

ABSTRACT
African swine fever (ASF), an acute, severe, highly contagious disease caused by African swine fever virus (ASFV) infection in domestic pigs and boars, has a mortality rate of up to 100%. Because effective vaccines and treatments for ASF are lacking, effective control of the spread of ASF remains a great challenge for the pig industry. Host epigenetic regulation is essential for the viral gene transcription. Bromodomain and extraterminal (BET) family proteins, including BRD2, BRD3, BRD4, and BRDT, are epigenetic “readers” critical for gene transcription regulation. Among these proteins, BRD4 recognizes acetylated histones via its two bromodomains (BD1 and BD2) and recruits transcription factors, thereby playing a pivotal role in transcriptional regulation and chromatin remodeling during viral infection. However, how BET/BRD4 regulates ASFV replication and gene transcription is unknown. Here, we randomly selected 12 representative BET family inhibitors and compared their effects on ASFV infection in pig primary alveolar macrophages (PAMs). These were found to inhibit viral infection by interfering viral replication. The four most effective inhibitors (ARV-825, ZL0580, I-BET-762, and PLX51107) were selected for further antiviral activity analysis. These BET/BRD4 inhibitors dose dependently decreased the ASFV titer, viral RNA transcription, and protein production in PAMs. Collectively, we report novel function of BET/BRD4 inhibitors in inducing suppression of ASFV infection, providing insights into the role of BET/BRD4 in the epigenetic regulation of ASFV and potential new strategies for ASF prevention and control.
IMPORTANCE Due to the continuing spread of the ASFV in the world and the lack of commercial vaccines, the development of improved control strategies, including antiviral drugs, is urgently needed. BRD4 is an important epigenetic factor and has been commonly used for drug development for tumor treatment. Furthermore, the latest research showed that BET/BRD4 inhibition could suppress replication of virus. In this study, we first showed the inhibitory effect of agents targeting BET/BRD4 on ASFV infection with no significant host cytotoxicity. Then, we found four BET/BRD4 inhibitors that can inhibit ASFV replication, RNA transcription, and protein synthesis. Our findings support the hypothesis that BET/BRD4 can be considered as attractive host targets in antiviral drug discovery against ASFV.
KEYWORDS: African swine fever virus, BET, BRD4, inhibitors, antiviral effect
INTRODUCTION
African swine fever (ASF), a highly contagious viral disease in swine infected with African swine fever virus (ASFV), exhibits a high mortality rate approaching 100% and has severe economic consequences for affected countries (1). ASF is clinically characterized by high fever, spotty skin, cyanosis, extensive bleeding of the internal organs, and disturbance of the respiratory and nervous systems (2). ASF was first introduced to Liaoning Province of China in August 2018, when genotype II ASFV resulted in numerous outbreaks within domestic pigs (http://www.oie.int/). These outbreaks resulted in economic losses of several billion dollars to China’s pig breeding industry and national economy, seriously affecting the lives of Chinese residents, national economic development and the pig industry (3).
The ASFV genome is large and complex, and the mechanism by which replication is regulated is unclear thus far. Although ASFV was discovered nearly a hundred years ago, no commercial vaccines or cost-effective antiviral drugs are available to effectively prevent ASF worldwide. ASFV, a tick-borne, double-stranded DNA virus and the only member of the Asfarviridae family, genus Asfivirus, mainly infects myeloid lineage cells, especially monocytes/macrophages and dendritic cells (4). The replication of ASFV primarily occurs in the cytoplasm, but a transient nuclear progress occurs at the early stage (5–10). However, the nuclear replication mechanism is not clear. ASFV encodes 150 to 167 proteins (11), including at least 54 structural proteins and more than 100 nonstructural proteins, which are involved in replication of the genome and assembly of the virion, respectively, and also regulate host cell function and immune evasion (12).
Histone lysine acetylation is a key mechanism in chromatin processes and the regulation of gene transcription (13). BET family proteins include BRD2, BRD3, BRD4, and BRDT, which have important biological functions, such as their ability to mediate transcriptional regulation and chromatin remodeling (14). BRD4 recruits positive transcriptional elongation factor (P-TEFb) complex, which plays an essential role in transcriptional regulation by RNA polymerase II (RNA Pol II) in eukaryotes (15). BRD4 is one of the most important proteins in the BET family, and contains two bromo-domains (BD1 and BD2). BRD4 is not only a chromatin reader protein but also an epigenetic regulatory factor and transcription cofactor closely related to gene transcription, the cell cycle, and apoptosis (16, 17). Abnormal BRD4 protein expression can lead to the disordered expression of various genes and thus affects the function of related genes. BRD4 also plays an important role in DNA replication, transcription, and repair (18). Among host molecules, BRD4 can be used by DNA viruses to regulate the transcription of viral genes during viral replication through critical protein-protein interactions. BRD4 and its inhibitors have been widely studied as potential antitumor therapies. The latest research showed that BRD4 inhibition activated the cGAS-STING pathway of the antiviral innate immune response by leading to DNA damage-dependent signaling and attenuated viral attachment of pseudorabies virus (suid herpesvirus 1) (19). In addition, a BRD4 inhibitor was found to suppress human immunodeficiency virus (HIV) by inhibiting Tat transactivation and transcription elongation and by inducing a repressive chromatin structure at the HIV promoter (20).
However, the potential effect of BET/BRD4 on ASFV replication and viral transcription has not been evaluated. ASFV may alter the epigenetic status of host chromatin to modulate cellular gene expression for its own benefit. Therefore, we focused on the biological effects of representative BET/BRD4 inhibitors on the replication and transcription of ASFV in vitro, and our results may open new avenues for the effective prevention and control of ASF.
RESULTS
Cytotoxicity of BET inhibitors in PAMs.
The inhibitors were used at nine different concentrations ranging from 0.5 to 240 μM to evaluate cytotoxicity by using a CCK-8 assay. The results indicated that at least seven of the inhibitors did not cause a significant increase in cell death, and the cell viability reached more than 60% when the concentrations of the inhibitors were up to 20 μM, but significant cytotoxic effects on the primary alveolar macrophages (PAMs) were observed at concentrations from 40 to 240 μM. Cell viability was still more than 50% when the inhibitors INCB054329 and CPI-203 were used at 80 μM. The inhibitors demonstrated potent cytotoxic effects on PAMs at 10 μM (ARV-825 and AZD5153), 20 μM [PLX51107, PFI-1, RVX-208, ZL0580, and (+)-JQ1], 40 μM (OTX051, MS436, and I-BET-762), and 80 μM (INCB054329 and CPI-203) (Fig. 1). The organic solvent dimethyl sulfoxide (DMSO) had no cytotoxic effect on PAMs (data not shown). Therefore, even though most of these BET inhibitors are commercially available as research tools, some of them show a certain degree of cytotoxicity against PAMs at high concentrations; at these concentrations, the primary cells are more sensitive to the inhibitors.
FIG 1.

Cytotoxicity of BET/BRD4 inhibitors against PAMs. PAMs were treated with BET/BRD4 inhibitors at a range of concentrations from 0.5 to 240 μM for 24 h, and cell viability was evaluated using a CCK-8 kit.
Effect of BET inhibitors on ASFV transcription in PAMs.
To determine whether the BET inhibitors could affect ASFV gene transcription by altering the functions of BET proteins, a time-of-addition assay was conducted to evaluate the effects of 12 BET/BRD4 inhibitors on specific step(s) of the ASFV life cycle. Cells were treated with the individual BET inhibitors at 5 μM, and the functional role of BET in BET inhibitor-induced ASFV gene transcription was evaluated using real-time PCR. The relative expression levels of CP204L (early), B646L (late), and GAPDH in cells treated with individual BET inhibitors were measured and compared to those in the control (DMSO; negative control [NC]) group. Pretreatment with the BET inhibitors potently suppressed ASFV gene transcription in the cells (Fig. 2A). A significant inhibitory effect on transcription of the CP204L gene, which is expressed early during the ASFV infection cycle, was observed when the inhibitors were applied simultaneously with ASFV infection, but the effect was less pronounced than that observed upon pretreatment (Fig. 2B). Moreover, neither CP204L nor B646L gene transcription was inhibited by BET inhibitor posttreatment (Fig. 2C). Interestingly, pretreatment with PLX51107 and ZL0580 almost completely inhibited ASFV gene transcription. Since accumulating evidence suggests that BET/BRD4 plays an important role in regulating viral transcriptional (21–23) and based on our results presented above, four representative inhibitors (PLX51107, I-BET762, ZL0580, and ARV-825) with the greatest inhibitory effects under both pretreatment and cotreatment conditions were selected for further experiments. Among these four inhibitors, the first two (PLX51107 and I-BET762) are broad-spectrum BET family inhibitors, while the latter two (ZL0580 and ARV-825) are BRD4-specific inhibitors. Collectively, these results suggest that BET/BRD4 inhibition results in decreased ASFV gene transcription in ASFV infection, and the expression of ASFV CP204L and B646L upon inhibitor treatment significantly differed from that in untreated cells (DMSO-treated group) in vitro. Thus, further experiments were performed.
