

Abstract
Food scientists have studied the many health benefits of polyphenols against pernicious human diseases. Evidence from scientific studies has shown that earlier healthy lifestyle changes, particularly in nutrition patterns, can reduce the burden of age-related diseases. In this context, a large number of plant-derived components belonging to the class of polyphenols have been reported to possess neuroprotective benefits. In this review, we examined studies on the effect of dietary polyphenols, notably from Punica granatum L., on neurodegenerative disease, including Alzheimer’s disease, which is symptomatically characterized by impairment of cognitive functions. Clinical trials are in favor of the role of some polyphenols in maintaining neuronal homeostasis and attenuating clinical presentations of the disease. However, discrepancies in study design often bring inconsistent findings on the same component and display differences in their effectiveness due to interindividual variability, bioavailability in the body after administration, molecular structures, cross-blood-brain barrier, and signaling pathways such as nuclear factor kappa B (NF-κB). Based on preclinical and clinical trials, it appears that pomegranate may prove valuable in treating neurodegenerative disorders, including Alzheimer’s disease (AD) and Parkinson’s disease (PD). Therefore, due to the lack of information on human clinical trials, future in-depth studies, focusing on human beings, of several bioactive components of pomegranate’s polyphenols and their synergic effects should be carried out to evaluate their curative treatment.
Keywords: Alzheimer’s disease, Punica granatum L., polyphenols, neurodegenerative diseases
1. Introduction
The improvement of living conditions and access to health care, as well as the progress achieved in the field of public health, have created an increase in the life expectancy of the world’s population. A corollary of this success is the emergence of many age-associated pathologies, such as Alzheimer’s disease (AD). Indeed, the past decades have been marked by a quasi-exponential increase in the prevalence of AD, and the future projections are dramatic. In light of this worrying situation and the absence of curative treatment, the repercussions will exceed the individual medical burden to become considerable social and economic challenges. These considerations, among others, have made AD a major public health problem and a worldwide medical preoccupation.
Although the etiology of AD is imperfectly understood, it is believed to be related to several brain abnormalities, including a defect in the phagocytosis of amyloid-beta (Aβ)-peptide by microglia and cytosolic aggregation in the axons/neurons of the hyperphosphorylated cytoskeletal Tau protein, leading, respectively, to senile plaques and neurofibrillary tangle formation. These abnormalities are closely connected and strongly associated with neuronal damages, including disruption of the neuronal communication at the synapses, cytoskeleton dissociation, and ultimately neuronal death (Figure 1). Several pathological circumstances, such as infections, ischemia, and toxic product release, stimulate microglia cells to secrete several proinflammatory cytokines and chemokines, which alter the neuronal homeostasis. Evidence regarding the overexpression of tumor necrosis factor alpha (TNF-α), interleukin-1β (IL-1β), interleukin-6 (IL-6), and transforming growth factor-β (TGF-β) in the brain of affected patients and transgenic animal models for AD [1,2,3], the association of some genetic polymorphisms of these mediators and the disease manifestation [4], in addition to the neuroprotective impact attributed to anti-inflammatory drugs [5], have accumulated to support the involvement of neuroinflammation in the pathogenesis of neurodegenerative diseases.
Figure 1.


(A) The involvement of neuro-inflammation in Alzheimer’s disease. (B) The proposed operative anti-neuroinflammatory mechanism and amyloidogenic inhibitory effects of pomegranate bioactive compounds.
Pomegranate, a rich source of secondary metabolites and natural compounds (Figure 2), has emerged as a complementary candidate to synthetic anti-inflammatory agents, and has already proved its effectiveness in the context of various chronic inflammatory diseases [6,7,8,9]. In the neuronal micro-environment, pomegranate polyphenols improve brain neurochemistry through their abilities to inhibit NF-κB actions [6], a redox-sensitive transcription factor that is strongly involved in the mRNA transcription of many pro-inflammatory and toxic biomolecules [7]. Additionally, pomegranate bioeffects also include the reduction of beta-site amyloid precursor protein (APP) cleaving enzyme 1 (BACE1) gene expression [6], cyclooxygenase 2 (Cox-2) enzymatic activities [8], and the catalytic activities of caspase enzymes [9]. These effects are thought to reduce neuroinflammation, restore, or at least maintain neuronal homeostasis, and attenuate clinical presentations of the disease.
Figure 2.


(A) Total phenolic content in peels, seeds, and leaves of pomegranate fruit growing in different regions of the world [49,50,52,54,55,56]. BR: Berkane region; SR: Statte region; BMR: Beni Mellal region; UC: unknown cultivar; MWE: methanol/water extract; n-BF1: n-butanol fraction of hydromethanolic extract; CE2: aqueous-acetone crude extract; EAF2: ethyl acetate fraction of aqueous-acetone extract; n-BF2: n-butanol fraction of aqueous-acetone extract; Chelfi-ES: chelfi exposed to sun; Chelfi IC: Chelfi in shade. (B) Total flavonoid content in pomegranate leaves, peels, seeds, and flowers, from different geographical areas of the world [49,52,53,54,56]. BMR: Beni-mellal region; SR: Statte region; BR: Berkane region; UC: unknown cultivar. As shown, the Iranian and Algerian varieties seem to accumulate very important quantities of these bioactive constituents. Furthermore, the final results are sensitive to different factors such as the solvent used for the extraction, genetic variabilities, variety, and the geoedaphic and climatic conditions that characterize each country.
This review describes the effects of the metabolite compounds of Punica granatum L on the attenuation of neurodegenerative diseases including AD and Parkinson’s disease (PD). Most results discuss the anti-inflammatory component of pomegranate in AD.
2. Microglia and Neuroinflammation in the Context of Neurodegenerative Diseases
Neurodegenerative diseases including AD, PD, and amyotrophic lateral sclerosis (ALS) are symptomatically characterized by the impairment of cognitive and/or motor functions. Neurodegeneration in these disorders affects neuronal death, which has been linked to the presence of toxic protein deposits in the central nervous system (CNS), including Aβ and Tau for AD, α-synuclein for PD, and superoxide dismutase 1 (SOD1) and TAR-DNA-binding protein (TDP-43) for ALS [10,11]. Interestingly, these events occur as a result of various neuroinflammatory processes involving glia–neuron cross-talk alterations. Although the genetic and environmental factors that initiate degeneration differ among these diseases, a shared biochemical cascade of inflammatory events plays a central role in mediating neuronal cell loss [12,13,14,15], though it has been well documented that neuroinflammation, largely mediated by microglia, the resident immune cells of the brain, contributes to the onset and progression of neurodegenerative diseases [16].
In a non-pathological context, microglia cells act as macrophages, participate in brain homeostasis, and accomplish several reparative and restorative neuronal functions. This includes the clearance of dendritic debris, synaptic organization, biological response to biotoxins, mediation of inflammation, and the phagocytosis of aggregated proteins [17]. However, the chronic activation of microglia, specifically mediated by microglia proinflammatory M1 phenotype, leads to the production of proinflammatory cytokines that contribute to the progression of various neurodegenerative diseases [18]. The microglia is able to release several signaling molecules required for the maintenance of brain homeostasis through many signaling pathways. Classical NF-κB signaling–NF-κB activation can prevent apoptosis of the cell in which it is activated; it may indirectly lead to apoptosis of other cells by promoting the production of cytotoxic agents and is a major regulator of inflammation, driving the gene expression of pro-inflammatory cytokines, including TNF-α, Il-6, and Il-1β [19], and enzymes such as cyclooxygenase-2 [20] and BACE1 gene expression [21], which have detrimental effects in neurodegenerative diseases (Figure 1A).
In AD patients, morphological changes of microglia surrounding the senile plaques can indicate the level of neuroinflammatory response, the phosphorylation of Tau increases, and the Tau and Aβ pathology exacerbation [22].
Similarly, in Parkinson disease (PD), the over-expression of α-synuclein released from dying dopaminergic neurons drives microglia into a reactive pro-inflammatory phenotype, altering the neuroinflammatory process and promoting neurodegeneration [23,24].
The infiltration of activated microglia observed in ALS patients is responsible for the toxicity caused by mutated SOD1 expression and increases of proinflammatory-activated microglia phenotypes [25].
The same suspects are also established for Huntington’s pathology, to incriminate this exaggerated innate immune response in the necrotic process of neurons and the progressive decline of brain vital capacities [26,27].
In summary, balancing and switching between the phenotypes of microglia at specific times and in specific patients may be important for modulating the progression of neurodegenerative diseases. The accumulation of Aβ and Tau protein, the superoxide dismutase 1 (SOD1) mutant microglia, and the aggregated α synuclein activate microglia through the release of neuroinflammatory mediators that promote neurodegeneration.
Many factors, including microglia phagocytosis incapacities, extracellular misfolded Aβ aggregation, IL-1, and TNF-α cytokines have been proposed to stimulate the NF-κB [28,29]. The activated form of this nuclear factor plays a crucial role in the neuro-inflammatory events that characterizes AD. ECSPF: extracellular senile plaques formation; HPTP: hyperphosphorylated Tau protein; PG-2: prostaglandin-2; IKK: inhibitor of the nuclear factor-κB kinase. Pomegranate polyphenols are thought to be effective in the modulation of the proinflammatory M1 phenotype of microglia. This is probably achieved through its abilities to inhibit NF-κB actions [6], BACE1 transcription [6], and Cox-2 enzymatic activities [8], and to reduce the catalytic activities of the caspase 3 enzyme [9].
3. Polyphenols in Clinical Research of Alzheimer’s Disease
Despite the scientific support of the preclinical results of the anti-neuroinflammatory effects of pomegranate bio-nutrients and products, clinical data are not yet available. However, several extracts and biopolyphenols from other natural resources, and the plant kingdom, have been sufficiently tested, in both preclinical and clinical conditions. Indeed, promising achievements in terms of enhancing neurological functions regarding the neuroprotectivity and antisenescence potential of many phytochemical ingredients from natural resources have attracted the attention of the neuroscientific community. In this regard, a placebo-controlled trial performed with melissa officinalis extract has suggested episodic memory and cognitive capacity enhancement in patients with mild to moderate AD [30]. Moreover, AD-related behavioral symptoms were attenuated in a group of patients treated with turmeric drugs, a traditional Indian medicine [31]. Furthermore, eight months of regular ingestion (200 mL/day) of a beverage rich in antioxidant constituents and polyphenols was seen to reduce homocysteine blood levels, a toxic amino acid that exerts direct neurotoxicity and influences AD pathophysiology and pathogenesis [32]. In the same way, a prospective, randomized, double-blind, placebo-controlled study, performed by Choudhary et al. [33], attributed promising efficacy of Withania somnifera L. Dunal root extracts against age-associated memory decline in Indian volunteers. Additionally, another prospective study carried out using Greek extra virgin olive oil demonstrated the effectiveness of this treatment in neutralizing cognitive impairments [34].
