

Abstract
Background CLEC-2 is a platelet receptor with an important role in thromboinflammation but a minor role in hemostasis. Two endogenous ligands of CLEC-2 have been identified, the transmembrane protein podoplanin and iron-containing porphyrin hemin, which is formed following hemolysis from red blood cells. Other exogenous ligands such as rhodocytin have contributed to our understanding of the role of CLEC-2.
Objectives To identify novel CLEC-2 small-molecule ligands to aid therapeutic targeting of CLEC-2.
Methods ALPHA screen technology has been used for the development of a high-throughput screening (HTS) assay recapitulating the podoplanin–CLEC-2 interaction. Light transmission aggregometry was used to evaluate platelet aggregation. Immunoprecipitation and western blot were used to evaluate direct phosphorylation of CLEC-2 and downstream protein phosphorylation. Autodock vina software was used to predict the molecular binding site of katacine and mass spectrometry to determine the polymeric nature of the ligand.
Results and Conclusion We developed a CLEC-2–podoplanin interaction assay in a HTS format and screened 5,016 compounds from a European Union-open screen library. We identified katacine, a mixture of polymers of proanthocyanidins, as a novel ligand for CLEC-2 and showed that it induces platelet aggregation and CLEC-2 phosphorylation via Syk and Src kinases. Platelet aggregation induced by katacine is inhibited by the anti-CLEC-2 monoclonal antibody fragment AYP1 F(ab)′2. Katacine is a novel nonprotein ligand of CLEC-2 that could contribute to a better understanding of CLEC-2 activation in human platelets.
Keywords: platelets, CLEC-2, high-throughput screening, small-molecule ligands, proanthocyanidin
Introduction
The C-type lectin-like platelet receptor, CLEC-2, is a novel target in thromboinflammatory diseases such as deep vein thrombosis and infection with a minimal role in hemostasis. 1 Hence, blocking agents are likely to cause less bleeding than current antiplatelets. The long-term targeting of CLEC-2 will be facilitated by development of a small molecule inhibitor that can be taken orally. 2
CLEC-2 is a single transmembrane receptor that is activated by multivalent ligands 3 4 leading to Src and Syk kinase phosphorylation of the single YxxL motif (hemITAM) in the CLEC-2 tail. This is followed by binding of Syk through its SH2 domains and crosslinking of two receptors. In turn, Syk promotes downstream signaling leading to phosphorylation of LAT and Btk/Tec tyrosine kinases. The signaling cascade culminates in the activation of PI 3-kinase and PLCγ2. 5
To date, only a handful of naturally occurring ligands have been shown to induce platelet aggregation via CLEC-2. The first was the snake venom rhodocytin which was used for its affinity purification. 6 Later, podoplanin was identified as an endogenous ligand. 6 7 Since then, the iron-containing porphyrin, hemin, which is formed from haem released from red blood cells, has been described as a CLEC-2 agonist. 8 9
Crystallography studies of rhodocytin and podoplanin in complex with CLEC-2 suggested a canonical binding site rich in arginine residues (Arg-107, Arg-118, Arg-152, Arg-157). 10 11 Podoplanin binding requires a carbohydrate motif but this is not the case for rhodocytin. Two small molecule antagonists of CLEC-2 have been identified, cobalt-hematoporphyrin and 2CP, with modeling suggesting that the former binds to an allosteric site and the latter to the four arginines. 12 13 The dependency on the four charged residues has led to the hypothesis that CLEC-2 may be a pattern recognition receptor (PRR) on platelets. 14
The identification of small-molecule ligands for CLEC-2 is essential for the development of orally available inhibitors as this is the preferred route of delivery for long-term antithrombotics. To address this objective, we set up a high-throughput screening (HTS) assay using a small-molecule library based on the podoplanin–CLEC-2 interaction. The screen identified the proanthocyanidin katacine as a novel inhibitor of this interaction but, when tested on platelets, katacine was found to activate platelets identifying it as the first exogenous small-molecule agonist.
Methods
Materials
Katacine was purchased from TargetMol (Boston, Massachusetts, United States), and recombinant human podoplanin-rFc and recombinant human CLEC-2–6xHis (Ser55-Pro229) were expressed in house HEK293T cells. Recombinant human CLEC-2 was also purchased from R&D Systems (Minnesota, United States) for optimization of the screening. Rhodocytin was purified from crude venom of Calloselasma rhodostoma as described previously. 15 Collagen-related peptide (CRP) was purchased from Cambcol Laboratories (Cambridge, United Kingdom). PP2 was from Sigma-Aldrich (Dorset, United Kingdom), and PRT-060318 was from ApexBio (Houston, Texas, United States). AYP1 was expressed and purified from hybridoma cells as previously reported. 16 Streptavidin donor beads, nickel-coated acceptor beads, and TruHits kit were purchased from Perkin Elmer (Waltham, United States). Rabbit anti-phospho-Syk (Y525/526), rabbit anti-FcR gamma, and rabbit anti-phospho LAT(Y200) were from Cell Signaling (Massachusetts, United States); Monoclonal phosphotyrosine antibody (clone 4G10) was from Upstate Biotechnologies (Lake Placid, New York, United States); and anti-CLEC-2 antibody (AF1718) was from R&D system (Minneapolis, United States).
For the HTS we have used the European Chemical Biology Library, a pilot library, constituted of 5,016 compounds, containing 2,464 bioactive molecules active against 1,039 different targets, and containing 654 approved drugs and 368 highly selective probes. A total of 2,464 representative compounds of the diversity library and 88 assay interference compounds in four dilutions were used.
