
Such research can be further improved by using twin data, which can help us understand where individual differences in adolescents’ daily mobility patterns come from and how they are related to potential explanatory variables like personality. Twin data allows researchers to measure the extent of genetic and environmental contributions to variation in a trait or behavior. Additionally, multivariate behavioral genetic models using twin data can assess whether associations between traits result from mutual genetic or environmental influences. Hence, twin studies can help alleviate the problem of confounding variables in observational research by providing additional understanding of the nature and origins of correlational patterns: helping to parse the extent to which associations between variables are explained by genetic, shared environmental, or non-shared environmental factors (McGue et al., 2010). Twin-based analyses can thereby offer evidence for whether adolescent mobility patterns stem more from heritable traits, such as their preferences for particular activities, or from aspects of their environment, such as how many kilometers away from school they live. Furthermore, measuring the degree of overlapping genetic influences on mobility and personality can offer further insight into why mobility might be heritable, perhaps partly due to the influence of other heritable behavioral traits, like personality.
The present study thus had three primary aims. First, to assess whether changes in autonomy and daily activities which occur during adolescence and emerging adulthood are reflected in adolescent mobility patterns. Second, to investigate how changes in mobility are related to adolescent personality. Third and finally, to estimate how mobility and its relationship to personality are influenced by genetic, shared environmental, and non-shared environmental factors.
Methods
Sample
Study participants were 709 adolescent and young adult twins (242 dizygotic twin pairs, 127 monozygotic twin pairs, 55.4% female; Mage= 16.3 at intake; 19.6 in December, 2019) recruited from birth records provided by the Colorado Department of Health and Human Services. The current study, the Colorado Online Twin Study (CoTwins), is a multi-year study with collection of genetic information, persistent location data, and both in-lab and smartphone-based surveys to understand the causes and risk trajectories for youth substance use and psychopathology. Participants had to have a smartphone to participate. Racial and ethnic diversity was somewhat lower than the adolescent population of the United States, (Puzzanchera, 2020) with Eighty percent of the sample identifying as white and fourteen percent identifying as Hispanic or Latino. See Table 1 for a detailed demographic breakdown.
Table 1.
Sample Demographic Characteristics
| Monozygotic Twins | Dizygotic Twins | Total | |
|---|---|---|---|
| Age | |||
| Age at intake (years) | 16.5 (1.3) | 16.1 (1.2) | 16.3 (1.2) |
| Length of Assessment (years) | 2.2 (1.3) | 2.3 (1.2) | 2.3 (1.2) |
| Sex | |||
| Female | 145 | 248 | 393 |
| Male | 98 | 218 | 316 |
| Race | |||
| American Indian/Alaska Native | 0 | 7 | 7 |
| Asian | 2 | 2 | 4 |
| Black/African American | 2 | 6 | 8 |
| White | 190 | 380 | 570 |
| Native-Hawaiian or other Pacific Islander | 2 | 0 | 2 |
| More than one Race | 22 | 49 | 71 |
| Declined to Answer | 25 | 22 | 47 |
| Hispanic/Latino Status | |||
| Hispanic/Latino | 35 | 64 | 99 |
| Non-Hispanic/Latino | 183 | 380 | 463 |
| Declined to Answer | 25 | 22 | 47 |
| Smartphone OS | |||
| Android | 43 | 78 | 121 |
| iOS | 198 | 387 | 585 |
| Total | 243 | 466 | 709 |
Measures
Participant location.
Location was collected via the CoTwins smartphone application (the “app”). The app was designed to provide persistent smartphone location tracking and to administer weekly and monthly surveys on several psychological outcomes. To reduce battery usage, iOS users’ locations were recorded using the Significant-Change Location API, which recorded a user’s location whenever a location change greater than 500m was detected. The Android version of the CoTwins smartphone app requested a participant’s location once every 5 minutes. Due to the inherently identifying nature of persistent GPS location data, significant protections needed to be put in place to ensure participant privacy. In support of ongoing open science efforts and as part of an agreement with the National Institute of Health, the authors have agreed to make this data publicly available after taking steps to ensure participants’ anonymity.
We restricted analyses to location data collected between June 2016 and December 31, 2019 to reduce confounding effects on mobility from the COVID-19 pandemic. Data collection yielded 17.1 million locations. Data cleaning consisted of removing duplicated points, points where a participant’s location could not be determined within 500m, and incomplete points missing either a latitude or longitude components. After cleaning, the data consisted of 13.2 million unique, reliable locations. Using the algorithm described in Zheng et al. (2010), these locations were further aggregated into points of interest, also referred to as “staypoints,” which were defined as places where participants spent at least 30 minutes within a 200m radius. Travel days, in which the total distance between staypoints exceeded 200km, were excluded from analyses to reduce the effect of outlying values. After aggregation and cleaning, the final dataset consisted of 1.6 million staypoints.
In addition to visually inspecting sensor data from a large minority of participants, several analyses were conducted to assess the reliability and validity of staypoint data. As, prior to age 18, participants were twins living in the same household in the state of Colorado, the proportion of points recorded within Colorado and the average distance between cotwins on a given day were assessed as validity checks. Prior to age 18, 97% of all staypoints were recorded within the state of Colorado. The median distance between two twins from the same family on a given day was 1.9 km while it was 52.4 km between randomly matched participants, suggesting a high degree of overlap in locations recorded by Cotwins on the same day. The reliability of the mobility measures was measured using repeated split-half reliabilities with randomly sampled days of the month (k = 10,000) was 0.79 for daily distance travelled and 0.88 for daily locations visited.
Daily Mobility Measures.
To ensure comparability to the existing literature, we selected two common measures used in psychological studies of human mobility: the number of locations visited per day and daily distance travelled. The number of locations visited per day was measured using the number of staypoints registered for a participant on a given day (Canzian & Musolesi, 2015). The total distance travelled by a participant on a given day was estimated by summing the distance between staypoints registered on that day (Canzian & Musolesi, 2015).
Big Five Inventory.
Participant personality was assessed at an in-person intake assessment using the Big Five Inventory (BFI) (John et al., 2008). The BFI consisted of 44 statements on how participants view themselves, which they rate on a five-point Likert scale from “Strongly Disagree” to “Strongly Agree.” Averaging items for five subscales yields personality trait scores for Openness, Conscientiousness, Extraversion, Agreeableness, and Neuroticism (see Table 2 for descriptive statistics). For ease of interpretation in regression models, participants’ personality trait scores were z-scored with respect to the overall sample’s mean and standard deviation.
Table 2.
Descriptive Statistics for participant location data, mobility measures and personality scores
| Mean (SD) | Minimum, Maximum | Skewness | Kurtosis | Missing Cases (%) | |
|---|---|---|---|---|---|
| Mobility Measures | |||||
| Raw Locations | 19,645.2 (24,899.4) | 2.0; 243,516.0 | 3.85 | 24.43 | 313,810 (38.4%) |
| Stay points | 1,890.9 (1,674.8) | 1.0; 7,841.0 | 0.81 | 2.71 | 347,932 (44.6%) |
| Locations per Day | 4.0 (0.7) | 2.0; 17.0 | 0.87 | 0.54 | 347,932 (44.6%) |
| Distance Travelled (km) | 19.4 (9.2) | 0.0; 200.0 | 2.75 | 9.06 | 347,932 (44.6%) |
| Big Five Inventory | |||||
| Openness | 3.5 (0.6) | 1.4; 5.0 | −0.17 | 0.20 | 3 (0.3%) |
| Conscientiousness | 3.6 (0.6) | 1.4; 5.0 | −0.02 | −0.31 | 3 (0.3%) |
| Extraversion | 3.3 (0.8) | 1.0; 5.0 | −0.21 | −0.50 | 3 (0.3%) |
| Agreeableness | 4.0 (0.6) | 2.0; 5.0 | −0.29 | −0.24 | 3 (0.3%) |
| Neuroticism | 2.8 (0.8) | 1.0; 5.0 | 0.20 | −0.19 | 3 (0.3%) |
Sex.
