

Abstract
Background
Pharmacotherapy with medicinal plants is a promising approach to treat cancer. Cinnamon is a medicinal plant whose properties have been proven in various fields of medical sciences. Among its biological activities, its antioxidant and antiviral effects can be mentioned. In this study, the antitumor effects of Cinnamon with a focus on glucose metabolism in bladder cancer carcinoma cell-line 5637 were investigated.
Methods
Aqueous extract of Cinnamon was prepared from Cinnamon bark. Bladder cancer 5637cell line were treated with different concentrations of aqueous extract of Cinnamon. MTT was used to evaluate cell viability at 24, 48, and 72 h. The concentration of 1.25, 2.50, and 5 mg/ml was used. Apoptosis was assessed with Hochest33258 staining. For evaluating of aqueous extract of Cinnamon effect on glycolysis, the gene expression of epidermal growth factor receptor 2 (ErbB2), heat shock protein transcription factor1 (HSF1), and lactate dehydrogenase A (LDHA), as well as protein levels of HSF1 and LDHA, LDH activity, glucose consumption, and lactate production, were measured.
Results
Aqueous extract of Cinnamon significantly decreased ErbB2, HSF1, and LDHA gene expression and also decreased the protein level of HSF1 and LDHA, LDH activity, glucose consumption, and lactate production dose-dependently (p < 0.05).
Conclusion
Our finding showed that the aqueous extract of Cinnamon can inhibit proliferation in 5637 cells by inhibition of glycolysis and induction of apoptosis.
1. Introduction
Bladder cancer is the 10th most common cancer worldwide, according to the World Health Organization. The rapid and progressive spread of this cancer has become a major concern in the world and kills millions of people every year [1]. An unhealthy lifestyle, alcohol, tobacco use, and other factors can play an important role in the development of many cancers and chronic diseases such as diabetes and cancer. Environment, nutritional, and occupational factors can activate oxidative stress in the body [2–4].
Oxidative stress is a stimulus for the onset and development of cancer because this process begins with the activation of inflammatory processes in the body and eventually ends with the development of chronic diseases such as diabetes, cardiovascular disease, cancer, and many disorders [5]. Second, oxidative stress helps convert normal cells to cancer cells, the progression, migration, recurrence of cancer, and many other features that are beneficial for cancer cells. Oxidative stress also helps convert normal cells to cancer cells, the progression, migration, recurrence of cancer, and many other features that are beneficial for cancer cells [5–7].
At first, the body's response to induced oxidative stress is mediated by a defense mechanism called the thermal shock response which is regulated by a transcription factor called heat shock protein transcription factor1 (HSF1). HSF1 is known as a powerful factor with different functions, that in addition to its role in the defense responses is also involved in regulating many pathways related to cell growth, proliferation, and metabolism such as glycolysis and lipid metabolism [8, 9]. In addition, various studies revealed that HSF1 is associated with metastasis, cancer cell survival, and tumor proliferation. In other words, elevated expression of HSF1 has been observed with a poor prognosis of cancer [10]. This association is due to the role of HSF1 in activating pathways such as protein kinase C (PKC) [11], nuclear factor-kappaB (NF-κB) [12], phosphoinositide 3-kinase (PI3K), Akt/mammalian (or mechanistic) target of rapamycin (mTOR) [8], which are essential for proliferation, resistance to apoptosis, migration, invasion, and cancer cell metastasis [8, 10, 13, 14].
Epidermal growth factor receptor 2 (ErbB2) is a potent oncogene associated with many cancers, including breast cancer, and its increased expression has been seen in the poor prognosis of cancer. This oncogene is a regulator of cell bioenergetic pathways such as ras, PI3K/AKT, and mTOR [15–17]. Inhibition of ErbB2 can reduce the expression of HSF1 [18].
Unlike normal cells, the energy supply in cancer cells is mainly through the anaerobic pathway or glycolysis [19]. Continuation of glycolysis in cancer cells requires the supply of NADH, produced by lactate dehydrogenase (LDH). One of that isoenzymes is LDHA, which can be increased in most cancers [20]. Increased HSF1 can act a rule as a transcription factor and cause evaluate the expression of lactate dehydrogenase A (LDHA), and oppositely reducing HSF1 can decrease the expression of LDHA, which targets one of the vital processes of cancer cells, which is actually glycolysis [21]. Therefore, due to the importance of glycolysis for cancer cells' survival, targeting these factors as regulators for glycolysis can be a good target for cancer treatment [22].
Since chemotherapy drugs have many side effects, researchers have always been looking for natural compounds that are the least harmful to the body. The Cinnamon plant, known as Cinnamon and Cinnamomum, belongs to the Lauraceae family and has been used as a spice in many countries for centuries. In addition to its use in cooking, it has been used as a gastrointestinal pain reliever in some diseases [23]. Cinnamon has various medicinal properties such as antioxidant [24, 25], anti-viral [26–28], anti-inflammatory [29], and anti-diabetic [30] effects.
In this study, we investigated the effects of the aqueous extract of Cinnamon on human bladder carcinoma cell line 5637, especially on the gene expression of HSF1, ErbB2, and LDHA and the level of LDHA and HSF1 proteins and apoptosis.
