

Abstract
Myotonic dystrophy type 2 (DM2) is caused by CCTG repeat expansions in the CNBP gene, comprising 75 to >11,000 units and featuring extensive mosaicism, making it challenging to sequence fully expanded alleles. To overcome these limitations, we used PCR-free Cas9-mediated nanopore sequencing to characterize CNBP repeat expansions at the single-nucleotide level in nine DM2 patients. The length of normal and expanded alleles can be assessed precisely using this strategy, agreeing with traditional methods, and revealing the degree of mosaicism. We also sequenced an entire ~50 kbp expansion, which has not been achieved previously for DM2 or any other repeat-expansion disorders. Our approach precisely counted the repeats and identified the repeat pattern for both short interrupted and uninterrupted alleles. Interestingly, in the expanded alleles, only two DM2 samples featured the expected pure CCTG repeat pattern, while the other seven presented also TCTG blocks at the 3′ end, which have not been reported before in DM2 patients, but confirmed hereby with orthogonal methods. The demonstrated approach simultaneously determines repeat length, structure/motif, and the extent of somatic mosaicism, promising to improve the molecular diagnosis of DM2 and achieve more accurate genotype–phenotype correlations for the better stratification of DM2 patients in clinical trials.
Research organism: Human
Introduction
Myotonic dystrophy type 2 (DM2; MIM#602668) is an autosomal dominant multisystem disorder characterized by progressive proximal muscle weakness, myotonia, myalgia, calf hypertrophy, and multiorgan involvement with cataract, cardiac conduction defects, and endocrine disorders (Meola and Cardani, 2015; Montagnese et al., 2017). The disease is caused by a (CCTG)n repeat expansion in intron 1 of the CNBP gene (previously ZNF9; MIM*116955) on chromosome 3q21.3 (Liquori et al., 2001). The CCTG repeat tract is part of a complex (TG)v (TCTG)w (CCTG)x motif that, in healthy-range alleles, is generally interrupted by one or more GCTG, TCTG, or ACTG (NCTG) motifs that confer repeat stability (Radvanszky et al., 2013; Mahyera et al., 2018; Guo and Lam, 2016). Nonpathogenic alleles contain up to 26 CCTG repeat units, whereas premutations are composed of <75 ‘pure’ CCTG blocks whose clinical significance remains unclear (Mahyera et al., 2018; Botta et al., 2021). In DM2 patients, the number of repeats is between ~75 and >11,000 units, among the largest reported so far in repeat-expansion disorders (Depienne and Mandel, 2021). The DM2 mutation shows marked somatic instability and tends to increase in length over time within the same individual, but it does not show a strong bias toward intergenerational expansion, and genetic anticipation is rarely seen in DM2 families (Liquori et al., 2001; Kamsteeg et al., 2012; Mahyera et al., 2018; Thornton, 2014; Udd et al., 2003).
The few genotype–phenotype studies reported so far in DM2 patients did not reveal any significant associations between the severity of the disease, including the age at onset, and the number of CCTG repeats (Day et al., 2003; Udd et al., 2003). The identification of such correlations is hindered by heterogeneity across tissues, somatic instability, and the technical challenges that must be overcome to measure repeat lengths accurately in such large microsatellite expansions. This has prevented the discovery of additional in cis genetic modifiers that may ameliorate or exacerbate DM2 disease symptoms. The genetic features of the CNBP microsatellite locus, as well as its extreme length and high CG content, have frustrated attempts to size and sequence expanded alleles in DM2 patients. Even the investigators in the original gene-discovery study were unable to sequence the entire CCTG array because of its length and the high level of somatic mosaicism (Liquori et al., 2001; Bachinski et al., 2003; Day et al., 2003).
According to international guidelines, current best practice for DM2 genetic testing relies mainly on PCR-based approaches (Botta et al., 2006; Kamsteeg et al., 2012). These include an initial short-range PCR (SR-PCR) to exclude a DM2 diagnosis when two normal-range alleles are detected. When only one allele is visible, (CCTG)n expansions can be identified by long-range PCR (LR-PCR) or quadruplet-repeat primed PCR (QP-PCR), leading to a ~99% detection rate (Kamsteeg et al., 2012). However, neither method can define the exact length of large DM2 expansions, which requires the Southern blot analysis of digested genomic DNA and has a sensitivity of ~80% (Day et al., 2003). The latter method is time consuming, requires large amounts of DNA, and is not included in the routine workflow of most diagnostic centers. Moreover, none of the methods described above can resolve the expansion to the single-nucleotide level and they have a limited ability to detect minor alleles and the degree of somatic mosaicism.
Third-generation long-read sequencing technologies provide an unprecedented opportunity to fully characterize DM2 expansions in terms of repeat size, allele configuration, and base composition. Whereas most second-generation sequencing methods produce short reads, third-generation methods such as Oxford Nanopore Technologies (ONT) and PacBio SMRT sequencing facilitate the analysis of DNA fragments multiple kilobases in length, including large repetitive elements and their flanking regions. Furthermore, third-generation methods are generally based on PCR-free workflows, thus avoiding amplification-related biases (Hommelsheim et al., 2014) and challenges caused by regions with a high CG content (such as the CNBP locus). However, these technologies are more expensive and error-prone than first- and second-generation methods. Targeted approaches that maximize data production on a selected region of interest can compensate for such errors and provide cost-effective alternatives to whole-genome sequencing.
