

Abstract
Autoimmune inflammatory diseases are primarily characterized by deregulated expression of cytokines, which drive pathogenesis of these diseases. A number of approved and experimental therapies utilize monoclonal antibodies against cytokine proteins. Cytokines can be classified into different families including the interleukins, which are secreted and act on leukocytes, the tumor necrosis factor (TNF) family, as well as chemokine proteins. In this review article, we focus on the interleukin family of cytokines, of which 39 members have been identified to this date. We outline the role of each of these interleukins in the immune system, and various dermatological inflammatory diseases with a focused discussion on the pathogenesis of psoriasis and atopic dermatitis. In addition, we describe the roles of various interleukins in psychiatric, cardiovascular, and gastrointestinal comorbidities. Finally, we review clinical efficacy and safety data from emerging late-phase anti-interleukin therapies under development for psoriasis and atopic dermatitis. Collectively, additional fundamental and clinical research remains necessary to fully elucidate the roles of various interleukin proteins in the pathogenesis of inflammatory dermatologic diseases, and treatment outcomes in patients.
Keywords: Psoriasis, Atopic dermatitis, Interleukin, Cytokine
Key Summary Points
| Pathogenesis of psoriasis and atopic dermatitis is largely driven by a complex cross-regulatory network of interleukins. |
| Interleukins can drive metabolic and psychiatric comorbidities associated with inflammatory dermatologic conditions. |
| Continued characterization of pathogenic roles of interleukin proteins will facilitate development of novel biologic therapies. |
Introduction
The immune system comprises various effector cells and associated cytokines that elicit adaptive and innate immune responses against pathogens. While the innate immune response is intrinsic to the human body since birth and is nonspecific, the adaptive immune response develops over time after exposure to an antigen either from a pathogen or a vaccination, and includes the production of specific antibodies against a foreign antigen [1]. Antigen presentation on dendritic cell surface along with the necessary costimulatory molecules can lead to activation of naïve T cells and their proliferation and subsequent differentiation to either CD8+ cytotoxic T cells or CD4+ helper T cells (Th cells) [1]. Thus, dendritic cells act as an important link between the innate and adaptive immune responses.
There are five major subtypes of CD4+ T cells: Th1 cells, Th2 cells, Th17 cells, T follicular helper cells, and regulatory T cells (Treg) [1]. Th1 cells are involved in intracellular immunity and release interferon-γ to activate macrophages. Th2 cells play a crucial role in mucosal and barrier immunity and release interleukins 4 and 5 (IL-4 and IL-5) to recruit eosinophils and basophils. Th17 cells produce Th17 cytokines such as IL-17 and IL-22 that activate stromal cells and epithelial cells, respectively, to produce various chemokines and antimicrobial peptides that recruit neutrophils to eliminate extracellular pathogens. T follicular helper cells interact with naïve B cells and regulate their antibody production capacity. Finally, Tregs suppress innate immunity and T-cell activation thus acting as a safeguard against development of autoimmunity. In addition, Treg pathways exist in close interaction with each other, and have associated feedback and feedforward regulatory mechanisms.
Both innate and adaptive immune cells are able to generate inflammatory mediators to combat infection and amplify or modulate the immune response. These mediators can be categorized into chemokines and cytokines that are secreted and act on other cells that harbor the respective cognate receptors. Cytokines are small proteins important in cell signaling and important in immune-mediated diseases, while chemokines are small cytokines or signaling proteins secreted by cells that induce directional movement of leukocytes, as well as other cell types. Cytokines are the principal determinants of CD4+ T-cell lineage specification [1]. Exposure of a naïve T cell to various combinations of cytokines will allow it to differentiate to any of the five mentioned subtypes above by activating various members of the STAT family of transcription factors as well as other lineage-specific transcription factors.
It has now become abundantly clear that interleukins play critical roles in the pathogenesis of inflammatory dermatologic conditions. In this article, we conducted a literature review to identify interleukins involved in the pathogenesis of dermatologic inflammatory diseases with a focus on psoriasis and atopic dermatitis. We first describe the various known families of interleukins and their main roles in the normal immune response. We then describe important preclinical and clinical data that support pathogenic roles of interleukins not only in psoriasis and atopic dermatitis, but also in a number of associated comorbidities. These include cardiovascular, psychiatric, and gastrointestinal comorbidities. Our last goal in this review article was to highlight emerging anti-interleukin therapies for the treatment of psoriasis and atopic dermatitis.
Compliance with Ethics Guidelines
This article is based on previously conducted studies, and does not contain any new studies with human participants or animals performed by any of the authors.
Interleukin Subfamilies and Structure
The term “interleukin” (between leukocytes) was first coined in Switzerland in 1979 at the second international lymphokine workshop to describe factors that influence function of leukocytes [2]. A number of nomenclatures have been proposed to classify interleukins based on sequence homology, structural similarity, bound receptors, as well as producing cells. These nomenclatures were adopted by the World Health Organization/International Union of Immunological Societies (WHO/IUIS) in 1990s [3, 4]. However, the advent of next-generation sequencing and continued discovery of additional interleukins increases the complexity of existing nomenclatures. In a recent report, Akdis et al. conducted a comprehensive review of existing interleukin factors and classified them into various families based on sequence homology, evolutionary relationship, common receptor chains, and immunological function [5]. From a structural standpoint, interleukins can be classified into class I or class II helical proteins. Their structural characteristics are described in Table 1 [6, 7]. Table 2 summarizes the main interleukin families, their structure, and main function in the immune system, their cognate receptors, and producing cells [5, 6, 8–14].
Table 3.
Interleukins involved in dermatological diseases, important associated comorbidities, and relevant experimental or approved therapeutics
| Interleukin | Important dermatological conditions | Important co-morbidities | Relevant therapeutics | References |
|---|---|---|---|---|
|
IL-1α/β IL-RA |
Hereditary systemic autoinflammatory diseasesa Systemic inflammatory diseasesb Hidradenitis suppurativa Pustular psoriasis Neutrophilic dermatoses Urticarial vasculitis Kawasaki disease Erdheim–Chester disease |
Diabetes Cardiac remodeling Gout RA Osteoarthritis GVHD Dry eye disease |
Rilonacept is an interleukin-1 alpha (IL-1α) and interleukin-1 beta (IL-1β) cytokine trap indicated for cryopyrin-associated periodic syndromes, familial cold autoinflammatory syndrome and Muckle–Wells syndrome, deficiency of IL-1 receptor antagonist, and recurrent pericarditis Canakinumab is a recombinant, human anti-human-IL-1β monoclonal antibody indicated for treatment of autoinflammatory periodic fever syndromes Anakinra is a recombinant, nonglycosylated form of IL-1RA indicated for rheumatoid arthritis, cryopyrin-associated periodic syndromes (CAPS), and deficiency of interleukin-1 receptor antagonist (DIRA) |
[113–118] |
| IL-2 |
Alopecia areata Malignant melanoma |
Aldesleukin is a recombinant IL-2 protein therapy approved for use in patients with stage IV melanoma | [119, 120] | |
| IL-4 |
Atopic dermatitis Scleroderma Bullous pemphigoid |
Atopic comorbidities | Dupilumab, an FDA approved anti-IL-4Rα monoclonal antibody for treatment of patients with atopic dermatitis or asthma | [70, 121–124] |
| IL-5 |
Atopic dermatitis Hypereosinophilic syndrome |
Asthma EoE Eosinophilic granulomatosis with polyangiitis Nasal polyposis |
Benralizumab, mepolizumab, and reslizumab are 3 FDA-approved anti-IL-5 antibodies for treatment of adult patients with severe eosinophilic asthma | [125–127] |
| IL-6 |
Psoriasis (–) Pustular psoriasis Hidradenitis suppurativa Wound Healing |
RA Psoriatic Arthritis (–) Cardiovascular disease Depression Diabetes |
Tocilizumab, monoclonal antibody against IL-6R, indicated for RA, giant cell arteritis, systemic sclerosis-associated interstitial lung disease, polyarticular juvenile idiopathic arthritis, systemic juvenile idiopathic arthritis, cytokine release syndrome Siltuximab, monoclonal antibody against IL-6, indicated for multicentric Castleman’s disease |
[26, 31, 128–133] |
| IL-7 |
CTCL (elevated levels) Generalized verrucosis and HPV-3 susceptibility (inherited IL-7 deficiency) |
T-cell acute lymphoblastic leukemia Sjögren’s syndrome Multiple sclerosis |
Safety of GSK2618960, an experimental anti-IL-7R antibody, has been assessed in healthy subjects (NCT02293161) RN168, an experimental anti-IL-7R antibody, is being assessed in patients with type I diabetes (NCT02038764) |
[134–140] |
| IL-8 |
Psoriasis Pyoderma gangrenosum |
IBD Ovarian cancer |
BMS-986253 is an experimental anti-IL-8 monoclonal antibody being assessed in patients with incurable metastatic or unresectable solid tumors (NCT02536469) | [32, 33, 141–143] |
| IL-9 |
Atopic dermatitis Allergic contact dermatitis Mycosis fungoides Important defense mechanism against Candida |
[75, 144–147] | ||
| IL-10 |
SLE Pemphigus vulgaris Bullous pemphigoid Alopecia areata Psoriasis Atopic dermatitis Allergic contact dermatitis Melanoma BCC/SCC Cutaneous lymphomas Hypertrophic scarring |
Important in viral diseases: HIV, EBV, Herpesviruses, CMV, Poxviruses Leishmaniasis Leprosy Systemic sclerosis |
SCH708980, a monoclonal anti-IL-10 antibody being assessed as an anti-immunosuppressive agent in visceral leishmaniasis (NCT01437020) | [148, 149] |
| IL-11 |
Psoriasis Atopic dermatitis (skin remodeling) |
Cardiovascular fibrosis | [34, 76, 150] | |
| IL-12 |
Psoriasis Atopic dermatitis Photocarcinogenesis |
Important in protection against infections and allergens | Ustekinumab, an anti-IL-12/23 antibody is indicated for moderate-to-severe plaque psoriasis, active psoriatic arthritis, moderate-to-severe Crohn’s disease, and ulcerative colitis | [36, 151–154] |
| IL-13 |
Atopic dermatitis Behçet’s disease Systemic sclerosis Localized scleroderma |
Dupilumab, see above Lebrikizumab, an experimental anti-IL-13 monoclonal antibody for treatment of atopic dermatitis (NCT03443024) or asthma (NCT02104674) Tralokinumab, anti-IL-13 antibody for treatment of patients with atopic dermatitis (NCT03131648 and NCT03160885) or asthma (NCT02161757 and NCT02194699). It has received regulatory approval by Health Canada |
[83, 155–162] | |
| IL-14 |
Sjögren’s syndrome SLE |
[163, 164] | ||
| IL-15 |
Vitiligo Mycosis fungoides |
RA Sarcoidosis IBD Transplant rejection Hematologic malignancies Infections |
IL-15 superagonist complex N-803 is an experimental treatment being assessed in BCG-unresponsive non-muscle invasive bladder cancer patients (NCT03022825) | [165–168] |
| IL-16 |
Cutaneous T-cell lymphoma Systemic sclerosis SLE Bullous pemphigoid |
HIV/immune reconstitution syndrome | [169–173] | |
| IL-17 |
Psoriasis Psoriatic arthritis Ankylosing spondylitis Tumor progression |
Asthma IBD |
Secukinumab and ixekizumab (IL-17A antibodies) and brodalumab (IL-17RA antibody) Bimekizumab, a monoclonal antibody that targets both IL-17A and IL-17F that was assessed in clinical trials in patients with plaque psoriasis (NCT03536884). It is approved by the EMA, and pending approval by the FDA and Health Canada Sonelokimab is an IL-17A/17F nanobody which neutralizes both IL-17A and IL-17F, and is currently being assessed in patients with plaque psoriasis (NCT03384745) |
[39, 40, 43, 174–178] |
| IL-18 |
Lupus Hemophagocytic syndrome |
IBD CVD |
GSK1070806, an experimental monoclonal antibody against IL-18 (NCT03522662) | [179–183] |
| IL-19 |
Psoriasis Atopic dermatitis |
[46, 184] | ||
| IL-20 | Psoriasis |
RA PsA |
[47, 49] | |
| IL-21 |
SLE Psoriasis Atopic dermatitis Alopecia areata Melanoma (anti-tumorigenic role) |
RA IBD Colon cancer |
[50, 185–192] | |
| IL-22 |
Psoriasis Atopic dermatitis Contact dermatitis Scleroderma CTCL Squamous cell carcinoma |
Multiple sclerosis RA |
Fezakinumab, a monoclonal anti-IL-22 antibody being assessed in atopic dermatitis (NCT01941537) | [51, 52, 77, 193–200] |
| IL-23 | Psoriasis |
Crohn’s disease RA PsA |
Ustekinumab, see above Guselkumab, risankizumab, tildrakizumab |
[35, 90, 201–203] |
| IL-24 |
Psoriasis Contact dermatitis Atopic dermatitis |
IBD (suppresses mucosal inflammation) | [48, 54, 204–206] | |
| IL-25 | Psoriasis | Asthma | [55, 207, 208] | |
| IL-26 |
Hidradenitis suppurativa Psoriasis Behçet’s disease |
RA Crohn’s disease Asthma COPD |
[56, 209–215] | |
| IL-27 |
Psoriasis Systemic sclerosis |
COPD Breast cancer Sjögren's syndrome Rheumatoid Arthritis |
[58, 216–220] | |
| IL-28 | SLE |
Sepsis HCV clearance Hashimoto’s thyroiditis |
[221–224] | |
| IL-29 |
SLE Psoriasis Atopic dermatitis Systemic sclerosis |
RA Osteoarthritis Uveitis Sjögren’s syndrome Hashimoto’s thyroiditis |
Peginterferon Lambda-1A or IL-29 is being assessed in clinical trials for treatment of patients with COVID-19 (NCT04354259) or hepatitis C (NCT01866930) | [62, 224–231] |
| IL-30 | Psoriasis |
Breast cancer Prostate cancer |
[59, 232, 233] | |
| IL-31 | Chronic pruritic skin disorders |
Allergic asthma and rhinitis IBD Osteoporosis |
[234] | |
| IL-32 |
Psoriasis Atopic dermatitis |
Grave’s disease RA Pulmonary arterial hypertension COPD |
[64, 235–239] | |
| IL-33 |
Psoriasis Atopic dermatitis Allergic contact dermatitis Vitiligo Chronic spontaneous urticaria |
Allergic rhinitis Asthma RA Ulcerative colitis |
Itepekimab, experimental monoclonal antibody against IL-33 in COPD (NCT04751487) | [60, 240–247] |
| IL-34 |
SLE Psoriasis |
RA Sjögren’s syndrome IBD Nonalcoholic fatty liver disease Lung cancer PsA |
[65, 248–253] | |
| IL-35 | Systemic sclerosis |
Sjögren's syndrome MS |
[254–256] | |
| IL-36 |
Psoriasis SLE AD |
RA PsA UC Sjögren’s syndrome |
Imsidolimab, a monoclonal antibody against IL-36R; hidradenitis suppurativa (NCT04856930), acneiform eruptions (NCT04697069), ichthyosis (NCT04697056) | [16, 17, 23, 257–261] |
| IL-37 |
Psoriasis SLE |
Ankylosing spondylitis Graves’ disease Guillain-Barré syndrome RA |
[66, 262–266] | |
| IL-38 | Psoriasis |
RA Idiopathic pulmonary lung fibrosis |
[18, 65, 67, 267, 268] | |
| IL-39 |
Neuromyelitis optica spectrum disorders ST-segment elevation myocardial infarction |
[269, 270] |
AD, atopic dermatitis; BCC, basal cell carcinoma; COPD, chronic obstructive pulmonary disease; CTCL, cutaneous T-cell lymphoma; CVD, cardiovascular disease; GVHD, graft-versus-host-disease; IBD, irritable bowel syndrome; MS, multiple sclerosis; NTK, natural killer T; PsA, psoriatic arthritis; SCC, squamous cell carcinoma; SLE, systemic lupus erythematosus; UC, ulcerative colitis
aHereditary systemic autoinflammatory diseases: familial Mediterranean fever (FMF), cryopyrin-associated periodic syndrome (CAPS), TNF receptor-associated periodic syndrome (TRAPS), hyper-IgD syndrome (HIDS), periodic fever, aphthous stomatitis, pharyngitis and adenitis (PFAPA), deficiency of interleukin-1 (IL-1) receptor antagonist (DIRA)
bSystemic inflammatory diseases: Systemic juvenile idiopathic arthritis, Adult-onset Still’s disease, Schnitzler syndrome, Behçet’s disease, PFAPA, synovitis, acne, pustulosis, hyperostosis, osteitis (SAPHO) syndrome, macrophage activation syndrome
Table 1.
| Structure | Subclassification | Interleukin |
|---|---|---|
| Class I: 4 α-helices in an “up-up-down-down” conformation | Short chain | IL-2, IL-3, IL-4, IL-5, IL-7, IL-9, IL-13, IL-15 and IL-21 |
| Long chain | IL-6, IL-11, IL-12, IL-23, IL-27, and IL-31 | |
| Class II: 6–7 α-helices | IL-10, IL-19, IL-20, IL-22, IL-24, IL-26, IL-28, IL-29 | |
| β-Trefoil structure made of 12 packed β-sheets | IL-1 family | |
| Neurotrophin-like cysteine-knot fold | IL-17 family |
Table 2.
