

Abstract
Background
Pappalysin 2 (PAPPA2) mutation, occurring most frequently in skin cutaneous melanoma (SKCM) and non‐small cell lung cancer (NSCLC), is found to be related to anti‐tumour immune response. However, the association between PAPPA2 and the efficacy of immune checkpoint inhibitors (ICIs) therapy remains unknown.
Methods
To analyse the performance of PAPPA2 mutation as an indicator stratifying beneficiaries of ICIs, seven public cohorts with whole‐exome sequencing (WES) data were divided into the NSCLC set (n = 165) and the SKCM set (n = 210). For further validation, 41 NSCLC patients receiving anti‐PD‐(L)1 treatment were enrolled in China cohort (n = 41). The mechanism was explored based on The Cancer Genome Atlas database (n = 1467).
Results
In the NSCLC set, patients with PAPPA2 mutation (PAPPA2‐Mut) demonstrated a significantly superior progress free survival (PFS, hazard ratio [HR], 0.28 [95% CI, 0.14–0.53]; p < 0.001) and objective response rate (ORR, 77.8% vs. 23.2%; p < 0.001) compared to those with wide‐type PAPPA2 (PAPPA2‐WT), consistent in the SKCM set (overall survival, HR, 0.49 [95% CI: 0.31–0.78], p < 0.001; ORR, 34.1% vs. 16.9%, p = 0.039) and China cohort. Similar results were observed in multivariable models. Accordingly, PAPPA2 mutation exhibited superior performance in predicting ICIs efficacy compared with other published ICIs‐related gene mutations, such as EPHA family, MUC16, LRP1B and TTN, etc. In addition, combined utilization of PAPPA2 mutation and tumour mutational burden (TMB) could expand the identification of potential responders to ICIs therapy in both NSCLC set (HR, 0.36 [95% CI: 0.23–0.57], p < 0.001) and SKCM set (HR, 0.51 [95% CI: 0.34–0.76], p < 0.001). Moreover, PAPPA2 mutation was correlated with enhanced anti‐tumour immunity including higher activated CD4 memory T cells level, lower Treg cells level, and upregulated DNA damage repair pathways.
Conclusions
Our findings indicated that PAPPA2 mutation could serve as a novel indicator to stratify beneficiaries from ICIs therapy in NSCLC and SKCM, warranting further prospective studies.
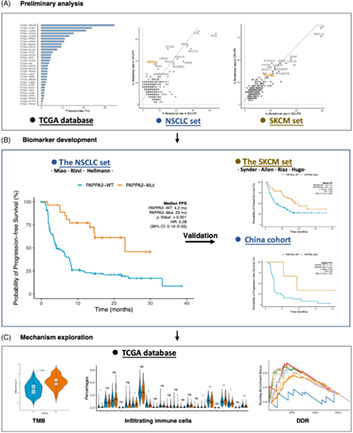
Flow diagram of the study. (A) Preliminary analysis. PAPPA2 mutated most frequently in skin cutaneous melanoma (SKCM) and non‐small cell lung cancer (NSCLC) in the The Cancer Genome Atlas (TCGA) database. PAPPA2 mutational rates in patients with objective response (CR + PR) versus without (SD + PD) were compared with other immune checkpoint inhibitors‐related gene mutations in the NSCLC and SKCM sets. (B) Biomarker development. Association between PAPPA2 mutation and clinical outcomes has been analysed in the NSCLC set, the SKCM set and China cohort. (C) Mechanism exploring. Based on the TCGA database, the correlation of PAPPA2 mutation with tumour mutation burden, infiltrating immune cells and DNA damage repair was explored for further immunogenicity and anti‐tumour activity mechanisms.
1. INTRODUCTION
Cancer is the leading cause of death in the world. In 2020, there were an estimated 19.3 million new cancer cases and nearly 10 million cancer deaths worldwide. 1 , 2 The advancement in medical technology has opened several avenues in the diagnosis and treatment of various diseases including cancer. 3 , 4 , 5 , 6
Immune checkpoint inhibitors (ICIs), targeting the programmed cell death 1 (PD‐1), programmed cell death ligand 1 (PD‐L1) or cytotoxic T lymphocyte‐associated protein 4 (CTLA‐4), have demonstrated impressive anti‐tumour efficacy in multiple cancers, in particular with non‐small cell lung cancer (NSCLC) 7 and skin cutaneous melanoma (SKCM). 8 However, the response rates of 10% ~ 20% in NSCLC 9 and 30% ~ 40% in SKCM 10 indicated that only a part of patients were therapeutic beneficiaries. Therefore, investigating predictive biomarkers of clinical outcomes from ICIs therapy is of great importance to identify the target population. 11 , 12
Even though the expression of PD‐L1, microsatellite instability, and tumour mutation burden (TMB) exhibited predictive utility to ICIs therapy response in some clinical practices, 13 , 14 , 15 the value of single gene prediction has been widely concerned, considering its relatively efficient and cost‐effective detection property. For instance, EPHA, 16 NOTCH4, 17 and LRP1B 18 mutations are all independent classifiers that could stratify beneficiaries of ICIs therapy. Although these biomarkers have been verified in some clinical trials, some limitations and indeterminacies remained. Therefore, it is necessary to explore other novel biomarkers to precisely maximize the identification of potential responders to ICIs treatment.
Pappalysin2 (PAPPA2) protein, a member of the pappalysin family of metzincin metalloproteinases, has been identified as a subset of insulin growth factor (IGF)‐binding proteins. 19 , 20 The decreasing of free IGF‐1 levels led by dysfunction of PAPPA2 protein could result in an imbalanced growth hormone (GH)/IGF‐1 signalling pathway, which was related to DNA damage repair (DDR) pathway, immune system maintenance and anti‐tumour immune activation. 21 , 22 , 23 , 24 Meantime, another study suggested that patients with PAPPA2 mutation (PAPPA2‐Mut) showed a prolonged survival time in lung adenocarcinoma (LUAD). 25 However, the understanding of the contribution of PAPPA2 mutation to the anti‐tumour immune system is still lacking and remains to be explored.
In this study, we observed that PAPPA2 mutated most frequently in NSCLC (22.2% mutant) and SKCM (34.3% mutant) based on the The Cancer Genome Atlas (TCGA) database. Therefore, to figure out the impact of PAPPA2 mutation on the clinical outcome of ICIs treatment, we investigated the association between PAPPA2 mutation and clinical efficacy of ICI in several NSCLC and SKCM cohorts. The underlying mechanisms were subsequently explored based on RNA expression and whole‐genome sequencing (WES) data from the TCGA database.
2. METHODS
2.1. Cohort description and data compilation
To investigate the association between PAPPA2 mutation and immunotherapy efficacy, seven public cohorts with WES data were collected and divided into an NSCLC set (n = 165) and a SKCM set (n = 210).
The NSCLC set (n = 165) was a pooled set consisting three independent cohorts (Rizvi cohort, 26 Hellmann cohort, 27 and Miao cohort 28 ). The SKCM set (n = 210) was a pooled set consisting four public cohorts (Synder cohort, 29 Allen cohort, 30 Riaz cohort, 31 and Hugo cohort 32 ).