FIG 2.

Effect of BET family inhibitors on ASFV gene transcription. The expression levels of CP204L and B646L mRNA from ASFV pretreatment (A), cotreatment (B), and posttreatment (C) with the inhibitors were detected by real-time PCR. Data were normalized to data from the DMSO-treated samples. PAMs in 12-well plates were pretreated, cotreated or posttreated with individual inhibitors or DMSO in relation to ASFV (MOI = 0.1) infection. The samples were collected at 24 hpi under pre- and cotreatment conditions with the inhibitors. For the posttreatment samples, the cells were first infected with ASFV for 4 h, followed by inhibitor treatment for 16 h, and then the samples were collected. The concentration of the BET/BRD4 inhibitors was 5 μM. Error bars show the SD of replicates qPCR experiments. All experiments were independently conducted at least three times. Statistical significance is denoted by asterisks (*, P < 0.05; **, P < 0.01; ***, P < 0.001; ****, P < 0.0001).
PLX51107, I-BET762, ZL0580, and ARV-825 inhibit viral infection in a time-dependent manner.
The structures of ARV-825, ZL0580, I-BET-762, and PLX51107 are shown in Fig. 3A. The 50% cell cytotoxicity (CC50) values, the concentrations of the four inhibitors at which they caused 50% cell death, were calculated in PAMs. The CC50 values of ARV-825, ZL0580, I-BET-762, and PLX51107 were determined to be 10.11 μM (95% confidence interval [CI] = 9.18 to 11.11), 25.3 μM (95% CI = 21.57 to 31.11), 35.86 μM (95% CI = 25.38 to 86.79), and 19.37 μM (95% CI = 15.91 to 24.68), respectively (Fig. 3B). In antiviral experiments, to mitigate their cytotoxic effects, ARV-825, ZL0580, PLX51107, and I-BET-762 were used at maximum concentrations of 1, 10, 5 and 10 μM, respectively, which were lower than the CC50 values. The duration over which the four inhibitors inhibited the replication of ASFV was further evaluated. The four inhibitors were added to PAM culture medium for 16 h prior to ASFV infection. Samples were collected at 4, 12, 24, and 48 h after infection. The copy number of the B646L gene was then detected by real-time PCR. The significant inhibitory effects of four inhibitors on late expressed gene B646L were observed started at 12 h after ASFV infection until 48 h postinfection (hpi) (Fig. 4). These results indicate that these four inhibitors significantly inhibited the replication of ASFV for a long time.
FIG 3.

Structures of inhibitors and CC50 values. (A) Structures of ARV-825, ZL0580, I-BET-762, and PLX51107. (B) CC50 values for ARV-825, ZL0580, I-BET-762, and PLX51107 calculated against PAMs.
FIG 4.
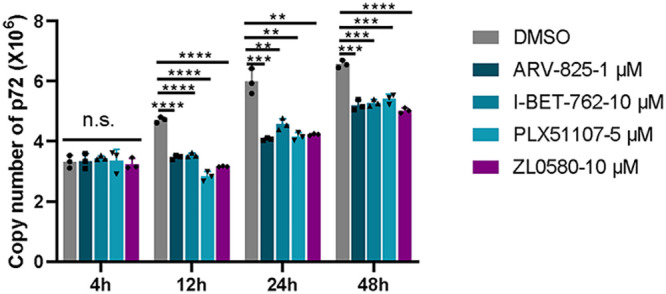
Time-dependent effect of four inhibitors on PAMs. The four inhibitors act throughout the whole ASFV infection cycle to decrease ASFV RNA levels. PAMs were treated with 1 μM ARV-825, 10 μM ZL0580, 10 μM I-BET-762, and 5 μM PLX51107 for 16 h prior to ASFV infection (MOI = 0.1). The ASFV B646L gene copy numbers at 4, 12, 24, and 48 h postinfection were determined and analyzed by qPCR. Data were normalized to data from DMSO-treated samples. Statistical significance is denoted by asterisks (*, P < 0.05; **, P < 0.01; ***, P < 0.001; ****, P < 0.0001).
Inhibitory effect of BET/BRD4 on ASFV infection of PAMs in a dose-dependent manner.
In an ASFV suppression model, PAMs were treated with four individual inhibitors, and their potential dose-dependent antiviral activity against ASFV was evaluated. We treated ASFV-infected PAMs with the individual inhibitors at increasing concentrations from 0.1 to 10 μM, depending on the inhibitor. As shown in Fig. 5A, the viral yields decreased significantly from 6 to 1.6 log HAD50/mL at a concentration of 1 μM (ARV-825), 5 μM (PLX51107), or 10 μM (ZL0580 and I-BET-762) (P < 0.05 or 0.001). At the gene transcription level, ZL0580 did not significantly suppress late ASFV B646L mRNA expression at concentrations lower than 2 μM. All four inhibitors clearly suppressed the early expression of ASFV CP204L mRNA (P < 0.05, 0.01, or 0.001) (Fig. 5B). Importantly, further analysis of protein expression levels revealed that viral p72 protein expression levels were clearly suppressed in ASFV-infected PAMs treated with the four inhibitors in a dose-dependent manner, especially upon treatment with 0.25 to 1 μM ARV-825, which fully inhibited expression of the p72 protein (Fig. 5C). These results indicated that the four BET/BRD4 inhibitors suppressed the ASFV titer, as well as mRNA and protein synthesis during replication. Based on these results, maximum concentrations of 1 μM (ARV-825), 10 μM (ZL0580 and I-BET-762), and 5 μM (PLX51107) were selected for further evaluation of the effects of the inhibitors against ASFV infection.
FIG 5.

Dose-dependent effects of ARV-825, I-BET-762, PLX51107, and ZL0580 on ASFV replication. (A) ASFV yield in PAMs decreased significantly in a dose-dependent manner with four inhibitor pretreatments. (B) ASFV CP204L and B646L mRNA levels were analyzed by RT qPCR. (C) The expression of p72 in the presence of four inhibitors at several concentrations was evaluated by WB analysis. PAMs in 12-well plates were treated with individual inhibitors or DMSO for 16 h prior to ASFV infection (MOI = 0.1). The samples were collected at 24 hpi. Data were normalized to data from DMSO-treated samples. Error bars show the SD of replicate qPCR experiments. All experiments were independently conducted at least three times. Statistical significance is denoted by asterisks (*, P < 0.05; **, P < 0.01; ***, P < 0.001).
ZL0580, PLX51107, I-BET762, and ARV-825 suppress ASFV protein synthesis.
To further confirm the antiviral effects of ZL0580, PLX51107, I-BET762, and ARV-825, early and late expression of the important viral structural proteins p30 and p72, respectively, was analyzed by Western blot (WB) analysis (Fig. 6A). Immunoblot analysis showed that in the presence of ZL0580, PLX51107, I-BET762, and ARV-825, the p30 and p72 protein levels in the PAMs were significantly reduced compared to those in DMSO-treated cells, especially after treatment with ARV-825, and the expression levels of both proteins were decreased by more than 50%. Similar results were observed when expression of the ASFV p30 protein was evaluated by immunofluorescence analysis (Fig. 6B). Clear fluorescent signals were detected, and the fluorescence density was higher in the DMSO-treated PAMs than in the inhibitor-treated PAMs. In contrast, the fluorescence intensity was significantly decreased in the four inhibitor treatment groups (Fig. 6B). The percentage of cells showing early expression of the p30 protein was lower among cells treated with the BET inhibitors, as shown by flow cytometry analysis. ARV-825 treatment (1 μM) led to the sharply loss of p30 expression in ASFV-infected PAMs, with this effect followed by the effects of PLX51107, I-BET-762, and ZL0580 treatment. Compared to the DMSO-treated group, which was used as a control, the inhibitors had an at least 40% inhibitory effect (Fig. 6C). These results indicated that BET/BRD4 inhibition affects early and late protein synthesis.