List of interleukin family members, their cognate receptors, their
source cells, and main functions in immune system [5, 6, 8–14]
| Interleukin family | Interleukin (s) | Producing cell(s) | Receptor(s) | Main role in immune system |
|---|---|---|---|---|
| IL-1 family | IL-1α/β | Macrophages, monocytes, dendritic cells, lymphocytes, keratinocytes, microglia, megakaryocytes, neutrophils, fibroblasts, synovial cells | IL-1 type I receptor (IL-1RI) | Potent proinflammatory cytokines, induce Th17 and B cells, induce chemokine production |
| IL-1Ra | Monocytes, fibroblasts, endothelial cells, keratinocytes, macrophages, neutrophils | IL-1RI | Inhibits IL-1α/β signaling | |
| IL-18 | Dendritic cells, macrophages, Kupffer cells, keratinocytes, osteoblasts, astrocytes | IL-18Rα/IL-18Rβ | Enhances NK cell cytotoxicity, promotes Th1 and Th2 immune responses, enhances IFN-γ production | |
| IL-33 | Fibroblasts, macrophages, damaged epithelial cells, dendritic cells | IL-1 receptor-related protein called ST2 | Released upon necrosis, inflammation, and cell damage, potent activator of Th2 immune responses | |
| Il-36α/β/γ/RA | Internal endothelial cells, keratinocytes, skin epithelia | IL-36R | NF-κB activation, activation of other proinflammatory pathways in response to tissue injury or infection, enhance Th17-immune response (inhibited by IL-36RA) | |
| IL-37 | NK cells, monocytes, B cells, keratinocytes, various human tissues | IL-18Rα, single immunoglobulin IL-1 receptor-related protein (SIGIRR) | Inhibits TLR-induced proinflammatory cytokines, suppress dendritic cell activation | |
| IL-38 | Basal epithelia of skin and in proliferating B cells of the tonsil | IL-1R1, IL-36R | Acts similar to IL-36RA, inhibits Th17 immune response, inhibits IL-36 signaling | |
| IL-6 family | IL-6 | B and T cells, eosinophils, granulocytes, mast cells, glial cells, keratinocytes, chondrocytes, osteoblasts | IL-6R, gp130 | T and B cell proliferation and differentiation, increase chemokine production, induce Th17 cells |
| IL-11 | Fibroblasts, epithelial cells, endothelial cells, synoviocytes, osteoblasts, bone marrow cells | IL-11Rα, gp130 | Stimulates hematopoiesis and platelet production, stimulates Th2 cytokines and inhibits Th1 cytokines | |
| IL-10 family | IL-10 | Monocytes, dendritic cells, Treg cells, B cells, macrophages | IL-10R1 and IL-10R2 | Immunosuppressive; inhibits TLR agonist-induced cytokines and chemokine production |
| IL-19 | Monocytes, macrophages, epithelial cells, endothelial cells | IL-20R1 and IL-20R2 | Induces Th2 responses; induces expression of IL-4, 5, 10, and 13 | |
| IL-20 | LPS-stimulated monocytes, dendritic cells, epithelial and endothelial cells | IL-20R1/IL-20R2 and IL-22R1/IL-20R2 | Regulates hematopoiesis; aid in clearance of bacterial infection via upregulation of psoriasin and β-defensin-2 | |
| IL-22 | Th17, Th22 and γδ T cells, activated NK cells | IL-22R1 and IL-10R2 | Antimicrobial defense, activate proliferative and anti-apoptotic pathways to prevent tissue damage | |
| IL-24 | B and T cells, monocytes, mast cells | IL-22R1/IL-20R2 or IL-20R1/IL-20R2 | Induces expression of IL-6, TNF-α, and IFN-γ in PBMCs, regulates balance between plasma and memory B cells | |
| IL-26 | Memory T cells, Th17 cells, and NK cells | IL-10R2/IL-20R1 | Induce proinflammatory cytokines IL-6, IL-8, and TNF-α in epithelial cells, inhibits IgG and IgA production in B cells | |
| IL-28A (IFN-λ2) | Dendritic cells, PBMCs, T reg cells, virus-infected cells, monocytes | IL-28R1/IL-10R2 | Have antiviral activities against various viruses | |
| IL-28B (IFN-λ3) | ||||
| IL-29 (IFN-λ1) | ||||
| IL-12 family | IL-12p40 and IL-12p70 | Monocytes, antigen presenting cells such as macrophages and dendritic cells, neutrophils | IL-12Rβ1 and IL-12Rβ2 | Development and maintenance of Th1 cells through induction of IFN-γ, enhance cytolytic activity of NK cells and antimicrobial activity of macrophages |
| IL-23 | Phagocytic cells, macrophages, activated DCs | IL-23R | Induction and differentiation of Th17 cells, NK cell activation, T-cell proliferation | |
| IL-27 | Antigen-presenting cells such as macrophages and dendritic cells, endothelial cells | WSX-1 (IL-27Ra) and gp130 | Development of Th1 cells, antagonizes Th17 cell response | |
| IL-35 | Treg cells, stimulated monocytes, B cells | IL-12Rβ2 and gp130 | Reduce proliferation of effector T cells and enhance that of Tregs, induce iTr35 cells, inhibit Th17 immune responses | |
| IL-17 family | IL-17A | Memory CD4+ T cells, Th17 cells, CD8+ T cell, γδ T cells, NK cells, neutrophils | IL-17RA, IL-17RC | Induce expression of proinflammatory cytokines, chemokines, and metalloproteases. Recruits neutrophils |
| IL-17F | Similar cell sources as IL-17A; more abundant in the skin compared to IL-17A | IL-17RA, IL-17RC | Similar to IL-17A, it induces chemokines, and proinflammatory cytokines | |
| IL-17B | Spinal cord cells, testis, small intestines, various nonimmune tissues | IL-17RB | May play a role in fracture healing, embryonic chondrogenesis, and neutrophil recruitment | |
| IL-17C | Synovial fluid mononuclear cells (SFMC), and PBMCs | IL-17RE | Enhances proinflammatory cytokines such as IL-1β, TNF-α, and IL-6, can induce IL-23 | |
| IL-17D | Skeletal muscle, brain, adipose tissue, heart, lung, and pancreas | Not known | Inhibits myeloid progenitor cell proliferation, enhances IL-6, IL-8, LIF, and MMP-3 production | |
| IL-17E/IL-25 | Th2 cells, alveolar macrophages, granulocytes, intestinal epithelial cells, brain endothelial cells | IL-17RA and IL-17RB | Enhances Th2 cell response, inhibits Th1 and Th17 cell response | |
| Th2 cytokines | IL-5 | Th2 cells, eosinophils, mast cells | IL-5Rα and a β chain subunit (CD131) recognized by IL-3 and GM-CSF | Activation, maturation, and survival of eosinophils |
| IL-13 | Th2 cells, T cells, NKT cells, mast cells, basophils, eosinophils | IL-4Rα and IL-13Rα1 | Promotes isotype switching of B cells, inhibits inflammatory cytokines, defends against nematodes | |
| IL-31 | Activated CD4+ Th2 cells, mast cells, monocytes, macrophages, dendritic cells | IL-31RA and oncostatin M receptor β (OSMRβ) | Modulates eosinophil function, induces chemokines in keratinocytes, induces itching upon engaging its receptors on sensory neurons | |
| Chemokines | IL-8 | Monocytes, macrophages, neutrophils, endothelial cells | CXC chemokine receptors: IL-8RA and IL-8RB | Recruitment of neutrophils to infection/injury site |
| IL-16 | T cells, mast cells, eosinophils, monocytes, dendritic cells | CD4 | Chemoattractant of CD4+ and CD8+ T cells, monocytes, mast cells and eosinophils | |
| Common γ chain cytokine family | IL-2 | CD4+ and CD8+ T cells | IL-2R composed of IL-2Rα, IL-2Rβ, and γc (CD132) | Proliferation and development of effector B and T cells |
| IL-4 | Th2 and type II innate lymphoid cells (ILC2) | Type I and II IL-4R | Development of Th2 cells from naïve T cells | |
| IL-7 | Stromal and epithelial cells of non-hematopoietic origin | IL-7R composed of IL-7Rα and CD132 | Lymphocyte development in the thymus, and naïve T-cell survival | |
| IL-9 | Th2 and Th9 cells | IL-9R composed of IL-9Rα and CD132 | Proliferation of CD8+ T cells and mast cells, reduction of Th1 cytokines, enhancing Th2 immune responses | |
| IL-15 | Monocytes, CD4+ T cells, macrophages, dendritic cells | IL-15R composed of IL-15Rα, IL-15Rβ, and CD132 | T cell and NK cell activation | |
| IL-21 | T cells and NKT cells | IL-21R | B cell differentiation and activation, NKT cell survival, CD4+ T-cell proliferation, NK cell maturation | |
| Other interleukins | IL-3 | T cells, NK cells, eosinophils, macrophages | IL-3Rα and CD131 | Hematopoietic cell differentiation, synergizes with GM-CSF to induce granulocyte–macrophage lineage, enhance macrophage function |
| IL-14 | Normal T cells and B cells, and cancerous B cells | IL-14R | Enhances B cell proliferation, inhibits antibody production | |
| IL-32 | T cells, NK cells, lung and intestinal epithelial cells | Unknown | Induces monocyte to macrophage differentiation | |
| IL-34 | Splenic cells, neuronal cells | CSF1R | Proliferation of monocytes and macrophages |
Interleukins in Pathogenesis of Psoriasis and Atopic Dermatitis
Interleukins produced by innate and adaptive immune cells can form cross-regulatory networks that are the driving force in many dermatologic inflammatory diseases. Table 2 summarizes dermatologic and non-dermatologic diseases in which various interleukins are implicated. In addition, it identifies relevant approved or experimental therapeutics targeting each interleukin. In this section, we discuss the roles of various interleukins in the pathogenesis of two of the most common dermatologic inflammatory diseases, psoriasis (Fig. 1) and atopic dermatitis (Fig. 2).
Fig. 1.
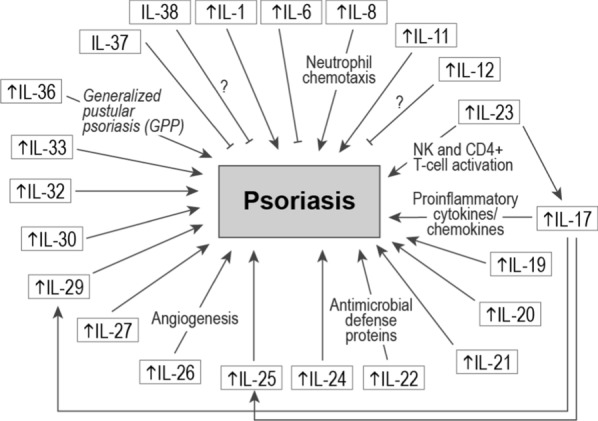
Interleukins involved in pathogenesis of psoriasis. An upwards arrow (↑) indicates elevated expression. Many interleukins contribute to psoriasis pathogenesis whereas others may play a protective role through increased expression of antimicrobial defense proteins
Fig. 2.
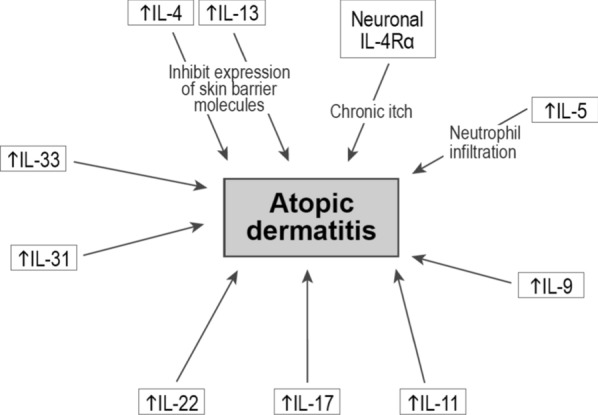
Interleukins involved in pathogenesis of atopic dermatitis. An upwards arrow (↑) indicates elevated expression
Psoriasis
In psoriasis, a number of stress triggers—ranging from environmental to genetic—can stimulate innate immune cells and keratinocytes to synthesize tumor necrosis factor-α (TNF-α), interferon-γ (IFN-γ), IFN-α, IL-6, and IL-1β [15]. These cytokines activate dermal dendritic cells to produce IL-12 and IL-23, which promote differentiation of naïve T cells into Th1 and Th17 cells, respectively. Production of TNF-α and IFN-γ by Th1 cells and IL17A-F and IL-22 by Th17 cells leads to activation and proliferation of keratinocytes, which in turn produce a wide range of proinflammatory cytokines and chemokines that results in the phenotypic manifestations of psoriasis. These inflammatory mediators exist in a positive feedback loop allowing transition from disease initiation to disease maintenance and chronicity [15].
Several additional interleukins have been documented to have complex roles in the pathogenesis of psoriasis. Gene expression profiling of generalized pustular psoriasis and psoriasis vulgaris lesional biopsies revealed elevated IL-1, IL-17A, and IL-36 expression levels compared to biopsies of normal skin [16]. Interestingly, IL-1 and IL-36 levels were more elevated in pustular psoriasis vs. psoriasis vulgaris. Immunohistochemical analysis showed that IL-36 was particularly expressed in keratinocytes surrounding neutrophilic pustules. Importantly, loss of function mutations in the IL36RN gene, which encodes for the IL-36 receptor antagonist, have been linked to pathogenesis of a debilitating form of psoriasis, generalized pustular psoriasis (GPP) in a mechanism that involves unrestrained IL-36 signaling that causes IL-1 activation [17, 18]. Anakinra, an IL-1R antagonist, has been described to improve pustular eruptions in generalized pustular psoriasis patients [19]. In addition, it has also proven effective in a patient with pustular psoriasis driven by IL36RN mutations, although it remains unclear what weight IL-1 vs. IL-36 have in pathogenesis of pustular psoriasis [20]. IL-36R inhibition blocks proinflammatory pathways underling GPP, and it has been suggested that IL-36 inhibiting therapies may provide more complete clinical benefit in patients harboring IL36RN mutations [21–23]. In addition, data from the 12-week Effisayil-1 trial, which examined efficacy of spesolimab, a humanized monoclonal IgG1 anti-IL-36R antibody, demonstrated superiority (vs. placebo) in clearing flares in patients with generalized pustular psoriasis [24]. Efficacy of imsidolimab, another IL-36R monoclonal antibody, was examined in the GALLOP phase 2 open-label trial. Seventy-five percent of patients with generalized pustular psoriasis (6/8) receiving imsidolimab achieved improvement in Clinical Global Impression (CGI) based on the modified Japanese Dermatology Association Severity Index (mJDA-SI) both at week 4 and week 16 [25].
IL-6 has been reported to be highly elevated in active psoriatic plaques from 35 psoriasis patients compared to non-lesional biopsies. In addition, IL-6 levels were increased in plasma of these patients, which may also contribute to epidermal hyperplasia given that IL-6 stimulates keratinocyte proliferation [26]. While this might indicate that targeting IL-6 could be beneficial in psoriasis treatment, clinical data reveals the opposite. Tocilizumab, a monoclonal anti-IL-6R antibody indicated for moderate-to-severe rheumatoid arthritis (RA), has been shown to induce psoriasis-like disease in a number of RA patients [27–29]. It is noteworthy that preclinical data originating from mouse studies have shown that genetic deletion of IL-6 can result in more severe psoriasis-like disease, potentially through a mechanism involving increased expression of proinflammatory cytokines that appear to compensate for IL-6 deletion [30, 31].
IL-8 levels are increased in psoriatic scale extracts as well as suction blister fluids in psoriatic skin lesions [32]. The major source of IL-8 in psoriatic tissue appears to be neutrophils located immediately under the parakeratotic stratum corneum (Munro’s microabcess) [33]. Given that IL-8 induces chemotaxis in neutrophils, and that neutrophils can synthesize IL-8 upon activation, it is plausible that IL-8 acts in an autocrine fashion to maintain neutrophil infiltration in psoriatic tissue thereby contributing to inflammation [33].
Given the anti-inflammatory properties of IL-11, a phase I open-label clinical trial assessed the efficacy of daily recombinant human IL-11 (rhIL-11) subcutaneous injections in 12 psoriasis vulgaris patients [34]. 11/12 patients had a 20–80% improvement in Psoriasis Area and Severity Index (PASI) score after 8 weeks of treatment, which was associated with reduced expression of proinflammatory cytokines such as IFN-γ, IL-8, IL-12, TNF-α, IL-1β. Large clinical trials remain necessary to confirm efficacy of rhIL-11 as an anti-inflammatory agent in psoriasis patients.
Ustekinumab acts by targeting the p40 subunit shared by IL-12 and IL-23, both of which are involved in inflammation and NK cell and CD4+ T-cell activation in psoriasis. Furthermore, both IL-23 and IL-23R are overexpressed in human psoriatic tissue [35]. The individual contribution of IL-12 and IL-23 in the imiquimod-induced psoriasis mouse model was recently examined [36]. As expected, mice deficient in the p40 subunit had a significant decrease in psoriatic plaque formation, however, mice that were engineered to be deficient of the IL-12 specific p35 subunit or the IL-12 receptor (i.e., intact IL-23 signaling) suffered from more severe inflammation and plaques compared to their wild type counterparts. This involved increased infiltration of IL-17A-producing Vγ4+ γδT cells. Importantly, this phenotype was alleviated by exposure to IL-12, suggesting that IL-12 has a protective effect and limits skin inflammation in this psoriasis mouse model. In human keratinocytes, IL-12 exposure also counteracted the psoriatic transcriptional signature, although the specific role of IL-12 in patients with psoriasis is still questionable as no monoclonal antibody targeting only IL-12 has been studied in humans. These data may suggest that concomitant targeting of IL-12 along with IL-23 may be counterproductive. Indeed, it has been reported that guselkumab and risankizumab, which target the p19 subunit of IL-23, were both associated with superior clinical responses in patients with moderate-to-severe plaque psoriasis, compared to ustekinumab [37, 38].
