

Abstract
Globally, kidney disease has become a serious health challenge, with approximately 10% of adults suffering with the disease, and increasing incidence and mortality rates every year. Small extracellular vesicles (sEVs) are 30 nm–100 nm sized nanovesicles released by cells into the extracellular matrix (ECM), which serve as mediators of intercellular communication. Depending on the cell origin, sEVs have different roles which depend on internal cargoes including, nucleic acids, proteins, and lipids. Mesenchymal stem cell (MSCs) exert anti-inflammatory, anti-aging, and wound healing functions mainly via sEVs in a stable and safe manner. MSC-derived sEVs (MSC-sEVs) exert roles in several kidney diseases by transporting renoprotective cargoes to reduce oxidative stress, inhibit renal cell apoptosis, suppress inflammation, and mediate anti-fibrosis mechanisms. Additionally, because MSC-sEVs efficiently target damaged kidneys, they have the potential to become the next generation cell-free therapies for kidney disease. Herein, we review recent research data on how MSC-sEVs could be used to treat kidney disease.
Keywords: small extracellular vesicles, mesenchymal stem cells, acute kidney injury, chronic kidney disease, renal cell carcinoma
Introduction
Globally, kidney disease affects approximately 10% of the world’s population, which equates approximately to more than 800 million people. Alarmingly, disease morbidity and mortality rates are increasing annually.1 Kidney diseases include acute kidney injury (AKI), chronic kidney disease (CKD), kidney stones, and renal cell carcinoma (RCC). When compared with the liver and other organs, the kidneys are less capable of self-repair, and currently the most common treatment methods for kidney disease are conservative in nature so as to slow down rather than reverse disease progression.
Mesenchymal Stem Cells (MSCs) are derived from the mesoderm and are a class of pluripotent stem cells widely used in clinical practice, with the potential for multidirectional cell differentiation and self-replenishment.2 MSCs are isolated from a variety of tissues, such as umbilical cords, endometrial polyps, muscle, dental pulp, bone marrow, and adipose tissue,3 and under certain conditions, they differentiate into different mesenchymal spectrum cell types, such as osteoblasts, adipocytes, and chondrocytes.4 MSCs localize to injury sites, secrete anti-inflammatory and growth factors via autocrine, paracrine, and endocrine mechanisms to accelerate wound healing and mediate damaged myocardium repair.5 They also suppressed lung inflammation and fibrosis in COVID-19 patients by secreting cytokines.6 Additionally, in the last decade, MSCs were instrumental in treating some musculoskeletal, neurodegenerative, and metabolic disorders.7 However, as some stem cell therapies are disadvantageous in terms of immune rejection, decreased proliferation and differentiation potential after transmission, and mutations in stem cell genetic material,8 researchers focused on MSC-small extracellular vesicles (MSC-sEVs) for the following reasons: they have the same functions as MSCs, they are more stable,9 and they do not induce serious immune rejection reactions. sEVs are lipid bilayer vesicles, they effectively protect bioactive cargoes from external degradation, they do not generate ethical issues, and they have significant potential for renal disease treatment. In recent years, these qualities have been recognized, therefore in this review, we summarized recent MSC-sEVs research advances in kidney disease treatments.
sEVs
As early as 1971, sEVs microscopic ultrastructures in Ochromonas Danica were discovered by Aaronson et al.10 Almost all human cells secrete sEVs; they are found in bodily fluids such as blood, urine, ascites, and amniotic fluid, and differ in structure and function depending on the tissue of origin.11 sEVs are composed of lipid bilayers that contain active molecules, including nucleic acids, lipids, proteins, and metabolites, which are related to different cell activities. sEVs act as carriers to transport these active molecules between cells,12 to cross natural barriers such as the blood-brain barrier,13 and regulate cell physiological functions while being relevant to numerous diseases.14
Structure and Biogenesis
sEVs ensure cell-to-cell or cell-to-extracellular matrix (ECM) connections by transporting proteins, nucleic acids, and lipids.15 Appearing like typical cup-like structures under transmission electron microscopy, sEVs measure 30 nm–100 nm in diameter, have a density of 1.10 g/mL–1.21 g/mL, and consist of lipid bilayers wrapped around an internal bioactive cargoes.16 The sEVs surface is rich in lipids (cholesterol, sphingomyelin, and ceramide), fusion-related proteins (annexin I, annexin II, and annexin IV), transmembrane 4 superfamily (CD9, CD63, and CD81), heat shock proteins (HSP) (HSP60, HSP70, HSP90, HSPA5, and CCT2), and some cell-specific proteins.17
Precise sEVs biogenesis mechanisms are unclear. Thus far, it is hypothesized that endosomal sorting complex required for transport (ESCRT)-dependent and ESCRT-independent pathways are required for sEVs formation, of which the ESCRT-dependent pathway is a classical pathway.18 ESCRT consists of approximately 20 proteins that bind to auxilin (Vps4, VTA1, and ALIX) to form ESCRT-0 (Hrs), ESCRT-I (TSG101, Vps28, and Vps37), ESCRT-II (Vps22, Vps36, Vps25, and ESCRT-III (Alix and Vps2) complexes.19 ESCRT-0 recognizes ubiquitinated proteins in endosomal membranes and interacts with clathrin to enrich their numbers; ESCRT-I and ESCRT-II act together to induce endosomal membrane loading specificity to bud inward; and ESCRT-III shears the bud neck to induce vesicle separation.20 The ESCRT-dependent sEVs formation pathway consists of two steps; firstly, cells undergo endocytosis and endocytosed cell membranes form multiple vesicles which fuse to form early endosomes (EEs).21 EEs gradually acidify and mature into late endosomes (LEs) which contain intraluminal vesicles (ILVs), while LEs further form multivesicular bodies (MVBs) rich in ILVs.16 Then, MVBs are divided into two types according to cholesterol content: MVBs with low cholesterol content are degraded through the lysosomal pathway22 while high cholesterol content MVBs are transported to and fuse with the cell membrane and release ILVs into the ECM. These ILVs are sEVs.
Biological Functions
By transporting proteins, nucleic acids, and lipids between cells, sEVs are key to most of the life activities, such as maintaining internal environmental homeostasis, immune regulation, angiogenesis, inflammatory responses, and apoptosis.16 Thus far, sEVs studies have identified two ways in which these entities elicit biological effects:23 firstly, sEVs directly activate target cell receptors via protein molecules or lipid ligands on the sEVs surface, which activates or inhibits some signaling pathways in target cells. Secondly, sEVs fuse directly with recipient cell plasma membranes or enter cells via endocytosis to release their cargoes, thereby regulating cell function and biological behaviors. By studying sEVs cargoes, we can identify disease development mechanisms and intervene with disease progression. With some diseases and associated development steps, cargo expression in sEVs is altered to some degree.24 As sEVs research intensifies, growing evidence now suggests that sEVs could function as potential drug carriers and diagnostic markers for several diseases.25
sEVs Isolation
To use sEVs in clinical settings, efficient, rapid, and stable extraction methods which maintain sEVs structure, biological function, and integrity are required. Currently, the most commonly used method is ultracentrifugation, which is simple and does not require complex technical support. However, the method is time-consuming, sEVs yields are low, and sEVs quantity and quality metrics are largely influenced by the type of rotor and the angle of settling of the rotor. The main issue is that precipitates or other vesicles, proteins, or protein/RNA aggregates cannot be easily distinguished from sEVs. Other methods include, sucrose density gradient centrifugation, size-exclusion chromatography, ultrafiltration, microfluidics, polymer precipitation, and commercial kits.26 However, to date, no standardized method has been developed to isolate sEVs, while sEV purity and yield levels from single isolation methods remain largely insufficient to meet downstream experimental requirements (Table 1). To ensure standardized extracellular vesicles (EVs) extraction techniques across clinical research, the International Society for Extracellular Vesicles (ISEV) recommended combined approaches to generate high sEVs purity and yields.27 Additionally, other extraction factors include but are not limited to; sample sources, EVs subpopulations, and their use in downstream applications. When considering sEVs purity, the ratio of quantitative methods can improve the exosome purity determinations and indicate exosome purity, eg, protein/lipid, protein/particle, and RNA/particle ratios.27 Although numerous sEVs extraction techniques are available and tend to be standardized and standardized, the establishment of sEVs collection and separation standards, as well as the standardization of product preparation processes, are still issues that need further clarification and resolution.
Table 1.