Sex differences can lead to artefactual correlations in twin analyses (McGue & Bouchard, 1984). Therefore, we included sex as a covariate to account for possible sex differences in mobility patterns. All participants in the sample self-reported identifying as either male or female at intake.
Seasonality.
To account for differences in mobility because of the school year calendar, binary seasonality was introduced as a covariate. Seasonality was coded as Summer/another season, where Summer was defined as the period between May 31st and September 1st each year.
Weekend.
To account for mobility differences between weekends and weekdays, weekend/weekday status was included as a binary covariate. Saturdays and Sundays were coded as weekend days while Monday-Friday were coded as weekdays.
Operating System.
Participants provided information on their phone’s operating system during recruitment. The Cotwins smartphone application collected location data differently in the iOS and Android operating systems. For iOS users, locations were recorded using the significant change API, which recorded a participant’s location whenever a change of more than a few hundred meters was detected. For Android users, participant locations were recorded once every five minutes. To account for mean differences in the mobility measures resulting from these different collection strategies, participant operating system was included as a fixed effect covariate.
Zygosity.
Participant zygosity was determined by two independent coders who assessed the twins for similarity across various physical traits such as hair color, eye color, and complexion. Similarities were rated on a five-point scale ranging from very similar to not at all similar. After discussing any rating discrepancies, testers made a consensus determination as to whether the twin pair was a monozygotic pair, a dizygotic pair, or an opposite-sex pair. Opposite sex twins were included as dizygotic twins for analytic purposes.
Analytic strategy
Given the novelty of our data, including years of GPS data, generalized additive mixture models (GAMM) were fit to characterize changes in mean mobility with age. These models were fit using the gamm4 package in R and included a smooth age term, fit by penalized spline regression, fixed effects of participant sex, operating system, seasonality, and weekend, and random intercepts for individuals and families (Wood & Scheipl, 2020). GAMMs can useful to characterize change dynamics as they utilize penalized spline regression to allow for smooth highly non-linear models of the mean without assuming a prespecified functional form, and unlike other smoothers (e.g., loess) can readily and appropriately accommodate our nested family data. We consider the GAMM results reported here to be descriptive, as they can be difficult to tune and unfortunately do not provide a model for the variance (Wood, 2017). To characterize the means and variance-covariance structure of age trajectories, we used linear mixed models with nested individual and family-level random effects of age using the nlme package in R (Faraway, 2005; Pinheiro et al., 2021). Both the generalized additive mixed models and mixed effects regression models included fixed effects of participant sex, operating system, seasonality, and weekend. Age was standardized to have a mean of zero and standard deviation of one across the entire sample to improve convergence. Binary covariates were dummy coded (0 and 1).
For both mobility measures, models with increasingly complex random effects structures were fit, up to and including linear, quadratic, and cubic effects of age. Ultimately, models with random intercepts and random age linear, quadratic, and cubic effects were chosen due to superior fit, as measured by AIC and BIC indices (Kenny, 2020). Ninety-five percent confidence intervals were estimated using non-parametric bootstrap resampling of twin pairs (1000 replications).
To maintain consistency with prior literature on personality and mobility (Ai et al., 2019; Alessandretti et al., 2018), all five personality factors were included simultaneously in these mixed effects models. While multicollinearity amongst personality traits can affect estimated regression coefficients, models that included each personality trait separately delivered highly similar results and thus we deemed it appropriate to interpret the results of these multivariate models (Rabe-Hesketh & Skrondal, 2012). The results of these univariate personality models are reported in Supplement A.
Additional multivariate “ACE” models, conducted in the OpenMx R package, were used to estimate the relative contribution of genetic and environmental factors to participant mobility patterns and personality traits as well as the genetic and environmental correlations between them (Boker et al., 2021). ACE models decompose variance in a trait into that attributable to additive genetic (A), shared environmental (C), and non-shared environmental (E) influences by comparing the within-family correlations for the trait between monozygotic (MZ) and dizygotic (DZ) twin pairs (Martin & Eaves, 1977; Neale, 2009). Shared environmental influences (C) are those that make members of the same family more similar, whereas non-shared environmental influences (E) are those that make members of the same family less similar. MZ and DZ twin pairs differ in their degree of genetic relatedness, with MZ pairs sharing 100% of their genomes identical by descent, and dizygotic pairs sharing on average 50% identical by descent. Both MZ and DZ twin pairs are assumed to share 100% of their C influences since both MZ and DZ twin pairs share their rearing environment. Finally, MZ and DZ twin pairs share none of their E influences. Because of this difference in within-pair genetic relatedness, greater trait similarity in MZ pairs relative to DZ pairs is attributed to A. C is observed when DZ twins are more similar on a trait than what would be expected considering their genetic similarity (when the DZ correlation is greater than half the MZ correlation). E is observed when the MZ correlation is less than 1. Additionally, multivariate ACE models can be extended to decompose the covariance in two phenotypes into genetic (rg), shared environmental (rc), and non-shared environmental (re) components representing the extent to which shared genetic and environmental factors influence both traits.
Latent intercepts for each mobility and personality measure were estimated from the observed data and these intercepts were biometrically decomposed in a multivariate ACE model. An analysis of both daily mobility measures and five personality sumscores was infeasible: the resulting random intercept model would have included 2,587 manifest variables and model complexity would have been unrealistically high; runtime was estimated by extrapolation at more than a decade on a single-threaded CPU.
To reduce computational complexity, we removed Agreeableness and Neuroticism from the models as they were not significantly related to any of the daily mobility measures. Next, the daily mobility variables were averaged within weeks. This reduced the average number of observations per participant from 346.3 (days) to 75.6 (weeks). Next, rather than estimating the genetic and environmental correlations between the mobility measures in a single model, these were estimated in a piecewise fashion. Two univariate mobility models, one for daily locations visited and one for daily distance travelled, were used to estimate the genetic and environmental variance components for both traits. Additionally, a bivariate mobility model which included both mobility measures was used to estimate the genetic and environmental correlations between the mobility measures. Finally, after initial models found that the shared environmental components for the three significant personality traits were equal to zero, these components and the shared environmental correlations between personality and mobility were removed from the model.
Point estimates and 95% confidence intervals for the variance components (taken from the univariate mobility models) were computed using bootstrap resampling with 1000 replications. Confidence intervals for the more computationally expensive covariance components (taken from the bivariate mobility models) were computed using bootstrap resampling with only 100 replications.