While the general consensus is that Th17 cells are a major source of IL-17 in psoriatic lesions, other cell types such as mast cells and γδ T cells are also important cellular players [39, 40]. IL-17 production by Th17 cells is stimulated by IL-23, which also promotes survival of these cells [41]. IL-17 stimulates keratinocytes to produce antimicrobial peptides as well as various proinflammatory cytokines (IL-1β, TNF-α, IL-6) and chemokines (CXCL1/3/5). The role of the IL-17/IL-23 axis in psoriasis pathogenesis has been examined in IL-17 deficient mice [42]. IL-23 exposure was sufficient to induce epidermal hyperplasia in wild-type mice but not in IL-17-deficient mice. In addition, preexposure to anti-IL-17A antibodies prevented IL-23-mediated epidermal hyperplasia. These data suggest that IL-17A plays a critical role in IL-23-mediated psoriatic pathology. However, it remains unclear whether IL-17 upregulation alone is sufficient to initiate psoriatic disease, and whether it can be counteracted by IL-23 targeting antibodies or in an IL-23 −/− genetic background. Finally, it’s noteworthy that inhibition of IL-17F in addition to IL-17A has been shown to provide more effective clinical responses compared to sole inhibition of IL-17A. Indeed, the BE RADIANT trial showed that 61.7% of patients who received bimekizumab (230/373), anti-IL17A/17F antibody, achieved a PASI100 response at week 16, compared to 48.9% in the secukinumab arm (181/370) [43]. Similar results were also seen with sonelokimab, a novel trivalent nanobody that comprises three monovalent camelid-derived (i.e., from the Camelidae family of mammals) monovalent nanobodies against IL-17A, IL-17F, and human serum albumin. Indeed, 76.5% of patients receiving sonelokimab achieved a PASI90 response (39/51) at week 12, compared to 64.2% in the secukinumab arm (34/53) [44].
The IL-10 family of cytokines is comprised of IL-10, IL-28, IL-29 and the IL-20 subfamily (IL-19, IL-20, IL-22, IL-24, and IL-26) [45]. These cytokines are implicated in psoriasis. Expression of IL-19 and IL-20 is elevated in basal and suprabasal psoriatic keratinocytes, although their expression was lower in the sera of psoriasis patients compared to healthy volunteers [46, 47]. Given that IL-19 is one of the main cytokines induced by IL-1β, its elevated expression may be a consequence of elevated expression of IL-1β in psoriatic tissue as mentioned earlier [48]. Both IL-19 and IL-20 can induce expression of keratinocyte growth factor (KGF) in CD8+ T cells. In a study of paired synovial tissue and psoriatic skin lesions of patients with psoriatic arthritis (PsA) and plaque psoriasis, IL-20 expression positively correlated with disease activity at baseline (PASI) and after treatment with alefacept [49]. Interestingly, alefacept treatment resulted in a significant reduction in IL-20 expression in skin lesions whereas its expression remained unchanged in synovial tissue, suggesting a role of CD4+ and CD8+ T cells in regulating its expression in psoriatic lesions.
IL-21 expression is also elevated in human psoriasis skin lesions, and intradermal exposure of mice to this interleukin results in keratinocyte hyperplasia [50]. Importantly, IL-21 antibody treatment was able to resolve inflammation and epidermal hyperplasia in a human psoriasis xenograft mouse model, suggesting that IL-21 targeting therapy may be a potential treatment avenue for psoriasis in the future.
IL-22 expression is increased in psoriatic skin lesions and sera of psoriasis patients compared to healthy volunteers [51]. It appears that IL-22 is responsible for expression of antimicrobial defense proteins such as psoriasin, calgranulin A, and calgranulin B, and keratinocyte mobility proteins such as matrix metalloproteinases (MMP1 and MMP3). In addition, IL-22 plasma levels positively correlated with disease severity as defined by PASI score, suggesting this interleukin may be involved in psoriasis pathogenesis, although a monoclonal antibody targeting IL-22, fezakinumab, was not effective in a phase 1 trial [52, 53].
One of the main target tissues of IL-24 is the skin, where its receptors IL-22R1/IL-20R2 or IL20R1/IL-20R2 are highly expressed. In addition, IL-24 expression is elevated in psoriatic lesions, and it is synthesized by both peripheral blood mononuclear cells (PBMCs) and keratinocytes [48]. Expression of IL-19, IL-20, and IL-24, all of which are of epidermal origin, was drastically reduced by etanercept treatment in psoriasis patients after 1 week [54]. In addition, expression of various keratinocyte-derived chemokines and proinflammatory interleukins was suppressed by etanercept. These changes took place concomitantly or prior to reduction in epidermal hyperplasia. Interestingly, expression of the IL-17/IL-23 interleukins IL-23p19, IL-12p40, IL-17A was also reduced although only after 3–4 weeks of treatment. These data highlight the complex regulation of various interleukins by TNF-α in the pathogenesis of psoriasis.
IL-25 or IL-17E is highly expressed in human psoriatic lesions, and is sufficient to cause psoriasis-like pathological features in mice such as acanthosis, parakeratosis, and immune cell infiltration [55]. Importantly, IL-25 (IL-17E) is induced by IL-17 and also positively autoregulates itself in keratinocytes via IL-17RB receptors leading to increased expression of proinflammatory cytokines and chemokines. This autoregulatory IL-25 circuit may therefore present a novel target in psoriasis, with potentially less adverse effects in patients particularly as its inhibition does not affect IL-17-dependent host defense mechanisms against infections [55].
IL-26 is increased in psoriatic tissue, and is implicated in vascularization and immune cell infiltration [56]. IL-26 can stimulate human keratinocytes to produce high levels of fibroblast growth factors that are involved in angiogenesis [56]. Not only are these factors known to be elevated in plasma of psoriasis patients, but they also correlate positively with disease severity [57]. IL-27, IL-30 and IL-33 serum levels were increased in psoriasis patients and correlated with disease PASI scores [58–60]. IL-29, which is mainly produced by Th17 cells, is also highly expressed in human psoriatic lesions [61]. Exposure of human keratinocytes to IL-27 or IL-29 results in enhanced expression of various chemokines such as CXCL9 and CXCL10 known to be induced in human psoriatic lesions [58, 62, 63]. IL-32, IL-33, and IL-36α are all elevated in the plasma of psoriasis patients or psoriatic lesions with less characterized roles in pathogenesis of the disease [64, 65].
IL-37 and IL-38 follow a distinct mechanism of action in psoriasis pathogenesis compared to previous interleukins. IL-37 is an inhibitor of innate immunity and has potent anti-inflammatory activity whereby it suppresses expression of proinflammatory genes CXCL8, IL-6, and S100A7 in human keratinocytes [66]. IL-37 exposure was also able to diminish psoriasis symptoms in the K14-VEGF-Tg human psoriasis mouse model [66]. Additional studies remain necessary to understand the anti-inflammatory properties of IL-37 and whether these can be harnessed for treatment of human psoriasis. Expression of IL-38, also known as a receptor antagonist for IL-36, is reduced in serum and psoriatic lesions in psoriasis patients [67]. Interestingly, IL-38 expression is enhanced by secukinumab (anti-IL-17A) treatment and is correlated with its therapeutic efficacy [67]. It remains important to investigate any potential clinical efficacy of IL-36 pathway inhibitors such as imsidolimab and spesolimab, or recombinant IL-38 in treatment of psoriasis.
Atopic Dermatitis
Atopic dermatitis (AD) is a common inflammatory disease that usually begins in childhood and continues into adulthood, although it may less often also start later in life [68]. AD is characterized by pruritus and disease flares, and its pathogenesis primarily involves skin barrier dysfunction together with immune cell dysregulation [68, 69]. Disruption of the epidermal layer along with environmental triggers activates keratinocytes to begin production of various proinflammatory cytokines, which in turn can activate dendritic cells. Dendritic cells subsequently stimulate production of Th2 cell cytokines IL-4, IL-5, IL-13, IL-31, and IL-33, which lead to barrier dysfunction, inhibition of keratinocyte differentiation and pruritus. The chronic phase of the disease is characterized by activation of additional Th cells such as Th17, Th1, and Th22 cells that produce IL-17, IFN-γ, and IL-22, respectively, leading to T-cell chemotaxis, abnormal keratinocyte proliferation, and thickening of the epidermal layer [69].
Consistent with a pathogenic role of IL-4 and IL-13 in AD pathogenesis, dupilumab, which is an approved anti-IL-4Rα monoclonal antibody, improved AD signs and symptoms, including pruritus, and quality of life, compared to placebo in the SOLO 1 and SOLO 2 trials [70]. As mentioned above, Th2 cytokines are the main hallmark of AD disease. Both IL-4 and IL-13 can inhibit expression of skin barrier molecules such as filaggrin, loricrin, and involucrin, which in turn further disrupts tight cell junctions, and may increase susceptibility to infections [71, 72]. In addition, neuronal IL-4Rα signaling has been shown to be a crucial component of chronic itch, given that Th2 cytokines can sensitize sensory neurons to a number of pruritogens [73]. IL-5, which plays an important role in eosinophil development, survival and proliferation, leads to increased infiltration of eosinophils in the epidermis in AD together with increased neuronal branching [74]. Th9 cell numbers, IL-9 expression in AD lesions, and IL-9 serum concentration were all elevated in AD patients compared to psoriasis patients and healthy volunteers [75]. IL-11 and type I collagen deposition consistent with papillary dermal fibrosis were found to be increased in chronic AD lesions [76]. IL-22 producing CD4+ and CD8+ T cells were significantly increased in AD skin lesions compared to psoriatic lesions, and correlated with AD disease severity [77]. IL-31, also known as the pruritic interleukin, was highly expressed in pruritic AD skin tissue compared to nonpruritic psoriatic skin lesions [78]. Mice treated with IL-31 had itch-associated scratching behavior similar to that observed in human AD [79]. IL-33 serum levels were elevated in AD patients compared to patients with urticaria and psoriasis or healthy volunteers, and correlated with disease severity [80]. Interestingly, transgenic mice that overexpress IL-33 develop itchy dermatitis with epidermal thickening and eosinophil infiltration, which are disease hallmarks reminiscent of human AD [81].
With regards to emerging therapies for treatment of AD, IL-13 inhibitors and IL-31 inhibitors have all demonstrated efficacy to reduce signs and symptoms of AD. Lebrikizumab is a monoclonal antibody against soluble IL-13 that is currently being assessed in the ADvocate 1 and ADvocate 2 phase 3 trials for treatment of patients with moderate-to-severe AD. It was recently announced that the trials met their primary endpoints, with EASI 75 achieved in > 50% of patients, and this was associated with significant itch improvement [82]. Tralokinumab, which has received regulatory approval by Health Canada, is a humanized IgG4 monoclonal antibody that binds with high affinity to IL-13, and prevents its interaction with IL-13R. Data from the ECZTRA 1 and ECZTRA 2 trials in patients with moderate-to-severe AD revealed that compared to placebo, significantly more patients who received tralokinumab achieved an IGA score of 0 or 1 (ECZTRA 1: 15.8% vs. 7.1%; ECZTRA 2: 22.2% vs. 10.9%) or EASI 75 (ECZTRA 1: 25% vs. 12.7%; ECZTRA 2: 33.2% vs. 11.4%) [83]. Nemolizumab is a humanized monoclonal antibody against IL-31 receptor A whose efficacy was examined in a double-blind phase 3 trial in Japan in patients with AD and moderate-to-severe pruritus. The mean % change in the visual-analogue scale (VAS) score at week 16 was − 42.8% in the nemolizumab group compared to − 21.4% with placebo (p < 0.001) [84].
Interleukins in Metabolic and Psychiatric Comorbidities Associated with Dermatological Diseases
Metabolic, cardiovascular, gastrointestinal, and psychiatric conditions are amongst the most common comorbidities known to be associated with dermatologic diseases. IL-1β has been linked to etiology of type 2 diabetes mellitus, which is characterized by β cell destruction. Activation of nod-like receptor protein 3 inflammasomes in macrophages and dendritic cells via islet amyloid polypeptides (IAPP) leads to enhanced production of IL-1β that acts on IL-1β receptors on β cells and causes β cell death, and defective insulin production [85]. Obesity can also lead to enhanced inflammasome activation in adipose tissue and increased IL-1β synthesis, which induces adipocyte differentiation and decreases fat oxidation [85]. Importantly, anakinra, a recombinant human IL-1-receptor antagonist, has been shown to improve glycemia and beta-cell secretory function in patients with type 2 diabetes [86]. IL-1β through activation of IL-6 signaling also plays a role in cardiovascular disease. Canakinumab, an anti-IL-1β antibody, results in reduction of plasma levels of IL-6 and high-sensitivity C-reactive protein, which are inflammatory biomarkers associated with increased risk of cardiovascular disease [87]. In addition, the CANTOS trial showed that canakinumab resulted in a significantly lower rate of recurrent cardiovascular events in patients with previous myocardial infarction, although it did not reduce incident diabetes [88, 89].
IL-23 plays an important role in inflammatory bowel disease (IBD). Genome-wide association studies have identified a strong association between IL23R and ulcerative colitis [90]. Furthermore, Th17 interleukins are increased in intestinal tissue of patients with IBD [91, 92]. This is consistent with increased activation and expression of IL-23, which as mentioned earlier, is a robust activator of Th17 signaling [93]. Indeed, ustekinumab resulted in significantly higher clinical response rates and remission rates compared to placebo in patients with Crohn’s disease or ulcerative colitis in the UNITI-1, UNITI-2, and IM-UNITI trials [94].
Psychosocial stress and some dermatologic conditions exist in a positive feedback loop where each condition exacerbates the other. Patients with psoriasis may develop anxiety because of associated chronic pruritus and skin disfiguration [95, 96]. Pruritus intensity positively correlates with patients’ quality of life, and feelings of stigmatization [97]. In addition, perceived poor social support has been associated with exacerbation of psoriasis [98]. Also, patients with moderate or severe depression had a significantly higher risk of developing psoriasis [99]. Early maladaptive schemas such as emotional deprivation, social isolation, failure, and vulnerability to harm were all more prevalent in patients with psoriasis [100]. On the other hand, there are several mechanisms by which psoriasis can trigger or exacerbate anxiety. Psychological stress can perturb homeostasis of epidermal permeability barrier [101]. Activation of the hypothalamus–pituitary–adrenal (HPA) axis and sympathetic adrenomedullary (SAM) axis is a response to stress. These processes are deregulated in psoriasis patients, which results in reduced secretion of cortisol and elevated epinephrine and norepinephrine [102, 103]. This in turn may trigger activation of mast cells and upregulation of proinflammatory cytokines, which creates a self-maintaining cycle [96].
A comprehensive literature review conducted by Ganança et al. to assess cytokine roles in suicidal behavior identified IL-6 elevations as the most robust association with suicidal ideation, trait aggression, impulsivity, and hopelessness [104]. Patients who have major depressive disorder have also been shown to have higher levels of IL-6, and other proinflammatory cytokines such as IL-1β, TNF-α, all of which may contribute to psoriatic pathogenesis [105–107]. In addition, several reports point to involvement of the nervous system in the development of psoriasis skin lesions, and as such IL-12 can also serve as a potential link between the skin immune system and the nervous system as it is produced by peripheral nerve cells [108, 109]. Cytokines are also known to stimulate activity of indoleamine 2,3-dioxygenase, which catalyzes conversion of tryptophan to kynurenine [110]. Kynurenine metabolism alterations are strongly associated with progression of various neurological diseases [111]. In addition, plasma kynurenine levels are increased in individuals who attempted suicide [112]. It remains unclear whether increased production of proinflammatory cytokines in dermatologic diseases influences kynurenine metabolism.
With increased prevalence of mental health disorders such as depression and suicidal ideation in psoriasis patients, it becomes important to utilize tools to measure quality of life and psychiatric comorbidities in the clinic. Some of these validated tools include the patient health questionnaire (PHQ), psoriasis quality of life questionnaire (PQOL-12), psychological general well-being (PGWB) index, Columbia suicide severity rating scale (C-SSRS), and Columbia classification algorithm for suicide assessment (C-CASA) [110].
Future Directions
Pathogenesis of psoriasis and AD is largely driven by a complex cross-regulatory network of interleukins (Figs. 1, 2). As previously discussed, expression of many interleukins is elevated in psoriatic and AD affected areas as well as in patient sera. Targeting these pathogenic interleukins for treatment of psoriasis and AD patients has proven successful in the clinic, although some patients may require dose adjustment, combination therapy, or switching to other biologic or nonbiologic agents due to intrinsic or acquired resistance. Trivalent antibodies that can target more than one interleukin may prove beneficial, although combined inhibition of several interleukins may be a concern from a safety standpoint. Continued discovery of interleukins and cytokines, elucidation of their structure and immune functions, along with next-generation sequencing of pathogenic tissues is critical in identifying new targets in pathogenesis of inflammatory skin diseases. In addition, identifying immune biomarkers that can predict patient response to therapy will be important in improving patient outcomes.
