

Abstract
The intestinal epithelium is responsible for food digestion and nutrient absorption and plays a critical role in hormone secretion, microorganism defense, and immune response. These functions depend on the integral single-layered intestinal epithelium, which shows diversified cell constitution and rapid self-renewal and presents powerful regeneration plasticity after injury. Derailment of homeostasis of the intestine epithelium leads to the development of diseases, most commonly including enteritis and colorectal cancer. Therefore, it is important to understand the cellular characterization of the intestinal epithelium at the molecular level and the mechanisms underlying its homeostatic maintenance. Single-cell technologies allow us to gain molecular insights at the single-cell level. In this review, we summarize the single-cell RNA sequencing applications to understand intestinal cell characteristics, spatiotemporal evolution, and intestinal disease development.
Keywords: scRNA-seq, Gut, Heterogeneity, Enteritis, Colorectal cancer
Background
The intestine plays an important role in nutrient digestion and absorption, hormone secretion, protection and immune regulation (Peterson and Artis, 2014; Sanger and Lee, 2008; Zorn and Wells, 2009). Also, the intestine is the main organ for microbiota residing that has been showing continuously emerging functions to influence other organs in the body (Tilg et al., 2020). These functions depend on the intestinal epithelium that constitutes the second-largest epithelial surface area of more than 30m2 in the human body (Helander and Fandriks, 2014). The intestinal epithelial surface is expanded by millions of special structures, crypt-villus units in the small intestine or crypt units in the large intestine (Fig. 1A). The intestinal epithelium undergoes a fast turnover, which is driven by the intestinal stem cells (ISCs) located at the bottom of crypts (Fu et al., 2021). ISCs continuously self-renew and meantime generate transient-amplifying (TA) cells and progenitors, which then differentiate into mature functional cell types. Briefly, TA cells from ISCs give rise to secretory progenitors and enterocyte progenitors. Then the secretory progenitors further differentiate into goblet cells, enteroendocrine cells (EECs), Paneth cells, and perhaps tuft cells, while enterocyte progenitors differentiate into enterocytes (Fig. 1B). However, it has been reported that tuft cells may be from enterocytes but not secretory progenitors (Herring et al., 2018). During the differentiation progress, Paneth cells move down to the bottom of small intestinal crypts and are long-lived, whereas other mature lineages move up to keep the crypt or villus structures. These cell types cooperate with each other to form a multifunctional network. After the functional performance, these mature cells undergo cell death in a cycle of 4–5 days (Cheng and Leblond, 1974), and up to 1011 epithelial cells are lost every day in the human intestine (Leblond and Walker, 1956). This dynamic renewal system provides the intestine epithelium the ability to endure continuous chemical stimulation and various damages.
Fig. 1.

The structure and homeostatic maintenance of the intestinal epithelium. A, The small intestine is enriched with enterocytes and contains the crypt and villus structures, while the large intestine is enriched with goblet cells and only has the crypt structures. The cell constitution of both small and large intestinal epithelia is driven by Lgr5 + intestinal stem cells (ISCs) at the bottom of the crypt. The fate of ISCs is regulated by niche factors from the surrounding stromal cells or the epithelium cells themselves, such as Paneth cells. Wnt and EGF signaling promote proliferation, and their activities are high in the crypt and decrease gradually towards the villus. In contrast, BMP signaling promotes differentiation, inhibits proliferation, and induces cell death, and its activity increases gradually towards the villus tip. B, The proliferation and differentiation trajectory of intestinal epithelium. ISCs undergo self-renewal and meanwhile generate transit-amplifying (TA) cells. TA cells are fast proliferating and produce enterocyte and secretory progenitors, which further differentiate into enterocytes and Paneth, goblet, enteroendocrine and tuft cells, respectively. Tuft cells have also been suggested to derive from enterocyte progenitors
The dynamic balance of intestinal epithelium is maintained by the underneath niche cells and the microenvironment signaling (Zhu et al., 2021a). Most importantly, epidermal growth factor (EGF) and Wnt are abundant in the crypt to promote the proliferation of ISCs and TA cells, while bone morphogenetic protein (BMP) activity is higher at the villus, promotes cell differentiation and induces cell death (Fig. 1A) (Qi and Chen, 2015; Wang and Chen, 2018). These microenvironment signals are mainly from the niche cells, including stromal cells, fibroblast cells, immune-related cells, and others (Fig. 1A). For example, Forkhead box L1 (Foxl1) positive trophocytes can provide a source of Wnt-activation and BMP-inhibition signals for ISC self-renewal (Shoshkes-Carmel et al., 2018), and T helper cells can support ISC renewal through IL-10 (Biton et al., 2018). In short, the surrounding niche cells and signals are necessary for the epithelial function performance; the destruction of the microenvironment homeostasis may lead to intestinal disease development.
With the discovery of the specific ISC marker Lgr5 (Barker et al., 2007), we have gained a great understanding of intestinal epithelium homeostasis in the last decade. A series of breakthroughs have been made in this field, such as intestinal organoid culture from single Lgr5+ cells (Clevers, 2016; Sato et al., 2009), stem cell contribution to intestinal tumorigenesis (Llado et al., 2015), and the repopulation of human colonic epithelium in vivo with potential therapeutic applications in the mouse model (Sugimoto et al., 2018). However, as in other scientific research fields, new findings beget new questions. For instance, the exact cell constitution and ratio of different cell types in small and large intestinal epithelia are unclear, the critical factors governing the fate trajectory of ISCs are not fully determined, and the cellular and molecular changes under intestinal pathological conditions are not well understood. In addition, cellular heterogeneity is an important theme for intestinal epithelium researches. For example, EECs can be divided into several subtypes (Gribble and Reimann, 2016), but the substantial crossover or the cellular heterogeneity is rarely revealed. Also, the mechanism of intestinal disease development is obscure, such as intestinal enteritis and colorectal cancer (CRC). These emerging questions require new advanced technologies to offer us a more comprehensive picture.
Although whole transcriptome analysis using bulk RNA-sequencing has yielded massive information on cell function and biological regulation, individual cells are the life identity and functional unit. Also, cellular heterogeneity is a general feature of biological tissues (Wen and Tang, 2016). To better understand the cellular composition and heterogeneity of primary tissues at the same time, single-cell PCR gene-expression analysis was developed about 10 years ago (Dalerba et al., 2011). Then, with the marked improvement of the single-cell RNA sequencing (scRNA-seq) technology during the past years (Grun et al., 2015; Magness et al., 2013), thousands of individual cells can be processed in a single experiment and more than 50,000 reads for each cell obtained (Chen et al., 2021b). Nowadays, scRNA-seq is widely used for routine transcriptional profiling at single-cell resolution (Zhao et al., 2021). Our cross-species analysis of the single-cell transcriptome of the ileum epithelium identified a new CA7 + cell type in pig, macaque, and human ileum and revealed the distinct expression pattern in enterocytes, EECs, and Paneth cells among different species (Li et al., 2022). In this review, we focus on an advanced understanding of the intestine biology gained from single-cell transcriptome analysis and provide an updated summary of the scRNA-seq database in the field.
ScRNA-seq reveals cellular heterogeneity of the intestinal epithelium
Multiple functions of the intestinal epithelium are based on complex cell constitution with each cell type performing specific functions while coordinating with each other (Liu and Chen, 2020). Therefore, the identification of cell types is critical for understanding their detailed functions. Before the scRNA-seq technology application, identification of different cellular signatures at the transcriptome level was usually achieved by bulk RNA sequencing after fluorescence-activated cell sorter (FACS) sorting by cell-type specific markers (Table 1). However, bulk RNA-seq cannot reveal the cellular heterogeneity among these cells. ScRNA-seq makes it possible to detect the subtypes and their function difference at the single-cell level of the intestinal epithelium and the surrounding niche cells. More information about ISCs can be found in a recent review (Fu et al., 2021).
Table 1.
Summary of the specific markers of intestinal epithelial cells
| Cell types | Marker genes | |
|---|---|---|
| Mouse intestine | Human intestine | |
| Enterocyte | Alpi, Fabp1, Slc26a3 (Apoa1 in SI *) | ALPI, FABP1, SLC26A3 (APOA1 in SI *) |
| Goblet cell | Muc2, Spdef, Clca1, Zg16, Spink4, Fcgbp | MUC2, SPDEF, CLCA1, ZG16, SPINK4, FCGBP |
| Enteroendocrine cell | Neurog3, Neurod1, Chga, Chgb | NEUROG3, NEUROD1, CHGA, CHGB |
| Paneth cell | Lyz1, Defa5, Defa6 | LYZ, DEFA5, DEFA6 |
| Tuft cell | Dclk1, Pou2f3, Trpm5, Il25 | POU2F3, TRPM5, IL25 |
| TA cell (Lgr5-negative) | Mki67, Hmgb2, Top2a, Ube2c, Stmn1 | MKI67, HMGB2, TOP2A, UBE2C, STMN1 |
| Stem cell | Lgr5, Ascl2, Rgmb, Smoc2, (Olfm4 in SI *) | LGR5, ASCL2, RGMB, SMOC2, (OLFM4 in SI *) |
| BEST4 cell | Absent in mice | BEST4, SPIB, CA7, OTOP2 |
| + 4 stem cell | Bmi1, Hopx, Tret, Lrig1, Mex3a | Need to be confirmed |
* SI small intestine
Enterocytes
Enterocytes, a predominant cell type in the intestine, are responsible for food digestion and nutrient absorption, and these cells are derived from enterocyte progenitors (Pinto et al., 2003). Several transcript factors were found to determine the differentiation of TA cells to enterocyte progenitors (or immature enterocytes), such as Hes1, Cdx2, and Hnf4 (Clevers and Batlle, 2013). Based on scRNA-seq, more transcription factors (Sox4, Foxm1, Mxd3, and Batf2) were identified to associate with enterocyte maturation in mouse small intestine (Haber et al., 2017). Similarly, FABP1 and KRT19 were found to participate in enterocyte maturation in the human small intestine (Fujii et al., 2018). In addition, a new subtype of enterocytes was found in the human colon, BEST4+ enterocytes, which comprise about 1% of the human ileal epithelium (Huang et al., 2019; Parikh et al., 2019; Smillie et al., 2019) and are enriched with the genes responsible for pH sensing and electrolyte transportation, including GUCA2A, OTOP2, and CA7 (Brenna et al., 2015; Tu et al., 2018). These studies expand our knowledge about the functions of enterocytes, which are responsible for nutrient absorption and electrolyte balance.
Goblet cells
Goblet cells synthesize and secrete mucins to prevent pathogen invasion and stabilize the bacterial biofilm (Zhang and Wu, 2020). Goblet cells are more enriched and contribute to two mucous layers in the large intestine: the outer layer is relatively loose and contains symbiotic bacteria, and the inner layer is tight and impervious to bacteria, while there is only one antibacterial gradient in the small intestine (Paone and Cani, 2020). The active mucin secretion in the large intestine is consistent with more goblet cells there compared with the small intestine (Wang et al., 2020b). The destruction of mucous layers and goblet cell functions may lead to enteritis development (Birchenough et al., 2016). ScRNA-seq trajectory analysis of mCherry-Muc2 cells showed that goblet cells segregate into two separate trajectories: one is enriched with canonical markers (Clca1 and Fcgbp), while the other one is typically associated with enterocytes (Dmbt1 and Gsdmc4) (Nystrom et al., 2021). This study also revealed a new subpopulation of goblet cells (intercrypt goblet cells, icGCs), which are especially located to the surface epithelium between crypts in the colon and contribute to a functional mucus barrier to protect the epithelium from microorganisms (Nystrom et al., 2021). Furthermore, scRNA-seq uncovered an immature goblet cell type, which is absent of TBX10 expression, and was reduced during inflammation (Smillie et al., 2019). Another scRNA-seq study revealed that goblet cells could be divided into two main states: proliferation and differentiation. HES6 was found in the early stage of goblet cell differentiation, which can be used to mark the goblet cells that are not yet morphologically identifiable as goblet cells (Zhang et al., 2019a). Similarly, mitotic goblet cells were identified by the co-expression of MKI67, UBE2C, ZG16, TFF3, and CLCA1 in the human colon (Huang et al., 2019). Mature TFF1+ goblet cells are found only in the villus of the human ileum and in the top zone of crypts of the human colon and rectum, and they are highly enriched with the function of MHC antigen processing (Wang et al., 2020b). These results suggest that goblet cells show more heterogeneity along the differentiation trajectory.
Paneth cells
Paneth cells, which are interspersed between Lgr5+ ISCs at the crypt base in the small intestine, secrete antimicrobial molecules modulating host-microbe interactions and provide the factors promoting Lgr5+ ISCs (Clevers and Bevins, 2013; Zhang and Liu, 2016). However, whether there are functionally equivalent cells in the large intestine is still under debate. With the application of single-cell technology, we have gained more insights into it. Single-cell PCR gene expression analysis identified a subset of cKit+ goblet cells in the mouse colon, which might have the equivalent function of Paneth cells in supporting Lgr5+ stem cells (Rothenberg et al., 2012). Furthermore, scRNA-seq of the human embryo’s digestive tract and adult intestinal segments unraveled the Paneth-like cells in human fetal and adult large intestine, respectively (Gao et al., 2018; Wang et al., 2020b). Interestingly, another scRNA-seq analysis revealed that LYZ, a Paneth cell marker, was upregulated in lower-crypt goblet cells and might mark the “deep crypt secretory cells” that are required to maintain the colonic stem cell niche and to protect stem cells from bacterial damage during colitis (Parikh et al., 2019). Moreover, scRNA-seq also uncovered the marker genes of Paneth cells in human ascending, transverse, and descending colons (Burclaff et al., 2022). Noteworthy, the cells designated as Paneth cells or Paneth-like cells in previous reports were designed as BEST4+ cells based on the expression of LYZ, SPIB, BEST4, and CA7 (Burclaff et al., 2022; Wang et al., 2020b). Therefore, in addition to the canonical Paneth cells in the small intestine, the large intestine hosts cKit+/BEST4+/Paneth-like cells.
Enteroendocrine cells
EECs can sense nutrients and secrete about 20 different kinds of hormones (Furness et al., 2013). They have been divided into several subtypes based on hormone secretion (Gribble and Reimann, 2016). The scRNA-seq analysis of mouse intestine has uncovered hormone cross-expression among these subtypes. Haber and colleagues have re-defined the subtypes of mouse intestinal EECs into 12 sub-clusters and provided the hormones’ secretion atlas (Fig. 2A) (Haber et al., 2017). However, 9 sub-clusters EECs were assigned in the human small and large intestine in a single-cell transcriptomic atlas (Beumer et al., 2020): motilin (M cells), gastrin (G cells), gip (K cells), and cholestocystokin (I cells) were enriched in the duodenum, while glucagon (L cells) enriched in the colon. These results reveal different enteroendocrine subtypes between human and mouse intestines, including gene expression, such as ASCL1 and MNX1 in the human but not in mouse EECs (Beumer et al., 2020). A hormone gradient along the crypt-to-villus axis was revealed by scRNA-seq. For instance, tachykinin 1 (Tac1) and glucagon (Gcg, induced by Glp1) are highly expressed in the crypt, while secretin (Sct) and neurotensin (Nts) are high in the villus (Fig. 2B) (Beumer et al., 2018). Moreover, a hormone gradient is also observed along the small intestine from the proximal to distal segment: cholecystokinin (Cck) and gastrin (Gast) are enriched in the proximal segment, while proglucagon (Gcg) and peptide YY (PYY) enriched in the distal segment (Fig. 2C) (Beumer et al., 2020). Moreover, 10 major EEC subtypes were identified in adult Drosophila midgut, and these subtypes produce approximately 14 different classes of hormone peptides with each subtype on average secreting approximately 2–5 classes (Guo et al., 2019), demonstrating the complexity of the endocrine system in the intestine.
Fig. 2.

Enteroendocrine subtypes and their spatial distribution. A, Main subtypes of enteroendocrine cells and their secreted representative hormones (9 main subtypes in the human intestine are marked by blue). B, Secreted hormones show the spatial difference along the villus-crypt axis. TRPA1, TAC1, and GLP1 are enriched in the crypt, NTS, SCT, and PYY are enriched in the villus, while GIP, CCK, SST, LAPP, and TPH1 show no significant difference. C, Intestinal segmental enrichments of secreted hormones. CCK, GAST, GHRL, and GIP are enriched in the proximal segment, GCG, NTS, PYY, GLP1, NTS, SCGN, UCN3, SST, and REG4 are enriched in the distal segment, while CHGA, CHGB, PCSK1N, SCG2, SCG3, SST, MLN, TPH1, and SCT show no significant difference between the proximal and distal segments
Along with the exploration of enteroendocrine subtypes, new marker genes are also identified by scRNA-seq. Reg4 may be a novel marker for both ChgA/ChgBhigh and ChgA/ChgBlow EECs (Grun et al., 2015) and Neurog3, Neurod1, and Sox4 as new markers for enteroendocrine precursors (Gehart et al., 2019). ScRNA-seq also revealed that the EECs marked by Bmi1 and Prox1 could serve as a reservoir to replenish homeostatic and injury-inducible ISCs (Yan et al., 2017).