These clinical investigations attribute a significant positive brain impact to pomegranate consumption. However, some studies present limitations, particularly in terms of study size, treatment duration, and negligence of nutrigenomics variabilities, in addition to the possible influences of hormonal fluctuations, as some trials included women participants.
Pure alkaloids with nitrogen groups and organic biomolecules with hydroxyl radicals may also exert neuroprotective effects regarding their central and peripheric biological activities. The healthy benefits of these bio-agents appear to involve multiple mechanisms and to act at different cellular and molecular locations (oxidant damages attenuation, interference with neuronal signaling pathways, toxic Aβ-deposits reduction, and mediation of parasympathetic activities). In this regard, Huperzine-A, a Chinese Huperzia Serrata naturally-occurring nutrient, with reversible, selective, and linear competitive acetylcholinesterase inhibitory activities, was effective in reducing task switching deficit and cognitive abnormalities in patients with Alzheimer’s pathology [35]. This evidence was supported by Chinese clinical research outcomes, which significantly suggest an enhancement of general cognitive capacities and a remarkable improvement of memory in elderly people suffering from the same disease [36]. Furthermore, Huperzine-A polyphenol interacts with AD-related pathophysiology to reduce clinical symptoms and mitigate memory loss in elderly subjects suffering from neurological diseases [37].
Other non-flavonoid compounds, such as resveratrol micronutrient, a natural food ingredient with higher bioavailability and brain blood barrier penetrability [38], were also investigated for their anti-neurodegeneration potential. In fact, a recent retrospective study attributed an anti-AD efficacy to the resveratrol compound, based on its ability to reduce cerebrospinal fluid matrix peptidases (such as metalloproteinase 9), neuroinflammation, and adaptative immune response [39]. However, opposite findings with resveratrol have been observed, suggesting unexpected results, manifesting in a greater reduction of cerebrospinal fluid and serum Aβ40 concentrations in the placebo group compared to the treated group [38]. In most clinical trials, the psychological effect of the placebo “the placebo effect” is neglected, but it could influence psychology and consequently impacts physiology to obtain such effects.
Even though the positive impact of dietary polyphenols on AD is noticeable, there is a clear gap provided from different results, which are conflicting for many reasons: the routes of administration, bioavailability in the body after administration, molecular structure and properties of polyphenols, the treatment duration, the size of trials and subjects, the study characteristics, and the pathways involvement. However, the new therapeutic approach should be exploited carefully for treating neurodegenerative disease. Ongoing study on polyphenol consumption in patients at risk for AD is significant, but it should be validated and transferred to humans without any risks.
4. Nutritional Characterization and Metabolites Compounds of Punica granatum L.
Secondary metabolites, such as flavonoids and ellagitannins, are synthesized through the shikimic pathway and have been, for some time, considered as non-necessary metabolites. It was required to wait for the development of a new understanding in the phytochemistry field to highlight their critical involvement in plant development and survival.
Chromatographic analysis of different varieties of pomegranate from different regions of the world shows both quantitative and qualitative diversity in its bioactive compounds (Table 1 and Table 2), with an abundance of the major polyphenolic classes, including, flavonoids (Figure 2B), tannins, and anthocyanin pigments.
Table 1.
Summary of the most important groups and subgroups of polyphenol compounds present in Punica granatum L.
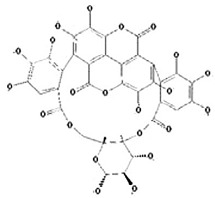
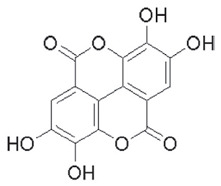
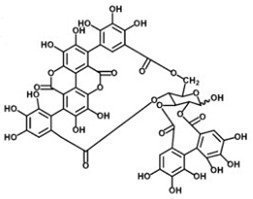
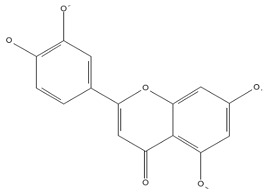
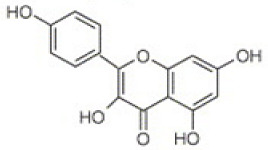
Table 2.
Summary of quantification findings in terms of total phenolic, total flavonoid, total anthocyanin, and total tannin content in different pomegranate parts and products.
| PG Part or Product |
Solvent | Variety | Country | TPC | TAC | TFC | TPA | TT | Ref |
|---|---|---|---|---|---|---|---|---|---|
| Data is presented in mg/L | |||||||||
| Juice | Aqueous | Malase Ashkzar |
Iran | 8130 ± 0.1 | 654,830 ± 0.1 | ni | ni | ni | [43] |
| Sweet Aalak | 2380 ± 0.0 | 815 ± 0.0 | ni | ni | ni | ||||
| Sooleghan | 7440 ± 0.1 | 5980 ± 0.1 | ni | ni | ni | ||||
| Malase Ardestan |
7920 ± 0.1 | 6430 ± 0.2 | ni | ni | ni | ||||
| Saveh Black Leather | 4200 ± 0.2 | 2750 ± 0.1 | ni | ni | ni | ||||
| Ardestan Black Leather | 5820 ± 0.1 | 4330 ± 0.0 | ni | ni | ni | ||||
| Saveh Sweet White Leather | 5490 ± 0.2 | 4070 ± 0.2 | ni | ni | ni | ||||
| Ostokhani Tabas | 9300 ± 0.1 | 7760 ± 0.1 | ni | ni | ni | ||||
| Juice | Aqueous | RPS | 1018 ± 22.1 | 0.87 ± 0.1 | 104.2 ± 12.5 | ni | 3.05 ± 0.34 | [44] | |
| RPGH | 1082 ± 12.8 | 0.18 ± 0.04 | 259.6 ± 9 | ni | 2.22 ± 0.9 | ||||
| ZZ | 960 ± 17. 5 | 0.44 ± 0.1 | 282.1 ± 11.0 | ni | 6.33 ± 0.1 | ||||
| SZ | 1026 ± 21.3 | 8.22 ± 1.9 | 342 ± 12.1 | ni | 4.12 ± 0.8 | ||||
| MY | 706 ± 23.9 | 0.63 ± 0.2 | 320.6 ± 9.9 | ni | 5.21 ± 1.1 | ||||
| MS | 1062 ± 14.1 | 0.48 ± 0.1 | 14.3 ± 4.8 | ni | 6.55 ± 0.5 | ||||
| B | 896 ± 12.1 | 1.13 ± 0.2 | 100 ± 10.1 | ni | 1.01 ± 0.4 | ||||
| Skin | Methanol | RPS | 785 ± 11.1 | 6.40 ± 1.3 | 749.5 ± 15.8 | ni | 3.23 ± 0.2 | ||
| RPGH | 645 ± 31.1 | 0.26 ± 0.03 | 328.2 ± 13.9 | ni | 4.46 ± 1 | ||||
| ZZ | 671 ± 20.02 | 1.57 ± 0.5 | 587.2 ± 10.2 | ni | 1.09 ± 0.7 | ||||
| SZ | 705 ± 23.3 | 1.69 ± 0.6 | 930 ± 16 | ni | 2.43 ± 1 | ||||
| MY | 553 ± 12.5 | 0.05 ± 0.02 | 502.4 ± 9.1 | ni | 4.43 ± 2.4 | ||||
| MS | 597 ± 18.8 | 0.16 ± 0.01 | 382.1 ± 17.5 | ni | 4.22 ± 2.2 | ||||
| B | 783 ± 10.02 | 11.20 ± 2.4 | 712.7 ± 9 | ni | 4.1 ± 0.8 | ||||
| Juice | Aqueous | unknown cultivar | Germany | 2015.2 ± 21.6 | 198.3 ± 0.8 | ni | 6.9 ± 0.0 a 21.4 ± 0.9 b |
423.8 ± 15.3 | [45] |
| 5186.0 ± 172.5 | 124.2 ± 2.2 | ni | 1.1 ± 0.0 a b: nd |
2074.4 ± 47.3 | |||||
| 2122.0 ± 0.0 | 557.7 ± 48.3 | ni | 10.6 ± 0.3 a 28.3 ± 1.4 b |
93.2 ± 3.6 | |||||
| Juice | Aqueous | Ermioni variety |
Greece | 1271 ± 40 | 382.8 ± 0.1 | ni | ni | ni | [46] |
| Data is presented in mg/kg | |||||||||
| Peel | Ethanol, formic acid, MeOH, water, and acetonitrile |
Italian cultivar | Italy | - | not detected | 16,156.4 ± 7450.9 | 5285.6 ± 9316.1 | ni | [47] |
| Pulp | - | 264.3 ± 184.7 | 344.4 ± 281.4 | 77.4 ± 33.3 | ni | ||||
| Peel | Aqueous | unknown cultivar |
Germany | 44,261.5 ± 414.0 | 447.1 ± 11.3 | ni | 270.4 ± 18.5 a b: nd |
43,991.2 ± 395.5 c | [45] |
| Mesocarp | 40,625.1 ± 4434.7 | nd | ni | a: nd b: nd |
40,625.1 ± 4434.7 c | ||||
| Data is presented in mg/g | |||||||||
| Peel | Ethanol, formic acid, methanol, water, and acetonitrile | Gaeta 1 | Italy | 179.92 ± 1.31 | - | - | - | ni | [47] |
| Gaeta 3 | 244.61 ± 1.41 | - | - | - | ni | ||||
| Gaeta 4 | 182.15 ± 1.57 | - | - | - | ni | ||||
| Tordimonte A | 89.68 ± 0.61 | - | - | - | ni | ||||
| Itri A | 141.14 ± 0.45 | - | - | - | ni | ||||
