

Abstract
Opioid exposure is known to cause transcriptomic changes in the nucleus accumbens (NAc). However, no studies to date have investigated cell type-specific transcriptomic changes associated with volitional opioid taking. Here, we use single nucleus RNA sequencing (snRNAseq) to comprehensively characterize cell type-specific alterations of the NAc transcriptome in rats self-administering morphine. One cohort of male Brown Norway rats was injected with acute morphine (10 mg/kg, i.p.) or saline. A second cohort of rats was allowed to self-administer intravenous morphine (1.0 mg/kg/infusion) for 10 consecutive days. Each morphine-experienced rat was paired with a yoked saline control rat. snRNAseq libraries were generated from NAc punches and used to identify cell type-specific gene expression changes associated with volitional morphine taking. We identified 1106 differentially expressed genes (DEGs) in the acute morphine group, compared to 2453 DEGs in the morphine self-administration group, across 27 distinct cell clusters. Importantly, we identified 1329 DEGs that were specific to morphine self-administration. DEGs were identified in novel clusters of astrocytes, oligodendrocytes, and D1R- and D2R-expressing medium spiny neurons in the NAc. Cell type-specific DEGs included Rgs9, Celf5, Oprm1, and Pde10a. Upregulation of Rgs9 and Celf5 in D2R-expressing neurons was validated by RNAscope. Approximately 85% of all oligodendrocyte DEGs, nearly all of which were associated with morphine taking, were identified in two subtypes. Bioinformatic analyses identified cell type-specific upstream regulatory mechanisms of the observed transcriptome alterations and downstream signaling pathways, including both novel and previously identified molecular pathways. These findings show that volitional morphine taking is associated with distinct cell type-specific transcriptomic changes in the rat NAc and highlight specific striatal cell populations and novel molecular substrates that could be targeted to reduce compulsive opioid taking.
Subject terms: Molecular neuroscience, Addiction
Introduction
Opioid use disorder (OUD) is an ongoing health crisis in the United States for which there is a critical need to understand the molecular basis of compulsive opioid taking [1]. Opioids, including morphine, cause transcriptomic changes throughout the brain. For example, an acute injection of morphine is sufficient to alter the expression of hundreds of genes in the rodent brain [2, 3]. Similarly, repeated, experimenter-delivered morphine has been shown to alter gene expression in multiple brain nuclei, including the nucleus accumbens (NAc) [3–9]. Despite this literature, no studies have investigated cell type-specific transcriptomic changes associated with the escalation of opioid taking, a DSM-5 criteria for diagnosing OUD. Thus, there are significant gaps in our understanding of the molecular mechanisms regulating the transition from acute/recreational opioid taking to compulsive opioid taking.
A notable limitation of gene expression studies to date is the use of bulk lysates. Neural tissue is a heterogenous mix of cell types and bulk transcriptomic approaches lack the resolution to identify changes in gene expression that are specific to individual cell populations. This is particularly problematic in the context of opioids because emerging evidence indicates that repeated opioid exposure induces cell-type-specific effects in the brain. For example, repeated experimenter-delivered morphine injections induce differential gene expression in dopamine receptor type 1 (D1R) and dopamine receptor type 2 (D2R)-expressing medium spiny neurons (MSNs) [10, 11] and astrocytes in the mouse NAc [12]. Thus, there is a critical need to investigate opioid-induced cell type-specific transcriptomic changes to advance our understanding of OUD and identify novel molecular substrates that could be targeted to reduce compulsive opioid taking.
Recent advances in the assay and analysis of neural transcriptomes using single nuclei RNA sequencing (snRNAseq) have provided unprecedented insights into the mRNA landscape of major neural cell types and subpopulations [13–19]. However, no studies have used these approaches to comprehensively characterize cell type-specific gene expression changes in the rat brain following volitional opioid taking. Here, we used snRNAseq to identify cell type-specific alterations of the rat NAc transcriptome after acute morphine exposure and repeated morphine self-administration. We identified cell type-specific differentially expressed genes (DEGs) unique to volitional morphine taking, including gene expression profiles specific to subtypes of astrocytes, oligodendrocytes, and D1R- and D2R-expressing MSNs. We identify cell type-specific affected canonical signaling pathways and gene ontologies, and predicted upstream regulatory mechanisms of the observed volitional morphine taking-induced differential expression. Together, these novel insights into striatal cell-subtype specific mechanisms underlying volitional opioid taking significantly advance our understanding of the molecular basis of OUD.
Methods and materials
Animals and housing
Brown Norway rats are an inbred strain, and their genomic homogeneity minimizes the potential for interindividual neural transcriptome differences. Male Brown Norway rats weighing 175–200 g were obtained from Charles River Laboratories (Wilmington, MA, USA). Rats were housed individually with food and water available ad libitum in their home cages. A reverse 12/12 h light/dark cycle was used with the lights on at 1900 h. All experimental procedures were performed during the dark cycle. The experimental protocols were consistent with the guidelines issued by the National Institutes of Health and were approved by the Institutional Animal Care and Use Committee of the University of Pennsylvania.
Acute morphine exposure
Rats were injected with morphine (10.0 mg/kg, i.p.) or saline two weeks after acclimating to their home cages (an age-matched timepoint that coincided with the initiation of morphine self-administration in the second experimental cohorts of rats described below; Fig. 1A). Rats were euthanized two hours post injection. Whole brains were then dissected, flash frozen in −20 °C isopentane, and stored at −80 °C. The dose of morphine and time course of administration were based on previous rodent studies investigating opioid-induced changes in gene expression in discrete brain nuclei [12, 20, 21].
Fig. 1. Morphine treatment.

Male Brown Norway rats were randomly assigned to the single experimenter-delivered experiment (A) or the chronic self-administration (Self-Admin) experiment (B). In the acute exposure experiment, rats received a single injection of morphine or saline (n = 5 each). In the chronic self-administration experiment, animals were allowed to self-administer morphine in daily test sessions or received yoked saline infusions (n = 6 each). The total number of active lever presses (C), total number of morphine infusions per day (D), and the total morphine infused per day (E). Following morphine and saline treatments, brains were frozen and the nucleus accumbens (F) was punched from coronal sections (G).
Morphine self-administration
A separate group of Brown Norway rats was implanted with indwelling jugular catheters as described in our previous intravenous drug self-administration studies (Fig. 1B) [22–24]. Briefly, rats were handled daily and allowed two weeks to acclimate to their home cages upon arrival and grow to the desired weight needed for surgery. Rats were then anesthetized using ketamine (100 mg/kg) and xylazine (10 mg/kg). An indwelling catheter (SAI Infusion Technologies, Lake Villa, IL) was inserted into the right jugular vein and sutured in place. The catheter was routed to a mesh back mount platform that was implanted subcutaneously dorsal to the shoulder blades. To prevent infection and maintain patency, catheters were flushed daily with 0.2 ml of the antibiotic Timentin (0.93 mg/ml; Fisher, Pittsburgh, PA) dissolved in heparinized 0.9% saline (Butler Schein, Dublin, OH). When not in use, catheters were sealed with plastic obturators.
Following 7 days of recovery, the rats were randomly assigned to one of two groups: morphine-experimental or yoked saline controls. Morphine-experimental rats were placed into operant conditioning chambers and allowed to lever-press for intravenous infusions of morphine (1.0 mg/kg/infusion, infused over 5 s) under a fixed-ratio 1 (FR1) schedule of reinforcement similar to our previous studies of opioid self-administration in rats [22, 23]. The unit dose of morphine was based on our pilot studies as well as previous studies of morphine self-administration in rats [25–27]. All self-administration sessions were 3 hours in duration [25–29] and were conducted over 10 consecutive days. Each morphine infusion was paired with a 20 s contingent light cue illuminated directly above the active lever (i.e., drug-paired lever). A 20 s time-out period followed each morphine infusion, during which time active lever responses were tabulated but had no scheduled consequences. Responses made on the inactive lever, which had no scheduled consequences, were also recorded during the self-administration sessions. Each rat that was allowed to respond for contingent morphine infusions was paired with a yoked rat that received infusions of saline. While lever pressing for the yoked saline rats had no scheduled consequences, these rats received the same number and temporal pattern of infusions as self-administered by their paired morphine-experimental rat. Rats were euthanized immediately after their last self-administration session. Whole brains were dissected, flash frozen in −20 °C isopentane, and stored at −80 °C.