Targeted enrichment approaches coupled to long-read sequencing have already been used for the in-depth characterization of repeat expansions in fragile X syndrome (FMR1), Huntington’s disease (HTT), and neuronal intranuclear inclusion disease (NOTCH2NLC), although these expansions are significantly shorter than the (CCTG)n repeats in DM2 patients (DeJesus-Hernandez et al., 2021; Ebbert et al., 2018; Giesselmann et al., 2019; Grosso et al., 2021; Hafford-Tear et al., 2019; Höijer et al., 2018; Mizuguchi et al., 2021; Sone et al., 2019; Tsai et al., 2017; Wallace et al., 2021; Wieben et al., 2019; Mitsuhashi and Matsumoto, 2020). The longest allele thus far characterized by third-generation sequencing is a 21-kbp allele of the C9orf72 gene that causes amyotrophic lateral sclerosis and frontotemporal dementia (DeJesus-Hernandez et al., 2021). In addition, many of the works cited above combined PacBio SMRT sequencing with LR-PCR (Mangin et al., 2021; Ciosi et al., 2021; Cumming et al., 2018), which is unsuitable for the analysis of DM2 expansions due to their extreme length and high GC content. Importantly, PacBio reads do not exceed 20 kbp in a PCR-free enrichment (DeJesus-Hernandez et al., 2021; Ebbert et al., 2018; Hafford-Tear et al., 2019; Höijer et al., 2018; Tsai et al., 2017; Wieben et al., 2019), and would not completely span DM2 pathogenic expansions (20 kbp on average, but up to 50 kbp).
To address these issues, we assessed the analysis of CNBP expansions using a combination of CRISPR/Cas9-based enrichment (Cas9-enrichment) and ONT sequencing. The latter can generate reads >100 kbp in length (Payne et al., 2019; Iyer et al., 2022) and recently demonstrated valuable for the analysis of very long repetitive elements, like telomeric and centromeric regions in the completion of human genome (Sergey et al., 2022; Consortium and The Telomere-to-telomere T T, 2022) and microsatellite expansions in the pathogenic range (Stevanovski et al., 2022). In this manner, we sequenced full-length (CCTG)n expansions in nine DM2 patients including one mutated allele 47 kbp in length. Because this approach achieves single-nucleotide resolution, we were able to detect a previously unreported (TCTG)n motif at the 3′ end of the CNBP expansion in seven of the DM2 patients. Our pilot study demonstrated that Cas9-mediated enrichment and long-read sequencing improves the DM2 diagnostic workflow, facilitating the in-depth characterization of CNBP expansions by accurately reporting the repeat length, structure/motif, and degree of somatic mosaicism in a single analysis. In the future, this approach may enable more precise genotype–phenotype correlations and thus improve patient stratifications in clinical trials for personalized therapies.
Results
Molecular characterization of DM2 patients using traditional methods
We analyzed nine DM2 patients (six males and three females, mean age = 46.4 ± 20 years) with existing molecular diagnoses based on a combination of PCR-based approaches (SR-PCR, LR-PCR, and QP-PCR) to detect the presence of (CCTG)n expansions in the CNBP gene (Table 1). Four patients were familial cases (A1–A4) (Figure 1—figure supplement 1) whereas patients B, C, D, E, and F were sporadic cases. The maximum length of the (CCTG)n expansion could not be determined using routine diagnostic methods, so we digested genomic DNA and estimated the size of each allele by Southern blot analysis (Figure 1A). This suggested that the size of the microsatellite ranged from 20 to about 40 kbp (Figure 1—figure supplement 2 and Table 1). As expected, no signal was detected for healthy control subjects (CTR) or myotonic dystrophy type 1 (DM1) patient (Figure 1—figure supplement 2). The characterization of normal CNBP alleles by SR-PCR and Sanger sequencing revealed the presence of eight short interrupted alleles with the structure (TG)17–24 (TCTG)6–9 (CCTG)5–7 GCTG CCTG TCTG (CCTG)7 and one short uninterrupted allele with the structure (TG)19 (TCTG)9 (CCTG)12, matching our previous results for an Italian population (Botta et al., 2021; Table 1).
Table 1. Demographic and molecular features of the dystrophy type 2 (DM2) patients.
For each patient, the table shows the ID, sex, age, age at onset, repeat length, and structure of the CNBP normal allele characterized by Sanger sequencing, and the estimated repeat length of the expanded allele determined by Southern blotting (–, not available).
| Normal allele | Expanded allele | |||||
|---|---|---|---|---|---|---|
| Sample ID | Sex | Age | Age at onset | Repeat length (bp) | Repeat structure | Repeat length (bp) |
| A1 | F | 75 | 70 | 136 | (TG)24 (TCTG)7 (CCTG)5 GCTG CCTG TCTG (CCTG)7 | 40,000 |
| A2 | M | 27 | 25 | 130 | (TG)17 (TCTG)9 (CCTG)5 GCTG CCTG TCTG (CCTG)7 | 20,000 |
| A3 | M | 21 | – | 132 | (TG)20 (TCTG)8 (CCTG)5 GCTG CCTG TCTG (CCTG)7 | 20,323 |
| A4 | M | 65 | 61 | 122 | (TG)19 (TCTG)9 (CCTG)12 | 32,745 |
| B | F | 49 | 44 | 134 | (TG)21 (TCTG)7 (CCTG)6 GCTG CCTG TCTG (CCTG)7 | 40,000 |
| C | M | 20 | – | 140 | (TG)24 (TCTG)6 (CCTG)7 GCTG CCTG TCTG (CCTG)7 | 29,027 |
| D | M | 44 | 39 | 134 | (TG)19 (TCTG)9 (CCTG)5 GCTG CCTG TCTG (CCTG)7 | 20,000 |
| E | F | 61 | 43 | 134 | (TG)19 (TCTG)9 (CCTG)5 GCTG CCTG TCTG (CCTG)7 | 30,000 |
| F | M | 56 | 50 | 138 | (TG)21 (TCTG)9 (CCTG)5 GCTG CCTG TCTG (CCTG)7 | 39,000 |
Figure 1. Cas9-mediated sequencing of the CNBP microsatellite.
(A) Experimental methods applied retrospectively to study the CNBP microsatellite in nine confirmed dystrophy type 2 (DM2) patients. The positions of AluI and HaeIII restriction sites (142 bp upstream and 108 bp downstream of the CCTG array, respectively) and the (CCTG)5 probe (orange rectangle) for Southern blot hybridization are shown, along with the gRNAs cleavage site for Cas9-mediated enrichment (boundaries of blue line = 4.2 kb) and the annealing position of P4TCTG primers for quadruplet-repeat primed PCR (QP-PCR). (B) Average coverage and (C) fold enrichment obtained by Cas9-mediated enrichment in DM2 patients following singleplex (n = 4) or multiplex (n = 4) runs. Numbers above bars represent the percentage of on-target reads. (D) Total and on-target number of PASS reads generated for each DM2 patient and (E) number of reads fully spanning the CNBP microsatellite and attributable to either the normal or expanded alleles. Data are plotted as means ± standard error of the mean (SEM). WG, whole genome. Sequencing statistics of singleplex and multiplex experiments are reported in detail in Supplementary file 1.