| Wonderful | 137.28 ± 1.19 | - | - | - | ni | ||||
| Formia | 191.59 ± 3.38 | - | - | - | ni | ||||
| Pulp | Gaeta 1 | 8.89 ± 0.08 | - | - | - | ni | |||
| Gaeta 3 | 5.40 ± 0.09 | - | - | - | ni | ||||
| Gaeta 4 | 4.95 ± 0.06 | - | - | - | ni | ||||
| Tordimonte A | 3.19 ± 0.07 | - | - | - | ni | ||||
| Itri A | 6.11 ± 0.07 | - | - | - | ni | ||||
| Wonderful | 6.14 ± 0.03 | - | - | - | ni | ||||
| Formia | 5.32 ± 0.05 | - | - | - | ni | ||||
| Aril | Aqueous | Unknown cultivar | 0.040 ± 0.0072 | ni | ni | ni | ni | [48] | |
| Ethyl acetate | 0.007 ± 0.0012 | ni | ni | ni | ni | ||||
| Juice | Aqueous | 0.052 ± 0.0001 | ni | ni | ni | ni | |||
| Ethyl acetate | 0.001 ± 0.0003 | ni | ni | ni | ni | ||||
| Rind | Aqueous | 0.907 ± 0.0757 | ni | ni | ni | ni | |||
| Ethyl acetate | 0.031 ± 0.0003 | ni | ni | ni | ni | ||||
| Seed | aqueous | Gabsi variety | Tunisia | 7.94 ± 1.25 | 19.62 ± 3.12 | 3.30 ± 0.52 | ni | 32.86 ± 4.24 c | [49] |
| Leave | 9.85 ± 0.82 | 40.91 ± 3.43 | 12.77 ± 0.23 | ni | 64.40 ± 4.85 c | ||||
| Flower | 42.70 ± 2.17 | 80.20 ± 7.02 | 21.45 ± 0.58 | ni | 57.04 ± 3.41 c | ||||
| Peel | 53.65 ± 4.13 | 51.02 ± 10.33 | 21.03 ± 1.62 | ni | 62.71 ± 11.32 c | ||||
| Seed | Methanol | 11.84 ± 1.92 | 40.84 ± 7.77 c | 6.79 ± 0.57 | ni | 29.57 ± 4.54 c | |||
| Leave | 14.78 ± 2.10 | 89.81 ± 7.50 | 26.08 ± 1.24 | ni | 128.02 ± 4.49 c | ||||
| Flower | 66.29 ± 3.06 | 168.91 ± 3.1 | 72.52 ± 5.59 | ni | 148.24 ± 10.29 c | ||||
| Peel | 85.60 ± 4.87 | 102.20 ± 16.42 | 51.52 ± 8.14 | ni | 139.63 ± 4.25 c | ||||
| Peel | Aqueous | Chelfi variety |
216.9 ± 7.3 | ni | ni | ni | ni | [50] | |
| Fruit | Methanol-water | Unknown cultivar | Algeria | 15.39 ± 0.08 | ni | 12.95 ± 0.07 | ni | ni | [51] |
| Seed | Methanol | Sefri | Morocco (BMR) |
67. 85 ± 1.98 | ni | 1.76 ± 0.02 | ni | ni | [52] |
| Morocco (SR) |
63.34 ± 0.7 | ni | 2.11 ± 0.28 | ni | ni | ||||
| Morocco (BR) |
62.17 ± 3.26 | ni | 1.94 ± 0.00 | ni | ni | ||||
| Peel | Morocco (BMR) |
224.39 ± 3 | ni | 62.63 ± 3.23 | ni | ni | |||
| Morocco (SR) |
223.21 ± 15 | ni | 52.12 ± 1.36 | ni | ni | ||||
| Morocco (BR) |
204.58 ± 1.96 | ni | 46.17 ± 2.18 | ni | ni | ||||
| Flowers | Hydromethanolic | Unknown cultivar | Iran | 348.81 ± 5.58 | ni | 225.776 ± 2.93 | ni | ni | [53] |
| Aqueous | 509.83 ± 11.61 | ni | 98.399 ± 1.15 | ni | ni | ||||
| n-Butanol | 866.47 ± 26.61 | ni | 258.127 ± 16.19 | ni | ni | ||||
| Ethyl acetate | 944.75 ± 8.27 | ni | 359.6 ± 13.91 | ni | ni | ||||
| Peel | Ethanol | Natanz | 276 ± 12.69 | ni | 36 ±3.56 | ni | ni | [54] | |
| Shahreza | 361 ± 12.87 | ni | 45 ± 6.25 | ni | ni | ||||
| Doorak | 413 ± 16.84 | ni | 54 ± 8.96 | ni | ni | ||||
| Seed | Natanz | 72.4 ± 10.02 | ni | 30.5 ± 6.38 | ni | ni | |||
| Shahreza | 73 ± 13.35 | ni | 7.55 ± 2.12 | ni | ni | ||||
| Doorak | 73 ± 9.45 | ni | 38 ± 6.38 | ni | ni | ||||
| Juice | Ethanol | Natanz | 23.8 ± 6.74 | ni | 1.8 ± 1.03 | ni | ni | ||
| Shahreza | 15.8 ± 5.81 | ni | 2.14 ± 0.92 | ni | ni | ||||
| Doorak | 12.4 ± 5.21 | ni | 8.7 ± 2.47 | ni | ni | ||||
| Peel | Methanol-water | Sweet-GP | China | 264.58 ± 8.74 | ni | ni | ni | ni | [55] |
| Flesh | 217.14 ± 16.80 | ni | ni | ni | ni | ||||
| Seeds | 9.04 ± 1.23 | ni | ni | ni | ni | ||||
| Juices | 8.62 ± 1.05 | ni | ni | ni | ni | ||||
| Leaves | 82.31 ± 2.26 | ni | ni | ni | ni | ||||
| Peel | Sweet-RP (Chinese cultivar) |
231.36 ± 4.40 | ni | ni | ni | ni | |||
| Flesh | 203.20 ± 12.68 | ni | ni | ni | ni | ||||
| Seeds | 6.17 ± 0.60 | ni | ni | ni | ni | ||||
| Juices | 6.12 ± 0.91 | ni | ni | ni | ni | ||||
| Leaves | 75.56 ± 4.39 | ni | ni | ni | ni | ||||
| Peel | Sour-RP (Chinese cultivar) |
255.31 ± 6.42 | ni | ni | ni | ni | |||
| Flesh | 208.39 ± 6.31 | ni | ni | ni | ni | ||||
| Seeds | 8.31 ± 0.83 | ni | ni | ni | ni | ||||
| Juices | 10.36 ± 1.12 | ni | ni | ni | ni | ||||
| Leaves | 75.61 ± 3.62 | ni | ni | ni | ni | ||||
| Peel | Sour-YRP | 302.43 ± 9.54 | ni | ni | ni | ni | |||
| Flesh | 272.10 ± 9.98 | ni | ni | ni | ni | ||||
| Seeds | 12.44 ± 0.96 | ni | ni | ni | ni | ||||
| Juices | 13.31 ± 0.87 | ni | ni | ni | ni | ||||
| Leaves | 88.33 ± 1.92 | ni | ni | ni | ni | ||||
| Peel | Sweet-TRP | 279.76 ± 13.32 | ni | ni | ni | ni | |||
| Flesh | 222.62 ± 7.72 | ni | ni | ni | ni | ||||
| Seeds | 8.59 ± 1.04 | ni | ni | ni | ni | ||||
| Juices | 12.91 ± 0.87 | ni | ni | ni | ni | ||||
| Leaves | 79.10 ± 2.74 | ni | ni | ni | ni | ||||
| Peel | CE1 | Unknown cultivar |
Algeria | 158.18 ± 0.66 | ni | 12.8 ± 2.2 | ni | - | [56] |
| CE2 | 221.54 ± 1.08 | ni | 30.9 ± 1.6 | ni | - | ||||
| EAF1 | 597.08 ± 3.9 | ni | 135.5 ± 2.0 | ni | - | ||||
| n-BF1 | 413.6 ± 1.48 | ni | 91.1 ± 0.88 | ni | - | ||||
| EAF2 | 602.8 ± 2.41 | ni | 149.4 ± 1.27 | ni | - | ||||
| n-BF2 | 417.56 ± 5.3 | ni | 115.56 ± 1.81 | ni | - | ||||
| Data is presented in µg/mg | |||||||||
| Fruit | Water | Unknown cultivar |
India | 11.99 | ni | ni | ni | ni | [57] |
| Ethanol | 28.13 | ni | ni | ni | ni | ||||
| Acetone | 6.44 | ni | ni | ni | ni | ||||
| Ether | 24.13 | ni | ni | ni | ni | ||||
| Ethanol-Ether | 24.2 | ni | ni | ni | ni | ||||
| Ethanol-water | 15.12 | ni | ni | ni | ni | ||||
| Ethanol-Ether-water | 24.5 | ni | ni | ni | ni | ||||
| Data is presented in mg/mL | |||||||||
| Juice | Aqueous | Gaeta 1 | Italy | 1.93 ± 0.05 | - | - | - | ni | [47] |
| Gaeta 3 | 1.34 ± 0.07 | - | - | - | ni | ||||
| Gaeta 4 | 0.87 ± 0.03 | - | - | - | ni | ||||
| Tordimonte A | 1.22 ± 0.03 | - | - | - | ni | ||||
| Itri A | 1.20 ± 0.05 | - | - | - | ni | ||||
| Wonderful | 1.58 ± 0.02 | - | - | - | ni | ||||
| Formia | 1.08 ± 0.06 | - | - | - | ni | ||||
Abbreviations: TPC: total phenolic content; TAC: total anthocyanin content; TFC: total flavonoid content; TPA: total phenolic acids; TT: total tannin; a: total hydroxybenzoic acids; b: total hydroxycinnamic acids; c: total hydrolyzable tannins; BR: Berkane region; BMR: Beni Mellal region; SR: Statte region; MeOH: Methanolic solution; PGS E1: Pomegranate seed extract protected from light; PGS E2: Pomegranate seed extract exposed to light; CE1: hydromethanolic crude extract; ni: not investigated; nd: not detected; -: investigated but reported by another unit; EAF1: ethyl acetate fraction of hydromethanolic crude extract; n-BF1: n-butanol fraction of hydromethanolic crude extract; CE2: aqueous-acetone crude extract; EAF2: ethyl acetate fraction of aqueous-acetone crude extract; n-BF2: n-butanol fraction of aqueous-acetone crude extract.
Anthocyanins from pomegranate have been sufficiently investigated and are represented by glucosides of pelargonidin, delphinidin, and cyanidin forms [58,59]. These functional compounds are among the major contributors to the pharmacological properties and color qualities of many vegetables and fruits. Indeed, the blue, red, and orange colors have been ascribed to delphinidin, cyanidin, and pelargonium structures, respectively. Progressively, with the advancement of the stage of maturity, an increase in anthocyanins’ proportions and a decrease in the total phenolic content have been recorded [60]. This is reported to be accompanied by a progressive change in the color properties and the decline of the antioxidant capacities [61]. The reported results for anthocyanins appear to vary from one variety to another, and from region to region (Table 2), suggesting the possible influence of genetic factors and environmental conditions.