Nuclei isolation, library preparation, and sequencing
Bilateral NAc tissue was punched from frozen brains and nuclei suspensions were prepared, as we described previously (see supplemental methods) [13, 30, 31].
Quality control and clustering of snRNAseq data
Seurat v3.1 was used to merge read count matrices from all samples. Nuclei with low numbers of detected genes (<539) or a high percentage of mitochondrial transcripts (≥5%) were considered low quality and removed. Nuclei with high numbers of UMIs (>11,365) were also removed to minimize the presence of potential multiplets in the data set. All mitochondrial transcripts were subsequently removed from the remaining data, reflecting the use of nuclei instead of whole cells.
Transcript counts were normalized to 10,000 per nucleus and scaled. Genes with highly variable expression and mean scaled expression between 0.003 and 2 (n = 1664) were identified using the mean.var.plot selection method. Principal components (PCs) were generated from these genes and nuclei were clustered in Seurat using the first 50 PCs. Clusters were identified using known cellular markers: Microglia—Arhgap15; Endothelial Cells—Cldn5, Ebf1; Astrocytes—Gja1; Oligodendrocyte Precursor Cells—Pdgfra; Oligodendrocytes—Mag; MSNs—Bcl11b, Drd2, Ebf1, Grm8, Ntng1, Nr4a1; Inhibitory Neurons—Gad1, Sst, Kit, Kcnc2; Cholinergic Neurons—Slc5a7.
Cell populations with mixed or unclear cell type markers, or with markers (e.g., Slc17a7) indicating nuclei of non-NAc origin were removed. A total of 40,223 nuclei were excluded during this step. Normalization, scaling, PC generation, and clustering were redone on the remaining data. An additional three clusters (870 nuclei) were removed due to low numbers of nuclei or lack of representation across samples, for a final total of 190,030 nuclei in 27 distinct cell clusters.
Identification of DEGs
Count data was converted to log2 counts per million. Differential expression between cases and controls in each cluster was analyzed for the acute injection and chronic self-administration groups separately. The R package MAST [32] was used to fit a linear mixed model with fixed effects for treatment group and the number of genes detected in each nucleus (gdr) and a random effect for subject:
Genes expressed in <20% of the nuclei in a cluster were excluded from the analysis. To optimize the random and fixed effects coefficients in the penalized iteratively reweighted least squares step, the integer scalar in the LME4 R package was set equal to zero, as previously described [33]. Gene expression differences associated with morphine compared to saline were identified with MAST using a Likelihood Ratio Test, which tests for differences between the model with and without the effect of the treatment group. P values were corrected for multiple testing using Benjamini-Hochberg correction (FDR = 0.05). Genes with at least a 10% difference in expression between the morphine and saline groups (log2 fold change ≥0.14) and an FDR corrected p value < 0.05 were considered differentially expressed.
Enrichment analyses
As in our prior description [30], the overrepresentation of cell-type-specific DEGs in GWAS phenotypes canonical pathways, gene ontologies, hallmark gene sets, microRNA targets, and transcription factor targets was determined by comparing cluster-specific DEGs, from all clusters with at least 50 DEGs in either the acute or chronic treatment group, from both treatment groups to GWAS Catalog Molecular and Signatures Database (MsigDB) v7.0 [34] using FUMA [35] (see supplemental methods).
Fluorescent in situ hybridization
To validate select DEGs identified by snRNAseq, a separate cohort of rats was allowed to self-administer intravenous morphine, with yoked saline controls, as described. Fluorescent in situ hybridization was conducted using the RNAscope, similar to our previously published studies [36, 37]. Data are presented as the average integrated density ± SEM (see supplemental methods).
Results
Volitional morphine taking
Male Brown Norway rats acquired and maintained robust morphine self-administration (Fig. 1C). Rats escalated their daily intake of morphine over the 10-day self-administration phase (Fig. 1D), similar to published studies from our lab and others investigating voluntary opioid taking in rats [22, 23, 38–40]. Rats self-administered almost double the amount of morphine on day 10 versus day 1 of the self-administration phase, and the total morphine infused was similar to previous rat morphine self-administration studies that used a three-hour daily access paradigm (Fig. 1E) [25, 41, 42]. Micropunches of the NAc (Fig. 1F, G) from the morphine-experienced rats and their respective controls were then processed for snRNAseq (see Table S1 for sample IDs).
Identification of novel striatal cell populations by single nuclei RNA sequencing
snRNAseq was performed on NAc samples using the 10x Genomics 3' gene expression assay. Nuclei were sequenced to an average depth of ~506 million reads per sample. We initially identified 293,735 putative nuclei, with average medians of ~2063 genes and ~3883 UMI per nucleus (Table S2). The number of sequencing reads per sample, nuclei per sample, average sequencing reads per nuclei per sample, median genes per nuclei per sample, and median UMI per nuclei per sample were consistent between the experimental and control rats in the acute and repeated morphine exposure groups (all Mann–Whitney U p values > 0.47; Fig. S1 and Table S2). Examination of sequencing-derived surrogates of RNA quality from the CellRanger output data showed no difference in the fraction of reads mapped confidently to the genome, intergenic regions, exonic regions, or the transcriptome between experimental and control rats in both the acute and repeated morphine exposure groups (all Mann–Whitney U p values > 0.13; Fig. S2 and Table S3). After quality controls (see Methods), 190,030 nuclei remained for downstream analysis.
Nuclei from all treatment groups were clustered together using Seurat, allowing for direct comparisons of the effects of different morphine exposures on the transcriptome of specific NAc cell types and subpopulations. Clustering with Seurat using principal components derived from variable gene expression identified 27 distinct cell clusters (Fig. 2A), which were then annotated using known expression markers for NAc tissue [43–45] (Fig. 2B). Our data set included all major cell types expected in the NAc and a notable number of D1R- and D2R-expressing MSN sub-populations (Fig. 2B). The number of nuclei from all major cell types were comparable between experimental and control rats in each exposure paradigm (Fig. 2C). Nuclei from all treatment groups were represented in each cluster, in terms of acute vs. self-administration treatments and morphine vs. saline groups (Fig. 2D) and from individual samples (Fig. S3).
Fig. 2. snRNAseq and clustering.

A After quality controls, ~190,000 nuclei profiles were used for unbiased clustering in Seurat and are presented as a uniform manifold approximation and projection (UMAP) dimension reduction plot of all nuclei color-coded by cluster. B Clusters were annotated with genes known to be markers for major neural cell types. The size and color of dots are proportional to the percentage of cells expressing the gene (Pct. Exp.) and the average expression level of the gene (Avg. Exp.), respectively. The cluster numbers and colors are matched to that of the UMAP. C The proportion of major cell types in the saline and morphine group for the acute and chronic self-administration (SA) experiment. D The proportion of nuclei in each cluster coming from either the acute or chronic self-administration experiment (left) and the morphine or saline group treatment (right). Clusters are in descending order.
Volitional morphine taking induces unique DEGs
To identify cell type-specific changes in NAc gene expression following acute morphine exposure and volitional morphine taking, snRNAseq data were analyzed for each cell cluster using a linear mixed model in the MAST R package. We found 1106 differential expression events in the acute morphine group (Fig. 3A, Table S4), with cell type-specific DEGs identified in 17 of 27 cell populations. DEGs were 64.6% upregulated and 35.4% were downregulated (Table S5). Similarly, 2,453 DEGs were detected in the morphine self-administration group (Fig. 3B, Table S6), with cell type-specific DEGs in 24 of 27 clusters. DEGs were 67.3% upregulated and 32.7% downregulated (Table S5). Acute morphine exposure and morphine self-administration were associated with a combination of shared and unique DEGs within different cell populations, with cell type-specific differences between upregulated and downregulated genes (Fig. 3C). We identified 514 cell subtype-specific differential expression events shared between the acute morphine and morphine self-administration groups (i.e., a DEG that was detected, in a given cell subtype, in both the acute morphine and morphine self-administration conditions) (Fig. 3C and Table S7). Nearly all shared differential expression events had the same direction of effect on transcript levels between the acute morphine and morphine self-administration treatments, suggesting that these cell subtype-specific transcription alterations were a result of morphine exposure, rather than volitional morphine taking (Fig. 3D). However, there were notable exceptions (Fig. 3D). In the Oligodendrocyte-3 and Oligodendrocyte-4 cell types, Ninj2 was upregulated after acute morphine exposure, but downregulated after volitional morphine taking. Similarly, AC096473.3, a putative isoform of Galr3, was upregulated after acute morphine exposure, but downregulated after volitional morphine taking in the Oligodendrocyte-3 cell type. These divergent expression patterns likely represent transcriptional changes associated with repeated morphine taking.