Tuft cells
Tuft cells, marked by Dclk1, Pou2f3, Trpm5, Il-25 (O'Leary et al., 2019), are rare solitary chemosensory cells in mucosal epithelia (Gerbe and Jay, 2016; Gerbe et al., 2012). These cells are involved in the initiation of immune type 2 responses (Gerbe and Jay, 2016) and produce a spectrum of biological effector molecules (Il-33, Il-25, prostaglandin D2, and the neurotransmitter acetylcholine) to mediate a cytokine-mediated cellular response (Gerbe and Jay, 2016; Schneider et al., 2019). Tuft cells closely interact with lymphoid cells (ILC2s) and recruit type 2 helper T cells (Th2 cells) to regulate immune activities in the intestine (Grencis, 2015; von Moltke et al., 2016). ScRNA-seq analysis has yielded new insights into tuft origin, differentiation progress, subtype, and functional subdivision.
Due to their secretion function, tuft cells were usually thought to originate from secretory progenitors that were regulated by Atoh1 (Gerbe et al., 2012). However, using scRNA-seq, Herring and colleagues reported that tuft cells in the small intestine appeared distinct from the secretory lineage goblet and Paneth cells while showing a common trajectory with enterocytes (Herring et al., 2018). This study suggests tuft cells may not originate from secretory lineage. ScRNA-seq revealed that Dclk1-positive tuft cells express high levels of Cox2 and Hopx in the mouse small intestine compared with the tuft cells in the colon (McKinley et al., 2017). ScRNA-seq studies also suggested that mature tuft cells may have two subtypes, one involved in the immune-related reaction and another related to neuronal development (Haber et al., 2017). These two subtypes present different enrichments in Il-33, the pan immune marker CD45, and the Th2-related cytokines Il-4 and Il-13 (Haber et al., 2017). Interestingly, tuft cell number increases after Paneth cell ablation and may act as novel niche cells in Paneth cell-ablated crypts (van Es et al., 2019). Moreover, the single-cell transcriptomic analysis showed that tuft cells may play an important role in intestinal enteritis—the enrichment of PSMA6 (Proteasome 20S subunit alpha 6, associated with Crohn’s disease risk) in tuft cells may lead to impaired epithelial integrity and stress responses (Huang et al., 2019).
ScRNA-seq unveils spatiotemporal regulation
Along the crypt-villus axis, cells display multifarious states, and the same cell type may have specialized functions (Beumer et al., 2022; Moor et al., 2018). The functional diversity also exists from the proximal to the distal small and large intestine (Wang et al., 2020b). In other words, each cell could be precisely regulated, and the same type of cells may show cellular heterogeneity along with the spatiotemporal points. This spatiotemporal regulation makes the epithelium flexible to adapt to specialized functions and various environments such as nutrient constitution (Wang et al., 2020b), microbiota (Boulange et al., 2016), pathogens (Peterson and Artis, 2014), hypoxia (Zheng et al., 2015), chemical concentration changes (Williamson and Clifford, 2017) and chymopoiesis (Bhat et al., 2020). Spatial difference along the crypt-villus axis has been explored with bulk RNA sequencing (George et al., 2008; Mariadason et al., 2005), but the effect of cell heterogeneity and precise differentiation state cannot be taken into consideration. The studies based on scRNA-seq have provided insightful information on the spatiotemporal regulation of the intestinal epithelium.
Enterocytes exhibit significant functional differences along the crypt-villus axis according to scRNA-seq combined with RNA-seq of laser capture micro-dissected tissue (LCM-RNA-seq) (Moor et al., 2018). Amino acid and carbohydrate transporters were enriched in the middle of the villus, while the proteins involved in lipoprotein and chylomicron biosynthesis are mainly expressed at the top of the villus. Reg family members and other peptides involved in the microbiota-host interactions are highly expressed at the bottom of the villus. The spatially functional difference of enterocytes, such as the increased lipid uptake at the top of the villus and the Reg expression at the bottom of the villus, is confirmed by the analysis of scRNA-seq and Bmpr1a knockout mouse model, and the zonated gene expression is controlled by BMP signaling along the crypt-villus axis (Beumer et al., 2022). ScRNA-seq analysis also unraveled that enterocytes at the bottom and middle of the villus show more plasticity than the ones at the tip (Ayyaz et al., 2019).
Different intestinal segments exhibit distinct activities in nutrient absorption. ScRNA-seq analysis of the intestinal epithelium of the human ileum, colon, and rectum reveals the differences of signature genes and nutrient transporters in enterocytes among these three segments (Wang et al., 2020b). Although the genes related to protein digestion and absorption, mineral and organic substance transports are evenly expressed in all three segments, the genes participating in lipid metabolism and drug metabolic processes are highly expressed in the ileum, while the genes related to small molecule transport were enriched in the large intestine. The scRNA-seq analysis of the gene expression profiles of the human embryo digestive tract between 6 and 25 weeks of gestation showed that the genes involved in protein digestion and absorption were enriched in the small intestine compared with the stomach and large intestine (Gao et al., 2018). A large-scale single-cell spatiotemporal atlas of the human embryo digestive tract ranging from 8 to 22 post-conceptual weeks further revealed that the spatial distribution of absorptive genes is established in development prior to crypt formation (Fawkner-Corbett et al., 2021). Another spatial scRNA-seq of epithelial cells from the human duodenum to descending colon also showed the segmental absorption difference—fatty acid, glucose, and cholesterol transporters were enriched in the small intestine, while sodium transporters were enriched in the colon (Burclaff et al., 2022).
Like enterocytes, EECs also display spatial distribution in mice and humans based on scRNA-seq. EECs show differential hormone secretion in the villus (Sct, PYY, Nts) and crypts (Tac1, Glp1, Trpa1), while other hormones (Cck, Sst, Lapp, Tph1, Gip) show no significant difference along the crypt-villus axis in the mouse small intestine (Fig. 2B) (Beumer et al., 2018). Also, some hormones show segmental enrichment. The hunger-related hormones Ghrl and Gcg are enriched in the mouse duodenum, while PYY, an appetite reducer upon feeding, is found mainly in the mouse ileum (Fig. 2C) (Haber et al., 2017). In addition, some of the hormones are highly expressed in the human ileum (SCT, NTS, and CCK), while some are enriched in the human colon and rectum (PAM, NMB, and INSL5) (Wang et al., 2020b). Gip-producing K cells are enriched in the proximal part of the mouse small intestine, while Glp1-producing L cells in the distal segment, Sst-producing D cells, and enterochromaffin cells are uniformly distributed (Beumer et al., 2018). Consistent with the mouse results, GAST-producing G cells and CCK-producing I cells are enriched in the proximal segment, whereas NTS-producing N cells and GHRL-producing X cells are in the human distal small intestine (Beumer et al., 2020).
Regional transcription factors may contribute to EEC diversity according to scRNA-seq of the Drosophila midgut. Class-specific Mirr and Ptx1 define Tk+ and AstC+ EEC types, respectively, while regional transcription factors contribute to segmental EEC identities. For instance, Drm defines the ITP+ EEC type in the posterior midgut, and Esg defines the NPLP2+ EEC type in the middle midgut (Guo et al., 2019). A time-resolution EEC subtype differentiation tree is given to depict common and lineage-specific transcriptional regulators during fate trajectory (Williamson and Clifford, 2017). Using a bi-fluorescent reporter (Neurog3-Chrono) to precisely position single-cell transcriptome along a time axis in mouse organoids, scRNA-seq identified temporal transcriptional regulators during enteroendocrine differentiation: Neurog3, Dll1, C1qbp function in the early stage, Pax4, Insm1, Arx in the intermediate stage, and Isl1, Pax6, Elf4 in the late stage (Williamson and Clifford, 2017).
Like other organs, the intestine also undergoes structural and functional changes during organism aging (Pentinmikko and Katajisto, 2020), for instance, disruption of the intestinal barrier (Parrish, 2017), increased risk of inflammatory conditions (Shemtov et al., 2022), the loss of the regenerative capacity of ISCs and epithelial renewal (Funk et al., 2020; Mihaylova et al., 2018; Nalapareddy et al., 2017). Like other organs, the intestine also undergoes structural and functional changes during organism aging (Pentinmikko and Katajisto, 2020), for instance, disruption of the intestinal barrier (Parrish, 2017), increased risk of inflammatory conditions (Shemtov et al., 2022), the loss of stem cell plasticity and epithelial renewal (Funk et al., 2020). ScRNA-seq analysis of cultured mouse organoids further confirmed the reduced regenerative capacity in senescent ISCs and identified three TFs (Egr1, Irf1, Fosb) that were downregulated in aged ISCs and might account for 80% of the age-specific ISC transcriptome changes (Nefzger et al., 2022). The reduced canonical Wnt signaling activity in ISCs, Paneth cells, and mesenchyme may cause impaired ISC function upon aging (Nalapareddy et al., 2017). Consistently, the number of crypts and TA cells were decreased in the aging mouse intestine, which may be driven by mTORC1 via a p38 MAPK-p53 pathway (He et al., 2020). ScRNA-seq analysis of aging mouse colon also revealed that aging might cause a shift from absorptive to secretory epithelial cells, thus contributing to age-associated intestinal disturbances, such as malabsorption (Sirvinskas et al., 2022).
ScRNA-seq reveals the mechanisms underlying enteritis
The gastrointestinal mucosa is the largest immunological organ in the body and represents a challenging environment where a meticulous balance must be maintained between tolerance and immune response from the huge microbiota burden (Abraham and Cho, 2009). Breakdown of the symbiotic relationship between the intestinal commensal microflora and the mucosal immune system leads to enteritis, such as inflammatory bowel disease (IBD) and other intestinal inflammation (Rathinam and Chan, 2018). Crohn’s disease (CD) and ulcerative colitis (UC) as two major types of IBD, both of which show defects of the epithelial barrier, accompanied with goblet cell decrease in UC and increase in CD (McCauley and Guasch, 2015). Various immune cells have been implicated in the pathogenesis of IBD, including macrophages (Chikina et al., 2020; Steinbach and Plevy, 2014), T cells (Lutter et al., 2018), innate lymphoid cells (Vivier et al., 2018), dendritic cells (Steinbach and Plevy, 2014), and plasma cells (Buckner et al., 2014). IBD is also associated with other events, such as deregulation of BMP and Wnt signaling (Kinchen et al., 2018), tumor necrosis factor (TNF) enrichment (Gaujoux et al., 2019), extracellular matrix remodeling (Zhang et al., 2020b), and genome instability (Wang et al., 2020a). Although the mechanisms underlying enteritis pathogenesis are partly uncovered, more comprehensive analyses are needed. In this regard, scRNA-seq provides a new insight into defining the disease-associated cell states and their possible interactions.
The altered immune response in enteritis development
The intestinal immune system undergoes dramatic alterations during IBD development and is perceived as a key for effective treatment (Gorreja et al., 2022). Several unique T cell subsets were found by scRNA-seq from two severe CD cases, including NKp30+ γδT cells expressing RORγt and IL-26 (Jaeger et al., 2021). This study also revealed the increase of activated CD8+ T cells and the decrease of Treg cells in the inflamed regions (Fig. 3). Although the accumulation of CD8+ T cells was confirmed by another scRNA-seq analysis of immune cell populations in checkpoint inhibitor-induced colitis (CPI), the percentage of FOXP3+ Treg cells was found to be significantly elevated in CPI patients (Luoma et al., 2020), and the increase of special FOXP3/BATF+ Treg cells and IL1B/LYZ+ myeloid cells were confirmed by another study in inflamed UC patients (Devlin et al., 2021). Along with CD8+ T cells, CD4+ T cells were increased in CD patients, accompanied by an increased expression of the chemokines and cytokines, such as CXCL2, CXCL10, CXCL13, CCL11, and IL-6 (Elmentaite et al., 2020). Furthermore, the disease-specific patterns and metabolic changes were observed in CD4+ effector T cells according to the scRNA-seq analysis of 6 CD and 6 UC donors, including the increase of Toll-like receptor (TCR), Janus kinase (JAK), IL-17A, IL-22, TNF, et al. (Huang et al., 2021). Interestingly, CD39+ intraepithelial T cells are decreased in the pediatric IBD group, which may exacerbate colonic inflammation via platelet aggregation and 5-hydroxytryptamine (5-HT) release (Huang et al., 2019). These results confirmed the accumulation of activated CD8+ T cells during enteritis development, while the change of Treg cells was still unclear.
Fig. 3.
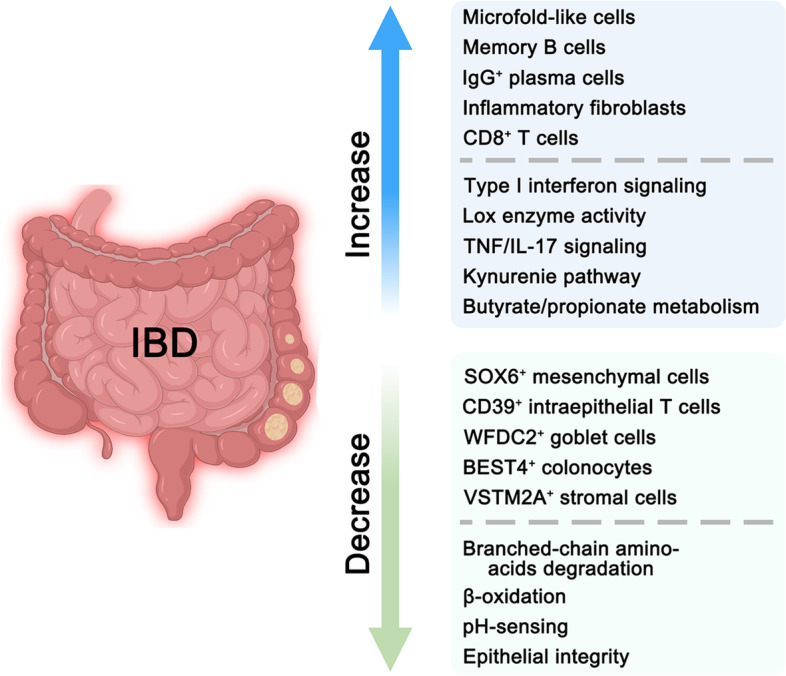
The alteration of cell types, signaling, and metabolism pathways in IBD. Some cell types are increased in IBD, such as microfold-like cells, memory B cells, IgG+ plasma cells, inflammatory fibroblasts, and CD8 + T cells, while SOX6 + mesenchymal cells, CD39 + intraepithelial T cells, WFDC2 + goblet cells, BEST4 + enterocytes, and VSTM2A + stromal cell are decreased in IBD. Similarly, type I interferon signaling, Lox enzyme activity, TNF/IL-17 signaling, et al. are increased in IBD, while β-oxidation, pH-sensing, epithelial integrity, et al. are decreased in IBD
For other immune cell types, the expansion of IgG+ plasma cells, myeloid cells, and memory B cells were observed in the pediatric IBD group (Huang et al., 2019), and the increased IgG+ plasma cells and myeloid cells were also verified in CD patients (Elmentaite et al., 2020). Pathogenic expansion of IgG+ plasma cells and naïve B cells was also confirmed by scRNA-seq of human UC samples, which may be regulated by intestinal CXCL13-expressing follicular helper (TFH)-like T peripheral helper cells (Uzzan et al., 2022). Additionally, a shift from plasma cells to follicular cells and a decrease in the frequencies of IgA+ relative to IgG+ plasma cells were observed in the scRNA-seq survey of the 366,650 stromal cells from 18 UC patients and 12 healthy individuals (Smillie et al., 2019). Moreover, enrichment of mononuclear phagocytes was also observed in both infliximab- and vedolizumab-treated non-responder samples from IBD patients by scRNA-seq analysis, which was validated in the dextran sulphate sodium (DSS)-induced colitis mouse model (Liu et al., 2019). Therefore, most immune cells are increased in enteritis, especially for IgG+ plasma cells and myeloid cells, but other cell types, such as naïve B cells and mononuclear phagocytes, need further investigation. These studies illustrate the detailed change in the intestinal immune system during enteritis development.