Flavonoids are another class of polyphenols and are among the most important secondary metabolites detected in vegetables and fruits. Flavonoids identified in pomegranate include, mainly, catechin [62], epicatechin [62], flavan-3-ol [62], quercetin [63], kaempferol [64], luteolin [64], naringin [65], pelargonidin [65], cyanidin [65], cyanidin 3-O-glucoside [66], cyanidin 3.5-di-O-glucoside [66], delphinidin 3-O-glucoside [66], delphinidin 3.5-di-O-glucoside [66], and punicaflavone [67]. The flavonoid-associated beneficial health effects, such as antioxidant [68], anti-mutagenic [68], and anti-inflammatory properties [69], have conferred an added value to this group, with a great impact on pharmaceutical and medicinal preparations.
Organic acids from pomegranate are mainly represented by citric acid, oxalic acid, shikimic acid, acetic acid, maleic acid, fumaric acid, succinic acid, tartaric acid, and ascorbic acid [70]. Phytochemical analysis of six cultivars growing in Georgia shows citric acid as a predominant organic acid in seed, pull, peel, and leaf (315.7 ± 1.0; 826.7 ± 5.4; 766.9 ± 3.2, and 130.2 ± 1.6 mg/100 g, respectively) [71]. Moreover, in the others twenty-five Iranian cultivars, and thirteen Turkish pomegranate varieties, the highest value of organic acid was also recorded for citric acid (3763.6 ± 144.8 mg/100 g, and 8.96 ± 0.45 g/L, respectively) [70,72]. Other organic acids, including oxalic, succinic, tartaric, and ascorbic acid, were also detected in pomegranate, but at lower amounts [70,72].
The tannin class is extensive, and tannins are the most significant phenolic compounds present in pomegranate. These include, mainly, pomegranatate [73], punicalin [74], 4.4-di-O-methylellagic acid [75], 3-O-methyl-3.4-methylenedioxy [75], 3-O-methylellagic acid [75], punicafolin [76], punicacortein [77], gallic acid [78], ellagic acid [79], and punicalagin [74,80]. In vitro and in vivo study has listed many neuroprotective actions against AD for this last compound [6], and it is suspected to be mediated by its gut microbiota-derived molecules, namely urolithins. This idea is supported by two concerns: the well-documented antioxidant and anti-inflammatory capacities attached to urolithin structures [81], and secondly, to their exclusive ability to penetrate the blood-brain barrier [82].
5. Pomegranate Consumption and Neurodegenerative Disorders
5.1. Anti-Inflammatory Effects of Pomegranate Intake on Alzheimer’s Disease
One of the first descriptions of the probable involvement of immune mechanisms in AD were made by Rogers et al. in 1988 [83]. As we discussed above, persistent activation of glial cells and astrocytes is suspected of being responsible for multi-neurological illnesses, including AD. This is supported by evidence from genetic investigations, which have linked the genetic polymorphisms of some pro-inflammatory cytokines to the pathological expression of the disease. Furthermore, a meta-analysis of observational studies suggests that chronic utilization of non-steroidal anti-inflammatory drugs could decrease the risk of developing AD [5]. However, most of these drugs are synthetic, expensive, and may present serious side effects. Therefore, it is of paramount importance to envisage and nominate new safe and effective agents that can be used as nutraceuticals and can be easily integrated into a healthy diet, such as the Mediterranean diet.
Bioactive compounds from medicinal plants offer an alternative and have emerged as a key strategy to surmount the mentioned limitations. Pomegranate is one such medicinal plant and contains a considerable number of polyphenols and natural pigments known for their antioxidant and anti-inflammatory properties. This last pharmacological activity of pomegranate has been extensively investigated in the context of several chronic inflammatory diseases [84,85,86,87,88]. Nevertheless, there are limited published investigations [3,6,89] that have focused their research interests on its capacities to attenuate the neuroinflammatory cascade involved in AD.
Evidence from transgenic animal models of AD (APPsw/Tg2576 mice) has proven the significant impact of long-term (15 months) diet supplementation with pomegranate fruit (4% w/w) on the level of the main pro-inflammatory cytokines, namely IL-2, IL-3, IL-4, IL-5, IL-6, IL-9, IL-10, IL-1β, and TNF-α, with interesting anti-amyloidogenic effects [3] (Figure 1B). Moreover, another study suggests that the same dose and duration of treatment were also effective in reducing the expression of BACE1, soluble amyloid precursor protein-β (sAPP-β), insulin-like growth factor-1 (IGF-1), inducible nitric oxide synthase (iNOS), C-terminal fragment-β (CTF-β), brain-derived neurotrophic factor (BDNF), postsynaptic density proteins-95 (PSD-95), and phosphorylation of calcium/calmodulin-dependent protein kinase II-α (p-CaMKII-α/CaMKII-α) [90]. Furthermore, Kim et al. [6] tested the therapeutical capacities of punicalagin, an ellagitannin-derived compound, on neuroinflammation and memory impairment. The research outcomes suggest that this compound may reduce Aβ 1-42, IL-1β, IL-6, and TNF-α levels, and neuronal expression of BACE1. Its capacities to attenuate NF-κB DNA binding activity and to inhibit the nuclear factor of kappa-B inhibitor alpha (IkB-α) phosphorylation were also reported by the same authors. To this anti-inflammatory potential, antioxidant activities have also been mentioned, and are exemplified mainly in the free radical-scavenging activities and the protection of brain lipids from the peroxidation cascade. However, all the animal models used in these studies present limitations. This usually manifests in their inability to reproduce the broad spectrum of the disease (low “face validity”) as occurring in humans, as well as in the structural and biochemical non-similarity of amyloid deposits between animal models of AD and affected human subjects [91].
Most of the in vivo-observed effects were also well-established in an in vitro context (Table 3). Pomegranate juice (25–200 μg/mL) treatment of human neuroblastoma cells (SK-N-SH), was correlated with the reduction of cyclooxygenase 2 expression, and consequently with prostaglandin E2 production [8]. An inhibitory effect of IκB, IKK, and BACE1 gene expression and Aβ peptide production were also registered. The same effects on cyclooxygenase 2 and BACE1 expressions were obtained in a co-culture of primary astrocytes and BV-2 cells cultivated with the punicalagin compound (10, 20, and 50 µM) [6]. Additionally, an inhibitory action is also observed against phosphorylation of IkB-α. This last effect is of fundamental importance to the inflammatory pathway, both on the capacity of IkB-α to govern NF-κB pathways and on related activities. The results from Rackova et al. [92] strengthen this evidence and attribute an important neuroprotective potential of pomegranate against glial cell-mediated neuroinflammation.
Table 3.
Summary of the main in vivo and in vitro findings related to the effect of the pomegranate intake on memory and neuro-inflammation implicated in Alzheimer’s disease.
| Animal Model/ Cell Line/CTI |
Induction of Neuro-Inflammation | Pomegranate Part or Product | Dose and Period of Treatment | Findings | Ref |
|---|---|---|---|---|---|
| Animal studies | |||||
| Male C57BI/6 mice | Chronic injection of Aβ peptide |
Pomegranate peel extract | 800 mg/kg/day for 35 days |
Improves spatial memory, and the expression of neurotrophin BDNF; reduce amyloid plaque density, AchE activity, lipid peroxidation and TNF-α level; no hepatic lesions. | [89] |
| Male ICR mice | LPS injection (250 µg/kg) | Punicalagin | 1.5 mg/kg for 4 weeks |
Memory improvement; no effect on swimming speed; reduction of BACE1 expression and Aβ 1-42 generation; no effect on APP expression; decrease the Il-1-β, IL-6 and TNF-α, H2O2 and MDA level; increase GSH/GSSG ratio; inhibition of iNOS and Co-2 activities; decrease NF-kB DNA binding activity; suppress IkB-α phosphorylation and P50 and P65 subunits translocation. | [6] |
| APPsw/Tg2576 mice | Experimental animal model overexpressing APP | Pomegranate fruit | 4% w/w, for 15 months | Decrease serum concentration of IL-2, IL-3, IL-4, IL-5, IL-9, IL-10, and Eotaxin; decrease the level of IL-1β, TNF-α, and IL-6, Aβ1–42 and 40, both in the cortex and hippocampus areas. | [3] |
| APPsw/Tg2576 mice | Transgenic female | Pomegranate extract | 4% w/w, for 15 months | Significant inhibition of TNF-α, Il-1-β, iNOS, ccl2, and Il-10 genes expression. | [90] |
| APP/PS1 transgenic mice | Transgenic female | Urolithin A | 300 mg/kg, for 14 days | Neuronal death attenuation; hippocampal neurogenesis improvement; diminished astrocytes and microglia activation; reduction in the level of IL-1β, IL-6, and TNF-α. | [94] |
| In vitro studies | |||||
| The human neuroblastoma (SK-N-SH) cells | IL-1 β (10 U/mL) | Standardized Pomegranate fruit juice (skins) | 25–200 μg/mL, for 24 h |
Inhibition of Co-2 activity, and PGE-2 production; significant inhibition of IκB and IKK phosphorylation; reduction of BACE1 gene expression and Aβ production. | [8] |
| HEK293 cells | TNFα (1 ng/mL) | 25–200 μg/mL, for 6 h | Significant inhibition of NF-κB transactivation. | ||
| Primary astrocyte and microglial BV-2 cells | LPS-(10, 20 and 50 µM), for 1 h | Punicalagin | (1 µg/mL), for 24h | Suppress the activation of NF-kB; inhibit IkB degradation and p50 and p65 translocation; inhibit APP and BACE1 transcription. | [6] |
| Microglial BV-2 cell | LPS | Pomegranate seed oil | 25 μg/mL, for 24h | Slight inhibition of TNF-α and iNOS gene expression; prevent microglia cells apoptosis. | [92] |
| Clinical trials | |||||
| NCT02093130 | _ | Pomegranate juice | 236.5 mL, for 12 months | Pomegranate juice supplementation stabilizes human capacities in terms of the acquisition of new visual information during advancement with age. | [93] |
CTI: clinical trial identifier; BDNF: brain-derived neurotrophic factor; TNF-α: tumor necrosis factor-alpha; Aβ1-42: amyloid beta 1-42; BACE1: beta-site APP cleaving enzyme 1; APP: Amyloid precursor protein; IL-1β: interleukin-1β; IL-6: interleukin-6; H2O2: hydrogen peroxide; MDA: malondialdehyde; GSH: reduced glutathione; GSSG: oxidized glutathione; iNOS: inducible nitric oxide synthase; Co-2: cyclooxygenase 2; AchE: acetylcholinesterase; NF-κB: nuclear factor-kappa B; IkB: inhibitor of nuclear factor-κB; P50: p50 protein; P65: p 65 protein; IL-2: interleukin-2; IL-3: interleukin-3; IL-4: interleukin-4; IL-5: interleukin-5; IL-9: interleukin-9; IL-10: interleukin-10; PGE-2: prostaglandin-2; IKK: inhibitor of nuclear factor-κB kinase; SK-N-SH: human neuroblastoma cell line. BV-2: murine microglial cell line; HEK293: human embryonic kidney cells-293; Ccl2: chemokine ligand 2; LPS: Lipopolysaccharide.