Fig. 3. Differential expression.
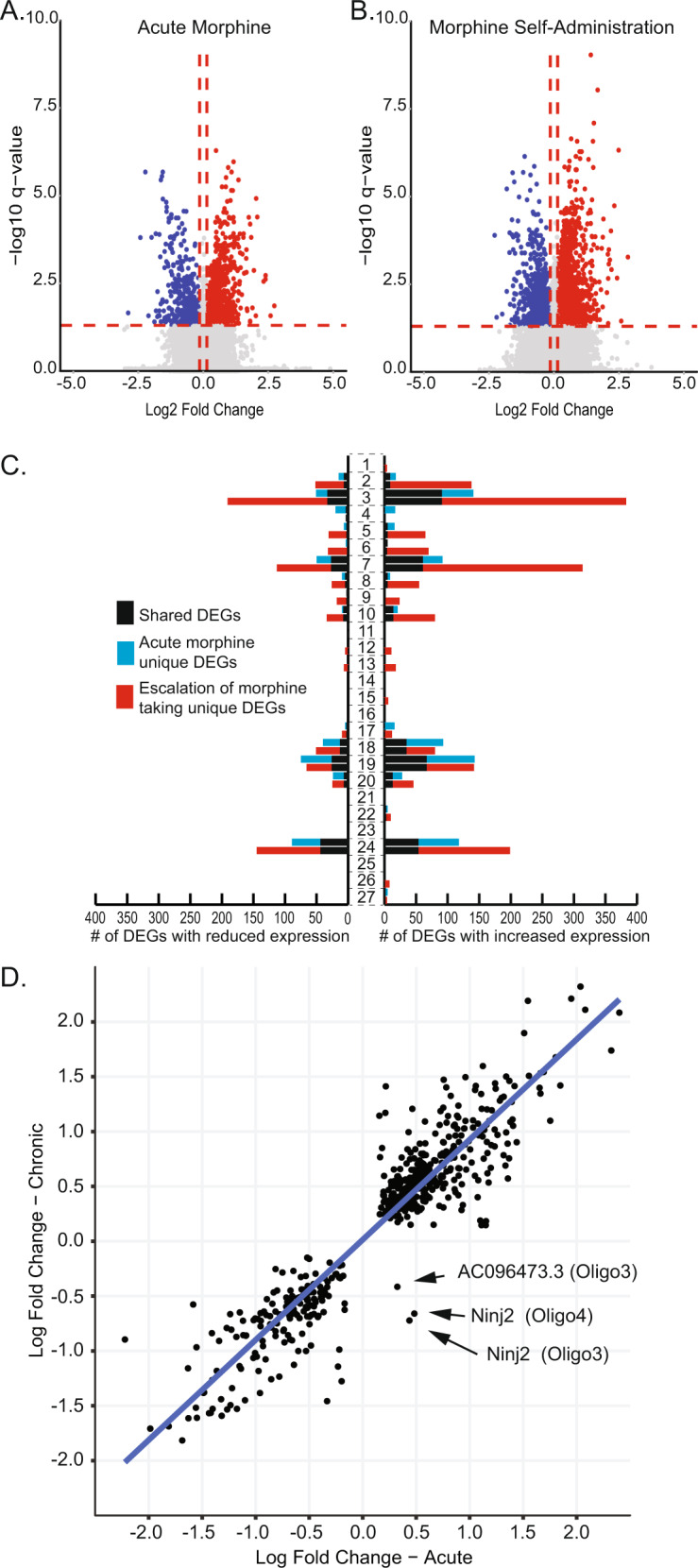
Volcano plots depicting the downregulated (blue) and up regulated (red) differential expressed genes (DEGs) from the acute morphine treatment group (A; n = 1106 DEGs) or the morphine self-administration (B; n = 2453 DEGs) experiment. C A stacked bar graph depicting the number of downregulated (left) or upregulated (right) shared (black bars) and unique (blue or red bars) DEGs per cluster. D Of the 514 differential expression event shared between the acute morphine and morphine self-administration groups, gene expression was altered in the same direction of effect in nearly all instances. Three examples of gene expression alteration changing direction of effect were identified between the acute and self-administration groups.
The majority of DEGs associated with morphine self-administration (54.2%) did not overlap with the acute morphine condition. These cell type-specific transcription alterations included 1,329 differential expression events in 869 unique genes (Table S8). These cell type-specific DEGs unique to volitional morphine taking offer new opportunities to better understand the neurobiological mechanisms underlying morphine self-administration. DEGs unique to volitional morphine taking were not evenly distributed amongst NAc cell types, with some subpopulations accounting for most or all the detected differential expression events within a major cell type (red bars, Fig. 3C; Table S5). To identify DEGs associated specifically with volitional morphine taking and not other aspects of the behavioral procedures (e.g. surgery, repeated handling, stress), we analyzed differential expression between the acute saline and yoked saline control animals. 204 DEGs were identified, with 134 occurring in a single cluster of D1R-expressing MSNs (Table S9). Only a single gene from the saline comparison was also a DEG with the same direction of effect in the morphine self-administration analysis, indicating that the DEGs associated with morphine taking are specifically associated with volitional drug taking. Having observed that differential gene expression patterns specific to volitional morphine taking were associated with distinct striatal cell subtypes, we next sought to define the transcriptome differences that demarcated these cell subtypes (Table S10). In neurons, DEGs specific to self-administration were concentrated in distinct D1R- or D2R-expressing cell subtypes (cell types D1R MSN-2 and D2R MSN-3, respectively; Fig. 4A, B). These neuronal subtypes are clearly defined by distinct patterns of gene expression. In oligodendrocytes, the Oligodendrocyte-4 and Oligodendrocyte-3 subtypes accounted for ~85% of all oligodendrocyte DEGs and nearly all oligodendrocyte DEGs were unique to morphine taking. Like D1R- and D2R-expressing neurons, the oligodendrocyte cell types can be clearly delineated by baseline patterns of gene expression (Fig. 4C). Similarly, the Astrocyte-2 subtype accounted for all astrocytic DEGs and is markedly characterized by a distinct pattern of baseline gene expression (Fig. 4D). Taken together, these studies identify, for the first time, distinct NAc neuronal and glial cell type-specific transcriptomic changes that are associated with volitional morphine taking. Importantly, the identification of these novel NAc cell types, and the transcriptomic markers by which they can be identified in future studies, presents opportunities for further investigating the neurobiological basis of OUD.
Fig. 4. Cell type identification.
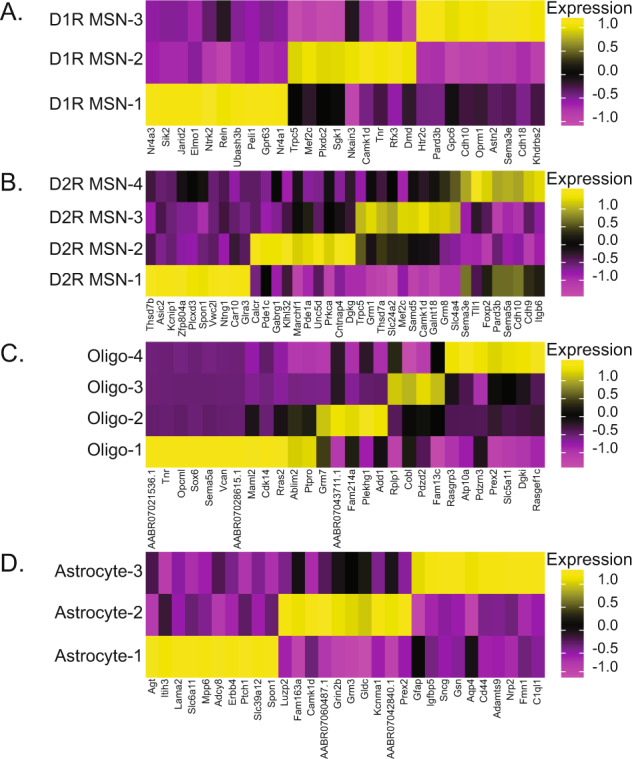
A D1R- and B D2R-expressing neuronal cell types are defined by increased expression of unique genes sets. Similarly, C oligodendrocyte (Oligo) and D astrocyte cell types can be identified by increased expression of unique sets of marker genes.