Power analyses were conducted for the multivariate ACE models using a simulation-based approach outlined in the literature (Verhulst, 2017). For traits without shared environmental influences (C = 0.00), the sample achieved 80% power when A = 0.51; for traits with modest shared environmental influences (C = 0.20), 80% power was achieved when A = 0.42. For genetic correlations, rg, assuming no shared environmental influences, 80% power was attained at rg = 0.69 for two moderately heritable traits (As = 0.50, Cs = 0.00), and at rg = 0.34 for two highly heritable traits (As = 0.75, Cs = 0.00). Assuming modest shared environmental influences (C = 0.20), the sample attained 80% power at rg = 0.50 for two moderately heritable traits (As = 0.50, Cs = 0.20), and at rg = 0.21 for two highly heritable traits (As = 0.75, Cs = 0.20).
Missing Data
Missing personality and demographic data were minimal, with no variable missing for more than 0.3% of participants. These 0.3% of participants contributed a total of 865 days of staypoint data, representing 0.3% of all recorded days. These data were assumed to be missing at random; the GAMMs and linear mixed effects models did not include these participants because they were missing on the independent variables. The multivariate ACE models used full information maximum likelihood, so did include all available data. To account for missing OS data (such as when participants switched cases phone operating system partway through the study), participant operating system was identified using logistic regression. This process was utilized for 55 participants whose operating system was not available for part or all of the study. When determining participant operating system, GPS metadata, such as the estimated accuracy of each point, the number of points per month, and the number of days with at least one point, as well as demographic factors such as participant sex, family income, and family education level, were included as predictors. The resulting logistic regression explained 52.1% of the variance in participant operating systems (as measured by McFadden’s pseudo-R2).
Validation analyses of the staypoint data revealed that, when data was missing, it was missing in chunks of multiple days. Staypoints were recorded on successive days 76% of the time while the remaining 24% were recorded after a gap of at least one day. Additionally, 458 participants exhibited at least one gap greater than one month in length where no staypoints were recorded, (median length = 80 days) possibly reflecting software updates, poor cell phone service, deleting the application, changing cell phones, or temporarily leaving the study. To assess whether results were biased by these recording problems, we controlled for the proportion of days on which no locations were recorded in supplemental sensitivity analyses for the mixed effects models and multivariate ACE models (Appendices C and D).
Results
Demographic information is provided in Table 1 and participants’ mean mobility and personality scores are reported in Table 2. Participants contributed an average of 19,645.2 unique GPS locations and 1,890.9 stay points over the course of the study. Means of the daily mobility measures were not significantly different between monozygotic and dizygotic twin pairs (min(p) = 0.47). Dizygotic twin pairs were significantly higher on Extraversion than monozygotic pairs (t(565) = 2.28, p = 0.02; Hedge’s g = 0.17) and marginally higher on Openness (t(604) = 1.83, p = 0.07, Hedge’s g = 0.07). Reliabilities for each of the big five personality factors were measured using McDonald’s omega (Hayes & Coutts, 2020). Omega values ranged from 0.73 (Agreeableness) to 0.81 (Extraversion).
Generalized Additive Mixture Models
GAMM results (Figure 1) revealed mean increases in both mobility measures from early to late adolescence and a levelling off or slight decrease in mobility from late adolescence into emerging adulthood. Daily locations visited increased between age 15 and age 18 before declining slightly between age 18 and age 21. Developmental changes in the average number of daily locations were modest. Participants registered an average of about four locations per day: increasing from a low of 3.6 at age 15 to a high of 4.4 per day at age 18 and declining slightly to about 4.0 locations a day at age 21. Contrastingly, daily distance travelled increased, though with decelerating growth after about age 18, over the full observation period, from a low of about 10km per day at age 15 to a high of about 20km per day at age 21. Patterns observed before age fifteen and after age twenty-one were less interpretable due to the small number of participants contributing data at these ages.
Figure 1: Smoothed means with 95% confidence intervals (obtained via GAMMs) and mixed effects model paths of mobility variables conditioned on age.
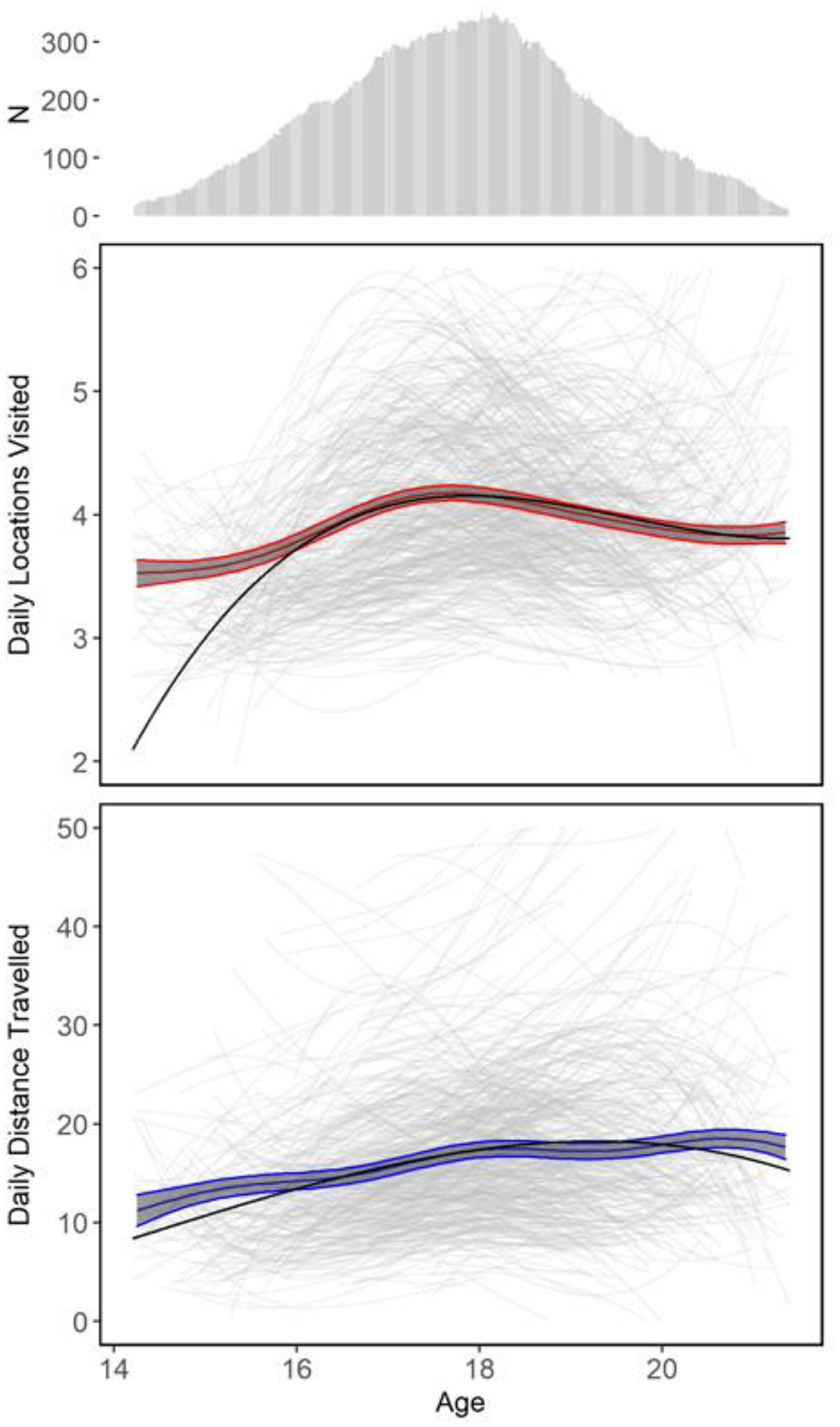
Smoothed means and mixed effects models of locations visited and distance travelled as a function of participant age. Smoothed mean models (represented by the colored curves) were calculated using generalized additive mixture models (GAMMs) with random intercepts of individuals nested in families. Participant operating system, weekday/weekend status, and school year/summer vacation status were included as fixed effect covariates. Uncertainty in the mean is given by 95% confidence intervals and represented by the shaded gray area between the colored lines. Mixed effect models are represented by the thicker black lines, the fixed effects, and transparent gray curves, the random effects. These included linear, quadratic, and cubic fixed and random effects of age with random effects nested within individuals and families. Participant sex, participant operating system, weekend/weekday status, and school year/summer vacation status were dummy-coded as fixed effect covariates. The number of participants providing data at each age is provided in the marginal histogram above both columns.