The association between environmental signaling and intestinal enteritis
Environment signaling also plays an important role in the pathogenesis of inflammation. Kinchen and colleagues surveyed the colonic mesenchymal atlas of health and IBD samples by scRNA-seq. They found that SOX6+ mesenchymal cells, which express TGF-β superfamily ligands BMP2 and BMP5, the non-canonical Wnt ligands WNT5A and WNT5B, and the Wnt antagonist FRZB, were decreased in inflamed UC colonic tissues (Kinchen et al., 2018). Similarly, PDGFRA, BMPs, WNT5A, SOX6, and matrix genes (ADAMDEC1 and GSN) in the crypt top fibroblasts were decreased in human IBD tissues and mouse DSS model (Elmentaite et al., 2020; Kinchen et al., 2018). Another study showed that MAP3K2-regulated intestinal stromal cells are the source of R-spondin 1 following intestinal injury, which can protect the intestine from DSS-induced colitis in mice (Wu et al., 2021). These results illustrate the importance of the microenvironment remodeling in intestinal enteritis.
A common therapeutic target for IBD is TNF, which is usually upregulated during enteritis development (Kinchen et al., 2018). Integration of scRNA-seq and proteomics revealed that activation of integrin signaling is associated with anti-TNF therapy resistance in IBD (Brubaker et al., 2020). Enrichments of IgG plasma cells, inflammatory mononuclear phagocytes, and activated T and stromal cells are also associated with resistance to anti-TNF therapy according to the scRNA-seq analysis of human ileal CD samples (Martin et al., 2019). ScRNA-seq of NOD2-driven CD in zebrafish suggests that gp130 blockade could be used to complement anti-TNF therapy (Nayar et al., 2021). Therefore, upregulated TNF signaling, reduced activities of BMPs, and non-canonical Wnts may be associated with intestinal enteritis, which may guide therapy.
Epithelial barrier and metabolism changes during enteritis development
The intestinal epithelium establishes the fundamental barrier for microbiota defense, intestinal luminal stress and mucosal immunity, and dysfunction of the epithelial barrier contributes to enteritis development. ScRNA-seq analysis of human colonic epithelia in health and clinically inflamed and noninflamed UC mucosa identified a novel pH-sensing absorptive colonocyte population marked by BEST4/OTOP2 (Parikh et al., 2019), and its reduction was associated with IBD development (Fig. 3). A disease-associated cluster of goblet cells was also observed to highly express WFDC2, KLF2, LAMC2, LAMB3, PLEC, and F3, and these genes may play an important role in the integrity maintenance of the epithelial barrier (Parikh et al., 2019). In addition, other genes, which may contribute to pediatric IBD pathogenesis by causing impaired epithelial integrity and stress responses, were reported to be enriched in IBD epithelial cells, including ERN1, PSMA6, DVL1, CASP7, and PIEZO1 (Huang et al., 2019).
Niche cells also contribute to the integrity maintenance of the intestinal epithelium. A stromal subset marked by VSTM2A, SOX6, and AGT was functionally related to the epithelial basement membrane at the physiological condition, and its deregulation contributes to epithelial barrier breakdown during colitis (Fig. 3) (Kinchen et al., 2018). Similarly, inflammation-driven fibroblasts could regulate mucosal matrix remodeling and healing in mouse DSS-induced colitis by producing IL-11 and the metalloprotease Adamdec1 (Jasso et al., 2022). The enrichment of fibroblasts and endothelial cells that participate in matrix remodeling and type I interferon signaling was also revealed by the single-cell transcriptomic analysis from the children samples with undifferentiated colitis, CD, and UC (Huang et al., 2019).
Interestingly, enteritis-associated inflammation also leads to metabolic changes in epithelial cells, such as induction of the kynurenine pathway and arginine biosynthesis enzymes, increased metabolism of butyrate and propionate, impaired degradation of branched-chain amino acids, and downregulation of β-oxidation (Fig. 3) (Smillie et al., 2019). The blockade of Lox enzyme activity in Lox/Loxl1+ mesenchymal cells could attenuate DSS colitis and reduce circulating markers of oxidative stress by reducing hydrogen peroxide (Kinchen et al., 2018). In addition, other changes may be pathogenic, such as 5-HT over-secretion (Huang et al., 2019), integrin signaling activation (Brubaker et al., 2020), and redox imbalances (Kinchen et al., 2018). Although the survey at the single-cell level surely advances our understanding of IBD development, the causal relationship between the dysfunction of the epithelial barrier, metabolism changes, and intestinal enteritis needs experimental verification.
Mice as a model for human enteritis
DSS treatment or irradiation of the mouse intestine is often used as enteritis models (Zhu et al., 2021b). Assessment of the difference and similarity between mouse and human enteritis is important for the proper application of mouse models. ScRNA-seq analysis of colon and colitis showed that embryonic-specific genes reappear in response to intestinal damage, which is conserved both in human IBD and mouse DSS-induced colitis, including MYO15B, S100A11, and CDV3 (Fazilaty et al., 2021). ScRNA-seq combined with spatial transcriptomic analyses also defined a subset of human IBD-risk genes that occur in the mouse DSS-treated colitis, such as the genes related to immune cell recruitment (e.g., Itgal, Icam1, Itga4), activation (e.g., Cd6, Plcg2, Ncf4, Il10ra), and antigen presentation (e.g., Tap1, Tap2, Psmb8) (Parigi et al., 2022).
However, different gene expression profiles were reported between human colitis and mouse model. For instance, Serpina1c (or PI3 in humans) was only increased in mice after intestinal damage (Fazilaty et al., 2021). ScRNA-seq analysis also revealed that a new skin-like epithelial population called squamous neo-epithelium marked by the expression of Sox2/Krt14/Krt7 could confer the resistance to colitis injury and rebuild the epithelial structure after colitis (Liu et al., 2022). This specialized population is only identified in the rectum of the DSS-treatment mouse model and need to be confirmed in human. These observations indicate that mouse colitis models should be cautiously used for human enteritis although some pathogenetic mechanisms are conserved.
Single-cell technologies reveal novel insights into gut cancer development
Deciphering tumor heterogeneity is a key to get a comprehensive understanding of tumor origination and to find better and more precise treatments. Our current knowledge of intra-tumoral heterogeneity is largely gained from the analysis of bulk tumor specimens, including bulk DNA/RNA sequencing. However, most bulk tumor specimens consist of a mixture of nonmalignant cells and diverse subpopulations of cancer cells. To gain more insight into intra-tumoral heterogeneity, single cancer cell clones have been generated (Roerink et al., 2018). But these clones still cannot cover all the cancer cells with distinct behavior. In this regard, single-cell technology exhibits its advantage in dissecting the multiple dimensions of intra-tumoral heterogeneity and their evolutionary relations (Francis et al., 2014; Navin et al., 2011). Combining single-cell DNA sequencing (scDNA-seq) and scRNA-seq, more details can be obtained at the single-cell resolution, such as tumor cell heterogeneity (Darmanis et al., 2017), epithelial-to-mesenchymal transition (EMT) (Pastushenko et al., 2018), cancer metastasis (Puram et al., 2017), and tumor microenvironment (Neal et al., 2018).
Transcriptomic heterogeneity of CRC
CRC development is influenced by numerous factors whose combinatory effects lead to various tumor subtypes and cellular heterogeneity (Dekker et al., 2019). ScRNA-seq has been used to profile various tumors and has revealed the cell constitution, tumor-initiating cells, and the microenvironment cells. For instance, it has been reported that stem-like cells accounted for 93% of tumor epithelial cells in CRC, but only 30% in normal mucosa epithelium, according to a new clustering method named reference component analysis (Li et al., 2017). This study also grouped colorectal tumors into subtypes with proliferating and differentiated states, which were previously assigned to a single cell type by bulk transcriptomics (Li et al., 2017; Wu et al., 2017). The distinct cell subpopulations resembling differentiation states of normal intestinal epithelial cells (ISCs-like, TA-like, and differentiated cells) were also revealed by other scRNA-seq analyses in CRC patients (Wu et al., 2017; Zowada et al., 2021). These ISCs-like and TA-like cells showed high levels of oxidative phosphorylation and mitochondrial membrane potential, which were linked to tumor-initiating activity (Zowada et al., 2021).
ScRNA-seq has also been employed to investigate the epithelial-mesenchymal interaction in CRC pathogenesis. Analysis of cancer stem cells in CRC in ApcMin/+ mouse adenoma at various time points during chemoradiotherapy suggests that tumor-initiating cells can shape the tumor environment into a landscape, which renders resistance to immunosuppression by forming an immune barrier against CD8+ T cells and promotes the proliferation of therapy-resistant cancer stem cells via Cox-2/PGE2 signaling (He et al., 2021).. Moreover, cancer stem cell activity is also regulated by the niche BMP and WNT signaling secreted by the PDGFRAlow/CD81+ stromal cells (Yum et al., 2021). The oncogene reporter mouse model combined with scRNA-seq analysis suggests that oncogene-expressing crypts can replace neighboring wild-type crypts over time, thereby leading to accelerated clonal drift and CRC development (Yum et al., 2021). In summary, these results implicate that the transformed epithelial cells can accelerate the tumorigenesis process themselves and cultivate the environment to facilitate tumor development.
The role of the immune system in CRC therapeutic outcomes
Apart from promoting tumor transformation of the intestinal epithelial cells, the immune cells also play an important role in CRC therapeutic outcomes and influence tumor metastases. ScRNA-seq transcriptomic profiling of T cells from peripheral blood, adjacent normal and tumor tissues of CRC patients showed that the CXCL13+/BHLHE40+ subset of IFNG+ Th1-like T cells was preferentially enriched in microsatellite instability tumors (Zhang et al., 2019b). Treatment of a CD40-activating antibody could expand Th1-like and CD8+ memory T cells in colon cancer (Zhang et al., 2020a). Furthermore, the scRNA-seq analysis of 37,931 T cells from 16 CRC patients suggests the phenotypically and functionally distinguishable CD4+ and CD8+ effector T cell types are associated with clinical outcomes (Masuda et al., 2022). The GZMK+/KLRG1+ cytotoxic CD8+ T cells with a less dysfunctional phenotype were enriched in CRC patients with good outcomes. For CD4+ T-cell infiltrates, Helios+ Treg cells were associated with good outcomes, while Helios−/CD38+ peripherally-induced Treg cells were strongly associated with bad outcomes. These immune cells may be good targets for advanced treatments and the prognosis of CRC.
It has been reported that specific macrophage and conventional dendritic cell subsets may be key factors in the development of colorectal adenoma and liver metastases, according to the scRNA-seq of 18 treatment-naive CRC patients (Zhang et al., 2020a). ScRNA-seq and spatial transcriptomic analysis further revealed that immune-suppressive cells, especially MRC1+/CCL18+ M2-like macrophages, were enriched in live metastases of CRC (Wu et al., 2022). Single-cell transcriptomic profiling of lung metastases from CRC patients uncovered that a special subtype of B cells (ERBIN+) was involved in cancer metastases, and targeting Erbin as well as the combinatory block of B cells and PD1 could suppress lung metastasis of CRC in mice (Shen et al., 2021). Therefore, special T cells may be associated with CRC development, and macrophages may play an important role in CRC metastases. These scRNA-seq results present a more comprehensive understanding of the immune roles in tumor development and metastasis and provide new insights to improve immune therapies.
Single-cell DNA-seq confirms tumor heterogeneity
In addition to scRNA-seq, scDNA-seq has also exhibited its power in the understanding of cellular heterogeneity of CRC (Jiang et al., 2021; Navin et al., 2011). An early study of 63 single tumor cells identified the SLC12A5 gene with a high frequency of mutation at the single-cell level but exhibited low prevalence at the bulk population level (Yu et al., 2014). By far, single-cell whole-exome sequencing of CRC has discovered many important gene mutations. One unique sub-clonal mutation of CSMD1 was detected exclusively in the adenomatous polyps by scDNA-seq, but not by bulk sequencing (Wu et al., 2017). A similar scDNA-seq analysis also revealed mutations and somatic copy number alterations hidden in bulk sequencing (Liu et al., 2017). ScDNA-seq also revealed that EpCAM/CD44 colonic cancer stem cells are critical for cancer development, metastasis, and drug resistance (Du et al., 2008; Went et al., 2004). Single-cell whole-genome sequencing of EpCAMhigh/CD44+ cancer stem cells and EpCAMhigh/CD44− differentiated cancer cells in colon cancer indicated that those cells from the same patient had a similar somatic copy number variation pattern. However, the somatic copy number variation was found to be different in certain patients and might confer different growth advantages at later stages (Liu et al., 2018). Moreover, scDNA-seq of the primary tumors and liver metastases of CRC unraveled the extensive intra-tumoral heterogeneity at the primary and metastatic tumor sites, as well as the late dissemination of metastases from early progenitor clones (Leung et al., 2017).
Multiple techniques were employed to simultaneously assess somatic copy number variation, DNA methylation, and transcriptome of 1,900 single cells from 10 CRC patients at the single-cell level (Bian et al., 2018). The results uncovered that the genome-wide DNA demethylation patterns of cancer cells were consistent in all 10 patients, while the DNA demethylation degrees were correlated with the density of the heterochromatin-associated H3K9me3 of normal tissues (Bian et al., 2018). Also, combined analyses of scDNA-seq and scRNA-seq on tumor niche cells from 21 colorectal patients showed that somatic copy number alterations were prevalent in immune cells, fibroblasts, and endothelial cells in tumors compared with adjacent tissues (Zhou et al., 2020). Furthermore, five genes (BGN, RCN3, TAGLN, MYL9, and TPM2) were identified as fibroblast-specific biomarkers for the poor prognosis of CRC. ScDNA-seq and scRNA-seq were also applied to the organoids, which were derived from single normal intestinal cells, subjected to RNAi-mediated downregulation of the tumor suppressor gene APC, and then transplanted to mice to investigate how intra-tumoral heterogeneity is generated. The results showed that the emergence of new transcriptional subpopulations was the key for the adaptation of cancer cells to drastic microenvironmental changes (Ono et al., 2021). All these studies clearly demonstrate that the single-cell studies provide novel insights into cancer initiation, metastases, heterogeneity, and relation to the microenvironment, and would advance disease diagnosis and treatment.
Conclusions and Perspectives
With the development of single-cell technology, we have gained a more comprehensive understanding on the intestinal cellular constitution, characterizations, and heterogeneity (Table 2). Single-cell sequencing has also yielded new insights into the pathogenesis of enteritis and intestinal cancer. Although a significant advance has been made, many questions remain. Single-cell transcriptomic analysis has unraveled new subtypes of intestinal tuft cells, enterocytes, and EECs, but the heterogeneity of other cell types and the landscape of the intestinal immune system need further investigation. Epithelial progenitors have been divided into absorptive and secretory subtypes (Clevers and Batlle, 2013), but no clear subpopulations are reported. It may be due to a vague definition of progenitors and unclear differences between progenitors and TA cells. Attention has also been paid to the differences between Lgr5high and Lgr5low subpopulations (Baulies et al., 2020), but the heterogeneity among intestinal stem cells is still unclear. Even the dispute over whether there are quiescent stem cells or + 4 stem cells is not settled. Microfold cells are indicated to be essential for antigens transport to the lymphoid cells underneath (Mabbott et al., 2013), but the atlas of microfold cells is rarely known because of their low cell number in the intestinal epithelium. The spatial heterogeneity of enterocytes and EECs has been explored along the crypt-villus axis and in different intestinal segments (Beumer et al., 2018; Wang et al., 2020b). More comprehensive investigations on spatial cell heterogeneity, pathophysiological significance, and the underlying mechanisms are needed. Also, the spatiotemporal transcriptional atlas of EECs has been reported. Similar studies should be applied to other epithelial cell types and niche cells during intestine development and regeneration, which would help address the question concerning what regulates the formation of the special structures. For instance, Hedgehog signaling has been reported to be critical for villus formation (Rao-Bhatia et al., 2020).
Table 2.