2. Material and Method
2.1. Preparation of Aqueous Extract of Cinnamon
Cinnamon sticks were purchased from a local market in this study. Cinnamomum cassia confirmed by an herbalist was used. It was then grounded. 250 g of the powder was taken and soaked in 2500 ml of water for 24 h at room temperature. Then it was boiled at 100°C for 30 min. Afterward, it was filtered and the resulting solution was lyophilized and stored at −20°C until used [31].
2.2. HPLC Analysis
HPLC analysis was performed to determine the amount of Cinnamaldehyde in the prepared extract. Commercial Cinnamaldehyde manufactured by Sigma (Cat. No. W228613) is used as standards makers for the quality control of the composition of Cinnamaldehyde in the aqueous extract of Cinnamon in each experiment. Chromatography was performed by 1% acetic acid (H20)-MeOH (50 : 50 v/v) at room temperature on a Phenomenex Luna 5u C18, 100 A pore size, 250 × 4.60 mm I.D. column. The flow rate of the mobile phase was 1 ml/min [32]. According to the results, the amount of Cinnamaldehyde in the prepared extract was 26 μg/ml.
2.3. Cell Culture
Human bladder carcinoma cell line 5637 was purchased from Pasteur Institute, Tehran, Iran. The cells were cultured in RPMI1640 (Gibco RL, Grand Island, NY) medium with 10% FBS (Gibco RL, Grand Island, NY), 20 mM HEPES, 2 mM glutamine, and 1% of penicillin-streptomycin in 37°C incubators under 5% CO2. When the confluency of cells reached 85%, experiments were performed.
2.4. MTT Colorimetric Assay
Cell viability was evaluated by the MTT method to determine the optimal concentration and time for treatment of cells [33] with aqueous extract of Cinnamon. 2 × 104 cells were seeded in 96 well plates. The cells incubated with concentrations of 0.16, 0.32, 0.64, 1.25, 2.50, 5, 10, and 20 mg/ml aqueous extract of Cinnamon at 37°C under 5% CO2 for 24, 48, and 72 h [34]. After treatment, 20 μl of MTT working solution was added to each well and incubated at 37° for 3 h. The supernatant was collected and the formazan crystals were dissolved in DMSO (100 μl/sample). Finally, cell viability was assessed by measuring light absorption at 590 nm by using a plate reader (Biotech Instruments Inc., Winooski, USA).
2.5. Hochest33258 Staining
The Hochest staining 33285 (Sigma, USA) was used to evaluate the occurrence of apoptosis. First, the cells were cultured in plates of 6 wells (106 cells/well). After 24 h treatment, the cells were trypsinized and the contents of each well were transferred to a separate microtube. Then, the tubes were centrifuged at 1500g for 5 min. After removing the supernatant, 100–200 μl of absolute cold methanol was added to the precipitate for cell fixation and incubated for 15 min at −20°C. After incubation, centrifuged again and added 50 μl PBS and 1 μl ready-to-use Hochest solution (1 μl Hochest stock per 100 μl PBS buffers) to the cell precipitate and incubate for 30 minutes at laboratory temperature. It was centrifuged again and 100 μl PBS was added to the precipitate. Then mixed and transferred to a slide for observation using a fluorescence microscope.
2.6. Total RNA Extraction and Real-Time PCR
Total RNA was extracted by KIAZOL (KIAZIST, Iran) after treating the cells with specific concentrations of aqueous extract of Cinnamon. The quantity and quality of the extracted RNA were determined by NanoDrop ND-1000 spectrophotometer and 1% agarose gel electrophoresis, respectively. Synthesis of cDNA from total RNA was performed with a cDNA synthesis kit (Biofact, Korea) according to the manufacturer's instructions. Polymerase chain reaction (PCR) was performed using gene-specific primers and SYBR Green kit (Biofact, Korea). Following primers were used; Glyceraldehyde 3-phosphate dehydrogenase(GAPDH) 5′-AAGGCTGTGGGCAAGGTCATC-3′ (forward) and 5′-GCGTCAAAGGTGGAGGAGTGG-3′ (reverse), HSF1: 5′-GAAGGGGAAGCAGGAGTG-3′ (forward) and 5′-GTTGACGACTTTCTGTTGCTG-3′ (reverse), ErbB2: 5′-TGATAGACACCAACCGCTCT-3′ (forward) and 5′-CAGAACTCTCTCCCCAGCA-3′ (reverse) and LDHA: 5′-GGTCCTTGGGGAACATGGAG-3′ (forward) and 5′-TAGCCCAGGATGTGTAGCCT-3′ (reverse). GAPDH is housekeeping gene and here it was used as internal control. The specificity of PCR products was verified by gel electrophoresis in 1% agarose. Finally, the relative genes expression was evaluated by 2−ΔΔCt method.
2.7. Western Blotting
A total of 5637 cells were treated and then lysed in RIPA buffer (KIAZIST, Hamadan, Iran) supplemented with protease inhibitor cocktail (KIAZIST, Iran). Then agitated for 30 min at 4°C. Next, centrifuged (10 min 12,000g at 4°C), the supernatant was collected and protein concentration was assayed by the bicinchoninic acid (BCA) method (BCA, Thermo Scientific pierce, USA). Protein (40 μg) was loaded on 10% SDS-polyacrylamide gels and then transferred to the nitrocellulose membrane. The membrane was blocked with 5% non-fat dried milk in TBS-T (50 mM Tris-HCl, pH 7.6, 100 Mm NaCl, and 0.1% Tween20) for 1 h. Subsequently, the membrane was incubated with anti-HSF1(1 : 2000 dilution), anti-LDHA (1 : 1000 dilution) and anti-βactin (1 : 2000 dilution) antibodies overnight at 4°C. For visualizing the immunoreactivity of proteins enhanced chemiluminescence (ECL) was used. Finally, the band intensity was quantified using ImageJ software (NIH, Bethesda, USA). β-actin was used as the internal control to normalize intensity values.