FIG 6.

Evaluation of the inhibitory activity of ARV-825, I-BET-762, PLX51107, and ZL0580 against ASFV protein synthesis. The expression of p30 and p72 in the presence of the four inhibitors ARV-825 (1 μM), I-BET-762 (10 μM), PLX51107 (5 μM), and ZL0580 (10 μM) was evaluated by WB analysis (A), confocal microscopy (B), and flow cytometry (C). PAMs in 6-well plates were treated with inhibitors or DMSO 16 h prior to ASFV infection (MOI = 1). The samples were collected at 24 hpi.
BET/BRD4 inhibitors suppress the ASFV RNA polymerase expression levels.
ASFV belongs to the nucleocytoplasmic large DNA virus family, the members of which utilize quite complex RNA polymerases. Studies have shown that, unlike the 14 subunits encoded by eukaryotic RNA Pol II, ASFV encodes nine subunits that are homologous to eukaryotic RNA Pol II subunits (24). BRD4 was proven to bind the positive transcription elongation factor (P-TEFb) to form a complex that is subsequently recruited to RNA Pol II of the host, which then regulates the transcription of host or viral genes (21). Therefore, we further analyzed the transcription levels of the nine subunits of ASFV RNA polymerase by real-time PCR. The results showed that ZL0580, PLX51107, I-BET-762, and ARV-825 significantly inhibited transcription of the ASFV RNA polymerase subunit genes compared to their transcription in the DMSO control group, and ARV-825 and ZL0580 treatment had a stronger inhibitory effect on gene transcription levels than treatment with the two other inhibitors (Fig. 7A). We then selected ARV-825 and ZL0580 (BRD4-specific inhibitors) and evaluated their suppressive effects on the pC315R and pH359L proteins, which are homologs of TFIIB and RPB3 of the eukaryotic RNA Pol II (25), and the p30 protein. WB analysis revealed that ZL0580 and ARV-825-mediated inhibition of BET/BRD4 suppressed ASFV pC315R, pH359L, and p30 protein expression (Fig. 7B).
FIG 7.

Effect of inhibitors on the putative subunits of ASFV RNA polymerase. The expression of ASFV RNA polymerase subunits was significantly decreased at the RNA level (A) and the protein level (B) with inhibitor treatment. PAMs in 12-well plates were treated with ZL0580 (10 μM), I-BET-762 (10 μM), PLX51107 (5 μM), ARV-825 (1 μM), or DMSO for 16 h prior to ASFV infection (MOI = 0.1), and the samples were collected at 24 hpi. The NP1450L, EP1242L, H359L, D205R, C147L, D339L, CP80R, C315R, and I243L genes of ASFV were analyzed by qRT-PCR. The pC315R and pH359L proteins of ASFV were analyzed by WB analysis. Error bars show the SD values of replicate qPCR experiments. All experiments were independently conducted at least three times. Statistical significance is denoted by asterisks (*, P < 0.05; **, P < 0.01; ***, P < 0.001; ****, P < 0.0001).
DISCUSSION
ASF, the most serious exotic pig disease, is listed as a class I animal disease in China. Since the first outbreak of ASF in Shenyang in August 2018 (26, 27), continuous infection has spread throughout the whole country, and ASF represents a serious threat for the global swine industry and the environment with grave economic consequences for stakeholders (28). The generation of vaccines can impede the global spread of ASF, in addition to the implementation of other measures, such as rapid diagnosis and control and eradication measures. However, commercialized vaccines for the prevention of ASFV infection remain lacking. In addition to vaccine development, the development of antiviral drugs is an important strategy to respond to ASF epidemics.
At present, the research and development of anti-ASFV drugs mainly focuses on two categories: (i) inhibitors that directly act on the proteins encoded by AFSV to affect its replication and (ii) inhibitors that act on host protein factors required for viral replication to indirectly exert an anti-ASFV effect (29). Antiviral agents against ASFV currently include interferon (30), antibiotics (31), nucleoside analogues (32), plant-derived products (33), and other compounds that have been reported to inhibit ASFV replication (29). However, the safety of action of these antiviral drugs has not been studied in depth. Therefore, the need to identify new antiviral drugs for controlling ASFV is urgent.
Similar to other viruses, the signs of host infection with ASFV depend on the interaction between viruses and the host. BET family members include BRD2, BRD3, BRD4, and BRDT, which are widely involved in regulating the expression of genes related to transcription, DNA repair, immunity, metabolism, and signal transduction; these proteins accomplish this by identifying acetylated histones or transcription factors via their two unique bromodomains and have become promising targets for tumor therapy and viral infection (14, 19). Small-molecule inhibitors of BET family proteins may provide a promising option for cancer treatment. To date, more than 10 BET inhibitors have entered clinical trials and have mainly been used for the treatment of human diseases (15, 34). However, the effects of currently available BET/BRD4 inhibitors on ASFV infection are unknown.
During viral infection, host epigenetic factors can be involved in epigenetic modifications that affect the transcription and expression of viral genes and host genes (35). Therefore, the elucidation of potential target genes of BET proteins may help reveal new functions of BET family members and provide new possibilities for clinical treatment and the combined application of BET inhibitors. The antiviral activity of BET inhibitors has been demonstrated against different viruses, including PRV (19), bovine papillomavirus (23), human papillomavirus (HPV) (36), HIV (20), respiratory syncytial virus (RSV) (37), and Epstein-Barr virus (38). Previous studies have reported that BET inhibitors suppress the infectivity of these related viruses by decreasing macrophage and neutrophil infiltration into the airway, suppressing key inflammatory cytokines, preventing the expression of viral immediate early proteins and/or effectively blocking BET/BRD4 phosphorylation-specific functions in transcription factor recruitment. Nevertheless, their antiviral effect on ASFV remains unknown.
Here, we evaluated for the first time the antiviral effect of 12 representative BET/BRD4 inhibitors against ASFV infection in vitro (Fig. 2). After screening for their cytotoxicity against PAMs by CCK-8 assay, four BET/BRD4 inhibitors were selected, and their roles in ASFV gene and protein expression were further studied. The cytotoxic effects of 12 BET/BRED4 inhibitors against PAMs were first evaluated by quantifying cell viability with a CCK-8 assay. Our results demonstrated that most of these BET inhibitors were less cytotoxic against PAMs at concentrations between 0.5 and 10 μM; therefore, we used doses of ≤10 μM (ARV-825, 1 μM; PLX51107, 5 μM; I-BET-762 and ZL0580, 10 μM) for further experiments. We determined the CC50 values for the four selected BET inhibitors to ensure their safety in PAMs (Fig. 3B). In general, primary cells are more sensitive to compound cytotoxic effect than cell lines. However, in a previous study, obvious cytotoxicity was observed when the cells were treated with BET/BRD4 inhibitors (JQ-1, OTX-015, and I-BET 151) at 30 μM in both PK15 and HEK293 cells, while concentrations of 0 to 10 μM were minimally toxic in both cell lines (19), consistent with our results obtained with PAMs. This suggests that these inhibitors are harmless at concentrations below 10 μM in both primary cells and cell lines.
We performed time-of-addition studies to investigate whether the BET/BRD4 inhibitors have a primary antiviral effect on ASFV CN/SC/2019, a viral strain that replicates efficiently in primary PAMs (Fig. 2). Early expression of the CP204L gene was significantly decreased when the inhibitors were applied prior to (pretreatment) or simultaneously with virus infection (P < 0.05), but the addition of inhibitors after ASFV infection (posttreatment) had no statistically significant effect on gene transcription levels. This suggests that the transcription of early viral genes is inhibited immediately by BET/BRD4 inhibitors when these genes begin to be largely transcribed. Interestingly, B646L gene expression was also obviously inhibited under cotreatment with all 12 inhibitors, but 2 inhibitors had no significant effect on B646L gene transcription. Earlier addition of the inhibitors had a more notable inhibitory effect on ASFV, indicating that these 12 inhibitors act over the whole ASFV transcription process. Remarkably, two BET inhibitors (I-BET-762 and PLX51107) and two BRD4-specific inhibitors (ARV-825 and ZL0580) largely inhibited ASFV infection when applied in two ways (Fig. 2A and B). Furthermore, the duration of action of I-BET-762, PLX51107, ARV-825 and ZL0580 was investigated in ASFV-infected cells for 4, 12, 24, and 48 h (Fig. 4). The cells were treated with individual inhibitors for 16 h prior to ASFV infection, and B646L gene copy number was then detected. The data indicated that four inhibitors suppressed B646L gene replication in ASFV-infected cells from 12 hpi. For ZL0580, a stronger effect was observed in cells infected with ASFV. In addition, the cells were treated by BRD4 inhibitors prior to infected with ASFV at different time points dependent on the infected stages of its life cycle. Since B646L is a late expressed gene during infection cycle, we found that the copy number of this gene was not significant difference in early infected time (4 hpi). The results indicated that the inhibitors could suppress ASFV infection. In general, the inhibitory effects were time dependent. It is likely that BET/BRD4 inhibition induces cell cycle arrest or different biological activities; thus, the effects of these inhibitors on ASFV infection may vary by times being added to the culture.