Summary of the scRNA-seq studies in the intestine
| Organism | Sample type | Sample source | Cell type | Library preparation method | Cell number | Data resource | Reference |
|---|---|---|---|---|---|---|---|
| Mus musculus | Jejunum | Labeled stem cells | Short-term and long-term label-retaining stem cells | Fluidigm | 558 | Supplementary Table 2 | (Li et al., 2016) |
| Mus musculus | Jejunum | Wildtype mice | Epithelial, mesenchymal and immune cells | MARS-seq | 329 | GSE134479 | (Bahar Halpern et al., 2020) |
| Mus musculus | Jejunum | Stem cells | Prox1-GFP+, Bmi1-GFP+, Lgr5-eGFP+, and Lgr5-eGFP− cells | 10X Chromium | 3,521 | GSE99457 | (Yan et al., 2017) |
| Mus musculus | Ileum | Massive small bowel resection | Epithelial cells | 10X Chromium | 19,245 | GSE130113 | (Seiler et al., 2019) |
| Mus musculus | Small intestine | Wildtype mice | Crypt cells | C1 | 76 | GSE146783 | (Sato et al., 2020) |
| Mus musculus | Small intestine | Labeled epithelial cells | Bmi1, Hopx or Lgr5 labeled cells | Fluidigm | 1,033 | Supplementary Table 2 | (Li et al., 2014) |
| Mus musculus | Small intestine | Labeled epithelial cells | Intestinal preproglucagon-expressing cells | Smart-seq2 | 288 | Not described | (Glass et al., 2017) |
| Mus musculus | Small intestine | Wildtype mice | Epithelial cells | 10X Chromium | 7,216 | GSE92332 | (Haber et al., 2017) |
| Mus musculus | Small intestine | Stem cells | Lgr5high cells | Smart-seq | 245 | GSE90856 | (Barriga et al., 2017) |
| Mus musculus | Small intestine | Stem cell depleted and normal epithelium | Epithelial cells | 10X Chromium | 192 | Not described | (Tetteh et al., 2016) |
| Mus musculus | Small intestine | Lgr5-eGFP+ intestinal stem cells | Stem cells | 10X Chromium | 13,247 | GSE92865 | (Yan et al., 2017) |
| Mus musculus | Small intestine | Wildtype and irradiated mice | Epithelial cells | 10X Chromium | 6,644 | GSE123516 | (Ayyaz et al., 2019) |
| Mus musculus | Small intestine | Small intestine mucositis | Epithelial cells | 10X Chromium | 12,653 | GSE131630 | (Zhao et al., 2020) |
| Mus musculus | Small intestine | Wildtype and Lats1/2 knockout mice | Pdgfrβ+ intestinal stromal cells and lymphatic endothelial cells | 10X Chromium | 7,906 | GSE124488 | (Hong et al., 2020) |
| Mus musculus | Small intestine | Normal diet and high-fat/high-sugar diet mice | Epithelial cells | 10X Chromium | 27,687 | GSE147319 | (Aliluev et al., 2021) |
| Mus musculus | Small intestine | Wildtype mice and β7-KO mice | Epithelial cells | 10X Chromium | 13,352 for WT; 10,763 for β7-KO | OEP000370 | (Chen et al., 2021a) |
| Mus musculus | Small intestine | Wildtype; FltpZV/+; Foxa2FVF/FVF mice | Epithelial cells | 10X Chromium | 60,000 | GSE152325 | (Bottcher et al., 2021) |
| Mus musculus | Small intestine | Wildtype crypts in Red2-KrasG12D and Red2-PIK3CAH1047R mice | Epithelial, mesenchymal and immune cells | 10X Chromium | 21,183 | E-MTAB-8656 | (Yum et al., 2021) |
| Mus musculus | Small intestine | Wildtype mice | Epithelial cells | MARS-seq | Not described | GSE178586 | (Zinina et al., 2022) |
| Mus musculus | Small intestine and colon | Casp3/7ΔIEC mice and Casp3/7FL/FL littermates | Epithelial cells and immune cells | 10X Chromium | 7,584 Casp3/7 fl/fl cells and 11,956 Casp3/7ΔIEC cells | GSE183885 | (Ghazavi et al., 2022) |
| Mus musculus | Distal small intestine and colon | Muc2-mCherry mice | Goblet cells | 10X Chromium | 6123 cells for the colon and 3552 cells for the small intestine | GSE144436 | (Nystrom et al., 2021) |
| Mus musculus | Colon | Wildtype mice | Gli1+ stromal cells | 10X Chromium | 4,464 | GSE113043 | (Degirmenci et al., 2018) |
| Mus musculus | Colon | LGR5+ lineage tracing and lethal injury (irradiation) mice | Regenerating crypt cells and Ascl2-deficient colonic stem cells | 10X Chromium | 3,254 | GSE130822 | (Murata et al., 2020) |
| Mus musculus | Colon | Wildtype mice | Pdgfra+ endothelial cells and epithelial cells | 10X Chromium | 6,358 | GSE130681 | (McCarthy et al., 2020c) |
| Mus musculus | Colon | NeuroD1-Cre x Rosa26-EYFP labeled mice | EECs | 10X Chromium | 1,560 | Not described | (Billing et al., 2019) |
| Mus musculus | Colon | Wildtype mice | Epithelium, endothelial and stromal cells | 10X Chromium | 7,395 | GSE151257 | (Brugger et al., 2020) |
| Mus musculus | Colon | Wildtype mice | T cells | 10X Chromium | 7,012 | GSE160055 | (Kiner et al., 2021) |
| Mus musculus | Colon | DSS-exposed 3 days | Epithelial and mesenchymal cells | 10X Chromium | 11,495 | GSE156245 | (Wu et al., 2021) |
| Mus musculus | Colon | MC38 xenografts upon SHP099 treatment | Cancer cells, stromal cells, and immune cells | 10X Chromium | 7,934(SHP099), 7,881(PBS) | GSE164908 | (Gao et al., 2022) |
| Mus musculus | Colon | Wildtype and DSS treated mice | Stromal cells | 10X Chromium | 34,197 | GSE172261 | (Jasso et al., 2022) |
| Mus musculus | Distal colon | Wildtype and DSS treated mice | Epithelial, stromal and immune cells | 10X Chromium | > 35,000 | GSE168033 | (Liu et al., 2022) |
| Mus musculus | Colorectal cancer | Primary tumor and lung metastases | Immune cells | 10X Chromium | 12,588 B220+ cells, 3,748 CD38+ cells, 1,588 CD79a+ cells | Not described | (Shen et al., 2021) |
| Mus musculus | Small intestine, colon | Wildtype mice | Endothelial cells | 10X Chromium | > 32,000 | E-MTAB-8077 | (Kalucka et al., 2020) |
| Mus musculus | Intestine | pLysDTR mice (DT-treated for 6 consecutive days) | Paneth cells | CEL-seq | 288 | Supplementary dataset S01 | (van Es et al., 2019) |
| Mus musculus | Intestine | Wildtype mice | Intestinal stromal cells | Drop-seq | 4,359 | GSE116514 | (Kim et al., 2020) |
| Mus musculus | Intestine | Endoderm | Definitive endoderm cells; trophectoderm cells; parietal endoderm cells; visceral endoderm cells; yolk sac endoderm cells | 10X Chromium | 112,217 | GSE123046 | (Nowotschin et al., 2019) |
| Mus musculus | Intestine | Embryo of wildtype mice | Epithelial and mesenchymal cells | STRT-seq | 217 | GSE87038 | (Dong et al., 2018) |
| Mus musculus | Intestine | Wildtype and irradiated mice | Epithelial cells | 10X Chromium | 2,329 | GSE145866 | (Sheng et al., 2020) |
| Mus musculus | Small intestinal organoids | Wildtype and Rfx6 conditional deletion organoids | Epithelial cells | BD™ Precise WTA Single Cell Kit | 290 | GSE133038 | (Piccand et al., 2019) |
| Mus musculus | Small intestinal organoids | Healthy and cancerous organoids | Epithelial cells | Multivariate-barcoded MC | > 1 million | https://community.cytobank.org/cytobank/login#project-id=1271 | (Qin et al., 2020) |
| Mus musculus | Organoids | Lgr5+ cell-derived organoids | EECs | CEL-seq2 | 384 | GSE114988 | (Beumer et al., 2018) |
| Mus musculus | Crypts and organoids | Wildtype mice and intestinal organoids | Epithelial cells | CEL-seq | 238 | GSE62270 | (Grun et al., 2015) |
| Mus musculus | Neurog3Chrono reporter mice and organoids | Neurog3 labeled mice | EECs | SORT-seq | 6,906 | GSE113561 | (Gehart et al., 2019) |
| Mus musculus | Organoids | Lgr5+ and Lgr5− single cells | Epithelial cells | 10X Chromium | 23,421 | GSE115956 | (Serra et al., 2019) |
| Mus musculus | Organoids upon RNAi-mediated APC and organoids transplanted into mice | APC mutated cells | Epithelial cells | C1 | 200 | https://figshare.com/articles/dataset/Single_Cell/14518056 | (Ono et al., 2021) |
| Mus musculus | Organoid | Organoids treated with forskolin, CFTRinh-172, or DMSO | Epithelial cells | inDrops | 18,303 | GSE164638 | (Tallapragada et al., 2021) |
| Mus musculus | Small intestinal organoids | 5-day differentiated BMP-off and BMP-on organoids | Epithelial cells | SORT-seq | Not described | GSE194004 | (Beumer et al., 2022) |
| Mus musculus | Transgene-inducible intestinal organoids | Induced organoids for KRAS, BRAF and CTNNB1 | Epithelial cells | BD™ Precise WTA Single Cell Kit | 160,000 | GSE115242 | (Brandt et al., 2019) |
| Mus musculus | Small intestine and organoids | Wildtype mice and organoids | Epithelial cells | 10X Chromium | 10,180 | GSE100274 | (Mead et al., 2018) |
| Mus musculus | Intestinal adenoma of ApcMin/+mice at various time points | Colorectal cancer | Epithelial and mesenchymal cells | 10X Chromium | 79,801 | GSE136256 | (Zowada et al., 2021) |
| Homo sapiens | Ileum | CD tissues | Lamina propria cells | 10X Chromium | 82,417 | GSE134809 | (Martin et al., 2019) |
| Homo sapiens | Ileum | Health and CD tissues | Immune cells | 10X Chromium | 16,731 | GSE157477 | (Jaeger et al., 2021) |
| Homo sapiens | Esophagus, stomach, duodenum | Healthy tissues | Epithelium; immune cells | SORT-seq | Not described | GSE157694/EGAS00001004695 | (Busslinger et al., 2021) |
| Homo sapiens | Small intestine | Donor- and recipient-derived cells after transplantation | T cells | 10X Chromium | 974 for scRNA-seq,196 for smart-seq | GSE162687 | (FitzPatrick et al., 2021) |
| Homo sapiens | Small intestine | Healthy tissues | Epithelial cells | 10X Chromium | 12,590 | GSE185224 | (Burclaff et al., 2022) |
| Homo sapiens | Ileum and ileum-derived organoids | Healthy and viral infected tissues | Epithelial cells | 10X Chromium | 25,482 | GSE171620 | (Triana et al., 2021) |
| Homo sapiens | Colon | Healthy and IBD tissues | Epithelial and mesenchymal cells | Smart-seq2 | 11,175 | GSE116222 | (Parikh et al., 2019) |
| Homo sapiens | Colon | Healthy and UC tissues | Epithelial cells & mesenchymal cells & immune cells | 10X Chromium | 366,650 | SCP259 | (Smillie et al., 2019) |
| Homo sapiens | Colon | Colorectal cancer | Cancer cells, stromal cells, and immune cells | 10X Chromium | 27,927 | GSE188711 | (Guo et al., 2022) |
| Homo sapiens | Colon | Colorectal cancer | Immune cells | 10X Chromium | 178,630 | OEP001756 | (Wu et al., 2022) |
| Homo sapiens | Colon | Tumors and adjacent tissues | T cells | 10X Chromium | 37,931 | EMTAB-9455 | (Masuda et al., 2022) |
| Homo sapiens | Colon | Healthy and UC tissues | Immune cells, stromal cells | 10X Chromium | 29,046 | GSE182270 | (Uzzan et al., 2022) |
| Homo sapiens | Colon | Healthy tissues | Macrophages | 10X Chromium | 63,970 | EGAD00001007765/EGAS00001005377 | (Domanska et al., 2022) |
| Homo sapiens | Colon and rectum | Familial adenomatous polyposis | Environmental cells (endothelial cells, fibroblasts, macrophages, mast cells, T cells and B cells) and epithelial cells | STRT-Seq (modified) | 8,757 | Not described | (Li et al., 2020) |
| Homo sapiens | Large intestine | Children with undifferentiated colitis, Crohn’s disease, and ulcerative colitis | Epithelium, mesenchyme and immune cells | 10X Chromium | 73,165 | GSE121380 | (Huang et al., 2019) |
| Homo sapiens | Large intestine | Colorectal cancer | Immune and non-immune cells | 10X Chromium and Smart-seq2 | 43,817 (Hi-seq 4000); 10,468 (Smart-seq2) | GSE146771 | (Zhang et al., 2020a) |
| Homo sapiens | Large intestine | Healthy; melanoma with Checkpoint inhibitor-induced colitis; melanoma without Checkpoint inhibitor-induced colitis | Immune cells | 10X Chromium | 51,652 | GSE144469 | (Luoma et al., 2020) |
| Homo sapiens | Large intestine | Healthy, CD and UC tissues | CD45+ immune cell | 10X Chromium | 63,314 | PRJCA003980 | (Huang et al., 2021) |
| Homo sapiens | Large intestine | Healthy and UC tissues | Immune cells | 10X Chromium | 20,678(UC) 16,678 (Health) | GSE162335 | (Devlin et al., 2021) |
| Homo sapiens | Ileum, colon, rectum | Precancerous tissues | Epithelial cells | 10X Chromium | 14,537 | GSE125970 | (Wang et al., 2020b) |
| Homo sapiens | Small intestine, large intestine | Fetal digestive tissues | Epithelial and mesenchymal cells | STRT-seq | 5,227 | GSE103239 | (Gao et al., 2018) |
| Homo sapiens | Embryonic gastrointestinal tract | Healthy tissues | Epithelial cells | Not described | 5,290 | Supplementary Table 1 and 2 | (Tan et al., 2019) |
| Homo sapiens | Human embryonic intestinal tract | Healthy tissues | Epithelial, mesenchymal and immune cells | 10X Chromium | 24,783 | E-MTAB-9489 | (Holloway et al., 2021) |
| Homo sapiens | Human embryonic and adult intestinal tract | Healthy tissues | Epithelial, mesenchymal and immune cells | 10X Chromium | 428,000 | E-MTAB-9543, E-MTAB-9536, E-MTAB-9532, E-MTAB-9533 and E-MTAB-10386 | (Elmentaite et al., 2021) |
| Homo sapiens | Small intestinal organoids | Crohn’s disease patient-derived tissues | Epithelial cells | C1 | 1,037 | Not described | (Suzuki et al., 2018) |
| Homo sapiens | Human pluripotent stem cell-derived intestinal organoids | Organoids | Epithelium, mesenchyme, endothelium, and neurons | 10X Chromium | 13,289 | E-MTAB-9228 | (Holloway et al., 2020) |
| Homo sapiens | Intestine and organoids | Intestinal tissues and organoids | EECs | 10X Chromium | 15,283 | GSE146799 | (Beumer et al., 2020) |
| Homo sapiens | Cell lines | Colorectal cancer | SW480 cells | 10X Chromium & SORT-seq | 192 | Not described | (Yi et al., 2020) |
| Homo sapiens | Cell lines | Control and 5FU-treated colon cancer | 5FU-treated RKO, HCT116, SW480 | Drop-seq | 10,421 | GSE149224 | (Park et al., 2020) |
| Homo sapiens | Large intestine and cell lines | Healthy tissues and colorectal cancer | Epithelial cells | C1 | 2,221 | GSE81861 | (Wu et al., 2017) |
| Homo sapiens | Primary and metastasis tumor | Colorectal cancer | Spheroids, Tumors, PDXs, PDOs | 10X Chromium and iCELL8 | 26,170 | EGAS00001004064 | (Zowada et al., 2021) |
| Homo sapiens | Tumors and adjacent normal tissues | Colorectal cancer | T cells | Smart-seq2 | 8,530 | EGAS00001002791 / GSE108989 | (Zhang et al., 2018) |
| Homo sapiens | Embryonic, fetal, childhood/adolescence ileum | Healthy and pediatric Crohn’s disease tissues | Epithelial, mesenchymal and immune cells | 10X Chromium | 74,106 | E-MTAB-8901 | (Elmentaite et al., 2020) |
| Homo sapiens | Human embryonic intestinal tract | Healthy tissues | Epithelial, mesenchymal and immune cells | 10X Chromium | 76,592 | GSE158328/GSE158702 | (Fawkner-Corbett et al., 2021) |
| Mus musculus; Homo sapiens | Intestine and organoids | Colonic mesenchyme; fibroblast-crypt cocultures; Ptger4-knockout crypt epithelial cells | Epithelial cells and mesenchymal cells | Drop-seq | 5,371 | GSE142431 | (Roulis et al., 2020) |
| Mus musculus; Homo sapiens | Colon | Healthy, UC and DSS treated tissues | Epithelial and mesenchymal cells | 10X Chromium | 11,549 | GSE114374 | (Kinchen et al., 2018) |
| Mus musculus; Homo sapiens | Human embryonic intestinal tract and mouse colon and colitis | Healthy and DSS treated tissues | Epithelial, mesenchymal and immune cells | 10X Chromium | 22,579 | GSE151257/GSE154007 | (Fazilaty et al., 2021) |
| Mus musculus; Homo sapiens | Intestinal organoids | Organoids | Epithelial cells | DisCo | 945 | GSE148093 | (Bues et al., 2022) |
| Rhesus macaques | Large intestine | Healthy and during acute graft-versus-host disease tissues | T cells | 10X Chromium | 21,490 | GSE142483 | (Tkachev et al., 2021) |
| Zebrafish | Whole intestine | Healthy and DSS treated tissues | Epithelial, mesenchymal and immune cells | 10X Chromium | 30,069 | https://github.com/Cho-lab-Sinai/Scripts_Nayar_et_al | (Nayar et al., 2021) |
| Swine | Ileum | 6 time points in the swine neonatal period | Epithelial cells | 10X Chromium | 40,186 | GSE162287 | (Meng et al., 2021) |
| Drosophila melanogaster | Midgut | Healthy tissues | EECs | 10X Chromium | 4,661 | GSE132274 | (Guo et al., 2019) |
Nowadays, we can perform single-cell sequencing analysis with millions of cells (Qin et al., 2020), but the sequencing depth in each cell is limited in the massive scRNA-seq. On another side, some sequencing technologies, such as Smart-seq2 (Picelli et al., 2013), shows a great sequencing depth, but can only apply to small cell number due to the cost. If the depth sequencing can be combined with massive single-cell numbers cost-effectively, more information will be gained (Lai et al., 2020). Furthermore, other single-cell technologies are under development, such as single-cell protein mass spectrum, single-cell sequencing of whole epigenomics, and combinatorial DNA/RNA/epigenetic sequencing in one cell (Bian et al., 2018; Hutchins, 2021), the low-input but high-resolution analysis scRNA-seq methods (Bues et al., 2022). If the breakthroughs of these technologies on the scale, sensitivity, and cost-efficiency can be made, biomedical research will enter the single-cell era.