Small Extracellular Vesicles (sEVs) Isolation Techniques: Advantages and Disadvantages
| Isolation Technique | Advantages | Disadvantages |
|---|---|---|
| Ultracentrifugation | High recovery rates, simplified operation, no chemical contamination. | Costly instrumentation and time-consuming. Low RNA yields, low purity, and may be mixed with other components. |
| Sucrose density gradient centrifugation | High sEVs purity. | Preparation work is tedious and time-consuming, sEVs levels are low and biological function easily lost. |
| Size-exclusion chromatography | High recovery rates, high purity, less time consuming, high sensitivity, intact structures and biological function preserved. No chemical contamination. It is currently a better method to extract sEVs. | Expensive instrumentation. |
| Ultrafiltration | High efficiency, free from chemical contamination, and low extraction costs. | Low elution efficiency, difficulty in removing the effects of proteins, purity, shape, and charge of the sample affecting the separation. Clogging of filter membranes and loss of biological function. |
| Polymer precipitation | Simple operation, high output, high integrity, no special equipment required, economical. | Cumbersome preparation work, low purity, and susceptibility to chemical contamination. |
| Microfluidics | Easy to operate, high throughput, automated, less sample volume required for highly pure extracted sEVs. | Costly, low yields, and prone to hole plugging. |
| Commercial kits | Easy to operate and short times required. | The kits are expensive, without uniform standards, and are not suitable for the extraction of massive samples. |
| Enzyme-linked immunosorbent assay | High purity, intact isolated sEVs, suitable for the separation of specific sEVs. High specificity. | Antibodies and equipment are costly, low throughput, low isolation efficiency, suitable for cell-free samples only. |
| Hydrostatic filtration dialysis | Low costs, no chemical pollution, sEVs can be isolated from highly diluted solutions, high throughput, high integrity, and biological functions preserved. | Large sample volumes show decreased efficiency |
sEVs Characterization
According to ISEV standards, when Western Blotting (WB) is used to characterize/identify extracted EVs proteins, the evaluation of transmembrane/lipid-bound proteins (CD63, CD81, CD82, HLA, and integrin), cytoplasmic proteins (TSG101/ALIX), and at least one negative protein marker is required to characterize extracted EVs and their purity.27 For individual sEVs populations, they must be analyzed by at least two methods, such as transmission electron microscopy (TEM) for sEVs morphology and Nanoparticle Tracking Analysis (NTA) to confirm sEVs size.
sEV Storage
sEVs do not store well for long periods; after 2 days at 37°C, sizes decreases by about 60%. Currently, the most common sEVs storage method is freezing at −80°C in phosphate buffer (PBS), in addition to freeze-drying and spray-drying methods.28
However, the −80°C storage method in PBS is not optimal as evidenced by Corso et al.29 The authors evaluated different EVs preservation strategies for up to 2 years, and found that over time, EVs in PBS had much lower recovery rates and that EVs degradation in PBS had already started within minutes of storage. The recommended buffer system, among several tested, was PBS plus human albumin and trehalose (PBS-HAT), which significantly improved short- and long-term EVs sample preservation at −80°C, maintained stability, and significantly improved EVs recovery rates during subsequent EVs studies. Additionally, another study recently analyzed the effects of different preservation strategies and times on EVs. Experiments had three components; firstly, they analyzed the purity and particle size of EVs preserved by different storage methods.; secondly, authors analyzed the effects of both fast and slow freeze-thaw cycles and the number of freeze-thaw cycles on EVs; and thirdly, authors explored EVs membrane fusion after freeze-thawing. This latter study component investigated membrane fusion phenomena after freeze-thawing. Study data were interesting; in the first study, storage at −80°C decreased EVs concentrations over time regardless of the storage state, EVs loading decreased significantly after 6 months, and EVs particle size increased over time at −80°C, which was negatively correlated with particle concentration. The second study showed that both fast and slow freeze-thawing cycles increased EVs particle sizes and decreased particle numbers, and that repeated freeze-thawing cycles increased EVs particle sizes. The third study demonstrated that freeze-thawing processes ruptured EVs films and the subsequent re-micellization of ruptured films into new particles.30 In summary, EVs extracted from fresh samples are optimal for use, but if long-term storage is required then repeated sample or EVs freeze-thawing must be avoided, and preferably PBS-HAT should be used for storage.
MSC-sEVs
Under TEM, like most other cell-secreted sEVs, MSC-sEVs show a classical cup-like morphology (Figure 1), but they not only express intrinsically expressed sEVs protein markers (CD9, CD63, CD81, TSG101, and ALIX), but also adhesive molecules on the MSC surface (CD90, CD29, CD44, CD73).31,32 Additionally, MSC-sEVs are rich in cholesterol and sphingomyelin; their fatty acids are predominantly arachidonic acid, docosahexaenoic acid, leukotrienes, phosphatidic acid, and prostaglandin lysophosphatidylcholine.4 MSC-sEVs also contain high levels of RNA molecules and low mitochondrial DNA levels,33 and it is these cargoes that allow MSC-sEVs exert their biological effects (Figure 2).
Figure 1.
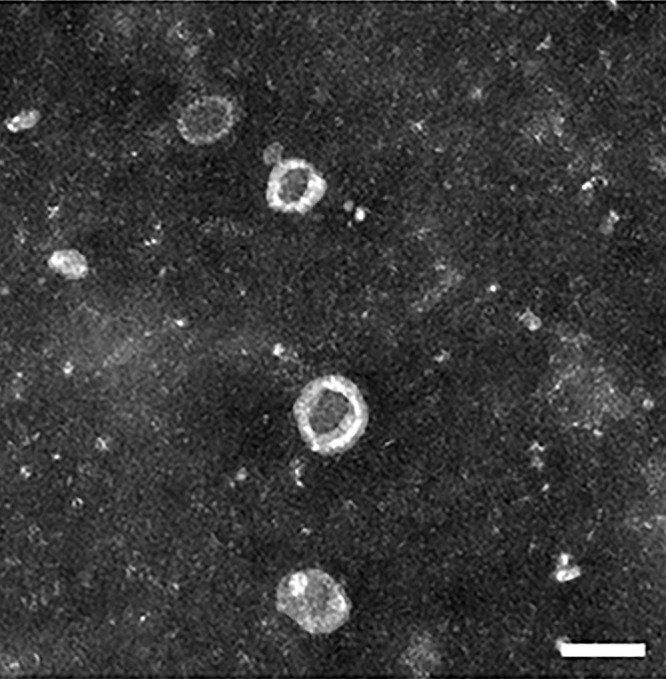
Transmission electron microscope (TEM) hucMSC-sEVs image.
Notes: HucMSC-sEVs show a typical cup-like morphology under TEM (scale bar = 100 nm). Reproduced with permission from He Q, Wang L, Zhao R et al. Mesenchymal stem cell-derived exosomes exert ameliorative effects in type 2 diabetes by improving hepatic glucose and lipid metabolism via enhancing autophagy. Stem Cell Res Ther. 2020;11(1):223.110 Copyright 2020 The Authors. Creative Commons Attribution License.
Abbreviations: hucMSC-sEVs, human umbilical cord Mesenchymal Stem Cell-derived small extracellular vesicles; TEM, transmission electron microscopy.
Figure 2.

MSC-sEVs cargoes functions in renal disease.
Abbreviations: MSC-sEVs, Mesenchymal Stem Cell-derived small extracellular vesicles; sEVs, Small extracellular vesicles; CCR2, C-C motif chemokine receptor-2; let7c, microRNA-let7c; GDNF-AMSC-sEVs, Glial cell line-derived neurotrophic factor to transfect human adipose mesenchymal stem cells and subsequently isolated sEVs.
As indicated, MSC-sEVs are safer, more stable, and easier to store than MSCs, and provide a unique cell-free therapeutic approach of great potential. Encouragingly, initial therapeutic MSC-sEVs studies generated promising results for different diseases.34,35 In the COVID-19 epidemic, MSC-sEVs reduced lymphocytes and decreased inflammatory molecules and neutrophils in COVID-19 patients, thereby reducing cytokine storms in individuals with severe infections.36 MSC-sEVs not only downregulated immune cells to reduce inflammatory responses, but also rescued damaged immune cells in some diseases. Myelin debris can directly inhibit axonal regeneration and impede neurological repair. After spinal cord injury, macrophage phagocytosis becomes impaired, and local myelin sheaths cannot be cleared, resulting in deteriorating motor functions after injury.37 Sheng et al38 reported that sEVs derived from bone marrow (BMSC-sEVs) reached injury sites and were phagocytosed by local macrophages, upregulates the expression of Macrophagereceptor with collagen structure, and restored normal physiological functions to macrophages, thereby removing myelin debris from damaged areas and improving neurological functions.
Thus far, MSC-sEVs have generated impressive results in wound healing and skin regeneration,39 tumor progression,40 nerve injury repair,41 and COVID-19.42 We next focused on recent MSC-sEVs research in kidney disease.
MSC-sEVs in Kidney Disease
Acute Kidney Injury
AKI etiology is related to ischemia, hypoxia, and reactive oxygen species. Also, because the kidneys require high oxygen consumption for water and electrolyte reabsorption, renal tubules are susceptible to oxidative stress injury.43 Without intervention, AKI predisposes patients to an increased risk of progressive CKD. MSC-sEVs protect the kidneys in several conditions, including ischemia-reperfusion injury (IRI), drug-induced nephropathy (DIN), and sepsis-induced AKI. The impact of MSC-sEVs on these conditions is discussed.