Protein Expression and Purification
For the recombinant human CLEC-2–6xHis tag and human podoplanin-rabbit Fc vector overexpression, the extracellular domain of the cDNA of human CLEC-2 and podoplanin were cloned into a pHLSEC (Addgene) and pFUSE (Invivogen) expression vectors. The constructs were used to transient transfection into human embryonic kidney HEK293T cells. Cells were grown in complete Dulbecco's modified eagle medium (DMEM; supplemented with 10% fetal bovine serum, glutamine 5%, and 100 U/mL of penicillin/streptomycin). Once 60% of cell confluence was achieved, the medium was exchanged for serum-free DMEM and transfected using the PEI method. Proteins secreted into the medium and were collected after 4 days. hCLEC-2–6xHis tag and h-podoplanin-rFc were purified using a Nickel-NTA Resin (Thermo Fisher) and protein A-coated beads (Thermo Fisher), respectively, in a gravity purification column. SDS-PAGE (sodium dodecyl sulfate–polyacrylamide gel electrophoresis) was used to characterize proteins. Protein coding sequences of recombinant proteins can be found in the Supplementary Information.
HTS Based on ALPHA Screen Assay
Each of the 5,016 chemicals from the chemical library (0.1% [v/v] DMSO for the vehicle controls) were dispensed into 1,536-well flat and transparent bottom white plates (Corning 3725) (Merck, Darmstadt, Germany) using an Echo 525 Liquid Handler (Labcyte, Indiana, United States). Followed by the addition of 50 nM of biotinylated h-podoplanin-rFc and 250 nM of h-CLEC-2–6xHis, the reagents were incubated for 30 minutes at 4°C. Then, 40 µg/mL of ALPHA screen streptavidin donor beads was added and incubated for 30 minutes at room temperature in the dark, followed by an additional incubation with 40 µg/mL nickel-coated acceptor beads. The plates were incubated in the dark for 60 minutes at room temperature and read using an EnVision Multimode Plate Reader (Perkin Elmer, Waltham, United States). AlphaScreen TruHits kits were used to identify false-positive results from the first screen (Perkin Elmer, Waltham, United States). Different concentrations of the potential ligands were used to estimate IC50 values.
Blood Collection and Platelet Isolation
Fresh blood was collected into 4% sodium citrate tubes from consenting healthy volunteers who had not taken antiplatelets in the previous 14 days. Washed human platelets were obtained by centrifugation of platelet-rich plasma in the presence of 0.2 μg/mL prostacyclin and resuspended in modified Tyrode's buffer (134 mM NaCl, 0.34 mM Na 2 HPO 4 , 2.9 mM KCl, 12 mM NaHCO 3 , 20 mM HEPES, 5 mM glucose, 1 mM MgCl 2 , pH 7.3) and allowed to rest for 30 minutes at room temperature.
Light Transmission Platelet Aggregation
Washed platelets at 2 × 10 8 /mL were stimulated with katacine (1–30 µM) in a ChronoLog model 700 aggregometer (ChronoLog, Havertown, Pennsylvania, United States) while stirring at 1,200 rpm at 37°C. Platelets were pretreated with PP2 (20 µM), PRT-060318 (1 µM), and AYP1 (10 µg/mL), or DMSO (0.1% [v/v]) as vehicle control for 5 minutes at 37°C and then stimulated by katacine (30–1 µM).
Phosphorylation Studies
Washed platelets at 4 × 10 8 /mL in the presence of 9 µM eptifibatide were stimulated with 10 µM katacine, 100 nM rhodocytin, or vehicle (DMSO 0.1%) for 5 minutes, while stirring at 1,200 rpm at 37°C, then lysed with 5× SDS buffer (10% [w/v] SDS, 0.5 M DTT, 50% glycerol, 0.125 M Tris). Samples were boiled for 5 minutes at 100°C and proteins were separated by SDS-polyacrylamide gel electrophoresis and subsequent immunoblotting using polyvinylidene difluoride membranes. After blocking with 5% (w/v) BSA in TBS-Tween, the membranes were incubated using the primary antibodies, as stated.
Immunoprecipitation
Washed platelets at 4 × 10 8 /mL in the presence of 9 µM eptifibatide were stimulated with 10 µM katacine, 100 nM rhodocytin, or vehicle for 5 minutes, while stirring at 1,200 rpm at 37°C, then lysed with 2× NP-40 lysis buffer in the presence of protease inhibitors (10 μg/mL aprotinin;1 μg/mL pepstatin; 10 μg/mL leupeptin; 400 μg/mL, AEBFS; 5mM sodium orthovanadate; pH 7.5). Whole lysates were precleared using Gamma bind Plus Sepharose (Cytiva, United States) and then incubated with 2 µg of AYP1 antibody for 30 minutes, and lysate–antibody complexes were then incubated for 60 minutes with Gamma bind Plus Sepharose. Immunoprecipitated proteins were resuspended on SDS lysis buffer. Further SDS-PAGE and western blotting were performed against 4G10 monoclonal antibody.
Computational Binding-Site Predictions
AutoDock tools (ADT) was used for protein and ligand preparation and binding site prediction was done using Autodock vina. 17 We used the three-dimensional (3D) crystal structure of the extracellular domain of human CLEC-2 (PDB: 2C6U) downloaded from RCSB and the 3D structure of katacine was obtained using Chemdraw software. Visualization and binding site analysis were done using PyMOL software.
Mass Spectrometry
A 10 µM of katacine resuspended in 200 mM ammonium acetate buffer was infused into a Q-Exactive HF mass spectrometer (ThermoFisher, California, United States) via nanoelectrospray ionization) using a Triversa NanoMate (Advion Biosciences, Ithaca, New York, United States) set at 1.4 kV with 0.30 psi backing pressure. The spectra were collected using source temperature of 250°C and collision energy of 200V for desolvation and with the resolution setting at 120 K (∼25,000 full-width half-maximum at m / z 1,000).