Acknowledgements
Funding
Publication fees were covered by Bausch Health Canada Inc.
Medical Writing and Editorial Assistance
The authors wish to acknowledge Houssam Ismail, PhD, for providing medical writing assistance with this manuscript, which was funded by Bausch Health Canada, Inc. Dr. Ismail is a freelance medical writer employed by STA HealthCare Communications. The authors also would like to thank Dr. Yuka Asai for critical reading of the manuscript.
Author Contributions
I.T. and M.B. conceived and designed the outline of the work, and co-wrote the manuscript.
Disclosures
Dr. Turchin was a consultant, speaker and investigator for AbbVie, Amgen, Arcutis, Aristea, Bausch Health, Boehringer Ingelheim, Celgene, Eli Lilly, Galderma, Incyte, Janssen, Kiniksa, LeoPharma, Novartis, Pfizer, Sanofi, UCB. Dr. Bourcier was a consultant speaker for AbbVie, Amgen-Wyeth, Bausch Health, Celgene, Eli-Lilly, GSK, Janssen, LEO Pharma, Pfizer Novartis, Sanofi, UCB and was an advisor for AbbVie, Amgen-Wyeth, Bausch Health, BMS, Celgene, Eli-Lilly, Janssen, LEO Pharma, Pfizer, Novartis, Sanofi, UCB. He has been a principal investigator for the following companies: AbbVie, Amgen, Celgene, Eli-Lilly, Janssen, Leo, Novartis.
Compliance with Ethics Guidelines
This article is based on previously conducted studies, and does not contain any new studies with human participants or animals performed by any of the authors.
Data Availability
There are no data associated with this review article apart from what is presented and referenced here.
References
- 1.Murphy K, Weaver C. Janeway’s Immunobiology. 9th edition. Garland Science.
- 2.Revised nomenclature for antigen-nonspecific T cell proliferation and helper factors. J Immunol. 1979;123:2928–9. [PubMed]
- 3.Nomenclature for secreted regulatory proteins of the immune system (interleukins). WHO-IUIS Nomenclature Subcommittee on Interleukin Designation. Bull World Health Organ. 1991;69:483–6. [PMC free article] [PubMed]
- 4.Nomenclature for secreted regulatory proteins of the immune system (interleukins): update. IUIS/WHO Standing Committee on Interleukin Designation. Bull World Health Organ. 1997;75:175. [PMC free article] [PubMed]
- 5.Akdis M, Aab A, Altunbulakli C, Azkur K, Costa RA, Crameri R, et al. Interleukins (from IL-1 to IL-38), interferons, transforming growth factor β, and TNF-α: receptors, functions, and roles in diseases. J Allergy Clin Immunol. 2016;138:984–1010. doi: 10.1016/j.jaci.2016.06.033. [DOI] [PubMed] [Google Scholar]
- 6.Brocker C, Thompson D, Matsumoto A, Nebert DW, Vasiliou V. Evolutionary divergence and functions of the human interleukin (IL) gene family. Hum Genom. 2010;5:30–55. doi: 10.1186/1479-7364-5-1-30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hymowitz SG, Filvaroff EH, Yin J, Lee J, Cai L, Risser P, et al. IL-17s adopt a cystine knot fold: structure and activity of a novel cytokine, IL-17F, and implications for receptor binding. EMBO J. 2001;20:5332–5341. doi: 10.1093/emboj/20.19.5332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Coondoo A. Cytokines in dermatology—a basic overview. Indian J Dermatol. 2011;56:368–374. doi: 10.4103/0019-5154.84717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Dinarello CA. Historical review of cytokines. Eur J Immunol. 2007;37:S34–45. doi: 10.1002/eji.200737772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Jia H, Liu J, Han B. Reviews of interleukin-37: functions, receptors, and roles in diseases. BioMed Res Int. 2018;2018:e3058640. doi: 10.1155/2018/3058640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lin H, Ho AS, Haley-Vicente D, Zhang J, Bernal-Fussell J, Pace AM, et al. Cloning and characterization of IL-1HY2, a novel interleukin-1 family member*. J Biol Chem. 2001;276:20597–20602. doi: 10.1074/jbc.M010095200. [DOI] [PubMed] [Google Scholar]
- 12.Zenewicz LA, Flavell RA. Recent advances in IL-22 biology. Int Immunol. 2011;23:159–163. doi: 10.1093/intimm/dxr001. [DOI] [PubMed] [Google Scholar]
- 13.Wynn TA. IL-13 effector functions. Annu Rev Immunol. 2003;21:425–456. doi: 10.1146/annurev.immunol.21.120601.141142. [DOI] [PubMed] [Google Scholar]
- 14.Skak K, Kragh M, Hausman D, Smyth MJ, Sivakumar PV. Interleukin 21: combination strategies for cancer therapy. Nat Rev Drug Discov. 2008;7:231–240. doi: 10.1038/nrd2482. [DOI] [PubMed] [Google Scholar]
- 15.Nestle FO, Kaplan DH, Barker J. Psoriasis. N Engl J Med. 2009;361:496–509. doi: 10.1056/NEJMra0804595. [DOI] [PubMed] [Google Scholar]
- 16.Johnston A, Xing X, Wolterink L, Barnes DH, Yin Z, Reingold L, et al. IL-1 and IL-36 are dominant cytokines in generalized pustular psoriasis. J Allergy Clin Immunol. 2017;140:109–120. doi: 10.1016/j.jaci.2016.08.056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Onoufriadis A, Simpson MA, Pink AE, Di Meglio P, Smith CH, Pullabhatla V, et al. Mutations in IL36RN/IL1F5 are associated with the severe episodic inflammatory skin disease known as generalized pustular psoriasis. Am J Hum Genet. 2011;89:432–437. doi: 10.1016/j.ajhg.2011.07.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Marrakchi S, Guigue P, Renshaw BR, Puel A, Pei X-Y, Fraitag S, et al. Interleukin-36-receptor antagonist deficiency and generalized pustular psoriasis. N Engl J Med. 2011;365:620–628. doi: 10.1056/NEJMoa1013068. [DOI] [PubMed] [Google Scholar]
- 19.Viguier M, Guigue P, Pagès C, Smahi A, Bachelez H. Successful treatment of generalized pustular psoriasis with the interleukin-1-receptor antagonist Anakinra: lack of correlation with IL1RN mutations. Ann Intern Med. 2010;153:66–67. doi: 10.7326/0003-4819-153-1-201007060-00030. [DOI] [PubMed] [Google Scholar]
- 20.Hüffmeier U, Wätzold M, Mohr J, Schön MP, Mössner R. Successful therapy with anakinra in a patient with generalized pustular psoriasis carrying IL36RN mutations. Br J Dermatol. 2014;170:202–204. doi: 10.1111/bjd.12548. [DOI] [PubMed] [Google Scholar]
- 21.Ganesan R, Raymond EL, Mennerich D, Woska JR, Caviness G, Grimaldi C, et al. Generation and functional characterization of anti-human and anti-mouse IL-36R antagonist monoclonal antibodies. MAbs. 2017;9:1143–1154. doi: 10.1080/19420862.2017.1353853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Tauber M, Viguier M, Le Gall C, Smahi A, Bachelez H. Is it relevant to use an interleukin-1-inhibiting strategy for the treatment of patients with deficiency of interleukin-36 receptor antagonist? Br J Dermatol. 2014;170:1198–1199. doi: 10.1111/bjd.12805. [DOI] [PubMed] [Google Scholar]
- 23.Bachelez H, Choon S-E, Marrakchi S, Burden AD, Tsai T-F, Morita A, et al. Inhibition of the interleukin-36 pathway for the treatment of generalized pustular psoriasis. N Engl J Med. 2019;380:981–983. doi: 10.1056/NEJMc1811317. [DOI] [PubMed] [Google Scholar]
- 24.EMA validates spesolimab marketing authorization application in GPP | Press [Internet]. boehringer-ingelheim.com [cited 2021 Nov 14]. https://www.boehringer-ingelheim.com/press-release/ema-filing-acceptance-and-validation-spesolimab.
- 25.Reich A, Barker J, Pink A, Reynolds NJ, Griffiths CEM, Kalapanda R, et al. Imsidolimab, an anti-IL-36 receptor monoclonal antibody, in the treatment of generalized pustular psoriasis: results from a phase 2 trial. 2021. p. 1–15.
- 26.Grossman RM, Krueger J, Yourish D, Granelli-Piperno A, Murphy DP, May LT, et al. Interleukin 6 is expressed in high levels in psoriatic skin and stimulates proliferation of cultured human keratinocytes. PNAS Natl Acad Sci. 1989;86:6367–6371. doi: 10.1073/pnas.86.16.6367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Palmou-Fontana N, Gaviño JAS, McGonagle D, García-Martinez E, Martín LIO. Tocilizumab-induced psoriasiform rash in rheumatoid arthritis. DRM. 2014;228:311–313. doi: 10.1159/000362266. [DOI] [PubMed] [Google Scholar]
- 28.Laurent S, Le Parc J-M, Clérici T, Bréban M, Mahé E. Onset of psoriasis following treatment with tocilizumab. Br J Dermatol. 2010;163:1364–1365. doi: 10.1111/j.1365-2133.2010.10005.x. [DOI] [PubMed] [Google Scholar]
- 29.Wendling D, Letho-Gyselinck H, Guillot X, Prati C. Psoriasis onset with tocilizumab treatment for rheumatoid arthritis. J Rheumatol. 2012;39:657. doi: 10.3899/jrheum.111166. [DOI] [PubMed] [Google Scholar]
- 30.Fritz Y, Klenotic PA, Swindell WR, Yin ZQ, Groft SG, Zhang L, et al. Induction of alternative proinflammatory cytokines accounts for sustained psoriasiform skin inflammation in IL-17C+IL-6KO mice. J Invest Dermatol. 2017;137:696–705. doi: 10.1016/j.jid.2016.10.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Blauvelt A. IL-6 differs from TNF-α: unpredicted clinical effects caused by IL-6 blockade in psoriasis. J Invest Dermatol. 2017;137:541–542. doi: 10.1016/j.jid.2016.11.022. [DOI] [PubMed] [Google Scholar]
- 32.Takematsu H, Tagami H. Quantification of chemotactic peptides (C5a anaphylatoxin and IL-8) in psoriatic lesional skin. Arch Dermatol. 1993;129:74–80. doi: 10.1001/archderm.1993.01680220086020. [DOI] [PubMed] [Google Scholar]
- 33.Duan H, Koga T, Kohda F, Hara H, Urabe K, Furue M. Interleukin-8-positive neutrophils in psoriasis. J Dermatol Sci. 2001;26:119–124. doi: 10.1016/S0923-1811(00)00167-5. [DOI] [PubMed] [Google Scholar]
- 34.Trepicchio WL, Ozawa M, Walters IB, Kikuchi T, Gilleaudeau P, Bliss JL, et al. Interleukin-11 therapy selectively downregulates type I cytokine proinflammatory pathways in psoriasis lesions. J Clin Invest. 1999;104:1527–1537. doi: 10.1172/JCI6910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Tonel G, Conrad C, Laggner U, Di Meglio P, Grys K, McClanahan TK, et al. Cutting edge: a critical functional role for IL-23 in psoriasis. J Immunol. 2010;185:5688–5691. doi: 10.4049/jimmunol.1001538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kulig P, Musiol S, Freiberger SN, Schreiner B, Gyülveszi G, Russo G, et al. IL-12 protects from psoriasiform skin inflammation. Nat Commun. 2016;7:13466. doi: 10.1038/ncomms13466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Diels J, Thilakarathne P, Cameron C, McElligott S, Schubert A, Puig L. Adjusted treatment COMPArisons between guSelkumab and uStekinumab for treatment of moderate-to-severe plaque psoriasis: the COMPASS analysis. Br J Dermatol. 2020;183:276–284. doi: 10.1111/bjd.18634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Papp KA, Blauvelt A, Bukhalo M, Gooderham M, Krueger JG, Lacour J-P, et al. Risankizumab versus ustekinumab for moderate-to-severe plaque psoriasis. N Engl J Med. 2017;376:1551–1560. doi: 10.1056/NEJMoa1607017. [DOI] [PubMed] [Google Scholar]
- 39.Lowes MA, Kikuchi T, Fuentes-Duculan J, Cardinale I, Zaba LC, Haider AS, et al. Psoriasis vulgaris lesions contain discrete populations of Th1 and Th17 T cells. J Invest Dermatol. 2008;128:1207–1211. doi: 10.1038/sj.jid.5701213. [DOI] [PubMed] [Google Scholar]
- 40.Blauvelt A, Chiricozzi A. The immunologic role of IL-17 in psoriasis and psoriatic arthritis pathogenesis. Clin Rev Allergy Immunol. 2018;55:379–390. doi: 10.1007/s12016-018-8702-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Stritesky GL, Yeh N, Kaplan MH. IL-23 promotes maintenance but not commitment to the Th17 lineage. J Immunol. 2008;181:5948–5955. doi: 10.4049/jimmunol.181.9.5948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Rizzo HL, Kagami S, Phillips KG, Kurtz SE, Jacques SL, Blauvelt A. IL-23-mediated psoriasis-like epidermal hyperplasia is dependent on IL-17A. J Immunol. 2011;186:1495–1502. doi: 10.4049/jimmunol.1001001. [DOI] [PubMed] [Google Scholar]
- 43.Reich K, Warren RB, Lebwohl M, Gooderham M, Strober B, Langley RG, et al. Bimekizumab versus secukinumab in plaque psoriasis. N Engl J Med. 2021;385:142–152. doi: 10.1056/NEJMoa2102383. [DOI] [PubMed] [Google Scholar]
- 44.Papp KA, Weinberg MA, Morris A, Reich K. IL17A/F nanobody sonelokimab in patients with plaque psoriasis: a multicentre, randomised, placebo-controlled, phase 2b study. Lancet. 2021;397:1564–1575. doi: 10.1016/S0140-6736(21)00440-2. [DOI] [PubMed] [Google Scholar]
- 45.Commins S, Steinke JW, Borish L. The extended IL-10 superfamily: IL-10, IL-19, IL-20, IL-22, IL-24, IL-26, IL-28, and IL-29. J Allergy Clin Immunol. 2008;121:1108–1111. doi: 10.1016/j.jaci.2008.02.026. [DOI] [PubMed] [Google Scholar]
- 46.Li H-H, Lin Y-C, Chen P-J, Hsiao C-H, Lee JY-Y, Chen W-C, et al. Interleukin-19 upregulates keratinocyte growth factor and is associated with psoriasis. Br J Dermatol. 2005;153:591–595. doi: 10.1111/j.1365-2133.2005.06665.x. [DOI] [PubMed] [Google Scholar]
- 47.Wei C-C, Chen W-Y, Wang Y-C, Chen P-J, Lee JY-Y, Wong T-W, et al. Detection of IL-20 and its receptors on psoriatic skin. Clin Immunol. 2005;117:65–72. doi: 10.1016/j.clim.2005.06.012. [DOI] [PubMed] [Google Scholar]
- 48.Kunz S, Wolk K, Witte E, Witte K, Doecke W-D, Volk H-D, et al. Interleukin (IL)-19, IL-20 and IL-24 are produced by and act on keratinocytes and are distinct from classical ILs. Exp Dermatol. 2006;15:991–1004. doi: 10.1111/j.1600-0625.2006.00516.x. [DOI] [PubMed] [Google Scholar]
- 49.Lebre MC, Jonckheere CL, Kraan MC, van Kuijk AW, Bos JD, de Rie M, et al. Expression of IL-20 in synovium and lesional skin of patients with psoriatic arthritis: differential response to alefacept treatment. Arthritis Res Ther. 2012;14:R200. doi: 10.1186/ar4038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Caruso R, Botti E, Sarra M, Esposito M, Stolfi C, Diluvio L, et al. Involvement of interleukin-21 in the epidermal hyperplasia of psoriasis. Nat Med. 2009;15:1013–1015. doi: 10.1038/nm.1995. [DOI] [PubMed] [Google Scholar]
- 51.Boniface K, Guignouard E, Pedretti N, Garcia M, Delwail A, Bernard F-X, et al. A role for T cell-derived interleukin 22 in psoriatic skin inflammation. Clin Exp Immunol. 2007;150:407–415. doi: 10.1111/j.1365-2249.2007.03511.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Wolk K, Witte E, Wallace E, Döcke W-D, Kunz S, Asadullah K, et al. IL-22 regulates the expression of genes responsible for antimicrobial defense, cellular differentiation, and mobility in keratinocytes: a potential role in psoriasis. Eur J Immunol. 2006;36:1309–1323. doi: 10.1002/eji.200535503. [DOI] [PubMed] [Google Scholar]
- 53.Pfizer. An ascending multiple dose study of the safety, tolerability, pharmacokinetics, pharmacodynamics, and clinical efficacy of ILV-094 administered subcutaneously or intravenously to subjects with psoriasis [Internet]. clinicaltrials.gov; 2011 Mar. Report No.: NCT00563524. https://clinicaltrials.gov/ct2/show/NCT00563524.