Ischemia-Reperfusion Injury
This condition is one of the leading causes of AKI, with approximately 50% of AKI hospitalizations due to IRI.44 IRI-induced AKI pathophysiology includes inflammation, oxidative stress, renin-angiotensin activation, and mitochondrial dysfunction, while blood reperfusion based on tissue and organ ischemia may lead to IRI.45
Early studies reported that adipose-derived MSC (ADMSC) minimized renal injury during renal ischemia-reperfusion injury (RIRI) by reducing oxidative stress and suppressing inflammatory responses.46 Several studies showed that MSC-sEVs had similar effects in alleviating AKI after IRI, eg, Li et al47 showed that MSC-sEVs minimized IRI-induced renal structural damage in rats by downregulating inflammatory factors and inhibiting nuclear factor kappa-B (NF-κB) signaling. Zhang et al48 demonstrated that human umbilical cord MSC-derived sEVs (hucMSC-sEVs) reduced renal pro-inflammatory factors like interleukin-1β(IL-1β), IL-6, tumor necrosis factor-α (TNF-α), and monocyte chemoattractant protein-1 (MCP-1) in IRI and reduced oxidative stress to mitigate renal injury. Huang et al49 established a porcine RIRI model and found that hucMSC-sEVs were beneficial for RIRI as they inhibited inflammatory factor release (Figure 3). Furthermore, by simultaneously administering ADMSC and ADMSC-derived sEVs (ADMSC-sEVs) in renal IRI rats, Lin et al50 showed that ADMSC and ADMSC-sEVs simultaneous-treatment better protected rat kidneys from acute IRI when compared with animals treated with ADMSC alone. Zhu et al51 used PKH26-labeled sEVs from human-bone-marrow-derived MSCs (hBMSC-sEVs), injected them into the tail vein of an RIRI mouse model, and found that hBMSC-sEVs accumulate well in the kidney and effectively protected the kidney. Additionally, miR-199a-3p was highly expressed in hBMSC-sEVs and activated phosphatidylinositol 3 kinase (PI3K) /protein kinase B(AKT) and extracellular receptor kinase (ERK) pathways by inhibiting downstream Sema3A expression, thus exerting protective effects toward the kidneys. Several studies reported that MSC-sEVs combinations with other therapies provided better protection for the kidneys. Melatonin (MT) is a hormone secreted by the pineal gland of the brain and improves sleep quality.52 However, because it has strong antioxidant and anti-inflammatory effects, MT has important protective roles in tissues undergoing IRI.53 An early 2008 study showed that MSC pretreatment with MT improved MSC survival in vivo,54 which exerted better renoprotective effect on MSCs. Alzahrani et al55 compared the renoprotective effects of MT-pretreated MSC-sEVs, MSC alone, and MSC-sEVs without MT pretreatment in kidneys. MT-pretreated MSC-sEVs maximized the protective effects on kidneys by enhancing antioxidant status, inhibiting renal cell apoptosis, suppressing inflammation, and inducing angiogenesis and renal tissue regeneration in IRI tissue. Similar studies reported that combined MT treatments with MSC and MSC-sEVs also minimized RIRI in rats.56 We previously indicated that sEVs mainly exerted their functions based on their cargoes (especially RNA). Due to sEVs heterogeneity, different sEVs have different cargoes,57,58 thus, MSC-sEVs can contain protective RNAs (eg, miR-199a-3p51) which protect the kidneys by reducing oxidative stress and inhibiting inflammation. Also, MSC pretreatment with MT not only induced MSC proliferation59 but also upregulated RNAs with renoprotective effects in MSC-sEVs.55 Shen et al60 identified increased CC motif chemokine receptor-2 (CCR2) levels in MSC-sEVs and demonstrated its ability to act as an endogenous CCL2 sponge to deplete ligands, thereby alleviating CCL2-induced macrophage infiltration in mice with IRI-induced AKI. Finally, a recent study reported that 3-dimensional cultured human placental stem cell EVs (hPMSC-EVs) more effectively inhibited apoptosis and inflammatory damage in RIRI when compared with 2-dimensional cultures.61 Collino et al62 identified an interesting phenomenon where ADMSCs secreted more EVs under hypoxic conditions, and these ADMSC-EVs were more protective of the kidneys under RIRI when compared with ADMSC-EVs secreted under normoxic conditions. Thus, MSC 3-dimension-culturing and culturing under hypoxic conditions may facilitate MSC-sEVs protective effects.
Figure 3.

Therapeutic efficacy of MSC-Exo in acute kidney injury.
Notes: Sham+Veh: Sham group treated with vehicle; Sham+Exo: Sham group treated with MSC-Exo; I/R+Veh: Ischemia-reperfusion model treated with vehicle; I/R+Exo: Ischemia-reperfusion model treated with MSC-Exo. (A) Kidney tissue hematoxylin and eosin (HE) staining in four groups. Scale bar = 100 µm. Injured tubules with intraluminal casts are indicated by arrows. (B) NGAL immunohistochemical staining. Scale bar = 100 µm. MSC-Exo administration caused significant relief in interstitial edema, tubular dilatation, vacuolar degeneration, and necrosis. Immunohistochemistry showed a decrease in NGAL expression after treatment. Reproduced with permission from Huang J, Cao H, Cui B et al. Mesenchymal Stem Cells-Derived Exosomes Ameliorate Ischemia/Reperfusion Induced Acute Kidney Injury in a Porcine Model. Front Cell Dev Biol. 2022;10:899869.49 Copyright © 2022 The Authors. Creative Commons Attribution License.
Abbreviations: MSC-Exo, Mesenchymal Stem Cell-derived exosome; NGAL, Neutrophil gelatinase-associated lipocalin.
Drug-Induced Nephropathy
In the body, the kidneys are the main drug metabolism and excretion organs; however, some drugs can seriously damage these organs. DIN is a common cause of AKI, and accounts for up to 60% of community- and hospital-acquired episodes.63
Cisplatin (CDDP) is a clinically effective broad-spectrum anticancer drug, and is regulated during renal transport by proximal tubular transport proteins such as OCT2 and MATE1, therefore, CDDP accumulates in proximal renal tubular cells, causes inflammation, injury, and cell death, and leads to nephrotoxicity in 30–40% of CDDP-treated patients.64 Although studies have highlighted the molecular mechanisms underpinning CDDP-induced AKI (CDDP-AKI),65 the pathway whereby CDDP regulates cell survival, metabolism, and immune responses is also involved in CDDP cytotoxicity in tumor cells, thus, intervening in this process may attenuate the antitumor effects of CDDP. Therefore, we anticipate that MSC-sEVs better protect renal cells without reducing drug efficacy. Wang et al66 showed that hucMS-sEVs reduced TNF-α, IL1-β, and IL6 inflammatory cytokine serum levels in a CDDP-AKI mouse model, thereby suppressing renal injury; similarly, hucMSc-sEVs inhibited CDDP-induced mitochondrial apoptosis and inflammatory cytokine secretion in in vitro renal tubular epithelial cells (TECs), with these effects effectively alleviating CDDP nephrotoxicity by inhibiting mechanistic target of rapamycin(mTOR) phosphorylation and activating autophagy in TECs. Additionally, the 14-3-3ζ protein from hucMSc-sEVs likewise induced autophagy in the CDDP-AKI model. Jia et al67 found that 14-3-3ζ from hucMSC-sEVs activated autophagy by binding to ATG16L and promoted its localization to the outer surface of phagocytes, thereby protecting the kidney in CDDP-AKI. Zhou et al68 used in vivo and in vitro approaches to show that hucMSC-sEVs ameliorated CDDP-induced AKI through antioxidant and anti-apoptotic effects and promoted cell proliferation, but did not explain its specific mechanisms. Zhang et al69 pretreated MSCs with the miR-1184 agomir to generate high miR-1184 expression in pretreated MSC-sEVs, with cells significantly upregulating CDDP-treated HK-2 cell viability, and MSC-sEVs-miR-1184 protective in CDDP-induced AKI via Forkhead Box O4.
Bruno et al conducted several studies using EVs derived from BMSC(BMSC-EVs) in a glycerol-induced AKI mouse model. Their early studies demonstrated that the intravenous administration of human BMSC-EVs had the same efficacy as MSC in restoring renal function and morphology in mice, and that mRNA molecules in BMSC-EVs promoted AKI recovery by activating anti-apoptotic and pro-regenerative programs in TECs.70 In another study, the authors demonstrated that miRNAs in BMSC-EVs had key roles in recovering glycerol-induced AKI.71 Interestingly, their recent study reported that in the same model, BMSC-EVs induced renal tubular cell proliferation and attenuated kidney injury (among other functions), which was mainly attributed to mRNA, miRNA, and growth factor cargoes in sEVs.72 Thus, the anti-apoptotic, regeneration-promoting, and other kidney-protective functions exerted by BMSC-EVs may depend on nucleic acids, proteins, and other cargoes in BMSC-sEVs, but more studies are required to identify which molecules act in which pathways.
In a rat gentamicin-induced AKI model, Reis et al73 observed that BMSC-sEVs had the same renoprotective role as BMSCs, and that RNAs in BMSC-sEVs mediated AKI recovery via a range of functions, including inhibiting apoptosis, promoting cell proliferation, downregulating pro-inflammatory cytokines, and upregulating anti-inflammatory cytokines. Unfortunately, no studies have identified which RNAs exerted roles in gentamicin-induced AKI, thus mechanisms require further investigation.
Sepsis-Induced Acute Kidney Injury
Gao et al74 used ADMSC-sEVs tail vein injections to treat a sepsis-AKI mouse model and found that ADMSC-sEVs improved renal function, reduced inflammatory cytokine release, and decreased mortality by activating Sirtuin 1 (SIRT1). hucMSC-sEVs reduced IRAK-1 target gene expression, which codes for a key protein that positively regulates NF-κB activity, by upregulating miR-146b levels in renal tissue (but not transporting miR-146b), which inhibited NF-κB activity and reduced pro-inflammatory factor levels during sepsis, thus effectively protecting the kidneys from sepsis-induced AKI.75
Chronic Kidney Disease
CKD refers to abnormalities in kidney structure and function, is caused by different mechanisms lasting 3 months or longer, and includes many types of kidney disease. Without timely and effective intervention, CKD leads to deterioration and disease progression, or during the disease course, it develops into chronic renal insufficiency, renal failure, and eventually end-stage renal disease (ESRD).76 One of the most common causes of CKD is diabetic nephropathy (DN); MSC-sEVs have been extensively studied in CKD, especially in DN and renal fibrosis.