Linear Mixed Effects Models
After fitting models with increasingly complex random effects structures, a model with random intercepts, age slopes, and quadratic and cubic age effects at the individual and family level was selected due to improved fit on AIC BIC, and log-likelihood fit indices (Kenny, 2020). Fit indices for the various models considered are presented in Supplement B.
The results of the linear mixed effects models are presented in Table 3. Participants contributed an average of 346.3 observations to the daily mobility models. Residuals were reasonably normal for the model of daily locations visited but were right skewed and leptokurtic for the distance travelled model. Fixed effects of age, age2, or age3 significantly predicted both daily distance travelled and daily locations visited, indicating that mobility changed significantly over the course of adolescence. However, large random effects of these age parameters indicate that there were substantial individual differences in the developmental trajectories of mobility (Figure 1). Changes in mobility with age observed in the mixed effects models were largely consistent with those obtained via GAMM (Figure 1). Locations visited per day were negatively associated with age2 and positively associated with age3. As is evident graphically, (Figure 1.) this corresponded to modest increases in daily locations visited during adolescence after which these declined slightly. Similarly, distance travelled was positively associated with age and negatively associated with age2: Distance travelled initially increased with age, but this increase decelerated throughout late adolescence and, unlike in the GAMM results, distance travelled was found to slightly decrease during participants’ late teens and early 20s.
Table 3.
Mixed effect model coefficients and bootstrapped 95% confidence intervals for mobility measures
| Locations Visited | Distance | |
|---|---|---|
| Random Effects, (sum of individual and family-level random effects) | ||
| Intercept | 0.63 (0.59, 0.67) | 6.74 (6.10, 7.30) |
| Age | 0.54 (0.49, 0.59) | 5.26 (4.55, 5.88) |
| Age2 | 0.25 (0.21, 0.31) | 1.91 (1.56, 2.25) |
| Age3 | 0.20 (0.17, 0.24) | 1.42 (1.03, 1.82) |
| Fixed Effects | ||
| Intercept | 4.15 (4.10, 4.22) | 17.24 (16.47,17.96) |
| Age | −0.01 (−0.06, 0.05) | 1.92 (1.32, 2.54) |
| Age2 | −0.18 (−0.22, −0.14) | −0.84 (−1.15, −0.54) |
| Age3 | 0.05 (0.02, 0.07) | −0.01 (−0.34, 0.16) |
| Openness | −0.06 (−0.98, −0.02) | −0.82 (−1.21, −0.38) |
| Conscientiousness | 0.06 (0.02, 0.09) | 0.42 (0.01, 0.79) |
| Extraversion | 0.15 (0.11, 0.18) | 1.03 (0.65, 1.42) |
| Agreeableness | 0.00 (−0.04, 0.03) | 0.15 (−0.26, 0.55) |
| Neuroticism | 0.04 (0.00, 0.08) | −0.06 (−0.61, 0.46) |
| Sex | −0.03 (−0.11, 0.06) | −0.67 (−1.67, 0.31) |
| Summer | 0.16 (0.13, 0.19) | 6.07 (5.61, 6.53) |
| OS | −0.09 (−0.23, 0.02) | −3.67 (−5.09, −2.25) |
| Weekend | −0.14 (−0.17, −0.11) | 8.44 (7.91, 9.00) |
| Correlation of Random Effects Terms | ||
| Locations Visited, Family Effects | |||
|---|---|---|---|
| Intercept | Age | Age2 | |
| Age | −0.31 (−0.49, −0.14) | ||
| Age2 | −0.69 (−0.79, −0.57) | 0.26 (−0.04, 0.50) | |
| Age3 | 0.30 (0.13, 0.47) | −0.88 (−0.95, −0.79) | −0.16 (−0.55, 0.27) |
| Locations Visited, Individual Effects | |||
| Age | 0.11 (−0.04, 0.27) | ||
| Age2 | −0.54 (−0.66, −0.41) | 0.11 (−0.12, 0.34) | |
| Age3 | −0.10 (−0.26, 0.06) | −0.89 (−0.92, −0.83) | 0.09 (−0.24, 0.36) |
| Distance, Family Effects | |||
| Age | 0.11 (−0.05, 0.30) | ||
| Age2 | −0.29 (−0.60, 0.00) | 0.22 (−0.10, 0.47) | |
| Age3 | 0.01 (−0.27, 0.25) | −0.87 (−0.94, −0.78) | −0.15 (−0.46, 0.22) |
| Distance, Individual Effects | |||
| Age | 0.20 (−0.03, 0.43) | ||
| Age2 | −0.43 (−0.60, −0.23) | 0.24 (0.06, 0.40) | |
| Age3 | −0.04 (−0.23, 0.14) | −0.76 (−0.85, −0.64) | 0.02 (−0.21, 0.29) |
Coefficients are unstandardized. Age and personality are rescaled with m =0 and sd = 1. Covariates are 0/1 dummy coded (Sex, 0 =female/1=male, OS, 0 =iOS/1=android, seasonality, 0 =other/1=summer, weekend, 0=weekday/1=weekend).
Over the summer, participants visited about one additional location per week and travelled about six additional kilometers per day. Compared to iOS users, Android users registered travelling about 3.7 fewer kilometers per week. Weekends were associated with slightly fewer locations visited but also with an additional 8.4 kilometers travelled per day. Sex was not a significant predictor of mobility. As a sensitivity analysis, an additional supplemental model was run for both mobility measures which included an age by sex interaction term. The effect of this interaction term was non-significant for both mobility measures (ps > 0.4).
Extraversion, Conscientiousness, and Openness significantly predicted both mobility measures. A one standard deviation increase in Extraversion was associated with 0.15 additional locations per day (about 1.05 additional locations per week), and an additional kilometer of distance travelled per day. Conscientiousness was also positively associated with both distance travelled and locations visited while Openness was negatively associated with the measures. All results are provided in Table 3.
The correlation between the proportion of missing cases and the big five personality traits is reported in supplement C. Missingness was significantly associated with Extraversion (r = −0.19, p < 1*10−5), Conscientiousness (r = −0.17, p < 1*10−5), and Openness (r = 0.09, p = 0.011). After including the proportion of missing days as a fixed effect covariate, the magnitude and significance of the intercept, age, age2, and age3 terms were unchanged in both models. Extraversion and openness remained significant predictors of both mobility measures with magnitudes comparable to those observed in the original models. However, conscientiousness was no longer a significant predictor of daily locations visited or daily distance travelled. The proportion of missing days was significantly, negatively related to the mobility measures, indicating that participants with more missing recording days registered fewer daily locations and less travel distance. Results of this supplementary analysis are reported in Supplement D.