- 54.Wang F, Smith N, Maier L, Xia W, Hammerberg C, Chubb H, et al. Etanercept suppresses regenerative hyperplasia in psoriasis by acutely downregulating epidermal expression of interleukin (IL)-19, IL-20 and IL-24. Br J Dermatol. 2012;167:92–102. doi: 10.1111/j.1365-2133.2012.10961.x. [DOI] [PubMed] [Google Scholar]
- 55.Xu M, Lu H, Lee Y-H, Wu Y, Liu K, Shi Y, et al. An interleukin-25-mediated autoregulatory circuit in keratinocytes plays a pivotal role in psoriatic skin inflammation. Immunity. 2018;48:787–798.e4. doi: 10.1016/j.immuni.2018.03.019. [DOI] [PubMed] [Google Scholar]
- 56.Itoh T, Hatano R, Komiya E, Otsuka H, Narita Y, Aune TM, et al. Biological effects of IL-26 on T cell-mediated skin inflammation, including psoriasis. J Invest Dermatol. 2019;139:878–889. doi: 10.1016/j.jid.2018.09.037. [DOI] [PubMed] [Google Scholar]
- 57.Andrys C, Borska L, Pohl D, Fiala Z, Hamakova K, Krejsek J. Angiogenic activity in patients with psoriasis is significantly decreased by Goeckerman’s therapy. Arch Dermatol Res. 2007;298:479–483. doi: 10.1007/s00403-006-0723-8. [DOI] [PubMed] [Google Scholar]
- 58.Shibata S, Tada Y, Kanda N, Nashiro K, Kamata M, Karakawa M, et al. Possible roles of IL-27 in the pathogenesis of psoriasis. J Invest Dermatol. 2010;130:1034–1039. doi: 10.1038/jid.2009.349. [DOI] [PubMed] [Google Scholar]
- 59.Omar NS, Long X, Xian J, Afewerky HK, Hussain SG, Peng X. Serum interleukin-30 level in patients with psoriasis and its correlation with psoriasis severity: a case-control study. J Int Med Res. 2021;49:3000605211004039. doi: 10.1177/03000605211004039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Mitsui A, Tada Y, Takahashi T, Shibata S, Kamata M, Miyagaki T, et al. Serum IL-33 levels are increased in patients with psoriasis. Clin Exp Dermatol. 2016;41:183–189. doi: 10.1111/ced.12670. [DOI] [PubMed] [Google Scholar]
- 61.Wolk K, Witte K, Witte E, Raftery M, Kokolakis G, Philipp S, et al. IL-29 is produced by T(H)17 cells and mediates the cutaneous antiviral competence in psoriasis. Sci Transl Med. 2013;5:204ra129. doi: 10.1126/scitranslmed.3006245. [DOI] [PubMed] [Google Scholar]
- 62.Witte E, Kokolakis G, Witte K, Warszawska K, Friedrich M, Christou D, et al. Interleukin-29 induces epithelial production of CXCR3A ligands and T-cell infiltration. J Mol Med (Berl) 2016;94:391–400. doi: 10.1007/s00109-015-1367-y. [DOI] [PubMed] [Google Scholar]
- 63.Flier J, Boorsma DM, van Beek PJ, Nieboer C, Stoof TJ, Willemze R, et al. Differential expression of CXCR3 targeting chemokines CXCL10, CXCL9, and CXCL11 in different types of skin inflammation. J Pathol. 2001;194:398–405. doi: 10.1002/1096-9896(200108)194:4<397::AID-PATH899>3.0.CO;2-S. [DOI] [PubMed] [Google Scholar]
- 64.Kempuraj D, Conti P, Vasiadi M, Alysandratos K-D, Tagen M, Kalogeromitros D, et al. IL-32 is increased along with tryptase in lesional psoriatic skin and is up-regulated by substance P in human mast cells. Eur J Dermatol. 2010;20:865–867. doi: 10.1684/ejd.2010.1110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Li J, Liu L, Rui W, Li X, Xuan D, Zheng S, et al. New interleukins in psoriasis and psoriatic arthritis patients: the possible roles of interleukin-33 to interleukin-38 in disease activities and bone erosions. Dermatology. 2017;233:37–46. doi: 10.1159/000471798. [DOI] [PubMed] [Google Scholar]
- 66.Teng X, Hu Z, Wei X, Wang Z, Guan T, Liu N, et al. IL-37 ameliorates the inflammatory process in psoriasis by suppressing proinflammatory cytokine production. J Immunol. 2014;192:1815–1823. doi: 10.4049/jimmunol.1300047. [DOI] [PubMed] [Google Scholar]
- 67.Mercurio L, Morelli M, Scarponi C, Eisenmesser EZ, Doti N, Pagnanelli G, et al. IL-38 has an anti-inflammatory action in psoriasis and its expression correlates with disease severity and therapeutic response to anti-IL-17A treatment. Cell Death Dis. 2018;9:1–13. doi: 10.1038/s41419-018-1143-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Ständer S. Atopic dermatitis. N Engl J Med. 2021;384:1136–1143. doi: 10.1056/NEJMra2023911. [DOI] [PubMed] [Google Scholar]
- 69.Kim J, Kim BE, Leung DYM. Pathophysiology of atopic dermatitis: clinical implications. Allergy Asthma Proc. 2019;40:84–92. doi: 10.2500/aap.2019.40.4202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Simpson EL, Bieber T, Guttman-Yassky E, Beck LA, Blauvelt A, Cork MJ, et al. Two phase 3 trials of dupilumab versus placebo in atopic dermatitis. N Engl J Med. 2016;375:2335–2348. doi: 10.1056/NEJMoa1610020. [DOI] [PubMed] [Google Scholar]
- 71.Kim BE, Leung DYM, Boguniewicz M, Howell MD. Loricrin and involucrin expression is down-regulated by Th2 cytokines through STAT-6. Clin Immunol. 2008;126:332–337. doi: 10.1016/j.clim.2007.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Howell MD, Kim BE, Gao P, Grant AV, Boguniewicz M, Debenedetto A, et al. Cytokine modulation of atopic dermatitis filaggrin skin expression. J Allergy Clin Immunol. 2007;120:150–155. doi: 10.1016/j.jaci.2007.04.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Oetjen LK, Mack MR, Feng J, Whelan TM, Niu H, Guo CJ, et al. Sensory neurons co-opt classical immune signaling pathways to mediate chronic itch. Cell. 2017;171:217–228.e13. doi: 10.1016/j.cell.2017.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Foster EL, Simpson EL, Fredrikson LJ, Lee JJ, Lee NA, Fryer AD, et al. Eosinophils increase neuron branching in human and murine skin and in vitro. PLoS ONE. 2011;6:e22029. doi: 10.1371/journal.pone.0022029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Ma L, Xue H-B, Guan X-H, Shu C-M, Zhang J-H, Yu J. Possible pathogenic role of T helper type 9 cells and interleukin (IL)-9 in atopic dermatitis. Clin Exp Immunol. 2014;175:25–31. doi: 10.1111/cei.12198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Toda M, Leung DYM, Molet S, Boguniewicz M, Taha R, Christodoulopoulos P, et al. Polarized in vivo expression of IL-11 and IL-17 between acute and chronic skin lesions. J Allergy Clin Immunol. 2003;111:875–881. doi: 10.1067/mai.2003.1414. [DOI] [PubMed] [Google Scholar]
- 77.Nograles KE, Zaba LC, Shemer A, Fuentes-Duculan J, Cardinale I, Kikuchi T, et al. IL-22-producing “T22” T cells account for upregulated IL-22 in atopic dermatitis despite reduced IL-17-producing TH17 T cells. J Allergy Clin Immunol. 2009;123:1244–1252.e2. doi: 10.1016/j.jaci.2009.03.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Sonkoly E, Muller A, Lauerma AI, Pivarcsi A, Soto H, Kemeny L, et al. IL-31: a new link between T cells and pruritus in atopic skin inflammation. J Allergy Clin Immunol. 2006;117:411–417. doi: 10.1016/j.jaci.2005.10.033. [DOI] [PubMed] [Google Scholar]
- 79.Arai I, Tsuji M, Takeda H, Akiyama N, Saito S. A single dose of interleukin-31 (IL-31) causes continuous itch-associated scratching behaviour in mice. Exp Dermatol. 2013;22:669–671. doi: 10.1111/exd.12222. [DOI] [PubMed] [Google Scholar]
- 80.Tamagawa-Mineoka R, Okuzawa Y, Masuda K, Katoh N. Increased serum levels of interleukin 33 in patients with atopic dermatitis. J Am Acad Dermatol. 2014;70:882–888. doi: 10.1016/j.jaad.2014.01.867. [DOI] [PubMed] [Google Scholar]
- 81.Imai Y, Yasuda K, Sakaguchi Y, Haneda T, Mizutani H, Yoshimoto T, et al. Skin-specific expression of IL-33 activates group 2 innate lymphoid cells and elicits atopic dermatitis-like inflammation in mice. Proc Natl Acad Sci USA. 2013;110:13921–13926. doi: 10.1073/pnas.1307321110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Lilly’s lebrikizumab significantly improved skin clearance and itch in people with moderate-to-severe atopic dermatitis in two Phase 3 trials | Eli Lilly and Company [Internet]. [cited 2021 Nov 15]. https://investor.lilly.com/news-releases/news-release-details/lillys-lebrikizumab-significantly-improved-skin-clearance-and.
- 83.Wollenberg A, Blauvelt A, Guttman-Yassky E, Worm M, Lynde C, Lacour J-P, et al. Tralokinumab for moderate-to-severe atopic dermatitis: results from two 52-week, randomized, double-blind, multicentre, placebo-controlled phase III trials (ECZTRA 1 and ECZTRA 2) Br J Dermatol. 2021;184:437–449. doi: 10.1111/bjd.19574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Kabashima K, Matsumura T, Komazaki H, Kawashima M, Nemolizumab-JP01 Study Group Trial of nemolizumab and topical agents for atopic dermatitis with pruritus. N Engl J Med. 2020;383:141–150. doi: 10.1056/NEJMoa1917006. [DOI] [PubMed] [Google Scholar]
- 85.Febbraio MA. Role of interleukins in obesity: implications for metabolic disease. Trends Endocrinol Metab. 2014;25:312–319. doi: 10.1016/j.tem.2014.02.004. [DOI] [PubMed] [Google Scholar]
- 86.Larsen CM, Faulenbach M, Vaag A, Vølund A, Ehses JA, Seifert B, et al. Interleukin-1–receptor antagonist in type 2 diabetes mellitus. N Engl J Med. 2007;356:1517–1526. doi: 10.1056/NEJMoa065213. [DOI] [PubMed] [Google Scholar]
- 87.Ridker PM, Howard CP, Walter V, Everett B, Libby P, Hensen J, et al. Effects of interleukin-1β inhibition with canakinumab on hemoglobin A1c, lipids, c-reactive protein, interleukin-6, and fibrinogen. Circulation. 2012;126:2739–2748. doi: 10.1161/CIRCULATIONAHA.112.122556. [DOI] [PubMed] [Google Scholar]
- 88.Ridker PM, Everett BM, Thuren T, MacFadyen JG, Chang WH, Ballantyne C, et al. Antiinflammatory therapy with canakinumab for atherosclerotic disease. N Engl J Med. 2017;377:1119–1131. doi: 10.1056/NEJMoa1707914. [DOI] [PubMed] [Google Scholar]
- 89.Everett BM, Donath MY, Pradhan AD, Thuren T, Pais P, Nicolau JC, et al. Anti-inflammatory therapy with canakinumab for the prevention and management of diabetes. J Am Coll Cardiol. 2018;71:2392–2401. doi: 10.1016/j.jacc.2018.03.002. [DOI] [PubMed] [Google Scholar]
- 90.Duerr RH, Taylor KD, Brant SR, Rioux JD, Silverberg MS, Daly MJ, et al. A genome-wide association study identifies IL23R as an inflammatory bowel disease gene. Science. 2006;314:1461–1463. doi: 10.1126/science.1135245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Fujino S, Andoh A, Bamba S, Ogawa A, Hata K, Araki Y, et al. Increased expression of interleukin 17 in inflammatory bowel disease. Gut. 2003;52:65–70. doi: 10.1136/gut.52.1.65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Kleinschek MA, Boniface K, Sadekova S, Grein J, Murphy EE, Turner SP, et al. Circulating and gut-resident human Th17 cells express CD161 and promote intestinal inflammation. J Exp Med. 2009;206:525–534. doi: 10.1084/jem.20081712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Liu Z, Yadav PK, Xu X, Su J, Chen C, Tang M, et al. The increased expression of IL-23 in inflammatory bowel disease promotes intraepithelial and lamina propria lymphocyte inflammatory responses and cytotoxicity. J Leukoc Biol. 2011;89:597–606. doi: 10.1189/jlb.0810456. [DOI] [PubMed] [Google Scholar]
- 94.Feagan BG, Sandborn WJ, Gasink C, Jacobstein D, Lang Y, Friedman JR, et al. Ustekinumab as induction and maintenance therapy for Crohn’s disease. N Engl J Med. 2016;375:1946–1960. doi: 10.1056/NEJMoa1602773. [DOI] [PubMed] [Google Scholar]
- 95.Ferreira BIRC, Abreu JLPDC, Reis JPGD, Figueiredo AMDC. Psoriasis and associated psychiatric disorders. J Clin Aesthet Dermatol. 2016;9:36–43. [PMC free article] [PubMed] [Google Scholar]
- 96.Heller MM, Lee ES, Koo JY. Stress as an influencing factor in psoriasis. Skin Ther Lett. 2011;16:1–4. [PubMed] [Google Scholar]
- 97.Reich A, Hrehorów E, Szepietowski JC. Pruritus is an important factor negatively influencing the well-being of psoriatic patients. Acta Derm Venereol. 2010;90:257–263. doi: 10.2340/00015555-0851. [DOI] [PubMed] [Google Scholar]
- 98.Janković S, Raznatović M, Marinković J, Maksimović N, Janković J, Djikanović B. Relevance of psychosomatic factors in psoriasis: a case-control study. Acta Derm Venereol. 2009;89:364–368. doi: 10.2340/00015555-0669. [DOI] [PubMed] [Google Scholar]
- 99.Devrimci-Ozguven H, Kundakci TN, Kumbasar H, Boyvat A. The depression, anxiety, life satisfaction and affective expression levels in psoriasis patients. J Eur Acad Dermatol Venereol. 2000;14:267–271. doi: 10.1046/j.1468-3083.2000.00085.x. [DOI] [PubMed] [Google Scholar]
- 100.Mizara A, Papadopoulos L, McBride SR. Core beliefs and psychological distress in patients with psoriasis and atopic eczema attending secondary care: the role of schemas in chronic skin disease. Br J Dermatol. 2012;166:986–993. doi: 10.1111/j.1365-2133.2011.10799.x. [DOI] [PubMed] [Google Scholar]
- 101.Garg A, Chren MM, Sands LP, Matsui MS, Marenus KD, Feingold KR, et al. Psychological stress perturbs epidermal permeability barrier homeostasis: implications for the pathogenesis of stress-associated skin disorders. Arch Dermatol. 2001;137:53–59. doi: 10.1001/archderm.137.1.53. [DOI] [PubMed] [Google Scholar]
- 102.Arnetz BB, Fjellner B, Eneroth P, Kallner A. Stress and psoriasis: psychoendocrine and metabolic reactions in psoriatic patients during standardized stressor exposure. Psychosom Med. 1985;47:528–541. doi: 10.1097/00006842-198511000-00003. [DOI] [PubMed] [Google Scholar]
- 103.Buske-Kirschbaum A, Ebrecht M, Kern S, Hellhammer DH. Endocrine stress responses in TH1-mediated chronic inflammatory skin disease (psoriasis vulgaris)—do they parallel stress-induced endocrine changes in TH2-mediated inflammatory dermatoses (atopic dermatitis)? Psychoneuroendocrinology. 2006;31:439–446. doi: 10.1016/j.psyneuen.2005.10.006. [DOI] [PubMed] [Google Scholar]
- 104.Ganança L, Oquendo MA, Tyrka AR, Cisneros-Trujillo S, Mann JJ, Sublette ME. The role of cytokines in the pathophysiology of suicidal behavior. Psychoneuroendocrinology. 2016;63:296–310. doi: 10.1016/j.psyneuen.2015.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Alesci S, Martinez PE, Kelkar S, Ilias I, Ronsaville DS, Listwak SJ, et al. Major depression is associated with significant diurnal elevations in plasma interleukin-6 levels, a shift of its circadian rhythm, and loss of physiological complexity in its secretion: clinical implications. J Clin Endocrinol Metab. 2005;90:2522–2530. doi: 10.1210/jc.2004-1667. [DOI] [PubMed] [Google Scholar]
- 106.Tuglu C, Kara SH, Caliyurt O, Vardar E, Abay E. Increased serum tumor necrosis factor-alpha levels and treatment response in major depressive disorder. Psychopharmacology. 2003;170:429–433. doi: 10.1007/s00213-003-1566-z. [DOI] [PubMed] [Google Scholar]
- 107.Owen BM, Eccleston D, Ferrier IN, Young AH. Raised levels of plasma interleukin-1beta in major and postviral depression. Acta Psychiatr Scand. 2001;103:226–228. doi: 10.1034/j.1600-0447.2001.00162.x. [DOI] [PubMed] [Google Scholar]
- 108.Saraceno R, Kleyn CE, Terenghi G, Griffiths CEM. The role of neuropeptides in psoriasis. Br J Dermatol. 2006;155:876–882. doi: 10.1111/j.1365-2133.2006.07518.x. [DOI] [PubMed] [Google Scholar]