Interestingly, some BET/BRD4 inhibitors do not affect PRV or PRRSV viral gene transcription (19), and, other previous studies have demonstrated that BRD4 facilitates viral infection through the regulation of HSV-1 and HSV-2 viral gene transcription and that, through inhibiting BRD4, HSV-1 and HSV-2 viral infection, gene transcription, and protein synthesis were significantly suppressed in a dose-dependent manner (21). This suggests that modulation of similar or same target proteins (e.g., BET protein family) or pathways by different regulatory agents (different BET/BRD4 inhibitors) may induce distinct functional outcomes in different viral infections. Epigenetic modifications in the cell may be changed by altering activity of BET/BRD4 to further affect the transcription and expression of both viral and host genes. Understanding the regulatory mechanism of BET/BRD4 inhibitors and their roles in ASFV infection needs further studies. Collectively, our results suggest that BET inhibitors have therapeutic potential for control of ASFV infection.
I-BET-762 inhibits BET proteins by occupying the acetyl-lysine-binding pocket of BET proteins, inhibiting the binding of BET proteins to acetylated histones, and thereby prevents the formation of chromatin complexes responsible for the expression of key inflammatory genes in activated macrophages and primary human monocytes (39). PLX51107 is a novel, structurally distinct BET inhibitor. In a group of cultured cells, treatment with PLX51107 for a short period (4 h) led to a sharp decrease in c-Myc levels but did not immediately cause an apoptotic response. After prolonged treatment time (continuous culture for 16 h or longer), PLX51107 induced apoptosis. Proteolysis targeting chimeric (PROTAC) molecules are a novel family of compounds with the ability to bind their target proteins and recruit a ubiquitin ligase, which promotes degradation of the targeted protein (40). ARV-825 is a PROTAC compound and BRD4 protein degrader that can recruit BRD4 to the E3 ubiquitin ligase cereblon to induce rapid, effective, and continuous degradation of the BRD4 protein, continuously lowering c-Myc levels (41, 42). Compared to other BRD4 inhibitors, ARV-825 treatment caused more significant changes in c-MYC levels and downstream cell proliferation and apoptosis induction (41). In our study, the dose-dependent inhibitory effects of ARV-825 were not as remarkable as those of ZL0580, PLX51107, or I-BET762. ARV-825 significantly inhibited ASFV CP204L and B646L mRNA and protein levels compared to those upon application of the other inhibitors at a lower concentration. To date, there have been no reports with respect to the effects of above three BET/BRD4 inhibitors on viral infection. ZL0580 is a BRD4-specific inhibitor that was designed by analyzing the crystal structures of available BRD4 modulators with the BRD4 BD1 domain (15). It displayed potent BRD4-binding activity with an IC50 of 163 nM against the BRD4 BD1 domain with 6.6-fold selectivity over the BRD4 BD2 domain. ZL0580 is a novel, BRD4-selective, small-molecule modulator that was reported to suppress both induced and basal HIV transcription and blocks viral reactivation events in human T cells and several latently infected myeloid cell lines. ZL0580 induces HIV suppression by inhibiting Tat-mediated transcription elongation and inducing a repressive chromatin structure at the HIV promoter (20, 43).
In our study, different assays (HAD, real-time PCR and WB analysis) showed that four inhibitors significantly inhibited ASFV infection in PAMs in a dose-dependent manner. A cumulative suppressive effect on ASFV infection was observed, suggesting that the BRD4 inhibitors specifically act on BRD4 to reduce ASFV infection (Fig. 5 and 6). After characterizing these four inhibitors, we speculate that BET/BRD4 is helpful for ASFV infection, and the virus may take advantage of BET/BRD4 that is released from chromatin to the viral genome to promote viral replication and gene transcription. ASFV encodes approximately 20 genes that are involved in the transcription and modification of its mRNA (24). Our results indicated that the transcript levels of nine related genes of ASFV were significantly decreased after treatment with the four individual BET/BRD4 inhibitors (Fig. 7). ASFV carries a set of enzymes similar to eukaryotic RNA Pol II, and their homology with RNA Pol II is higher than that with other nuclear or cytoplasmic large DNA molecules (28). Interestingly, ZL0580, PLX51107, I-BET762, and ARV-825 inhibit the nine subunits of ASFV RNA polymerase, which suggests that ZL0580, PLX51107, I-BET762, and ARV-825 exert their antiviral effects by altering ASFV transcription.
In summary, previous studies showed that the host nucleus is required for replication of ASFV, but the mechanism was not defined (5, 6). Since ASFV has itself RNA Pol II, host RNA Pol II is not required for ASFV gene transcription (44). However, even though the ASFV gene transcription predominantly occurred in cytoplasmic according to the description of earlier published studies (24), the roles of BET/BRD4 (which are located in nucleus) on ASFV replication remain unknown. The regulatory mechanisms and the role of epigenetic BET/BRD4 in ASFV infection need to be further investigated. This study provides multiple lines of evidence to support that the downregulation of early and late ASFV gene expression is associated with inhibition of BET/BRD4 activation and thus has a suppressive effect on ASFV infection. Extensive study of the role of BET/BRD4 in ASFV replication will be helpful to unravel the interactions between this virus and host cells and provide insights into the development of new approaches for the control of ASFV infection.
MATERIALS AND METHODS
Biosafety statement and facility.
All experiments carried out with live ASFV were performed in a biosafety level-3 (BSL-3) laboratory at the Lanzhou Veterinary Research Institute (LVRI), Chinese Academy of Agriculture and Sciences, and were accredited by the China National Accreditation Service for Conformity Assessment and approved by the Ministry of Agriculture and Rural Affairs. In the laboratory, to reduce any potential risk, all protocols were strictly followed, and all activities were monitored by the professional staff at LVRI and randomly inspected by local and central governmental authorities without advance notice.
Cells culture and ASFV.
Primary alveolar macrophages (PAMs) were isolated from 50- to 60-day-old specific-pathogen-free pigs and stored at the African Swine Fever Regional Laboratory (Lanzhou). The PAMs were cultured in RPMI 1640 medium (Thermo Scientific, USA) with l-glutamine and 25 mM HEPES (Gibco) supplemented with 10% fetal bovine serum (Gibco, Australia), 100 IU/mL penicillin, and 100 μg/mL streptomycin (Gibco/Life Technologies) at 37°C under 5% CO2. The ASFV strain used in this study (CN/SC/2019) was provided by the African Swine Fever Regional Laboratory (Lanzhou).
Antibodies and reagents.
For WB analysis, anti-p30, -p72, anti-pC315R, and anti-pH359L rabbit sera were raised against recombinant ASFV p30, p72 pC315R, and pH359L proteins and deposited at the African Swine Fever Regional Laboratory (Lanzhou), LVRI, of the Chinese Academy of Agricultural Sciences. Anti-CD2v mouse sera were kindly provided by Liguo Zhang from Institute of Biophysics, Chinese Academy of Sciences. Anti-β-actin (13E5) rabbit monoclonal antibody (catalog no. 4970) and anti-GAPDH (14C10) rabbit monoclonal antibody (catalog no. 2118) were purchased from Cell Signaling Technology. Anti-β-tubulin rabbit polyclonal antibody (catalog no. 10094-1-AP) and horseradish peroxidase (HRP)-conjugated AffiniPure goat anti-rabbit IgG(H+L) (catalog no. SA00001-2) were purchased from ProteinTech Group. Fluorescein isothiocyanate (FITC)-conjugated goat anti-rabbit IgG secondary antibody (catalog no. F0382) was purchased from Sigma-Aldrich. A Cell Counting Kit-8 (CCK-8; catalog no. K1018-30) was purchased from APExBIO (USA). TRIzol reagent (catalog no. 15596018), DAPI (4′,6′-diamidino-2-phenylindole; catalog no. 62248), octadecyl rhodamine B (R18, catalog no. O246), and radioimmunoprecipitation assay (RIPA) lysis and extraction buffers (catalog no. 89901) were purchased from Thermo Fisher Scientific. Filters (0.22-μm pore size) were purchased from Millipore.