Data Analysis
Data are presented as mean ± standard deviation of the mean with statistical significance taken as p <0.05 unless otherwise stated. Statistical analysis was performed using the Kruskal–Wallis one-way ANOVA (analysis of variance) test as stated. All statistical analyses were performed using GraphPad Prism 7 (GraphPad Software Inc., La Jolla, United States). IC50 values were calculated also using GraphPad Prism 7.
Results
Identification of Katacine as a CLEC-2 Ligand by HTS
To identify novel CLEC-2 ligands, we used a podoplanin–CLEC-2 interaction assay based on ALPHA screen technology that was further miniaturized to a HTS format ( Fig. 1A ). In this assay, a high signal indicates an interaction between the targets. We identified 40 hits from a library of 5,016 compounds able to interrupt podoplanin and CLEC-2 interaction ( Fig. 1B-i ). Evaluating false positives using the TruHits Kit reduced the number of hits to 17 ( Fig. 1B-ii ). The compounds were ranked according to their potency with the two most potent compounds, sennoside A (IC50 1.3 µM) ( Fig. 1C-i ) and katacine (IC50 2.7 µM) ( Fig. 1C-ii ) selected for functional studies using light transmission aggregometry. The other less potent compounds ( Fig. 1C[iii–vi] ) were not tested. Sennoside A (10 and 30 µM) has no effect on platelet aggregation or the response to rhodocytin (100 nM; data not shown). On the other hand, katacine (10 µM) stimulated rapid and full platelet aggregation (77.7 ± 9%) as shown in Fig. 2A .
Fig. 1.
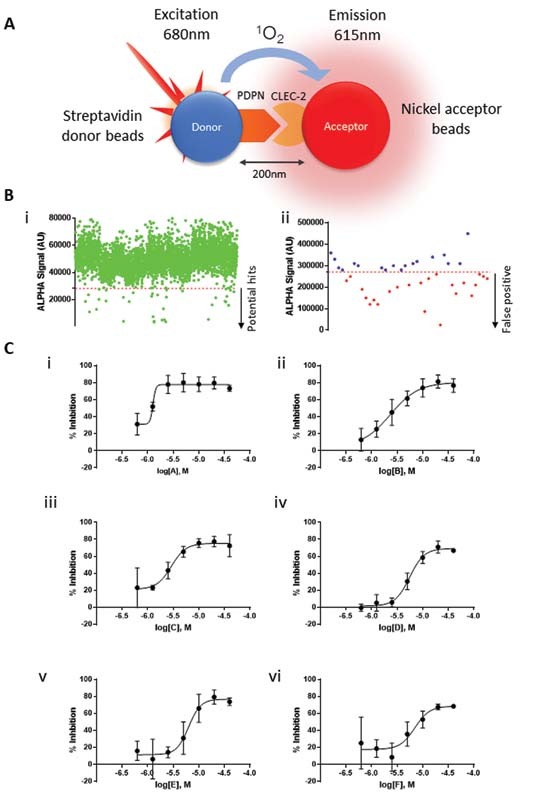
H igh-throughput screening based on CLEC-2 and podoplanin interaction. ( A ) Schematic representation of ALPHA screen based on podoplanin and CLEC-2 interaction. ALPHA screen assay is based on the proximity of the donor and acceptor beads mediated by the protein–protein interaction. After laser excitation, podoplanin-conjugated donor beads transform ambient oxygen to singlet oxygen, which is absorbed by CLEC-2-conjugated nickel acceptor beads, then ALPHA signal is emitted and detected by the plate reader. ( B-i ) Scatter plot showing the effect of each compound on the ALPHA signal values emitted by CLEC-2 and podoplanin interaction; dots below the cut off ( red line ) represent compounds that significantly decreased signal and potential hits. ( B-ii ) Scatter plot showing TruHits assay to identify potential false-positive compounds on the screening; dots over the cut-off indicate potential hits. ( C ) The dose–response curves of potential hits and IC50 of each potential compound (i–vi). C-i and C-ii show the dose–response curves of the two most potent compounds, further evaluated them in functional assays. Supplementary Table S1 shows a chemical description of the compounds tested.
Fig. 2.