Multivariate ACE Models
The results of the multivariate ACE models revealed that both genetic and environmental effects contributed to daily mobility (Table 4). Daily locations visited was influenced by both moderate genetic and shared environmental components (A = 0.56, C = 0.28). Heritability estimates for distance travelled were large and the 95% confidence bound for the shared environmental term included zero (A = 0.81, C = 0.12).
Table 4.
ACE decomposition of mobility measure random intercepts and their correlations with Big Five personality traits (bootstrapped 95% confidence intervals)
| Locations Visited | Daily Distance Travelled | |
|---|---|---|
| Correlations Between Random Intercepts of Mobility Measures and Personality Traits | ||
| Locations Visited | 1 | |
| Distance | 0.56 (0.51, 0.64) | 1 |
| Openness | −0.11 (−0.18, −0.04) | −0.13 (−0.19, −0.06) |
| Extraversion | 0.25 (0.20, 0.31) | 0.17 (0.11, 0.23) |
| Conscientiousness | 0.16 (0.10, 0.22) | 0.10 (0.05, 0.15) |
| ACE Decomposition of Random Intercepts of Mobility Measures | ||
| Additive Genetic (A) | 0.56 (0.38, 0.74) | 0.81 (0.58, 0.95) |
| Shared Environment (C) | 0.28 (0.14, 0.45) | 0.12 (0.00, 0.34) |
| Non-Shared Environment (E) | 0.15 (0.09, 0.21) | 0.07 (0.05, 0.10) |
| Personality-Mobility Genetic Correlations (rg) | ||
| Locations Visited | 1 | |
| Distance | 0.75 (0.64, 0.87) | 1 |
| Openness | −0.27 (−0.45, −0.09) | −0.27 (−0.39, −0.13) |
| Extraversion | 0.53 (0.39, 0.71) | 0.31 (0.22, 0.47) |
| Conscientiousness | 0.29 (0.17, 0.45) | 0.13 (0.02, 0.26) |
| Mobility Shared Environmental Correlations (rc) | ||
| Locations Visited | 1 | |
| Distance | −0.10 (−1.00, 0.82) | 1 |
| Personality-Mobility Non-Shared Environmental Correlations (re) | ||
| Locations Visited | 1 | |
| Distance | 0.58 (0.41, 0.73) | 1 |
| Openness | 0.05 (−0.09, 0.19) | 0.15 (−0.03, 0.40) |
| Extraversion | 0.05 (−0.10, 0.15) | −0.02 (−0.20, 0.16) |
| Conscientiousness | 0.02 (−0.10, 0.15) | 0.10 (−0.13, 0.30) |
| AE Decomposition of Big Five Personality Traits | ||
| Additive Genetic (A) | Non-Shared Environment (E) | |
| Openness | 0.44 (0.32, 0.53) | 0.56 (0.47, 0.68) |
| Extraversion | 0.38 (0.25, 0.59) | 0.62 (0.50, 0.75) |
| Conscientiousness | 0.48 (0.36, 0.61) | 0.52 (0.39, 0.64) |
| Agreeableness | 0.32 (0.26, 0.40) | 0.68 (0.60, 0.74) |
| Neuroticism | 0.53 (0.48, 0.58) | 0.47 (0.42, 0.52) |
Shared environmental contributions to personality were fixed at 0. Bolded values indicate that bootstrapped 95% confidence intervals did not include zero. AE estimates for agreeableness and neuroticism were estimated in separate models.
As expected in variance components tests with smaller sample sizes (Maes, 2014), bootstrapped confidence intervals revealed substantial uncertainty around these point estimates. Genetic and environmental correlations between personality and daily mobility are presented in Table 4. Daily locations visited and distance travelled were highly genetically correlated with one another, (rg = 0.75) suggesting highly similar, though not identical, genetic influences on these two measures. Significant genetic correlations were observed between the daily mobility measures and the three personality traits included in the models: Openness, Extraversion, and Conscientiousness (Agreeableness and Neuroticism were not included in the models as they were not found to be significantly related to mobility). Genetic correlations between personality and mobility measures were generally moderate in magnitude and ranged from 0.13 between Conscientiousness and daily distance travelled to 0.53 between extraversion and locations visited.
The shared environmental correlation between the mobility measures was imprecisely estimated with wide bootstrapped confidence intervals which included 0 (rc = −0.10). Conversely, their non-shared environmental correlation was re = 0.58, suggesting substantially overlapping non-shared environmental influences. Non-shared environmental correlations between personality traits and mobility measures were all non-significant. Shared environmental correlations were not estimated as the shared-environmental variance in big five personality traits were estimated to be 0.
As a further sensitivity analysis to assess whether heritability estimates were influenced by the inclusion of opposite sex dizygotic twins in the models, the multivariate ACE models were run again with opposite sex twin pairs excluded from the analyses. These models yielded standardized ACE estimates of A = 0.49, C = 0.36, and E = 0.15 for daily locations visited and A = 0.63, C = 0.30, and E = 0.07 for daily distance travelled. To test whether estimates of A, C, and E were significantly different after removing opposite sex twin pairs, we compared the fit of freely estimated models in the full sample to reduced models where A, C, and E were fixed at the values obtained after excluding opposite sex twin pairs. Log-likelihood ratio tests comparing the model fits obtained in these analyses revealed that differences in the two ACE estimates were negligible (all ps > 0.9) and so results did not appear biased by the inclusion of opposite sex twins. Lastly, results of supplementary analyses conducted after controlling for the proportion of missing days were highly similar to those obtained in table 4. Results of these analyses are reported in Supplement E.
Discussion
This study investigated the genetic and environmental influences on personality and mobility patterns over the course of adolescence and emerging adulthood, a period characterized by substantial lifestyle changes and increases in personal autonomy. We found mean increases in overall mobility over the course of adolescence (though with substantial individual variation in these trajectories) as well as small to moderate relationships between mobility and three of five big five personality factors, Extraversion, Openness, and Conscientiousness. Additionally, we found that both daily distance travelled and daily locations visited were moderately or highly heritable and that daily locations visited was modestly influenced by shared-environmental factors. We observed large genetic correlations between mobility measures, indicating that genetic influences are largely shared between them. Additionally, significant genetic correlations of variable magnitude were observed between each of the personality and mobility measures, indicating that mobility is heritable in part due to similar genetic influences to personality.
As our sample of Colorado youth grew older, on average they visited more locations per day and travelled further each day. The number of locations per day increased by about 20%, from a low of 3.6 at age 15 to 4.4 at age 18, while daily distance travelled doubled from roughly 10km/day at age 15 to about 20km/day at age 21. While there were clear mean changes in these behaviors over time, we observed substantial individual differences in these trajectories. This suggests that increases in autonomy and exploration during adolescence are on average reflected in increasing mobility but that there is a great deal of variability in the way that adolescents move through and engage with their environment.
Three of five big five personality factors were significantly related to mobility. The largest associations were with extraversion, where a one standard deviation increase in extraversion predicted about one additional location visited per week and an additional kilometer per day of distance travelled. Smaller, significant relationships were also observed between mobility and conscientiousness, which was positively associated with the mobility measures, and openness, which was negatively associated with them. Extraversion’s robust, small to moderate sized positive relationship with mobility is broadly consistent with the existing literature, which has repeatedly found that Extraversion is the personality trait most strongly related to mobility patterns (Ai et al., 2019; Chorley et al., 2015; Stachl et al., 2017). The relationship between higher extraversion and increased mobility may reflect greater socializing behavior or seeking out different, socially stimulating environments.