The neuroprotective ability of pomegranate and its abilities to improve, or at least to maintain, cognitive performances have been scarcely investigated in human subjects. Until now, and to the best of our knowledge, there is only one clinical trial [93] that has investigated the effects of long-term pomegranate juice supplementation on healthy middle-aged and older adults. In this randomized placebo-controlled research, the pomegranate molecules were effective in stabilizing visual memory capacities. However, no other conclusive effects were observed in these healthy subjects.
The discussed benevolent effects are thought to be associated with its capacity to inhibit the cyclooxygenase pathway and the enzymatic activities of β and γ secretase [8]. The inflammatory lowering effect is well documented as being related to the modulation of the NFκB signaling pathways (Figure 1B) [6,8]. Other mechanisms, such as radical scavenging properties, and the organism’s antioxidant status improvement may also contribute to the observed neuroprotective impact. Human studies are badly needed to corroborate the in vitro and animal studies.
5.2. Anti-Inflammatory Effects of Pomegranate Intake against Parkinson’s Disease
To date, there are few studies that have investigated the possible link between pomegranate intake and neuro-inflammation implicated in PD [95,96]. The short-term (two weeks) diet supplementation with pomegranate juice (6.5–7.5 mL/day) of male Lewis rats was found to be associated with the aggravation of the inflammatory response, dopaminergic neuronal death, and caspase activity improvement, with no effect on postural instability [95]. These unexpected results have attracted more attention, regarding the anti-inflammatory and antioxidant activities attached to pomegranate consumption [97,98,99]. Numerous factors have been proposed to explain these interesting findings, among them the fact that polyphenol constituents may act as a pro-oxidant compound with all known associated deleterious consequences on neuronal components. Five years later, a paper appeared to suggest opposing results [100]. In this research, the anti-Parkinson’s disease effects are attributed to pomegranate consumption (500 mg/kg/day), and evidenced by anti-α-synuclein aggregation, oxidative damage attenuation, neuronal survival improvement, and postural stability enhancement. Similar findings have been obtained by [96] in a rat model of Parkinson’s disease, using standardized pomegranate extract (40% ellagic acid). The pomegranate-treated groups exhibit a reduction in malondialdehyde, ILβ-1, and TNF-α levels, in addition to the reduction of iNOS activity and apoptotic caspase-3 expression. Furthermore, this phenolic extract also increases cellular brain viability and locomotor performances. However, all these studies have used Rotenone to induce Parkinson’s disease-like-pathology, and they are not based on the same dose, duration of treatment, and animal model (Table 4). Additionally, the artificial method used for the disease induction does not simulate the normal pathological process. Consequently, this may impact the results and contribute to explaining, at least in part, the contradictory reported outcomes.
Table 4.
Summary of the main findings related to Pomegranate supplementation effect on Parkinson’s disease development.
| Animal Model | Induction of Parkinson’s Disease |
Pomegranate Part or Product | Dose And Period Of Treatment | Findings | Ref |
|---|---|---|---|---|---|
| Male Lewis rats | intraperitoneal administration of Rotenone (3 mg/kg) | Pomegranate juice | 6.5–7.5 mL/day, for 2 weeks | ↑ iNOS expression and activation; ↔ IL-1β; ↔ TNF-α level; ↔ Cox2 expression; ↑ p65 catalytic subunit of the NF-κB; ↑caspase-3 activity. | [95] |
| Male albino rats | Rotenone (2.5 mg/kg), for 30 days |
Pomegranate extract | 150 mg/kg/day, for 30 days | ↑ locomotor activity; ↓ AchE activities; ↑ BDNF and glutamate; ↓ lipid peroxidation; ↑ SOD; ↑ TAA; ↓ IL-1β; ↓ TNF-α; ↓ iNOS. | [96] |
| Male albino Wistar rats | subcutaneous injection of Rotenone (1.3 mg/kg), for 35 days |
Pomegranate juice |
500 mg/kg/day, for 45 days | ↑ postural stability; ↑ neuronal survival; ↓ MDA; ↓ α-synuclein; ↑ mitochondrial ALDH activity; ↑ antiapoptotic Bcl-xL protein. | [100] |
↑: increase; ↓: decrease; ↔: no effect; iNOS: inducible nitric oxide synthase; IL-1β: interleukin 1-β; TNFα: tumor necrosis factor-alpha; Cox-2: cyclooxygenase 2; NF-κB: nuclear factor-kappa B; AchE: acetylcholinesterase; BDNF: brain-derived neurotrophic factor; TAA: total antioxidant activity; SOD: superoxide dismutase; P65: p 65 protein; MDA: malondialdehyde; ALDH: aldehyde dehydrogenases.
5.3. The Preventive Role of Pomegranate against Multiple Sclerosis
Limited research has focused on the ability of pomegranate polyphenols to prevent the expression and development of multiple sclerosis abnormalities. Scopus, web of science, and PubMed database research show the existence of only one paper that has concentrated its focus of interest on pomegranate intake and neuro-inflammation implicated in multiple sclerosis [101]. In this publication, pomegranate peel extract was effective in reducing the level of Il-17. This may interfere with multiple sclerosis expression besides its ability to attenuate the activation of encephalitogenic T-cells. Such an event is fundamental in multiple sclerosis and encephalomyelitis disease development [102]. Moreover, pomegranate seed oil has been proven to inhibit demyelination, a seminal feature of the disease, in addition to its great capacities to attenuate brain lipid peroxidation [103]. However, it is very difficult to generalize these outcomes due to the limited amount of research, and it will be essential to await future studies for probable confirmation or information of the mentioned effects. Therefore, we have highlighted the need to perform more research to further explore and to clarify the effect of pomegranate intake on this pathology.
6. Conclusions
We reviewed the roles of polyphenols from pomegranate in neurodegenerative diseases, mostly focusing on AD and PD. In addition, clinical studies on pomegranate treatment associated with the neuroinflammation involved in neurodegenerative diseases were discussed, even though the data was insufficient. From that, it appears that a balance between the neuroprotective glial cells involved in glia-neurons cross-talks may be critical in the progression of neurodegenerative pathologies. Disease stage and complexity, the microglial phenotypes, the polyphenols potents, the routes of administration, and the bioavailability of polyphenols in the brain may influence the neuroprotective benefits of pomegranate’s components against neurodegeneration. Further in-depth studies should be conducted in the future based on human beings and carried out to investigate the curative treatment of pomegranate’s polyphenols, established on the new synergic approaches between different components of pomegranate and neuroinflammation in neurodegenerative disorders.
Author Contributions
Conceptualization: A.M. and B.H.; methodology: A.M., B.H. and B.S.; software: A.M., M.R. and B.H.; schematization: A.M. and B.L.; writing—original draft preparation: A.M., B.L., B.H. and B.S.; writing—review and editing: A.M., B.H., F.T. and K.A.; visualization, K.A., F.T. and M.R.; supervision, B.H. and B.S.; project administration, B.H.; funding acquisition, B.H. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Funding Statement
This research was funded by (Agence Nationale des Plantes Médicinales et Aromatiques; Centre National de Recherche Scientifique et Technique and Université Sultan Moulay Slimane), (project number: 3/2021-348/20. Morocco (H.B.).