Acknowledgements
We thank Mengxian Zhang, Huidong Liu, and Ruoyu Lou for critical reading of the manuscript.
Abbreviations
- 5-HT
5-Hydroxytryptamine
- BMP
Bone morphogenetic protein
- CCK
Cholecystokinin
- CD
Crohn’s disease
- ChgA
Chromogranin-A
- CPI
Checkpoint inhibitor-induced colitis
- CRC
Colorectal cancer
- DISCO
Digital microfluidic isolation of single cells for-omics
- DSS
Dextran sulphate sodium
- EECs
Enteroendocrine cells
- EGF
Epidermal growth factor
- EMT
Epithelial-to-mesenchymal transition
- FACS
Fluorescence activated cell sorter
- Foxl1
Forkhead box L1
- GAST
Gastrin
- GCG
Glucagon
- GHRL
Ghrelin
- GIP
Glucose-dependent insulinotropic polypeptide
- HIST
Histamine
- IBD
Inflammatory bowel disease
- icGCs
Intercrypt goblet cells
- ISCs
Intestinal stem cells
- JAK
Janus kinase
- LCM-RNA-seq
RNA-sequencing of laser capture micro-dissected tissue
- MARS-seq
Massively parallel single-cell RNA-sequencing
- MLN
Motilin
- NTS
Neurotensin
- PSMA6
Proteasome 20S subunit alpha 6
- PYY
Peptide YY
- scDNA-seq
Single-cell DNA sequencing
- scRNA-seq
Single-cell RNA sequencing
- SCT
Secretin
- SORT-seq
FACS sorting followed by single-cell RNA-sequencing
- SST
Somatostatin
- STRT-seq
Single-cell tagged reverse transcription sequencing
- TA cells
Transient-amplifying cells
- TAC1
Tachykinin 1
- TCR
Toll-like receptor
- Th2 cells
Type 2 helper T cells
- TNF
Tumor necrosis factor
- UC
Ulcerative colitis
Authors’ contributions
YW and YGC wrote the manuscript; WS integrated the reported single-cell data. SY and YL helped with the manuscript writing. The author(s) read and approved the final manuscript.
Funding
This work was supported by grants from the National Natural Science Foundation of China (31988101 and 31900550), the Guangdong Postdoctoral Research Foundation (O0390302), Guangdong Basic and Applied Basic Research Foundation (2021A1515111215) and China Postdoctoral Science Foundation (2021M703230 and 2022T150653). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Availability of data and materials
Not applicable.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing financial interests. YGC is the Editor-in-Chief of Cell Regeneration. He was not involved in the review or decision related to this manuscript.
Footnotes
Yalong Wang and Wanlu Song contributed equally to this paper.
References
- Abraham C, Cho JH. Inflammatory bowel disease. N Engl J Med. 2009;361:2066–2078. doi: 10.1056/NEJMra0804647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aliluev A, Tritschler S, Sterr M, Oppenlander L, Hinterdobler J, Greisle T, Irmler M, Beckers J, Sun N, Walch A, et al. Diet-induced alteration of intestinal stem cell function underlies obesity and prediabetes in mice. Nat Metab. 2021;3:1202–1216. doi: 10.1038/s42255-021-00458-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ayyaz A, Kumar S, Sangiorgi B, Ghoshal B, Gosio J, Ouladan S, Fink M, Barutcu S, Trcka D, Shen J, et al. Single-cell transcriptomes of the regenerating intestine reveal a revival stem cell. Nature. 2019;569:121–125. doi: 10.1038/s41586-019-1154-y. [DOI] [PubMed] [Google Scholar]
- Bahar Halpern K, Massalha H, Zwick RK, Moor AE, Castillo-Azofeifa D, Rozenberg M, Farack L, Egozi A, Miller DR, Averbukh I, et al. Lgr5+ telocytes are a signaling source at the intestinal villus tip. Nat Commun. 2020;11:1936. doi: 10.1038/s41467-020-15714-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barker N, van Es JH, Kuipers J, Kujala P, van den Born M, Cozijnsen M, Haegebarth A, Korving J, Begthel H, Peters PJ, et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature. 2007;449:1003–1007. doi: 10.1038/nature06196. [DOI] [PubMed] [Google Scholar]
- Barriga FM, Montagni E, Mana M, Mendez-Lago M, Hernando-Momblona X, Sevillano M, Guillaumet-Adkins A, Rodriguez-Esteban G, Buczacki SJA, Gut M, et al. Mex3a Marks a Slowly Dividing Subpopulation of Lgr5+ Intestinal Stem Cells. Cell Stem Cell. 2017;20(801–816):e807. doi: 10.1016/j.stem.2017.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baulies A, Angelis N, Foglizzo V, Danielsen ET, Patel H, Novellasdemunt L, Kucharska A, Carvalho J, Nye E, De Coppi P, et al. The Transcription Co-Repressors MTG8 and MTG16 Regulate Exit of Intestinal Stem Cells From Their Niche and Differentiation Into Enterocyte vs Secretory Lineages. Gastroenterology. 2020;159(1328–1341):e1323. doi: 10.1053/j.gastro.2020.06.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beumer J, Artegiani B, Post Y, Reimann F, Gribble F, Nguyen TN, Zeng H, Van den Born M, Van Es JH, Clevers H. Enteroendocrine cells switch hormone expression along the crypt-to-villus BMP signalling gradient. Nat Cell Biol. 2018;20:909–916. doi: 10.1038/s41556-018-0143-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beumer J, Puschhof J, Bauza-Martinez J, Martinez-Silgado A, Elmentaite R, James KR, Ross A, Hendriks D, Artegiani B, Busslinger GA, et al. High-Resolution mRNA and Secretome Atlas of Human Enteroendocrine Cells. Cell. 2020;182:1062–1064. doi: 10.1016/j.cell.2020.08.005. [DOI] [PubMed] [Google Scholar]
- Beumer J, Puschhof J, Yengej FY, Zhao L, Martinez-Silgado A, Blotenburg M, Begthel H, Boot C, van Oudenaarden A, Chen YG, et al. BMP gradient along the intestinal villus axis controls zonated enterocyte and goblet cell states. Cell Rep. 2022;38:110438. doi: 10.1016/j.celrep.2022.110438. [DOI] [PubMed] [Google Scholar]
- Bhat S, Sharma P, Cameron NR, Bissett IP, O'Grady G. Chyme Reinfusion for Small Bowel Double Enterostomies and Enteroatmospheric Fistulas in Adult Patients: A Systematic Review. Nutr Clin Pract. 2020;35:254–264. doi: 10.1002/ncp.10417. [DOI] [PubMed] [Google Scholar]
- Bian S, Hou Y, Zhou X, Li X, Yong J, Wang Y, Wang W, Yan J, Hu B, Guo H, et al. Single-cell multiomics sequencing and analyses of human colorectal cancer. Science. 2018;362:1060–1063. doi: 10.1126/science.aao3791. [DOI] [PubMed] [Google Scholar]
- Billing LJ, Larraufie P, Lewis J, Leiter A, Li J, Lam B, Yeo GSH, Goldspink DA, Kay RG, Gribble FM, et al. Single cell transcriptomic profiling of large intestinal enteroendocrine cells in mice - Identification of selective stimuli for insulin-like peptide-5 and glucagon-like peptide-1 co-expressing cells. Molecular Metabolism. 2019;29:158–169. doi: 10.1016/j.molmet.2019.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Birchenough GM, Nystrom EE, Johansson ME, Hansson GC. A sentinel goblet cell guards the colonic crypt by triggering Nlrp6-dependent Muc2 secretion. Science. 2016;352:1535–1542. doi: 10.1126/science.aaf7419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biton M, Haber AL, Rogel N, Burgin G, Beyaz S, Schnell A, Ashenberg O, Su CW, Smillie C, Shekhar K, et al. T Helper Cell Cytokines Modulate Intestinal Stem Cell Renewal and Differentiation. Cell. 2018;175(1307–1320):e1322. doi: 10.1016/j.cell.2018.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bottcher A, Buttner M, Tritschler S, Sterr M, Aliluev A, Oppenlander L, Burtscher I, Sass S, Irmler M, Beckers J, et al. Non-canonical Wnt/PCP signalling regulates intestinal stem cell lineage priming towards enteroendocrine and Paneth cell fates. Nat Cell Biol. 2021;23:23–31. doi: 10.1038/s41556-020-00617-2. [DOI] [PubMed] [Google Scholar]
- Boulange CL, Neves AL, Chilloux J, Nicholson JK, Dumas ME. Impact of the gut microbiota on inflammation, obesity, and metabolic disease. Genome Med. 2016;8:42. doi: 10.1186/s13073-016-0303-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brandt R, Sell T, Luthen M, Uhlitz F, Klinger B, Riemer P, Giesecke-Thiel C, Schulze S, El-Shimy IA, Kunkel D, et al. Cell type-dependent differential activation of ERK by oncogenic KRAS in colon cancer and intestinal epithelium. Nat Commun. 2019;10:2919. doi: 10.1038/s41467-019-10954-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brenna O, Bruland T, Furnes MW, Granlund A, Drozdov I, Emgard J, Bronstad G, Kidd M, Sandvik AK, Gustafsson BI. The guanylate cyclase-C signaling pathway is down-regulated in inflammatory bowel disease. Scand J Gastroenterol. 2015;50:1241–1252. doi: 10.3109/00365521.2015.1038849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brugger MD, Valenta T, Fazilaty H, Hausmann G, Basler K. Distinct populations of crypt-associated fibroblasts act as signaling hubs to control colon homeostasis. PLoS Biol. 2020;18:e3001032. doi: 10.1371/journal.pbio.3001032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brubaker, D.K., Kumar, M.P., Chiswick, E.L., Gregg, C., Starchenko, A., Vega, P.N., Southard-Smith, A.N., Simmons, A.J., Scoville, E.A., Coburn, L.A., et al. (2020). An interspecies translation model implicates integrin signaling in infliximab-resistant inflammatory bowel disease. Sci Signal 13, eaay3258. [DOI] [PMC free article] [PubMed]
- Buckner CM, Moir S, Kardava L, Ho J, Santich BH, Kim LJ, Funk EK, Nelson AK, Winckler B, Chairez CL, et al. CXCR4/IgG-expressing plasma cells are associated with human gastrointestinal tissue inflammation. J Allergy Clin Immunol. 2014;133(1676–1685):e1675. doi: 10.1016/j.jaci.2013.10.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bues J, Biocanin M, Pezoldt J, Dainese R, Chrisnandy A, Rezakhani S, Saelens W, Gardeux V, Gupta R, Sarkis R, et al. Deterministic scRNA-seq captures variation in intestinal crypt and organoid composition. Nat Methods. 2022;19:323–330. doi: 10.1038/s41592-021-01391-1. [DOI] [PubMed] [Google Scholar]
- Burclaff J, Bliton RJ, Breau KA, Ok MT, Gomez-Martinez I, Ranek JS, Bhatt AP, Purvis JE, Woosley JT, Magness ST. A Proximal-to-Distal Survey of Healthy Adult Human Small Intestine and Colon Epithelium by Single-Cell Transcriptomics. Cell Mol Gastroenterol Hepatol. 2022;13:1554–1589. doi: 10.1016/j.jcmgh.2022.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Busslinger GA, Weusten BLA, Bogte A, Begthel H, Brosens LAA, Clevers H. Human gastrointestinal epithelia of the esophagus, stomach, and duodenum resolved at single-cell resolution. Cell Rep. 2021;34:108819. doi: 10.1016/j.celrep.2021.108819. [DOI] [PubMed] [Google Scholar]
- Chen S, Zheng Y, Ran X, Du H, Feng H, Yang L, Wen Y, Lin C, Wang S, Huang M, et al. Integrin alphaEbeta7(+) T cells direct intestinal stem cell fate decisions via adhesion signaling. Cell Res. 2021;31:1291–1307. doi: 10.1038/s41422-021-00561-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen W, Zhao Y, Chen X, Yang Z, Xu X, Bi Y, Chen V, Li J, Choi H, Ernest B, et al. A multicenter study benchmarking single-cell RNA sequencing technologies using reference samples. Nat Biotechnol. 2021;39:1103–1114. doi: 10.1038/s41587-020-00748-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng H, Leblond CP. Origin, differentiation and renewal of the four main epithelial cell types in the mouse small intestine. I Columnar Cell Am J Anat. 1974;141:461–479. doi: 10.1002/aja.1001410403. [DOI] [PubMed] [Google Scholar]
- Chikina AS, Nadalin F, Maurin M, San-Roman M, Thomas-Bonafos T, Li XV, Lameiras S, Baulande S, Henri S, Malissen B, et al. Macrophages Maintain Epithelium Integrity by Limiting Fungal Product Absorption. Cell. 2020;183(411–428):e416. doi: 10.1016/j.cell.2020.08.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clevers H. Modeling Development and Disease with Organoids. Cell. 2016;165:1586–1597. doi: 10.1016/j.cell.2016.05.082. [DOI] [PubMed] [Google Scholar]
- Clevers H, Batlle E. SnapShot: the intestinal crypt. Cell. 2013;152(1198–1198):e1192. doi: 10.1016/j.cell.2013.02.030. [DOI] [PubMed] [Google Scholar]
- Clevers HC, Bevins CL. Paneth cells: maestros of the small intestinal crypts. Annu Rev Physiol. 2013;75:289–311. doi: 10.1146/annurev-physiol-030212-183744. [DOI] [PubMed] [Google Scholar]
- Dalerba P, Kalisky T, Sahoo D, Rajendran PS, Rothenberg ME, Leyrat AA, Sim S, Okamoto J, Johnston DM, Qian D, et al. Single-cell dissection of transcriptional heterogeneity in human colon tumors. Nat Biotechnol. 2011;29:1120–1127. doi: 10.1038/nbt.2038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darmanis S, Sloan SA, Croote D, Mignardi M, Chernikova S, Samghababi P, Zhang Y, Neff N, Kowarsky M, Caneda C, et al. Single-Cell RNA-Seq Analysis of Infiltrating Neoplastic Cells at the Migrating Front of Human Glioblastoma. Cell Rep. 2017;21:1399–1410. doi: 10.1016/j.celrep.2017.10.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Degirmenci B, Valenta T, Dimitrieva S, Hausmann G, Basler K. GLI1-expressing mesenchymal cells form the essential Wnt-secreting niche for colon stem cells. Nature. 2018;558:449–453. doi: 10.1038/s41586-018-0190-3. [DOI] [PubMed] [Google Scholar]
- Dekker E, Tanis PJ, Vleugels JLA, Kasi PM, Wallace MB. Colorectal cancer. Lancet. 2019;394:1467–1480. doi: 10.1016/S0140-6736(19)32319-0. [DOI] [PubMed] [Google Scholar]
- Devlin JC, Axelrad J, Hine AM, Chang S, Sarkar S, Lin JD, Ruggles KV, Hudesman D, Cadwell K, Loke P. Single-Cell Transcriptional Survey of Ileal-Anal Pouch Immune Cells From Ulcerative Colitis Patients. Gastroenterology. 2021;160:1679–1693. doi: 10.1053/j.gastro.2020.12.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Domanska D, Majid U, Karlsen VT, Merok MA, Beitnes AR, Yaqub S, Baekkevold ES, Jahnsen FL. Single-cell transcriptomic analysis of human colonic macrophages reveals niche-specific subsets. J Exp Med. 2022;219:e20211846. doi: 10.1084/jem.20211846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dong J, Hu Y, Fan X, Wu X, Mao Y, Hu B, Guo H, Wen L, Tang F. Single-cell RNA-seq analysis unveils a prevalent epithelial/mesenchymal hybrid state during mouse organogenesis. Genome Biol. 2018;19:31. doi: 10.1186/s13059-018-1416-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du L, Wang H, He L, Zhang J, Ni B, Wang X, Jin H, Cahuzac N, Mehrpour M, Lu Y, et al. CD44 is of functional importance for colorectal cancer stem cells. Clin Cancer Res. 2008;14:6751–6760. doi: 10.1158/1078-0432.CCR-08-1034. [DOI] [PubMed] [Google Scholar]