2.8. LDH Activity Assay
Treated cells were lysed by freeze-thawing and then used PBS buffer supplemented with antiprotease. afterward, it was centrifuged at 15000g for 15 min. The obtained lysates were used to measure activity LDH according to the manufacturers' instructions kit (Pars Azmun, Iran). Total protein was also quantified by bicinchoninic acid assay (BCA, Thermo Scientific Pierce, USA).
2.9. Measurement of the Glucose Consumption and Lactate Production
Glucose intake and lactate production was measured in the supernatant of media after treatment. The cells were treated with certain concentrations of aqueous extract of Cinnamon for 24 h, then the supernatant was collected and used to evaluate lactate and glucose according to the kit manufacturer's instructions (Pars Azmun, Iran).
2.10. Statistical Analysis
Data analysis was performed by IBM SPSS Statistics 16.0 (IBM, USA) and Microsoft Graph Pad Prism 6 (Graph Pad Prism Inc., USA) used for drawing graphs. To determine statistical differences, we performed one-way ANOVA and Tukey's post-test. Results were represented as mean ± standard division (SD). P < 0.05 was considered statistically significant.
3. Result
3.1. MTT Assay and Cell Viability
The results obtained from the MTT assay showed that aqueous extract of Cinnamon has time and dose-dependent effects on 5637 cells, as shown in Figure 1. After 24, 48, and 72 h treatment of 5637 cells with (0.16, 0.32, 0.64, 1.25, 2.50, 5, 10, and 20 mg/ml) aqueous extract of Cinnamon concentrations MTT test was performed. At 72 h, concentrations above 2.50 mg/ml were toxic to cells. At 48 hours after treatment, concentrations above 5 mg/ml showed cell viability below 50% but cell viability at 24 h, for all used concentrations was above 50%, except 10 and 20 mg/ml. According to the results obtained from MTT, the IC50 was calculated at 10 mg/ml. Therefore, concentrations of 1.25, 2.50, and 5 mg/ml and 24 h were selected to continue the experiments.
Figure 1.
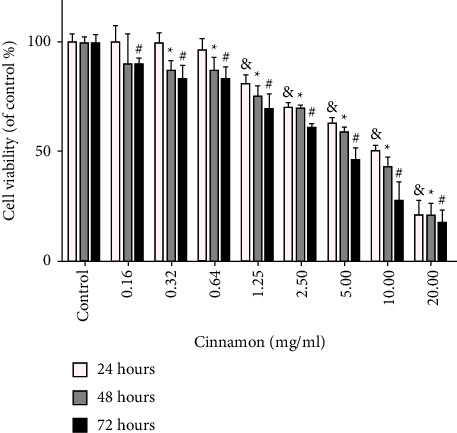
Effect of different concentrations of Cinnamon aqueous extract on 5637cell viability. 5637cell were treated with concentrations (0.16, 0.32, 0.64, 1.25, 2.50, 5, 10, and 20 mg/ml) for 24, 48, and 72 h Results are expressed as mean ± SD and each value is the average of at least three independent replicates. & denoted the significant difference in comparison with the control group in 24 h (P < 0.05), ∗ in 48 h (P < 0.05), # in 72 h. (P < 0.05) compared to the control group.
3.2. Apoptosis Assay and Hochest33258 Staining
The Hochest staining is used for investigating apoptosis in cell culture. In this method, normal cells are seen uniformly and have a normal nucleus, whereas apoptotic cells' nuclei have dense Chromatin and nuclear fragmentation and seem irregular appearance, also they seem as bright spots. For this purpose, 5637 cells were treated for 24 h with 1.25, 2.50, and 5 mg/ml aqueous extract of Cinnamon. Then cells were stained with Hochest33258 followed by observation under a fluorescence microscope. As shown in Figure 2, bright spots that represented apoptosis in cells and irregular luminosity compared to control were seen. Therefore, it can be concluded that aqueous extracts of Cinnamon induce apoptosis at these concentrations.
Figure 2.

Effect of different concentrations of Cinnamon aqueous extract on 5637cell apoptosis by Hochest33258 staining. All ×40 magnification images are obtained by fluorescence microscopy.
3.3. mRNA Expression of HSF1, ErbB2, and LDHA in 5637 Cells
To determine the effect of the aqueous extract of Cinnamon on the inhibition of glucose metabolism in 5637 cells, the expression of genes HSF1, ErbB2 and LDHA were evaluated. Our results showed that different aqueous extracts of Cinnamon in selected concentrations could reduce the expression of all three genes in a dose-dependent condition compared to the control group. As shown in Figure 3(a), gene expression of HSF1 was significantly reduced at concentrations of 2.50 (P < 0.05) and 5 mg/ml (P < 0.001). At a concentration of 1.25 mg/ml, the decrease was not significant compared to the control group (P > 0.05). Aqueous extract of Cinnamon reduced, significantly the expression of ErbB2 for 2.50 and 5 mg/ml (P < 0.001). There was no significant change for 1.25 mg/ml compared to the control group (P > 0.05) (Figure 3(b)). LDHA gene expression was significantly lowered in all three concentrations (Figure 3(c)), thus there were significant changes in concentrations of 1.25 mg/ml (P < 0.05), 2.50, and 5 mg/ml (P < 0.001).