We also obtained TCGA data of LUAD and SKCM to explore the mechanism underlying the association between PAPPA2 mutation and immunotherapy. The RNA sequencing (RNA‐Seq) data were retrieved from Genomic Data Commons (GDC) Data Portal (https://portal.gdc.cancer.gov/). The genomic data of WES and TMB in each TCGA tumour sample were obtained from Hoadley et al. 33 Survival data were retrieved from UCSC Xena data portal (https://xenabrowser.net). Information regarding the neoantigen load (NAL) and CIBERSORT‐inferred values in each TCGA tumour sample was obtained from Thorsson et al. 34
2.2. Patient enrollment
For further validation, we included NSCLC patients treated with anti‐PD‐(L)1 at the National Cancer Center/National Clinical Research Center for Cancer/Cancer Hospital and Chinese Academy of Medical Sciences and Peking Union Medical College (NCC) and Sun Yat‐sen University Cancer Center (SYUCC) from December 2016 to December 2018 (China cohort, n = 41). Eligible patients were 18 years of age or older with advanced or recurrent NSCLC diagnosed using Response Evaluation Criteria in Solid Tumours (RECIST) version 1.1 by investigator review, and an Eastern Cooperative Oncology Group performance status of 0 or 1. Key exclusion criteria included known active central nervous system metastases, diagnosis of immunodeficiency, prior immunotherapy for other diseases, autoimmune disease or active infection that required systemic therapy. WES analysis was performed on the tissue samples of all 41 patients. This study was approved by the ethics committees of the participating centres and all patients provided written informed consent.
2.3. Clinical outcomes
The primary clinical outcomes were progress free survival (PFS) and overall survival (OS). The secondary clinical outcomes were objective response rate (ORR), disease control rate (DCR) and durable clinical benefit (DCB). ORR and DCR were assessed using RECIST 1.1 (irRECIST for Hugo cohort, irRC for Rizvi cohort). The survival data available in the NSCLC cohorts (Rizvi, Hellmann, Miao and China cohorts) is PFS, while the survival data in common for SKCM cohorts (Synder, Allen, Riaz, and Hugo cohorts) is OS. The details of DCB definition are shown in Table S1.
2.4. PAPPA2 gene mutation
Patients with nonsynonymous somatic mutations in the coding region of the PAPPA2 gene were defined as PAPPA2‐mutant (PAPPA2‐Mut) and patients without were defined as PAPPA2‐wildtype (PAPPA2‐WT).
2.5. TMB data analysis
TMB for immunotherapy cohorts and TCGA datasets was defined as the total number of nonsynonymous somatic mutations and the total number of nonsynonymous somatic mutations per megabase of genome examined, respectively. The cutoff value for high and low TMB in this study was the top 25% TMB within each set. 35 , 36
2.6. DDR pathways and gene sets
The core genes associated with DDR pathways were obtained from Knijnenburg et al. 37 The details of DDR core genes are shown in Table S2. The DDR gene sets were obtained from the Reactome Knowledgebase (https://reactome.org). 38 The details of DDR gene sets are shown in Table S3.
2.7. Gene set enrichment analysis
R package DESeq2 was conducted for differential gene expression analysis. 39 Reactome pathways analysis based on Gene set enrichment analysis (GSEA) was performed by the R package ClusterProfiler. 40 Gene sets with an adjusted p value (Benjamini‐Hochberg method) lower than 0.05 were considered significantly enriched.
2.8. Statistical analysis
The differences in TMB, NAL, tumour‐infiltrating leukocytes and gene expressions between PAPPA2‐Mut and PAPPA2‐WT groups were examined using the Mann–Whitney U test. Comparisons of ORR, DCR, DCB, and PD‐L1 expression in different sets were conducted with Fisher's exact test. PFS and OS were estimated by Kaplan–Meier method, with the p value determined by a log‐rank test. The Cox regression was applied for univariable and multivariable survival analyses. Variables with p < 0.05 in the univariable regression and those which have been reported associated with the effect of ICIs were also included in multivariable Cox regression. All the statistical analyses were performed using R version 4.1.1 (https://www.r-project.org). All reported p values were two‐tailed, and p < 0.05 was considered statistically significant.
3. RESULTS
3.1. Association between PAPPA2 mutation and the clinical benefit of ICIs therapy in the NSCLC set
The flow diagram of this study is depicted in Figure 1. In this study, we observed that PAPPA2 mutation was mostly enriched in patients with NSCLC and SKCM while strongly differenced in patients with objective response to ICIs versus without in the NSCLC and SKCM sets. To identify whether PAPPA2 mutation was associated with the response to ICIs therapy, we integrated the mutational and clinical data of three NSCLC cohorts (Rizvi, Hellmann and Miao cohorts) and four SKCM cohorts (Synder, Allen, Riaz cohort and Hugo cohorts) to form the NSCLC and SKCM set, respectively. Tables S4 and S5 summarized the clinical characteristics of patients in the NSCLC set and the SKCM set, respectively.
FIGURE 1.
Flow diagram of the study. (A) Preliminary analysis. PAPPA2 mutated most frequently in SKCM and NSCLC in the TCGA database. PAPPA2 mutational rates in patients with objective response (CR + PR) versus without (SD + PD) were compared with other ICIs‐related gene mutations in the NSCLC and SKCM sets. (B) Biomarker development. Association between PAPPA2 mutation and clinical outcomes has been analysed in the NSCLC set, the SKCM set and China cohort. (C) Mechanism exploring. Based on the TCGA database, the correlations of PAPPA2 mutation with TMB, infiltrating immune cells and DDR were explored for further immunogenicity and anti‐tumour activity mechanisms. DDR, DNA damage repair; NSCLC, non‐small cell lung cancer; SKCM, skin cutaneous melanoma; TCGA, The Cancer Genome Atlas; TMB, tumour mutation burden
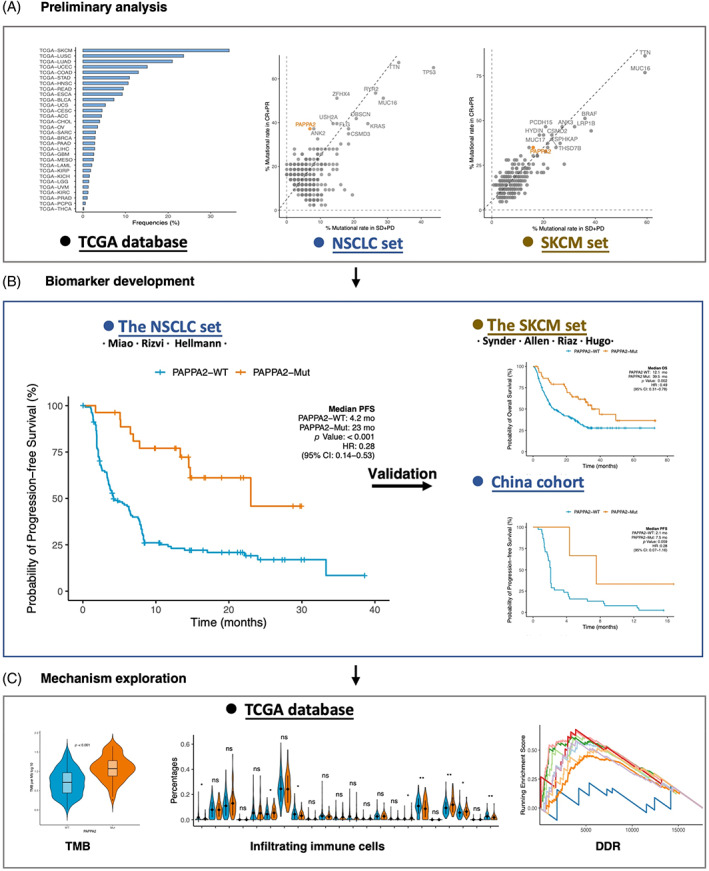
Among all candidates, PAPPA2 mutation was discovered to be enriched in patients with objective response (39.6%) versus without (6.3%) in the NSCLC set (Figure 2A). As expected, compared to that in the PAPPA2‐WT group, longer PFS was observed in the PAPPA2‐Mut group (hazard ratio [HR], 0.26 [95% CI: 0.14–0.53]; p < 0.001, Figure 2B). Patients harbouring PAPPA2‐Mut had a higher ORR (77.8% vs. 23.2%; p < 0.001; Figure 2C) and a higher DCR (100.0% vs. 60.9%; p < 0.001; Figure 2C). Meanwhile, a higher DCB (85.2% vs. 39.9%; p < 0.001; Figure 2D) in PAPPA2‐Mut compared with those PAPPA2‐WT was confirmed. The result of prolonged PFS in PAPPA2‐Mut patients was consistently observed across all three cohorts (Rizvi, Hellmann, and Miao cohorts; Figure 2E). The favourable survival for PAPPA2‐Mut was also verified after considering confounding factors (Table 1, multivariable analysis, HR, 0.28 [95% CI, 0.14–0.53], p < 0.001). These results suggested that PAPPA2 mutation was associated with the clinical benefit of immunotherapy.