- 109.Turka LA, Goodman RE, Rutkowski JL, Sima AA, Merry A, Mitra RS, et al. Interleukin 12: a potential link between nerve cells and the immune response in inflammatory disorders. Mol Med. 1995;1:690–699. doi: 10.1007/BF03401609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Koo J, Marangell LB, Nakamura M, Armstrong A, Jeon C, Bhutani T, et al. Depression and suicidality in psoriasis: review of the literature including the cytokine theory of depression. J Eur Acad Dermatol Venereol. 2017;31:1999–2009. doi: 10.1111/jdv.14460. [DOI] [PubMed] [Google Scholar]
- 111.Maddison DC, Giorgini F. The kynurenine pathway and neurodegenerative disease. Semin Cell Dev Biol. 2015;40:134–141. doi: 10.1016/j.semcdb.2015.03.002. [DOI] [PubMed] [Google Scholar]
- 112.Sublette ME, Galfalvy HC, Fuchs D, Lapidus M, Grunebaum MF, Oquendo MA, et al. Plasma kynurenine levels are elevated in suicide attempters with major depressive disorder. Brain Behav Immun. 2011;25:1272–1278. doi: 10.1016/j.bbi.2011.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Jesus AA, Goldbach-Mansky R. IL-1 blockade in autoinflammatory syndromes. Annu Rev Med. 2014;65:223–244. doi: 10.1146/annurev-med-061512-150641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Witte-Händel E, Wolk K, Tsaousi A, Irmer ML, Mößner R, Shomroni O, et al. The IL-1 pathway is hyperactive in hidradenitis suppurativa and contributes to skin infiltration and destruction. J Invest Dermatol. 2019;139:1294–1305. doi: 10.1016/j.jid.2018.11.018. [DOI] [PubMed] [Google Scholar]
- 115.Zhou J, Luo Q, Cheng Y, Wen X, Liu J. An update on genetic basis of generalized pustular psoriasis (review) Int J Mol Med. 2021;47:1–12. doi: 10.3892/ijmm.2021.4951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Dinarello CA, Simon A, van der Meer JWM. Treating inflammation by blocking interleukin-1 in a broad spectrum of diseases. Nat Rev Drug Discov. 2012;11:633–652. doi: 10.1038/nrd3800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Dusser P, Koné-Paut I. IL-1 inhibition may have an important role in treating refractory Kawasaki disease. Front Pharmacol. 2017;8:1–8. doi: 10.3389/fphar.2017.00163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Aouba A, Georgin-Lavialle S, Pagnoux C, Martin Silva N, Renand A, Galateau-Salle F, et al. Rationale and efficacy of interleukin-1 targeting in Erdheim–Chester disease. Blood. 2010;116:4070–4076. doi: 10.1182/blood-2010-04-279240. [DOI] [PubMed] [Google Scholar]
- 119.Castela E, Le Duff F, Butori C, Ticchioni M, Hofman P, Bahadoran P, et al. Effects of low-dose recombinant interleukin 2 to promote T-regulatory cells in alopecia areata. JAMA Dermatol. 2014;150:748–751. doi: 10.1001/jamadermatol.2014.504. [DOI] [PubMed] [Google Scholar]
- 120.Atkins MB, Kunkel L, Sznol M, Rosenberg SA. High-dose recombinant interleukin-2 therapy in patients with metastatic melanoma: long-term survival update. Cancer J Sci Am. 2000;6(Suppl 1):S11–14. [PubMed] [Google Scholar]
- 121.Chiricozzi A, Maurelli M, Peris K, Girolomoni G. Targeting IL-4 for the treatment of atopic dermatitis. Immunotargets Ther. 2020;9:151–156. doi: 10.2147/ITT.S260370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Salmon-Ehr V, Serpier H, Nawrocki B, Gillery P, Clavel C, Kalis B, et al. Expression of interleukin-4 in scleroderma skin specimens and scleroderma fibroblast cultures. Potential role in fibrosis. Arch Dermatol. 1996;132:802–806. doi: 10.1001/archderm.1996.03890310088013. [DOI] [PubMed] [Google Scholar]
- 123.Schmidt E, Bastian B, Dummer R, Tony HP, Bröcker EB, Zillikens D. Detection of elevated levels of IL-4, IL-6, and IL-10 in blister fluid of bullous pemphigoid. Arch Dermatol Res. 1996;288:353–357. doi: 10.1007/BF02507102. [DOI] [PubMed] [Google Scholar]
- 124.Abdat R, Waldman RA, de Bedout V, Czernik A, Mcleod M, King B, et al. Dupilumab as a novel therapy for bullous pemphigoid: a multicenter case series. J Am Acad Dermatol. 2020;83:46–52. doi: 10.1016/j.jaad.2020.01.089. [DOI] [PubMed] [Google Scholar]
- 125.Roufosse F. Targeting the interleukin-5 pathway for treatment of eosinophilic conditions other than asthma. Front Med (Lausanne) 2018;5:1–27. doi: 10.3389/fmed.2018.00049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Wechsler ME, Akuthota P, Jayne D, Khoury P, Klion A, Langford CA, et al. Mepolizumab or placebo for eosinophilic granulomatosis with polyangiitis. N Engl J Med. 2017;376:1921–1932. doi: 10.1056/NEJMoa1702079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Nakajima S, Tie D, Nomura T, Kabashima K. Novel pathogenesis of atopic dermatitis from the view of cytokines in mice and humans. Cytokine. 2021;148:155664. doi: 10.1016/j.cyto.2021.155664. [DOI] [PubMed] [Google Scholar]
- 128.Xu H, Xiao X, He Y, Zhang X, Li C, Mao Q, et al. Increased serum interleukin-6 levels in patients with hidradenitis suppurativa. Postepy Dermatol Alergol. 2017;34:82–84. doi: 10.5114/ada.2017.65626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Ogata A, Kumanogoh A, Tanaka T. Pathological role of interleukin-6 in psoriatic arthritis. Arthritis. 2012;2012:1–6. doi: 10.1155/2012/713618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Ting EY-C, Yang AC, Tsai S-J. Role of interleukin-6 in depressive disorder. Int J Mol Sci. 2020;21:1–22. doi: 10.3390/ijms21062194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Lin Z-Q, Kondo T, Ishida Y, Takayasu T, Mukaida N. Essential involvement of IL-6 in the skin wound-healing process as evidenced by delayed wound healing in IL-6-deficient mice. J Leukoc Biol. 2003;73:713–721. doi: 10.1189/jlb.0802397. [DOI] [PubMed] [Google Scholar]
- 132.Markousis-Mavrogenis G, Tromp J, Ouwerkerk W, Devalaraja M, Anker SD, Cleland JG, et al. The clinical significance of interleukin-6 in heart failure: results from the BIOSTAT-CHF study. Eur J Heart Fail. 2019;21:965–973. doi: 10.1002/ejhf.1482. [DOI] [PubMed] [Google Scholar]
- 133.Pradhan AD, Manson JE, Rifai N, Buring JE, Ridker PM. C-reactive protein, interleukin 6, and risk of developing type 2 diabetes mellitus. JAMA. 2001;286:327–334. doi: 10.1001/jama.286.3.327. [DOI] [PubMed] [Google Scholar]
- 134.Yamanaka K, Clark R, Rich B, Dowgiert R, Hirahara K, Hurwitz D, et al. Skin-derived interleukin-7 contributes to the proliferation of lymphocytes in cutaneous T-cell lymphoma. Blood. 2006;107:2440–2445. doi: 10.1182/blood-2005-03-1139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Horev L, Unger S, Molho-Pessach V, Meir T, Maly A, Stepensky P, et al. Generalized verrucosis and HPV-3 susceptibility associated with CD4 T-cell lymphopenia caused by inherited human interleukin-7 deficiency. J Am Acad Dermatol. 2015;72:1082–1084. doi: 10.1016/j.jaad.2015.02.1118. [DOI] [PubMed] [Google Scholar]
- 136.Gregory SG, Schmidt S, Seth P, Oksenberg JR, Hart J, Prokop A, et al. Interleukin 7 receptor alpha chain (IL7R) shows allelic and functional association with multiple sclerosis. Nat Genet. 2007;39:1083–1091. doi: 10.1038/ng2103. [DOI] [PubMed] [Google Scholar]
- 137.Zenatti PP, Ribeiro D, Li W, Zuurbier L, Silva MC, Paganin M, et al. Oncogenic IL7R gain-of-function mutations in childhood T-cell acute lymphoblastic leukemia. Nat Genet. 2011;43:932–939. doi: 10.1038/ng.924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Bikker A, van Woerkom JM, Kruize AA, Wenting-van Wijk M, de Jager W, Bijlsma JWJ, et al. Increased expression of interleukin-7 in labial salivary glands of patients with primary Sjögren’s syndrome correlates with increased inflammation. Arthritis Rheum. 2010;62:969–977. doi: 10.1002/art.27318. [DOI] [PubMed] [Google Scholar]
- 139.Herold KC, Bucktrout SL, Wang X, Bode BW, Gitelman SE, Gottlieb PA, et al. Immunomodulatory activity of humanized anti-IL-7R monoclonal antibody RN168 in subjects with type 1 diabetes. JCI Insight. 2019;4:126054. doi: 10.1172/jci.insight.126054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Ellis J, van Maurik A, Fortunato L, Gisbert S, Chen K, Schwartz A, et al. Anti-IL-7 receptor α monoclonal antibody (GSK2618960) in healthy subjects—a randomized, double-blind, placebo-controlled study. Br J Clin Pharmacol. 2019;85:304–315. doi: 10.1111/bcp.13748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Oka M, Berking C, Nesbit M, Satyamoorthy K, Schaider H, Murphy G, et al. Interleukin-8 overexpression is present in pyoderma gangrenosum ulcers and leads to ulcer formation in human skin xenografts. Lab Invest. 2000;80:595–604. doi: 10.1038/labinvest.3780064. [DOI] [PubMed] [Google Scholar]
- 142.Bilusic M, Heery CR, Collins JM, Donahue RN, Palena C, Madan RA, et al. Phase I trial of HuMax-IL8 (BMS-986253), an anti-IL-8 monoclonal antibody, in patients with metastatic or unresectable solid tumors. J Immunother Cancer. 2019;7:1–8. doi: 10.1186/s40425-019-0706-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Daig R, Andus T, Aschenbrenner E, Falk W, Schölmerich J, Gross V. Increased interleukin 8 expression in the colon mucosa of patients with inflammatory bowel disease. Gut. 1996;38:216–222. doi: 10.1136/gut.38.2.216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Liu J, Harberts E, Tammaro A, Girardi N, Filler RB, Fishelevich R, et al. IL-9 regulates allergen-specific Th1 responses in allergic contact dermatitis. J Invest Dermatol. 2014;134:1903–1911. doi: 10.1038/jid.2014.61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Gutin L, Tammaro A, Fishelevich R, Gaspari AA. Elevation of IL-9 in extreme patch test reactions suggests it is an inflammatory mediator in allergic contact dermatitis. Dermatitis. 2016;27:35–36. doi: 10.1097/DER.0000000000000159. [DOI] [PubMed] [Google Scholar]
- 146.Vieyra-Garcia PA, Wei T, Naym DG, Fredholm S, Fink-Puches R, Cerroni L, et al. STAT3/5-dependent IL9 overexpression contributes to neoplastic cell survival in mycosis fungoides. Clin Cancer Res. 2016;22:3328–3339. doi: 10.1158/1078-0432.CCR-15-1784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Schlapbach C, Gehad A, Yang C, Watanabe R, Guenova E, Teague JE, et al. Human TH9 cells are skin-tropic and have autocrine and paracrine proinflammatory capacity. Sci Transl Med. 2014;6:219ra8. doi: 10.1126/scitranslmed.3007828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Weiss E, Mamelak AJ, La Morgia S, Wang B, Feliciani C, Tulli A, et al. The role of interleukin 10 in the pathogenesis and potential treatment of skin diseases. J Am Acad Dermatol. 2004;50:657–675. doi: 10.1016/j.jaad.2003.11.075. [DOI] [PubMed] [Google Scholar]
- 149.Shi J, Li J, Guan H, Cai W, Bai X, Fang X, et al. Anti-fibrotic actions of interleukin-10 against hypertrophic scarring by activation of PI3K/AKT and STAT3 signaling pathways in scar-forming fibroblasts. PLoS ONE. 2014;9:e98228. doi: 10.1371/journal.pone.0098228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Schafer S, Viswanathan S, Widjaja AA, Lim W-W, Moreno-Moral A, DeLaughter DM, et al. IL-11 is a crucial determinant of cardiovascular fibrosis. Nature. 2017;552:110–115. doi: 10.1038/nature24676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Meeran SM, Mantena SK, Meleth S, Elmets CA, Katiyar SK. Interleukin-12-deficient mice are at greater risk of UV radiation-induced skin tumors and malignant transformation of papillomas to carcinomas. Mol Cancer Ther. 2006;5:825–832. doi: 10.1158/1535-7163.MCT-06-0003. [DOI] [PubMed] [Google Scholar]
- 152.Gee K, Guzzo C, Che Mat NF, Ma W, Kumar A. The IL-12 family of cytokines in infection, inflammation and autoimmune disorders. Inflamm Allergy Drug Targets. 2009;8:40–52. doi: 10.2174/187152809787582507. [DOI] [PubMed] [Google Scholar]
- 153.Hamid QA, Schotman E, Jacobson MR, Walker SM, Durham SR. Increases in IL-12 messenger RNA+ cells accompany inhibition of allergen-induced late skin responses after successful grass pollen immunotherapy. J Allergy Clin Immunol. 1997;99:254–260. doi: 10.1016/S0091-6749(97)70106-4. [DOI] [PubMed] [Google Scholar]
- 154.Biedermann T, Lametschwandtner G, Tangemann K, Kund J, Hinteregger S, Carballido-Perrig N, et al. IL-12 instructs skin homing of human Th2 cells. J Immunol. 2006;177:3763–3770. doi: 10.4049/jimmunol.177.6.3763. [DOI] [PubMed] [Google Scholar]
- 155.Hamid Q, Naseer T, Minshall EM, Song YL, Boguniewicz M, Leung DY. In vivo expression of IL-12 and IL-13 in atopic dermatitis. J Allergy Clin Immunol. 1996;98:225–231. doi: 10.1016/S0091-6749(96)70246-4. [DOI] [PubMed] [Google Scholar]
- 156.Tsoi LC, Rodriguez E, Degenhardt F, Baurecht H, Wehkamp U, Volks N, et al. Atopic dermatitis Is an IL-13-dominant disease with greater molecular heterogeneity compared to psoriasis. J Invest Dermatol. 2019;139:1480–1489. doi: 10.1016/j.jid.2018.12.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Aridogan BC, Yildirim M, Baysal V, Inaloz HS, Baz K, Kaya S. Serum levels of IL-4, IL-10, IL-12, IL-13 and IFN-gamma in Behçet’s disease. J Dermatol. 2003;30:602–607. doi: 10.1111/j.1346-8138.2003.tb00442.x. [DOI] [PubMed] [Google Scholar]
- 158.Hasegawa M, Fujimoto M, Kikuchi K, Takehara K. Elevated serum levels of interleukin 4 (IL-4), IL-10, and IL-13 in patients with systemic sclerosis. J Rheumatol. 1997;24:328–332. [PubMed] [Google Scholar]
- 159.Hasegawa M, Sato S, Nagaoka T, Fujimoto M, Takehara K. Serum levels of tumor necrosis factor and interleukin-13 are elevated in patients with localized scleroderma. Dermatology. 2003;207:141–147. doi: 10.1159/000071783. [DOI] [PubMed] [Google Scholar]
- 160.Korenblat P, Kerwin E, Leshchenko I, Yen K, Holweg CTJ, Anzures-Cabrera J, et al. Efficacy and safety of lebrikizumab in adult patients with mild-to-moderate asthma not receiving inhaled corticosteroids. Respir Med. 2018;134:143–149. doi: 10.1016/j.rmed.2017.12.006. [DOI] [PubMed] [Google Scholar]
- 161.Guttman-Yassky E, Blauvelt A, Eichenfield LF, Paller AS, Armstrong AW, Drew J, et al. Efficacy and safety of lebrikizumab, a high-affinity interleukin 13 inhibitor, in adults with moderate to severe atopic dermatitis: a phase 2b randomized clinical trial. JAMA Dermatol. 2020;156:411–420. doi: 10.1001/jamadermatol.2020.0079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Panettieri RA, Sjöbring U, Péterffy A, Wessman P, Bowen K, Piper E, et al. Tralokinumab for severe, uncontrolled asthma (STRATOS 1 and STRATOS 2): two randomised, double-blind, placebo-controlled, phase 3 clinical trials. Lancet Respir Med. 2018;6:511–525. doi: 10.1016/S2213-2600(18)30184-X. [DOI] [PubMed] [Google Scholar]
- 163.Shen L, Suresh L, Li H, Zhang C, Kumar V, Pankewycz O, et al. IL-14 alpha, the nexus for primary Sjögren’s disease in mice and humans. Clin Immunol. 2009;130:304–312. doi: 10.1016/j.clim.2008.10.006. [DOI] [PubMed] [Google Scholar]
- 164.Kiratiseavee S, Shen L, Baer AN, Ambrus JL. IL-14 RNA levels in the peripheral blood leukocytes of patients with SLE. J Allergy Clin Immunol. 2006;117:S18. doi: 10.1016/j.jaci.2005.12.076. [DOI] [Google Scholar]