Diabetic Nephropathy
DN is a common microvascular complication of diabetes mellitus and a major cause of ESRD. Disease pathogenesis includes abnormal glucose metabolism, renal hyperperfusion hyperfiltration, oxidative stress, immune inflammation, and genetic factors. Currently, the clinical treatment is to slow down DN progression by administering drugs that improve blood pressure or blood glucose levels. Once DN progresses to irreversible stages, alternative treatments such as hemodialysis are inevitable77. Li et al78 reported that mouse umbilical cord MSCs (MUC-MSCs) blocked transforming growth factor-β(TGF-β)-triggered myofibroblast transdifferentiation through paracrine sEVs, inhibited PI3K/Akt and mitogen-activated protein kinase (MAPK) signaling-mediated mesangial cell proliferation, increased matrix metalloproteinase levels in mesangial cells, and reduced renal fibrosis in a DN mouse model. Additionally, hucMSC-sEVs inhibited TGF-β, IL-6, IL-1β, and TNF-α inflammatory cytokine production in HK2 cells when induced by high glucose, thus effectively improving renal function, inhibiting inflammation and fibrosis, and preventing early DN progression.79 In their DN rat model, Hao et al showed that ADMSC-sEVs significantly reduced blood glucose, serum creatinine, 24-h urine protein, urine albumin creatinine ratio, and kidney weight, and inhibited renal fibrosis and thylakoid hyperplasia when compared with control animals. Moreover, ADMSC-sEVs downregulated IL-6 expression in in vitro DN models, while IL-6 promoted mesangial proliferation. The authors also observed that processes were putatively mediated by miR-125a in ADMSC-sEVs, which bound histone deacetylase 1 (HDAC1) and down-regulated endothelin-1 expression, thereby reducing DN symptoms and kidney injury.80 Nagaishi et al81 showed that MSC-sEVs inhibited tubular interstitial fibrosis and glomerular mesangial matrix expansion by reducing p38-MAPK and TGF-β expression, preventing Z0-1 degradation in TECs, and reducing urinary albumin excretion in DN mice. Jin et al82 indicated that miR-486 in ADMSC-sEVs could be transferred to podocytes, as autophagy activators, to ameliorate podocyte injury by regulating miR-486/Smad1/mTOR signaling to alleviate high glucose-induced cell injury in MPC5 cells and DN mice.
Renal Fibrosis
Renal fibrosis is an irreversible pathological change in end stage chronic kidney disease; its pathogenesis is very complex and the specific mechanisms remain unclear. The main pathological causes may be due to ECM deposition, TEC epithelial-mesenchymal transition (EMT),83 and vascular sclerosis. In previous research, MSC-sEVs transported miRNA-let7c to damaged TECs and inhibited collagen Iα1, IVα1, α-smooth muscle actin, and TGF-β1 expression by inducing increased let7c expression. Ultimately, this process attenuated renal fibrosis and renal injury.84 Yang et al85 showed that miR-186-5p from MSC-sEVs suppressed ECM deposition and EMT by downregulating Smad5, and inhibiting renal fibrosis progression both in vitro and in vivo. BMSC-sEVs reduced M1 and M2 macrophage polarization by activating EP2 receptors, thereby inhibiting pro-inflammatory factor (IL-6) secretion, promoting anti-inflammatory factor (IL-10) release, and ultimately reducing ECM deposits and renal fibrosis (Figure 4).86 Additionally, combined BMSC-sEVs and serelaxin (antifibrotic drug) effectively treated mice with renal fibrosis while reducing hypertension, renal macrophage levels, T-cell infiltration, and TEC injury.87 BMSC-sEVs also inhibited TGF-β1-induced EMT in HK-2 cells by transporting Nedd4L, which also improved in vivo renal fibrosis.88 Jin et al89 used anti-let-7i-5p to transfect MSCs and observed that MSC-sEVs delivered anti-let-7i-5p to target cells in vitro and attenuated TGF-β1-induced fibrotic responses; in vivo, anti-let-7i-5p exerted anti-fibrotic effects in a renal fibrosis model by activating TSC1/mTOR signaling. Chen et al90 used glial cell line-derived neurotrophic factor (GDNF) to transfect human adipose MSCs and subsequently isolated sEVs (GDNF-AMSC-sEVs), which inhibited apoptosis and stimulated angiogenesis by activating SIRT1 and upregulating phosphorylated endothelial nitric oxide synthase levels, thereby improving renal fibrosis. MSC-sEV pretreatments with different drugs effectively inhibited renal fibrosis and could be used as MSC-sEV applications in the clinic.
Figure 4.

Therapeutic efficacy of BMSC-exo in a renal fibrosis model.
Notes: (A) HE, Masson, Sirius red picric acid staining and α-SMA immunohistochemical staining were used to detect renal histopathological changes in the different treatment groups (X400). (B) Compared with the control group, the UUO model group showed dilated renal tubules, infiltration of renal interstitial inflammatory cells and increased extracellular matrix deposition. Compared with the UUO group, the BMSC-exo treatment group significantly reduced interstitial matrix deposition and significantly decreased α-SMA expression, with effects similar to those of the EP2 agonist butaprost. *p < 0.05, **p < 0.01. Reproduced with permission from Lu Y, Yang L, Chen X, Liu J, Nie A, Chen X. Bone marrow mesenchymal stem cell-derived exosomes improve renal fibrosis by reducing the polarisation of M1 and M2 macrophages through the activation of EP2 receptors. IET Nanobiotechnol. 2022;16(1):14–24.86 Copyright 2020 The Authors. Creative Commons Attribution License.
Abbreviations: BMSC-exo, exosome derived from bone marrow mesenchymal stem cells; α-SMA, α-smooth muscle actin; UUO, unilateral ureteral occlusion.
Metformin is the drug of choice for treating type 2 diabetes. In addition to anti-hyperglycemic effects, the drug exerts impacts inflammation, autophagy, oxidative damage, apoptosis, and aging, and has protective functions toward the kidneys. After treating CKD MSCs with metformin, Kim et al91 reported that the paracrine effects of pretreated MSCs were enhanced and inhibited MSC senescence by suppressing SA-β-gal activity, p16 Ink4a expression, and p53 and NF-κB activation, thus effectively reducing CKD inflammation and fibrosis. Furthermore, Liao et al92 observed that MSCs treated with metformin enhanced EVs production via autophagy-related pathways and showed that elevated ITIH4 levels in EVs, which are critical for regulating cell growth and inflammatory responses, improved disc cell senescence. Although MSC-sEV pretreatment studies with metformin in CKD remain underreported, we hypothesize potential feasibility for treating CKD.
Rencal Cell Carcinoma
RCC is one of the most common urological malignancies,93 with annual RCC cases at approximately 400,000, and deaths at approximately 175,000. Incidence rates are increasing year on year, with higher rates in men than women.94 The most common disease subtype is clear cell renal cell carcinomas (ccRCC),95 which accounts for 70–80% of all RCC cases.96 To date, several sEVs-related studies have provided new concepts for RCC treatment.25,97 Li et al98 used hucMSC-sEVs to treat ccRCC mice via intravenous administration and found not only enhanced in vivo immune responses, but also inhibited in vivo ccRCC growth and metastasis via vascular endothelial growth factor (VEGFA) targeting by miR-182 in hucMSC-sEVs. Another study showed that human liver stem cell-derived EVs upregulated miRNAs with antitumor effects, including miR-145, miR-200b and miR-200c, which slowed down tumor progression in mice by promoting apoptosis and reducing tumor angiogenesis.99 These studies identified MSC-sEVs superiority in treating RCC; however, MSC-sEVs are a double-edged sword in cancer. Qi et al100 reported that BMSC-sEVs promoted osteosarcoma and gastric cancer cell proliferation by activating Hedgehog signaling, while Zhang et al101 observed that BMSC-sEVs carrying miR-193a-3p, miR-210-3p, and miR-5100 to recipient cells, activated STAT3 signaling to induce EMT and enhance lung cancer cell invasion. Therefore, the main issues are how to safely and stably exploit MSC-sEVs for RCC treatment. As the research on MSC-sEVs progresses, it is believed that the application of MSCs-sEVs in the treatment of RCC can achieve more satisfactory results and be more widely used in the clinic in the future.
Kidney Transplantation
Kidney transplantation is the ideal treatment for ESRD. After more than half a century of development, kidney transplantation has become the most common organ transplantation procedure with high success rates, and has significantly reduced ESRD mortality rates to approximately 29 deaths out of 1000 patients/year.102 However, some patients may suffer from transplanted kidney failure or progressive renal insufficiency after renal transplantation, which may lead to death, therefore, prognostics for renal transplantation are critical in clinical research. Critically, Wu et al103 demonstrated that the long non-coding RNA DANCR in BMSC-sEVs promoted Treg cell differentiation by reducing SIRT1 levels in CD4+ T cells, which induced immune tolerance after kidney transplantation.