- Elmentaite R, Ross ADB, Roberts K, James KR, Ortmann D, Gomes T, Nayak K, Tuck L, Pritchard S, Bayraktar OA, et al. Single-Cell Sequencing of Developing Human Gut Reveals Transcriptional Links to Childhood Crohn's Disease. Dev Cell. 2020;55(771–783):e775. doi: 10.1016/j.devcel.2020.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elmentaite R, Kumasaka N, Roberts K, Fleming A, Dann E, King HW, Kleshchevnikov V, Dabrowska M, Pritchard S, Bolt L, et al. Cells of the human intestinal tract mapped across space and time. Nature. 2021;597:250–255. doi: 10.1038/s41586-021-03852-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fawkner-Corbett D, Antanaviciute A, Parikh K, Jagielowicz M, Geros AS, Gupta T, Ashley N, Khamis D, Fowler D, Morrissey E, et al. Spatiotemporal analysis of human intestinal development at single-cell resolution. Cell. 2021;184(810–826):e823. doi: 10.1016/j.cell.2020.12.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fazilaty H, Brugger MD, Valenta T, Szczerba BM, Berkova L, Doumpas N, Hausmann G, Scharl M, Basler K. Tracing colonic embryonic transcriptional profiles and their reactivation upon intestinal damage. Cell Rep. 2021;36:109484. doi: 10.1016/j.celrep.2021.109484. [DOI] [PubMed] [Google Scholar]
- FitzPatrick MEB, Provine NM, Garner LC, Powell K, Amini A, Irwin SL, Ferry H, Ambrose T, Friend P, Vrakas G, et al. Human intestinal tissue-resident memory T cells comprise transcriptionally and functionally distinct subsets. Cell Rep. 2021;34:108661. doi: 10.1016/j.celrep.2020.108661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Francis JM, Zhang CZ, Maire CL, Jung J, Manzo VE, Adalsteinsson VA, Homer H, Haidar S, Blumenstiel B, Pedamallu CS, et al. EGFR variant heterogeneity in glioblastoma resolved through single-nucleus sequencing. Cancer Discov. 2014;4:956–971. doi: 10.1158/2159-8290.CD-13-0879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fu X, He Q, Tao Y, Wang M, Wang W, Wang Y, Yu QC, Zhang F, Zhang X, Chen YG, et al. Recent advances in tissue stem cells. Science China Life Science. 2021;64:1998–2029. doi: 10.1007/s11427-021-2007-8. [DOI] [PubMed] [Google Scholar]
- Fujii M, Matano M, Toshimitsu K, Takano A, Mikami Y, Nishikori S, Sugimoto S, Sato T. Human Intestinal Organoids Maintain Self-Renewal Capacity and Cellular Diversity in Niche-Inspired Culture Condition. Cell Stem Cell. 2018;23(787–793):e786. doi: 10.1016/j.stem.2018.11.016. [DOI] [PubMed] [Google Scholar]
- Funk MC, Zhou J, Boutros M. Ageing, metabolism and the intestine. EMBO Rep. 2020;21:e50047. doi: 10.15252/embr.202050047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furness JB, Rivera LR, Cho HJ, Bravo DM, Callaghan B. The gut as a sensory organ. Nat Rev Gastroenterol Hepatol. 2013;10:729–740. doi: 10.1038/nrgastro.2013.180. [DOI] [PubMed] [Google Scholar]
- Gao S, Yan L, Wang R, Li J, Yong J, Zhou X, Wei Y, Wu X, Wang X, Fan X, et al. Tracing the temporal-spatial transcriptome landscapes of the human fetal digestive tract using single-cell RNA-sequencing. Nat Cell Biol. 2018;20:721–734. doi: 10.1038/s41556-018-0105-4. [DOI] [PubMed] [Google Scholar]
- Gao J, Wu Z, Zhao M, Zhang R, Li M, Sun D, Cheng H, Qi X, Shen Y, Xu Q, et al. Allosteric inhibition reveals SHP2-mediated tumor immunosuppression in colon cancer by single-cell transcriptomics. Acta Pharm Sin B. 2022;12:149–166. doi: 10.1016/j.apsb.2021.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaujoux R, Starosvetsky E, Maimon N, Vallania F, Bar-Yoseph H, Pressman S, Weisshof R, Goren I, Rabinowitz K, Waterman M, et al. Cell-centred meta-analysis reveals baseline predictors of anti-TNFalpha non-response in biopsy and blood of patients with IBD. Gut. 2019;68:604–614. doi: 10.1136/gutjnl-2017-315494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gehart H, van Es JH, Hamer K, Beumer J, Kretzschmar K, Dekkers JF, Rios A, Clevers H. Identification of Enteroendocrine Regulators by Real-Time Single-Cell Differentiation Mapping. Cell. 2019;176(1158–1173):e1116. doi: 10.1016/j.cell.2018.12.029. [DOI] [PubMed] [Google Scholar]
- George MD, Wehkamp J, Kays RJ, Leutenegger CM, Sabir S, Grishina I, Dandekar S, Bevins CL. In vivo gene expression profiling of human intestinal epithelial cells: analysis by laser microdissection of formalin fixed tissues. BMC Genomics. 2008;9:209. doi: 10.1186/1471-2164-9-209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerbe F, Jay P. Intestinal tuft cells: epithelial sentinels linking luminal cues to the immune system. Mucosal Immunol. 2016;9:1353–1359. doi: 10.1038/mi.2016.68. [DOI] [PubMed] [Google Scholar]
- Gerbe F, Legraverend C, Jay P. The intestinal epithelium tuft cells: specification and function. Cell Mol Life Sci. 2012;69:2907–2917. doi: 10.1007/s00018-012-0984-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghazavi F, Huysentruyt J, De Coninck J, Kourula S, Martens S, Hassannia B, Wartewig T, Divert T, Roelandt R, Popper B, et al. Executioner caspases 3 and 7 are dispensable for intestinal epithelium turnover and homeostasis at steady state. Proc Natl Acad Sci U S A. 2022;119:e2024508119. doi: 10.1073/pnas.2024508119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glass LL, Calera-Nieto FJ, Jawaid W, Larraufle P, Kay RG, Gottgens B, Relmann F, Gribble FM. Single-cell RNA-sequencing reveals a distinct population of proglucagon-expressing cells specific to the mouse upper small intestine. Molecular Metabolism. 2017;6:1296–1303. doi: 10.1016/j.molmet.2017.07.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gorreja F, Caer C, Rush STA, Forsskal SK, Hartlova A, Magnusson MK, Bexe Lindskog E, Borjesson LG, Block M, Wick MJ. MEFV and NLRP3 Inflammasome Expression Is Attributed to Immature Macrophages and Correlates with Serum Inflammatory Proteins in Crohn s Disease Patients. Inflammation. 2022;45:1631–1650. doi: 10.1007/s10753-022-01647-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grencis RK. Immunity to helminths: resistance, regulation, and susceptibility to gastrointestinal nematodes. Annu Rev Immunol. 2015;33:201–225. doi: 10.1146/annurev-immunol-032713-120218. [DOI] [PubMed] [Google Scholar]
- Gribble FM, Reimann F. Enteroendocrine Cells: Chemosensors in the Intestinal Epithelium. Annu Rev Physiol. 2016;78:277–299. doi: 10.1146/annurev-physiol-021115-105439. [DOI] [PubMed] [Google Scholar]
- Grun D, Lyubimova A, Kester L, Wiebrands K, Basak O, Sasaki N, Clevers H, van Oudenaarden A. Single-cell messenger RNA sequencing reveals rare intestinal cell types. Nature. 2015;525:251–255. doi: 10.1038/nature14966. [DOI] [PubMed] [Google Scholar]
- Guo X, Yin C, Yang F, Zhang Y, Huang H, Wang J, Deng B, Cai T, Rao Y, Xi R. The Cellular Diversity and Transcription Factor Code of Drosophila Enteroendocrine Cells. Cell Rep. 2019;29(4172–4185):e4175. doi: 10.1016/j.celrep.2019.11.048. [DOI] [PubMed] [Google Scholar]
- Guo W, Zhang C, Wang X, Dou D, Chen D, Li J. Resolving the difference between left-sided and right-sided colorectal cancer by single-cell sequencing. JCI Insight. 2022;7:e152616. doi: 10.1172/jci.insight.152616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haber AL, Biton M, Rogel N, Herbst RH, Shekhar K, Smillie C, Burgin G, Delorey TM, Howitt MR, Katz Y, et al. A single-cell survey of the small intestinal epithelium. Nature. 2017;551:333–339. doi: 10.1038/nature24489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- He D, Wu H, Xiang J, Ruan X, Peng P, Ruan Y, Chen YG, Wang Y, Yu Q, Zhang H, et al. Gut stem cell aging is driven by mTORC1 via a p38 MAPK-p53 pathway. Nat Commun. 2020;11:37. doi: 10.1038/s41467-019-13911-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- He X, Smith SE, Chen S, Li H, Wu D, Meneses-Giles PI, Wang Y, Hembree M, Yi K, Zhao X, et al. Tumor-initiating stem cell shapes its microenvironment into an immunosuppressive barrier and pro-tumorigenic niche. Cell Rep. 2021;36:109674. doi: 10.1016/j.celrep.2021.109674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Helander HF, Fandriks L. Surface area of the digestive tract - revisited. Scand J Gastroenterol. 2014;49:681–689. doi: 10.3109/00365521.2014.898326. [DOI] [PubMed] [Google Scholar]
- Herring CA, Banerjee A, McKinley ET, Simmons AJ, Ping J, Roland JT, Franklin JL, Liu Q, Gerdes MJ, Coffey RJ, et al. Unsupervised Trajectory Analysis of Single-Cell RNA-Seq and Imaging Data Reveals Alternative Tuft Cell Origins in the Gut. Cell Syst. 2018;6(37–51):e39. doi: 10.1016/j.cels.2017.10.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holloway EM, Wu JH, Czerwinski M, Sweet CW, Wu A, Tsai YH, Huang S, Stoddard AE, Capeling MM, Glass I, et al. Differentiation of Human Intestinal Organoids with Endogenous Vascular Endothelial Cells. Dev Cell. 2020;54(516–528):e517. doi: 10.1016/j.devcel.2020.07.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holloway EM, Czerwinski M, Tsai YH, Wu JH, Wu A, Childs CJ, Walton KD, Sweet CW, Yu Q, Glass I, et al. Mapping Development of the Human Intestinal Niche at Single-Cell Resolution. Cell Stem Cell. 2021;28(568–580):e564. doi: 10.1016/j.stem.2020.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hong SP, Yang MJ, Cho H, Park I, Bae H, Choe K, Suh SH, Adams RH, Alitalo K, Lim D, et al. Distinct fibroblast subsets regulate lacteal integrity through YAP/TAZ-induced VEGF-C in intestinal villi. Nat Commun. 2020;11:4102. doi: 10.1038/s41467-020-17886-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang B, Chen Z, Geng L, Wang J, Liang H, Cao Y, Chen H, Huang W, Su M, Wang H, et al. Mucosal Profiling of Pediatric-Onset Colitis and IBD Reveals Common Pathogenics and Therapeutic Pathways. Cell. 2019;179(1160–1176):e1124. doi: 10.1016/j.cell.2019.10.027. [DOI] [PubMed] [Google Scholar]
- Huang LJ, Mao XT, Li YY, Liu DD, Fan KQ, Liu RB, Wu TT, Wang HL, Zhang Y, Yang B, et al. Multiomics analyses reveal a critical role of selenium in controlling T cell differentiation in Crohn's disease. Immunity. 2021;54(1728–1744):e1727. doi: 10.1016/j.immuni.2021.07.004. [DOI] [PubMed] [Google Scholar]
- Hutchins AP. Single cells and transposable element heterogeneity in stem cells and development. Cell Regen. 2021;10:23. doi: 10.1186/s13619-021-00085-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaeger N, Gamini R, Cella M, Schettini JL, Bugatti M, Zhao S, Rosadini CV, Esaulova E, Di Luccia B, Kinnett B, et al. Single-cell analyses of Crohn's disease tissues reveal intestinal intraepithelial T cells heterogeneity and altered subset distributions. Nat Commun. 2021;12:1921. doi: 10.1038/s41467-021-22164-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jasso GJ, Jaiswal A, Varma M, Laszewski T, Grauel A, Omar A, Silva N, Dranoff G, Porter JA, Mansfield K, et al. Colon stroma mediates an inflammation-driven fibroblastic response controlling matrix remodeling and healing. PLoS Biol. 2022;20:e3001532. doi: 10.1371/journal.pbio.3001532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang H, Zhang H, Zhang X. Single-cell genomic profile-based analysis of tissue differentiation in colorectal cancer. Sci China Life Sci. 2021;64:1311–1325. doi: 10.1007/s11427-020-1811-5. [DOI] [PubMed] [Google Scholar]
- Kalucka J, de Rooij L, Goveia J, Rohlenova K, Dumas SJ, Meta E, Conchinha NV, Taverna F, Teuwen LA, Veys K, et al. Single-Cell Transcriptome Atlas of Murine Endothelial Cells. Cell. 2020;180(764–779):e720. doi: 10.1016/j.cell.2020.01.015. [DOI] [PubMed] [Google Scholar]
- Kim JE, Fei L, Yin WC, Coquenlorge S, Rao-Bhatia A, Zhang X, Shi SSW, Lee JH, Hahn NA, Rizvi W, et al. Single cell and genetic analyses reveal conserved populations and signaling mechanisms of gastrointestinal stromal niches. Nat Commun. 2020;11:334. doi: 10.1038/s41467-019-14058-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kinchen J, Chen HH, Parikh K, Antanaviciute A, Jagielowicz M, Fawkner-Corbett D, Ashley N, Cubitt L, Mellado-Gomez E, Attar M, et al. Structural Remodeling of the Human Colonic Mesenchyme in Inflammatory Bowel Disease. Cell. 2018;175(372–386):e317. doi: 10.1016/j.cell.2018.08.067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiner E, Willie E, Vijaykumar B, Chowdhary K, Schmutz H, Chandler J, Schnell A, Thakore PI, LeGros G, Mostafavi S, et al. Gut CD4(+) T cell phenotypes are a continuum molded by microbes, not by TH archetypes. Nat Immunol. 2021;22:216–228. doi: 10.1038/s41590-020-00836-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lai S, Ma L, E, W., Ye, F., Chen, H., Han, X., and Guo, G. Mapping a mammalian adult adrenal gland hierarchy across species by microwell-seq. Cell Regen. 2020;9:11. doi: 10.1186/s13619-020-00042-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leblond CP, Walker BE. Renewal of cell populations. Physiol Rev. 1956;36:255–276. doi: 10.1152/physrev.1956.36.2.255. [DOI] [PubMed] [Google Scholar]
- Leung ML, Davis A, Gao R, Casasent A, Wang Y, Sei E, Vilar E, Maru D, Kopetz S, Navin NE. Single-cell DNA sequencing reveals a late-dissemination model in metastatic colorectal cancer. Genome Res. 2017;27:1287–1299. doi: 10.1101/gr.209973.116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li N, Yousefi M, Nakauka-Ddamba A, Jain R, Tobias J, Epstein JA, Jensen ST, Lengner CJ. Single-Cell Analysis of Proxy Reporter Allele-Marked Epithelial Cells Establishes Intestinal Stem Cell Hierarchy. Stem Cell Reports. 2014;3:876–891. doi: 10.1016/j.stemcr.2014.09.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li N, Nakauka-Ddamba A, Tobias J, Jensen ST, Lengner CJ. Mouse Label-Retaining Cells Are Molecularly and Functionally Distinct From Reserve Intestinal Stem Cells. Gastroenterology. 2016;151(298–310):e297. doi: 10.1053/j.gastro.2016.04.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li H, Courtois ET, Sengupta D, Tan Y, Chen KH, Goh JJL, Kong SL, Chua C, Hon LK, Tan WS, et al. Reference component analysis of single-cell transcriptomes elucidates cellular heterogeneity in human colorectal tumors. Nat Genet. 2017;49:708–718. doi: 10.1038/ng.3818. [DOI] [PubMed] [Google Scholar]