- 165.Fehniger TA, Caligiuri MA. Interleukin 15: biology and relevance to human disease. Blood. 2001;97:14–32. doi: 10.1182/blood.V97.1.14. [DOI] [PubMed] [Google Scholar]
- 166.Richmond JM, Strassner JP, Zapata L, Garg M, Riding RL, Refat MA, et al. Antibody blockade of IL-15 signaling has the potential to durably reverse vitiligo. Sci Transl Med. 2018;10:eaam7710. doi: 10.1126/scitranslmed.aam7710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Willerslev-Olsen A, Litvinov IV, Fredholm SM, Petersen DL, Sibbesen NA, Gniadecki R, et al. IL-15 and IL-17F are differentially regulated and expressed in mycosis fungoides (MF) Cell Cycle. 2014;13:1306–1312. doi: 10.4161/cc.28256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Chamie K, Chang S, Gonzalgo ML, Kramolowsky EV, Sexton WJ, Reddy SK, et al. Phase II/III clinical results of IL-15RαFc superagonist N-803 with BCG in BCG-unresponsive non-muscle invasive bladder cancer (NMIBC) carcinoma in situ (CIS) patients. JCO. 2021;39:510–510. doi: 10.1200/JCO.2021.39.6_suppl.510. [DOI] [Google Scholar]
- 169.Asadullah K, Haeussler-Quade A, Gellrich S, Hanneken S, Hansen-Hagge TE, Döcke WD, et al. IL-15 and IL-16 overexpression in cutaneous T-cell lymphomas: stage-dependent increase in mycosis fungoides progression. Exp Dermatol. 2000;9:248–251. doi: 10.1034/j.1600-0625.2000.009004248.x. [DOI] [PubMed] [Google Scholar]
- 170.Kawabata K, Makino T, Makino K, Kajihara I, Fukushima S, Ihn H. IL-16 expression is increased in the skin and sera of patients with systemic sclerosis. Rheumatology (Oxford) 2020;59:519–523. doi: 10.1093/rheumatology/kez318. [DOI] [PubMed] [Google Scholar]
- 171.Frezzolini A, Cianchini G, Ruffelli M, Cadoni S, Puddu P, De Pità O. Interleukin-16 expression and release in bullous pemphigoid. Clin Exp Immunol. 2004;137:595–600. doi: 10.1111/j.1365-2249.2004.02570.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Lard LR, Roep BO, Verburgh CA, Zwinderman AH, Huizinga TWJ. Elevated IL-16 levels in patients with systemic lupus erythematosus are associated with disease severity but not with genetic susceptibility to lupus. Lupus. 2002;11:181–185. doi: 10.1191/0961203302lu176sr. [DOI] [PubMed] [Google Scholar]
- 173.Amiel C, Darcissac E, Truong MJ, Dewulf J, Loyens M, Mouton Y, et al. Interleukin-16 (IL-16) inhibits human immunodeficiency virus replication in cells from infected subjects, and serum IL-16 levels drop with disease progression. J Infect Dis. 1999;179:83–91. doi: 10.1086/314550. [DOI] [PubMed] [Google Scholar]
- 174.Bowness P, Ridley A, Shaw J, Chan AT, Wong-Baeza I, Fleming M, et al. Th17 cells expressing KIR3DL2+ and responsive to HLA-B27 homodimers are increased in ankylosing spondylitis. J Immunol. 2011;186:2672–2680. doi: 10.4049/jimmunol.1002653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Molet S, Hamid Q, Davoine F, Nutku E, Taha R, Pagé N, et al. IL-17 is increased in asthmatic airways and induces human bronchial fibroblasts to produce cytokines. J Allergy Clin Immunol. 2001;108:430–438. doi: 10.1067/mai.2001.117929. [DOI] [PubMed] [Google Scholar]
- 176.Hohenberger M, Cardwell LA, Oussedik E, Feldman SR. Interleukin-17 inhibition: role in psoriasis and inflammatory bowel disease. J Dermatolog Treat. 2018;29:13–18. doi: 10.1080/09546634.2017.1329511. [DOI] [PubMed] [Google Scholar]
- 177.Zhao J, Chen X, Herjan T, Li X. The role of interleukin-17 in tumor development and progression. J Exp Med. 2020;217:e20190297. doi: 10.1084/jem.20190297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Chang SH, Chung Y, Dong C. Vitamin D suppresses Th17 cytokine production by inducing C/EBP homologous protein (CHOP) expression. J Biol Chem. 2010;285:38751–38755. doi: 10.1074/jbc.C110.185777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Wong CK, Li EK, Ho CY, Lam CW. Elevation of plasma interleukin-18 concentration is correlated with disease activity in systemic lupus erythematosus. Rheumatology (Oxford) 2000;39:1078–1081. doi: 10.1093/rheumatology/39.10.1078. [DOI] [PubMed] [Google Scholar]
- 180.Mazodier K, Marin V, Novick D, Farnarier C, Robitail S, Schleinitz N, et al. Severe imbalance of IL-18/IL-18BP in patients with secondary hemophagocytic syndrome. Blood. 2005;106:3483–3489. doi: 10.1182/blood-2005-05-1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Pizarro TT, Michie MH, Bentz M, Woraratanadharm J, Smith MF, Foley E, et al. IL-18, a novel immunoregulatory cytokine, is up-regulated in Crohn’s disease: expression and localization in intestinal mucosal cells. J Immunol. 1999;162:6829–6835. [PubMed] [Google Scholar]
- 182.Monteleone G, Trapasso F, Parrello T, Biancone L, Stella A, Iuliano R, et al. Bioactive IL-18 expression is up-regulated in Crohn’s disease. J Immunol. 1999;163:143–147. [PubMed] [Google Scholar]
- 183.Mallat Z, Heymes C, Corbaz A, Logeart D, Alouani S, Cohen-Solal A, et al. Evidence for altered interleukin 18 (IL)-18 pathway in human heart failure. FASEB J. 2004;18:1752–1754. doi: 10.1096/fj.04-2426fje. [DOI] [PubMed] [Google Scholar]
- 184.Oka T, Sugaya M, Takahashi N, Nakajima R, Otobe S, Kabasawa M, et al. Increased interleukin-19 expression in cutaneous T-cell lymphoma and atopic dermatitis. Acta Derm Venereol. 2017;97:1172–1177. doi: 10.2340/00015555-2723. [DOI] [PubMed] [Google Scholar]
- 185.Wong CK, Wong PTY, Tam LS, Li EK, Chen DP, Lam CWK. Elevated production of B cell chemokine CXCL13 is correlated with systemic lupus erythematosus disease activity. J Clin Immunol. 2010;30:45–52. doi: 10.1007/s10875-009-9325-5. [DOI] [PubMed] [Google Scholar]
- 186.Festen EM, Goyette P, Scott R, Annese V, Zhernakova A, Lian J, et al. Genetic variants in the region harbouring IL2/IL21 associated with ulcerative colitis. Gut. 2009;58:799–804. doi: 10.1136/gut.2008.166918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Jin H, Oyoshi MK, Le Y, Bianchi T, Koduru S, Mathias CB, et al. IL-21R is essential for epicutaneous sensitization and allergic skin inflammation in humans and mice. J Clin Invest. 2009;119:47–60. doi: 10.1172/JCI32310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Petukhova L, Duvic M, Hordinsky M, Norris D, Price V, Shimomura Y, et al. Genome-wide association study in alopecia areata implicates both innate and adaptive immunity. Nature. 2010;466:113–117. doi: 10.1038/nature09114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Wang G, Tschoi M, Spolski R, Lou Y, Ozaki K, Feng C, et al. In vivo antitumor activity of interleukin 21 mediated by natural killer cells. Cancer Res. 2003;63:9016–9022. [PubMed] [Google Scholar]
- 190.Costanzo A, Chimenti MS, Botti E, Caruso R, Sarra M, Monteleone G. IL-21 in the pathogenesis and treatment of skin diseases. J Dermatol Sci. 2010;60:61–66. doi: 10.1016/j.jdermsci.2010.08.016. [DOI] [PubMed] [Google Scholar]
- 191.Stolfi C, Rizzo A, Franzè E, Rotondi A, Fantini MC, Sarra M, et al. Involvement of interleukin-21 in the regulation of colitis-associated colon cancer. J Exp Med. 2011;208:2279–2290. doi: 10.1084/jem.20111106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Niu X, He D, Zhang X, Yue T, Li N, Zhang JZ, et al. IL-21 regulates Th17 cells in rheumatoid arthritis. Hum Immunol. 2010;71:334–341. doi: 10.1016/j.humimm.2010.01.010. [DOI] [PubMed] [Google Scholar]
- 193.Gittler JK, Shemer A, Suárez-Fariñas M, Fuentes-Duculan J, Gulewicz KJ, Wang CQF, et al. Progressive activation of T(H)2/T(H)22 cytokines and selective epidermal proteins characterizes acute and chronic atopic dermatitis. J Allergy Clin Immunol. 2012;130:1344–1354. doi: 10.1016/j.jaci.2012.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Ricciardi L, Minciullo PL, Saitta S, Trombetta D, Saija A, Gangemi S. Increased serum levels of IL-22 in patients with nickel contact dermatitis. Contact Dermat. 2009;60:57–58. doi: 10.1111/j.1600-0536.2008.01454.x. [DOI] [PubMed] [Google Scholar]
- 195.Larsen JM, Bonefeld CM, Poulsen SS, Geisler C, Skov L. IL-23 and T(H)17-mediated inflammation in human allergic contact dermatitis. J Allergy Clin Immunol. 2009;123:486–492. doi: 10.1016/j.jaci.2008.09.036. [DOI] [PubMed] [Google Scholar]
- 196.Mathian A, Parizot C, Dorgham K, Trad S, Arnaud L, Larsen M, et al. Activated and resting regulatory T cell exhaustion concurs with high levels of interleukin-22 expression in systemic sclerosis lesions. Ann Rheum Dis. 2012;71:1227–1234. doi: 10.1136/annrheumdis-2011-200709. [DOI] [PubMed] [Google Scholar]
- 197.Miyagaki T, Sugaya M, Suga H, Kamata M, Ohmatsu H, Fujita H, et al. IL-22, but not IL-17, dominant environment in cutaneous T-cell lymphoma. Clin Cancer Res. 2011;17:7529–7538. doi: 10.1158/1078-0432.CCR-11-1192. [DOI] [PubMed] [Google Scholar]
- 198.Zhang S, Fujita H, Mitsui H, Yanofsky VR, Fuentes-Duculan J, Pettersen JS, et al. Increased Tc22 and Treg/CD8 ratio contribute to aggressive growth of transplant associated squamous cell carcinoma. PLoS ONE. 2013;8:e62154. doi: 10.1371/journal.pone.0062154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Perriard G, Mathias A, Enz L, Canales M, Schluep M, Gentner M, et al. Interleukin-22 is increased in multiple sclerosis patients and targets astrocytes. J Neuroinflamm. 2015;12:1–18. doi: 10.1186/s12974-015-0335-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Ikeuchi H, Kuroiwa T, Hiramatsu N, Kaneko Y, Hiromura K, Ueki K, et al. Expression of interleukin-22 in rheumatoid arthritis: potential role as a proinflammatory cytokine. Arthritis Rheum. 2005;52:1037–1046. doi: 10.1002/art.20965. [DOI] [PubMed] [Google Scholar]
- 201.Geremia A, Arancibia-Cárcamo CV, Fleming MPP, Rust N, Singh B, Mortensen NJ, et al. IL-23-responsive innate lymphoid cells are increased in inflammatory bowel disease. J Exp Med. 2011;208:1127–1133. doi: 10.1084/jem.20101712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Brentano F, Ospelt C, Stanczyk J, Gay RE, Gay S, Kyburz D. Abundant expression of the interleukin (IL)23 subunit p19, but low levels of bioactive IL23 in the rheumatoid synovium: differential expression and Toll-like receptor-(TLR) dependent regulation of the IL23 subunits, p19 and p40, in rheumatoid arthritis. Ann Rheum Dis. 2009;68:143–150. doi: 10.1136/ard.2007.082081. [DOI] [PubMed] [Google Scholar]
- 203.Gottlieb A, Menter A, Mendelsohn A, Shen Y-K, Li S, Guzzo C, et al. Ustekinumab, a human interleukin 12/23 monoclonal antibody, for psoriatic arthritis: randomised, double-blind, placebo-controlled, crossover trial. Lancet. 2009;373:633–640. doi: 10.1016/S0140-6736(09)60140-9. [DOI] [PubMed] [Google Scholar]
- 204.Van Belle AB, Cochez PM, de Heusch M, Pointner L, Opsomer R, Raynaud P, et al. IL-24 contributes to skin inflammation in para-phenylenediamine-induced contact hypersensitivity. Sci Rep. 2019;9:1–11. doi: 10.1038/s41598-018-38156-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Mitamura Y, Nunomura S, Nanri Y, Ogawa M, Yoshihara T, Masuoka M, et al. The IL-13/periostin/IL-24 pathway causes epidermal barrier dysfunction in allergic skin inflammation. Allergy. 2018;73:1881–1891. doi: 10.1111/all.13437. [DOI] [PubMed] [Google Scholar]
- 206.Andoh A, Shioya M, Nishida A, Bamba S, Tsujikawa T, Kim-Mitsuyama S, et al. Expression of IL-24, an activator of the JAK1/STAT3/SOCS3 cascade, is enhanced in inflammatory bowel disease. J Immunol. 2009;183:687–695. doi: 10.4049/jimmunol.0804169. [DOI] [PubMed] [Google Scholar]
- 207.Beale J, Jayaraman A, Jackson DJ, Macintyre JDR, Edwards MR, Walton RP, et al. Rhinovirus-induced IL-25 in asthma exacerbation drives type 2 immunity and allergic pulmonary inflammation. Sci Transl Med. 2014;6:256ra134. doi: 10.1126/scitranslmed.3009124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Tang W, Smith SG, Salter B, Oliveria JP, Mitchell P, Nusca GM, et al. Allergen-induced increases in interleukin-25 and interleukin-25 receptor expression in mature eosinophils from atopic asthmatics. Int Arch Allergy Immunol. 2016;170:234–242. doi: 10.1159/000449248. [DOI] [PubMed] [Google Scholar]
- 209.Scala E, Di Caprio R, Cacciapuoti S, Caiazzo G, Fusco A, Tortorella E, et al. A new T helper 17 cytokine in hidradenitis suppurativa: antimicrobial and proinflammatory role of interleukin-26. Br J Dermatol. 2019;181:1038–1045. doi: 10.1111/bjd.17854. [DOI] [PubMed] [Google Scholar]
- 210.Corvaisier M, Delneste Y, Jeanvoine H, Preisser L, Blanchard S, Garo E, et al. IL-26 is overexpressed in rheumatoid arthritis and induces proinflammatory cytokine production and Th17 cell generation. PLoS Biol. 2012;10:e1001395. doi: 10.1371/journal.pbio.1001395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Fujii M, Nishida A, Imaeda H, Ohno M, Nishino K, Sakai S, et al. Expression of Interleukin-26 is upregulated in inflammatory bowel disease. World J Gastroenterol. 2017;23:5519–5529. doi: 10.3748/wjg.v23.i30.5519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Dambacher J, Beigel F, Zitzmann K, De Toni EN, Göke B, Diepolder HM, et al. The role of the novel Th17 cytokine IL-26 in intestinal inflammation. Gut. 2009;58:1207–1217. doi: 10.1136/gut.2007.130112. [DOI] [PubMed] [Google Scholar]
- 213.Konradsen JR, Nordlund B, Levänen B, Hedlin G, Linden A. The cytokine interleukin-26 as a biomarker in pediatric asthma. Respir Res. 2016;17:1–4. doi: 10.1186/s12931-016-0351-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Kaabachi W, Bouali E, Berraïes A, Dhifallh IB, Hamdi B, Hamzaoui K, et al. Interleukin-26 is overexpressed in Behçet’s disease and enhances Th17-related -cytokines. Immunol Lett. 2017;190:177–184. doi: 10.1016/j.imlet.2017.08.008. [DOI] [PubMed] [Google Scholar]
- 215.Savchenko L, Mykytiuk M, Cinato M, Tronchere H, Kunduzova O, Kaidashev I. IL-26 in the induced sputum is associated with the level of systemic inflammation, lung functions and body weight in COPD patients. Int J Chron Obstruct Pulmon Dis. 2018;13:2569–2575. doi: 10.2147/COPD.S164833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Yoshizaki A, Yanaba K, Iwata Y, Komura K, Ogawa A, Muroi E, et al. Elevated serum interleukin-27 levels in patients with systemic sclerosis: association with T cell, B cell and fibroblast activation. Ann Rheum Dis. 2011;70:194–200. doi: 10.1136/ard.2009.121053. [DOI] [PubMed] [Google Scholar]
- 217.Cao J, Zhang L, Li D, Xu F, Huang S, Xiang Y, et al. IL-27 is elevated in patients with COPD and patients with pulmonary TB and induces human bronchial epithelial cells to produce CXCL10. Chest. 2012;141:121–130. doi: 10.1378/chest.10-3297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Lu D, Zhou X, Yao L, Liu C, Jin F, Wu Y. Clinical implications of the interleukin 27 serum level in breast cancer. J Investig Med. 2014;62:627–631. doi: 10.2310/JIM.0000000000000046. [DOI] [PubMed] [Google Scholar]
- 219.Xia L, Shen H, Zhao L, Lu J. Elevated levels of interleukin-27 in patients with Sjögren’s syndrome. Scand J Rheumatol. 2012;41:73–74. doi: 10.3109/03009742.2011.620574. [DOI] [PubMed] [Google Scholar]