BET/BRD4 chemical inhibitors.
Apabetalone (RVX-208; BET inhibitor, S7295), ARV-825 (BRD4 specific inhibitor, S8297), AZD-5153 6-hydroxy-2-naphthoic acid (BET/BRD4 inhibitor, S8344), CPI-203 (BET inhibitor, S7304), Molibresib (I-BET-762; BET inhibitor, S7189), (+)-JQ1 (BET inhibitor, S7110), INCB054329 (BET inhibitor, S8753), MS436 (BET inhibitor, S7305), Birabresib (OTX015; BET inhibitor, S7360), PLX51107 (a new BET inhibitor, S8739), and PFI-1 (PF-6405761; BRD2/BRD4 inhibitor, S1216) were purchased from Selleck.cn, and ZL0580 (a BRD4-specific inhibitor) was prepared as previously described (20, 45). The structures and functions of these inhibitors are shown in Fig. 3A, Text S1 in the supplemental material (see also Selleck.cn [https://www.selleck.cn]), and Table 1 (20, 41, 46–55).
TABLE 1.
BET/BRD4 chemical inhibitors used in this study
| Inhibitor | Functions | Reference(s) |
|---|---|---|
| Apabetalone (RVX-208) | An effective BET bromodomain inhibitor, acts on BD2. | 46 |
| ARV-825 | Recruit BRD4 to E3 ubiquitin ligase cereblon to induce rapid, effective, and continuous degradation of BRD4. | 41 |
| AZD5153 | BET/BRD4 bromodomain BD2 inhibitor, inhibits the target gene expression of nuclear receptor binding SET domain protein 3 (NSD3). | 47 |
| CPI-203 | Potent BET bromodomain inhibitor. | 48 |
| Molibresib (I-BET-762) | A highly selective inhibitor of BET family. | 49 |
| INCB-054329 | A BET family bromodomain inhibitor. | 50 |
| (+)-JQ1 | BET bromodomain inhibitor, (+)-JQ1 inhibits cell proliferation by inducing autophagy; (+)-JQ1 can inhibit the target gene expression of nuclear receptor binding SET domain protein 3 (NSD3). | 51 |
| MS436 | A BET bromodomain inhibitor. | 52 |
| Birabresib (OTX015) | Specifically binds to BRD2/3/4, inhibit the target gene expression of nuclear receptor binding SET domain protein 3 (NSD3). | 53 |
| PLX51107 | A novel BET inhibitor. Among non-BET proteins, PLX51107 only has a significant interaction with the bromine region of CBP and EP300 (p300). | 54 |
| PFI-1 (PF-6405761) | A highly selective BET inhibitor that acts on BRD4 and BRD2. | 55 |
| ZL0580 | ZL0580 is selectively bound to the BRD4 BD1 domain that induced epigenetic suppression of HIV via BRD4. | 20, 45 |
Cytotoxicity assay.
The cytotoxicity of 12 representative inhibitors in PAMs was evaluated by using a CCK-8 kit. Briefly, PAMs (2 × 105 cells per well) in 96-well cell culture plates were treated with the inhibitors at increasing concentrations (from 0.5 to 240 μM). The experiments included three replicates, and a blank and DMSO control were also included. The treated cells were incubated for 24 h at 37°C in 5% CO2 and, after incubation, 10 μL of CCK-8 reagent was added to each well, followed by incubation at 37°C for 1 to 4 h. The absorbance at 450 nm was measured using a microplate reader. The viability of the PAMs was calculated according to the following formula: cell viability (%) = [(ODinhibitor – ODblank)/(ODcontrol – ODblank)] × 100.
Virus HAD50 assay.
PAMs were seeded in 96-well plates and cultured overnight at 37°C under 5% CO2. The cells were then pretreated with DMSO, PLX51107, ARV-825, ZL0580, and I-BET-762 for 16 h; then, 10-fold serial dilutions (100 to 10−12) of virus were inoculated into each well (with eight replicates for each dilution), with pig erythrocytes (1:1,000) added to each well at the same time. The ASFV was quantified by the formation of characteristic rosettes formed through hemadsorption (HAD) of erythrocytes around the infected cells. HAD activity was observed for 7 consecutive days after inoculation, and the 50% HAD dose (HAD50) was calculated using the Reed and Muench method (56).
Time-of-addition assay.
PAMs in 12-well plates (2 × 106 cells/well) were seeded for ASFV infection. In the pretreatment assay, PAMs were treated with 12 individual BET/BRD4 inhibitors for 16 h before infection with ASFV CN/SC/2019 (MOI = 0.1). In the cotreatment assay, PAMs were exposed to 12 individual BET/BRD4 inhibitors at the same time that the ASFV was added to the plates. The plates were then incubated at 37°C under 5% CO2 for 24 h. In the posttreatment assay, cells were infected with ASFV, and the inhibitors were then added 4 h after infection. The plates were then incubated at 37°C under 5% CO2 for 16 h. DMSO-treated cells when then infected with ASFV for different assays in individual wells. The viruses were collected, titrated by HAD assay, and quantified by real-time PCR and WB analysis.
Quantification of cell-associated ASFV DNA or mRNA.
To quantify ASFV DNA or mRNA in ASFV-infected PAMs, total DNA or RNA was extracted from different PAM samples using a standard protocol with the QIAamp DNA blood kit (Qiagen, USA) according to the manufacturer’s instructions or TRIzol reagent (Life Technologies), followed by chloroform extraction and precipitation with isopropyl alcohol and ethanol. cDNA was synthesized from the RNA using a PrimeScript RT reagent kit with gDNA Eraser (TaKaRa Bio, Inc., Shiga, Japan) according to the manufacturer’s instructions. Gene expression in cDNA samples was measured by one-step qRT-PCR using a OneStep PrimeScript RT-PCR kit (Perfect Real Time) according to the manufacturer’s specifications (TaKaRa, Dalian, China). Quantitative real-time PCR was performed on the CFX Connect real-time PCR detection system (Bio-Rad). The sequences of primers and probes specific for the B646L gene were obtained according to the OIE-recommended sequence described in King et al. (57). Primer and probe sequences specific for other genes were designed in this study (Table 2). All samples were run and analyzed in duplicate. The B646L gene was amplified from DNA, and the copy number was calculated according to the description by King et al. (56). The RNA expression of each target gene in the PAMs was normalized to GAPDH expression and then calculated using the 2−ΔΔCT method.
TABLE 2.
Primers and probes used in this study
| Target | Primer (R and F) and probe sequences (5′–3′)a |
|---|---|
| CP204L | F: GAGGAGACGGAATCCTCAGC R: GCAAGCATATACAGCTTGGAGT FAM–ACCTCCGATGAGGGCTCTTGCT–TAMRA |
| B646L | F: CTGCTCATGGTATCAATCTTATCGA R: GATACCACAAGATCRGCCGT FAM–CCAGGAGCGAGATCCCGCCA–TAMRA |
| NP1450L | F: GGCTGGAGGTAGGAGACATC R: CCTATGCTGCTTCGTTCGAG FAM–CGTCACTGGCGACGTCGCGT–TAMRA |
| EP1242L | F: GAAACCACGGTTGGTCTAGC R: TGAAGATGGCCGCATCAAAG FAM–CAACGGCCAGACCGGCGAGT–TAMRA |
| H359L | F: AGGATTCCACGGACCTGTTT R: TTTAAGCTTAGGGCCTGCCA FAM–CCGCAGAGCAAATACCAGTGTCTCGT–TAMRA |
| D205R | F: ATCCCTACCACCTGTTCTGC R: TGACGCGCTAATTTGCATGA FAM–ACTCCTGCGCCTCCTCCTGAGT–TAMRA |
| CP80R | F: TATTGGAACCTACGCGGCAA R: AATGAGTGCGACAACACACC FAM–TTGCGGCAATGTTCCGCCCA–TAMRA |
| C315R | F: GGATCTTCTGCGCTCCCTAT R: CGCCGATGTTCTTCTCATCC FAM–ACAAATCCACCAAGAACTGCAGGAGGA–TAMRA |
| D339L | F: AATATGGAAAGGGCCCAAGG R: AACCCTAGGCTGCTGTTCTT FAM–TGTCGCGGCTTAAGCCTTGCA–TAMRA |
| C147L | F: TCATGGATGACCTCGTGGAG R: ACGATCTCGTCCTTGTCCTC FAM–ACTCCTCCTCACTGTCGACGAGGT–TAMRA |
| I243L | F: CGTTGTGGGACGATCAATCA R: ACGTCATGCTACCAATTGCC FAM–TCACCAACAACAGGATAACGATGCCCT–TAMRA |
| GAPDH | F: TGGAAAGGCCATCACCATCT R: ATGGTCGTGAAGACACCAGT FAM–CCAGGAGCGAGATCCCGCCA–TAMRA |
R, reverse; F, forward.