Figure 1—figure supplement 1. Pedigree of the dystrophy type 2 (DM2) family analyzed.

Figure 1—figure supplement 2. Southern blot analysis of expanded alleles in dystrophy type 2 (DM2) patients.

CNBP repeat-expansion analysis by Cas9-mediated enrichment coupled to ONT sequencing
We characterized the full-length CNBP expansions at single-nucleotide resolution by ONT sequencing following Cas9-mediated enrichment. Accordingly, we designed two gRNAs to excise a 4.2-kbp fragment spanning the CNBP repeat on chromosome 3q21.3 (Figure 1A and Supplementary file 1).
Genomic DNA from the nine DM2 patients was analyzed in four singleplex and four multiplex runs, the latter applied to clinical samples here for the first time (Supplementary file 1). Cas9-mediated sequencing achieved good target coverage in all experiments, with 346 ± 64 reads (mean ± standard error of the mean) on the CNBP locus (Figure 1B and Supplementary file 1). Singleplex runs had consistently lower background than multiplex runs (0.08× vs. 0.57×), and thus achieved a higher average fold enrichment (3521- vs. 637-fold) (Figure 1B, C and Supplementary file 1). Collectively, for each DM2 patient, we generated a mean total of 105,737 PASS reads, 308 of which were on target (Figure 1D) and 186 of which completely spanned the normal or expanded alleles (Figure 1E). Only these ‘complete sequences’ were used for subsequent analysis, representing ~78% and~22% of the normal and expanded repeat-spanning reads, respectively (Figure 1E).
The de novo assembly of reads derived from the normal CNBP alleles in DM2 patients (145 on average per sample, IQR = 67; Figure 1E) showed that the complex (TG)v (TCTG)w (CCTG)x (NCTG)y (CCTG)z repeat ranged in size from 122 to 141 bp, corresponding to 12–15 CCTG quadruplets (Table 2 and Figure 2B). The size and repeat pattern in each patient were largely consistent with the Sanger sequencing data (99.5% mean accuracy, Pearson’s r=0.971, p < 0.0001; Figure 2C), with six patients showing a perfect match, two differing at a single-nucleotide position and only one differing at two nucleotide positions (Table 2).
Table 2. CNBP repeat analysis based on Cas9-mediated sequencing of the normal alleles.
For each patient, the table shows the characteristics of normal CNBP alleles based on the analysis of ONT sequencing data, in terms of length and structure. The table reports the percentage identity of consensus sequences reconstructed from ONT or Sanger sequencing data and the incongruences in ONT-derived sequences are highlighted in bold.
| Normal allele | Sample ID | Repeat length (bp) | Repeat structure | Identity with Sanger sequence |
|---|---|---|---|---|
| A1 | 136 | (TG)24 (TCTG)7 (CCTG)5 GCTG CCTG TCTG (CCTG)7 | 100.0% | |
| A2 | 131 | (TG)17 TGCTG (TCTG)8 (CCTG)5 GCTG CCTG TCTG (CCTG)7 | 99.2% | |
| A3 | 132 | (TG)20 (TCTG)8 (CCTG)5 GCTG CCTG TCTG (CCTG)7 | 100.0% | |
| A4 | 122 | (TG)19 (TCTG)9 (CCTG)12 | 100.0% | |
| B | 134 | (TG)21 (TCTG)7 (CCTG)6 GCTG CCTG TCTG (CCTG)7 | 100.0% | |
| C | 141 | (TG)24 TGCTG (TCTG)5 (CCTG)7 GCTG CCTG TCTG (CCTG)7 | 99.3% | |
| D | 138 | (TG)21 (TCTG)9 (CCTG)5 GCTG CCTG TCTG (CCTG)7 | 97.1% | |
| E | 134 | (TG)19 (TCTG)9 (CCTG)5 GCTG CCTG TCTG (CCTG)7 | 100.0% | |
| F | 138 | (TG)21 (TCTG)9 (CCTG)5 GCTG CCTG TCTG (CCTG)7 | 100.0% |
Figure 2. Analysis of ONT sequencing data from normal and expanded CNBP alleles.

(A) Integrative Genomics Viewer (IGV) visualization of ONT sequencing data at the CNBP locus of a representative dystrophy type 2 (DM2) patient following Cas9-mediated enrichment. Reads generated from the normal allele feature clear cuts on both sides of the CNBP repeat, whereas those derived from the expanded allele are longer, soft-clipped and do not match the reference genome, as expected. Length distributions of reads derived from the normal alleles (B) and expanded alleles (D) of each patient. Boxes represent the interquartile range (IQR) of lengths, the horizontal line is the median, whiskers and outliers are plotted according to Tukey’s method. (C) Correlation between the length of ONT and Sanger consensus sequences for the normal allele (n = 9). (E) Correlations between the maximum length of ONT sequences (longest complete read) and the upper edge of the Southern blot trace for the expanded allele (n = 9). Numbers on top of panels (B) and (D) indicate the coefficient of variation of normal and expanded alleles, respectively.