FIGURE 2.
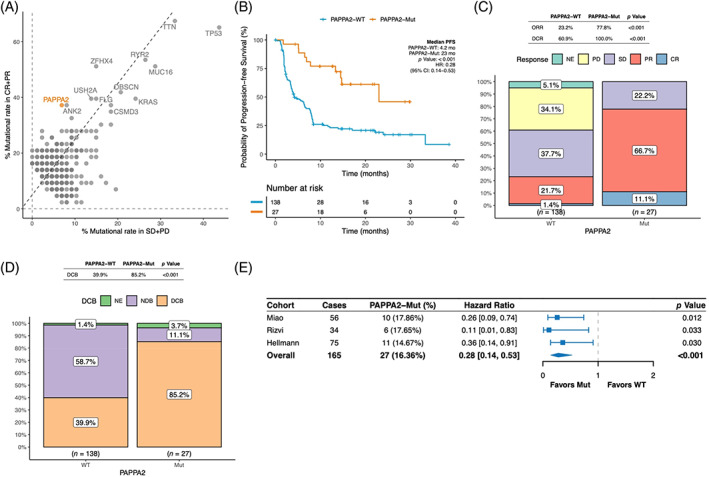
Association between PAPPA2 mutation and clinical benefits of ICIs therapy in the NSCLC set. (A) A scatter diagram displaying the mutational rate of different ICIs‐related gene mutations in patients with objective response (CR + PR) versus without (SD + PD) in the NSCLC set. PAPPA2 mutation (39.6% vs. 6.3%) is highlighted in orange. (B) Longer PFS observed in the PAPPA2‐Mut group compared to the PAPPA2‐MT group in the NSCLC set. (C) The response data on ORR and DCR of patients evaluated in the NSCLC set. (D) The response data on DCB of patients evaluated in the NSCLC set. (E) The prolonged PFS in the PAPPA2‐Mut group consistently observed among three cohorts included in the NSCLC set. DCB, durable clinical benefit; DCR, disease control rate; ICIs, immune checkpoint inhibitors; NSCLC, non‐small cell lung cancer; ORR, objective response rate; PFS, progress free survival
TABLE 1.
Univariable and multivariable analyses of PFS in the NSCLC set
| Characteristic | Univariable | Multivariable | ||||
|---|---|---|---|---|---|---|
| HR | 95% CI | p value | HR | 95% CI | p value | |
| Cohort | ||||||
| Hellmann | ‐ | ‐ | ‐ | ‐ | ||
| Miao | 1.28 | 0.85, 1.93 | 0.2 | |||
| Rizvi | 1.08 | 0.65, 1.78 | 0.8 | |||
| Gender | ||||||
| Female | ‐ | ‐ | ‐ | ‐ | ||
| Male | 1.26 | 0.87, 1.82 | 0.2 | |||
| Age | ||||||
| <65 | ‐ | ‐ | ‐ | ‐ | ||
| ≥65 | 0.91 | 0.61, 1.35 | 0.6 | |||
| Unknown | 1.58 | 0.89, 2.82 | 0.12 | |||
| Smoking | ||||||
| Never | ‐ | ‐ | ‐ | ‐ | ||
| Current | 0.69 | 0.38, 1.24 | 0.2 | 1.07 | 0.57, 2.00 | 0.8 |
| Former | 0.58 | 0.37, 0.92 | 0.019 | 0.83 | 0.52, 1.33 | 0.4 |
| Histology | ||||||
| Non‐squamous | ‐ | ‐ | ‐ | ‐ | ||
| Squamous | 0.92 | 0.55, 1.54 | 0.7 | |||
| NSCLC NOS | 0.36 | 0.05, 2.56 | 0.3 | |||
| Treatment | ||||||
| Anti‐PD‐(L)1 | ‐ | ‐ | ‐ | ‐ | ||
| Anti‐PD‐(L)1 + Anti‐CTLA4 | 0.83 | 0.57, 1.21 | 0.3 | |||
| Line | ||||||
| First | ‐ | ‐ | ‐ | ‐ | ||
| Second or subsequent | 1.16 | 0.66, 2.02 | 0.6 | |||
| Unknown | 1.29 | 0.86, 1.93 | 0.2 | |||
| PDL1 | ||||||
| <1% | ‐ | ‐ | ‐ | ‐ | ||
| 1%–49% | 0.91 | 0.53, 1.56 | 0.7 | 0.85 | 0.48, 1.50 | 0.6 |
| ≥50% | 0.38 | 0.17, 0.86 | 0.02 | 0.38 | 0.16, 0.85 | 0.019 |
| Unknown | 0.98 | 0.59, 1.63 | >0.9 | 0.85 | 0.51, 1.43 | 0.5 |
| TMB | ||||||
| Others | ‐ | ‐ | ‐ | ‐ | ||
| Top 25% | 0.35 | 0.22, 0.58 | <0.001 | 0.47 | 0.27, 0.80 | 0.005 |
| PAPPA2 | ||||||
| WT | ‐ | ‐ | ‐ | ‐ | ||
| Mut | 0.28 | 0.14, 0.53 | <0.001 | 0.37 | 0.18, 0.78 | 0.009 |
Abbreviations: CI, confidence interval; HR, hazard ratio; NSCLC, non‐small cell lung cancer; PFS, progress free survival; TMB, tumour mutational burden.
Factors of TMB (HR, 0.35 [95% CI, 0.22–0.58]; p < 0.001; Table 1) and PD‐L1 (≥50%; HR, 0.38 [95% CI, 0.17–0.86]; p = 0.02; Table 1) were also significantly associated with superior PFS. In 100 patients with known PD‐L1 status, 35.3% and 16.9% had PD‐L1 ≥50% in patients with PAPPA2‐Mut and PAPPA2‐WT, respectively (Figure S1A). Patients with positive PD‐L1 expression (≥1%) and PAPPA2‐Mut tended to have a superior PFS (HR, 0.11 [95% CI, 0.02–0.46]; p = 0.003; Figure S1B).
3.2. Association between PAPPA2 mutation and clinical benefits of ICIs therapy in the SKCM set
In the SKCM set, PAPPA2 mutation was also discovered to be enriched in patients with objective response (34.9%) versus without (16%; Figure 3A). We further validated the association between PAPPA2 mutation and clinical benefits. Expectedly, patients with SKCM harbouring PAPPA2 mutation had superior OS (HR, 0.49 [95% CI: 0.31–0.78]; p = 0.002; Figure 3B), higher ORR (34.1% vs. 16.9%; p = 0.039; Figure 3C) and higher DCB + LB (50.0% vs. 30.7%; p = 0.036; Figure 3D) compared with those with PAPPA2‐WT group. The prolonged OS in PAPPA2 mutation patients with SKCM was consistently observed across four datasets included in the validation set (Figure 3E). The prolonged OS in PAPPA2‐Mut patients was also consistently observed in multivariable analysis (HR, 0.54 [95% CI, 0.32–0.91]; p = 0.021; Table 2). These results suggested that PAPPA2 mutation was associated with the clinical benefit of immunotherapy.