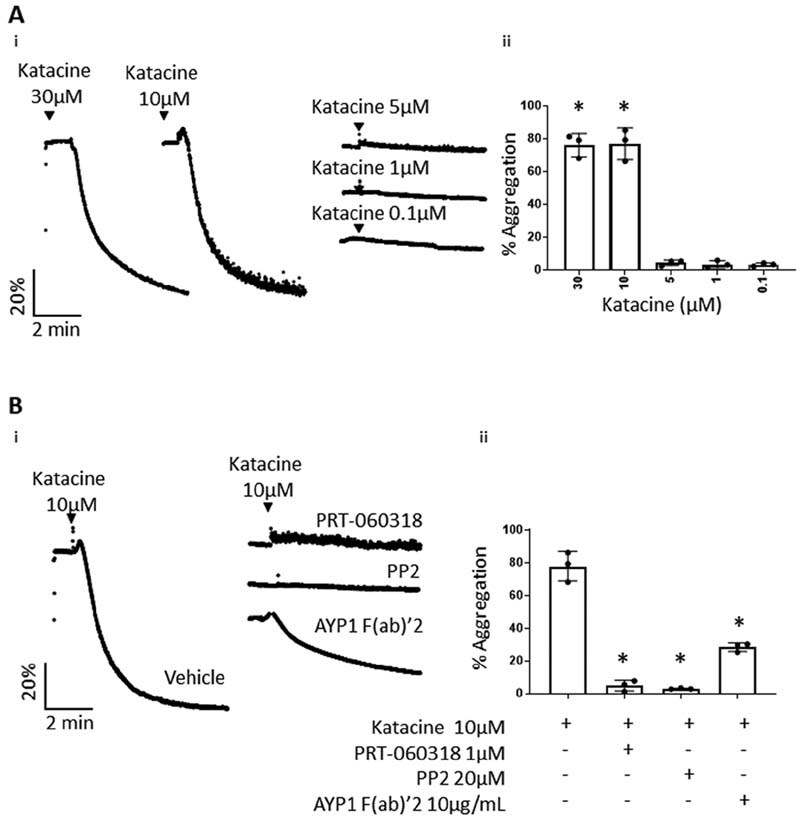
Katacine induces platelet aggregation. ( A-i ) Representative traces of washed platelets (2 × 10 8 platelets/mL) stimulated by different doses of katacine (0.1–10 µM) and panel ( A-ii ) represents bar columns of the mean + SD of three independent experiments. ( B-i ) Representative traces of washed platelets pretreated by the Syk and Src inhibitors, PRT-060318 at 1 µM and PP2 at 20 µM, respectively, or by the mouse anti-CLEC-2 antibody, AYP1 (10 µg/mL), and panel ( B-ii ) shows the mean ± SD of three independent experiments. The Kruskal–Wallis one-way ANOVA test was used to evaluate significant differences between the treatment in relation with the control; the symbol “*” represents significance differences where p < 0.05, as stated. ANOVA, analysis of variance; SD, standard deviation.
These results illustrate the power of the ALPHA screen to detect novel ligands for CLEC-2 and reveal katacine as a novel platelet agonist.
Katacine Activates CLEC-2 via Syk- and Src-Dependent Mechanism
We first determined the concentration–response curve for katacine. We observed maximal aggregation at 10 and 30 μМ, but no response at 5 μM and below ( Fig. 2A ). To investigate if activation is mediated through Src and Syk tyrosine kinases, platelets were pretreated with the Syk inhibitor PRT-060318 (1 µM) and Src inhibitor, PP2 (20 µM). Both inhibitors completely blocked platelet aggregation induced by 10 µM katacine ( Fig. 2B ). Aggregation was also partially reduced to 28.3% ± 2.6 by the anti-CLEC-2 monoclonal antibody fragment AYP1 F(ab)′2 (10 μg/mL) demonstrating that activation is partially mediated by CLEC-2 ( Fig. 2B ). In line with these results, katacine (10 µM) stimulated a similar increase in protein tyrosine phosphorylation in platelet lysates to an intermediate dose of rhodocytin (100 nM). Using phospho-specific antibodies, katacine was shown to stimulate a similar or slightly smaller increase in phosphorylation of Syk Y525/526 (4.5 ± 0.9-fold) and LAT Y200 (18.8 ± 5.9-fold) compared with rhodocytin (100 nM) (4.5 ± 1.0 and 41.5 ± 4.5-fold, respectively) relative to vehicle ( Fig. 3A[ii–iii] ). Moreover, katacine induced a marked increase in phosphorylation levels of CLEC-2 (6.6 ± 4.6-fold) relative to platelets treated with the vehicle. A similar level of increase in phosphorylation of CLEC-2 was observed with rhodocytin (6.1 ± 4.6-fold) ( Fig. 3B[i–ii] ). These results demonstrate that katacine is a novel ligand of CLEC-2 that stimulates activation of platelets through Src and Syk tyrosine kinases. The partial inhibitory effect of the CLEC-2 AYP1 F(ab)′2 relative to the full inhibition induced by the Src and Syk inhibitors indicates that katacine binds to more than one site on CLEC-2 or to another platelet glycoprotein receptor that signals through the same two kinases.
Fig. 3.
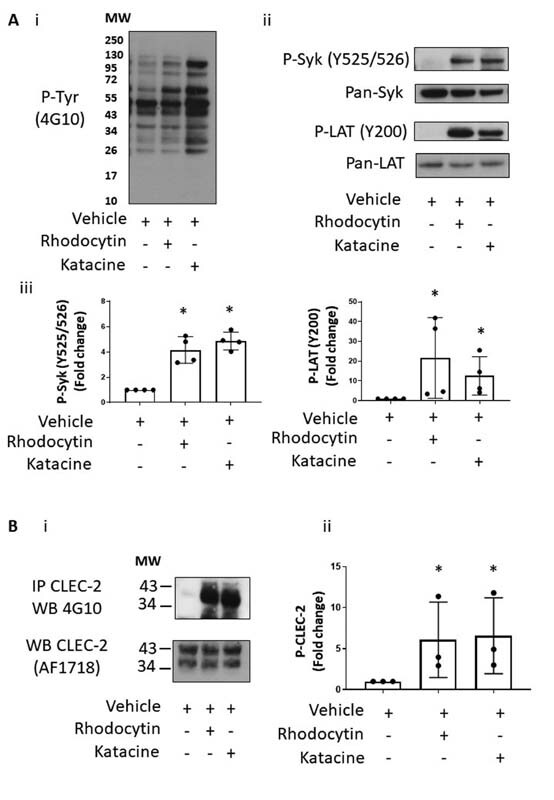
Katacine induces phosphorylation of CLEC-2 and key signaling protein on CLEC-2 signaling pathway. ( A ) Representative immunoblots of platelet lysates (4 × 10 8 ) stimulated with DMSO 0.1% (negative control), rhodocytin at 100 nM (positive control) or katacine (10 µM). (A-i) Tyrosine phosphorylation profiles in platelets stimulated with katacine or rhodocytin have similar profile. (A-ii) An increase of the phosphorylation levels of Syk (Y525/526) and LAT (Y200) in platelets treated with rhodocytin 100 nM and katacine 10 µM is shown compared with the DMSO 0.1% control. (A-iii) The mean and SD of four independent experiments. ( B ) A representative blot of an immunoprecipitation of CLEC-2 from lysed platelets (4 × 10 8 platelets/mL) and probed against the tyrosine phosphorylation antibody (4G10), demonstrating a significant increase on the CLEC-2 phosphorylation levels in platelets stimulated with 100 nM rhodocytin or 10 µM katacine compared with the DMSO 0.1% control. (B-i) The mean ± SD values from three independent experiments. The Kruskal–Wallis one-way ANOVA test was used to evaluate significant differences between the treatment in relation with the control; the symbol “*” represents significance difference where p < 0.05, as stated. SD, standard deviation.