Figure 3.

Effect of different concentrations of Cinnamon aqueous extract on mRNA expression (fold change) of HSF1 (a), ErbB2 (b), and LDHA (c). Data are represented as the mean ± SD. Each value is the average of at least three independent replicates. (∗P < 0.05, ∗∗∗P < 0.001compared to the control group). ErbB2: human epidermal growth factor receptor2, HSF1: heat-shock transcription factor1, LDHA: lactate dehydrogenase A.
3.4. Level of HSF1 and LDHA Proteins in 5637 Cells
The results showed that level of HSF1 and LDHA protein in treated cells with aqueous extract of Cinnamon was significantly reduced. As shown in Figure 4(b), the level of HSF1 in 2.50 (P < 0.05) and 5 mg/ml (P < 0.01) was significantly decreased. The concentration of 1.25 mg/ml showed no significant change compared to the control group (P > 0.05). LDHA protein expression was significantly reduced in three used concentrations. This diminution in protein levels was seen as dose-dependent significantly compared to lower concentrations (Figure 4(c)), which was for 1.25 mg/ml (P < 0.05), 2.50 mg/ml, and 5 mg/ml (P < 0.001).
Figure 4.

Effect of different concentrations of Cinnamon aqueous extract on 5637cell. (a) The image of protein extraction by the immunoblotting result, (b) HSF1, and (c) LDHA protein expression. Data are reported as the mean ± SD and each value is the average of at least three independent replicates. (∗P < 0.05, ∗∗P < 0.01 and ∗∗∗P < 0.001 compared to the control group). HSF1: heat-shock transcription factor1, LDHA: lactate dehydrogenase A.
3.5. LDH Activity
To confirm the effects of the aqueous extract of Cinnamon on the reduction of glycolysis, an LDH activity assay was performed. According to Figure 5, aqueous extract of Cinnamon could reduce LDH activity. LDH activity was significantly reduced at 1.25 (P < 0.05), 2.5 (P < 0.01), and 5 mg/ml (P < 0.001) compared to the control group.
Figure 5.

Effect of different concentrations of Cinnamon aqueous extract on LDH activity in5637cells. Data are reported as the mean ± SD. All reported value is the average of at least three independent replicates. (∗P < 0.05, ∗∗P < 0.01 and ∗∗∗P < 0.001 compared to control group). LDH: lactate dehydrogenase.
3.6. Glucose Consumption and Lactate Production
Glucose uptake and lactate production in media was assayed to ensure glycolysis inhibition. Results were shown (Figure 6) that glucose consumption and lactate production were significantly decreased in three used concentrations time and dose-dependently. Glucose intake for 5637 cells from media after 24 h treatment was significantly decreased in all used concentrations (P < 0.01) (Figure 6(a)). Analysis of data obtained from lactate consumption by 5637 cells in concentrations of 1.25 (P < 0.01), 2.5 and 5 mg/ml (P < 0.001) showed a significant decrease in lactate consumption compared to the control group (Figure 6(b)).
Figure 6.

Effect of different concentrations of Cinnamon aqueous extract on glucose consumption (a) and lactate production (b) in 5637 cells. Data are represented as the mean ± SD. All value is the average of at least three independent replicates. (∗∗P < 0.01 and ∗∗∗P < 0.001 compared to the control group).
4. Discussion
The World Health Organization estimates cancer as the first or second leading cause of death before the age of 70 in most countries. This issue caused finding the best way for controlling or treating cancer be the priority for researchers who work in this field. Although chemotherapy is routinely used to treat many cancers, sometimes effective yet mostly has hazardous side effects that are limiting its therapeutic potential [35–37].