FIGURE 3.
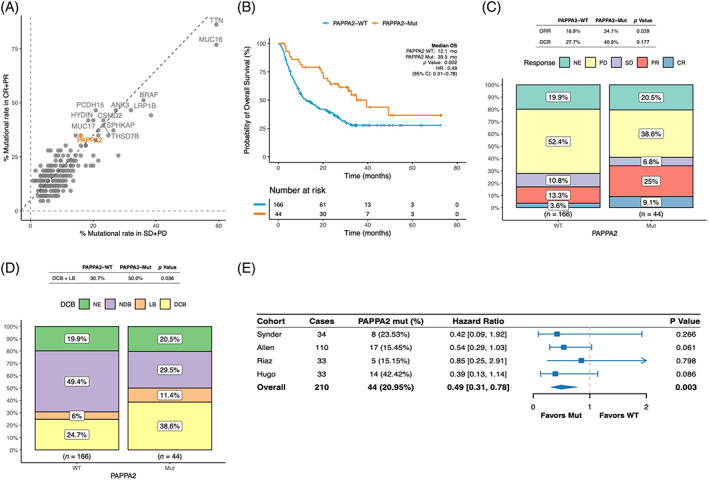
Association between PAPPA2 mutation and clinical benefits of ICIs therapy in the SKCM set. (A) A scatter diagram displaying the mutational rate of different ICIs‐related gene mutations in patients with objective response (CR + PR) versus without (SD + PD) in the SKCM set, with PAPPA2 mutation(34.9% vs. 16%) highlighted in orange. (B) Longer OS observed in the PAPPA2‐Mut group compared to the PAPPA2‐MT group in the SKCM set. (C) The response data on ORR and DCR of patients evaluated in the SKCM set. (D) The response data on DCB + LB of patients evaluated in the SKCM set. (E) The trend of prolonged OS in the PAPPA2‐Mut group consistently observed among four cohorts included in the SKCM set. DCB, durable clinical benefit; DCR, disease control rate; ICIs, immune checkpoint inhibitors; ORR, objective response rate; PFS, progress free survival; SKCM, skin cutaneous melanoma
TABLE 2.
Univariable and multivariable analyses of overall survival in the SKCM set
| Characteristic | Univariable | Multivariable | ||||
|---|---|---|---|---|---|---|
| HR | 95% CI | p value | HR | 95% CI | p value | |
| Cohort | ||||||
| Riaz | ‐ | ‐ | ||||
| Synder | 0.51 | 0.25, 1.03 | 0.06 | |||
| Allen | 1.58 | 0.97, 2.58 | 0.067 | |||
| Hugo | 0.75 | 0.39, 1.45 | 0.4 | |||
| Gender | ||||||
| Female | ‐ | ‐ | ||||
| Male | 0.87 | 0.58, 1.29 | 0.5 | |||
| Unknown | 0.8 | 0.46, 1.39 | 0.4 | |||
| Age | ||||||
| <65 | ‐ | ‐ | ||||
| ≥65 | 1.1 | 0.76, 1.59 | 0.6 | |||
| Unknown | 0.92 | 0.56, 1.54 | 0.8 | |||
| M class | ||||||
| M0 | ‐ | ‐ | ‐ | ‐ | ||
| M1a | 1.72 | 0.55, 5.40 | 0.4 | 1.81 | 0.58, 5.69 | 0.3 |
| M1b | 2.39 | 0.81, 7.08 | 0.11 | 2.6 | 0.88, 7.69 | 0.084 |
| M1c | 3.71 | 1.36, 10.1 | 0.01 | 4.39 | 1.61, 12.0 | 0.004 |
| Unknown | 1.57 | 0.39, 6.27 | 0.5 | 1.53 | 0.38, 6.14 | 0.5 |
| Treatment | ||||||
| Anti‐CTLA‐4 | ‐ | ‐ | ||||
| Anti‐PD‐1 | 0.71 | 0.48, 1.04 | 0.075 | |||
| Line | ||||||
| First | ‐ | ‐ | ||||
| Second | 0.62 | 0.34, 1.15 | 0.13 | |||
| Third or subsequent | 0 | 0.00, Inf | >0.9 | |||
| Unknown | 0 | 0.00, Inf | >0.9 | |||
| TMB | ||||||
| Others | ‐ | ‐ | ‐ | ‐ | ||
| Top 25% | 0.56 | 0.36, 0.87 | 0.01 | 0.62 | 0.38, 1.02 | 0.06 |
| PAPPA2 | ||||||
| WT | ‐ | ‐ | ‐ | ‐ | ||
| Mut | 0.49 | 0.31, 0.78 | 0.003 | 0.54 | 0.32, 0.91 | 0.021 |
Abbreviations: CI, confidence interval; HR, hazard ratio; SKCM, skin cutaneous melanoma; TMB, tumour mutational burden.
3.3. The association between PAPPA2 mutation and TMB
Based on the TCGA database, findings in the TCGA‐NSCLC dataset showed that the PAPPA2‐Mut group had higher TMB (p < 0.001) and NAL (p < 0.001) levels than the PAPPA2‐WT group (Figure 4A,B), consistent in the TCGA‐SKCM dataset (TMB, p < 0.001; NAL, p = 0.048; Figure 4C,D).
FIGURE 4.