MSC-sEVs Imaging Studies in Kidney Disease
Despite the potential shown by MSC-sEVs in regenerative medicine, their non-invasive in vivo tracking is highly complex due to variations/changes induced by interactions with recipient cells.104 Tracking in vivo sEVs distribution, migratory capacity, toxicity, biological effects, communication capacity, and mechanisms may provide a theoretical basis for assessing sEVs levels for treating organs and predicting therapeutic responses.105
The first labeling quandary is whether fluorescent labeling should be performed directly or indirectly on MSCs to obtain fluorescently labeled MSC-sEVs. This question was addressed by Grange et al106 who used the lipophilic fluorescent dye DiD to label MSC-EV to obtain direct EV labeling (DL-EV) and labeled EVs by donor cells pre-treated with the dye (LCD-EV). The authors then administered labeled MSC-EV groups to a glycerol-induced AKI mouse model to assess fluorescence signals using optical imaging. Both labeling methods detected EVs in whole-body images and damaged kidneys, while DL-EV showed higher and brighter fluorescence when compared with LCD-EV whose signals were detectable in dissected kidneys only. These data suggested that LCD-EV exerted higher specificity but lower fluorescence intensity than DL-EVs, therefore, direct EVs labeling appeared to be the better option.
Gupta et al107 used luciferase NanoLuc and ThermoLuc systems to label MSC-EVs with CD63, and injected cells into mice to determine in vivo MSC-EVs pharmacokinetics. ThermoLuc-labeled MSC-EVs were more suitable for non-invasive EVs follow-up, while NanoLuc was only suitable for in vivo quantification under ex vivo conditions due to poorer substrate distributions, emission wavelength, and toxicity issues, even though it possesses a broader dynamic range. However, fluorescent molecule signals suffer from tissue scattering, poor penetration depth, and low resolution, which can be better facilitated by nuclear and magnetic resonance imaging (MRI)-based methods.
Han et al108 performed the first MRI tracking study of systemically administered EVs, and improved on previous studies by using electroporation to directly label EVs with superparamagnetic iron oxide. The authors generated magneto-EVs of high purity and sufficient MRI sensitivity by purifying labeled EVs using Ni-NTA columns. Using this method, authors labeled pluripotent stem cell-derived EVs (iPSC-EVs) and injected intravenously them into a mouse AKI model. Unequivocally, iPSC-EVs treatment significantly improved kidney function and increased animal survival. Notably, using MRI signals from labeled EVs, authors observed that iPSC-EVs targeted and rapidly accumulated in damaged kidneys. When compared with previous imaging methods, this method was highly sensitive and allowed for real-time MRI ER tracking in a non-invasive manner, without compromising MSC-EVs therapeutic efficacy. Although MRI facilitated excellent tissue penetration and quantification, it is expensive and time-consuming, and may be difficult to scale-up in future studies, therefore more options are required.
Cao et al109 labeled isolated MSC-EVs by incubating cells with DPA-SCP and injecting preparations into a RIRI mouse model. DPA-SCP exhibited deeper tissue penetration, lower biological background interference, and higher in vivo biosafety profiles than the conventional dye pkh26, and imaging MSC-EVs by aggregation-induced emission luminogens did not affect the function of EVs membranes and damaged kidneys. Using this method, the authors found that MSC-EVs were rapidly absorbed by damaged kidneys in RIRI mice to reach TECs and exert nephroprotective effects. Protective effect mechanisms suggested that MSC-EVs transported miRNA-200a-3p to TECs, targeted Keap1-Nrf2 signaling, and targeted and protected TEC mitochondrial functions by reducing mitochondrial fragmentation and normalizing membrane potential in AKI.
These studies demonstrated the following commonalities; firstly, MSC-EVs were rapidly absorbed after entering the body and the highest absorption occurred in the liver; secondly, MSC-EVs had excellent abilities to target damaged kidneys. The above describes some current applications of Imaging study in MSC-EVs for renal diseases. In conclusion, there is an urgent need to develop suitable non-invasive in vivo imaging methods to track MSC-sEVs secretion, transport, and uptake in clinical regenerative medicine (Figure 5).
Figure 5.

Imaging MSC-sEVs in kidney disease – a flow chart.
Notes: Labeling methods include chemical dye labeling, aptamer labeling, labeling extracellular vesicle surface proteins by genetic engineering, and labeling with radioisotope or magnetic resonance imaging contrast agents.104 Created with BioRender.com.
Abbreviation: sEVs, Small extracellular vesicles.
Conclusion and Future Perspectives
With recent advances in regenerative medicine, MSC-sEVs have the potential to become the next generation of cell-free therapies for kidney disease (Figure 6). EVs have great potential as natural drug delivery systems and EVs bilayer membranes protect cargoes from body clearance thereby prolonging circulating half-lives. From our review, MSC-sEVs have four main application areas. Firstly, MSC-sEVs can transport renoprotective miRNAs to target cells to alleviate AKI by reducing oxidative stress, inhibiting renal cell apoptosis, suppressing inflammation, and inducing renal tissue regeneration. Secondly, MSC-sEVs can slow down DN progression by anti-fibrosis actions, reducing EMT, inhibiting apoptosis, and reducing inflammation. Thirdly, MSC-sEVs have the potential to become anti-tumor vaccines to inhibit RCC progression by enhancing in vivo immune responses. Fourthly, MSC-sEVs appear to improve survival rates in kidney transplanted mice by inducing immune tolerance.
Figure 6.

Mesenchymal stem cells secrete MSC-sEVs to protect the kidney.
Notes: MSCs undergo endocytosis to form EEs, which mature into LEs and eventually form MVBs for transport to the cell membrane to release sEVs outside the cell. MSC-sEVs enter renal cells by endocytosis or bind to receptors, and exert protective effects on the kidney via anti-oxidative stress, anti-apoptosis, anti-inflammation, anti-fibrosis, cell proliferation, and immunomodulation actions.
Abbreviations: EEs, Early endosomes; LEs, Late endosomes; MVBs, Multivesicular bodies; RAB, Ras-associated GTP-binding; Alix, Alg-2 interacting protein x; HSP, Heat shock proteins; TSG101, Tumor susceptibility gene 101; MHC, Major histocompatibility complex; LcRNA, Long non-coding RNA; mRNA, Messenger RNAs.
We also reviewed some of the tools currently available for EVs labeling. These technological advances will facilitate a greater understanding of transport trajectories and targeting ability of MSC-sEVs in vivo, with a view to early applications for clinical medicine. Although MSC-sEVs are important for renal disease therapy, many unknowns and challenges remain. Firstly, MSC-sEVs research in kidney disease treatment is still focused on cell and animal studies, and there remains a lack of knowledge on MSC-sEVs duration effects, administered amounts, and therapeutic effects in kidney disease, thus it is unclear if proteins, lipids, mRNAs, and miRNAs in MSC-sEVs may cause other harmful effects in humans. Secondly, due to sEVs heterogeneity, different sEVs sources have different therapeutic effects on kidney disease, so it is important to screen for the most effective sEVs tissue sources. Thirdly, to ensure further research development, more sophisticated isolation and purification techniques are required to generate highly pure and stable sEVs within shorter periods.
Unequivocally, MSC-sEVs have enormous potential for treating renal disease, and its efficiency, safety, pharmacokinetics and specific mechanisms for the treatment of renal diseases will be the focus of the next studies.
Acknowledgments
We deeply appreciate the support from all participants.
Funding Statement
This research was supported by the Natural Science Foundation of Jiangxi Province (No. 20192BAB205075 and No. 20202BABL206119) and the Health Commission of Jiangxi Province (No. 20204518).
Author Contributions
Y.L. collected and analyzed the relevant literature, the writing of the first draft, figure design, and comment revision and feedback. L.W. performed analyses and organized the literature, figure design, and manuscript revision. M.Z. helped write the first draft, graph design, and data collection. Z.C. identified the topic, guided the writing of the manuscript, helped with diagramming, made grammatical corrections to the article and identified the journal for submission. All authors contributed to data analysis, drafting or revising the article, have agreed on the journal to which the article will be submitted, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
We have no conflicts of interest to disclose.