Different molecular mechanisms regulate transcription following acute morphine exposure versus volitional morphine taking
To understand the regulation of the gene expression patterns induced by acute morphine and morphine self-administration, we next sought to identify transcriptional regulatory mechanisms associated with cell subtype-specific patterns in differential expression. We analyzed cell type-specific DEGs lists from the acute morphine treatment group, the morphine self-administration group, the overlap between these phenotypes, and the cell type-specific DEGs unique to the self-administration group. Cell subtype-specific DEG lists for all groups were compared to MSigDB to identify microRNAs (Table S11) and transcription factors (Table S12) whose target genes were statistically overrepresented amongst the cellular subtype-specific DEGs. The cell type-specific microRNA and transcription factors identified for the acute morphine treatment group and the DEGs overlapping both treatment groups are highly similar, suggesting that these transcriptional regulatory mechanisms are invoked in response to the presence of morphine. For example, miR181 and miR141 were identified as regulators of gene expression in neuronal and glial cell types in response to morphine and are linked to the effects of opioids and the regulation of pain [46–50]. Interestingly, the analysis of the DEGs unique to morphine self-administration identified 28 microRNAs and 52 transcription factors as cell type-specific transcriptome regulatory mechanisms that were unique to volitional morphine taking and suggests that volitional morphine taking induces NAc neuronal and glial cellular subtypes to utilize distinct mechanisms for transcription regulation. These results identify direct regulators of transcriptome alterations. To validate the direct regulators and identify indirect upstream regulators of the observed cell type-specific transcriptome alterations, IPA was used to predict activated and inhibited upstream regulatory network master regulators for D1R-expressing MSNs, D2R-expressing MSNs, oligodendrocytes, and astrocytes for the acute morphine and morphine self-administration groups, and for the DEGs unique to the self-administration group (Table S13). Supporting the microRNA and transcription factor analyses, IPA validated many of the cell type-specific direct regulators of transcription, suggesting face validity of these results. Additionally, the analysis identified shared upstream regulators for the acute morphine and morphine self-administration groups, and upstream regulators specific to volitional morphine taking. Taken together, these results identify cell type-specific mechanisms by which volitional morphine taking alters NAc cellular subtype-specific transcriptomes; providing intriguing targets for interventions.
Volitional morphine intake is associated with cell type-specific neuroadaptations
Observing that acute morphine injection and repeated morphine self-administration altered gene expression in ways that were unique to morphine exposure and volitional morphine taking, we next sought to examine the cell type-specific neurobiological functions affected by these transcriptome alterations. Comparison of cluster-specific DEGs for the acute morphine group, the repeated morphine self-administration group, and the cell type-specific DEGs unique to the self-administration group were conducted. Comparison of cluster-specific DEGs to canonical pathways (Table S14) and gene ontologies (Table S15) identified cell type-specific pathways associated with morphine exposure (i.e., significant in both treatment groups) and pathways unique to volitional morphine taking. For example, the canonical pathways “response to stress”, “response to heat stress”, and “response to external stimuli” were significant in D1R- and D2R-expressing MSNs, oligodendrocyte, and astrocyte clusters in both the acute morphine exposure and repeated morphine self-administration groups, while the canonical pathway “opioid signaling” was significant in a population of D1R-expressing MSNs after morphine self-administration. Additionally, comparison of the cell type specific DEGs to GWAS Catalog identified cell type-specific overrepresentation of genes associated with GWAS phenotypes for both the acute exposure (29 GWAS phenotypes) and repeated self-administration (105 GWAS phenotypes) treatment groups (Table S16). Importantly, morphine self-administration was also associated with cell type-specific GWAS phenotypes (73 GWAS phenotypes) (Table S16). These GWAS phenotypes included “smoking status” and “general risk tolerance”, both of which have been previously associated with substance use disorders. Taken together, these data suggest that the NAc cell subtypes identified in this analysis may represent a portion of the neural circuits associated with these GWAS phenotypes.
Cell-type-specific validation of target genes in D2R-expressing MSNs
To validate cell type-specific differential expression, we utilized florescence in situ hybridization (RNAscope) to quantify changes in the expression of two genes in D2R-expressing MSNs. Given the emerging role of NAc MSN populations in opioid-mediated behaviors [51–53], we identified DEGs specific to morphine self-administration in all four D2R-expressing MSN subtypes. We selected Rgs9 for validation due to its regulation of mu-opioid receptors and considerable literature linking it to addiction-like phenotypes (reviewed [54, 55]). We also selected a gene with no known role in opioid taking, Celf5, the protein product of which is involved in regulating mRNA splicing [56, 57]. NAc slices from rats self-administering morphine and their yoked saline controls (n = 4; four slices per rat) were imaged following RNAscope. Quantification of FISH revealed significantly increased expression of Rgs9 (Control 414.49 ± 3.53; Morphine Self-administration 510.88 ± 4.73; t(30) = 3.9521, p < 0.001; Fig. 5A) and Celf5 (Control 435.43 ± 10.62; Morphine Self-administration 594.25 ± 15.45; t(30) = 2.0504, p < 0.05; Fig. 5B) in NAc D2R-expressing MSNs of rats self-administering morphine. These results are consistent with the direction of effect observed in the snRNAseq results.
Fig. 5. Fluorescence in situ hybridization.

RNAscope was used to validate the differential expression of Rgs9 (A) and Celf5 (B) in Drd2 expressing medium spiny neurons in rats self-administering morphine or their saline yoked controls. Each gene was assessed in four animals (n = 4 slices each), and example images for the yoked saline or morphine self-administration are presented. Quantification of integrated florescence density (see Methods) confirmed the increased expression of both genes in Drd2 expressing neurons of animals that self-administered morphine. *p < 0.05. ***p < 0.001.
Discussion
We used snRNAseq to comprehensively characterize the cell type-specific longitudinal effects of morphine exposure on NAc transcriptomes. We identified novel transcriptomic changes in unique striatal cell populations that are associated with morphine self-administration. These studies represent the first insight into striatal cell type-specific gene expression patterns associated with volitional morphine taking. Furthermore, we were able to identify cell type-specific regulatory mechanisms and affected neurobiological signaling pathways associated with acute morphine exposure or volitional morphine taking. We believe these data are critical to the field, as they provide novel insights into the neural mechanisms associated with a widely employed behavioral model for OUD.
Although there are no prior snRNAseq studies of morphine self-administration for comparison, single-cell transcriptomics in the NAc following an intraperitoneal morphine injection has previously been studied in mice [43]. Our acute morphine data are consistent with the overall trends observed by Avey et al., who found significant gene expression changes primarily in oligodendrocytes, neurons, and astrocytes. Selective transcriptomically distinct subtypes of these three major cell types were the major sources of DEGs in our analysis of acute morphine as well, including the D1R MSN-2, D2R MSN-3, Oligodendrocyte-3, Oligodendrocyte-4, and Astrocyte-2 populations. Critically, these NAc cell types are defined by their transcriptome (Fig. 4), which serves as cell type-specific markers for follow-up studies investigating the functional role of these striatal cell types in opioid-mediated responses. The variance between the cell type-specific DEGs in this study and prior work [43] is likely due to species and technical differences. The pharmacokinetics of opioids differ between mice and rats, including their half-lives, and different time points were applied for analyses between the studies. Additionally, this analysis utilized the 10x Genomics platform for snRNAseq, compared to Drop-Seq [58].