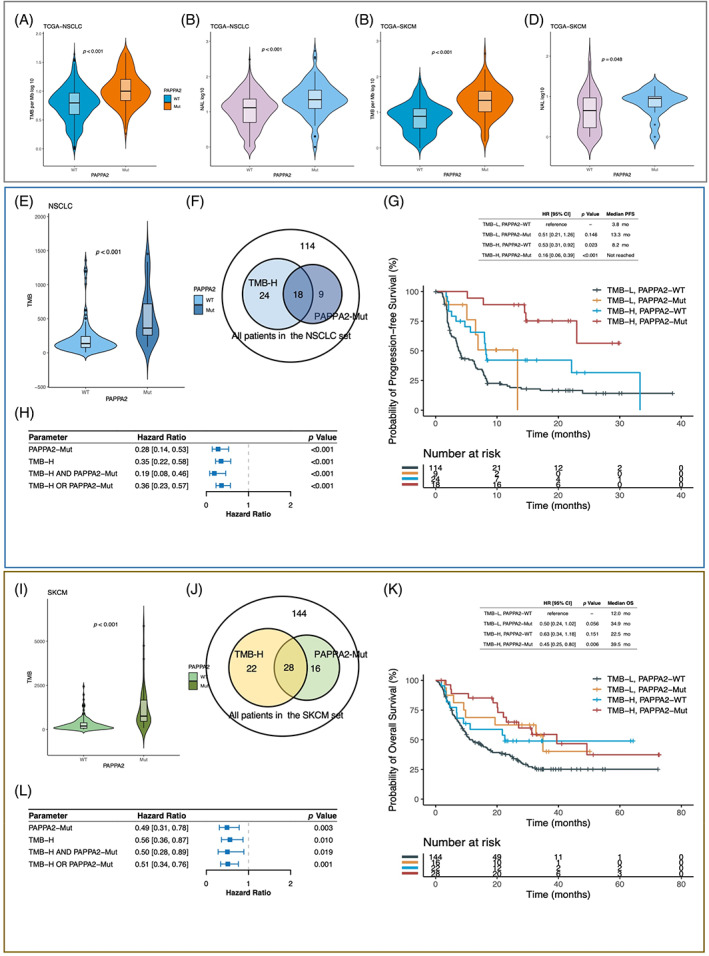
The association between PAPPA2 mutation and TMB. (A–D) A comparison of TMB and NAL between PAPPA2‐Mut and PAPPA2‐WT groups in NSCLC and SKCM based on the TCGA database. (E) A comparison of TMB between PAPPA2‐Mut and PAPPA2‐WT groups in the NSCLC set. (F) A Venn diagram showing the concomitant presence of TMB‐H and PAPPA2‐Mut in the NSCLC set. (G) The HRs and p values of parameters including TMB‐H, PAPPA2‐Mut, TMB‐H and PAPPA2‐Mut, and TMB‐H or PAPPA2‐Mut in the NSCLC set. (H) Kaplan–Meier curves comparing PFS among TMB‐H and PAPPA2‐Mut, TMB‐H and PAPPA2‐WT, TMB‐L and PAPPA2‐Mut, with TMB‐L and PAPPA2‐WT as a reference. (I) A comparison of TMB between PAPPA2‐Mut and PAPPA2‐WT groups in the SKCM set. (J) A Venn diagram showing the concomitant presence of TMB‐H and PAPPA2‐Mut in the SKCM set. (K) The HRs and p values of parameters including TMB‐H, PAPPA2‐Mut, TMB‐H and PAPPA2‐Mut, TMB‐H or PAPPA2‐Mut. (L) The Kaplan–Meier curves comparing OS among TMB‐H & PAPPA2‐Mut, TMB‐H & PAPPA2‐WT, TMB‐L & PAPPA2‐Mut with TMB‐L and PAPPA2‐WT as a reference. NSCLC, non‐small cell lung cancer; SKCM, skin cutaneous melanoma; TMB, tumour mutational burden
For the NSCLC set, the PAPPA2‐Mut group had significantly higher TMB (p < 0.001) than the PAPPA2‐WT group (Figure 4E). In addition, a significant longer PFS was observed in patients with TMB‐H and PAPPA2‐Mut compared to those with TMB‐L and PAPPA2‐WT (HR, 0.16 [95% CI, 0.06–0.39]; p < 0.001; Figure 4F–G). Among all patients in the NSCLC set, those with TMB‐H or PAPPA2‐Mut (53/165) achieved significantly longer PFS (HR, 0.36 [95% CI, 0.23–0.57]; p < 0.001; Figure 4H) than counterparts. Similarly, the TMB level was significantly higher in PAPPA2‐Mut tumours in the SKCM set (p < 0.001, Figure 4I). In addition, a significant longer PFS was observed in patients with TMB‐H and PAPPA2‐Mut compared to those with TMB‐L and PAPPA2‐WT (HR, 0.45 [95% CI, 0.25–0.80]; p = 0.006; Figure 4I–K). Among all patients in the SKCM set, those with TMB‐H or PAPPA2‐Mut (66/210) achieved significantly longer OS (HR, 0.51 [95% CI, 0.34–0.76]; p < 0.001; Figure 4L). Results above revealed that combined utilization of PAPPA2 mutation and TMB could expand the identification of ICIs therapy response in both NSCLC and SKCM patients.
3.4. Association between PAPPA2 mutation and clinical benefits of ICIs in China cohort
In China cohort, which comprised 41 Chinese patients with NSCLC, we further investigated the association between PAPPA2 mutation and clinical benefits. The trend of prolonged PFS in PAPPA2‐Mut was observed (HR, 0.28 [95% CI, 0.07–1.16]; p = 0.059; Figure S2A). Meanwhile, patients with PAPPA2 mutation had a higher ORR (66.7% vs. 13.2%; p = 0.070; Figure S2B) and DCB (66.7% vs. 22.1%; p = 0.142; Figure S2C) compared with those in the PAPPA2‐WT group. Additionally, the TMB level in PAPPA2‐Mut was significantly higher compared to that in PAPPA2‐WT (p = 0.015, Figure S2D). Table S6 summarized the clinical characteristics of patients in China cohort.
3.5. Comparison to known predictive gene mutations of ICIs benefit
Among PAPPA2 and other established predictive gene mutations, mutations of PAPPA2, EPHA family, MUC16, LRP1B and TTN brought superior prediction of ICIs benefit in both sets by univariable analysis (Table 3). PAPPA2‐Mut presented the lowest risk of progression or death in both sets (NSCLC: HR, 0.28 [95% CI, 0.04–0.53]; p < 0.001; SKCM set: HR, 0.49 [95% CI, 0.31–0.78]; p = 0.003). These results confirmed the remarkable prediction value of PAPPA2 mutation in ICIs benefit.
TABLE 3.
Compared with other known predictive gene mutations with univariable Cox analysis
| Gene | NSCLC | SKCM | ||||
|---|---|---|---|---|---|---|
| Mutation (%) | Hazard ratio | p value | Mutation (%) | Hazard ratio | p value | |
| ARID1A | 6.67 | 0.53 [0.22, 1.30] | 0.167 | 6.67 | 0.69 [0.34, 1.42] | 0.313 |
| CDKN2A | 6.67 | 0.38 [0.14, 1.04] | 0.060 | 5.08 | 0.65 [0.27, 1.60] | 0.348 |
| EGFR | 15.76 | 2.03 [1.28, 3.23] | 0.003 | 5.08 | 0.84 [0.39, 1.81] | 0.656 |
| EPHA_family | 19.39 | 0.46 [0.28, 0.78] | 0.004 | 39.05 | 0.54 [0.37, 0.78] | 0.001 |
| KEAP1 | 16.97 | 0.65 [0.37, 1.13] | 0.126 | 1.82 | 0.73 [0.18, 3.01] | 0.662 |
| KMT2_family | 15.76 | 0.56 [0.32, 0.98] | 0.041 | 28.57 | 0.68 [0.46, 1.01] | 0.058 |
| KRAS | 28.48 | 0.61 [0.40, 0.94] | 0.026 | 3.47 | 0.46 [0.11, 1.88] | 0.281 |
| LRP1B | 26.06 | 0.53 [0.33, 0.84] | 0.007 | 36.19 | 0.55 [0.37, 0.80] | 0.002 |
| MUC16 | 35.15 | 0.53 [0.35, 0.79] | 0.002 | 64.29 | 0.63 [0.44, 0.89] | 0.009 |
| NOTCH_family | 10.30 | 0.82 [0.45, 1.50] | 0.521 | 24.29 | 0.66 [0.44, 1.01] | 0.053 |
| PAPPA2 | 16.36 | 0.28 [0.14, 0.53] | <0.001 | 20.95 | 0.49 [0.31, 0.78] | 0.003 |
| POLD1 | 7.34 | 0.50 [0.18, 1.36] | 0.174 | 3.33 | 0.58 [0.21, 1.56] | 0.279 |
| POLE | 1.82 | 0.00 [0.00, Inf] | 0.995 | 3.95 | 0.63 [0.23, 1.72] | 0.368 |
| STK11 | 12.12 | 1.67 [0.96, 2.88] | 0.068 | 0.70 | 2.40 [0.33, 17.42] | 0.385 |
| TP53 | 52.73 | 0.73 [0.50, 1.05] | 0.092 | 11.43 | 1.58 [0.95, 2.64] | 0.077 |
| TTN | 44.85 | 0.43 [0.29, 0.63] | <0.001 | 66.67 | 0.66 [0.46, 0.94] | 0.021 |
Abbreviations: NSCLC, non‐small cell lung cancer; SKCM, skin cutaneous melanoma.