References
- 1.Kovesdy CP. Epidemiology of chronic kidney disease: an update 2022. Kidney Int Suppl. 2022;12(1):7–11. doi: 10.1016/j.kisu.2021.11.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Samsonraj RM, Raghunath M, Nurcombe V, et al. Concise review: multifaceted characterization of human mesenchymal stem cells for use in regenerative medicine. Stem Cells Transl Med. 2017;6(12):2173–2185. doi: 10.1002/sctm.17-0129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Li Y, Cheng Q, Hu G, et al. Extracellular vesicles in mesenchymal stromal cells: a novel therapeutic strategy for stroke. Exp Ther Med. 2018;15(5):4067–4079. doi: 10.3892/etm.2018.5993 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Deng H, Sun C, Sun Y, et al. Lipid, protein, and MicroRNA composition within mesenchymal stem cell-derived exosomes. Cell Reprogram. 2018;20(3):178–186. doi: 10.1089/cell.2017.0047 [DOI] [PubMed] [Google Scholar]
- 5.Sid-Otmane C, Perrault LP, Ly HQ. Mesenchymal stem cell mediates cardiac repair through autocrine, paracrine and endocrine axes. J Transl Med. 2020;18(1):336. doi: 10.1186/s12967-020-02504-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Henriques-Pons A, Beghini DG, Silva V, Iwao HS, Da SF. Pulmonary mesenchymal stem cells in mild cases of COVID-19 are dedicated to proliferation; in severe cases, they control inflammation, make cell dispersion, and tissue regeneration. Front Immunol. 2021;12:780900. doi: 10.3389/fimmu.2021.780900 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Warrier S, Mohana SS, Varier L, Balasubramanian A. Stalling SARS-CoV2 infection with stem cells: can regenerating perinatal tissue mesenchymal stem cells offer a multi-tiered therapeutic approach to COVID-19? Placenta. 2022;117:161–168. doi: 10.1016/j.placenta.2021.12.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Trohatou O, Roubelakis MG. Mesenchymal stem/stromal cells in regenerative medicine: past, present, and future. Cell Reprogram. 2017;19(4):217–224. doi: 10.1089/cell.2016.0062 [DOI] [PubMed] [Google Scholar]
- 9.Zou XY, Yu Y, Lin S, et al. Comprehensive miRNA analysis of human umbilical cord-derived mesenchymal stromal cells and extracellular vesicles. Kidney Blood Press Res. 2018;43(1):152–161. doi: 10.1159/000487369 [DOI] [PubMed] [Google Scholar]
- 10.Aaronson S, Behrens U, Orner R, Haines TH. Ultrastructure of intracellular and extracellular vesicles, membranes, and myelin figures produced by Ochromonas danica. J Ultrastruct Res. 1971;35(5):418–430. doi: 10.1016/S0022-5320(71)80003-5 [DOI] [PubMed] [Google Scholar]
- 11.Rashed HM, Bayraktar E, Helal KG, et al.Exosomes: from garbage bins to promising therapeutic targets. Int J Mol Sci. 2017;18(3):538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Denzer K, Kleijmeer MJ, Heijnen HF, Stoorvogel W, Geuze HJ. Exosome: from internal vesicle of the multivesicular body to intercellular signaling device. J Cell Sci. 2000;113(Pt 19):3365–3374. doi: 10.1242/jcs.113.19.3365 [DOI] [PubMed] [Google Scholar]
- 13.Jiang XC, Gao JQ. Exosomes as novel bio-carriers for gene and drug delivery. Int J Pharm. 2017;521(1–2):167–175. doi: 10.1016/j.ijpharm.2017.02.038 [DOI] [PubMed] [Google Scholar]
- 14.Xu R, Rai A, Chen M, et al. Extracellular vesicles in cancer - implications for future improvements in cancer care. Nat Rev Clin Oncol. 2018;15(10):617–638. doi: 10.1038/s41571-018-0036-9 [DOI] [PubMed] [Google Scholar]
- 15.Villata S, Canta M, Cauda V. EVs and bioengineering: from cellular products to engineered nanomachines. Int J Mol Sci. 2020;21(17):17. doi: 10.3390/ijms21176048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367:6478. doi: 10.1126/science.aau6977 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mathivanan S, Simpson RJ. ExoCarta: a compendium of exosomal proteins and RNA. Proteomics. 2009;9(21):4997–5000. doi: 10.1002/pmic.200900351 [DOI] [PubMed] [Google Scholar]
- 18.Mao W, Wang K, Wu Z, Xu B, Chen M. Current status of research on exosomes in general, and for the diagnosis and treatment of kidney cancer in particular. J Exp Clin Cancer Res. 2021;40(1):305. doi: 10.1186/s13046-021-02114-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Mosesso N, Nagel MK, Isono E. Ubiquitin recognition in endocytic trafficking - with or without ESCRT-0. J Cell Sci. 2019;132(16):16. doi: 10.1242/jcs.232868 [DOI] [PubMed] [Google Scholar]
- 20.McCullough J, Frost A, Sundquist WI. Structures, functions, and dynamics of ESCRT-III/Vps4 membrane remodeling and fission complexes. Annu Rev Cell Dev Biol. 2018;34(1):85–109. doi: 10.1146/annurev-cellbio-100616-060600 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Fujita Y, Kosaka N, Araya J, Kuwano K, Ochiya T. Extracellular vesicles in lung microenvironment and pathogenesis. Trends Mol Med. 2015;21(9):533–542. doi: 10.1016/j.molmed.2015.07.004 [DOI] [PubMed] [Google Scholar]
- 22.Vietri M, Radulovic M, Stenmark H. The many functions of ESCRTs. Nat Rev Mol Cell Biol. 2020;21(1):25–42. doi: 10.1038/s41580-019-0177-4 [DOI] [PubMed] [Google Scholar]
- 23.French KC, Antonyak MA, Cerione RA. Extracellular vesicle docking at the cellular port: extracellular vesicle binding and uptake. Semin Cell Dev Biol. 2017;67:48–55. doi: 10.1016/j.semcdb.2017.01.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Xiao Y, Zhong J, Zhong B, et al. Exosomes as potential sources of biomarkers in colorectal cancer. Cancer Lett. 2020;476:13–22. doi: 10.1016/j.canlet.2020.01.033 [DOI] [PubMed] [Google Scholar]
- 25.Greenberg JW, Kim H, Moustafa AA, et al. Repurposing ketoconazole as an exosome directed adjunct to sunitinib in treating renal cell carcinoma. Sci Rep. 2021;11(1):32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Wu X, Showiheen S, Sun AR, et al. Exosomes extraction and identification. Methods Mol Biol. 2019;2054:81–91. [DOI] [PubMed] [Google Scholar]
- 27.Thery C, Witwer KW, Aikawa E, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International society for extracellular vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles. 2018;7(1):1535750. doi: 10.1080/20013078.2018.1535750 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zhang Y, Bi J, Huang J, et al. Exosome: a review of its classification, isolation techniques, storage, diagnostic and targeted therapy applications. Int J Nanomedicine. 2020;15:6917–6934. doi: 10.2147/IJN.S264498 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Gorgens A, Corso G, Hagey DW, et al. Identification of storage conditions stabilizing extracellular vesicles preparations. J Extracell Vesicles. 2022;11(6):e12238. doi: 10.1002/jev2.12238 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Gelibter S, Marostica G, Mandelli A, et al. The impact of storage on extracellular vesicles: a systematic study. J Extracell Vesicles. 2022;11(2):e12162. doi: 10.1002/jev2.12162 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Yu B, Zhang X, Li X. Exosomes derived from mesenchymal stem cells. Int J Mol Sci. 2014;15(3):4142–4157. doi: 10.3390/ijms15034142 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Cheng L, Zhang K, Wu S, Cui M, Xu T. Focus on mesenchymal stem cell-derived exosomes: opportunities and challenges in cell-free therapy. Stem Cells Int. 2017;2017:6305295. doi: 10.1155/2017/6305295 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Vakhshiteh F, Atyabi F, Ostad SN. Mesenchymal stem cell exosomes: a two-edged sword in cancer therapy. Int J Nanomedicine. 2019;14:2847–2859. doi: 10.2147/IJN.S200036 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ha DH, Kim HK, Lee J, et al. Mesenchymal stem/stromal cell-derived exosomes for immunomodulatory therapeutics and skin regeneration. Cells-Basel. 2020;9(5):1157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Moghadasi S, Elveny M, Rahman HS, et al. A paradigm shift in cell-free approach: the emerging role of MSCs-derived exosomes in regenerative medicine. J Transl Med. 2021;19(1):302. doi: 10.1186/s12967-021-02980-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Sengupta V, Sengupta S, Lazo A, et al. Exosomes derived from bone marrow mesenchymal stem cells as treatment for severe COVID-19. Stem Cells Dev. 2020;29(12):747–754. doi: 10.1089/scd.2020.0080 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Gensel JC, Zhang B. Macrophage activation and its role in repair and pathology after spinal cord injury. Brain Res. 2015;1619:1–11. doi: 10.1016/j.brainres.2014.12.045 [DOI] [PubMed] [Google Scholar]
- 38.Sheng X, Zhao J, Li M, et al. Bone marrow mesenchymal stem cell-derived exosomes accelerate functional recovery after spinal cord injury by promoting the phagocytosis of macrophages to clean myelin debris. Front Cell Dev Biol. 2021;9:772205. doi: 10.3389/fcell.2021.772205 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Hu P, Yang Q, Wang Q, et al. Mesenchymal stromal cells-exosomes: a promising cell-free therapeutic tool for wound healing and cutaneous regeneration. Burns Trauma. 2019;7:38. doi: 10.1186/s41038-019-0178-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Vallabhaneni KC, Penfornis P, Dhule S, et al. Extracellular vesicles from bone marrow mesenchymal stem/stromal cells transport tumor regulatory microRNA, proteins, and metabolites. Oncotarget. 2015;6(7):4953–4967. doi: 10.18632/oncotarget.3211 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Mu J, Wu J, Cao J, et al. Rapid and effective treatment of traumatic spinal cord injury using stem cell derived exosomes. Asian J Pharm Sci. 2021;16(6):806–815. doi: 10.1016/j.ajps.2021.10.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Yousefi DM, Goodarzi N, Azhdari MH, Doroudian M. Mesenchymal stem cells and their derived exosomes to combat Covid-19. Rev Med Virol. 2022;32(2):e2281. doi: 10.1002/rmv.2281 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Liu B, Hu D, Zhou Y, et al. Exosomes released by human umbilical cord mesenchymal stem cells protect against renal interstitial fibrosis through ROS-mediated P38MAPK/ERK signaling pathway. Am J Transl Res. 2020;12(9):4998–5014. [PMC free article] [PubMed] [Google Scholar]