Prior findings reported in the literature on the relationship between conscientiousness and mobility were more mixed. In contrast to this sample, in which conscientiousness was positively associated with daily mobility, Ai et al. (2019) found a negative relationship between conscientiousness and daily locations visited. Contrastingly, in a social network-based study on personality and mobility, Chorley et al. (2015) found that conscientiousness was positively associated with the number of locations visited (Chorley et al., 2015). Additionally, conscientiousness, unlike extraversion and openness, was not significantly related to mobility in the supplementary analysis controlling for gaps in recording.
Conscientiousness measures behaviors related to achievement motivation, self-control, and adherence to rules and behavioral norms. Conceivably, these traits may influence mobility differently across different developmental periods or cultural contexts, leading to these inconsistencies across studies. Positive relationships between mobility and conscientiousness amongst US adolescents may, for example, reflect taking on additional family responsibilities that require travel, increased engagement in extracurricular activities, or starting a job. Additionally, the fact that this relationship becomes non-significant after controlling for missing data may suggest that more conscientious individuals may have reported greater mobility because they were more diligent in ensuring their locations were being recorded properly.
Prior studies that have looked at relationships between openness and daily locations visited or daily distance travelled have not reported a negative relationship between them (Ai et al., 2019; Chorley et al., 2015). Openness in part measures interest in intellectual and artistic pursuits which, similarly to conscientiousness, could exhibit relationships to mobility that are moderated by cultural or developmental factors. For example, a negative relationship between openness and mobility in US adolescents may result from increased engagement in sedentary leisure activities like reading or making art (Barnett, 2006; Caprara et al., 2011; Proctor & McCord, 2009).
Neither Agreeableness nor Neuroticism were significantly related to mobility in our sample. Agreeableness seems particularly weakly related to mobility; we found no significant relationships between agreeableness and our mobility measures reported in the literature. Stachl et al. (2017) reported that, despite their use of a sophisticated machine learning algorithm, they were entirely unable to predict agreeableness from smartphone sensor data. The lack of association in our study and in prior studies suggests that agreeableness is likely unrelated to mobility.
Similarly, there were few significant mobility-Neuroticism relationships reported in the literature, apart from Chorley et al. (2015), who found a significant relationship between Neuroticism and daily locations visited (as measured by check ins on a location-based social network). Notably, mobility measures have been used to predict depression symptoms, which are strongly associated with Neuroticism (Canzian & Musolesi, 2015; Kotov et al., 2010; Ren et al., 2022). This relationship between mobility and depression symptoms suggests that mobility may be related to neuroticism at clinical levels, when it manifests in depression symptoms like lethargy, loss of interest in activities, or excessive sleep, but that the relationship is not found at subclinical levels of Neuroticism where such symptoms are less evident.
Both mobility traits were moderately to highly heritable (As = 0.56–0.81), demonstrating that mobility, like nearly all human behaviors and traits, is influenced by genetic factors (Polderman et al., 2015). The heritability of these mobility measures suggest that they are validly capturing aspects of individuals daily behaviors which may reflect other heritable traits, such as adolescent personality, academic engagement, or work and leisure preferences.
Significant shared environmental effects were observed for daily locations visited but not daily distance travelled. Shared environmental effects are uncommon for complex traits in behavioral genetics research (Bouchard & McGue, 2003; Turkheimer, 2000). Nonetheless, adolescents of the same age in the same family likely experience similar pressures and constraints on their movement patterns like living in the same home, attending the same schools, and having access to similar recreational opportunities. Consistent with this hypothesis, time allocation in leisure activities in adolescence displays high estimates of shared environmental contributions (Haberstick et al., 2014). Shared environmental effects observed in this sample may be less evident later in adulthood when such constraints are absent.
Genetic correlations amongst the mobility measures were large, suggesting there is a shared genetic basis for mobility patterns more generally. Large re correlations were also observed, suggesting this is also true for non-shared environmental effects. It should be noted however that this is influenced by how distance travelled was measured; these large correlations in part reflect that the number of locations visited influences the computation of daily distance travelled.
Relationships between personality and mobility were partially explained by shared genetic effects. This pattern was particularly pronounced for extraversion, which showed moderate genetic correlations with the mobility measures, suggesting a shared genetic relationship between extraversion and mobility. This is consistent with our hypothesis that mobility patterns are heritable in part because they reflect heritable preferences measured by personality inventories like the BFI. Adolescents’ genetic predispositions to seek out socially stimulating environments, commit to responsibilities, achieve academically or at work, and to engage with artistically or intellectually stimulating activities all appear to influence how they move about and engage with the world around them.
Strengths and Limitations
This investigation had several important strengths. To our knowledge, no previous study has investigated mobility trajectories during the multi-year transition from adolescence into emerging adulthood. The CoTwins sample is relatively large and includes multiple years of persistent GPS location data. This novel sample also allowed us to conduct twin based analyses to estimate the relative genetic and environmental contributions to mobility measures and their relationships to personality. This work therefore represents the first effort to characterize the developmental course and heritability of common GPS mobility measures in a youth sample; it has demonstrated that these mobility measures are strongly influenced by genetic factors, in part through their relationship to personality.
Nonetheless, several important limitations must be noted. Particularly for daily locations visited, much of the developmental change in daily mobility occurred prior to age 16, when the study sample size was relatively low. This suggests that changes estimated at these ages may be somewhat less reliable than those measured later in development. In part due to the study’s long timeframe and the use of a battery-conserving location request API, location data are sparser in the CoTwins sample than in many shorter-term GPS studies and contains significant gaps in recording. Though we have attempted to account for these gaps through statistical controls, such controls may not be fully capturing how missingness influences the results of this investigation. As missingness was significantly negatively related to the mobility measures, these analyses may reflect an underestimate of the true extent of adolescent mobility. Additionally, this finding may be indicative that days were not missing at random which may be a further source of bias in the results obtained here. Additionally, the CoTwins sample is whiter, wealthier, and more educated than the population of United States youth, which may reduce the generalizability of the findings to more diverse populations. While personality is a relatively stable construct, personality changes occur during development (Soto et al., 2011), leading to the possibility that participants’ personalities throughout the study may have differed somewhat from their measures at intake. Additionally, though the big five inventory has previously been shown to exhibit metric invariance across gender in community, adult samples (Ock et al., 2020), it is nonetheless possible that our personality construct may have differed by gender in our sample, especially given that these were adolescents and not adults. Both daily distance travelled and daily locations visited exhibited right-skewed distributions and the residuals for the mixed effects models of distance were non-normally distributed. While bootstrapping was used to limit the effect of this non-normality on the estimation of confidence intervals, point estimates in the linear mixed effects models may have been inflated somewhat by the presence of outlying values. When estimating genetic and environmental effects, ACE models make use of several assumptions, namely that environmental effects are of equal magnitude for monozygotic and dizygotic twins, that parents do not engage in assortative mating, and that genes and environments do not statistically interact for the trait of interest, all of which may be violated to some extent for complex behavioral traits like mobility and personality.