Footnotes
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Swardfager W., Lanctôt K., Rothenburg L., Wong A., Cappell J., Herrmann N. A meta-analysis of cytokines in Alzheimer’s disease. Biol. Psychiatry. 2010;68:930–941. doi: 10.1016/j.biopsych.2010.06.012. [DOI] [PubMed] [Google Scholar]
- 2.Forlenza O.V., Diniz B.S., Talib L.L., Mendonça V.A., Ojopi E.B., Gattaz W.F., Teixeira A.L. Increased serum IL-1beta level in Alzheimer’s disease and mild cognitive impairment. Dement. Geriatr. Cogn. Disord. 2009;28:507–512. doi: 10.1159/000255051. [DOI] [PubMed] [Google Scholar]
- 3.Essa M.M., Subash S., Akbar M., Al-Adawi S., Guillemin G.J. Guillemin Long-Term dietary supplementation of pomegranates, figs and dates alleviate neuroinflammation in a transgenic mouse model of alzheimer’s disease. PLoS ONE. 2015;10:e120964. doi: 10.1371/journal.pone.0120964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Flex A., Giovannini S., Biscetti F., Liperoti R., Spalletta G., Straface G., Landi F., Angelini F., Caltagirone C., Ghirlanda G., et al. Effect of proinflammatory gene polymorphisms on the risk of Alzheimer’s disease. Neurodegener. Dis. 2014;13:230–236. doi: 10.1159/000353395. [DOI] [PubMed] [Google Scholar]
- 5.Stroup D.F., Berlin J.A., Morton S.C., Olkin I., Williamson G.D., Rennie D., Moher D., Becker B.J., Sipe T.A., Thacker S.B. Effect of non-steroidal anti-inflammatory drugs on risk of Alzheimer’s disease. BMJ. 2003;327:128. [Google Scholar]
- 6.Kim Y.E., Hwang C.J., Lee H.P., Kim C.S., Son D.J., Ham Y.W., Hellström M., Han S.B., Kim H.S., Park E.K., et al. Inhibitory effect of punicalagin on lipopolysaccharide-induced neuroinflammation, oxidative stress and memory impairment via inhibition of nuclear factor-kappaB. Neuropharmacology. 2017;117:21–32. doi: 10.1016/j.neuropharm.2017.01.025. [DOI] [PubMed] [Google Scholar]
- 7.Sun S.C., Chang J.H., Jin J. Regulation of nuclear factor-kB in autoimmunity. Trends Immunol. 2013;34:282–289. doi: 10.1016/j.it.2013.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Velagapudi R., Baco G., Khela S., Okorji U., Olajide O. Pomegranate inhibits neuroinflammation and amyloidogenesis in IL-1 β-stimulated SK-N-SH cells. Eur. J. Nutr. 2015;55:1653–1660. doi: 10.1007/s00394-015-0984-0. [DOI] [PubMed] [Google Scholar]
- 9.Loren D.J., Seeram N.P., Schulman R.N., Holtzman D.M. Maternal dietary supplementation with pomegranate juice is neuroprotective in an animal model of neonatal hypoxic-ischemic brain injury. Pediatric Res. 2005;57:858–864. doi: 10.1203/01.PDR.0000157722.07810.15. [DOI] [PubMed] [Google Scholar]
- 10.Yoo H.J., Kwon M. Aged Microglia in Neurodegenerative Diseases: Microglia Lifespan and Culture Methods. Front. Aging Neurosci. 2022;13:766267. doi: 10.3389/fnagi.2021.766267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cuní-López C., Stewart R., Quek H., White A.R. Recent Advances in Microglia Modelling to Address Translational Outcomes in Neurodegenerative Diseases. Cells. 2022;11:1662. doi: 10.3390/cells11101662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Marinelli S., Basilico B., Marrone M.C., Ragozzino D. Microglia-neuron crosstalk: Singh naling mechanism and control of synaptic transmission. Semin. Cell Dev. Biol. 2019;94:138–151. doi: 10.1016/j.semcdb.2019.05.017. [DOI] [PubMed] [Google Scholar]
- 13.Correa F.C. Miriam Hernangómez, Carmen Guaza. Understanding Microglia–Neuron Cross Talk: Relevance of the Microglia–Neuron Cocultures. Microglia; Totowa, NJ, USA: 2013. [DOI] [PubMed] [Google Scholar]
- 14.Damulewicz M., Szypulski K., Pyza E. Glia-Neurons Cross-Talk Regulated Through Autophagy. Front. Physiol. 2022;13:886273. doi: 10.3389/fphys.2022.886273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ortiz G.G., Flores Alvarado L.J., Pacheco Moisés F.P., Mireles Ramírez M.A., González Renovato E.D., Sánchez López A.L., Sánchez Romero L., Santoscoy J.F., Velázquez Brizuela I.E., Sánchez González V.J. Cross-talk between glial cells and neurons: Relationship in Multiple Sclerosis. Clin. Case Rep. Rev. 2016;2:565–571. [Google Scholar]
- 16.Kwon H.S., Koh S.H. Neuroinflammation in neurodegenerative disorders: The roles of microglia and astrocytes. Transl. Neurodegener. 2020;9:42. doi: 10.1186/s40035-020-00221-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Neumann H., Kotter M.R., Franklin R.J.M. Debris clearance by microglia: An essential link between degeneration and regeneration. Brain. 2009;132:288–295. doi: 10.1093/brain/awn109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lull M.E., Block M.L. Microglial Activation and Chronic Neurodegeneration. Neurotherapeutics. 2010;7:354–365. doi: 10.1016/j.nurt.2010.05.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Pires B.R., Silva R.C., Ferreira G.M., Abdelhay E. NF-kappaB: Two Sides of the Same Coin. Genes. 2018;9:24. doi: 10.3390/genes9010024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Tsatsanis C., Androulidaki A., Venihaki M., Margioris A.N. Signalling networks regulating cyclooxygenase-2. Int. J. Biochem. Cell Biol. 2006;38:1654–1661. doi: 10.1016/j.biocel.2006.03.021. [DOI] [PubMed] [Google Scholar]
- 21.Chen C.H., Zhou W., Liu S., Deng Y., Cai F., Tone M., Tone Y., Tong Y., Song W. Increased NF-kB signalling up-regulates BACE1 expression and its therapeutic potential in Alzheimer’s disease. Int. J. Neuropsychopharmacol. 2012;15:77–90. doi: 10.1017/S1461145711000149. [DOI] [PubMed] [Google Scholar]
- 22.Perlmutter L.S., Scott S.A., Barron E., Chui H.C. MHC class II-positive microglia in human brain: Association with alzheimer lesions. J. Neurosci. Res. 1992;33:549–558. doi: 10.1002/jnr.490330407. [DOI] [PubMed] [Google Scholar]
- 23.YOuchi Y., Yoshikawa E., Sekine Y., Futatsubashi M., Kanno T., Ogusu T., Torizuka T. Microglial activation and dopamine terminal loss in early Parkinson’s disease. Ann. Neurol. 2005;57:168–175. doi: 10.1002/ana.20338. [DOI] [PubMed] [Google Scholar]
- 24.Ouchi Y., Yagi S., Yokokura M., Sakamoto M. Neuroinflammation in the living brain of Parkinson’s disease. Park. Relat. Disord. 2009;15:S200–S204. doi: 10.1016/S1353-8020(09)70814-4. [DOI] [PubMed] [Google Scholar]
- 25.Brites D., Vaz A.R. Microglia centered pathogenesis in ALS: Insights in cell interconnectivity. Front. Cell. Neurosci. 2014;8:117. doi: 10.3389/fncel.2014.00117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Tai Y.F., Pavese N., Gerhard A., Tabrizi S.J., Barker R.A., Brooks D.J., Piccini P. Microglial activation in presymptomatic Huntington’s disease gene carriers. Brain. 2007;130:1759–1766. doi: 10.1093/brain/awm044. [DOI] [PubMed] [Google Scholar]
- 27.Singhrao S.K., Neal J.W., Morgan B.P., Gasque P. Increased complement biosynthesis by microglia and complement activation on neurons in Huntington’s disease. Exp. Neurol. 1999;159:362–376. doi: 10.1006/exnr.1999.7170. [DOI] [PubMed] [Google Scholar]
- 28.Kaltschmidt B., Uherek M., Volk B., Baeuerle P.A., Kaltschmidt C. Transcription factor NF-kB is activated in primary neurons by amyloid b peptides and in neurons surrounding early plaques from patients with Alzheimer disease. Proc. Natl. Acad. Sci. USA. 1997;94:2642–2647. doi: 10.1073/pnas.94.6.2642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Patrick A. Baeuerle and Thomas Henkel. Function and activation of NF-kB in the immune system. Annu. Rev. Immunol. 1994;12:141–179. doi: 10.1146/annurev.iy.12.040194.001041. [DOI] [PubMed] [Google Scholar]
- 30.Akhondzadeh S., Noroozian M., Mohammadi M., Ohadinia S., Jamshidi A.H., Khani M. Melissa officinalis extract in the treatment of patients with mild to moderate Alzheimer’s disease: A double blind, randomised, placebo-controlled trial. J. Neurol. Neurosurg. Psychiatry. 2003;74:863–866. doi: 10.1136/jnnp.74.7.863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hishikawa N., Takahashi Y., Amakusa Y., Tanno Y., Tuji Y., Niwa H., Murakami N., Krishna U.K. Effects of turmeric on Alzheimer′s disease with behavioral and psychological symptoms of dementia. AYU Int. Q. J. Res. Ayurveda. 2012;33:499. doi: 10.4103/0974-8520.110524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Morillas-Ruiz J.M., Rubio-Perez J.M., Albaladejo M.D., Zafrilla P., Parra S., Vidal-Guevara M.L. Effect of an antioxidant drink on homocysteine levels in Alzheimer’s patients. J. Neurol. Sci. 2010;299:175–178. doi: 10.1016/j.jns.2010.08.050. [DOI] [PubMed] [Google Scholar]
- 33.Choudhary D., Bhattacharyya S., Bose S. Efficacy and Safety of Ashwagandha (Withania somnifera (L.) Dunal) Root Extract in Improving Memory and Cognitive Functions. J. Diet. Suppl. 2017;14:599–612. doi: 10.1080/19390211.2017.1284970. [DOI] [PubMed] [Google Scholar]
- 34.Tsolaki M., Lazarou E., Kozori M., Petridou N., Tabakis I., Lazarou I., Karakota M., Saoulidis I., Melliou E., Magiatis P. A Randomized Clinical Trial of Greek High Phenolic Early Harvest Extra Virgin Olive Oil in Mild Cognitive Impairment: The MICOIL Pilot Study. J. Alzheimer’s Dis. 2020;78:801–817. doi: 10.3233/JAD-200405. [DOI] [PubMed] [Google Scholar]
- 35.Gul A., Bakht J., Mehmood F. Huperzine-A response to cognitive impairment and task switching deficits in patients with Alzheimer’s disease. J. Chin. Med. Assoc. 2019;82:40–43. doi: 10.1016/j.jcma.2018.07.004. [DOI] [PubMed] [Google Scholar]