- Li J, Wang R, Zhou X, Wang W, Gao S, Mao Y, Wu X, Guo L, Liu H, Wen L, et al. Genomic and transcriptomic profiling of carcinogenesis in patients with familial adenomatous polyposis. Gut. 2020;69:1283–1293. doi: 10.1136/gutjnl-2019-319438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li H, Wang X, Wang Y, Zhang M, Hong F, Wang H, Cui A, Zhao J, Ji W, Chen YG. Cross-species single-cell transcriptomic analysis reveals divergence of cell composition and functions in mammalian ileum epithelium. Cell Regen. 2022;11:19. doi: 10.1186/s13619-022-00118-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Y, Chen YG. Intestinal epithelial plasticity and regeneration via cell dedifferentiation. Cell Regen. 2020;9:14. doi: 10.1186/s13619-020-00053-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu M, Liu Y, Di J, Su Z, Yang H, Jiang B, Wang Z, Zhuang M, Bai F, Su X. Multi-region and single-cell sequencing reveal variable genomic heterogeneity in rectal cancer. BMC Cancer. 2017;17:787. doi: 10.1186/s12885-017-3777-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu M, Di J, Liu Y, Su Z, Jiang B, Wang Z, Su X. Comparison of EpCAM(high)CD44(+) cancer stem cells with EpCAM(high)CD44(-) tumor cells in colon cancer by single-cell sequencing. Cancer Biol Ther. 2018;19:939–947. doi: 10.1080/15384047.2018.1456605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu H, Dasgupta S, Fu Y, Bailey B, Roy C, Lightcap E, Faustin B. Subsets of mononuclear phagocytes are enriched in the inflamed colons of patients with IBD. BMC Immunol. 2019;20:42. doi: 10.1186/s12865-019-0322-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu CY, Girish N, Gomez ML, Dube PE, Washington MK, Simons BD, Polk DB. Transitional Anal Cells Mediate Colonic Re-epithelialization in Colitis. Gastroenterology. 2022;S0016–5085(0022):00196–192. doi: 10.1053/j.gastro.2022.02.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Llado V, Nakanishi Y, Duran A, Reina-Campos M, Shelton PM, Linares JF, Yajima T, Campos A, Aza-Blanc P, Leitges M, et al. Repression of Intestinal Stem Cell Function and Tumorigenesis through Direct Phosphorylation of beta-Catenin and Yap by PKCzeta. Cell Rep. 2015;10:740–754. doi: 10.1016/j.celrep.2015.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luoma AM, Suo S, Williams HL, Sharova T, Sullivan K, Manos M, Bowling P, Hodi FS, Rahma O, Sullivan RJ, et al. Molecular Pathways of Colon Inflammation Induced by Cancer Immunotherapy. Cell. 2020;182(655–671):e622. doi: 10.1016/j.cell.2020.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lutter L, Hoytema van Konijnenburg DP, Brand EC, Oldenburg B, van Wijk F. The elusive case of human intraepithelial T cells in gut homeostasis and inflammation. Nat Rev Gastroenterol Hepatol. 2018;15:637–649. doi: 10.1038/s41575-018-0039-0. [DOI] [PubMed] [Google Scholar]
- Mabbott NA, Donaldson DS, Ohno H, Williams IR, Mahajan A. Microfold (M) cells: important immunosurveillance posts in the intestinal epithelium. Mucosal Immunol. 2013;6:666–677. doi: 10.1038/mi.2013.30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magness ST, Puthoff BJ, Crissey MA, Dunn J, Henning SJ, Houchen C, Kaddis JS, Kuo CJ, Li L, Lynch J, et al. A multicenter study to standardize reporting and analyses of fluorescence-activated cell-sorted murine intestinal epithelial cells. Am J Physiol Gastrointest Liver Physiol. 2013;305:G542–551. doi: 10.1152/ajpgi.00481.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mariadason JM, Nicholas C, L'Italien KE, Zhuang M, Smartt HJ, Heerdt BG, Yang W, Corner GA, Wilson AJ, Klampfer L, et al. Gene expression profiling of intestinal epithelial cell maturation along the crypt-villus axis. Gastroenterology. 2005;128:1081–1088. doi: 10.1053/j.gastro.2005.01.054. [DOI] [PubMed] [Google Scholar]
- Martin JC, Chang C, Boschetti G, Ungaro R, Giri M, Grout JA, Gettler K, Chuang LS, Nayar S, Greenstein AJ, et al. Single-Cell Analysis of Crohn's Disease Lesions Identifies a Pathogenic Cellular Module Associated with Resistance to Anti-TNF Therapy. Cell. 2019;178(1493–1508):e1420. doi: 10.1016/j.cell.2019.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masuda K, Kornberg A, Miller J, Lin S, Suek N, Botella T, Secener KA, Bacarella AM, Cheng L, Ingham M, et al. Multiplexed single-cell analysis reveals prognostic and nonprognostic T cell types in human colorectal cancer. JCI Insight. 2022;7:e154646. doi: 10.1172/jci.insight.154646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCauley HA, Guasch G. Three cheers for the goblet cell: maintaining homeostasis in mucosal epithelia. Trends Mol Med. 2015;21:492–503. doi: 10.1016/j.molmed.2015.06.003. [DOI] [PubMed] [Google Scholar]
- McKinley ET, Sui Y, Al-Kofahi Y, Millis BA, Tyska MJ, Roland JT, Santamaria-Pang A, Ohland CL, Jobin C, Franklin JL, et al. Optimized multiplex immunofluorescence single-cell analysis reveals tuft cell heterogeneity. JCI Insight. 2017;2:e93487. doi: 10.1172/jci.insight.93487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mead BE, Ordovas-Montanes J, Braun AP, Levy LE, Bhargava P, Szucs MJ, Ammendolia DA, MacMullan MA, Yin X, Hughes TK, et al. Harnessing single-cell genomics to improve the physiological fidelity of organoid-derived cell types. BMC Biol. 2018;16:62. doi: 10.1186/s12915-018-0527-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meng, Q., Chen, L., Xiong, B., Kang, B., Zhang, P., Tang, S., Han, H., Shen, W., Feng, X., Feng, S., et al. (2021). Single-Cell Transcriptome Sequencing and Proteomics Reveal Neonatal Ileum Dynamic Developmental Potentials. mSystems, e0072521. [DOI] [PMC free article] [PubMed]
- Mihaylova MM, Cheng CW, Cao AQ, Tripathi S, Mana MD, Bauer-Rowe KE, Abu-Remaileh M, Clavain L, Erdemir A, Lewis CA, et al. Fasting Activates Fatty Acid Oxidation to Enhance Intestinal Stem Cell Function during Homeostasis and Aging. Cell Stem Cell. 2018;22(769–778):e764. doi: 10.1016/j.stem.2018.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moor AE, Harnik Y, Ben-Moshe S, Massasa EE, Rozenberg M, Eilam R, Bahar Halpern K, Itzkovitz S. Spatial Reconstruction of Single Enterocytes Uncovers Broad Zonation along the Intestinal Villus Axis. Cell. 2018;175(1156–1167):e1115. doi: 10.1016/j.cell.2018.08.063. [DOI] [PubMed] [Google Scholar]
- Murata K, Jadhav U, Madha S, van Es J, Dean J, Cavazza A, Wucherpfennig K, Michor F, Clevers H, Shivdasani RA. Ascl2-Dependent Cell Dedifferentiation Drives Regeneration of Ablated Intestinal Stem Cells. Cell Stem Cell. 2020;26(377–390):e376. doi: 10.1016/j.stem.2019.12.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nalapareddy K, Nattamai KJ, Kumar RS, Karns R, Wikenheiser-Brokamp KA, Sampson LL, Mahe MM, Sundaram N, Yacyshyn MB, Yacyshyn B, et al. Canonical Wnt Signaling Ameliorates Aging of Intestinal Stem Cells. Cell Rep. 2017;18:2608–2621. doi: 10.1016/j.celrep.2017.02.056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Navin N, Kendall J, Troge J, Andrews P, Rodgers L, McIndoo J, Cook K, Stepansky A, Levy D, Esposito D, et al. Tumour evolution inferred by single-cell sequencing. Nature. 2011;472:90–94. doi: 10.1038/nature09807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nayar S, Morrison JK, Giri M, Gettler K, Chuang LS, Walker LA, Ko HM, Kenigsberg E, Kugathasan S, Merad M, et al. A myeloid-stromal niche and gp130 rescue in NOD2-driven Crohn's disease. Nature. 2021;593:275–281. doi: 10.1038/s41586-021-03484-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neal JT, Li X, Zhu J, Giangarra V, Grzeskowiak CL, Ju J, Liu IH, Chiou SH, Salahudeen AA, Smith AR, et al. Organoid Modeling of the Tumor Immune Microenvironment. Cell. 2018;175(1972–1988):e1916. doi: 10.1016/j.cell.2018.11.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nefzger CM, Jarde T, Srivastava A, Schroeder J, Rossello FJ, Horvay K, Prasko M, Paynter JM, Chen J, Weng CF, et al. Intestinal stem cell aging signature reveals a reprogramming strategy to enhance regenerative potential. NPJ Regen Med. 2022;7:31. doi: 10.1038/s41536-022-00226-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nowotschin S, Setty M, Kuo YY, Liu V, Garg V, Sharma R, Simon CS, Saiz N, Gardner R, Boutet SC, et al. The emergent landscape of the mouse gut endoderm at single-cell resolution. Nature. 2019;569:361–367. doi: 10.1038/s41586-019-1127-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nystrom, E.E.L., Martinez-Abad, B., Arike, L., Birchenough, G.M.H., Nonnecke, E.B., Castillo, P.A., Svensson, F., Bevins, C.L., Hansson, G.C., and Johansson, M.E.V. (2021). An intercrypt subpopulation of goblet cells is essential for colonic mucus barrier function. Science 372, eabb1590. [DOI] [PMC free article] [PubMed]
- O'Leary CE, Schneider C, Locksley RM. Tuft Cells-Systemically Dispersed Sensory Epithelia Integrating Immune and Neural Circuitry. Annu Rev Immunol. 2019;37:47–72. doi: 10.1146/annurev-immunol-042718-041505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ono H, Arai Y, Furukawa E, Narushima D, Matsuura T, Nakamura H, Shiokawa D, Nagai M, Imai T, Mimori K, et al. Single-cell DNA and RNA sequencing reveals the dynamics of intra-tumor heterogeneity in a colorectal cancer model. BMC Biol. 2021;19:207. doi: 10.1186/s12915-021-01147-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paone P, Cani PD. Mucus barrier, mucins and gut microbiota: the expected slimy partners? Gut. 2020;69:2232–2243. doi: 10.1136/gutjnl-2020-322260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parigi SM, Larsson L, Das S, Ramirez Flores RO, Frede A, Tripathi KP, Diaz OE, Selin K, Morales RA, Luo X, et al. The spatial transcriptomic landscape of the healing mouse intestine following damage. Nat Commun. 2022;13:828. doi: 10.1038/s41467-022-28497-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parikh K, Antanaviciute A, Fawkner-Corbett D, Jagielowicz M, Aulicino A, Lagerholm C, Davis S, Kinchen J, Chen HH, Alham NK, et al. Colonic epithelial cell diversity in health and inflammatory bowel disease. Nature. 2019;567:49–55. doi: 10.1038/s41586-019-0992-y. [DOI] [PubMed] [Google Scholar]
- Park SR, Namkoong S, Friesen L, Cho CS, Zhang ZZ, Chen YC, Yoon E, Kim CH, Kwak H, Kang HM, et al. Single-Cell Transcriptome Analysis of Colon Cancer Cell Response to 5-Fluorouracil-Induced DNA Damage. Cell Rep. 2020;32:108077. doi: 10.1016/j.celrep.2020.108077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parrish AR. The impact of aging on epithelial barriers. Tissue Barriers. 2017;5:e1343172. doi: 10.1080/21688370.2017.1343172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pastushenko I, Brisebarre A, Sifrim A, Fioramonti M, Revenco T, Boumahdi S, Van Keymeulen A, Brown D, Moers V, Lemaire S, et al. Identification of the tumour transition states occurring during EMT. Nature. 2018;556:463–468. doi: 10.1038/s41586-018-0040-3. [DOI] [PubMed] [Google Scholar]
- Pentinmikko N, Katajisto P. The role of stem cell niche in intestinal aging. Mech Ageing Dev. 2020;191:111330. doi: 10.1016/j.mad.2020.111330. [DOI] [PubMed] [Google Scholar]
- Peterson LW, Artis D. Intestinal epithelial cells: regulators of barrier function and immune homeostasis. Nat Rev Immunol. 2014;14:141–153. doi: 10.1038/nri3608. [DOI] [PubMed] [Google Scholar]
- Piccand J, Vagne C, Blot F, Meunier A, Beucher A, Strasser P, Lund ML, Ghimire S, Nivlet L, Lapp C, et al. Rfx6 promotes the differentiation of peptide-secreting enteroendocrine cells while repressing genetic programs controlling serotonin production. Molecular Metabolism. 2019;29:24–39. doi: 10.1016/j.molmet.2019.08.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Picelli S, Bjorklund AK, Faridani OR, Sagasser S, Winberg G, Sandberg R. Smart-seq2 for sensitive full-length transcriptome profiling in single cells. Nat Methods. 2013;10:1096–1098. doi: 10.1038/nmeth.2639. [DOI] [PubMed] [Google Scholar]
- Pinto D, Gregorieff A, Begthel H, Clevers H. Canonical Wnt signals are essential for homeostasis of the intestinal epithelium. Genes Dev. 2003;17:1709–1713. doi: 10.1101/gad.267103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Puram SV, Tirosh I, Parikh AS, Patel AP, Yizhak K, Gillespie S, Rodman C, Luo CL, Mroz EA, Emerick KS, et al. Single-Cell Transcriptomic Analysis of Primary and Metastatic Tumor Ecosystems in Head and Neck Cancer. Cell. 2017;171(1611–1624):e1624. doi: 10.1016/j.cell.2017.10.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qi Z, Chen YG. Regulation of intestinal stem cell fate specification. Sci China Life Sci. 2015;58:570–578. doi: 10.1007/s11427-015-4859-7. [DOI] [PubMed] [Google Scholar]
- Qin X, Sufi J, Vlckova P, Kyriakidou P, Acton SE, Li VSW, Nitz M, Tape CJ. Cell-type-specific signaling networks in heterocellular organoids. Nat Methods. 2020;17:335–342. doi: 10.1038/s41592-020-0737-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rao-Bhatia A, Zhu M, Yin WC, Coquenlorge S, Zhang X, Woo J, Sun Y, Dean CH, Liu A, Hui CC, et al. Hedgehog-Activated Fat4 and PCP Pathways Mediate Mesenchymal Cell Clustering and Villus Formation in Gut Development. Dev Cell. 2020;52(647–658):e646. doi: 10.1016/j.devcel.2020.02.003. [DOI] [PubMed] [Google Scholar]
- Rathinam VAK, Chan FK. Inflammasome, Inflammation, and Tissue Homeostasis. Trends Mol Med. 2018;24:304–318. doi: 10.1016/j.molmed.2018.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roerink SF, Sasaki N, Lee-Six H, Young MD, Alexandrov LB, Behjati S, Mitchell TJ, Grossmann S, Lightfoot H, Egan DA, et al. Intra-tumour diversification in colorectal cancer at the single-cell level. Nature. 2018;556:457–462. doi: 10.1038/s41586-018-0024-3. [DOI] [PubMed] [Google Scholar]
- Rothenberg ME, Nusse Y, Kalisky T, Lee JJ, Dalerba P, Scheeren F, Lobo N, Kulkarni S, Sim S, Qian D, et al. Identification of a cKit(+) colonic crypt base secretory cell that supports Lgr5(+) stem cells in mice. Gastroenterology. 2012;142(1195–1205):e1196. doi: 10.1053/j.gastro.2012.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roulis M, Kaklamanos A, Schernthanner M, Bielecki P, Zhao J, Kaffe E, Frommelt LS, Qu R, Knapp MS, Henriques A, et al. Paracrine orchestration of intestinal tumorigenesis by a mesenchymal niche. Nature. 2020;580:524–529. doi: 10.1038/s41586-020-2166-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanger GJ, Lee K. Hormones of the gut-brain axis as targets for the treatment of upper gastrointestinal disorders. Nat Rev Drug Discov. 2008;7:241–254. doi: 10.1038/nrd2444. [DOI] [PubMed] [Google Scholar]
- Sato T, Vries RG, Snippert HJ, van de Wetering M, Barker N, Stange DE, van Es JH, Abo A, Kujala P, Peters PJ, et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature. 2009;459:262–265. doi: 10.1038/nature07935. [DOI] [PubMed] [Google Scholar]