- 44.Bonventre JV, Yang L. Cellular pathophysiology of ischemic acute kidney injury. J Clin Invest. 2011;121(11):4210–4221. doi: 10.1172/JCI45161 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Rajan D, Wu R, Shah KG, et al. Human ghrelin protects animals from renal ischemia-reperfusion injury through the vagus nerve. Surgery. 2012;151(1):37–47. doi: 10.1016/j.surg.2011.06.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Chen YT, Sun CK, Lin YC, et al. Adipose-derived mesenchymal stem cell protects kidneys against ischemia-reperfusion injury through suppressing oxidative stress and inflammatory reaction. J Transl Med. 2011;9(1):51. doi: 10.1186/1479-5876-9-51 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Li L, Wang R, Jia Y, et al. Exosomes derived from mesenchymal stem cells ameliorate renal ischemic-reperfusion injury through inhibiting inflammation and cell apoptosis. Front Med. 2019;6:269. doi: 10.3389/fmed.2019.00269 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Zhang Y, Wang C, Bai Z, Li P. Umbilical cord mesenchymal stem cell exosomes alleviate the progression of kidney failure by modulating inflammatory responses and oxidative stress in an ischemia-reperfusion mice model. J Biomed Nanotechnol. 2021;17(9):1874–1881. doi: 10.1166/jbn.2021.3155 [DOI] [PubMed] [Google Scholar]
- 49.Huang J, Cao H, Cui B, et al. Mesenchymal stem cells-derived exosomes ameliorate ischemia/reperfusion induced acute kidney injury in a porcine model. Front Cell Dev Biol. 2022;10:899869. doi: 10.3389/fcell.2022.899869 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Lin KC, Yip HK, Shao PL, et al. Combination of adipose-derived mesenchymal stem cells (ADMSC) and ADMSC-derived exosomes for protecting kidney from acute ischemia-reperfusion injury. Int J Cardiol. 2016;216:173–185. doi: 10.1016/j.ijcard.2016.04.061 [DOI] [PubMed] [Google Scholar]
- 51.Zhu G, Pei L, Lin F, et al. Exosomes from human-bone-marrow-derived mesenchymal stem cells protect against renal ischemia/reperfusion injury via transferring miR-199a-3p. J Cell Physiol. 2019;234(12):23736–23749. doi: 10.1002/jcp.28941 [DOI] [PubMed] [Google Scholar]
- 52.Fatemeh G, Sajjad M, Niloufar R, et al. Effect of melatonin supplementation on sleep quality: a systematic review and meta-analysis of randomized controlled trials. J Neurol. 2022;269(1):205–216. doi: 10.1007/s00415-020-10381-w [DOI] [PubMed] [Google Scholar]
- 53.Reiter RJ, Tan DX. Melatonin: a novel protective agent against oxidative injury of the ischemic/reperfused heart. Cardiovasc Res. 2003;58(1):10–19. doi: 10.1016/S0008-6363(02)00827-1 [DOI] [PubMed] [Google Scholar]
- 54.Mias C, Trouche E, Seguelas MH, et al. Ex vivo pretreatment with melatonin improves survival, proangiogenic/mitogenic activity, and efficiency of mesenchymal stem cells injected into ischemic kidney. Stem Cells. 2008;26(7):1749–1757. doi: 10.1634/stemcells.2007-1000 [DOI] [PubMed] [Google Scholar]
- 55.Alzahrani FA. Melatonin improves therapeutic potential of mesenchymal stem cells-derived exosomes against renal ischemia-reperfusion injury in rats. Am J Transl Res. 2019;11(5):2887–2907. [PMC free article] [PubMed] [Google Scholar]
- 56.Zahran R, Ghozy A, Elkholy SS, El-Taweel F, El-Magd MA. Combination therapy with melatonin, stem cells and extracellular vesicles is effective in limiting renal ischemia-reperfusion injury in a rat model. Int J Urol. 2020;27(11):1039–1049. doi: 10.1111/iju.14345 [DOI] [PubMed] [Google Scholar]
- 57.Allmang C, Kufel J, Chanfreau G, et al. Functions of the exosome in rRNA, snoRNA and snRNA synthesis. EMBO J. 1999;18(19):5399–5410. doi: 10.1093/emboj/18.19.5399 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Collino F, Pomatto M, Bruno S, et al. Exosome and microvesicle-enriched fractions isolated from mesenchymal stem cells by gradient separation showed different molecular signatures and functions on renal tubular epithelial cells. Stem Cell Rev Rep. 2017;13(2):226–243. doi: 10.1007/s12015-016-9713-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Luchetti F, Canonico B, Bartolini D, et al. Melatonin regulates mesenchymal stem cell differentiation: a review. J Pineal Res. 2014;56(4):382–397. doi: 10.1111/jpi.12133 [DOI] [PubMed] [Google Scholar]
- 60.Shen B, Liu J, Zhang F, et al. CCR2 positive exosome released by mesenchymal stem cells suppresses macrophage functions and alleviates ischemia/reperfusion-induced renal injury. Stem Cells Int. 2016;2016:1240301. doi: 10.1155/2016/1240301 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Zhang X, Wang N, Huang Y, et al. Extracellular vesicles from three dimensional culture of human placental mesenchymal stem cells ameliorated renal ischemia/reperfusion injury. Int J Artif Organs. 2022;45(2):181–192. doi: 10.1177/0391398820986809 [DOI] [PubMed] [Google Scholar]
- 62.Collino F, Lopes JA, Correa S, et al. Adipose-derived mesenchymal stromal cells under hypoxia: changes in extracellular vesicles secretion and improvement of renal recovery after ischemic injury. Cell Physiol Biochem. 2019;52(6):1463–1483. doi: 10.33594/000000102 [DOI] [PubMed] [Google Scholar]
- 63.Yun CW, Lee SH. Potential and therapeutic efficacy of cell-based therapy using mesenchymal stem cells for acute/chronic kidney disease. Int J Mol Sci. 2019;20(7):1619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Holditch SJ, Brown CN, Lombardi AM, Nguyen KN, Edelstein CL. Recent advances in models, mechanisms, biomarkers, and interventions in cisplatin-induced acute kidney injury. Int J Mol Sci. 2019;20(12):3011. doi: 10.3390/ijms20123011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Volarevic V, Markovic BS, Jankovic MG, et al. Galectin 3 protects from cisplatin-induced acute kidney injury by promoting TLR-2-dependent activation of IDO1/Kynurenine pathway in renal DCs. Theranostics. 2019;9(20):5976–6001. doi: 10.7150/thno.33959 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Wang B, Jia H, Zhang B, et al. Pre-incubation with hucMSC-exosomes prevents cisplatin-induced nephrotoxicity by activating autophagy. Stem Cell Res Ther. 2017;8(1):75. doi: 10.1186/s13287-016-0463-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Jia H, Liu W, Zhang B, et al. HucMSC exosomes-delivered 14-3-3zeta enhanced autophagy via modulation of ATG16L in preventing cisplatin-induced acute kidney injury. Am J Transl Res. 2018;10(1):101–113. [PMC free article] [PubMed] [Google Scholar]
- 68.Zhou Y, Xu H, Xu W, et al. Exosomes released by human umbilical cord mesenchymal stem cells protect against cisplatin-induced renal oxidative stress and apoptosis in vivo and in vitro. Stem Cell Res Ther. 2013;4(2):34. doi: 10.1186/scrt194 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Zhang J, He W, Zheng D, et al. ExosomalmiR1184 derived from mesenchymal stem cells alleviates cisplatin associated acute kidney injury. Mol Med Rep. 2021;24(5):1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Bruno S, Grange C, Deregibus MC, et al. Mesenchymal stem cell-derived microvesicles protect against acute tubular injury. J Am Soc Nephrol. 2009;20(5):1053–1067. doi: 10.1681/ASN.2008070798 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Collino F, Bruno S, Incarnato D, et al. AKI recovery induced by mesenchymal stromal cell-derived extracellular vesicles carrying MicroRNAs. J Am Soc Nephrol. 2015;26(10):2349–2360. doi: 10.1681/ASN.2014070710 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Bruno S, Tapparo M, Collino F, et al. Renal regenerative potential of different extracellular vesicle populations derived from bone marrow mesenchymal stromal cells. Tissue Eng Part A. 2017;23(21–22):1262–1273. doi: 10.1089/ten.tea.2017.0069 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Reis LA, Borges FT, Simoes MJ, et al. Bone marrow-derived mesenchymal stem cells repaired but did not prevent gentamicin-induced acute kidney injury through paracrine effects in rats. PLoS One. 2012;7(9):e44092. doi: 10.1371/journal.pone.0044092 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Gao F, Zuo B, Wang Y, et al. Protective function of exosomes from adipose tissue-derived mesenchymal stem cells in acute kidney injury through SIRT1 pathway. Life Sci. 2020;255:117719. doi: 10.1016/j.lfs.2020.117719 [DOI] [PubMed] [Google Scholar]
- 75.Zhang R, Zhu Y, Li Y, et al. Human umbilical cord mesenchymal stem cell exosomes alleviate sepsis-associated acute kidney injury via regulating microRNA-146b expression. Biotechnol Lett. 2020;42(4):669–679. doi: 10.1007/s10529-020-02831-2 [DOI] [PubMed] [Google Scholar]