Reads derived from the expanded alleles (41 on average per sample, IQR = 11; Figure 1E) ranged from 344 bp to as much as 46.6 kbp (Figure 2D), confirming the presence of extremely large expansions in these patients. To our knowledge, the latter is the longest repeat expansion analyzed thus far at single-nucleotide resolution (Mizuguchi et al., 2021; Sone et al., 2019; Giesselmann et al., 2019; Wallace et al., 2021) and is one of the longest DNA fragments captured by Cas9-mediated enrichment with no specific adjustment (Gilpatrick et al., 2020; Iyer et al., 2020). Considering average values per sample, the number of repetitive quadruplets varied from 1371 to 4421, corresponding to expansion lengths of 5485–17,685 bp. Moreover, in each patient, the longest molecule sequenced with ONT (derived from the expanded allele) largely agreed with the Southern blot estimates (Pearson’s r = 0.6840, p = 0.0422, Figure 2E), with the exception of sample (B). Even within the same individual, the mean size of reads derived from the expanded allele was consistently more variable than that from the normal allele (65% ± 0.091 vs. 4% ± 0.002; Figure 2B–D, Figure 3). Such pronounced variability within each DM2 patient indicates extensive mosaicism, in agreement with previous reports (Liquori et al., 2001; Bachinski et al., 2003; Day et al., 2003).
Figure 3. Analysis of the expanded-repeat CNBP alleles in dystrophy type 2 (DM2) patients.
Integrative Genomics Viewer (IGV) visualization (35-kbp windows) of ONT-targeted sequencing data from the expanded alleles of four representative DM2 patients. Complete reads were aligned at the 5′ end (A) and then at the 3′ end (B) in order to identify the repeat pattern that characterizes the expanded microsatellite locus. Each motif in the expanded alleles was visualized using a different color, as indicated in the key. Samples C and E contained a ‘pure’ CCTG expansion (blue) whereas samples A4 and A2 also contained the unexpected TCTG motif (red) downstream of CCTG. (C) Abundance of quadruplets identified in each patient. The y-axis shows the number of ONT reads with a certain number of repeats, whereas the x-axis shows the number of quadruplet repeats identified. ONT reads were grouped into 500 bp bins. The gray line represents the estimated kernel density of the underlying solid gray distribution of ONT reads.

Figure 3—figure supplement 1. Analysis of the CNBP repeat motif for the expanded alleles in dystrophy type 2 (DM2) patients.

Figure 3—figure supplement 2. Analysis of the CNBP 5′-end (TG)v repeat motif of the CNBP expanded alleles of A1–A4 dystrophy type 2 (DM2) family members.

To characterize the repeat pattern across the expanded microsatellite locus, we identified the quadruplet motifs in each individual read, and highlighted them with distinct colors after aligning ‘complete sequences’ at the 5′ and 3′ ends (Figure 3 and Figure 3—figure supplement 1A, B). The total number of quadruplets was highly variable within each patient, thus confirming the extensive donor-dependent mosaicism described above (Figure 3C and Figure 3—figure supplement 1). For example, reads from patient C carried on average 3000 quadruplets (Figure 3C), but the number of (CCTG) repeats in individual reads varied from 150 up to 8000 (Table 3). Since we analyzed only those reads containing at least 600 bp up-/downstream the CNBP microsatellite, and thus comprising the repeat entirely, we excluded that such large variability could be ascribed to the analysis of fragmented DNA molecules.
Table 3. CNBP repeat analysis based on Cas9-mediated sequencing of the expanded alleles.
For each patient, the table shows the characteristics of expanded CNBP alleles based on the analysis of ONT sequencing data, in terms of length and structure. The expanded (CCTG)x are colored in blue and (TCTG)y in red. Potential incongruences in ONT-derived sequences are highlighted in bold. The table indicates also the fraction of reads carrying the unexpected TCTG motif at the 3′ end of the CNBP microsatellite expansion.
| Expanded allele | Sample ID | Repeat length (bp) (min–max) | Repeat structure | Number of reads carrying the TCTG motif |
|---|---|---|---|---|
| A1 | 3241–46,685 | (TG)20 (TCTG)7 (CCTG)1000–12,000 (TCTG)0–10 | 14 (45%) | |
| A2 | 864–23,779 | (TG)18 (TCTG)7 (CCTG)1000–4500 (TCTG)0–2000 | 92 (86%) | |
| A3 | 4429–18,983 | (TG)19 (TCTG)7 (CCTG)3000–5000 (TCTG)0–25 | 8 (73%) | |
| A4 | 660–34,284 | (TG)18 (TCTG)7 (CCTG)250–8000 (TCTG)0–1500 | 9 (29%) | |
| B | 344–23,358 | (TG)18 (TCTG)7 (CCTG)300–4000 (TCTG)0–400 | 6 (23%) | |
| C | 700–31,753 | (TG)20 (TCTG)7 (CCTG)150–8000 | 0 (0%) | |
| D | 383–19,143 | (TG)18 (TCTG)7 (CCTG)100–4000 (TCTG)0–1000 | 3 (11%) | |
| E | 848–25,162 | (TG)18 (TCTG)6 (CCTG)200–6200 | 0 (0%) | |
| F | 1533–32,824 | (TG)15 (TCTG)10 (CCTG)400–6000 (TCTG)0–2000 | 15 (43%) |
A variable number of (TG)v repetitions upstream of the (CCTG)n array was also observed in the familiar cases A1–A4 (Table 3). Since this microsatellite tract is supposed to be stably transmitted within the same family, the observed discrepancy was likely an artefact due to ONT accuracy, as reported for the healthy alleles. Manual inspection of sequencing data indeed confirmed that all family members show an equivalent pattern of (TG)v repetitions (Figure 3—figure supplement 2).
The uninterrupted (TG)v (TCTG)w (CCTG)x motif that characterizes expanded CNBP alleles was found at the 5′ end of the repeat locus in all nine patients (Figure 3A, Figure 3—figure supplement 1, and Table 3). However, only two patients featured a ‘pure’ pattern of (CCTG)n repeats. In the remaining seven, we observed additional (TCTG)n arrays (colored in red) at the 3′ end of the CCTG expansion, which has been never reported in DM2 patients before (Figure 3B, Figure 3—figure supplement 1, and Table 3). When present, the TCTG motif was detected in a highly variable fraction of sequences (11%–86% of the expanded allele reads, Table 3), and differed widely in length (40–8000 bp) between donors and within the same individual (Figure 3, Figure 3—figure supplement 1, and Table 3).