In the current study, the cell types with the most DEGs were similar between acute morphine and morphine self-administration, with astrocytes, oligodendrocytes, and neuronal populations having the largest number of significant changes in gene expression. While cell type-specific gene expression alterations overlapped the treatment groups, suggesting an enduring neurobiological response to morphine, the majority of DEGs in response to self-administration were not shared with the DEGs for investigator-delivered morphine, suggesting that they are gene expression alterations specific to volitional morphine taking. Furthermore, the majority of oligodendrocyte and astrocyte DEGs were concentrated in Oligodendrocyte-3, Oligodendrocyte-4, and Astrocyte-2 cell subtypes, suggesting that these non-neuronal cells shift their gene expression profiles during the transition from acute morphine exposure to repeated morphine taking. Astrocytes have been implicated in opioid addiction [59], with prior evidence showing that morphine alters astrocytic branching [60], and astrocyte regulation of neural maturation [61] and synapse formation [62]. NAc astrocytes have also been shown to regulate morphine-conditioned place preference and synaptic plasticity through morphine-induced alterations of astrocytic metabolic processes [12]. Taken together, these findings suggest that the Astrocyte-2 population may play a role in opioid reinforcement and provide an exciting target for future research. Altered white matter integrity has also been observed in patients with OUDs [63–65] and these changes have been associated with relapse risk [64]. Given the significant transcriptomic changes observed in the Oligodendrocyte-3 and Oligodendrocyte-4 clusters, these specific cell types may be important to this known neuropathological feature of chronic opioid use. Future studies examining additional time points between first exposure and 10 days of self-administration could help better define transcriptomic trajectories in the implicated cell types and explain the specific time frames in which genes and pathways are relevant. The alterations in gene expression associated with volitional morphine taking may include altered genes associated with volitional morphine taking and/or the pharmacological effects of repeated morphine exposure. While a yoked morphine group would have controlled for the number and temporal pattern of repeated intravenous morphine infusion, interpreting results from this potential control group would be limited due to the repeated unpredictable stress associated with yoked morphine infusions. Similarly, while repeated experimenter-delivered injections of morphine avoid the unpredictable stress associated with yoked administration, it poorly recapitulates the pharmacokinetics and temporal pattern of intravenous drug self-administration. Thus, a portion of the DEGs in our dataset may be related to the pharmacological effects of repeated morphine exposure and not volitional morphine taking. Furthermore, the data described here were generated using only male rats and potentially relevant sex differences could not be addressed. Future work should focus on this, as well as the potential role of the estrus cycle.
Morphine self-administration resulted in a greater number of DEGs than acute morphine. Many DEGs associated with morphine self-administration have been implicated in opioid phenotypes. For example, expression of Oprm1, the gene encoding the mu-opioid receptor, was downregulated in cluster D2R MSN-2 (cluster #8, Fig. 2A, B). An emerging literature identifies a role for NAc D2R-expressing MSNs in opioid withdrawal [66–68]. Morphine tolerance is regulated by reductions in mu-opioid receptor availability at the cell surface, either by transcriptional regulation or internalization of the receptor [69, 70]. Studies have reported downregulation of Oprm1 mRNA following chronic experimenter-delivered morphine due to either promoter hypermethylation [71] or an increase in miRNAs that cause degradation of Oprm1 transcript [72]. Our data suggest that morphine tolerance in the D2R MSN-2 cell type is regulated at the level of transcription, by the downregulation of Oprm1 transcripts. Whether the morphine-induced downregulation of Oprm1 in D2R MSN-2 is regulated by promoter epigenetics or miRNA expression remains an open question and an enticing target for future studies. Furthermore, the lack of Oprm1 gene expression changes amongst the other three D2R-expressing MSN cell types indicates that they may regulate morphine tolerance by internalization of the receptor or that they may not play a morphine tolerance. These potential mechanisms should be further explored in the context of D2R-expressing MSN function and opioid-mediated behaviors.
In contrast to Oprm1, Rgs9 was upregulated in all four D2R-expressing MSN subtypes (clusters #6–9, Fig. 2A, B) following volitional morphine taking. This result was validated in the second cohort of rats (Fig. 5A). A splice variant of Rgs9, known as Rgs9-2, is highly expressed in the NAc and involved in opioid reward and tolerance [55, 73, 74]. Constitutive Rgs9 knockout alters the rewarding effects of morphine, as well as morphine-induced analgesic responses [55]. Furthermore, optogenetic activation of Rgs9+ neurons in the NAc increased development of morphine tolerance [73] and data suggest this tolerance is related to interactions between Rgs9 and G-protein subunits [75, 76]. Taken together, these data suggest that regulation of Rgs9 gene expression in NAc D2R-expresssing MSNs is a ubiquitous mechanism of regulating opioid reward and tolerance. Pde10a was also upregulated after morphine self-administration in Sst+ interneurons (cluster #13) and Astrocyte-2 (cluster #24, Fig. 2A, B). Inhibiting Pde10a has been shown to suppress the acquisition of morphine-conditioned place preference in rats [77]. Pde10a inhibition also facilitated the extinction of conditioned place preference, suggesting that Pde10a is involved in morphine-induced learning and memory [77]. These results suggest that inhibiting volitional morphine taking induced an increase of Pde10a in the Sst+ interneuron cell type, the Astrocyte-2 cell type, or both cell types could be a potential future pharmacological target to reduce morphine conditioned responses. In total, the identification of cell type-specific DEGs associated with morphine self-administration in genes previously associated with opioid phenotypes provides face validity for our findings and suggests that the cell type-specific alterations in gene expression represent opportunities for mechanistic study.
The DEGs lists were enriched for large numbers of different GWAS hits, gene ontology (GO) terms, and pathways. While acute morphine exposure and morphine self-administration shared many of these results (e.g., genes related to stress response), there were also distinct findings for volitional morphine taking. For example, cell type-specific microRNA and transcription factors that are putative regulators of DEGs associated with morphine self-administration were identified. Similarly, the GO terms “regulation of glutamate receptor signaling pathway” and “regulation of NMDA receptor activity”, which were driven in part by the presence of Oprm1 and Rgs9 in the DEGs of D2R MSN-2 in response to volitional morphine taking. NMDA receptors directly interact with the mu-opioid receptor in the striatum and other brain regions [78]. Furthermore, NMDA receptor antagonists inhibit the development of morphine tolerance without blocking the analgesic effects of acute morphine injection [79]. Studies also suggest NMDA receptor antagonists may be useful in reducing symptoms associated with opioid withdrawal [80–82]. These previous findings fit well with the results of the present study and support the idea that alterations to NMDA receptor activity occur in the context of volitional morphine taking, but not acute morphine exposure.
In conclusion, we identified unique cell type-specific changes in the NAc transcriptome that were associated with volitional morphine taking. These DEGs may represent possible cell type-specific molecular targets for novel therapeutic approaches to treating OUD. Future studies investigating the functional significance of these genes will be required to understand the therapeutic potential of these targets. Future snRNAseq studies can also extend our findings in a number of directions. Analyses of tissue from rats self-administering different opioids may reveal differences in cell type-specific effects between common opioids of abuse. Studying single nuclei transcriptomics during morphine withdrawal and the reinstatement of opioid seeking, a model of relapse, will also expand our understanding of the longitudinal transcriptomic changes that promote relapse during acute and prolonged withdraw.
Supplementary information
Acknowledgements
This work was supported in part by a State of Pennsylvania Department of Health Nonformula Tobacco Settlement Act Grant, Pharmacogenetics of Opioid Use Disorder (W.H.B.), R01 DA044015 (W.H.B.), R01 DK105155 (M.R.H.), R01 DA037897 (H.D.S.) and R21 DA045792 (H.D.S.) H.D.S. was also supported by the Dr. Dorothy Mereness Endowed Research Fund through a Faculty Grant Award in the School of Nursing. H.D.S and J.B.N. were supported by the Grant for Faculty Mentoring Undergraduate Research (GfFMUR) at the University of Pennsylvania.
Author contributions
Conceptualization: BCR, WHB, HDS, RCC. Methodology: BCR, LMS, HDS, RCC. Software: BCR, LMS, EDP, RCC. Validation: BCR, LMS, MRH, RCC. Formal Analysis: BCR, YZ, LMS, EDP, HDS, RCC. Investigation: BCR, YZ, LMS, GA, JBN, KR. Resources: BCR, YZ, LMS, MRH, WHB, HDS, RCC. Data Curation: BCR, YZ, LMS, EDP, HDS, RCC. Writing—original draft: BCR, RCC. Writing—review & editing: BCR, YZ, LMS, EDP, GA, JBN, KR, MRH, TNF, WHB, HDS, RCC. Visualization: BCR, LMS, EDP, HDS, RCC. Supervision: BCR, MRH, WHB, HDS, RCC. Project administration: BCR, HDS, RCC. Funding acquisition: BCR, MRH, TNF, WHB, HDS, RCC.