Conclusion
This study has shown how mobility patterns change over time in a community sample of American youth, how genetic and environmental factors contribute to these behaviors, and how psychological differences are reflected in differences in mobility patterns. In doing so, these findings may offer an interesting window into the life transition from adolescence into emerging adulthood. As adolescents grow increasingly independent from their parents, obtain new privileges such as driving, and increase in their needs for autonomy, exploration, and social connectedness, we see a corresponding increase in their mobility. In adulthood, as these opportunities grow less novel and responsibilities such as working or attending college increase, we see corresponding reductions in their mobility. However, these patterns only describe mean trends around which very large individual differences exist. No single pattern or narrative can explain the ways in which adolescents move through and engage with their environments. Nonetheless, this research demonstrates that passive GPS measures reflect meaningful differences in the genes, environments, and psychologies of our participants. Human mobility research thus represents a promising avenue for characterizing the environments in which we live, the ways in which we engage with them and how we are influenced by them. Learning how and why these behaviors develop during a critical period in human development can help shed light on this fundamental question in psychology: “how do our experiences shape who we become?”
Supplementary Material
Acknowledgements:
We would like to thank the CoTwins Research team, including Amy Ledbetter, Enning Tan, Jacob Anderson, Rio Hindami, Ming Jiang, Robin Corley, and Yan Luo for their invaluable contributions to this project. We would also like to acknowledge the individual contributions of Dr. Nathan Helwig and Stephanie Zellers for their help and feedback on the analytic strategy. This project was funded by the National Institute on Drug Abuse award U01 DA 046413.
Footnotes
Preregistration Statement
This study was not preregistered with any independent institutional registry.
References
- Ai P, Liu Y, & Zhao X (2019). Big Five personality traits predict daily spatial behavior: Evidence from smartphone data. Personality and Individual Differences, 147, 285–291. 10.1016/j.paid.2019.04.027 [DOI] [Google Scholar]
- Alberga AS, Sigal RJ, Goldfield G, Prud’ homme D, & Kenny GP (2012). Overweight and obese teenagers: Why is adolescence a critical period? Pediatric Obesity, 7(4), 261–273. 10.1111/j.2047-6310.2011.00046.x [DOI] [PubMed] [Google Scholar]
- Alessandretti L, Lehmann S, & Baronchelli A (2018). Understanding the interplay between social and spatial behaviour. EPJ Data Science, 7(1), 1–17. 10.1140/epjds/s13688-018-0164-6 [DOI] [Google Scholar]
- Andrade T, Cancela B, & Gama J (2019). Mining Human Mobility Data to Discover Locations and Habits. ArXiv:1909.11406 [Cs, Stat]. http://arxiv.org/abs/1909.11406 [Google Scholar]
- Barnett LA (2006). Accounting for Leisure Preferences from Within: The Relative Contributions of Gender, Race or Ethnicity, Personality, Affective Style, and Motivational Orientation. Journal of Leisure Research, 38(4), 445–474. 10.1080/00222216.2006.11950087 [DOI] [Google Scholar]
- Billig JP, Hershberger SL, Iacono WG, & McGue M (1996). Life events and personality in late adolescence: Genetic and environmental relations. Behavior Genetics, 26(6), 543–554. 10.1007/BF02361227 [DOI] [PubMed] [Google Scholar]
- Boker S, Neale M, Maes H, Wilde M, Spiegel M, Brick T, Spies J, Estabrook R, Kenny S, Bates T, Mehta P, & Fox J (2011). OpenMx: An Open Source Extended Structural Equation Modeling Framework. Psychometrika, 76(2), 306–317. 10.1007/s11336-010-9200-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouchard TJ, & McGue M (2003). Genetic and environmental influences on human psychological differences. Journal of Neurobiology, 54(1), 4–45. 10.1002/neu.10160 [DOI] [PubMed] [Google Scholar]
- Bureau U. C. (n.d.). 2019. Population Estimates by Age, Sex, Race and Hispanic Origin. Census.Gov. Retrieved August 1, 2022, from https://www.census.gov/newsroom/press-kits/2020/population-estimates-detailed.html
- Canzian L, & Musolesi M (2015). Trajectories of depression: Unobtrusive monitoring of depressive states by means of smartphone mobility traces analysis. Proceedings of the 2015 ACM International Joint Conference on Pervasive and Ubiquitous Computing - UbiComp ‘15, 1293–1304. 10.1145/2750858.2805845 [DOI] [Google Scholar]
- Caprara GV, Vecchione M, Alessandri G, Gerbino M, & Barbaranelli C (2011). The contribution of personality traits and self-efficacy beliefs to academic achievement: A longitudinal study. British Journal of Educational Psychology, 81(1), 78–96. 10.1348/2044-8279.002004 [DOI] [PubMed] [Google Scholar]
- Chambers RA, Taylor JR, & Potenza MN (2003). Developmental Neurocircuitry of Motivation in Adolescence: A Critical Period of Addiction Vulnerability. American Journal of Psychiatry, 160(6), 1041–1052. 10.1176/appi.ajp.160.6.1041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chorley MJ, Whitaker RM, & Allen SM (2015). Personality and location-based social networks. Computers in Human Behavior, 46, 45–56. 10.1016/j.chb.2014.12.038 [DOI] [Google Scholar]
- Csikszentmihalyi M, Larson R, & Prescott S (2014). The Ecology of Adolescent Activity and Experience. In Csikszentmihalyi M (Ed.), Applications of Flow in Human Development and Education: The Collected Works of Mihaly Csikszentmihalyi (pp. 241–254). Springer; Netherlands. 10.1007/978-94-017-9094-9_12 [DOI] [Google Scholar]
- DeYoung CG, Peterson JB, Séguin JR, & Tremblay RE (2008). Externalizing behavior and the higher order factors of the Big Five. Journal of Abnormal Psychology, 117(4), 947–953. 10.1037/a0013742 [DOI] [PubMed] [Google Scholar]
- Faraway JJ (2005). Extending the Linear Model with R: Generalized Linear, Mixed Effects and Nonparametric Regression Models (1st edition). Chapman and Hall/CRC. [Google Scholar]
- Grimes DA, & Schulz KF (2002). Bias and causal associations in observational research. The Lancet, 359(9302), 248–252. 10.1016/S0140-6736(02)07451-2 [DOI] [PubMed] [Google Scholar]
- Haberstick BC, Zeiger JS, & Corley RP (2014). Genetic and Environmental Influences on the Allocation of Adolescent Leisure Time Activities. BioMed Research International, 2014, 1–12. 10.1155/2014/805476 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harari GM, Lane ND, Wang R, Crosier BS, Campbell AT, & Gosling SD (2016). Using Smartphones to Collect Behavioral Data in Psychological Science: Opportunities, Practical Considerations, and Challenges. Perspectives on Psychological Science, 11(6), 838–854. 10.1177/1745691616650285 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayes AF, & Coutts JJ (2020). Use Omega Rather than Cronbach’s Alpha for Estimating Reliability. But…. Communication Methods and Measures, 14(1), 1–24. 10.1080/19312458.2020.1718629 [DOI] [Google Scholar]