Western blotting analysis.
PAMs were seeded in 6-well plates overnight and treated with ZL0580, I-BET-76, PLX51107, ARV-825, or DMSO 16 h prior to inoculation, followed by infection with the ASFV CN/SC/2019 strain (MOI = 1) for 48 h. The cells were harvested, washed, and then lysed in RIPA lysis and extraction buffers supplemented with a protease inhibitor cocktail and 1 mM phenylmethylsulfonyl fluoride by rotation at room temperature for 1 h. The total protein concentration was measured by using a microplate BCA protein assay kit (Pierce; Thermo Fisher Scientific). Proteins were separated on an SDS-PAGE gel and then transferred to a nitrocellulose membrane (Merck Millipore, ISEQ00010), which was incubated with individual protein-specific primary antibodies at 4°C overnight on a shaker. The membrane was then incubated with HRP-linked secondary antibodies for 1 h at room temperature. The reaction was detected with Immobilon Western HRP substrate (B1911-100ML; Sigma). The corresponding grayscale value for each expressed protein band was analyzed using ImageJ software.
Confocal microscopy and flow cytometry.
Native ASFV P30 or P72 was identified in infected PAMs by using an indirect fluorescent antibody test. Briefly, PAMs seeded in a 2-cm laser confocal dish were pretreated with ZL0580 (10 μM), I-BET-762 (2 μM), PLX51107 (5 μM), ARV-825 (0.5 μM), or DMSO for 16 h. After treatment, the cells were infected with the ASFV CN/SC/2019 strain (MOI = 1) for 24 h and then washed twice with cold phosphate-buffered saline (PBS). The cells were fixed in a buffer containing 4% paraformaldehyde and 10 mM piperazine-N,N′-bis(2-ethanesulfonic acid) in PBS at pH 6.4 for 10 min. After one wash and incubation with specific anti-ASFV P30 or P72 sera (1:100) diluted in blocking buffer without Triton X-100 at 4°C overnight, the cells were stained with FITC-conjugated goat anti-rabbit IgG secondary antibody for 1 h at room temperature. Cells were acquired using confocal microscopy (Leica, TCS SP8) or the BD Accuri C6 Plus instrument, and the data were analyzed using the program FlowJo v10.6.2.
Statistical analysis.
Statistical analyses of all data were performed using Prism 8.0 (GraphPad Software, Inc.). Statistical comparisons between groups were performed using paired or nonpaired t tests. Two-tailed P values were determined, and a P value of <0.05 was considered to indicate statistical significance (*, P < 0.05; **, P < 0.01; ***, P < 0.001; ****, P < 0.0001). The quantitative data in all figures are expressed as the means ± the standard deviations (SD; indicated by the error bars). The CC50 was calculated by a linear regression analysis of dose-response curves generated from the obtained data. The 95% CIs for CC50 values were calculated using IBM SPSS Statistics v19.0.
ACKNOWLEDGMENTS
This study was financially supported by the National Natural Science Foundation of China (grant 32072830); the Gansu Provincial Major Project for Science and Technology Development (grant 20ZD7NA006); the State Key Laboratory of Veterinary Etiological Biology, Lanzhou Veterinary Research Institute, Chinese Academy of Agricultural Sciences (grant SKLVEB2020CGPY02); and the Basic Scientific Research Business Expenses Budget Incremental Project, Chinese Academy of Agricultural Sciences, Lanzhou Veterinary Research Institute (grant 1610312021002).
Q.N., G.G., Z.L., J.L., and H.Y. conceived and designed the study. Y.Z., S.Y., and Q.N. participated in the whole experiments. Q.N. wrote the manuscript. J.Y., Z.Z., S.G., and J.F. isolated the PAM cells. Z.L. and J.Z. synthesized the BRD4 inhibitor ZL0580 and revised the manuscript. H.H. analyzed the data and revised the manuscript.
We thank Yongxiang Fang from Lanzhou Veterinary Research Institute, Chinese Academy of Agriculture and Sciences, for help with the flow cytometry assay and AJE for English language editing. All authors reviewed the manuscript and declared that they have no competing interests.
Footnotes
Supplemental material is available online only.
[This article was published on 27 June 2022 with an incorrect affiliation listed for author Hong Yin. The affiliations for Hong Yin were updated in the current version, posted on 7 July 2022.]
Contributor Information
Qingli Niu, Email: niuqingli@caas.cn.
Hong Yin, Email: yinhong@caas.cn.
Daniel R. Perez, University of Georgia
REFERENCES
- 1.De la Torre A, Bosch J, Iglesias I, Muñoz MJ, Mur L, Martínez-López B, Martínez M, Sánchez-Vizcaíno JM. 2015. Assessing the risk of African swine fever introduction into the European Union by wild boar. Transbound Emerg Dis 62:272–279. doi: 10.1111/tbed.12129. [DOI] [PubMed] [Google Scholar]
- 2.Revilla Y, Pérez-Núñez D, Richt JA. 2018. African swine fever virus biology and vaccine approaches. Adv Virus Res 100:41–74. doi: 10.1016/bs.aivir.2017.10.002. [DOI] [PubMed] [Google Scholar]
- 3.Zhao D, Liu R, Zhang X, Li F, Wang J, Zhang J, Liu X, Wang L, Zhang J, Wu X, Guan Y, Chen W, Wang X, He X, Bu Z. 2019. Replication and virulence in pigs of the first African swine fever virus isolated in China. Emerg Microbes Infect 8:438–447. doi: 10.1080/22221751.2019.1590128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Gómez-Villamandos JC, Bautista MJ, Sánchez-Cordón PJ, Carrasco L. 2013. Pathology of African swine fever: the role of monocyte-macrophage. Virus Res 173:140–149. doi: 10.1016/j.virusres.2013.01.017. [DOI] [PubMed] [Google Scholar]
- 5.García-Beato R, Salas ML, Viñuela E, Salas J. 1992. Role of the host cell nucleus in the replication of African swine fever virus DNA. Virology 188:637–649. doi: 10.1016/0042-6822(92)90518-T. [DOI] [PubMed] [Google Scholar]
- 6.Rojo G, Garcia-Beato R, Vinuela E, Salas ML, Salas J. 1999. Replication of African swine fever virus DNA in infected cells. Virology 257:524–536. doi: 10.1006/viro.1999.9704. [DOI] [PubMed] [Google Scholar]
- 7.Simões M, Rino J, Pinheiro I, Martins C, Ferreira F. 2015. Alterations of nuclear architecture and epigenetic signatures during African swine fever virus infection. Viruses 7:4978–4996. doi: 10.3390/v7092858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Simões M, Freitas FB, Leitão A, Martins C, Ferreira F. 2019. African swine fever virus replication events and cell nucleus: new insights and perspectives. Virus Res 270:197667. doi: 10.1016/j.virusres.2019.197667. [DOI] [PubMed] [Google Scholar]
- 9.Ballester M, Rodríguez-Cariño C, Pérez M, Gallardo C, Rodríguez JM, Salas ML, Rodriguez F. 2011. Disruption of nuclear organization during the initial phase of African swine fever virus infection. J Virol 85:8263–8269. doi: 10.1128/JVI.00704-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Frouco G, Freitas FB, Martins C, Ferreira F. 2017. Sodium phenylbutyrate abrogates African swine fever virus replication by disrupting the virus-induced hypoacetylation status of histone H3K9/K14. Virus Res 242:24–29. doi: 10.1016/j.virusres.2017.09.009. [DOI] [PubMed] [Google Scholar]
- 11.Dixon LK, Chapman DA, Netherton CL, Upton C. 2013. African swine fever virus replication and genomics. Virus Res 173:3–14. doi: 10.1016/j.virusres.2012.10.020. [DOI] [PubMed] [Google Scholar]