Data availability
Single-nuclei RNA-sequencing data are available at the NCBI Gene Expression Omnibus (GEO) under accession number GSE171165.
Competing interests
BCR and MRH receive research funding from Novo Nordisk and Boehringer Ingelheim that was not used in support of these studies. MRH receives research funding from Eli Lilly & Co. that was not used in support of these studies. MRH is CEO of Cantius Therapeutics, LLC which pursues biological work unrelated to the current study. All other authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
These authors contributed equally: Heath D. Schmidt, Richard C. Crist.
Supplementary information
The online version contains supplementary material available at 10.1038/s41398-022-02135-1.
References
- 1.Volkow ND, Blanco C. The changing opioid crisis: development, challenges and opportunities. Mol Psychiatry. 2021;26:218–33. doi: 10.1038/s41380-020-0661-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Loguinov AV, Anderson LM, Crosby GJ, Yukhananov RY. Gene expression following acute morphine administration. Physiol Genomics. 2001;6:169–81. doi: 10.1152/physiolgenomics.2001.6.3.169. [DOI] [PubMed] [Google Scholar]
- 3.Korostynski M, Piechota M, Kaminska D, Solecki W, Przewlocki R. Morphine effects on striatal transcriptome in mice. Genome Biol. 2007;8:R128. doi: 10.1186/gb-2007-8-6-r128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Albertson DN, Schmidt CJ, Kapatos G, Bannon MJ. Distinctive profiles of gene expression in the human nucleus accumbens associated with cocaine and heroin abuse. Neuropsychopharmacology. 2006;31:2304–12. doi: 10.1038/sj.npp.1301089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Heller EA, Kaska S, Fallon B, Ferguson D, Kennedy PJ, Neve RL, et al. Morphine and cocaine increase serum- and glucocorticoid-inducible kinase 1 activity in the ventral tegmental area. J Neurochem. 2015;132:243–53. doi: 10.1111/jnc.12925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Marie-Claire C, Courtin C, Robert A, Gidrol X, Roques BP, Noble F. Sensitization to the conditioned rewarding effects of morphine modulates gene expression in rat hippocampus. Neuropharmacology. 2007;52:430–5. doi: 10.1016/j.neuropharm.2006.08.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sanchis-Segura C, Lopez-Atalaya JP, Barco A. Selective boosting of transcriptional and behavioral responses to drugs of abuse by histone deacetylase inhibition. Neuropsychopharmacology. 2009;34:2642–54. doi: 10.1038/npp.2009.125. [DOI] [PubMed] [Google Scholar]
- 8.Tapocik JD, Letwin N, Mayo CL, Frank B, Luu T, Achinike O, et al. Identification of candidate genes and gene networks specifically associated with analgesic tolerance to morphine. J Neurosci. 2009;29:5295–307. doi: 10.1523/JNEUROSCI.4020-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Seney ML, Kim SM, Glausier JR, Hildebrand MA, Xue X, Zong W, et al. Transcriptional alterations in dorsolateral prefrontal cortex and nucleus accumbens implicate neuroinflammation and synaptic remodeling in opioid use disorder. Biol Psychiatry. 2021;90:550–62. doi: 10.1016/j.biopsych.2021.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hearing MC, Jedynak J, Ebner SR, Ingebretson A, Asp AJ, Fischer RA, et al. Reversal of morphine-induced cell-type-specific synaptic plasticity in the nucleus accumbens shell blocks reinstatement. Proc Natl Acad Sci USA. 2016;113:757–62. doi: 10.1073/pnas.1519248113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Madayag AC, Gomez D, Anderson EM, Ingebretson AE, Thomas MJ, Hearing MC. Cell-type and region-specific nucleus accumbens AMPAR plasticity associated with morphine reward, reinstatement, and spontaneous withdrawal. Brain Struct Funct. 2019;224:2311–24. doi: 10.1007/s00429-019-01903-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Skupio U, Tertil M, Bilecki W, Barut J, Korostynski M, Golda S, et al. Astrocytes determine conditioned response to morphine via glucocorticoid receptor-dependent regulation of lactate release. Neuropsychopharmacology. 2020;45:404–15. doi: 10.1038/s41386-019-0450-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Borner T, Geisler CE, Fortin SM, Cosgrove R, Alsina-Fernandez JA, Dogra M, et al. GIP receptor agonism attenuates GLP-1 receptor agonist induced nausea and emesis in preclinical models. Diabetes 2021;70:2545–53. [DOI] [PMC free article] [PubMed]
- 14.He L, Vanlandewijck M, Mae MA, Andrae J, Ando K, Del Gaudio F, et al. Single-cell RNA sequencing of mouse brain and lung vascular and vessel-associated cell types. Sci Data. 2018;5:180160. doi: 10.1038/sdata.2018.160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lake BB, Ai R, Kaeser GE, Salathia NS, Yung YC, Liu R, et al. Neuronal subtypes and diversity revealed by single-nucleus RNA sequencing of the human brain. Science. 2016;352:1586–90. doi: 10.1126/science.aaf1204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Mathys H, Davila-Velderrain J, Peng Z, Gao F, Mohammadi S, Young JZ, et al. Single-cell transcriptomic analysis of Alzheimer’s disease. Nature. 2019;570:332–7. doi: 10.1038/s41586-019-1195-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nagy C, Maitra M, Tanti A, Suderman M, Theroux JF, Davoli MA, et al. Single-nucleus transcriptomics of the prefrontal cortex in major depressive disorder implicates oligodendrocyte precursor cells and excitatory neurons. Nat Neurosci 2020;23:771–81. [DOI] [PubMed]
- 18.Tepe B, Hill MC, Pekarek BT, Hunt PJ, Martin TJ, Martin JF, et al. Single-cell RNA-seq of mouse olfactory bulb reveals cellular heterogeneity and activity-dependent molecular census of adult-born neurons. Cell Rep. 2018;25:2689–703.e2683. doi: 10.1016/j.celrep.2018.11.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zeisel A, Hochgerner H, Lonnerberg P, Johnsson A, Memic F, van der Zwan J, et al. Molecular architecture of the mouse nervous system. Cell. 2018;174:999–1014 e1022. doi: 10.1016/j.cell.2018.06.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ahtee L, Attila LM, Carlson KR. Augmentation of morphine-induced changes in brain monoamine metabolism after chronic naltrexone treatment. J Pharmacol Exp Ther. 1990;255:803–8. [PubMed] [Google Scholar]
- 21.Piechota M, Korostynski M, Solecki W, Gieryk A, Slezak M, Bilecki W, et al. The dissection of transcriptional modules regulated by various drugs of abuse in the mouse striatum. Genome Biol. 2010;11:R48. doi: 10.1186/gb-2010-11-5-r48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zhang Y, Kahng MW, Elkind JA, Weir VR, Hernandez NS, Stein LM, et al. Activation of GLP-1 receptors attenuates oxycodone taking and seeking without compromising the antinociceptive effects of oxycodone in rats. Neuropsychopharmacology. 2020;45:451–61. doi: 10.1038/s41386-019-0531-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zhang Y, Rahematpura S, Ragnini KH, Moreno A, Stecyk KS, Kahng MW, et al. A novel dual agonist of glucagon-like peptide-1 receptors and neuropeptide Y2 receptors attenuates fentanyl taking and seeking in male rats. Neuropharmacology. 2021;192:108599. doi: 10.1016/j.neuropharm.2021.108599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Schmidt HD, Mietlicki-Baase EG, Ige KY, Maurer JJ, Reiner DJ, Zimmer DJ, et al. Glucagon-like peptide-1 receptor activation in the ventral tegmental area decreases the reinforcing efficacy of cocaine. Neuropsychopharmacology. 2016;41:1917–28. doi: 10.1038/npp.2015.362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lucantonio F, Kambhampati S, Haney RZ, Atalayer D, Rowland NE, Shaham Y, et al. Effects of prior cocaine versus morphine or heroin self-administration on extinction learning driven by overexpectation versus omission of reward. Biol Psychiatry. 2015;77:912–20. doi: 10.1016/j.biopsych.2014.11.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ucha M, Coria SM, Nunez AE, Santos-Toscano R, Roura-Martinez D, Fernandez-Ruiz J, et al. Morphine self-administration alters the expression of translational machinery genes in the amygdala of male Lewis rats. J Psychopharmacol. 2019;33:882–93. doi: 10.1177/0269881119836206. [DOI] [PubMed] [Google Scholar]
- 27.Kruzich PJ, Chen AC, Unterwald EM, Kreek MJ. Subject-regulated dosing alters morphine self-administration behavior and morphine-stimulated [35S]GTPgammaS binding. Synapse. 2003;47:243–9. doi: 10.1002/syn.10173. [DOI] [PubMed] [Google Scholar]
- 28.Neelakantan H, Holliday ED, Fox RG, Stutz SJ, Comer SD, Haney M, et al. Lorcaserin suppresses oxycodone self-administration and relapse vulnerability in rats. ACS Chem Neurosci. 2017;8:1065–73. doi: 10.1021/acschemneuro.6b00413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.You ZB, Bi GH, Galaj E, Kumar V, Cao J, Gadiano A, et al. Dopamine D 3 R antagonist VK4-116 attenuates oxycodone self-administration and reinstatement without compromising its antinociceptive effects. Neuropsychopharmacology 2018;44:1415–24. [DOI] [PMC free article] [PubMed]
- 30.Reiner BC, Crist RC, Borner T, Doyle RP, Hayes MR, De Jonghe BC. Single nuclei RNA sequencing of the rat AP and NTS following GDF15 treatment. Mol Metab. 2022;56:101422. [DOI] [PMC free article] [PubMed]
- 31.Reiner BC, Crist RC, Stein LM, Weller AE, Doyle GA, Arauco-Shapiro G, et al. Single-nuclei transcriptomics of schizophrenia prefrontal cortex primarily implicates neuronal subtypes. BioRxiv 2020; https://www.biorxiv.org/content/10.1101/2020.07.29.227355v2.