- 36.Wang R., Yan H., Tang X.C. Progress in studies of huperzine A, a natural cholinesterase inhibitor from Chinese herbal medicine. Acta Pharmacol. Sin. 2006;27:1–26. doi: 10.1111/j.1745-7254.2006.00255.x. [DOI] [PubMed] [Google Scholar]
- 37.Xu S.S., Cai Z.Y., Qu Z.W., Yang R.M., Cai Y.L., Wang G.Q., Su X.Q., Zhong X.S., Cheng R.Y., Xu W.A., et al. Huperzine-A in capsules and tablets for treating patients with Alzheimer disease. Zhongguo Yao Li Xue Bao. 1999;20:486–490. [PubMed] [Google Scholar]
- 38.Turner R.S., Thomas R.G., Craft S., Van Dyck C.H., Mintzer J., Reynolds B.A., Brewer J.B., Rissman R.A., Raman R., Aisen P.S. Placebo-Controlled trial of resveratrol for Alzheimer disease. Neurology. 2015;85:1383–1391. doi: 10.1212/WNL.0000000000002035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Moussa C., Hebron M., Huang X., Ahn J., Rissman R.A., Aisen P.S., Turner R.S. Resveratrol regulates neuro-inflammation and induces adaptive immunity in Alzheimer’s disease. J. Neuroinflamm. 2017;14:1–10. doi: 10.1186/s12974-016-0779-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hasnaoui N., Jbir R., Mars M., Trifi M., Kamal-Eldin A., Melgarejo P., Hernandez F. Organic acids, sugars, and anthocyanins contents in juices of Tunisian pomegranate fruits. Int. J. Food Prop. 2011;14:741–757. doi: 10.1080/10942910903383438. [DOI] [Google Scholar]
- 41.Akhtar S., Ismail T., Fraternale D., Sestili P. Pomegranate peel and peel extracts: Chemistry and food features. Food Chem. 2015;174:417–425. doi: 10.1016/j.foodchem.2014.11.035. [DOI] [PubMed] [Google Scholar]
- 42.Du L., Li J., Zhang X., Wang L., Zhang W. Pomegranate peel polyphenols inhibits inflammation in LPS-induced RAW264.7 macrophages via the suppression of MAPKs activation. J. Funct. Foods. 2018;43:62–69. doi: 10.1016/j.jff.2018.01.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Mousavinejad G., Emam-Djomeh Z., Rezaei K., Khodaparast M.H.H. Identification and quantification of phenolic compounds and their effects on antioxidant activity in pomegranate juices of eight Iranian cultivars. Food Chem. 2009;115:1274–1278. doi: 10.1016/j.foodchem.2009.01.044. [DOI] [Google Scholar]
- 44.Mottaghipisheh J., Ayanmanesh M., Babadayei-Samani R., Javid A., Sanaeifard M., Vitalini S., Iriti M. Total anthocyanin, flavonoid, polyphenol and tannin contents of seven pomegranate cultivars grown in Iran [pdf] Acta Sci. Pol. Technol. Aliment. 2018;17:211–217. doi: 10.17306/J.AFS.2018.0584. [DOI] [PubMed] [Google Scholar]
- 45.Fischer U.A., Carle R., Kammerer D.R. Identification and quantification of phenolic compounds from pomegranate (Punica granatum L.) peel, mesocarp, aril and differently produced juices by HPLC-DAD-ESI/MSn. Food Chem. 2011;127:807–821. doi: 10.1016/j.foodchem.2010.12.156. [DOI] [PubMed] [Google Scholar]
- 46.Gardeli C., Varela K., Krokida E., Mallouchos A. Investigation of Anthocyanins Stability from. Medicines. 2019;6:90. doi: 10.3390/medicines6030090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Russo M., Fanali C., Tripodo G., Dugo P., Muleo R., Dugo L., De Gara L., Mondello L. Analysis of phenolic compounds in different parts of pomegranate (Punica granatum) fruit by HPLC-PDA-ESI/MS and evaluation of their antioxidant activity: Application to different Italian varieties. Anal. Bioanal. Chem. 2018;410:3507–3520. doi: 10.1007/s00216-018-0854-8. [DOI] [PubMed] [Google Scholar]
- 48.Ricci D., Giamperi L., Bucchini A., Fraternale D. Antioxidant activity of Punica granatum fruits. Fitoterapia. 2006;77:310–312. doi: 10.1016/j.fitote.2006.01.008. [DOI] [PubMed] [Google Scholar]
- 49.Elfalleh W. Total phenolic contents and antioxidant activities of pomegranate peel, seed, leaf and flower. J. Med. Plants Res. 2012;6:4724–4730. doi: 10.5897/JMPR11.995. [DOI] [Google Scholar]
- 50.Ben Nasr C., Ayed N., Metche M. Quantitative determination of the polyphenolic content of pomegranate peel. Z. Fur Leb. Unters. Und Forsch. 1996;203:374–378. doi: 10.1007/BF01231077. [DOI] [PubMed] [Google Scholar]
- 51.Zeghad N., Ahmed E., Belkhiri A., Vander Heyden Y., Demeyer K. Antioxidant activity of Vitis vinifera, Punica granatum, Citrus aurantium and Opuntia ficus indica fruits cultivated in Algeria. Heliyon. 2019;5:e01575. doi: 10.1016/j.heliyon.2019.e01575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Sabraoui T., Khider T., Nasser B., Eddoha R., Moujahid A., Benbachir M., Essamadi A. Determination of Punicalagins Content, Metal Chelating, and Antioxidant Properties of Edible Pomegranate (Punica granatum L.) Peels and Seeds Grown in Morocco. Int. J. Food Sci. 2020;2020:4–11. doi: 10.1155/2020/8885889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Amjad L., Shafighi M. Evaluation of Antioxidant Activity, Phenolic and Flavonoid Content in Punica granatum var. Isfahan Malas Flowers. Int. J. Agric. Crop Sci. 2013;5:1133–1139. [Google Scholar]
- 54.Derakhshan Z., Ferrante M., Tadi M., Ansari F., Heydari A., Hosseini M.S., Conti G.O., Sadrabad E.K. Antioxidant activity and total phenolic content of ethanolic extract of pomegranate peels, juice and seeds. Food Chem. Toxicol. 2018;114:108–111. doi: 10.1016/j.fct.2018.02.023. [DOI] [PubMed] [Google Scholar]
- 55.Li R., Chen X.G., Jia K., Liu Z.P., Peng H.Y. A systematic determination of polyphenols constituents and cytotoxic ability in fruit parts of pomegranates derived from five Chinese cultivars. Springerplus. 2016;5:914. doi: 10.1186/s40064-016-2639-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Belkacem N., Djaziri R., Lahfa F., El-Haci I.A., Boucherit Z. Screening phytochimique et activité antioxidante in vitro de différents extraits de l’épicarpe de Punica granatum L. d’Algérie: Étude comparative. Phytotherapie. 2014;12:372–379. doi: 10.1007/s10298-014-0850-x. [DOI] [Google Scholar]
- 57.Singh M., Jha A., Kumar A., Hettiarachchy N., Rai A.K., Sharma D. Influence of the solvents on the extraction of major phenolic compounds (Punicalagin, ellagic acid and gallic acid) and their antioxidant activities in pomegranate aril. J. Food Sci. Technol. 2014;51:2070–2077. doi: 10.1007/s13197-014-1267-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Kim H., Banerjee N., Ivanov I., Pfent C.M., Prudhomme K.R., Bisson W.H., Dashwood R.H., Talcott S.T., Mertens-Talcott S.U. Comparison of anti-inflammatory mechanisms of mango (Mangifera Indica L.) and pomegranate (Punica Granatum L.) in a preclinical model of colitis. Mol. Nutr. Food Res. 2016;60:1912–1923. doi: 10.1002/mnfr.201501008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Türkyilmaz M. Anthocyanin and organic acid profiles of pomegranate (Punica granatum L.) juices from registered varieties in Turkey. Int. J. Food Sci. Technol. 2013;48:2086–2095. doi: 10.1111/ijfs.12190. [DOI] [Google Scholar]
- 60.Fawole O.A., Opara U.L. Changes in physical properties, chemical and elemental composition and antioxidant capacity of pomegranate (cv. Ruby) fruit at five maturity stages. Sci. Hortic. Amst. 2013;150:37–46. doi: 10.1016/j.scienta.2012.10.026. [DOI] [Google Scholar]
- 61.Fawole O.A., Opara U.L. Effects of maturity status on biochemical content, polyphenol composition and antioxidant capacity of pomegranate fruit arils (cv. ’Bhagwa’) South Afr. J. Bot. 2013;85:23–31. doi: 10.1016/j.sajb.2012.11.010. [DOI] [Google Scholar]
- 62.de Pascual-Teresa S., Santos-Buelga C., Rivas-Gonzalo J.C. Quantitative analysis of flavan-3-ols in Spanish foodstuffs and beverages. J. Agric. Food Chem. 2000;48:5331–5337. doi: 10.1021/jf000549h. [DOI] [PubMed] [Google Scholar]
- 63.Gómez-Caravaca A.M., Verardo V., Toselli M., Segura-Carretero A., Fernández-Gutiérrez A., Caboni M.F. Determination of the major phenolic compounds in pomegranate juices by HPLC-DAD-ESI-MS. J. Agric. Food Chem. 2013;61:5328–5337. doi: 10.1021/jf400684n. [DOI] [PubMed] [Google Scholar]
- 64.Van Elswijk D.A., Schobel U.P., Lansky E.P., Irth H., Van Der Greef J. Rapid dereplication of estrogenic compounds in pomegranate (Punica granatum) using on-line biochemical detection coupled to mass spectrometry. Phytochemistry. 2004;65:233–241. doi: 10.1016/j.phytochem.2003.07.001. [DOI] [PubMed] [Google Scholar]
- 65.Noda Y., Kaneyuki T., Mori A., Packer L. Antioxidant activities of pomegranate fruit extract and its anthocyanidins: Delphinidin, cyanidin, and pelargonidin. J. Agric. Food Chem. 2002;50:166–171. doi: 10.1021/jf0108765. [DOI] [PubMed] [Google Scholar]
- 66.Hernandez F., Melgarejo P., Tomas-Barberan F.A., Artes F. Evolution of juice anthocyanins during ripening of new selected pomegranate (Punica granatum) clones. Eur. Food Res. Technol. 1999;210:39–42. doi: 10.1007/s002170050529. [DOI] [Google Scholar]
- 67.Ali M., Sharma N. Phytochemical investigation of the flowers of Punica granatum. Indian J. Chem. Sect. B Org. Med. Chem. 2006;45:1681–1685. doi: 10.1002/chin.200648207. [DOI] [Google Scholar]
- 68.Negi P.S., Jayaprakasha G.K., Jena B.S. Antioxidant and antimutagenic activities of pomegranate peel extracts. Food Chem. 2003;80:393–397. doi: 10.1016/S0308-8146(02)00279-0. [DOI] [Google Scholar]
- 69.Maleki S.J., Crespo J.F., Cabanillas B. Anti-inflammatory effects of flavonoids. Food Chem. 2019;299:125124. doi: 10.1016/j.foodchem.2019.125124. [DOI] [PubMed] [Google Scholar]
- 70.Aarabi A., Barzegar M., Azizi M.H. Effect of cultivar and cold storage of pomegranate (Punica granatum L.) Juices on organic acid composition. Int. Food Res. J. 2008;15:45–55. [Google Scholar]