- 12.Correia S, Ventura S, Parkhouse RM. 2013. Identification and utility of innate immune system evasion mechanisms of ASFV. Virus Res 173:87–100. doi: 10.1016/j.virusres.2012.10.013. [DOI] [PubMed] [Google Scholar]
- 13.Yu L, Wang Z, Zhang Z, Ren X, Lu X, Ding K. 2015. Small-molecule BET inhibitors in clinical and preclinical development and their therapeutic potential. Curr Top Med Chem 15:776–794. doi: 10.2174/1568026615666150302110135. [DOI] [PubMed] [Google Scholar]
- 14.Fu LL, Tian M, Li X, Li JJ, Huang J, Ouyang L, Zhang Y, Liu B. 2015. Inhibition of BET bromodomains as a therapeutic strategy for cancer drug discovery. Oncotarget 6:5501–5516. doi: 10.18632/oncotarget.3551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Liu Z, Wang P, Chen H, Wold EA, Tian B, Brasier AR, Zhou J. 2017. Drug discovery targeting bromodomain-containing protein 4. J Med Chem 60:4533–4558. doi: 10.1021/acs.jmedchem.6b01761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Devaiah BN, Gegonne A, Singer DS. 2016. Bromodomain 4: a cellular Swiss army knife. J Leukoc Biol 100:679–686. doi: 10.1189/jlb.2RI0616-250R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hsu SC, Blobel GA. 2017. The role of bromodomain and extraterminal motif (BET) Proteins in chromatin structure. Cold Spring Harbor Symp Quant Biol 82:37–43. doi: 10.1101/sqb.2017.82.033829. [DOI] [PubMed] [Google Scholar]
- 18.Wu SY, Chiang CM. 2007. The double bromodomain-containing chromatin adaptor Brd4 and transcriptional regulation. J Biol Chem 282:13141–13145. doi: 10.1074/jbc.R700001200. [DOI] [PubMed] [Google Scholar]
- 19.Wang J, Li GL, Ming SL, Wang CF, Shi LJ, Su BQ, Wu HT, Zeng L, Han YQ, Liu ZH, Jiang DW, Du YK, Li XD, Zhang GP, Yang GY, Chu BB. 2020. BRD4 inhibition exerts anti-viral activity through DNA damage-dependent innate immune responses. PLoS Pathog 16:e1008429. doi: 10.1371/journal.ppat.1008429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Niu Q, Liu Z, Alamer E, Fan X, Chen H, Endsley J, Gelman BB, Tian B, Kim JH, Michael NL, Robb ML, Ananworanich J, Zhou J, Hu H. 2019. Structure-guided drug design identifies a BRD4-selective small molecule that suppresses HIV. J Clin Invest 129:3361–3373. doi: 10.1172/JCI120633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ren K, Wei Z, Chen X, Ma Y, Dai Y, Fan Y, Hou Y, Tan RX, Li E. 2016. An epigenetic compound library screen identifies BET inhibitors that promote HSV-1 and -2 replication by bridging P-TEFb to viral gene promoters through BRD4. PLoS Pathog 12:e1005950. doi: 10.1371/journal.ppat.1005950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Wu SY, Lee AY, Hou SY, Kemper JK, Erdjument-Bromage H, Tempst P, Chiang CM. 2006. Brd4 links chromatin targeting to HPV transcriptional silencing. Genes Dev 20:2383–2396. doi: 10.1101/gad.1448206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.You J, Croyle JL, Nishimura A, Ozato K, Howley PM. 2004. Interaction of the bovine papillomavirus E2 protein with Brd4 tethers the viral DNA to host mitotic chromosomes. Cell 117:349–360. doi: 10.1016/s0092-8674(04)00402-7. [DOI] [PubMed] [Google Scholar]
- 24.Rodríguez JM, Salas ML. 2013. African swine fever virus transcription. Virus Res 173:15–28. doi: 10.1016/j.virusres.2012.09.014. [DOI] [PubMed] [Google Scholar]
- 25.Iyer LM, Balaji S, Koonin EV, Aravind L. 2006. Evolutionary genomics of nucleo-cytoplasmic large DNA viruses. Virus Res 117:156–184. doi: 10.1016/j.virusres.2006.01.009. [DOI] [PubMed] [Google Scholar]
- 26.Zhou X, Li N, Luo Y, Liu Y, Miao F, Chen T, Zhang S, Cao P, Li X, Tian K, Qiu HJ, Hu R. 2018. Emergence of African swine fever in China, 2018. Transbound Emerg Dis 65:1482–1484. doi: 10.1111/tbed.12989. [DOI] [PubMed] [Google Scholar]
- 27.Wang Q, Ren W, Bao J, Ge S, Li J, Li L, Fan X, Liu C, Wang H, Zhang Y, Xu T, Duan Y, Gu G, Zhou C, Wu X. 2018. The first outbreak of African swine fever was confirmed in China. China Animal Health Inspection 35:1–4. [Google Scholar]
- 28.Cackett G, Matelska D, Sýkora M, Portugal R, Malecki M, Bähler J, Dixon L, Werner F. 2020. The African swine fever virus transcriptome. J Virol 94:e00119-20. doi: 10.1128/JVI.00119-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Arabyan E, Kotsynyan A, Hakobyan A, Zakaryan H. 2019. Antiviral agents against African swine fever virus. Virus Res 270:197669. doi: 10.1016/j.virusres.2019.197669. [DOI] [PubMed] [Google Scholar]
- 30.Fan W, Jiao P, Zhang H, Chen T, Zhou X, Qi Y, Sun L, Shang Y, Zhu H, Hu R, Liu W, Li J. 2020. Inhibition of African swine fever virus replication by porcine type I and type II interferons. Front Microbiol 11:1203. doi: 10.3389/fmicb.2020.01203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mottola C, Freitas FB, Simões M, Martins C, Leitão A, Ferreira F. 2013. In vitro antiviral activity of fluoroquinolones against African swine fever virus. Vet Microbiol 165:86–94. doi: 10.1016/j.vetmic.2013.01.018. [DOI] [PubMed] [Google Scholar]
- 32.Hakobyan A, Galindo I, Nañez A, Arabyan E, Karalyan Z, Chistov AA, Streshnev PP, Korshun VA, Alonso C, Zakaryan H. 2018. Rigid amphipathic fusion inhibitors demonstrate antiviral activity against African swine fever virus. J Gen Virol 99:148–156. doi: 10.1099/jgv.0.000991. [DOI] [PubMed] [Google Scholar]
- 33.Arabyan E, Hakobyan A, Kotsinyan A, Karalyan Z, Arakelov V, Arakelov G, Nazaryan K, Simonyan A, Aroutiounian R, Ferreira F, Zakaryan H. 2018. Genistein inhibits African swine fever virus replication in vitro by disrupting viral DNA synthesis. Antiviral Res 156:128–137. doi: 10.1016/j.antiviral.2018.06.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Brasier AR, Zhou J. 2020. Validation of the epigenetic reader bromodomain-containing protein 4 (BRD4) as a therapeutic target for treatment of airway remodeling. Drug Discov Today 25:126–132. doi: 10.1016/j.drudis.2019.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Weller SK, Coen DM. 2012. Herpes simplex viruses: mechanisms of DNA replication. Cold Spring Harb Perspect Biol 4:a013011. doi: 10.1101/cshperspect.a013011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wu SY, Nin DS, Lee AY, Simanski S, Kodadek T, Chiang CM. 2016. BRD4 phosphorylation regulates HPV E2-mediated viral transcription, origin replication, and cellular MMP-9 expression. Cell Rep 16:1733–1748. doi: 10.1016/j.celrep.2016.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Nguyen TH, Maltby S, Eyers F, Foster PS, Yang M. 2016. Bromodomain and extra terminal (BET) inhibitor suppresses macrophage-driven steroid-resistant exacerbations of airway hyper-responsiveness and inflammation. PLoS One 11:e0163392. doi: 10.1371/journal.pone.0163392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Keck KM, Moquin SA, He A, Fernandez SG, Somberg JJ, Liu SM, Martinez DM, Miranda JL. 2017. Bromodomain and extraterminal inhibitors block the Epstein-Barr virus lytic cycle at two distinct steps. J Biol Chem 292:13284–13295. doi: 10.1074/jbc.M116.751644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Nicodeme E, Jeffrey KL, Schaefer U, Beinke S, Dewell S, Chung CW, Chandwani R, Marazzi I, Wilson P, Coste H, White J, Kirilovsky J, Rice CM, Lora JM, Prinjha RK, Lee K, Tarakhovsky A. 2010. Suppression of inflammation by a synthetic histone mimic. Nature 468:1119–1123. doi: 10.1038/nature09589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Wang P, Zhou J. 2018. Proteolysis targeting chimera (PROTAC): a paradigm-shifting approach in small molecule drug discovery. Curr Top Med Chem 18:1354–1356. doi: 10.2174/1568026618666181010101922. [DOI] [PubMed] [Google Scholar]