- Sato T, Sase M, Ishikawa S, Kajita M, Asano J, Sato T, Mori Y, Ohteki T. Characterization of radioresistant epithelial stem cell heterogeneity in the damaged mouse intestine. Sci Rep. 2020;10:8308. doi: 10.1038/s41598-020-64987-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider C, O'Leary CE, Locksley RM. Regulation of immune responses by tuft cells. Nat Rev Immunol. 2019;19:584–593. doi: 10.1038/s41577-019-0176-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seiler KM, Waye SE, Kong W, Kamimo K, Bajinting A, Goo WH, Onufer EJ, Courtney C, Guo J, Warner BW, et al. Single-Cell Analysis Reveals Regional Reprogramming During Adaptation to Massive Small Bowel Resection in Mice. Cell Mol Gastroenterol Hepatol. 2019;8:407–426. doi: 10.1016/j.jcmgh.2019.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Serra D, Mayr U, Boni A, Lukonin I, Rempfler M, Challet Meylan L, Stadler MB, Strnad P, Papasaikas P, Vischi D, et al. Self-organization and symmetry breaking in intestinal organoid development. Nature. 2019;569:66–72. doi: 10.1038/s41586-019-1146-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shemtov, S.J., Emani, R., Bielska, O., Covarrubias, A.J., Verdin, E., Andersen, J.K., and Winer, D.A. (2022). The intestinal immune system and gut barrier function in obesity and aging. FEBS J. [DOI] [PMC free article] [PubMed]
- Shen T, Liu JL, Wang CY, Rixiati Y, Li S, Cai LD, Zhao YY, Li JM. Targeting Erbin in B cells for therapy of lung metastasis of colorectal cancer. Signal Transduct Target Ther. 2021;6:115. doi: 10.1038/s41392-021-00501-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheng X, Lin Z, Lv C, Shao C, Bi X, Deng M, Xu J, Guerrero-Juarez CF, Li M, Wu X, et al. Cycling Stem Cells Are Radioresistant and Regenerate the Intestine. Cell Rep. 2020;32:107952. doi: 10.1016/j.celrep.2020.107952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shoshkes-Carmel M, Wang YJ, Wangensteen KJ, Toth B, Kondo A, Massasa EE, Itzkovitz S, Kaestner KH. Subepithelial telocytes are an important source of Wnts that supports intestinal crypts. Nature. 2018;557:242–246. doi: 10.1038/s41586-018-0084-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sirvinskas, D., Omrani, O., Lu, J., Rasa, M., Krepelova, A., Adam, L., Kaeppel, S., Sommer, F., and Neri, F. (2022). Single-cell atlas of the aging mouse colon. iScience 25, 104202. [DOI] [PMC free article] [PubMed]
- Smillie CS, Biton M, Ordovas-Montanes J, Sullivan KM, Burgin G, Graham DB, Herbst RH, Rogel N, Slyper M, Waldman J, et al. Intra- and Inter-cellular Rewiring of the Human Colon during Ulcerative Colitis. Cell. 2019;178(714–730):e722. doi: 10.1016/j.cell.2019.06.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinbach EC, Plevy SE. The role of macrophages and dendritic cells in the initiation of inflammation in IBD. Inflamm Bowel Dis. 2014;20:166–175. doi: 10.1097/MIB.0b013e3182a69dca. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sugimoto S, Ohta Y, Fujii M, Matano M, Shimokawa M, Nanki K, Date S, Nishikori S, Nakazato Y, Nakamura T, et al. Reconstruction of the Human Colon Epithelium In Vivo. Cell Stem Cell. 2018;22(171–176):e175. doi: 10.1016/j.stem.2017.11.012. [DOI] [PubMed] [Google Scholar]
- Suzuki K, Murano T, Shimizu H, Ito G, Nakata T, Fujii S, Ishibashi F, Kawamoto A, Anzai S, Kuno R, et al. Single cell analysis of Crohn's disease patient-derived small intestinal organoids reveals disease activity-dependent modification of stem cell properties. J Gastroenterol. 2018;53:1035–1047. doi: 10.1007/s00535-018-1437-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tallapragada NP, Cambra HM, Wald T, Keough Jalbert S, Abraham DM, Klein OD, Klein AM. Inflation-collapse dynamics drive patterning and morphogenesis in intestinal organoids. Cell Stem Cell. 2021;28(1516–1532):e1514. doi: 10.1016/j.stem.2021.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan P, Ren Y, Zhang Y, Lin Y, Cui T, Huang Y, Zhang Y, Ning L, Yu J, Gao S, et al. Dissecting dynamic expression of autophagy-related genes during human fetal digestive tract development via single-cell RNA sequencing. Autophagy. 2019;15:2019–2021. doi: 10.1080/15548627.2019.1656956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tetteh PW, Basak O, Farin HF, Wiebrands K, Kretzschmar K, Begthel H, van den Born M, Korving J, de Sauvage F, van Es JH, et al. Replacement of Lost Lgr5-Positive Stem Cells through Plasticity of Their Enterocyte-Lineage Daughters. Cell Stem Cell. 2016;18:203–213. doi: 10.1016/j.stem.2016.01.001. [DOI] [PubMed] [Google Scholar]
- Tilg H, Zmora N, Adolph TE, Elinav E. The intestinal microbiota fuelling metabolic inflammation. Nat Rev Immunol. 2020;20:40–54. doi: 10.1038/s41577-019-0198-4. [DOI] [PubMed] [Google Scholar]
- Tkachev, V., Kaminski, J., Potter, E.L., Furlan, S.N., Yu, A., Hunt, D.J., McGuckin, C., Zheng, H., Colonna, L., Gerdemann, U., et al. (2021). Spatiotemporal single-cell profiling reveals that invasive and tissue-resident memory donor CD8(+) T cells drive gastrointestinal acute graft-versus-host disease. Sci Transl Med 13, eabc0227. [DOI] [PMC free article] [PubMed]
- Triana S, Stanifer ML, Metz-Zumaran C, Shahraz M, Mukenhirn M, Kee C, Serger C, Koschny R, Ordonez-Rueda D, Paulsen M, et al. Single-cell transcriptomics reveals immune response of intestinal cell types to viral infection. Mol Syst Biol. 2021;17:e9833. doi: 10.15252/msb.20209833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tu YH, Cooper AJ, Teng B, Chang RB, Artiga DJ, Turner HN, Mulhall EM, Ye W, Smith AD, Liman ER. An evolutionarily conserved gene family encodes proton-selective ion channels. Science. 2018;359:1047–1050. doi: 10.1126/science.aao3264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uzzan M, Martin JC, Mesin L, Livanos AE, Castro-Dopico T, Huang R, Petralia F, Magri G, Kumar S, Zhao Q, et al. Ulcerative colitis is characterized by a plasmablast-skewed humoral response associated with disease activity. Nat Med. 2022;28:766–779. doi: 10.1038/s41591-022-01680-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Es JH, Wiebrands K, Lopez-Iglesias C, de Wetering, M.v., Zeinstra, L., van den Born, M., Korving, J., Sasaki, N., Peters, P.J., van Oudenaarden, A., , et al. Enteroendocrine and tuft cells support Lgr5 stem cells on Paneth cell depletion. Proc Natl Acad Sci USA. 2019;116:26599–26605. doi: 10.1073/pnas.1801888117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vivier E, Artis D, Colonna M, Diefenbach A, Di Santo JP, Eberl G, Koyasu S, Locksley RM, McKenzie ANJ, Mebius RE, et al. Innate Lymphoid Cells: 10 Years On. Cell. 2018;174:1054–1066. doi: 10.1016/j.cell.2018.07.017. [DOI] [PubMed] [Google Scholar]
- von Moltke J, Ji M, Liang HE, Locksley RM. Tuft-cell-derived IL-25 regulates an intestinal ILC2-epithelial response circuit. Nature. 2016;529:221–225. doi: 10.1038/nature16161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang S, Chen YG. BMP signaling in homeostasis, transformation and inflammatory response of intestinal epithelium. Sci China Life Sci. 2018;61:800–807. doi: 10.1007/s11427-018-9310-7. [DOI] [PubMed] [Google Scholar]
- Wang R, Li H, Wu J, Cai ZY, Li B, Ni H, Qiu X, Chen H, Liu W, Yang ZH, et al. Gut stem cell necroptosis by genome instability triggers bowel inflammation. Nature. 2020;580:386–390. doi: 10.1038/s41586-020-2127-x. [DOI] [PubMed] [Google Scholar]
- Wang Y, Song W, Wang J, Wang T, Xiong X, Qi Z, Fu W, Yang X, Chen YG. Single-cell transcriptome analysis reveals differential nutrient absorption functions in human intestine. J Exp Med. 2020;217:e20191130. doi: 10.1084/jem.20191130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wen L, Tang F. Single-cell sequencing in stem cell biology. Genome Biol. 2016;17:71. doi: 10.1186/s13059-016-0941-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Went PT, Lugli A, Meier S, Bundi M, Mirlacher M, Sauter G, Dirnhofer S. Frequent EpCam protein expression in human carcinomas. Hum Pathol. 2004;35:122–128. doi: 10.1016/j.humpath.2003.08.026. [DOI] [PubMed] [Google Scholar]
- Williamson G, Clifford MN. Role of the small intestine, colon and microbiota in determining the metabolic fate of polyphenols. Biochem Pharmacol. 2017;139:24–39. doi: 10.1016/j.bcp.2017.03.012. [DOI] [PubMed] [Google Scholar]
- Wu H, Zhang XY, Hu Z, Hou Q, Zhang H, Li Y, Li S, Yue J, Jiang Z, Weissman SM, et al. Evolution and heterogeneity of non-hereditary colorectal cancer revealed by single-cell exome sequencing. Oncogene. 2017;36:2857–2867. doi: 10.1038/onc.2016.438. [DOI] [PubMed] [Google Scholar]
- Wu N, Sun H, Zhao X, Zhang Y, Tan J, Qi Y, Wang Q, Ng M, Liu Z, He L, et al. MAP3K2-regulated intestinal stromal cells define a distinct stem cell niche. Nature. 2021;592:606–610. doi: 10.1038/s41586-021-03283-y. [DOI] [PubMed] [Google Scholar]
- Wu Y, Yang S, Ma J, Chen Z, Song G, Rao D, Cheng Y, Huang S, Liu Y, Jiang S, et al. Spatiotemporal Immune Landscape of Colorectal Cancer Liver Metastasis at Single-Cell Level. Cancer Discov. 2022;12:134–153. doi: 10.1158/2159-8290.CD-21-0316. [DOI] [PubMed] [Google Scholar]
- Yan KS, Gevaert O, Zheng GXY, Anchang B, Probert CS, Larkin KA, Davies PS, Cheng ZF, Kaddis JS, Han A, et al. Intestinal Enteroendocrine Lineage Cells Possess Homeostatic and Injury-Inducible Stem Cell Activity. Cell Stem Cell. 2017;21(78–90):e76. doi: 10.1016/j.stem.2017.06.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yi H, Li G, Long Y, Liang W, Cui H, Zhang B, Tan Y, Li Y, Shen L, Deng D, et al. Integrative multi-omics analysis of a colon cancer cell line with heterogeneous Wnt activity revealed RUNX2 as an epigenetic regulator of EMT. Oncogene. 2020;39:5152–5164. doi: 10.1038/s41388-020-1351-z. [DOI] [PubMed] [Google Scholar]
- Yu C, Yu J, Yao X, Wu WK, Lu Y, Tang S, Li X, Bao L, Li X, Hou Y, et al. Discovery of biclonal origin and a novel oncogene SLC12A5 in colon cancer by single-cell sequencing. Cell Res. 2014;24:701–712. doi: 10.1038/cr.2014.43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yum MK, Han S, Fink J, Wu SS, Dabrowska C, Trendafilova T, Mustata R, Chatzeli L, Azzarelli R, Pshenichnaya I, et al. Tracing oncogene-driven remodelling of the intestinal stem cell niche. Nature. 2021;594:442–447. doi: 10.1038/s41586-021-03605-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, M., and Wu, C. (2020). The relationship between intestinal goblet cells and the immune response. Biosci Rep 40, BSR20201471. [DOI] [PMC free article] [PubMed]
- Zhang Z, Liu Z. Paneth cells: the hub for sensing and regulating intestinal flora. Sci China Life Sci. 2016;59:463–467. doi: 10.1007/s11427-016-5018-5. [DOI] [PubMed] [Google Scholar]
- Zhang L, Yu X, Zheng L, Zhang Y, Li Y, Fang Q, Gao R, Kang B, Zhang Q, Huang JY, et al. Lineage tracking reveals dynamic relationships of T cells in colorectal cancer. Nature. 2018;564:268–272. doi: 10.1038/s41586-018-0694-x. [DOI] [PubMed] [Google Scholar]
- Zhang P, Yang M, Zhang Y, Xiao S, Lai X, Tan A, Du S, Li S. Dissecting the Single-Cell Transcriptome Network Underlying Gastric Premalignant Lesions and Early Gastric Cancer. Cell Rep. 2019;27(1934–1947):e1935. doi: 10.1016/j.celrep.2019.04.052. [DOI] [PubMed] [Google Scholar]
- Zhang Y, Zheng L, Zhang L, Hu X, Ren X, Zhang Z. Deep single-cell RNA sequencing data of individual T cells from treatment-naive colorectal cancer patients. Sci Data. 2019;6:131. doi: 10.1038/s41597-019-0131-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang L, Li Z, Skrzypczynska KM, Fang Q, Zhang W, O'Brien SA, He Y, Wang L, Zhang Q, Kim A, et al. Single-Cell Analyses Inform Mechanisms of Myeloid-Targeted Therapies in Colon Cancer. Cell. 2020;181(442–459):e429. doi: 10.1016/j.cell.2020.03.048. [DOI] [PubMed] [Google Scholar]
- Zhang Y, Li X, Luo Z, Ma L, Zhu S, Wang Z, Wen J, Cheng S, Gu W, Lian Q, et al. ECM1 is an essential factor for the determination of M1 macrophage polarization in IBD in response to LPS stimulation. Proc Natl Acad Sci U S A. 2020;117:3083–3092. doi: 10.1073/pnas.1912774117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao Y, Feng Y, Liu M, Chen L, Meng Q, Tang X, Wang S, Liu L, Li L, Shen W, et al. Single-cell RNA sequencing analysis reveals alginate oligosaccharides preventing chemotherapy-induced mucositis. Mucosal Immunol. 2020;13:437–448. doi: 10.1038/s41385-019-0248-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao Z, Wang Y, Wu Y, Li D, Zhang T, Ma Y, Teng X, Zuo W. Single-cell analysis defines the lineage plasticity of stem cells in cervix epithelium. Cell Regen. 2021;10:36. doi: 10.1186/s13619-021-00096-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng L, Kelly CJ, Colgan SP. Physiologic hypoxia and oxygen homeostasis in the healthy intestine. A Review in the Theme: Cellular Responses to Hypoxia. Am J Physiol Cell Physiol. 2015;309:C350–360. doi: 10.1152/ajpcell.00191.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou Y, Bian S, Zhou X, Cui Y, Wang W, Wen L, Guo L, Fu W, Tang F. Single-Cell Multiomics Sequencing Reveals Prevalent Genomic Alterations in Tumor Stromal Cells of Human Colorectal Cancer. Cancer Cell. 2020;38(818–828):e815. doi: 10.1016/j.ccell.2020.09.015. [DOI] [PubMed] [Google Scholar]
- Zhu G, Hu J, Xi R. The cellular niche for intestinal stem cells: a team effort. Cell Regen. 2021;10:1. doi: 10.1186/s13619-020-00061-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu, X., Yang, M., Lin, Z., Mael, S.K., Li, Y., Zhang, L., Kong, Y., Zhang, Y., Ren, Y., Li, J., et al. (2021b). REGgamma drives Lgr5(+) stem cells to potentiate radiation induced intestinal regeneration. Sci China Life Sci. [DOI] [PubMed]
- Zinina VV, Ruehle F, Winkler P, Rebmann L, Lukas H, Mockel S, Diefenbach A, Mendez-Lago M, Soshnikova N. ID2 controls differentiation of enteroendocrine cells in mouse small intestine. Acta Physiol (oxf) 2022;234:e13773. doi: 10.1111/apha.13773. [DOI] [PubMed] [Google Scholar]
- Zorn AM, Wells JM. Vertebrate endoderm development and organ formation. Annu Rev Cell Dev Biol. 2009;25:221–251. doi: 10.1146/annurev.cellbio.042308.113344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zowada MK, Tirier SM, Dieter SM, Krieger TG, Oberlack A, Chua RL, Huerta M, Ten FW, Laaber K, Park J, et al. Functional States in Tumor-Initiating Cell Differentiation in Human Colorectal Cancer. Cancers (basel) 2021;13:1097. doi: 10.3390/cancers13051097. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.