3.6. PAPPA2 mutation was not a prognostic factor
To evaluate whether the survival benefit of ICIs therapy in patients with PAPPA2‐Mut was simply resulted from the general prognostic impact of PAPPA2 mutation, we further assessed the PFS and OS difference between PAPPA2‐Mut and PAPPA2‐WT patients with NSCLC or SKCM in TCGA database (Figure S3). Obviously, there was no PFS or OS difference owing to PAPPA2 mutation in lung adenocarcinoma (LUAD), lung squamous carcinoma (LUSC), NSCLC or SKCM. Therefore, PAPPA2 mutation may be a predictive but not a prognostic factor in ICIs treatment for patients with NSCLC as well as SKCM.
3.7. Potential mechanisms associated with PAPPA2 mutation in anti‐tumour immunity
To investigate the potential mechanisms associated with PAPPA2 mutation, we used the CIBERSORT algorithm to estimate the immune cell infiltration status based on the TCGA database. A comparing analysis in both TCGA‐LUAD and TCGA‐SKCM datasets showed that the PAPPA2‐Mut group had revealed higher activated CD4 memory T cells and lower Treg cells than PAPPA2‐WT tumours (Figure 5A,B).
FIGURE 5.
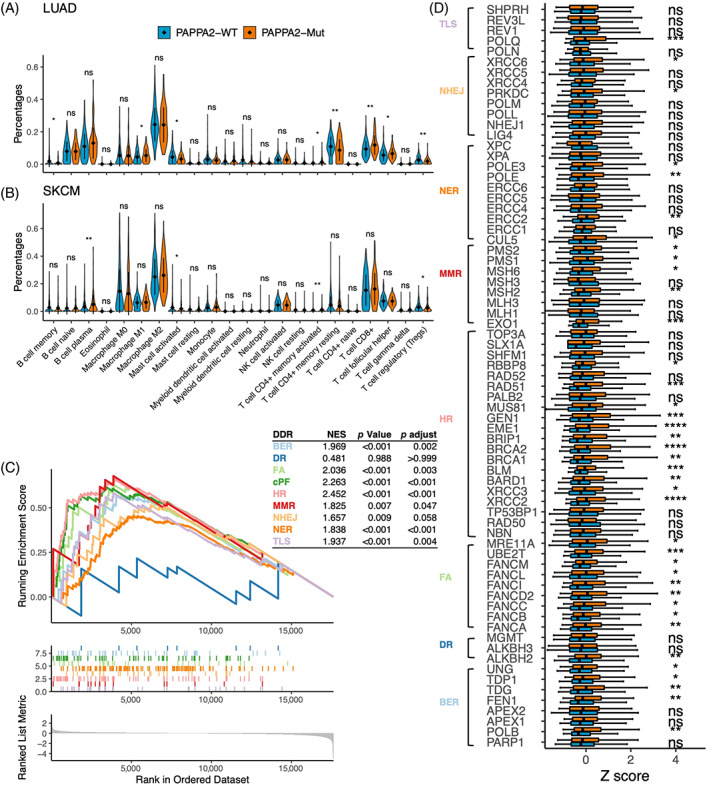
PAPPA2 mutation was associated with enhanced anti‐tumour immunity in the TCGA database. (A,B) Violin plots depicting the infiltration of immune cells in PAPPA2‐Mut tumours and PAPPA2‐WT tumours in TCGA‐LUAD and TCGA‐SKCM. (C) The enrichment of DDR pathways between PAPPA2‐Mut and PAPPA2‐WT groups in TCGA‐LUAD. (D) Box plots comparing the expression of DDR‐related genes between PAPPA2‐Mut and PAPPA2‐WT groups in TCGA‐LUAD (*p < 0.05, **p < 0.01, ***p < 0.001)
DDR signalling pathways and related genes based on RNA‐Seq data from the TCGA database were analysed. GSEA of Reactome revealed that gene sets related to the DDR pathways (the nonhomologous end‐joining (NHEJ) pathway, homologous recombination repair (HR) pathway, etc.) were significantly enriched in PAPPA2‐Mut tumours (p < 0.001, Figure 5C) in TCGA‐LUAD. Moreover, TCGA‐LUAD tumours with PAPPA2 mutation had increased mRNA expression of DDR‐related genes (Figure 5D).
4. DISCUSSION
The application of advanced technologies and bioinformatic tools has enabled us to investigate the role of certain factors in various diseases. 41 , 42 Likewise, in this study for preliminary analysis, we observed an enrichment of PAPPA2 mutation in NSCLC and SKCM, as well as a discrepancy in patients with objective response versus without. In addition, independent of PD‐L1 expression or TMB status, PAPPA2 mutation displayed a strong association with better clinical outcomes in patients with NSCLC and SKCM after receiving ICIs therapy. However, the correlation between PAPPA2 mutation and prognosis has not drawn much attention even though Ayako Suzuki et al identified PAPPA2 mutation as prolonged‐prognosis‐related gene of LUAD in 2013. 25 So far as we know, this is the first study to elucidate the role of PAPPA2 mutation in stratifying the efficacy of ICIs therapy.
Currently, PD‐L1 expression and TMB status were the most utilized predictive biomarkers of ICIs. However, the prediction value could be affected by various factors, such as different cut‐off points, calculating algorithms and detecting assays, etc. 43 , 44 Hence, limitations remained in practice due to inconsistency and heterogeneity. 12 , 45 Our results revealed the improved survival in patients with PAPPA2 mutation, suggesting PAPPA2 mutation was a potential predictive biomarker of ICIs, complementing to PD‐L1 and TMB. As a single gene biomarker, the qualitative detection of PAPPA2 mutation made it objective to identify potential beneficiaries. Noteworthy, a comparison of PAPPA2 mutation with other published ICIs‐related gene mutations showed a prominently potential predicting ability, such as EPHA family, MUC16, LRP1B and TTN, etc. 16 , 18 , 46 , 47 However, it also revealed the instability of single genes to some extent. Intriguingly, patients revealed the prediction instability or TMB‐H demonstrated a beneficial significance in both sets, suggesting a combination of PAPPA2 mutation and TMB could expand the identification of potential responders to ICIs therapy.
We utilized a multidimensional TCGA database to explore how PAPPA2‐Mut tumours respond to immunotherapy. PAPPA2‐Mut tumours were discovered with higher levels of TMB and NAL, which are associated with increased tumour immunogenicity. Then RNA‐Seq data revealed that PAPPA2 mutation was significantly associated with higher activated CD4 memory T cells and lower Treg cells, suggesting an enhanced anti‐tumour immunity. 48 , 49 Moreover, PAPPA2‐Mut tumours were enriched with multiple DDR pathways, which are associated with the efficacy of ICIs treatment in tumours. 50 Alterations of DDR‐related genes are closely correlated with higher TMB. 51 Hence, PAPPA2‐Mut tumours were correlated with increased immunogenicity and enhanced anti‐tumour immunity, implying a better performance in ICIs therapy.
Our study has several limitations. First, to our knowledge, PAPPA2 mutation has not been added to existing targeted gene panels. The retrospective study design and limited public data might introduce information bias. To minimize it, we extracted common clinical characteristics across data sets for NSCLC and SKCM respectively, and unified the category definition and processing algorithms. We also conducted multivariable models and consistent results were found. Secondly, interestingly enough, in the TCGA database, we did not find the PFS/OS difference between PAPPA2‐Mut and PAPPA2‐WT neither in LUAD, LUSC, NSCLC or SKCM, suggesting it was not a prognosis biomarker, contrary to its role in Ayako Suzuki cohort (7/90, about 7.7%) as a prognosis biomarker. 25 In addition, though varying among cohorts, PAPPA2 mutation rates in the NSCLC set (14.67%–17.86%) are much higher than Asian cohorts, like Ayako Suzuki et al cohort (Japanese, 7.7%) and China cohort (Chinese, 7.1%). It seems that PAPPA2 mutation rates and corresponding functions vary by race. Hence, several parameters may contribute to the insignificance of PAPPA2 mutation in this relatively small‐sized China cohort, though the tendency supported the prediction value of PAPPA2 mutation to some extent. All in all, larger prospective clinical trials with multidimensional data and mechanism‐exploring experiments are needed to clarify and validate the predictive capacity and functional alterations of PAPPA2 mutation.