To explore the specificity of katacine for CLEC-2, we investigated if it also induces phosphorylation of the FcR γ-chain which is expressed as a complex with glycoprotein VI (GPVI). Katacine did not stimulate an increase in phosphorylation of FcR γ-chain as measured by western blotting in a whole cell lysate, in contrast to a marked increased by CRP (3 µg/mL) ( Supplementary Fig. S1 , available in the online version).
Katacine Is a Mixture of Polymers with Different Sizes
Proanthocyanidins are known to polymerize which may explain why katacine is an agonist rather than an antagonist, as expected for a monomeric ligand. We used the monomeric structure of the proanthocyanidin, flavan-3-ol (C 15 H 14 O 7; MW: 306Da), which is the basic unit of katacine, to calculate the size of oligomers of katacine ( Fig. 4 ). Using mass spectrometry, we identified the minimum size of katacine as a trimer of ∼914 Da ( Fig. 5A ), as well as the presence of tetramers (∼1,218 Da), pentamers (∼1,522 Da), hexamers (∼1,826 Da), and higher order species up to dodecamers (∼3,660 Da) ( Fig. 5A ). The mass spectra also indicate that trimers and hexamers of katacine are sodiated and potassiated ( Fig. 5B–E ), suggesting that medium-sized oligomeric species have a polar nature. The negative charge of katacine could be required for the interaction with the positively charged surface of the extracellular CLEC-2 domain.
Fig. 4.
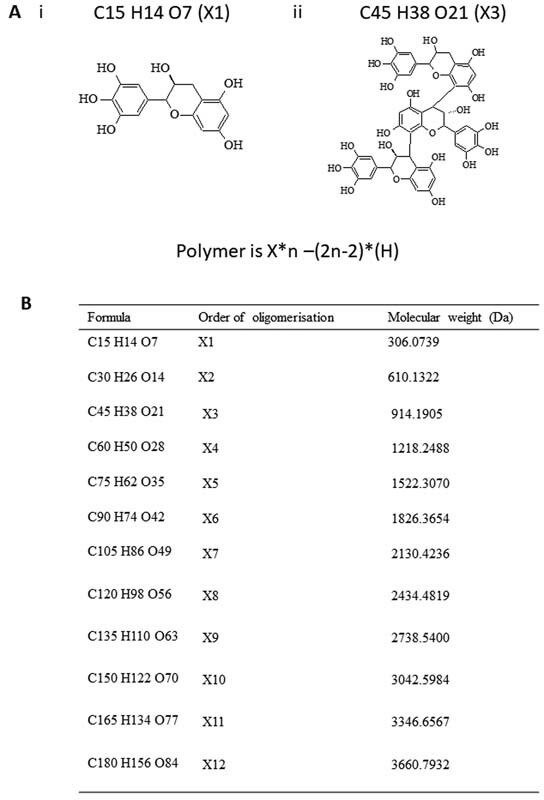
Different oligomeric conformations of katacine. ( A ) Theoretical monomeric form of katacine (flavan-3-ol) (i) and the minimum oligomeric size observed identified for katacine (trimer). Theoretical masses of the different polymers of katacine are calculated based on the equation X * n –(2 n -2)*( H ), where X is the mass of the monomer and n is the size of the polymer. ( B ) The theoretical mass of the different order of oligomerization derived from katacine.
Fig. 5.
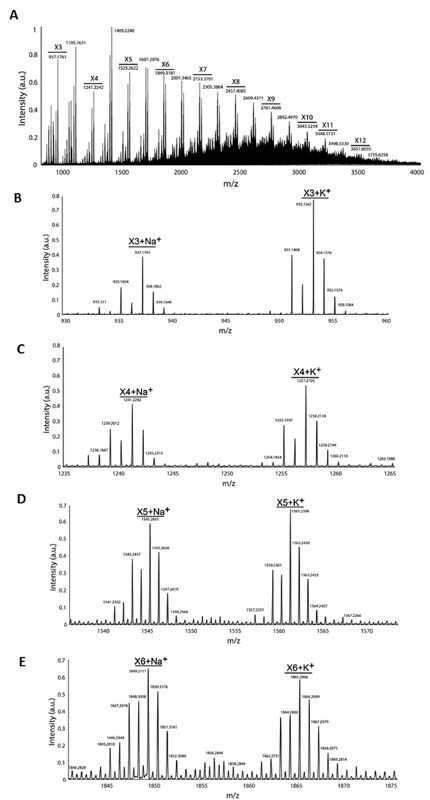
Katacine is a mixture of polymers with different sizes. 10 µM of katacine resuspended in 200 mM ammonium acetate was injected into a HPLC for separation and electrosprayed into an Orbitrap mass spectrometer in Intact protein mode for mass spectrometry analysis. ( A ) The global mass spectra of katacine, X1–X12 indicate the size of the polymer of katacine. A decrease on the intensity of the signal was observed with the size of polymer. m / z indicates the mass of the species. The size of the oligomers was calculated based on the monomeric structure of flavan-3-ol. ( B–E ) The zoomed-in peaks belonging to trimers (B), tetramers (C), pentamers (D), and hexamer (E) in their sodiated and potassiated forms. HPLC, high-performance liquid chromatography.