- 76.Tong X, Yu Q, Ankawi G, et al. Insights into the role of renal biopsy in patients with T2DM: a Literature review of global renal biopsy results. Diabetes Ther. 2020;11(9):1983–1999. doi: 10.1007/s13300-020-00888-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Wang Y, Shan SK, Guo B, et al. The multi-therapeutic role of MSCs in diabetic nephropathy. Front Endocrinol (Lausanne). 2021;12:671566. doi: 10.3389/fendo.2021.671566 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Li H, Rong P, Ma X, et al. Mouse umbilical cord mesenchymal stem cell paracrine alleviates renal fibrosis in diabetic nephropathy by reducing myofibroblast transdifferentiation and cell proliferation and upregulating MMPs in mesangial cells. J Diabetes Res. 2020;2020:3847171. doi: 10.1155/2020/3847171 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Xiang E, Han B, Zhang Q, et al. Human umbilical cord-derived mesenchymal stem cells prevent the progression of early diabetic nephropathy through inhibiting inflammation and fibrosis. Stem Cell Res Ther. 2020;11(1):336. doi: 10.1186/s13287-020-01852-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Hao Y, Miao J, Liu W, et al. Mesenchymal stem cell-derived exosomes carry MicroRNA-125a to protect against diabetic nephropathy by targeting histone deacetylase 1 and downregulating endothelin-1. Diabetes Metab Syndr Obes. 2021;14:1405–1418. doi: 10.2147/DMSO.S286191 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Nagaishi K, Mizue Y, Chikenji T, et al. Mesenchymal stem cell therapy ameliorates diabetic nephropathy via the paracrine effect of renal trophic factors including exosomes. Sci Rep. 2016;6(1):34842. doi: 10.1038/srep34842 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Jin J, Shi Y, Gong J, et al. Exosome secreted from adipose-derived stem cells attenuates diabetic nephropathy by promoting autophagy flux and inhibiting apoptosis in podocyte. Stem Cell Res Ther. 2019;10(1):95. doi: 10.1186/s13287-019-1177-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Yang J, Liu Y. Dissection of key events in tubular epithelial to myofibroblast transition and its implications in renal interstitial fibrosis. Am J Pathol. 2001;159(4):1465–1475. doi: 10.1016/S0002-9440(10)62533-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Wang B, Yao K, Huuskes BM, et al. Mesenchymal stem cells deliver exogenous MicroRNA-let7c via exosomes to attenuate renal fibrosis. Mol Ther. 2016;24(7):1290–1301. doi: 10.1038/mt.2016.90 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Yang Y, Wang J, Zhang Y, et al. Exosomes derived from mesenchymal stem cells ameliorate renal fibrosis via delivery of miR-186-5p. Hum Cell. 2022;35(1):83–97. doi: 10.1007/s13577-021-00617-w [DOI] [PubMed] [Google Scholar]
- 86.Lu Y, Yang L, Chen X, et al. Bone marrow mesenchymal stem cell-derived exosomes improve renal fibrosis by reducing the polarisation of M1 and M2 macrophages through the activation of EP2 receptors. Iet Nanobiotechnol. 2022;16(1):14–24. doi: 10.1049/nbt2.12071 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Li Y, Chakraborty A, Broughton B, et al. Comparing the renoprotective effects of BM-MSCs versus BM-MSC-exosomes, when combined with an anti-fibrotic drug, in hypertensive mice. Biomed Pharmacother. 2021;144:112256. doi: 10.1016/j.biopha.2021.112256 [DOI] [PubMed] [Google Scholar]
- 88.Yin S, Zhou S, Ren D, et al. Mesenchymal stem cell-derived exosomes attenuate epithelial-mesenchymal transition of HK-2 cells. Tissue Eng Part A. 2022. doi: 10.1089/ten.tea.2021.0190 [DOI] [PubMed] [Google Scholar]
- 89.Jin J, Qian F, Zheng D, et al. Mesenchymal stem cells attenuate renal fibrosis via exosomes-mediated delivery of microRNA Let-7i-5p antagomir. Int J Nanomedicine. 2021;16:3565–3578. doi: 10.2147/IJN.S299969 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Chen L, Wang Y, Li S, et al. Exosomes derived from GDNF-modified human adipose mesenchymal stem cells ameliorate peritubular capillary loss in tubulointerstitial fibrosis by activating the SIRT1/eNOS signaling pathway. Theranostics. 2020;10(20):9425–9442. doi: 10.7150/thno.43315 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Kim H, Yu MR, Lee H, et al. Metformin inhibits chronic kidney disease-induced DNA damage and senescence of mesenchymal stem cells. Aging Cell. 2021;20(2):e13317. doi: 10.1111/acel.13317 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Liao Z, Li S, Lu S, et al. Metformin facilitates mesenchymal stem cell-derived extracellular nanovesicles release and optimizes therapeutic efficacy in intervertebral disc degeneration. Biomaterials. 2021;274:120850. doi: 10.1016/j.biomaterials.2021.120850 [DOI] [PubMed] [Google Scholar]
- 93.Hu G, Ma J, Zhang J, et al. Hypoxia-induced lncHILAR promotes renal cancer metastasis via ceRNA for the miR-613/206/ 1-1-3p/Jagged-1/Notch/CXCR4 signaling pathway. Mol Ther. 2021;29(10):2979–2994. doi: 10.1016/j.ymthe.2021.05.020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Bray F, Ferlay J, Soerjomataram I, et al. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi: 10.3322/caac.21492 [DOI] [PubMed] [Google Scholar]
- 95.Lu L, Li Y, Wen H, Feng C. Overexpression of miR-15b promotes resistance to sunitinib in renal cell carcinoma. J Cancer. 2019;10(15):3389–3396. doi: 10.7150/jca.31676 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Rini BI, Campbell SC, Escudier B. Renal cell carcinoma. Lancet. 2009;373(9669):1119–1132. doi: 10.1016/S0140-6736(09)60229-4 [DOI] [PubMed] [Google Scholar]
- 97.Himbert D, Zeuschner P, Ayoubian H, et al. Characterization of CD147, CA9, and CD70 as tumor-specific markers on extracellular vesicles in clear cell renal cell carcinoma. Diagnostics. 2020;10(12):1034. doi: 10.3390/diagnostics10121034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Li D, Lin F, Li G, Zeng F. Exosomes derived from mesenchymal stem cells curbs the progression of clear cell renal cell carcinoma through T-cell immune response. Cytotechnology. 2021;73(4):593–604. doi: 10.1007/s10616-021-00480-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Brossa A, Fonsato V, Grange C, et al. Extracellular vesicles from human liver stem cells inhibit renal cancer stem cell-derived tumor growth in vitro and in vivo. Int J Cancer. 2020;147(6):1694–1706. doi: 10.1002/ijc.32925 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Qi J, Zhou Y, Jiao Z, et al. Exosomes derived from human bone marrow mesenchymal stem cells promote tumor growth through hedgehog signaling pathway. Cell Physiol Biochem. 2017;42(6):2242–2254. doi: 10.1159/000479998 [DOI] [PubMed] [Google Scholar]
- 101.Zhang X, Sai B, Wang F, et al. Hypoxic BMSC-derived exosomal miRNAs promote metastasis of lung cancer cells via STAT3-induced EMT. Mol Cancer. 2019;18(1):40. doi: 10.1186/s12943-019-0959-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Saran R, Robinson B, Abbott KC, et al. US renal data system 2017 annual data report: epidemiology of kidney disease in the United States. Am J Kidney Dis. 2018;71(3 Suppl 1):A7. doi: 10.1053/j.ajkd.2018.01.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Wu X, Wang Z, Wang J, et al. Exosomes secreted by mesenchymal stem cells induce immune tolerance to mouse kidney transplantation via transporting LncRNA DANCR. Inflammation. 2022;45(1):460–475. doi: 10.1007/s10753-021-01561-5 [DOI] [PubMed] [Google Scholar]
- 104.Han C, Qin G. Reporter systems for assessments of extracellular vesicle transfer. Front Cardiovasc Med. 2022;9:922420. doi: 10.3389/fcvm.2022.922420 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Buschmann D, Mussack V, Byrd JB. Separation, characterization, and standardization of extracellular vesicles for drug delivery applications. Adv Drug Deliv Rev. 2021;174:348–368. doi: 10.1016/j.addr.2021.04.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Grange C, Tapparo M, Bruno S, et al. Biodistribution of mesenchymal stem cell-derived extracellular vesicles in a model of acute kidney injury monitored by optical imaging. Int J Mol Med. 2014;33(5):1055–1063. doi: 10.3892/ijmm.2014.1663 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Gupta D, Liang X, Pavlova S, et al. Quantification of extracellular vesicles in vitro and in vivo using sensitive bioluminescence imaging. J Extracell Vesicles. 2020;9(1):1800222. doi: 10.1080/20013078.2020.1800222 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Han Z, Liu S, Pei Y, et al. Highly efficient magnetic labelling allows MRI tracking of the homing of stem cell-derived extracellular vesicles following systemic delivery. J Extracell Vesicles. 2021;10(3):e12054. doi: 10.1002/jev2.12054 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Cao H, Cheng Y, Gao H, et al. In vivo tracking of mesenchymal stem cell-derived extracellular vesicles improving mitochondrial function in renal ischemia-reperfusion injury. Acs Nano. 2020;14(4):4014–4026. doi: 10.1021/acsnano.9b08207 [DOI] [PubMed] [Google Scholar]
- 110.He Q, Wang L, Zhao R, et al. Mesenchymal stem cell-derived exosomes exert ameliorative effects in type 2 diabetes by improving hepatic glucose and lipid metabolism via enhancing autophagy. Stem Cell Res Ther. 2020;11(1):223. doi: 10.1186/s13287-020-01731-6 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]