- 32.Finak G, McDavid A, Yajima M, Deng J, Gersuk V, Shalek AK, et al. MAST: a flexible statistical framework for assessing transcriptional changes and characterizing heterogeneity in single-cell RNA sequencing data. Genome Biol. 2015;16:278. doi: 10.1186/s13059-015-0844-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Velmeshev D, Schirmer L, Jung D, Haeussler M, Perez Y, Mayer S, et al. Single-cell genomics identifies cell type-specific molecular changes in autism. Science. 2019;364:685–9. doi: 10.1126/science.aav8130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Liberzon A, Subramanian A, Pinchback R, Thorvaldsdottir H, Tamayo P, Mesirov JP. Molecular signatures database (MSigDB) 3.0. Bioinformatics. 2011;27:1739–40. doi: 10.1093/bioinformatics/btr260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Watanabe K, Taskesen E, van Bochoven A, Posthuma D. Functional mapping and annotation of genetic associations with FUMA. Nat Commun. 2017;8:1826. doi: 10.1038/s41467-017-01261-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Hernandez NS, Weir VR, Ragnini K, Merkel R, Zhang Y, Mace K, et al. GLP-1 receptor signaling in the laterodorsal tegmental nucleus attenuates cocaine seeking by activating GABAergic circuits that project to the VTA. Mol Psychiatry 2020;26:4394–408 [DOI] [PMC free article] [PubMed]
- 37.Fortin SM, Lipsky RK, Lhamo R, Chen J, Kim E, Borner T, et al. GABA neurons in the nucleus tractus solitarius express GLP-1 receptors and mediate anorectic effects of liraglutide in rats. Sci Transl Med 2020;12:eaay8071. [DOI] [PMC free article] [PubMed]
- 38.Wade CL, Vendruscolo LF, Schlosburg JE, Hernandez DO, Koob GF. Compulsive-like responding for opioid analgesics in rats with extended access. Neuropsychopharmacology. 2015;40:421–8. doi: 10.1038/npp.2014.188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Vendruscolo LF, Schlosburg JE, Misra KK, Chen SA, Greenwell TN, Koob GF. Escalation patterns of varying periods of heroin access. Pharmacol Biochem Behav. 2011;98:570–4. doi: 10.1016/j.pbb.2011.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Blackwood CA, Leary M, Salisbury A, McCoy MT, Cadet JL. Escalated oxycodone self-administration causes differential striatal mRNA expression of FGFs and IEGs following abstinence-associated incubation of oxycodone craving. Neuroscience. 2019;415:173–83. doi: 10.1016/j.neuroscience.2019.07.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Hou YY, Cai YQ, Pan ZZ. Persistent pain maintains morphine-seeking behavior after morphine withdrawal through reduced MeCP2 repression of GluA1 in rat central amygdala. J Neurosci. 2015;35:3689–700. doi: 10.1523/JNEUROSCI.3453-14.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Ellis AS, Toussaint AB, Knouse MC, Thomas AS, Bongiovanni AR, Mayberry HL, et al. Paternal morphine self-administration produces object recognition memory deficits in female, but not male offspring. Psychopharmacology (Berl) 2020;237:1209–21. doi: 10.1007/s00213-019-05450-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Avey D, Sankararaman S, Yim AKY, Barve R, Milbrandt J, Mitra RD. Single-cell RNA-seq uncovers a robust transcriptional response to morphine by glia. Cell Rep. 2018;24:3619–29.e3614. doi: 10.1016/j.celrep.2018.08.080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Gokce O, Stanley GM, Treutlein B, Neff NF, Camp JG, Malenka RC, et al. Cellular taxonomy of the mouse striatum as revealed by single-cell RNA-seq. Cell Rep. 2016;16:1126–37. doi: 10.1016/j.celrep.2016.06.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Zhang Y, Chen K, Sloan SA, Bennett ML, Scholze AR, O’Keeffe S, et al. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J Neurosci. 2014;34:11929–47. doi: 10.1523/JNEUROSCI.1860-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Dai Z, Chu H, Ma J, Yan Y, Zhang X, Liang Y. The regulatory mechanisms and therapeutic potential of microRNAs: from chronic pain to morphine tolerance. Front Mol Neurosci. 2018;11:80. doi: 10.3389/fnmol.2018.00080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Xu C, Zhang Y, Zheng H, Loh HH, Law PY. Morphine modulates mouse hippocampal progenitor cell lineages by upregulating miR-181a level. Stem Cells. 2014;32:2961–72. doi: 10.1002/stem.1774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Chen J, Liang L, Li Y, Zhang Y, Zhang M, Yang T, et al. Naloxone regulates the differentiation of neural stem cells via a receptor-independent pathway. FASEB J. 2020;34:5917–30. doi: 10.1096/fj.201902873R. [DOI] [PubMed] [Google Scholar]
- 49.Hauser KF, Knapp PE. Opiate drugs with abuse liability hijack the endogenous opioid system to disrupt neuronal and glial maturation in the central nervous system. Front Pediatr. 2017;5:294. doi: 10.3389/fped.2017.00294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Zhang Y, Loh HH, Law PY. Effect of opioid on adult hippocampal neurogenesis. ScientificWorldJournal. 2016;2016:2601264. doi: 10.1155/2016/2601264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.McDevitt DS, Jonik B, Graziane NM. Morphine differentially alters the synaptic and intrinsic properties of D1R- and D2R-expressing medium spiny neurons in the nucleus accumbens. Front Synaptic Neurosci. 2019;11:35. doi: 10.3389/fnsyn.2019.00035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.James AS, Chen JY, Cepeda C, Mittal N, Jentsch JD, Levine MS, et al. Opioid self-administration results in cell-type specific adaptations of striatal medium spiny neurons. Behav Brain Res. 2013;256:279–83. doi: 10.1016/j.bbr.2013.08.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Severino AL, Mittal N, Hakimian JK, Velarde N, Minasyan A, Albert R, et al. mu-opioid receptors on distinct neuronal populations mediate different aspects of opioid reward-related behaviors. eNeuro 2020;7:ENEURO.0146-20.2020. [DOI] [PMC free article] [PubMed]
- 54.Senese NB, Kandasamy R, Kochan KE, Traynor JR. Regulator of G-protein signaling (RGS) protein modulation of opioid receptor signaling as a potential target for pain management. Front Mol Neurosci. 2020;13:5. doi: 10.3389/fnmol.2020.00005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Zachariou V, Georgescu D, Sanchez N, Rahman Z, DiLeone R, Berton O, et al. Essential role for RGS9 in opiate action. Proc Natl Acad Sci USA. 2003;100:13656–61. doi: 10.1073/pnas.2232594100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ladd AN, Charlet N, Cooper TA. The CELF family of RNA binding proteins is implicated in cell-specific and developmentally regulated alternative splicing. Mol Cell Biol. 2001;21:1285–96. doi: 10.1128/MCB.21.4.1285-1296.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Pique L, Martinez de Paz A, Pineyro D, Martinez-Cardus A, Castro de Moura M, Llinas-Arias P, et al. Epigenetic inactivation of the splicing RNA-binding protein CELF2 in human breast cancer. Oncogene. 2019;38:7106–12. doi: 10.1038/s41388-019-0936-x. [DOI] [PubMed] [Google Scholar]