Molecular Docking Studies Predict that Katacine Binds to the Same CLEC-2 Binding Site as Podoplanin
We used computational docking studies to predict the binding site of katacine. This suggested that katacine may bind to Arg-117, Arg-118, Arg-152, and Arg-157 on the extracellular domain of CLEC-2, as is the case for rhodocytin and podoplanin ( Fig. 6 ). In addition, the prediction identifies a second site to residues Asn-194 and Ser-166 on the opposite side of the canonical binding site ( Supplementary Fig. S2 , available in the online version). These data may suggest the possible existence of an allosteric binding site for CLEC-2 activation as has been reported for cobalt-hematoporphyrin. 12 The estimated binding affinities of these two conformations (−6.6 and −6.9 kcal/mol, respectively) are similar.
Fig. 6.
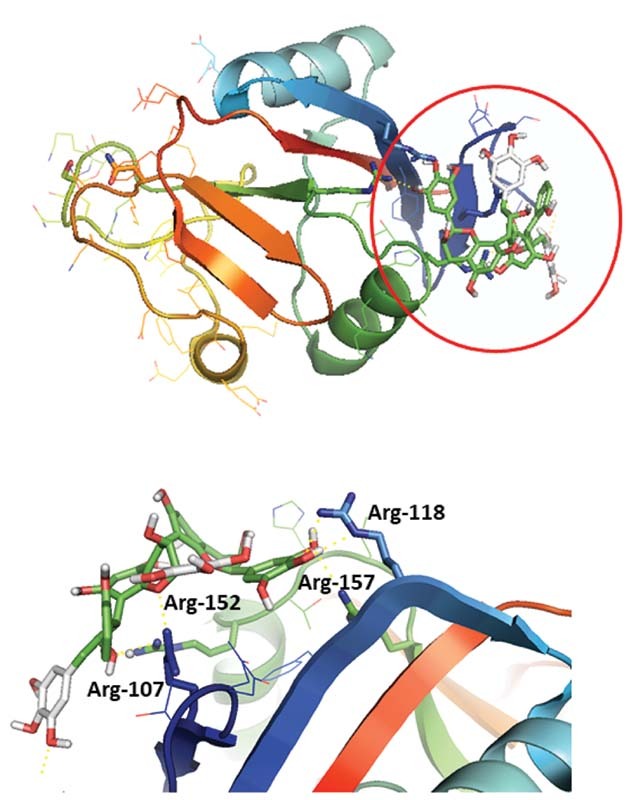
Katacine binds to the same binding site as rhodocytin based on molecular binding site prediction. Molecular docking has been performed using AutoDock vina and ADT for protein preparation. Grid boxes for binding prediction have covered the extracellular domain of CLEC-2, using the crystal structure of human CLEC-2 (PDB: 2C6U). Katacine can bind to a positive charge surface rich of arginine in the CLEC-2 extracellular domain (Arg-107, Arg-118, Arg-152, Arg-157) previously reported as the binding site for podoplanin and rhodocytin (enclosed in a red circle ), with a predicted binding energy of −6.6 kcal/mol. ADT, AutoDock tools.
Discussion
This study revealed that katacine induces platelet activation through Src- and Syk-mediated phosphorylation of CLEC-2 in a similar way to the heterodimeric C-type lectin ligand, rhodocytin. As for rhodocytin, we observed that platelet aggregation induced by katacine occurs in an all-or-none response mechanism, 18 with activation inhibited by the AYP1 F(ab)′2. Thus, katacine is the first exogenous small-molecule-based ligand of CLEC-2 which acts as a platelet agonist.
Katacine is a proanthocyanidin extracted from the polygonaceae family of flowering plants known as the knotweed family. These forms of polyphenols are highly distributed in the Plantae kingdom as oligomers or polymers and most have antioxidant properties and have significant affinity for protein binding. 19 This is the first report describing a member of this family as a ligand of CLEC-2.
Proanthocyanidin polymers of katacine are planar, negatively charged structures raising a possible interaction with the arginine-rich residues on CLEC-2 which mediate binding to rhodocytin and podoplanin. Indeed, molecular docking predicts that katacine (trimer) has the potential to bind to the same site on CLEC-2 as that for podoplanin or rhodocytin. Additionally, our predictions show a potential binding on Asn-194 and Ser-166 on the C-type lectin-like domain, suggesting a possible allosteric binding site. The recognition of this ligand pattern may aid the design of more potent ligands. The co-crystallization of katacine and CLEC-2 is required to map the binding site(s). Furthermore, an additional binding site may explain why AYP1 is not able to fully block platelet aggregation induced by katacine, considering that AYP1 may bind to the canonical binding site of rhodocytin.
To gain further knowledge on the above, we extended our studies to a mouse model; however, we did not observe platelet aggregation or an increase in protein tyrosine phosphorylation levels induced by katacine in the murine model ( Supplementary Fig. S3 , available in the online version). This may be due to the lack of homology between human and mouse CLEC-2 (62%). 4 These data may be in line with the hypothesis of katacine binding to an allosteric binding site since the amino acid residues adjacent to S166 in this region are not conserved between human CLEC-2 (QK S 16 6 NEV) and mouse CLEC-2 (QN S KKD).
CLEC-2 activation requires clustering of the receptor as recently reviewed. 20 This mechanism explains why podoplanin, which is expressed as multiple copies on a cell surface, and rhodocytin, which is an oligomer, stimulate platelet activation. In line with this, we have shown that katacine is a mixture of polymers as is the case for other proanthocyanidins. 19 This suggests a model in which katacine could promote oligomerization of CLEC-2, inducing activation.