The use of plants such as Cinnamon has always been considered in medical sciences due to their proven properties. Various compounds in the Cinnamon plant, such as Cinnamaldehyde, have made it possible to be effective in cancer therapy or chemoprevention by various mechanisms such as inhibition of NF-KB or activated protien1 (AP1) [32], angiogenesis [38], and vascular endothelial growth factor (VEGF) pathways [39]. In this study, we showed that Cinnamon reduced the expression of ErbB2, HSF1, and LDHA at transcriptional and translational levels. In the present study, it was seen that cinnamon extract reduced all three ErbB2, HSF1, and LDHA genes. The relationship between these three genes has been proven in a study conducted in 2009 by Zhao et al. They observed that with the decrease in the expression of ErbB2, the expression level of LDHA gene and enzyme activity of LDH both decrease. They also reported that when siRNA was used to inhibit LDHA gene expression, in addition to inhibiting glucose uptake into cell cytoplasm and lactate production, it can reduce cell growth and prevent overexpression of ErbB2. They also observed that the amount of LDHA also decreased following a decrease in HSF1 in the cell, and this could be due to the direct effect of the expression of HSF1 as a transcription factor [21]. Different mechanisms are related to the ErbB2-HSF1-LDHA pathway. Overexpression of ErbB2 was seen in bladder cancer and is associated with poor prognosis [40–42]. ErbB2 calls PI3K and then AKT and mTOR through the signaling cascade it activates. HSF1 is an activation target for mTOR that activates HSF1 by phosphorylation of serine 326. By activating HSF1, proteins called HSPs are activated. Each of the HSPs serves the cancer cell in certain ways. HSPs proteins participate in many processes such as angiogenesis, metastasis, and cancer cell metabolism [43, 44]. HSP27,70 and 90 have an important role in the reactivation of ErbB2, which reactivates the entire mTOR pathway as an important way for cancer cell survival. HSP90 expression is directly affected by HSF1. HSP90 stabilizes many proteins such as platelet-derived growth factor receptor (PDGFR), vascular endothelial growth factor receptor (VEGFR), and even ErbB2 [10]. Ras, raf, ERK, and the MAPK pathway are all activated by ErbB2. All of these events occur in the form of direct feedback, followed by activation one after the other. The result of these events is an increase in transcription from intracellular glucose transporter genes and other genes that are involved in energy production, such as lactate dehydrogenase, phosphofructokinase, and pyruvate kinase, which is further evidence of altered metabolism in cancer cells known as the Warburg effect [45]. On the other hand, HSF1 plays an important role as a transcription factor in regulating a process called metabolic reprogramming in cancer cells. HSF1 also directly increases its expression by acting on the LDHA gene. The pyruvate dehydrogenase kinase 3 gene is another target for HSF1, which inhibits the conversion of glucose to Acetyl CoA by inactivating the pyruvate dehydrogenase complex, thus ensuring the continuation of glycolysis in cancer cells [46]. On the other hand, HSF1 recruits methyltransferase DNMT3 to the MN317 lncRNA gene region. These methyltransferases prevent the activation of MIR317 by hypermethylating this gene region. Because MIR317 causes the activation of glutaminase GLS1, which helps to reactivate mTOR by hydrolyzing glutamine. Thus, HSF1, which is a target for mTOR, is also reactivated [10, 47, 48] for the first time Current study showed an aqueous extract of Cinnamon, is able to reduce the expression of ErbB2 and HSF1; consequently, the expression of LDHA is also reduced. Also, we measured LDH activity, glucose uptake, and lactate production, which indicate glycolysis and lactate dehydrogenase activity, respectively. Results confirmed an aqueous extract of Cinnamon can reduce glucose metabolism in the 5637 cell line.
Moreover, we are using Hochest23258 to detect apoptotic cells and four finding shows Cinnamon could induce apoptosis in 5637 cells, which is consistent with previous studies. Soumya et al. showed that in cervical cancer cells aqueous extract of Cinnamon increases intracellular calcium signaling and alters the permeability of mitochondrial potential consequently inducing apoptosis [49]. Gwang et al. demonstrated water extract of Cinnamon can induce apoptosis in a colorectal cancer cell by ROS-dependent NF-κB and ATF3 activation [50]. Another point is the importance of HSF1, it has a vital role in protecting cancer cells from apoptosis and helping them to invade from the immune system by metastasis. Therefore, decreased expression and inactivation of HSF1, has protecting effect on the host [51, 52]. In fact, Cinnamon and its active components kill cancer cells directly by their regulating act on transcription factors such as HSF1, and kill indirectly by increasing of ROS production and mitochondrial dysfunction. Cinnamaldehyde is the active ingredient in Cinnamon extract, which can induce apoptosis in different cancer cells. In this regard, we can refer to the study of Hyeon et al. in 2003. It is one of the first published reports about the inhibitory properties of Cinnamaldehyde in cancer cells in HL-60 cells. They showed Cinnamaldehyde can impair the cell's oxidative system by increasing ROS production and subsequently induce apoptosis by increasing mitochondrial membrane permeability and cytochrome C releasing [53]. In the next years after 2003, a lot of research was done about the inhibitory effect of Cinnamaldehyde and Cinnamon extract, they confirmed the report of Hyeon et al with different mechanisms. In 2020, Lee et al. showed that Cinnamaldehyde induces cell apoptosis by interfering with mitochondrial function, disrupting mitochondrial membrane balance, and increasing permeability (∆Mψ), as well as increasing intracellular calcium [54].
5. Conclusion
Our results suggest that the aqueous extract of Cinnamon suppresses 5637 cells proliferation through inhibition of glycolysis, and induces apoptosis. From these findings, the aqueous extract of Cinnamon can act as a natural candidate for the development of chemoprevention or therapeutic agents in human bladder cancer.
Acknowledgments
The study was funded by Vice-Chancellor for Research and Technology, Hamadan University of Medical Sciences, Iran (No. 9804182856).
Data Availability
All data have sufficient accuracy and precision. All the reported results are obtained from the detailed analysis of the raw data obtained from the experiments.
Ethical Approval
Hamadan University of Medical Sciences approved this research by ID: IR.UMSHA.REC.1398.227 (approval date: 2019-06-15).
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Authors' Contributions
Z.A. performed the experiments and wrote the manuscript, N.A. generated and developed the study hypothesis and design, R.A. revised the manuscript, S.D. performed part of the experiments, S.H. analyzed the data, and A.M. completed the final version of the manuscript. The authors declare that all data were generated in-house and that no paper mill was used.