Analysis of the TCTG repeat using orthogonal methods
To confirm the presence of the (TCTG)n motifs in the CNBP expanded alleles, we used a traditional QP-PCR method for the selective amplification of TCTG blocks at the 3′ end of the (CCTG)n array (Figure 1A). In agreement with the ONT data, QP-PCR analysis using primer P4TCTG revealed an electrophoretic profile compatible with the presence of (TCTG)n downstream of the (CCTG)n expansion in seven DM2 samples (Figure 4A). The intensity and pattern of fluorescent peaks obtained using primer P4TCTG were more variable across samples compared to the routine protocol using the standard P4CCTG primer, possibly due to different levels of somatic mosaicism in the TCTG and CCTG expansions. A 260 bp signal was visible in all samples, including the DM2 negative controls (one DM1 positive patient and one healthy subject CTR), suggesting it was a PCR artifact (Figure 4). Interestingly, the two patients with ‘pure’ (CCTG)n expansions based on ONT data (patients C and E) did not yield amplification peaks within the expansion range with the P4TCTG primer (Figure 4B). Fluorescent signals <140 bp were visible in these samples because normal CNBP alleles also contain (TCTG)n repeats at their 5′ end (Table 1). The direct sequencing of QP-PCR products generated using primer P4TCTG confirmed the presence of (TCTG)n motif in DM2 patients A1, A2, A3, A4, B, D, and F, thus supporting the ONT data (Figure 4 and Figure 4—figure supplement 1).
Figure 4. Analysis of the TCTG motif by quadruplet-repeat primed PCR (QP-PCR) and Sanger sequencing.
(A) Representative QP-PCR profiles of genomic DNA samples from patients A2 and A4 and showing the presence of the TCTG block. Upper panels show QP-PCR results using the conventional P4CCTG primer. Lower panels show QP-PCR results using primer P4TCTG. (B) Sanger sequencing of the QP-PCR products from P4TCTG reaction confirming the presence of the TCTG sequence. (C) QP-PCR profiles of genomic DNA samples from patients C and E showing only the traditional dystrophy type 2 (DM2) motif CCTG. For each patient, the composition of CNBP expanded alleles above the QP-PCR tracks reflects the ONT sequencing data. Asterisks (*) indicate nonspecific signals, also visible in the QP-PCR profiles of DM1 and CTR samples (D).
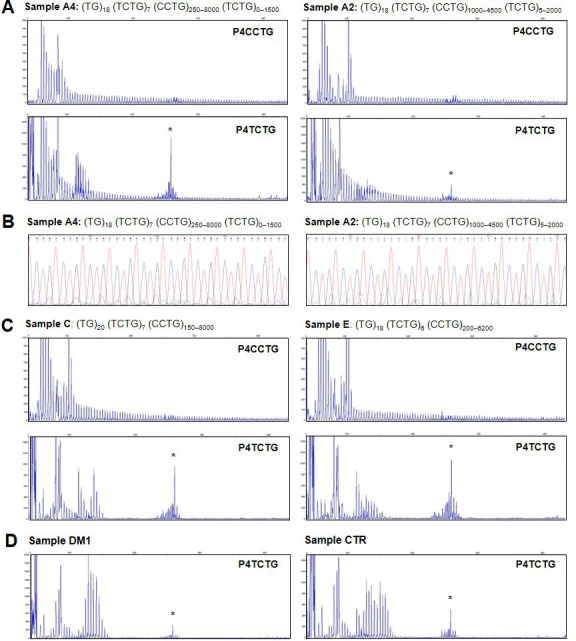
Figure 4—figure supplement 1. Sanger sequencing of quadruplet-repeat primed PCR (QP-PCR) products showing the presence of the (TCTG)n motif.

Discussion
The analysis of extremely long microsatellite expansions is challenging, preventing the in-depth characterization of CNBP mutations underlying DM2 and its relationship with the clinical phenotype. To date, the genotype–phenotype correlation issue in DM2 is still unsolved and relies on a single study from Day et al., 2003 in which Southern blot analysis was used to determine the length of DM2 mutation. Because of the extremely large size of the CCTG expansions and somatic instability of the repeat, Southern blot fails to detect the DM2 mutation in about 20% of known carriers, whose expansion length remains undeterminable. Moreover, currently no diagnostic method allows to sequence through fully expanded CNBP microsatellites. Here, we have demonstrated for the first time the use of Cas9-mediated enrichment and ONT long-read sequencing to analyze the full (CCTG)n expansion in DM2 patients at single-nucleotide resolution. We were able to characterize normal and expanded CNBP alleles, simultaneously revealing the repeat length, structure/motif, and extent of somatic mosaicism, which is not possible with traditional methods, even if several are used in combination.
The paucity of genotype–phenotype data for DM2 reflects the historical inability to determine the size of CCTG repeats, especially in the largest expansions, which traditionally was based on the Southern blot analysis of genomic DNA. This labor-intensive and time-consuming technique is becoming obsolete because it requires large amounts of high-molecular-weight DNA. LR-PCR can be used instead, but the performance of this technique is poor in regions with a high CG content, and it cannot accommodate the very large expanded alleles (>15 kbp) often found in DM2. Therefore, although LR-PCR achieves the sensitive detection of DM2 mutations, the full length of the (CCTG)n expansion cannot be determined in all patients. Our Cas9-targeted sequencing protocol overcomes these limits by focusing long-read sequencing data on the CNBP microsatellite, with reads spanning the entire expansion. The length of normal and expanded alleles in DM2 patients determined using this new approach closely matched the values obtained with the traditional reference methods. The mean size of normal CNBP alleles was 134 bp whereas expanded alleles ranged from 344 to 46,685 bp, in agreement with previous reports (Meola and Cardani, 2015; Montagnese et al., 2017; Botta et al., 2021). A single incongruence was observed for one expanded allele in patient B, where ONT sequencing underestimated the size determined by Southern blotting (~20 vs. ~40 kbp). A possible explanation is the presence of damaged DNA in the sample, reflecting its long-term storage in a biobank. Southern blotting involves the fractionation of double-stranded DNA, which would be unaffected by the presence of nicked strands, whereas ONT sequencing involves the analysis of single DNA strands, so the presence of nicks would have a profound effect (Oxford Nanopore Community, 2021). Even so, Cas9-mediated enrichment allowed us to sequence DM2 alleles up to ~50 kbp in length at single-nucleotide resolution, which has not been reported previously for DM2 or any other repeat-expansion disorder using this approach (Giesselmann et al., 2019; Mizuguchi et al., 2021; Sone et al., 2019; Wallace et al., 2021; Gilpatrick et al., 2020; Iyer et al., 2020). The analysis of such long and repetitive alleles required the coupling of a PCR-free enrichment protocol to ONT sequencing because even other long-read sequencing technologies cannot accommodate this read length in targeted sequencing experiments. For example, PacBio long-read sequencing was previously used to sequence repeat expansions in DM1, which is also characterized by long alleles of 4–6 kbp, but the microsatellites were first amplified by PCR (Mangin et al., 2021; Cumming et al., 2018). Even when coupled to PCR-free enrichment approach based on Cas9, the length of PacBio sequencing reads could not exceed 20 kbp (DeJesus-Hernandez et al., 2021; Ebbert et al., 2018; Hafford-Tear et al., 2019; Höijer et al., 2018; Tsai et al., 2017; Wieben et al., 2019).