We suspect that katacine may bind to other glycoprotein receptors apart from CLEC-2 since we observed a partial inhibition of platelet aggregation by AYP1 but full inhibition by inhibitors of Syk and Src kinases. Alternatively, this could reflect binding to two sites on CLEC-2. GPVI and FcγRIIA are the other two ITAM receptors on platelets that signal through Src and Syk tyrosine kinases. Recently, Montague et al (2021) have proposed the three platelet ITAM receptors as PRRs as they are activated by a miscellaneous group of polyvalent charged ligands. 14 The charged nature of katacine is consistent with this mechanism. However, we did not observe clear phosphorylation on the FcR gamma chain (∼15 kDa) on whole lysates when treated with katacine, while a marked increase on the phosphorylation was observed when platelets were treated with the GPVI ligand, CRP. This finding indicates a degree of specificity of katacine for CLEC-2 and excludes the possibility that katacine is acting by a nonspecific tyrosine phosphorylation mechanism. 21
In contrast, the most potent agent identified in the HTS, sennoside A, does not alter platelet function. The ALPHA screen identifies disruptors of the podoplanin and CLEC-2 interaction and does not discriminate between agents interacting with CLEC-2 or podoplanin. We therefore speculate that it may bind to podoplanin rather than to CLEC-2. The other ligands identified were not tested due to their lower potency.
In summary, we have identified katacine as a novel CLEC-2 ligand that activates platelets by a Syk- and Src-dependent mechanism. This opens a new field to investigate if nonpolymeric forms of proanthocyanidins may represent a novel scaffold for the rational development of CLEC-2 antagonists.
Acknowledgement
The authors would like to thank Ms. Rocío Piña, from the BioFarma research group, for her technical assistance during the high-throughput screening and automatization of the HTS. We also thank to Dr. Lourdes Garcia for the advises on CLEC-2 inmunoprecipitation experiments and the personnel from the Servizo de Vixilancia da Saúde, Universidade de Santiago de Compostela, and Dr. Beata Grygielska, University of Birmingham, for the assistance on blood collection from healthy volunteers.
Funding Statement
Funding L.A.M. and M.S. are supported by the European Union's Horizon 2020 Research and Innovation Program (Marie Sklodowska-Curie grant agreement No. 766118); S.P.W. is a British Heart Foundation Chair (CH03/003); and E.M. is supported by the Wellcome Trust (204951/Z/16/Z). A.G. is supported by the Spanish Ministry of Science and Innovation (Grant No. PID2019–108727RB-I00). J.A.E. is financially supported by the Interdisciplinary Center of Clinical Research (IZKF) of the University of Münster (grant no. Ebl-A/009/21). L.H-.N. receives financial support from the Consellería de Cultura, Educación e Ordenación Universitaria, Xunta de Galicia (Centro Singular de investigación de Galicia accreditation 2019–2022, ED431G 2019/02; predoctoral grant 2018 Call).
Conflict of Interest None declared.
Informed Consent Statement
Informed consents were obtained from all the individuals involved in the study under the local ethics ERN_11–0175: the regulation of activation of platelets; and also following the approval by the Galician Investigation Ethics Committee (2009/270). The study was developed according to the principles outlined in the Declaration of Helsinki. Mouse experiments were performed in accordance with the UK law (Animal Scientific Procedures Act 1986) with approval of the local ethics committee and UK Home Office approval under PP9677279 to the University of Birmingham.
Author Contributions
L.A.M. performed and designed the experiments, analyzed the data, wrote and edited the manuscript. Y.D., M.A.S., L.H-.N., M.N.B., E.M., J.C.C., T.H.M., and D.M. performed the experiments; M.I.L. provided key reagents and analytical tools; and A.Y.P. provided supervision and edited the manuscript. E.D. designed the study, analyzed the data, and wrote the manuscript; S.P.W and A.G. provided supervision, funding, designed the study, reviewed data, and wrote and edited the manuscript. J.A.E. purified and provided rhodocytin.
Current affiliation: Vascular Biology Program, Boston Children's Hospital, Harvard Medical School, Boston, United States.
What is known about this topic?
CLEC-2 is a promising target for the development of novel antiplatelet agents due to its minor role during hemostasis and relevant impact on thrombus formation and deep vein thrombosis.
CLEC-2 requires clustering for its activation, but the precise way this is done is not fully understood yet. The research on new CLEC-2 ligands may expand the understanding of the mechanisms of activation of CLEC-2.
Few agonists for CLEC-2 have been described so far, including the protein snake venom rhodocytin, the glycopeptide podoplanin, and the coordinated ring-like compound heme.
What does this paper add?
We present ALPHA screen as a potential tool to investigate new disruptors of CLEC-2 and podoplanin interaction. This is a quick tool to screen thousands of compounds in a selective and robust biochemical assay for the development of new agonists or antagonists of CLEC-2.
We have identified katacine, a mixture of oligomers with different sizes, to be able to induce human platelet aggregation, and phosphorylation of the CLEC-2 receptor. The new agonist also suggests an interesting scaffold for the development of further nonpolymeric ligands of CLEC-2 with potential antagonist effect.