5. CONCLUSION
In summary, our study explored the association between PAPPA2 mutation and the clinical benefit of ICIs therapy in NSCLC and SKCM. Our results demonstrated that patients with PAPPA2 mutation were associated with better clinical outcomes in ICIs treatment via activated immunogenicity and enhanced anti‐tumour immunity. Thus, PAPPA2 mutation could act as a potential predictive biomarker for ICIs therapy in NSCLC and SKCM, warranting further prospective studies.
AUTHOR CONTRIBUTIONS
Zhijie Wang and Jie Wang developed the concept and designed the study. Lele Zhao, Jianchun Duan, Qin Zhang and Hua Bai performed the statistical analysis. Yiting Dong, Yangyang Yu, Lele Zhao, Dongsheng Chen and Si Li wrote the manuscript. Mingzhe Xiao, Qianqian Duan, Jie Wang, Zhijie Wang, Tingting Sun and Chuang Qi performed a critical revision of the manuscript for the intellectual content. All authors read and approved the final manuscript.
FUNDING INFORMATION
Support for the study was provided by CAMS Innovation Fund for Medical Sciences (No. 2021‐1‐I2M‐012 to Dr Zhijie Wang); CAMS Key lab of translational research on lung cancer (No. 2018PT31035 to Dr Jie Wang), the National Natural Sciences Foundation (Nos. 81871889 and 82072586 to Dr Zhijie Wang) and Beijing Natural Science Foundation (No. 7212084 to Dr Zhijie Wang).
CONFLICT OF INTEREST
Lele Zhao, Dongsheng Chen, Si Li, Yangyang Yu, Mingzhe Xiao, Qin Zhang, Qianqian Duan, Tingting Sun and Chuang Qi were employed by Jiangsu Simcere Diagnostics Co., Ltd. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supporting information
Appendix S1 Supporting information.
ACKNOWLEDGEMENTS
The authors would like to thank the staff members of the TCGA Research Network, the cBioportal data portal, the GDC data portal as well as all the authors for making their valuable research data public. We would like to thank all patients, all colleagues, doctors and nurses of the department of NCC and SYUCC for this work.
Dong Y, Zhao L, Duan J, et al. PAPPA2 mutation as a novel indicator stratifying beneficiaries of immune checkpoint inhibitors in skin cutaneous melanoma and non‐small cell lung cancer. Cell Prolif. 2022;55(9):e13283. doi: 10.1111/cpr.13283
Yiting Dong and Lele Zhao contributed equally to this study.
Funding information Beijing Natural Science Foundation, Grant/Award Number: 7212084; CAMS Innovation Fund for Medical Sciences, Grant/Award Number: 2021‐1‐I2M‐012; CAMS Key lab of translational research on lung cancer, Grant/Award Number: 2018PT31035; National Natural Sciences Foundation, Grant/Award Numbers: 81871889, 82072586
Contributor Information
Jie Wang, Email: zlhuxi@163.com.
Zhijie Wang, Email: jie_969@163.com.
DATA AVAILABILITY STATEMENT
The public cohorts used in this study were publicly available as described in the ‘Materials and methods’ section. The China cohort is available from the corresponding author on reasonable request.
REFERENCES
- 1. Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209‐249. [DOI] [PubMed] [Google Scholar]
- 2. Ma W, Yang Y, Zhu J, et al. Biomimetic Nanoerythrosome‐coated aptamer‐DNA tetrahedron/Maytansine conjugates: pH‐responsive and targeted cytotoxicity for HER2‐positive breast cancer. Adv Mater. 2022;e2109609. [DOI] [PubMed] [Google Scholar]
- 3. Zhang T, Tian T, Lin Y. Functionalizing framework nucleic‐acid‐based nanostructures for biomedical application. Adv Mater. 2021;e2107820. [DOI] [PubMed] [Google Scholar]
- 4. Zhang B, Tian T, Xiao D, Gao S, Cai X, Lin Y. Facilitating in situ tumor imaging with a tetrahedral DNA framework‐enhanced hybridization chain reaction probe. Adv Funct Mater. 2022;32(16):2109728. [Google Scholar]
- 5. Zhou M, Zhang T, Zhang B, et al. A DNA nanostructure‐based Neuroprotectant against neuronal apoptosis via inhibiting toll‐like receptor 2 signaling pathway in acute ischemic stroke. ACS Nano. 2021;16:1456‐1470. [DOI] [PubMed] [Google Scholar]
- 6. Li J, Lai Y, Li M, et al. Repair of infected bone defect with clindamycin‐tetrahedral DNA nanostructure complex‐loaded 3D bioprinted hybrid scaffold. Chem Eng J. 2022;134855:134855. [Google Scholar]
- 7. Doroshow DB, Sanmamed MF, Hastings K, et al. Immunotherapy in non‐small cell lung cancer: facts and hopes. Clin Cancer Res. 2019;25(15):4592‐4602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Jenkins RW, Fisher DE. Treatment of advanced melanoma in 2020 and beyond. J Invest Dermatol. 2021;141(1):23‐31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Pu X, Wu L, Su D, Mao W, Fang B. Immunotherapy for non‐small cell lung cancers: biomarkers for predicting responses and strategies to overcome resistance. BMC Cancer. 2018;18(1):1082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Yarchoan M, Hopkins A, Jaffee EM. Tumor mutational burden and response rate to PD‐1 inhibition. N Engl J Med. 2017;377(25):2500‐2501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Hegde PS, Chen DS. Top 10 challenges in cancer immunotherapy. Immunity. 2020;52(1):17‐35. [DOI] [PubMed] [Google Scholar]
- 12. Havel JJ, Chowell D, Chan TA. The evolving landscape of biomarkers for checkpoint inhibitor immunotherapy. Nat Rev Cancer. 2019;19(3):133‐150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Ott PA, Bang YJ, Piha‐Paul SA, et al. T‐cell‐inflamed gene‐expression profile, programmed death ligand 1 expression, and tumor mutational burden predict efficacy in patients treated with Pembrolizumab across 20 cancers: KEYNOTE‐028. J Clin Oncol. 2019;37(4):318‐327. [DOI] [PubMed] [Google Scholar]
- 14. Ready N, Hellmann MD, Awad MM, et al. First‐line nivolumab plus ipilimumab in advanced non‐small‐cell lung cancer (CheckMate 568): outcomes by programmed death ligand 1 and tumor mutational burden as biomarkers. J Clin Oncol. 2019;37(12):992‐1000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Marabelle A, Le DT, Ascierto PA, et al. Efficacy of pembrolizumab in patients with noncolorectal high microsatellite instability/mismatch repair‐deficient cancer: results from the phase II KEYNOTE‐158 study. J Clin Oncol. 2020;38(1):1‐10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Bai H, Duan J, Li C, et al. EPHA mutation as a predictor of immunotherapeutic efficacy in lung adenocarcinoma EPHA mutation as a predictor of immunotherapeutic efficacy in lung adenocarcinoma. J Immunother Cancer. 2020;8(2):e001315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Long J, Wang D, Yang X, et al. Identification of NOTCH4 mutation as a response biomarker for immune checkpoint inhibitor therapy. BMC Med. 2021;19(1):154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Brown LC, Tucker MD, Sedhom R, et al. LRP1B mutations are associated with favorable outcomes to immune checkpoint inhibitors across multiple cancer types. J Immunother Cancer. 2021;9(3):e001792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Overgaard MT, Boldt HB, Laursen LS, Sottrup‐Jensen L, Conover CA, Oxvig C. Pregnancy‐associated plasma protein‐A2 (PAPP‐A2), a novel insulin‐like growth factor‐binding protein‐5 proteinase. J Biol Chem. 2001;276(24):21849‐21853. [DOI] [PubMed] [Google Scholar]