Although ONT sequencing had been already utilized for the analysis of the microsatellite within the CNBP gene, this was confined to CNBP alleles in the normal range only (Stevanovski et al., 2022; Mohammad et al., 2022). Moreover, the work of Mitsuhashi et al. exploited ONT whole-genome sequencing, that is not applicable in the routine due to the very high costs (Mohammad et al., 2022). The group of Stevanovski utilized the recently introduced ‘Read Until’ feature of ONT sequencing for the analysis of microsatellites in 37 disease-associated loci. This allows the selective sequencing of predefined genomic regions, thus enabling a targeted sequencing with similar advantages of the Cas9-mediated sequencing presented hereby. However, enrichment levels achieved by ‘Read Until’ (5×) are consistently lower than those obtained with the Cas9 approach (500×), due to higher background (Stevanovski et al., 2022). This may constitute an important issue when dealing with extremely long CNBP alleles that can be disadvantaged in sequencing as compared to shorter contaminating fragments (Holgersen et al., 2021).
From a technical perspective, we achieved >500× enrichment on CNBP using the Cas9 protocol, which is robust and comparable to similar assays for the assessment of microsatellite length (Giesselmann et al., 2019; Mizuguchi et al., 2021; Sone et al., 2019; Wallace et al., 2021). We also compared Cas9-mediated singleplex versus multiplex enrichment protocols for the first time on clinical samples and observed a consistently lower performance (10-fold lower enrichment, with 70% unclassified reads) in the multiplex environment, as reported by other ONT users for other type of samples (Oxford Nanopore Community, 2022). Further improvements are therefore required before the multiplexing protocol is suitable, for example by combining the Cas9 protocol with a second enrichment step using the ‘Read Until’ feature of ONT sequencing to exclude the background noise. Alternatively, costs could be optimized by analyzing single samples using Flongles, which produce less sequencing data than regular ONT flow cells but reduce costs by 90%.
The bioinformatic pipeline we used to analyze the CNBP microsatellite sequences also allowed us to recognize the repeat pattern and precisely count the repeats, including the short interrupted and uninterrupted alleles typifying the Italian population (Botta et al., 2021). Consensus sequences generated from the normal allele shared a high degree of identity (99.5%) and significant correlation with state-of-the-art Sanger sequences, aside from a few nucleotide positions. Similar small discrepancies have also been reported in characterization of the (TG)v motif upstream of the (CCTG)n repeated array in familial cases A1–A4. These inconsistences probably reflect ONT sequencing errors and could be addressed by using the most recent base-calling algorithm and eventually the more accurate Q20+ chemistry.
In the expanded alleles of seven DM2 patients, the anticipated (CCTG)n repeat was accompanied by a previously unreported (TCTG)n repeat located at the 3′ end of the (CCTG)n array. When present, the atypical motif varied in length between donors and within each sample (40–8000 bp). Repeat interruptions within the expanded array have been reported in 3%–8% of DM1 patients (Tomé et al., 2018; Braida et al., 2010; Santoro et al., 2017; Santoro et al., 2013; Ballester-Lopez et al., 2020; Pešović et al., 2018; Miller et al., 2020; Radvansky et al., 2011; Siena et al., 2018; Botta et al., 2017; Santoro et al., 2015; Addis et al., 2012; Fontana et al., 2020; Lian et al., 2016; Leeflang and Arnheim, 1995; Musova et al., 2009; Cumming et al., 2018), but have not been described in DM2 before. This may reflect the challenge of sequencing complete CNBP expanded alleles and/or the use of a primer containing ‘pure’ (CCTG)n repeats for diagnostic QP-PCR. Given that (TCTG)n repeats were present in a highly variable proportion of expanded alleles (11%–86%), always in the presence of the typical (CCTG)n repeats, technical bias may have revealed only the ‘pure’ (CCTG)n repeats. Indeed, a modified QP-PCR protocol using a primer containing five TCTG units – (TCTG)5T – was able to confirm the ONT sequencing data. From a biological perspective, the (TCTG)n motif may have arisen through DNA duplication/repair errors or spontaneous DNA damage in the somatic cells of DM2 patients. Although the presence of this motif may be biologically relevant in the context of DM2, our data must be interpreted with caution. First, the motif was discovered in a small set of patients, most belonging to the same family, so confirmation requires a larger prospective DM2 cohort enrolled in multicenter studies, in which DNA samples are collected in order to ensure the optimal quality for ONT sequencing. Second, considering the known limitations of ONT sequencing when presented with low-complexity regions such as homopolymers, the length and recurrence of such motifs should be investigated using other long-read methods when they are sufficiently advanced to sequence the expanded alleles completely.