- Jacobson NC, & Bhattacharya S (2022). Digital biomarkers of anxiety disorder symptom changes: Personalized deep learning models using smartphone sensors accurately predict anxiety symptoms from ecological momentary assessments. Behaviour Research and Therapy, 149, 104013. 10.1016/j.brat.2021.104013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- John OP, Naumann LP, & Soto CJ (2008). Paradigm shift to the integrative Big Five trait taxonomy: History, measurement, and conceptual issues. In Handbook of personality: Theory and research, 3rd ed (pp. 114–158). The Guilford Press. [Google Scholar]
- Johnson DP, Rhee SH, Whisman MA, Corley RP, & Hewitt JK (2013). Genetic and Environmental Influences on Negative Life Events From Late Childhood to Adolescence. Child Development, 84(5), 1823–1839. 10.1111/cdev.12055 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kenny DA (2020, June). Measuring Model Fit. Davidakenny.Net. http://davidakenny.net/cm/fit.htm
- Kotov R, Gamez W, Schmidt F, & Watson D (2010). Linking “big” personality traits to anxiety, depressive, and substance use disorders: A meta-analysis. Psychological Bulletin, 136(5), 768–821. 10.1037/a0020327 [DOI] [PubMed] [Google Scholar]
- Maes HH (2014). ACE Model. In Balakrishnan N, Colton T, Everitt B, Piegorsch W, Ruggeri F, & Teugels JL (Eds.), Wiley StatsRef: Statistics Reference Online (1st ed.). Wiley. 10.1002/9781118445112.stat06752 [DOI] [Google Scholar]
- Martin NG, & Eaves LJ (1977). The genetical analysis of covariance structure. Heredity, 38(1), 79–95. 10.1038/hdy.1977.9 [DOI] [PubMed] [Google Scholar]
- McAdams TA, Gregory AM, & Eley TC (2013). Genes of Experience: Explaining the Heritability of Putative Environmental Variables Through Their Association with Behavioural and Emotional Traits. Behavior Genetics, 43(4), 314–328. 10.1007/s10519-013-9591-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGue M, & Bouchard TJ (1984). Adjustment of twin data for the effects of age and sex. Behavior Genetics, 14(4), 325–343. 10.1007/BF01080045 [DOI] [PubMed] [Google Scholar]
- McGue M, Osler M, & Christensen K (2010). Causal Inference and Observational Research: The Utility of Twins. Perspectives on Psychological Science, 5(5), 546–556. 10.1177/1745691610383511 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller G (2012). The Smartphone Psychology Manifesto. Perspectives on Psychological Science, 7(3), 221–237. 10.1177/1745691612441215 [DOI] [PubMed] [Google Scholar]
- Neale MC (2009). Biometrical Models in Behavioral Genetics. In Kim Y-K (Ed.), Handbook of Behavior Genetics (pp. 15–33). Springer; New York. 10.1007/978-0-387-76727-7_2 [DOI] [Google Scholar]
- Ock J, McAbee ST, Mulfinger E, & Oswald FL (2020). The Practical Effects of Measurement Invariance: Gender Invariance in Two Big Five Personality Measures. Assessment, 27(4), 657–674. 10.1177/1073191119885018 [DOI] [PubMed] [Google Scholar]
- Pinheiro J, Bates D, DebRoy D, Sarkar D, Heisterkamp S, & Van Willigen B (2021). Package “nlme” (3.1-152) [R]. http://cran.rapporter.net/web/packages/nlme/nlme.pdf
- Polderman TJC, Benyamin B, de Leeuw CA, Sullivan PF, van Bochoven A, Visscher PM, & Posthuma D (2015). Meta-analysis of the heritability of human traits based on fifty years of twin studies. Nature Genetics, 47(7), 702–709. 10.1038/ng.3285 [DOI] [PubMed] [Google Scholar]
- Proctor S, & McCord D (2009). Correlates of the Openness to Experience Domain. Individual Differences Research, 7, 222–227. [Google Scholar]
- Rabe-Hesketh S, & Skrondal A (2012). Multilevel and Longitudinal Modeling Using Stata, Third Edition (Third, Vol. 1). Stata Press. [Google Scholar]
- Remschmidt H (1994). Psychosocial Milestones in Normal Puberty and Adolescence. Hormone Research, 41(2), 19–29. 10.1159/000183955 [DOI] [PubMed] [Google Scholar]
- Ren B, Balkind EG, Pastro B, Israel E, Pizzagalli DA, Rahimi-Eichi H, Baker JT, & Webb CA (2022). Predicting states of elevated negative affect in adolescents from smartphone sensors: A novel personalized machine learning approach [Preprint]. PsyArXiv. 10.31234/osf.io/zjm4d [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santani D, Do T-M-T, Labhart F, Landolt S, Kuntsche E, & Gatica-Perez D (2018). DrinkSense: Characterizing Youth Drinking Behavior Using Smartphones. IEEE Transactions on Mobile Computing, 17(10), 2279–2292. 10.1109/TMC.2018.2797901 [DOI] [Google Scholar]
- Sathyanarayana A, Joty S, Fernandez-Luque L, Ofli F, Srivastava J, Elmagarmid A, Arora T, & Taheri S (2016). Sleep Quality Prediction From Wearable Data Using Deep Learning. JMIR MHealth and UHealth, 4(4), e125. 10.2196/mhealth.6562 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sawyer SM, Azzopardi PS, Wickremarathne D, & Patton GC (2018). The age of adolescence. The Lancet Child & Adolescent Health, 2(3), 223–228. 10.1016/S2352-4642(18)30022-1 [DOI] [PubMed] [Google Scholar]
- Soto CJ, John OP, Gosling SD, & Potter J (2011). Age differences in personality traits from 10 to 65: Big Five domains and facets in a large cross-sectional sample. Journal of Personality and Social Psychology, 100(2), 330–348. 10.1037/a0021717 [DOI] [PubMed] [Google Scholar]
- Stachl C, Au Q, Schoedel R, Gosling SD, Harari GM, Buschek D, Völkel ST, Schuwerk T, Oldemeier M, Ullmann T, Hussmann H, Bischl B, & Bühner M (2020). Predicting personality from patterns of behavior collected with smartphones. Proceedings of the National Academy of Sciences, 117(30), 17680–17687. 10.1073/pnas.1920484117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stachl C, Hilbert S, Au J, Buschek D, De Luca A, Bischl B, Hussmann H, & Bühner M (2017). Personality Traits Predict Smartphone Usage. European Journal of Personality, 31(6), 701–722. 10.1002/per.2113 [DOI] [Google Scholar]
- Steinberg L, & Morris AS (2001). Adolescent Development. Annual Review of Psychology, 52(1), 83–110. 10.1146/annurev.psych.52.1.83 [DOI] [PubMed] [Google Scholar]
- Turkheimer E (2000). Three Laws of Behavior Genetics and What They Mean. Current Directions in Psychological Science, 9(5), 160–164. 10.1111/1467-8721.00084 [DOI] [Google Scholar]
- Verhulst B (2017). A Power Calculator for the Classical Twin Design. Behavior Genetics, 47(2), 255–261. 10.1007/s10519-016-9828-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wood SN (2017). Generalized additive models: An introduction with R (Second edition). CRC Press/Taylor & Francis Group. [Google Scholar]
- Wood S, & Scheipl F (2020). Gamm4 (0.2–6) [R]. https://cran.r-project.org/web/packages/gamm4/gamm4.pdf
- Wrzus C, Hänel M, Wagner J, & Neyer FJ (2013). Social network changes and life events across the life span: A meta-analysis. Psychological Bulletin, 139(1), 53–80. 10.1037/a0028601 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.