- 20. Fujimoto M, Hwa V, Dauber A. Novel modulators of the growth hormone ‐ insulin‐like growth factor Axis: pregnancy‐associated plasma protein‐A2 and Stanniocalcin‐2. J Clin Res Pediatr Endocrinol. 2017;9(Suppl 2):1‐8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Yang S, Chintapalli J, Sodagum L, et al. Activated IGF‐1R inhibits hyperglycemia‐induced DNA damage and promotes DNA repair by homologous recombination. Am J Physiol Renal Physiol. 2005;289(5):F1144‐F1152. [DOI] [PubMed] [Google Scholar]
- 22. Dauber A, Munoz‐Calvo MT, Barrios V, et al. Mutations in pregnancy‐associated plasma protein A2 cause short stature due to low IGF‐I availability. EMBO Mol Med. 2016;8(4):363‐374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. de Mello‐Coelho V, Villa‐Verde DM, Dardenne M, Savino W. Pituitary hormones modulate cell‐cell interactions between thymocytes and thymic epithelial cells. J Neuroimmunol. 1997;76(1–2):39‐49. [DOI] [PubMed] [Google Scholar]
- 24. Baserga R, Hongo A, Rubini M, Prisco M, Valentinis B. The IGF‐I receptor in cell growth, transformation and apoptosis. Biochim Biophys Acta. 1997;1332(3):F105‐F126. [DOI] [PubMed] [Google Scholar]
- 25. Suzuki A, Mimaki S, Yamane Y, et al. Identification and characterization of cancer mutations in Japanese lung adenocarcinoma without sequencing of normal tissue counterparts. PLoS One. 2013;8(9):e73484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Rizvi NA, Hellmann MD, Snyder A, et al. Mutational landscape determines sensitivity to PD‐1 blockade in non‐small cell lung cancer. Science. 2015;348(6230):124‐128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Hellmann MD, Nathanson T, Rizvi H, et al. Genomic features of response to combination immunotherapy in patients with advanced non‐small‐cell lung cancer. Cancer Cell. 2018;33(5):843‐852 e844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Miao D, Margolis CA, Vokes NI, et al. Genomic correlates of response to immune checkpoint blockade in microsatellite‐stable solid tumors. Nat Genet. 2018;50(9):1271‐1281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Snyder A, Makarov V, Merghoub T, et al. Genetic basis for clinical response to CTLA‐4 blockade in melanoma. N Engl J Med. 2014;371(23):2189‐2199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Van Allen EM, Miao D, Schilling B, et al. Genomic correlates of response to CTLA‐4 blockade in metastatic melanoma. Science. 2015;350(6257):207‐211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Riaz N, Havel JJ, Makarov V, et al. Tumor and microenvironment evolution during immunotherapy with Nivolumab. Cell. 2017;171(4):934‐949 e916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Hugo W, Zaretsky JM, Sun L, et al. Genomic and transcriptomic features of response to anti‐PD‐1 therapy in metastatic melanoma. Cell. 2016;165(1):35‐44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Hoadley KA, Yau C, Hinoue T, et al. Cell‐of‐origin patterns dominate the molecular classification of 10,000 tumors from 33 types of cancer. Cell. 2018;173(2):291‐304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Thorsson V, Gibbs DL, Brown SD, et al. The immune landscape of cancer. Immunity. 2018;48(4):812‐830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Shim JH, Kim HS, Cha H, et al. HLA‐corrected tumor mutation burden and homologous recombination deficiency for the prediction of response to PD‐(L)1 blockade in advanced non‐small‐cell lung cancer patients. Ann Oncol. 2020;31(7):902‐911. [DOI] [PubMed] [Google Scholar]
- 36. Jia Q, Chu Q, Zhang A, et al. Mutational burden and chromosomal aneuploidy synergistically predict survival from radiotherapy in non‐small cell lung cancer. Commun Biol. 2021;4(1):131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Knijnenburg TA, Wang L, Zimmermann MT, et al. Genomic and molecular landscape of DNA damage repair deficiency across the cancer genome atlas. Cell Rep. 2018;23(1):239‐254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Jassal B, Matthews L, Viteri G, et al. The reactome pathway knowledgebase. Nucleic Acids Res. 2020;48(D1):D498‐D503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA‐seq data with DESeq2. Genome Biol. 2014;15(12):550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Yu G, Wang LG, Han Y, He QY. clusterProfiler: an R package for comparing biological themes among gene clusters. Omics. 2012;16(5):284‐287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Zhang M, Zhang X, Tian T, et al. Anti‐inflammatory activity of curcumin‐loaded tetrahedral framework nucleic acids on acute gouty arthritis. Bioact Mater. 2022;8:368‐380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Qin X, Xiao L, Li N, et al. Tetrahedral framework nucleic acids‐based delivery of microRNA‐155 inhibits choroidal neovascularization by regulating the polarization of macrophages. Bioact Mater. 2022;14:134‐144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. de Miguel M, Calvo E. Clinical challenges of immune checkpoint inhibitors. Cancer Cell. 2020;38(3):326‐333. [DOI] [PubMed] [Google Scholar]
- 44. Jardim DL, Goodman A, de Melo GD, Kurzrock R. The challenges of tumor mutational burden as an immunotherapy biomarker. Cancer Cell. 2021;39(2):154‐173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Davis AA, Patel VG. The role of PD‐L1 expression as a predictive biomarker: an analysis of all US Food and Drug Administration (FDA) approvals of immune checkpoint inhibitors. J Immunother Cancer. 2019;7(1):278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Mariathasan S, Turley SJ, Nickles D, et al. TGFβ attenuates tumour response to PD‐L1 blockade by contributing to exclusion of T cells. Nature. 2018;554(7693):544‐548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Jia Q, Wang J, He N, He J, Zhu B. Titin mutation associated with responsiveness to checkpoint blockades in solid tumors. JCI Insight. 2019;4(10):e127901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Zuazo M, Arasanz H, Bocanegra A, et al. Systemic CD4 immunity: a powerful clinical biomarker for PD‐L1/PD‐1 immunotherapy. EMBO Mol Med. 2020;12(9):e12706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Kamada T, Togashi Y, Tay C, et al. PD‐1(+) regulatory T cells amplified by PD‐1 blockade promote hyperprogression of cancer. Proc Natl Acad Sci U S A. 2019;116(20):9999‐10008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Wang Z, Zhao J, Wang G, et al. Comutations in DNA damage response pathways serve as potential biomarkers for immune checkpoint blockade. Cancer Res. 2018;78(22):6486‐6496. [DOI] [PubMed] [Google Scholar]
- 51. Zhang H, Deng YM, Chen ZC, et al. Clinical significance of tumor mutation burden and DNA damage repair in advanced stage non‐small cell lung cancer patients. Eur Rev Med Pharmacol Sci. 2020;24(14):7664‐7672. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Appendix S1 Supporting information.
Data Availability Statement
The public cohorts used in this study were publicly available as described in the ‘Materials and methods’ section. The China cohort is available from the corresponding author on reasonable request.