References
- 1.Powles T., Bellmunt J., Comperat E., et al. Bladder cancer: ESMO clinical practice guideline for diagnosis, treatment and follow-up. Annals of Oncology . 2022;33(3):244–258. doi: 10.1016/j.annonc.2021.11.012. [DOI] [PubMed] [Google Scholar]
- 2.Sasco A., Secretan M., Straif K. Tobacco smoking and cancer: a brief review of recent epidemiological evidence. Lung Cancer . 2004;45:S3–S9. doi: 10.1016/j.lungcan.2004.07.998. [DOI] [PubMed] [Google Scholar]
- 3.Weiderpass E. Lifestyle and cancer risk. Journal of Preventive Medicine and Public Health . 2010;43(6):459–471. doi: 10.3961/jpmph.2010.43.6.459. [DOI] [PubMed] [Google Scholar]
- 4.Møller P., Wallin H., Knudsen L. E. Oxidative stress associated with exercise, psychological stress and life-style factors. Chemico-Biological Interactions . 1996;102(1):17–36. doi: 10.1016/0009-2797(96)03729-5. [DOI] [PubMed] [Google Scholar]
- 5.Sosa V., Moliné T., Somoza R., Paciucci R., Kondoh H., LLeonart M. E. Oxidative stress and cancer: an overview. Ageing Research Reviews . 2013;12(1):376–390. doi: 10.1016/j.arr.2012.10.004. [DOI] [PubMed] [Google Scholar]
- 6.Visconti R., Grieco D. New insights on oxidative stress in cancer. Current Opinion in Drug Discovery & Development . 2009;12(2):240–245. [PubMed] [Google Scholar]
- 7.Toyokuni S., Okamoto K., Yodoi J., Hiai H. Persistent oxidative stress in cancer. FEBS Letters . 1995;358(1):1–3. doi: 10.1016/0014-5793(94)01368-b. [DOI] [PubMed] [Google Scholar]
- 8.Jiang S., Tu K., Fu Q., et al. Multifaceted roles of HSF1 in cancer. Tumor Biology . 2015;36(7):4923–4931. doi: 10.1007/s13277-015-3674-x. [DOI] [PubMed] [Google Scholar]
- 9.Vihervaara A., Sistonen L. HSF1 at a glance. Journal of Cell Science . 2014;127(2):261–266. doi: 10.1242/jcs.132605. [DOI] [PubMed] [Google Scholar]
- 10.Wang G., Cao P., Fan Y., Tan K. Emerging roles of HSF1 in cancer: cellular and molecular episodes. Biochimica et Biophysica Acta (BBA)—Reviews on Cancer . 2020;1874(1) doi: 10.1016/j.bbcan.2020.188390.188390 [DOI] [PubMed] [Google Scholar]
- 11.Kumar S., Tomar M. S., Acharya A. HSF1-mediated regulation of tumor cell apoptosis: a novel target for cancer therapeutics. Future Oncology . 2013;9(10):1573–1586. doi: 10.2217/fon.13.106. [DOI] [PubMed] [Google Scholar]
- 12.Rao R., Home T., Yacoub A., et al. Heat shock factor 1 promotes NF-κb and B-cell signaling in a preclinical model of chronic lymphocytic leukemia. Blood . 2015;126 [Google Scholar]
- 13.Carpenter R. L., Gökmen-Polar Y. HSF1 as a cancer biomarker and therapeutic target. Current Cancer Drug Targets . 2019;19(7):515–524. doi: 10.2174/1568009618666181018162117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Calderwood S. K. HSF1, a versatile factor in tumorogenesis. Current Molecular Medicine . 2012;12(9):1102–1107. doi: 10.2174/156652412803306675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Slamon D. J., Godolphin W., Jones L. A., et al. Studies of the HER-2/neu proto-oncogene in human breast and ovarian cancer. Science . 1989;244(4905):707–712. doi: 10.1126/science.2470152. [DOI] [PubMed] [Google Scholar]
- 16.Omenn G. S., Guan Y., Menon R. A new class of protein cancer biomarker candidates: differentially expressed splice variants of ERBB2 (HER2/neu) and ERBB1 (EGFR) in breast cancer cell lines. Journal of Proteomics . 2014;107:103–112. doi: 10.1016/j.jprot.2014.04.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Harari D., Yarden Y. Molecular mechanisms underlying ErbB2/HER2 action in breast cancer. Oncogene . 2000;19(53):6102–6114. doi: 10.1038/sj.onc.1203973. [DOI] [PubMed] [Google Scholar]
- 18.De Thonel A., Mezger V., Garrido C. Implication of heat shock factors in tumorigenesis: therapeutical potential. Cancers . 2011;3(1):1158–1181. doi: 10.3390/cancers3011158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hsu P. P., Sabatini D. M. Cancer cell metabolism: warburg and beyond. Cell . 2008;134(5):703–707. doi: 10.1016/j.cell.2008.08.021. [DOI] [PubMed] [Google Scholar]
- 20.Ooi A. T., Gomperts B. N. Molecular pathways: targeting cellular energy metabolism in cancer via inhibition of SLC2A1 and LDHA. Clinical Cancer Research . 2015;21(11):2440–2444. doi: 10.1158/1078-0432.ccr-14-1209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zhao Y. H., Zhou M., Liu H., et al. Upregulation of lactate dehydrogenase A by ErbB2 through heat shock factor 1 promotes breast cancer cell glycolysis and growth. Oncogene . 2009;28(42):3689–3701. doi: 10.1038/onc.2009.229. [DOI] [PubMed] [Google Scholar]