- 220.Tanida S, Yoshitomi H, Ishikawa M, Kasahara T, Murata K, Shibuya H, et al. IL-27-producing CD14(+) cells infiltrate inflamed joints of rheumatoid arthritis and regulate inflammation and chemotactic migration. Cytokine. 2011;55:237–244. doi: 10.1016/j.cyto.2011.04.020. [DOI] [PubMed] [Google Scholar]
- 221.Luo Q, Liu Y, Liu S, Yin Y, Xu B, Cao J. Interleukin 28 is a potential therapeutic target for sepsis. Clin Immunol. 2019;205:29–34. doi: 10.1016/j.clim.2019.05.012. [DOI] [PubMed] [Google Scholar]
- 222.Lin S-C, Kuo C-C, Tsao J-T, Lin L-J. Profiling the expression of interleukin (IL)-28 and IL-28 receptor α in systemic lupus erythematosus patients. Eur J Clin Invest. 2012;42:61–69. doi: 10.1111/j.1365-2362.2011.02557.x. [DOI] [PubMed] [Google Scholar]
- 223.Jiménez-Sousa MA, Fernández-Rodríguez A, Guzmán-Fulgencio M, García-Álvarez M, Resino S. Meta-analysis: implications of interleukin-28B polymorphisms in spontaneous and treatment-related clearance for patients with hepatitis C. BMC Med. 2013;11:1–18. doi: 10.1186/1741-7015-11-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Arpaci D, Karakas Celik S, Can M, Cakmak Genc G, Kuzu F, Unal M, et al. Increased serum levels of IL-28 and IL-29 and the protective effect of IL28B rs8099917 polymorphism in patients with Hashimoto’s thyroiditis. Immunol Invest. 2016;45:668–678. doi: 10.1080/08820139.2016.1208215. [DOI] [PubMed] [Google Scholar]
- 225.Wu Q, Yang Q, Lourenco E, Sun H, Zhang Y. Interferon-lambda1 induces peripheral blood mononuclear cell-derived chemokines secretion in patients with systemic lupus erythematosus: its correlation with disease activity. Arthritis Res Ther. 2011;13:R88. doi: 10.1186/ar3363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Fedenko ES, Elisyutina OG, Filimonova TM, Boldyreva MN, Burmenskaya OV, Rebrova OY, et al. Cytokine gene expression in the skin and peripheral blood of atopic dermatitis patients and healthy individuals. Self Nonself. 2011;2:120–124. doi: 10.4161/self.2.2.16939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Dantas AT, Gonçalves SMC, Pereira MC, de Almeida AR, Marques CDL, Rego MJBM, et al. Interferons and systemic sclerosis: correlation between interferon gamma and interferon-lambda 1 (IL-29) Autoimmunity. 2015;48:429–433. doi: 10.3109/08916934.2015.1054028. [DOI] [PubMed] [Google Scholar]
- 228.Wang F, Xu L, Feng X, Guo D, Tan W, Zhang M. Interleukin-29 modulates proinflammatory cytokine production in synovial inflammation of rheumatoid arthritis. Arthritis Res Ther. 2012;14:R228. doi: 10.1186/ar4067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229.Xu L, Peng Q, Xuan W, Feng X, Kong X, Zhang M, et al. Interleukin-29 enhances synovial inflammation and cartilage degradation in osteoarthritis. Mediat Inflamm. 2016;2016:9631510. doi: 10.1155/2016/9631510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Haasnoot A-MJW, Kuiper JJW, Hiddingh S, Schellekens PAWJF, de Jager W, Imhof SM, et al. Ocular fluid analysis in children reveals interleukin-29/interferon-λ1 as a biomarker for juvenile idiopathic arthritis-associated uveitis. Arthritis Rheumatol. 2016;68:1769–1779. doi: 10.1002/art.39621. [DOI] [PubMed] [Google Scholar]
- 231.Apostolou E, Kapsogeorgou EK, Konsta OD, Giotakis I, Saridaki MI, Andreakos E, et al. Expression of type III interferons (IFNλs) and their receptor in Sjögren’s syndrome. Clin Exp Immunol. 2016;186:304–312. doi: 10.1111/cei.12865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Airoldi I, Cocco C, Sorrentino C, Angelucci D, Di Meo S, Manzoli L, et al. Interleukin-30 promotes breast cancer growth and progression. Cancer Res. 2016;76:6218–6229. doi: 10.1158/0008-5472.CAN-16-0189. [DOI] [PubMed] [Google Scholar]
- 233.Di Meo S, Airoldi I, Sorrentino C, Zorzoli A, Esposito S, Di Carlo E. Interleukin-30 expression in prostate cancer and its draining lymph nodes correlates with advanced grade and stage. Clin Cancer Res. 2014;20:585–594. doi: 10.1158/1078-0432.CCR-13-2240. [DOI] [PubMed] [Google Scholar]
- 234.Bağci IS, Ruzicka T. IL-31: a new key player in dermatology and beyond. J Allergy Clin Immunol. 2018;141:858–866. doi: 10.1016/j.jaci.2017.10.045. [DOI] [PubMed] [Google Scholar]
- 235.Yao Q, Wang B, Jia X, Li Q, Yao W, Zhang J-A. Increased human interleukin-32 expression is related to disease activity of Graves’ disease. Front Endocrinol (Lausanne) 2019;10:1–7. doi: 10.3389/fendo.2019.00613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Di Benedetto P, Guggino G, Manzi G, Ruscitti P, Berardicurti O, Panzera N, et al. Interleukin-32 in systemic sclerosis, a potential new biomarker for pulmonary arterial hypertension. Arthritis Res Ther. 2020;22:1–9. doi: 10.1186/s13075-020-02218-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Meyer N, Zimmermann M, Bürgler S, Bassin C, Woehrl S, Moritz K, et al. IL-32 is expressed by human primary keratinocytes and modulates keratinocyte apoptosis in atopic dermatitis. J Allergy Clin Immunol. 2010;125:858–865.e10. doi: 10.1016/j.jaci.2010.01.016. [DOI] [PubMed] [Google Scholar]
- 238.Joosten LAB, Netea MG, Kim S-H, Yoon D-Y, Oppers-Walgreen B, Radstake TRD, et al. IL-32, a proinflammatory cytokine in rheumatoid arthritis. Proc Natl Acad Sci USA. 2006;103:3298–3303. doi: 10.1073/pnas.0511233103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 239.Calabrese F, Baraldo S, Bazzan E, Lunardi F, Rea F, Maestrelli P, et al. IL-32, a novel proinflammatory cytokine in chronic obstructive pulmonary disease. Am J Respir Crit Care Med. 2008;178:894–901. doi: 10.1164/rccm.200804-646OC. [DOI] [PubMed] [Google Scholar]
- 240.Savinko T, Matikainen S, Saarialho-Kere U, Lehto M, Wang G, Lehtimäki S, et al. IL-33 and ST2 in atopic dermatitis: expression profiles and modulation by triggering factors. J Invest Dermatol. 2012;132:1392–1400. doi: 10.1038/jid.2011.446. [DOI] [PubMed] [Google Scholar]
- 241.Mattii M, Ayala F, Balato N, Filotico R, Lembo S, Schiattarella M, et al. The balance between pro- and anti-inflammatory cytokines is crucial in human allergic contact dermatitis pathogenesis: the role of IL-1 family members. Exp Dermatol. 2013;22:813–819. doi: 10.1111/exd.12272. [DOI] [PubMed] [Google Scholar]
- 242.Kay AB, Clark P, Maurer M, Ying S. Elevations in T-helper-2-initiating cytokines (interleukin-33, interleukin-25 and thymic stromal lymphopoietin) in lesional skin from chronic spontaneous ('idiopathic’) urticaria. Br J Dermatol. 2015;172:1294–1302. doi: 10.1111/bjd.13621. [DOI] [PubMed] [Google Scholar]
- 243.Li P, Ma H, Han D, Mou K. Interleukin-33 affects cytokine production by keratinocytes in vitiligo. Clin Exp Dermatol. 2015;40:163–170. doi: 10.1111/ced.12464. [DOI] [PubMed] [Google Scholar]
- 244.Sakashita M, Yoshimoto T, Hirota T, Harada M, Okubo K, Osawa Y, et al. Association of serum interleukin-33 level and the interleukin-33 genetic variant with Japanese cedar pollinosis. Clin Exp Allergy. 2008;38:1875–1881. doi: 10.1111/j.1365-2222.2008.03114.x. [DOI] [PubMed] [Google Scholar]
- 245.Préfontaine D, Lajoie-Kadoch S, Foley S, Audusseau S, Olivenstein R, Halayko AJ, et al. Increased expression of IL-33 in severe asthma: evidence of expression by airway smooth muscle cells. J Immunol. 2009;183:5094–5103. doi: 10.4049/jimmunol.0802387. [DOI] [PubMed] [Google Scholar]
- 246.Matsuyama Y, Okazaki H, Tamemoto H, Kimura H, Kamata Y, Nagatani K, et al. Increased levels of interleukin 33 in sera and synovial fluid from patients with active rheumatoid arthritis. J Rheumatol. 2010;37:18–25. doi: 10.3899/jrheum.090492. [DOI] [PubMed] [Google Scholar]
- 247.Kobori A, Yagi Y, Imaeda H, Ban H, Bamba S, Tsujikawa T, et al. Interleukin-33 expression is specifically enhanced in inflamed mucosa of ulcerative colitis. J Gastroenterol. 2010;45:999–1007. doi: 10.1007/s00535-010-0245-1. [DOI] [PubMed] [Google Scholar]
- 248.Moon S-J, Hong Y-S, Ju JH, Kwok S-K, Park S-H, Min J-K. Increased levels of interleukin 34 in serum and synovial fluid are associated with rheumatoid factor and anticyclic citrullinated peptide antibody titers in patients with rheumatoid arthritis. J Rheumatol. 2013;40:1842–1849. doi: 10.3899/jrheum.130356. [DOI] [PubMed] [Google Scholar]
- 249.Xie HH, Shen H, Zhang L, Cui MY, Xia LP, Lu J. Elevated serum interleukin-34 level in patients with systemic lupus erythematosus is associated with disease activity. Sci Rep. 2018;8:1–6. doi: 10.1038/s41598-018-21859-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250.Ciccia F, Alessandro R, Rodolico V, Guggino G, Raimondo S, Guarnotta C, et al. IL-34 is overexpressed in the inflamed salivary glands of patients with Sjögren’s syndrome and is associated with the local expansion of pro-inflammatory CD14(bright)CD16+ monocytes. Rheumatology (Oxford) 2013;52:1009–1017. doi: 10.1093/rheumatology/kes435. [DOI] [PubMed] [Google Scholar]
- 251.Franzè E, Monteleone I, Cupi ML, Mancia P, Caprioli F, Marafini I, et al. Interleukin-34 sustains inflammatory pathways in the gut. Clin Sci (Lond) 2015;129:271–280. doi: 10.1042/CS20150132. [DOI] [PubMed] [Google Scholar]
- 252.Shoji H, Yoshio S, Mano Y, Kumagai E, Sugiyama M, Korenaga M, et al. Interleukin-34 as a fibroblast-derived marker of liver fibrosis in patients with non-alcoholic fatty liver disease. Sci Rep. 2016;6:28814. doi: 10.1038/srep28814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Baghdadi M, Endo H, Takano A, Ishikawa K, Kameda Y, Wada H, et al. High co-expression of IL-34 and M-CSF correlates with tumor progression and poor survival in lung cancers. Sci Rep. 2018;8:1–10. doi: 10.1038/s41598-017-18796-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254.Fogel O, Rivière E, Seror R, Nocturne G, Boudaoud S, Ly B, et al. Role of the IL-12/IL-35 balance in patients with Sjögren syndrome. J Allergy Clin Immunol. 2018;142:258–268.e5. doi: 10.1016/j.jaci.2017.07.041. [DOI] [PubMed] [Google Scholar]
- 255.Jafarzadeh A, Jamali M, Mahdavi R, Ebrahimi HA, Hajghani H, Khosravimashizi A, et al. Circulating levels of interleukin-35 in patients with multiple sclerosis: evaluation of the influences of FOXP3 gene polymorphism and treatment program. J Mol Neurosci. 2015;55:891–897. doi: 10.1007/s12031-014-0443-z. [DOI] [PubMed] [Google Scholar]
- 256.Tomcik M, Zerr P, Palumbo-Zerr K, Storkanova H, Hulejova H, Spiritovic M, et al. Interleukin-35 is upregulated in systemic sclerosis and its serum levels are associated with early disease. Rheumatology (Oxford) 2015;54:2273–2282. doi: 10.1093/rheumatology/kev260. [DOI] [PubMed] [Google Scholar]
- 257.Chu M, Wong CK, Cai Z, Dong J, Jiao D, Kam NW, et al. Elevated expression and pro-inflammatory activity of il-36 in patients with systemic lupus erythematosus. Molecules. 2015;20:19588–19604. doi: 10.3390/molecules201019588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258.Frey S, Derer A, Messbacher M-E, Baeten DLP, Bugatti S, Montecucco C, et al. The novel cytokine interleukin-36α is expressed in psoriatic and rheumatoid arthritis synovium. Ann Rheum Dis. 2013;72:1569–1574. doi: 10.1136/annrheumdis-2012-202264. [DOI] [PubMed] [Google Scholar]
- 259.Russell SE, Horan RM, Stefanska AM, Carey A, Leon G, Aguilera M, et al. IL-36α expression is elevated in ulcerative colitis and promotes colonic inflammation. Mucosal Immunol. 2016;9:1193–1204. doi: 10.1038/mi.2015.134. [DOI] [PubMed] [Google Scholar]
- 260.Ciccia F, Accardo-Palumbo A, Alessandro R, Alessandri C, Priori R, Guggino G, et al. Interleukin-36α axis is modulated in patients with primary Sjögren’s syndrome. Clin Exp Immunol. 2015;181:230–238. doi: 10.1111/cei.12644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 261.Suárez-Fariñas M, Ungar B, da Rosa JC, Ewald DA, Rozenblit M, Gonzalez J, et al. RNA sequencing atopic dermatitis transcriptome profiling provides insights into novel disease mechanisms with potential therapeutic implications. J Allergy Clin Immunol. 2015;135:1218–1227. doi: 10.1016/j.jaci.2015.03.003. [DOI] [PubMed] [Google Scholar]
- 262.Ye L, Ji L, Wen Z, Zhou Y, Hu D, Li Y, et al. IL-37 inhibits the production of inflammatory cytokines in peripheral blood mononuclear cells of patients with systemic lupus erythematosus: its correlation with disease activity. J Transl Med. 2014;12:1–8. doi: 10.1186/1479-5876-12-69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263.Chen B, Huang K, Ye L, Li Y, Zhang J, Zhang J, et al. Interleukin-37 is increased in ankylosing spondylitis patients and associated with disease activity. J Transl Med. 2015;13:1–9. doi: 10.1186/s12967-015-0394-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 264.Li Y, Wang Z, Yu T, Chen B, Zhang J, Huang K, et al. Increased expression of IL-37 in patients with Graves’ disease and its contribution to suppression of proinflammatory cytokines production in peripheral blood mononuclear cells. PLoS ONE. 2014;9:e107183. doi: 10.1371/journal.pone.0107183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 265.Li C, Zhao P, Sun X, Che Y, Jiang Y. Elevated levels of cerebrospinal fluid and plasma interleukin-37 in patients with Guillain-Barré syndrome. Mediat Inflamm. 2013;2013:639712. doi: 10.1155/2013/639712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 266.Zhao P-W, Jiang W-G, Wang L, Jiang Z-Y, Shan Y-X, Jiang Y-F. Plasma levels of IL-37 and correlation with TNF-α, IL-17A, and disease activity during DMARD treatment of rheumatoid arthritis. PLoS ONE. 2014;9:e95346. doi: 10.1371/journal.pone.0095346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 267.Boutet M-A, Bart G, Penhoat M, Amiaud J, Brulin B, Charrier C, et al. Distinct expression of interleukin (IL)-36α, β and γ, their antagonist IL-36Ra and IL-38 in psoriasis, rheumatoid arthritis and Crohn’s disease. Clin Exp Immunol. 2016;184:159–173. doi: 10.1111/cei.12761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 268.Tominaga M, Okamoto M, Kawayama T, Matsuoka M, Kaieda S, Sakazaki Y, et al. Overexpression of IL-38 protein in anticancer drug-induced lung injury and acute exacerbation of idiopathic pulmonary fibrosis. Respir Investig. 2017;55:293–299. doi: 10.1016/j.resinv.2017.06.001. [DOI] [PubMed] [Google Scholar]
- 269.Yang M-G, Tian S, Zhang Q, Han J, Liu C, Zhou Y, et al. Elevated serum interleukin-39 levels in patients with neuromyelitis optica spectrum disorders correlated with disease severity. Mult Scler Relat Disord. 2020;46:102430. doi: 10.1016/j.msard.2020.102430. [DOI] [PubMed] [Google Scholar]
- 270.Luo Y, Liu F, Liu H, Chen H, Cheng W, Dong S, et al. Elevated serum IL-39 in patients with ST-segment elevation myocardial infarction was related with left ventricular systolic dysfunction. Biomark Med. 2017;11:419–426. doi: 10.2217/bmm-2016-0361. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
There are no data associated with this review article apart from what is presented and referenced here.