The Cas9-targeted sequencing approach also allowed us to estimate the degree of somatic mosaicism for the mutated alleles, either ‘pure’ or ‘interrupted’. On average, the allele length within each patient varied by 65%. Mosaicism plays an important role in the development of disease symptoms, so establishing the relative proportion of expanded alleles in the lower and upper mutation range could add prognostic value, significantly improving genetic counseling for DM2. Extreme mosaicism (more than expected based on previous studies) has also been detected when long-read sequencing is applied to other repeat-expansion disorders, suggesting that such techniques achieve higher resolution (Loomis et al., 2013; Mizuguchi et al., 2021; Mangin et al., 2021). As already demonstrated for DM1 (Cumming et al., 2019; Monckton et al., 1995), the progenitor allele length (i.e., the length of the CCTG repeat transmitted by the affected parent) is one genetic determinant that influences the age at onset of DM2 symptoms, and that age is further modified by individual-specific differences in the level of somatic instability. Notably, our method accurately distinguished between the shortest expanded allele and the normal allele.
Another advantage of the approach demonstrated hereby is that PCR-free analysis potentially allows the direct assessment of DNA methylation, as already reported for other repeat-linked diseases (Fukuda et al., 2021; Giesselmann et al., 2019). This can provide additional information concerning the impact of expansions on the functionality of the CNBP gene. The methylation of the CNBP gene has been analyzed using a pyrosequencing method, revealing hypomethylation of CpG sites upstream and hypermethylation of CpG sites downstream of the (CCTG)n expansion in DM2 patients and healthy individuals, with no significant differences between these groups (Santoro et al., 2018). However, it remains possible that the DM2 mutation could have epigenetic effects in other regulatory regions of the CNBP gene and/or in different tissues.
Given the ability of our method to simultaneously determine the size, single-nucleotide composition and degree of somatic mosaicism of DM2 repeat expansions, ONT sequencing could be included in the DM2 diagnostic workflow to improve the information content available for genetic counseling. To date, the cost for Cas9-mediated sequencing of a single patient is relatively high and not comparable with the PCR-based approaches used in the routine of the DM2 molecular diagnostics. Nevertheless, targeted long-read sequencing might help to solve unusual large and complex (CCTG)n expansions not detectable with conventional methods and identifies noncanonical repetitive motif conformations and sequence interruptions. Taken together, this information will allow more precise correlation between the length and composition of DM2 expansion and the clinical phenotype. In a next future, further evolution of ONT chemistry and the optimization of multiplexing strategies are expected to drastically decrease the costs of the analysis, making the Cas9-mediated sequencing more easily accessible in the clinical practice.
Taken together, our pilot study has demonstrated the potential of PCR-free long-read sequencing for the genetic assessment of DM2, allowing us to investigate both the length and genetic features of normal and expanded alleles in a single round of analysis. The use of such an approach in larger cohorts will increase the accuracy of genotype–phenotype correlations and enhance the information content available for DM2 genetic counseling.
Materials and methods
Key resources table.
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Gene (Homo sapiens) | CNBP | Ensembl | HGNC:13164 | Hg38 |
| Biological sample (Homo sapiens) | Anti-coagulated peripheral blood | Policlinico Tor Vergata, Rome, Italy | Patient A1, A2, A3, A4, B, C, D, E, F | |
| Sequence-based reagent | Digoxigenin (DIG)-labeled locked nucleic acid (LNA) probe | Nakamori et al., 2009 | DIG-LNA probe | (CCTG)5 |
| Sequence-based reagent | P4TCTG | This paper | PCR primers | agc gga taa caa ttt cac aca gga TCT GTC TGT CTG TCT GTC TGT |
| Sequence-based reagent | CL3N58_DR-[FAM] | This paper | PCR primers | GCC TAG GGG ACA AAG TGA GA |
| Sequence-based reagent | P3 | This paper | PCR primers | AGC GGA TAA CAA TTT CAC ACA GGA |
| Sequence-based reagent | crRNA1_CNBP | This paper | CRISPR RNA | CCA CCT GAT TCA CTG CGA TA |
| Sequence-based reagent | crRNA2_CNBP | This paper | CRISPR RNA | GGC TTC TCA TTC CAC GAC CA |
| Sequence-based reagent | Native barcodes | Oxford Nanopore Technologies (ONT) | EXP-NBD104 | |
| Commercial assay or kit | DIG-High Prime DNA Labeling and Detection Starter Kit II | Roche | Cat. No. 11585614910 | |
| Commercial assay or kit | Flexigene DNA Kit | Qiagen | Cat. No. 51206 | |
| Commercial assay or kit | BigDye Terminator v3.1 Cycle Sequencing Kit | Thermo Fisher | Cat. No. 4337458 | |
| Commercial assay or kit | Nanobind CBB Big DNA HMW Kit | Circulomics | SKU 102-301-900 | |
| Commercial assay or kit | NucleoSpin Blood L Kit | Macherey-Nagel | Item number: 740954.20 | |
| Commercial assay or kit | Qubit dsDNA BR Assay Kit | Thermo Fisher Scientific | Cat. No. Q32853 | |
| Commercial assay or kit | TapeStation DNA ScreenTape & Reagents | Agilent Technologies | Cat. No. 5067–5365 5067–5366 |
|
| Software, algorithm | GeneMapper Software 6 | Applied Biosystems | Cat. No. 4475074 | |
| Software, algorithm | CHOPCHOP | Labun et al., 2019 | chopchop.cbu.uib.no/ | |
| Software, algorithm | Guppy v3.4.5 | Computational Biology Research Center – AIST | ||
| Software, algorithm | NanoFilt v2.7.1 | De Coster et al., 2018 | ||
| Software, algorithm | BBMap suite v38.87 | https://sourceforge.net/projects/bbmap/ | ||
| Software, algorithm | Tandem Repeat Finder v4.09 | Benson, 1999 | ||
| Software, algorithm | Minimap2 v2.17-r941 | Li, 2018 | ||