- 22.Zhao Y., Butler E. B., Tan M. Targeting cellular metabolism to improve cancer therapeutics. Cell Death & Disease . 2013;4 doi: 10.1038/cddis.2013.60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Gruenwald J., Freder J., Armbruester N. Cinnamon and health. Critical Reviews in Food Science and Nutrition . 2010;50(9):822–834. doi: 10.1080/10408390902773052. [DOI] [PubMed] [Google Scholar]
- 24.Mancini-Filho J., Van-Koiij A., Mancini D. A., Cozzolino F. F., Torres R. P. Antioxidant activity of cinnamon (Cinnamomum zeylanicum, breyne) extracts. Bollettino Chimico Farmaceutico . 1998;137(11):443–447. [PubMed] [Google Scholar]
- 25.Mathew S., Abraham T. E. Studies on the antioxidant activities of cinnamon (Cinnamomum verum) bark extracts, through various in vitro models. Food Chemistry . 2006;94(4):520–528. doi: 10.1016/j.foodchem.2004.11.043. [DOI] [Google Scholar]
- 26.Sadeghi S., Davoodvandi A., Pourhanifeh M. H., et al. Anti-cancer effects of cinnamon: insights into its apoptosis effects. European Journal of Medicinal Chemistry . 2019;178:131–140. doi: 10.1016/j.ejmech.2019.05.067. [DOI] [PubMed] [Google Scholar]
- 27.Fatima M., Zaidi N. U S. S., Amraiz D., Afzal F. In vitro antiviral activity of Cinnamomum cassia and its nanoparticles against H7N3 influenza A virus. Journal of Microbiology and Biotechnology . 2016;26(1):151–159. doi: 10.4014/jmb.1508.08024. [DOI] [PubMed] [Google Scholar]
- 28.Tamam S., Abd el Hamid M., Samah M. H., Marwa A. N. The anti-viral and immunomodulatory activity of cinnamon zeylanicum against “NDV” newcastle disease virus in chickens. Group . 2017;1 [Google Scholar]
- 29.Mohammed G. J., Hameed I. H. Anti-fungal, antitumor and anti-inflammatory activity of acorus calamus. Indian Journal of Public Health Research & Development . 2018;9(3):254–258. doi: 10.5958/0976-5506.2018.00218.8. [DOI] [Google Scholar]
- 30.Bandara T., Uluwaduge I., Jansz E. R. Bioactivity of cinnamon with special emphasis on diabetes mellitus: a review. International Journal of Food Sciences & Nutrition . 2012;63(3):380–386. doi: 10.3109/09637486.2011.627849. [DOI] [PubMed] [Google Scholar]
- 31.Shen Y., Honma N., Kobayashi K., et al. Cinnamon extract enhances glucose uptake in 3T3-L1 adipocytes and C2C12 myocytes by inducing LKB1-AMP-activated protein kinase signaling. PLoS One . 2014;9(2) doi: 10.1371/journal.pone.0087894.e87894 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kwon H.-K., Hwang J.-S., So J.-S., et al. Cinnamon extract induces tumor cell death through inhibition of NF-κB and AP1. BMC Cancer . 2010;10(1):392–410. doi: 10.1186/1471-2407-10-392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Dash S. G., Kantevari S., Guru S. K., Naik P. K. Combination of docetaxel and newly synthesized 9-Br-trimethoxybenzyl-noscapine improve tubulin binding and enhances antitumor activity in breast cancer cells. Computers in Biology and Medicine . 2021;139 doi: 10.1016/j.compbiomed.2021.104996.104996 [DOI] [PubMed] [Google Scholar]
- 34.Singh R., Koppikar S. J., Paul P., Gilda S., Paradkar A. R., Kaul-Ghanekar R. Comparative analysis of cytotoxic effect of aqueous cinnamon extract from Cinnamomum zeylanicum bark with commercial cinnamaldehyde on various cell lines. Pharmaceutical Biology . 2009;47(12):1174–1179. doi: 10.3109/13880200903019242. [DOI] [Google Scholar]
- 35.Kamat A. M., Hahn N. M., Efstathiou J. A., et al. Bladder cancer. The Lancet . 2016;388(10061):2796–2810. doi: 10.1016/s0140-6736(16)30512-8. [DOI] [PubMed] [Google Scholar]
- 36.Sanli O., Dobruch J., Knowles M. A., et al. Bladder cancer. Nature Reviews Disease Primers . 2017;3(1):17022–17119. doi: 10.1038/nrdp.2017.22. [DOI] [PubMed] [Google Scholar]
- 37.Stein J. P., Lieskovsky G., Cote R., et al. Radical cystectomy in the treatment of invasive bladder cancer: long-term results in 1, 054 patients. Journal of Clinical Oncology . 2001;19(3):666–675. doi: 10.1200/jco.2001.19.3.666. [DOI] [PubMed] [Google Scholar]
- 38.Kwon H.-K., Jeon W. K., Hwang J.-S., et al. Cinnamon extract suppresses tumor progression by modulating angiogenesis and the effector function of CD8+ T cells. Cancer Letters . 2009;278(2):174–182. doi: 10.1016/j.canlet.2009.01.015. [DOI] [PubMed] [Google Scholar]
- 39.Zhang K., Han E. S., Dellinger T. H., et al. Cinnamon extract reduces VEGF expression via suppressing HIF‐1α gene expression and inhibits tumor growth in mice. Molecular Carcinogenesis . 2017;56(2):436–446. doi: 10.1002/mc.22506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Groenendijk F. H., De Jong J., Fransen Van De Putte E. E., et al. ERBB2 mutations characterize a subgroup of muscle-invasive bladder cancers with excellent response to neoadjuvant chemotherapy. European Urology . 2016;69(3):384–388. doi: 10.1016/j.eururo.2015.01.014. [DOI] [PubMed] [Google Scholar]