- 58.Campbell JN, Macosko EZ, Fenselau H, Pers TH, Lyubetskaya A, Tenen D, et al. A molecular census of arcuate hypothalamus and median eminence cell types. Nat Neurosci. 2017;20:484–96. doi: 10.1038/nn.4495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Narita M, Suzuki M, Kuzumaki N, Miyatake M, Suzuki T. Implication of activated astrocytes in the development of drug dependence: differences between methamphetamine and morphine. Ann N Y Acad Sci. 2008;1141:96–104. doi: 10.1196/annals.1441.032. [DOI] [PubMed] [Google Scholar]
- 60.Lazriev IL, Kiknadze GI, Kutateladze II, Nebieridze MI. Effect of morphine on the number and branching of astrocytes in various regions of rat brain. Bull Exp Biol Med. 2001;131:248–50. doi: 10.1023/A:1017699315355. [DOI] [PubMed] [Google Scholar]
- 61.Stiene-Martin A, Gurwell JA, Hauser KF. Morphine alters astrocyte growth in primary cultures of mouse glial cells: evidence for a direct effect of opiates on neural maturation. Brain Res Dev Brain Res. 1991;60:1–7. doi: 10.1016/0165-3806(91)90149-D. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Ikeda H, Miyatake M, Koshikawa N, Ochiai K, Yamada K, Kiss A, et al. Morphine modulation of thrombospondin levels in astrocytes and its implications for neurite outgrowth and synapse formation. J Biol Chem. 2010;285:38415–27. doi: 10.1074/jbc.M110.109827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Bora E, Yucel M, Fornito A, Pantelis C, Harrison BJ, Cocchi L, et al. White matter microstructure in opiate addiction. Addict Biol. 2012;17:141–8. doi: 10.1111/j.1369-1600.2010.00266.x. [DOI] [PubMed] [Google Scholar]
- 64.Li W, Zhu J, Li Q, Ye J, Chen J, Liu J, et al. Brain white matter integrity in heroin addicts during methadone maintenance treatment is related to relapse propensity. Brain Behav. 2016;6:e00436. doi: 10.1002/brb3.436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Liu H, Li L, Hao Y, Cao D, Xu L, Rohrbaugh R, et al. Disrupted white matter integrity in heroin dependence: a controlled study utilizing diffusion tensor imaging. Am J Drug Alcohol Abuse. 2008;34:562–75. doi: 10.1080/00952990802295238. [DOI] [PubMed] [Google Scholar]
- 66.Capasso A, Sorrentino L. Differential influence of D1 and D2 dopamine receptors on acute opiate withdrawal in guinea-pig isolated ileum. Br J Pharmacol. 1997;120:1001–6. doi: 10.1038/sj.bjp.0700995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Harris GC, Aston-Jones G. Involvement of D2 dopamine receptors in the nucleus accumbens in the opiate withdrawal syndrome. Nature. 1994;371:155–7. doi: 10.1038/371155a0. [DOI] [PubMed] [Google Scholar]
- 68.Walters CL, Aston-Jones G, Druhan JP. Expression of fos-related antigens in the nucleus accumbens during opiate withdrawal and their attenuation by a D2 dopamine receptor agonist. Neuropsychopharmacology. 2000;23:307–15. doi: 10.1016/S0893-133X(00)00113-5. [DOI] [PubMed] [Google Scholar]
- 69.Koch T, Hollt V. Role of receptor internalization in opioid tolerance and dependence. Pharmacol Ther. 2008;117:199–206. doi: 10.1016/j.pharmthera.2007.10.003. [DOI] [PubMed] [Google Scholar]
- 70.Zuo Z. The role of opioid receptor internalization and beta-arrestins in the development of opioid tolerance. Anesth Analg. 2005;101:728–34. doi: 10.1213/01.ANE.0000160588.32007.AD. [DOI] [PubMed] [Google Scholar]
- 71.Chorbov VM, Todorov AA, Lynskey MT, Cicero TJ. Elevated levels of DNA methylation at the OPRM1 promoter in blood and sperm from male opioid addicts. J Opioid Manag. 2011;7:258–64. doi: 10.5055/jom.2011.0067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Wu Q, Hwang CK, Zheng H, Wagley Y, Lin HY, Kim DK, et al. MicroRNA 339 down-regulates mu-opioid receptor at the post-transcriptional level in response to opioid treatment. FASEB J. 2013;27:522–35. doi: 10.1096/fj.12-213439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Gaspari S, Papachatzaki MM, Koo JW, Carr FB, Tsimpanouli ME, Stergiou E, et al. Nucleus accumbens-specific interventions in RGS9-2 activity modulate responses to morphine. Neuropsychopharmacology. 2014;39:1968–77. doi: 10.1038/npp.2014.45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Gaspari S, Cogliani V, Manouras L, Anderson EM, Mitsi V, Avrampou K, et al. RGS9-2 modulates responses to oxycodone in pain-free and chronic pain states. Neuropsychopharmacology. 2017;42:1548–56. doi: 10.1038/npp.2017.4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Psifogeorgou K, Terzi D, Papachatzaki MM, Varidaki A, Ferguson D, Gold SJ, et al. A unique role of RGS9-2 in the striatum as a positive or negative regulator of opiate analgesia. J Neurosci. 2011;31:5617–24. doi: 10.1523/JNEUROSCI.4146-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Garzon J, Rodriguez-Munoz M, Lopez-Fando A, Sanchez-Blazquez P. Activation of mu-opioid receptors transfers control of Galpha subunits to the regulator of G-protein signaling RGS9-2: role in receptor desensitization. J Biol Chem. 2005;280:8951–60. doi: 10.1074/jbc.M407005200. [DOI] [PubMed] [Google Scholar]
- 77.Mu Y, Ren Z, Jia J, Gao B, Zheng L, Wang G, et al. Inhibition of phosphodiesterase10A attenuates morphine-induced conditioned place preference. Mol Brain. 2014;7:70. doi: 10.1186/s13041-014-0070-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Garzon J, Rodriguez-Munoz M, Sanchez-Blazquez P. Direct association of Mu-opioid and NMDA glutamate receptors supports their cross-regulation: molecular implications for opioid tolerance. Curr Drug Abuse Rev. 2012;5:199–226. doi: 10.2174/1874473711205030199. [DOI] [PubMed] [Google Scholar]
- 79.Trujillo KA, Akil H. Inhibition of morphine tolerance and dependence by the NMDA receptor antagonist MK-801. Science. 1991;251:85–87. doi: 10.1126/science.1824728. [DOI] [PubMed] [Google Scholar]
- 80.Bisaga A, Comer SD, Ward AS, Popik P, Kleber HD, Fischman MW. The NMDA antagonist memantine attenuates the expression of opioid physical dependence in humans. Psychopharmacology. 2001;157:1–10. doi: 10.1007/s002130100739. [DOI] [PubMed] [Google Scholar]
- 81.Harris AC, Rothwell PE, Gewirtz JC. Effects of the NMDA receptor antagonist memantine on the expression and development of acute opiate dependence as assessed by withdrawal-potentiated startle and hyperalgesia. Psychopharmacology. 2008;196:649–60. doi: 10.1007/s00213-007-0998-2. [DOI] [PubMed] [Google Scholar]
- 82.Kolik LG, Konstantinopolsky MA. Comparative assessment of the effectiveness of noncompetitive NMDA receptor antagonists amantadine and hemantane in morphine withdrawal syndrome model. Bull Exp Biol Med. 2019;166:739–43. doi: 10.1007/s10517-019-04430-2. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Single-nuclei RNA-sequencing data are available at the NCBI Gene Expression Omnibus (GEO) under accession number GSE171165.