- 71.Garima P., Akoh C.C. Antioxidant capacity and lipid characterization of six georgia-grown pomegranate cultivars. J. Agric. Food Chem. 2009;57:9427–9436. doi: 10.1021/jf901880p. [DOI] [PubMed] [Google Scholar]
- 72.Poyrazoğlu E., Gökmen V., Artιk N. Organic Acids and Phenolic Compounds in Pomegranates (Punica granatum L.) Grown in Turkey. J. Food Compos. Anal. 2002;15:567–575. doi: 10.1016/S0889-1575(02)91071-9. [DOI] [Google Scholar]
- 73.Wang R., Wang W., Wang L., Liu R., Ding Y., Du L. Constituents of the flowers of Punica granatum. Fitoterapia. 2006;77:534–537. doi: 10.1016/j.fitote.2006.06.011. [DOI] [PubMed] [Google Scholar]
- 74.Gil M.I., Tomás-Barberán F.A., Hess-Pierce B., Holcroft D.M., Kader A.A. Antioxidant activity of pomegranate juice and its relationship with phenolic composition and processing. J. Agric. Food Chem. 2000;48:4581–4589. doi: 10.1021/jf000404a. [DOI] [PubMed] [Google Scholar]
- 75.El-Toumy S.A.A., Rauwald H.W. Two new ellagic acid rhamnosides from Punica granatum heartwood. Planta Med. 2003;69:682–684. doi: 10.1055/s-2003-41107. [DOI] [PubMed] [Google Scholar]
- 76.Nawwar M.A., Hussein S.A., Merfort I. NMR spectral analysis of polyphenols from Punica granatum. Phytochemistry. 1994;36:793–798. doi: 10.1016/S0031-9422(00)89820-9. [DOI] [Google Scholar]
- 77.Tanaka T., Nonaka G.I., Nishioka I. Tannins and related compounds.XL1. Revision of the structures of Punicalin and Punicalagin and isolation and charcterization of 2-O-Galloylpunicalin from the Bark of Punica granatum L. Chem. Pharm. Bull. 1986;34:650–655. doi: 10.1248/cpb.34.650. [DOI] [Google Scholar]
- 78.Huang T.H.W., Peng G., Kota B.P., Li G.Q., Yamahara J., Roufogalis B.D., Li Y. Anti-diabetic action of Punica granatum flower extract: Activation of PPAR-γ and identification of an active component. Toxicol. Appl. Pharmacol. 2005;207:160–169. doi: 10.1016/j.taap.2004.12.009. [DOI] [PubMed] [Google Scholar]
- 79.Wang R.F., Xie W.D., Zhang Z., Xing D.M., Ding Y., Wang W., Ma C., Du L.J. Bioactive compounds from the seeds of Punica granatum (pomegranate) J. Nat. Prod. 2004;67:2096–2098. doi: 10.1021/np0498051. [DOI] [PubMed] [Google Scholar]
- 80.Kulkarni A.P., Aradhya S.M., Divakar S. Isolation and identification of a radical scavenging antioxidant-Punicalagin from pith and carpellary membrane of pomegranate fruit. Food Chem. 2004;87:551–557. doi: 10.1016/j.foodchem.2004.01.006. [DOI] [Google Scholar]
- 81.Ishimoto H., Shibata M., Myojin Y., Ito H., Sugimoto Y., Tai A., Hatano T. In vivo anti-inflammatory and antioxidant properties of ellagitannin metabolite urolithin A. Bioorg. Med. Chem. Lett. 2011;21:5901–5904. doi: 10.1016/j.bmcl.2011.07.086. [DOI] [PubMed] [Google Scholar]
- 82.Yuan T., Ma H., Liu W., Niesen D.B., Shah N., Crews R., Rose K.N., Vattem D.A., Seeram N.P. Pomegranate’s Neuroprotective Effects against Alzheimer’s Disease Are Mediated by Urolithins, Its Ellagitannin-Gut Microbial Derived Metabolites. ACS Chem. Neurosci. 2016;7:26–33. doi: 10.1021/acschemneuro.5b00260. [DOI] [PubMed] [Google Scholar]
- 83.Rogers J., Luber-Narod J., Styren S.D., Civin W.H. Expression of immune system-associated antigens by cells of the human central nervous system: Relationship to the pathology of Alzheimer’s disease. Neurobiol. Aging. 1988;9:339–349. doi: 10.1016/S0197-4580(88)80079-4. [DOI] [PubMed] [Google Scholar]
- 84.Shukla M., Gupta K., Rasheed Z., Khan K.A., Haqqi T.M. Consumption of hydrolyzable tannins-rich pomegranate extract suppresses inflammation and joint damage in rheumatoid arthritis. Nutrition. 2008;24:733–743. doi: 10.1016/j.nut.2008.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Husari A., Hashem Y., Bitar H., Dbaibo G., Zaatari G., El Sabban M. Antioxidant activity of pomegranate juice reduces emphysematous changes and injury secondary to cigarette smoke in an animal model and human alveolar cells. Int. J. COPD. 2016;11:227–237. doi: 10.2147/COPD.S97027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Bachoual R., Talmoudi W., Boussetta T., Braut F., El-Benna J. An aqueous pomegranate peel extract inhibits neutrophil myeloperoxidase in vitro and attenuates lung inflammation in mice. Food Chem. Toxicol. 2011;49:1224–1228. doi: 10.1016/j.fct.2011.02.024. [DOI] [PubMed] [Google Scholar]
- 87.Asghari G., Sheikholeslami S., Mirmiran P., Chary A., Hedayati M., Shafiee A., Azizi F. Effect of pomegranate seed oil on serum TNF-α level in dyslipidemic patients. Int. J. Food Sci. Nutr. 2012;63:368–371. doi: 10.3109/09637486.2011.631521. [DOI] [PubMed] [Google Scholar]
- 88.Rasheed Z., Akhtar N., Anbazhagan A.N., Ramamurthy S., Shukla M., Haqqi T.M. Polyphenol-rich pomegranate fruit extract (POMx) suppresses PMACI-induced expression of pro-inflammatory cytokines by inhibiting the activation of MAP kinases and NF-κB in human KU812 cells. J. Inflamm. 2009;6:1–12. doi: 10.1186/1476-9255-6-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Morzelle M.C., Salgado J.M., Telles M., Mourelle D., Bachiega P., Buck H.S., Viel T.A. Neuroprotective effects of pomegranate peel extract after chronic infusion with amyloid-β peptide in mice. PLoS ONE. 2016;11:e166123. doi: 10.1371/journal.pone.0166123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Braidy N., Essa M.M., Poljak A., Selvaraju S., Al-Adawi S., Manivasagm T., Thenmozhi A.J., Ooi L., Sachdev P., Guillemin G.J. Consumption of pomegranates improves synaptic function in a transgenic mice model of Alzheimer’s disease. Oncotarget. 2016;7:64589–64604. doi: 10.18632/oncotarget.10905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Murphy M.P., Levine H. Alzheimer’s disease and the amyloid-β peptide. J. Alzheimer’s Dis. 2010;19:311–323. doi: 10.3233/JAD-2010-1221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Račková L., Ergin V., Burcu Bali E., Kuniakova M., Karasu Ç. Pomegranate seed oil modulates functions and survival of BV-2 microglial cells in vitro. Int. J. Vitam. Nutr. Res. 2014;84:295–309. doi: 10.1024/0300-9831/a000216. [DOI] [PubMed] [Google Scholar]
- 93.Siddarth P., Li Z., Miller K.J., Ercoli L.M., Merril D.A., Henning S.M., Heber D., Small G.W. Randomized placebo-controlled study of the memory effects of pomegranate juice in middle-aged and older adults. Am. J. Clin. Nutr. 2020;111:170–177. doi: 10.1093/ajcn/nqz241. [DOI] [PubMed] [Google Scholar]
- 94.Gong Z., Huang J., Xu B., Ou Z., Zhang L., Lin X., Ye X., Kong X., Long D., Sun X., et al. Urolithin A attenuates memory impairment and neuroinflammation in APP/PS1 mice. J. Neuroinflamm. 2019;16:62. doi: 10.1186/s12974-019-1450-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Tapias V., Cannon J.R., Greenamyre J.T. Pomegranate juice exacerbates oxidative stress and nigrostriatal degeneration in Parkinson’s disease. Neurobiol. Aging. 2014;35:1162–1176. doi: 10.1016/j.neurobiolaging.2013.10.077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Ali A.A., Kamal M.M., Khalil M.G., Ali S.A., Elariny H.A., Bekhit A., Wahid A. Behavioral, Biochemical and Histopathological effects of Standardised Pomegranate extract with Vinpocetine, Propolis or Cocoa in a rat model of Parkinson’s disease. BioRxiv. 2020 doi: 10.1080/0361073X.2021.1959823. [DOI] [PubMed] [Google Scholar]
- 97.Amer O.S., Dkhil M.A., Hikal W.M., Al-Quraishy S. Antioxidant and anti-inflammatory activities of pomegranate (punica granatum) on eimeria papillata -induced infection in mice. Biomed Res. Int. 2015;2015:219670. doi: 10.1155/2015/219670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Bekir J., Mars M., Vicendo P., Fterrich A., Bouajila J. Chemical composition and antioxidant, anti-inflammatory, and antiproliferation activities of pomegranate (Punica granatum) flowers. J. Med. Food. 2013;16:544–550. doi: 10.1089/jmf.2012.0275. [DOI] [PubMed] [Google Scholar]
- 99.Bekir J., Mars M., Souchard J.P., Bouajila J. Assessment of antioxidant, anti-inflammatory, anti-cholinesterase and cytotoxic activities of pomegranate (Punica granatum) leaves. Food Chem. Toxicol. 2013;55:470–475. doi: 10.1016/j.fct.2013.01.036. [DOI] [PubMed] [Google Scholar]
- 100.Kujawska M., Jourdes M., Kurpik M., Szulc M., Szaefer H., Chmielarz P., Kreiner G., Krajka-Kuźniak V. Neuroprotective Effects of Pomegranate Juice against Parkinson’s Disease and Presence of Ellagitannins- Derived Metabolite—Urolithin A—In the Brain. Int. J. Mol. Sci. 2020;21:202. doi: 10.3390/ijms21010202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Stojanović I., Šavikin K., Đedović N., Živković J., Saksida T., Momčilović M., Koprivica I., Vujičić M., Stanisavljević S., Miljković Đ., et al. Pomegranate peel extract ameliorates autoimmunity in animal models of multiple sclerosis and type 1 diabetes. J. Funct. Foods. 2017;35:522–530. doi: 10.1016/j.jff.2017.06.021. [DOI] [Google Scholar]
- 102.Dendrou C.A., Fugger L., Friese M.A. Immunopathology of multiple sclerosis. Nat. Rev. Immunol. 2015;15:545–558. doi: 10.1038/nri3871. [DOI] [PubMed] [Google Scholar]
- 103.Binyamin O., Larush L., Frid K., Keller G., Friedman-Levi Y., Ovadia H., Abramsky O., Magdassi S., Gabizon R. Treatment of a multiple sclerosis animal model by a novel nanodrop formulation of a natural antioxidant. Int. J. Nanomed. 2015;10:7165–7174. doi: 10.2147/IJN.S179354. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.