- 41.De Martino M., Zhuang D., Klatte T., et al. Impact of ERBB2 mutations on in vitro sensitivity of bladder cancer to lapatinib. Cancer Biology & Therapy . 2014;15(9):1239–1247. doi: 10.4161/cbt.29687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Chow N.-H., Chan S.-H., Tzai T.-S., Ho C.-L., Liu H.-S. Expression profiles of ErbB family receptors and prognosis in primary transitional cell carcinoma of the urinary bladder. Clinical Cancer Research: An Official Journal of the American Association for Cancer Research . 2001;7(7):1957–1962. [PubMed] [Google Scholar]
- 43.Yallowitz A., Ghaleb A., Garcia L., Alexandrova E. M., Marchenko N. Heat shock factor 1 confers resistance to lapatinib in ERBB2-positive breast cancer cells. Cell Death & Disease . 2018;9(6):621–713. doi: 10.1038/s41419-018-0691-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Tan M., Lan K.-H., Yao J., et al. Selective inhibition of ErbB2-overexpressing breast cancer in vivo by a novel TAT-based ErbB2-targeting signal transducers and activators of transcription 3-blocking peptide. Cancer Research . 2006;66(7):3764–3772. doi: 10.1158/0008-5472.can-05-2747. [DOI] [PubMed] [Google Scholar]
- 45.Li D., Marchenko N. D. ErbB2 inhibition by lapatinib promotes degradation of mutant p53 protein in cancer cells. Oncotarget . 2017;8(4):5823–5833. doi: 10.18632/oncotarget.12878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Yoshida K., Tsuda M., Matsumoto R., et al. Exosomes containing ErbB2/CRK induce vascular growth in premetastatic niches and promote metastasis of bladder cancer. Cancer Science . 2019;110(7):2119–2132. doi: 10.1111/cas.14080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Li J., Song P., Jiang T., et al. Heat shock factor 1 epigenetically stimulates glutaminase-1-dependent mTOR activation to promote colorectal carcinogenesis. Molecular Therapy . 2018;26(7):1828–1839. doi: 10.1016/j.ymthe.2018.04.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Tan M., Li P., Klos K. S., et al. ErbB2 promotes Src synthesis and stability: novel mechanisms of Src activation that confer breast cancer metastasis. Cancer Research . 2005;65(5):1858–1867. doi: 10.1158/0008-5472.can-04-2353. [DOI] [PubMed] [Google Scholar]
- 49.Soumya K. J., Choudhari A. S., Suryavanshi S. A., Kumari S., Chattopadhyay S., Kaul-Ghanekar R. Aqueous cinnamon extract (ACE-c) from the bark of Cinnamomum cassia causes apoptosis in human cervical cancer cell line (SiHa) through loss of mitochondrial membrane potential. BMC Cancer . 2010;10(1):210–212. doi: 10.1186/1471-2407-10-210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Gwang P. H., Song H. M., Park S. B., et al. Cytotoxic activity of the twigs of Cinnamomum cassia through the suppression of cell proliferation and the induction of apoptosis in human colorectal cancer cells. BMC Complementary and Alternative Medicine . 2018;18(1):28–13. doi: 10.1186/s12906-018-2096-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Choi Y. H. Trans-cinnamaldehyde protects C2C12 myoblasts from DNA damage, mitochondrial dysfunction and apoptosis caused by oxidative stress through inhibiting ROS production. Genes & Genomics. . 2021;43(4):303–312. doi: 10.1007/s13258-020-00987-9. [DOI] [PubMed] [Google Scholar]
- 52.Aminzadeh Z., Ziamajidi N., Abbasalipourkabir R. Anticancer effects of cinnamaldehyde through inhibition of ErbB2/HSF1/LDHAPathway in 5637 cell line of bladder cancer. Anti-Cancer Agents in Medicinal Chemistry . 2022;22(6):1139–1148. doi: 10.2174/1871520621666210726142814. [DOI] [PubMed] [Google Scholar]
- 53.Hyeon K., Park H.-J., Jung H.-J., et al. Cinnamaldehyde induces apoptosis by ROS-mediated mitochondrial permeability transition in human promyelocytic leukemia HL-60 cells. Cancer Letters . 2003;196(2):143–152. doi: 10.1016/s0304-3835(03)00238-6. [DOI] [PubMed] [Google Scholar]
- 54.Wang Z., Wang H., Wang Z., et al. Associated analysis of PER1/TUBB2B with endometrial cancer development caused by circadian rhythm disorders. Medical Oncology . 2020;37(10):90–11. doi: 10.1007/s12032-020-01415-4. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All data have sufficient accuracy and precision. All the reported results are obtained from the detailed analysis of the raw data obtained from the experiments.