Supplementary Material
References
- 1.Payne H, Ponomaryov T, Watson S P, Brill A. Mice with a deficiency in CLEC-2 are protected against deep vein thrombosis. Blood. 2017;129(14):2013–2020. doi: 10.1182/blood-2016-09-742999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Damaskinaki F N, Moran L A, Garcia A, Kellam B, Watson S P. Overcoming challenges in developing small molecule inhibitors for GPVI and CLEC-2. Platelets. 2021;32(06):744–752. doi: 10.1080/09537104.2020.1863939. [DOI] [PubMed] [Google Scholar]
- 3.Watson A A, Christou C M, James J R. The platelet receptor CLEC-2 is active as a dimer. Biochemistry. 2009;48(46):10988–10996. doi: 10.1021/bi901427d. [DOI] [PubMed] [Google Scholar]
- 4.Martin E M, Zuidscherwoude M, Morán L A, Di Y, García A, Watson S P. The structure of CLEC-2: mechanisms of dimerization and higher-order clustering. Platelets. 2021;32(06):733–743. doi: 10.1080/09537104.2021.1906407. [DOI] [PubMed] [Google Scholar]
- 5.Izquierdo I, Barrachina M N, Hermida-Nogueira L. A comprehensive tyrosine phosphoproteomic analysis reveals novel components of the platelet CLEC-2 signaling cascade. Thromb Haemost. 2020;120(02):262–276. doi: 10.1055/s-0039-3400295. [DOI] [PubMed] [Google Scholar]
- 6.Suzuki-Inoue K, Fuller G L, García A. A novel Syk-dependent mechanism of platelet activation by the C-type lectin receptor CLEC-2. Blood. 2006;107(02):542–549. doi: 10.1182/blood-2005-05-1994. [DOI] [PubMed] [Google Scholar]
- 7.Suzuki-Inoue K, Kato Y, Inoue O. Involvement of the snake toxin receptor CLEC-2, in podoplanin-mediated platelet activation, by cancer cells. J Biol Chem. 2007;282(36):25993–26001. doi: 10.1074/jbc.M702327200. [DOI] [PubMed] [Google Scholar]
- 8.Bourne J H, Colicchia M, Di Y. Heme induces human and mouse platelet activation through C-type-lectin-like receptor-2. Haematologica. 2021;106(02):626–629. doi: 10.3324/haematol.2020.246488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Manne B K, Getz T M, Hughes C E. Fucoidan is a novel platelet agonist for the C-type lectin-like receptor 2 (CLEC-2) J Biol Chem. 2013;288(11):7717–7726. doi: 10.1074/jbc.M112.424473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Watson A A, Brown J, Harlos K, Eble J A, Walter T S, O'Callaghan C A. The crystal structure and mutational binding analysis of the extracellular domain of the platelet-activating receptor CLEC-2. J Biol Chem. 2007;282(05):3165–3172. doi: 10.1074/jbc.M610383200. [DOI] [PubMed] [Google Scholar]
- 11.Watson A A, Eble J A, O'Callaghan C A. Crystal structure of rhodocytin, a ligand for the platelet-activating receptor CLEC-2. Protein Sci. 2008;17(09):1611–1616. doi: 10.1110/ps.035568.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Tsukiji N, Osada M, Sasaki T. Cobalt hematoporphyrin inhibits CLEC-2-podoplanin interaction, tumor metastasis, and arterial/venous thrombosis in mice. Blood Adv. 2018;2(17):2214–2225. doi: 10.1182/bloodadvances.2018016261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chang Y W, Hsieh P W, Chang Y T. Identification of a novel platelet antagonist that binds to CLEC-2 and suppresses podoplanin-induced platelet aggregation and cancer metastasis. Oncotarget. 2015;6(40):42733–42748. doi: 10.18632/oncotarget.5811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Montague S J, Patel P, Martin E M. Platelet activation by charged ligands and nanoparticles: platelet glycoprotein receptors as pattern recognition receptors. Platelets. 2021;32(08):1018–1030. doi: 10.1080/09537104.2021.1945571. [DOI] [PubMed] [Google Scholar]
- 15.Eble J ABB, Beermann B, Hinz H J, Schmidt-Hederich A. alpha 2beta 1 integrin is not recognized by rhodocytin but is the specific, high affinity target of rhodocetin, an RGD-independent disintegrin and potent inhibitor of cell adhesion to collagen. J Biol Chem. 2001;276(15):12274–12284. doi: 10.1074/jbc.M009338200. [DOI] [PubMed] [Google Scholar]
- 16.Gitz E, Pollitt A Y, Gitz-Francois J J. CLEC-2 expression is maintained on activated platelets and on platelet microparticles. Blood. 2014;124(14):2262–2270. doi: 10.1182/blood-2014-05-572818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Trott O, Olson A J. AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J Comput Chem. 2010;31(02):455–461. doi: 10.1002/jcc.21334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Shin Y, Morita T. Rhodocytin, a functional novel platelet agonist belonging to the heterodimeric C-type lectin family, induces platelet aggregation independently of glycoprotein Ib. Biochem Biophys Res Commun. 1998;245(03):741–745. doi: 10.1006/bbrc.1998.8516. [DOI] [PubMed] [Google Scholar]
- 19.Rue E A, Rush M D, van Breemen R B. Procyanidins: a comprehensive review encompassing structure elucidation via mass spectrometry. Phytochem Rev. 2018;17(01):1–16. doi: 10.1007/s11101-017-9507-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Pollitt A Y, Poulter N S, Gitz E. Syk and Src family kinases regulate C-type lectin receptor 2 (CLEC-2)-mediated clustering of podoplanin and platelet adhesion to lymphatic endothelial cells. J Biol Chem. 2014;289(52):35695–35710. doi: 10.1074/jbc.M114.584284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gibbins J, Asselin J, Farndale R, Barnes M, Law C L, Watson S P. Tyrosine phosphorylation of the Fc receptor gamma-chain in collagen-stimulated platelets. J Biol Chem. 1996;271(30):18095–18099. doi: 10.1074/jbc.271.30.18095. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.