- 41.Lu J, Qian Y, Altieri M, Dong H, Wang J, Raina K, Hines J, Winkler JD, Crew AP, Coleman K, Crews CM. 2015. Hijacking the E3 ubiquitin ligase cereblon to efficiently target BRD4. Chem Biol 22:755–763. doi: 10.1016/j.chembiol.2015.05.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Noblejas-López MDM, Nieto-Jimenez C, Burgos M, Gómez-Juárez M, Montero JC, Esparís-Ogando A, Pandiella A, Galán-Moya EM, Ocaña A. 2019. Activity of BET-proteolysis targeting chimeric (PROTAC) compounds in triple negative breast cancer. J Exp Clin Cancer Res 38:383. doi: 10.1186/s13046-019-1387-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Alamer E, Zhong C, Liu Z, Niu Q, Long F, Guo L, Gelman BB, Soong L, Zhou J, Hu H. 2020. Epigenetic suppression of HIV in myeloid cells by the BRD4-selective small molecule modulator ZL0580. J Virol 94:e01880-19. doi: 10.1128/JVI.01880-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Salas J, Salas ML, Viñuela E. 1988. Effect of inhibitors of the host cell RNA polymerase II on African swine fever virus multiplication. Virology 164:280–283. doi: 10.1016/0042-6822(88)90646-0. [DOI] [PubMed] [Google Scholar]
- 45.Liu ZQ, Li Y, Chen H, Lai HT, Wang PY, Wu SY, Wold EA, Leonard PG, Joseph S, Hu HT, Chiang CM, Brasier AR, Tian B, Zhou J. 2022. Discovery, X-ray crystallography, and anti-inflammatory activity of bromodomain-containing protein 4 (BRD4) BD1 inhibitors targeting a distinct new binding site. J Med Chem 65:2388–2408. doi: 10.1021/acs.jmedchem.1c01851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Picaud S, Wells C, Felletar I, Brotherton D, Martin S, Savitsky P, Diez-Dacal B, Philpott M, Bountra C, Lingard H, Fedorov O, Müller S, Brennan PE, Knapp S, Filippakopoulos P. 2013. RVX-208, an inhibitor of BET transcriptional regulators with selectivity for the second bromodomain. Proc Natl Acad Sci USA 110:19754–19759. doi: 10.1073/pnas.1310658110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Bradbury RH, Callis R, Carr GR, Chen H, Clark E, Feron L, Glossop S, Graham MA, Hattersley M, Jones C, Lamont SG, Ouvry G, Patel A, Patel J, Rabow AA, Roberts CA, Stokes S, Stratton N, Walker GE, Ward L, Whalley D, Whittaker D, Wrigley G, Waring MJ. 2016. Optimization of a series of bivalent triazolopyridazine based bromodomain and extraterminal inhibitors: the discovery of (3R)-4-[2-[4-[1–(3-methoxy-[1,2,4]triazolo[4,3-b]pyridazin-6-yl)-4-piperidyl]phenoxy]ethyl]-1,3-dimethyl-piperazin-2-one (AZD5153). J Med Chem 59:7801–7817. doi: 10.1021/acs.jmedchem.6b00070. [DOI] [PubMed] [Google Scholar]
- 48.Hultmark S, Baudet A, Schmiderer L, Prabhala P, Palma-Tortosa S, Sandén C, Fioretos T, Sasidharan R, Larsson C, Lehmann S, Juliusson G, Ek F, Magnusson M. 2020. Combinatorial molecule screening identifies a novel diterpene and the BET inhibitor CPI-203 as differentiation inducers of primary acute myeloid leukemia cells. Haematologica 106:2566–2577. doi: 10.3324/haematol.2020.249177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Bandukwala HS, Gagnon J, Togher S, Greenbaum JA, Lamperti ED, Parr NJ, Molesworth AM, Smithers N, Lee K, Witherington J, Tough DF, Prinjha RK, Peters B, Rao A. 2012. Selective inhibition of CD4+ T-cell cytokine production and autoimmunity by BET protein and c-Myc inhibitors. Proc Natl Acad Sci USA 109:14532–14537. doi: 10.1073/pnas.1212264109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Drusbosky LM, Vidva R, Gera S, Lakshminarayana AV, Shyamasundar VP, Agrawal AK, Talawdekar A, Abbasi T, Vali S, Tognon CE, Kurtz SE, Tyner JW, McWeeney SK, Druker BJ, Cogle CR. 2019. Predicting response to BET inhibitors using computational modeling: a BEAT AML project study. Leuk Res 77:42–50. doi: 10.1016/j.leukres.2018.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Filippakopoulos P, Qi J, Picaud S, Shen Y, Smith WB, Fedorov O, Morse EM, Keates T, Hickman TT, Felletar I, Philpott M, Munro S, McKeown MR, Wang Y, Christie AL, West N, Cameron MJ, Schwartz B, Heightman TD, La Thangue N, French CA, Wiest O, Kung AL, Knapp S, Bradner JE. 2010. Selective inhibition of BET bromodomains. Nature 468:1067–1073. doi: 10.1038/nature09504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Zhang G, Plotnikov AN, Rusinova E, Shen T, Morohashi K, Joshua J, Zeng L, Mujtaba S, Ohlmeyer M, Zhou MM. 2013. Structure-guided design of potent diazobenzene inhibitors for the BET bromodomains. J Med Chem 56:9251–9264. doi: 10.1021/jm401334s. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Berthon C, Raffoux E, Thomas X, Vey N, Gomez-Roca C, Yee K, Taussig DC, Rezai K, Roumier C, Herait P, Kahatt C, Quesnel B, Michallet M, Recher C, Lokiec F, Preudhomme C, Dombret H. 2016. Bromodomain inhibitor OTX015 in patients with acute leukaemia: a dose-escalation, phase 1 study. Lancet Haematol 3:e186-95–e195. doi: 10.1016/S2352-3026(15)00247-1. [DOI] [PubMed] [Google Scholar]
- 54.Ozer HG, El-Gamal D, Powell B, Hing ZA, Blachly JS, Harrington B, Mitchell S, Grieselhuber NR, Williams K, Lai TH, Alinari L, Baiocchi RA, Brinton L, Baskin E, Cannon M, Beaver L, Goettl VM, Lucas DM, Woyach JA, Sampath D, Lehman AM, Yu L, Zhang J, Ma Y, Zhang Y, Spevak W, Shi S, Severson P, Shellooe R, Carias H, Tsang G, Dong K, Ewing T, Marimuthu A, Tantoy C, Walters J, Sanftner L, Rezaei H, Nespi M, Matusow B, Habets G, Ibrahim P, Zhang C, Mathé EA, Bollag G, Byrd JC, Lapalombella R. 2018. BRD4 profiling identifies critical chronic lymphocytic leukemia oncogenic circuits and reveals sensitivity to PLX51107, a novel structurally distinct BET inhibitor. Cancer Discov 8:458–477. doi: 10.1158/2159-8290.CD-17-0902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Picaud S, Da Costa D, Thanasopoulou A, Filippakopoulos P, Fish PV, Philpott M, Fedorov O, Brennan P, Bunnage ME, Owen DR, Bradner JE, Taniere P, O’Sullivan B, Müller S, Schwaller J, Stankovic T, Knapp S. 2013. PFI-1, a highly selective protein interaction inhibitor, targeting BET Bromodomains. Cancer Res 73:3336–3346. doi: 10.1158/0008-5472.CAN-12-3292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Reed LJ, Muench H. 1938. A simple method of examining fifty percent endpoints. Am J Hyg 27:493–497. [Google Scholar]
- 57.King DP, Reid SM, Hutchings GH, Grierson SS, Wilkinson PJ, Dixon LK, Bastos AD, Drew TW. 2003. Development of a TaqMan PCR assay with internal amplification control for the detection of African swine fever virus. J Virol Methods 107:53–61. doi: 10.1016/s0166-0934(02)00189-1. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplemental material. Download spectrum.02419-21-s0001.pdf, PDF file, 0.2 MB (180.6KB, pdf)