| Software, algorithm | Integrative Genomics Viewer (IGV) v2.8.3 | Robinson, 2011 | ||
| Software, algorithm | Scripts for the generation of consensus sequences and repeat annotations for the normal allele | https://github.com/MaestSi/CharONT2 | Script Name: CharONT2 | |
| Software, algorithm | Scripts for the annotation of repeats and the generation of simplified reads for the expanded allele | https://github.com/MaestSi/MosaicViewer_CNBP | Script Name: MosaicViewer_CNBP v1.0.0 | |
| Software, algorithm | MinKNOW V20.06.5 | Oxford Nanopore Technologies | ||
| Commercial assay or kit | Alt-R S.p. HiFi Cas9 Nuclease v3 | IDT | Cat. No. 1081060 | Recombinant Cas9 nuclease for target excision (see M&M) |
| Commercial assay or kit | Alt-R CRISPR-Cas9 tracrRNA | IDT | Cat. No. 1072532 | Structural RNA for gRNA formation (see M&M) |
| Commercial assay or kit | AMPure XP Beads | Beckman-Coulter | Product No. A63881 | Magnetic beads for nucleic acid purification (see M&M) |
| Commercial assay or kit | CutSmart buffer 10× | New England BioLabs | Cat. No. B7204 | Buffer for gDNA dephosphorylation, RNP formation, and target excision (see M&M) |
| Commercial assay or kit | Blunt/TA Ligase Master Mix | New England BioLabs | Cat. No.: M0367S | T4 DNA ligase for native barcode ligation to dA-tailed ends (see M&M) |
| Commercial assay or kit | FLO-MIN106D (R9.4.1) flow cell | Oxford Nanopore Technologies | FLO-MIN106D | Flowcell for ONT sequencing (see M&M) |
DM2 patients
We retrospectively analyzed nine genetically confirmed DM2 Italian patients, whose enrollment in the study was approved by the institutional review board of Policlinico Tor Vergata (document no. 232/19). All experimental procedures were carried out according to The Code of Ethics of the World Medical Association (Declaration of Helsinki). Informed consent was obtained from all nine participants and all samples and clinical information were anonymized immediately after collection using a unique alphanumeric identification code. Sociodemographic data for the DM2 patients are summarized in Table 1.
Southern blotting
Genomic DNA was extracted from 500 µl of anticoagulated peripheral blood using a Flexigene DNA Kit (Qiagen, Hilden, Germany) and diluted to a final volume of 25 μl with double-distilled water. The quality and quantity of DNA were assessed using a Denovix spectrophotometer and by 1% agarose gel electrophoresis. CNBP expanded alleles were detected as previously described (Nakamori et al., 2009), with modifications. Briefly, 2 μg of genomic DNA was digested with AluI and HaeIII and the fragments were resolved by 0.4% agarose gel electrophoresis at 40 V for 40 hr. After denaturation and neutralization, the DNA was transferred to a nylon membrane (MilliporeSigma, Burlington, MA) and fixed by UV cross-linking using a Stratalinker 2400 (Stratagene, San Diego, CA). The membrane was hybridized for 16 hr at 65°C with a digoxigenin (DIG)-labeled locked nucleic acid (LNA) probe (CCTG)5 at a concentration of 10 pmol/ml. After washing at high stringency, the signal was revealed using the DIG High Prime DNA Labeling and Detection Starter Kit II (Roche, Basel, Switzerland) and visualized using an ImageQuant LAS 4000 device (GE Healthcare, Chicago, IL). Bands were sized by running two sets of molecular weight markers alongside the samples: DNA Molecular Weight Marker XV (Expand DNA Molecular Weight Marker, Roche) and λ DNA-HindIII Digest (New England Biolabs, Ipswich, MA).
SR-PCR, QP-PCR, and Sanger sequencing
SR-PCR products were generated as reported earlier (Kamsteeg et al., 2012; Botta et al., 2006). QP-PCR targeting the 3′ end of the (CCTG)n repeat array was carried out as previously described (Catalli et al., 2010; Musova et al., 2009), with modifications. Specifically, the repeat primer P4TCTG-agc gga taa caa ttt cac aca gga TCT GTC TGT CTG TCT GTC TGT (lower case letters indicate the primer tail that does not complement the repeat) was combined with primers CL3N58_DR-[FAM]-GCC TAG GGG ACA AAG TGA GA and P3-AGC GGA TAA CAA TTT CAC ACA GGA to target the most 3′ (TCTG)n interruptions. The length of the CNBP unexpanded alleles and QP-PCR products were determined by capillary electrophoresis on the 3500 Genetic Analyzer followed by analysis using GeneMapper 6 (Applied Biosystems, Waltham, MA). The SR-PCR and QP-PCR products were purified using the ExoSAP protocol, directly sequenced using the Big Dye Terminator Cycle Sequencing Kit v3.1 (Thermo Fisher Scientific, Waltham, MA) and visualized by capillary electrophoresis on the 3500 Genetic Analyzer as above.
Cas9-mediated enrichment coupled to ONT sequencing
For DM2 patients A1–A4, B, and C, genomic DNA was extracted from 0.2 to 0.5 ml of peripheral blood using the Nanobind CBB Big DNA HMW Kit (Circulomics, Baltimore, MD), designed for HMW DNA extraction. For DM2 patients D, E, and F, the Nanobind CBB Big DNA HMW Kit failed, likely due to the presence of partially degraded DNA consequent to long-term blood storage. For DM2 patients D, E, and F genomic DNA was thus extracted from 1 to 2 ml whole blood using the NucleoSpin Blood L Kit (Macherey-Nagel, Düren, Germany), an extraction kit providing higher yield thanks to the capability of retaining both long and short DNA molecules. Regardless of the extraction method, the DNA was resuspended in Tris–EDTA buffer (pH 8.0) and the quantity was determined using a Qubit fluorometer (Thermo Fisher Scientific) and Qubit dsDNA BR Assay Kit (Thermo Fisher Scientific). DNA integrity was assessed using a TapeStation 4150 device, Genomic DNA ScreenTape and Genomic DNA Reagents (ladder and sample buffer) all from Agilent Technologies (Santa Clara, CA).
We designed crRNAs using the online tool CHOPCHOP (https://chopchop.cbu.uib.no/) following ONT’s recommendations (https://community.nanoporetech.com/info_sheets/targeted-amplification-free-dna-sequencing-using-crispr-cas/v/eci_s1014_v1_reve_11dec2018), and making sure that the excised fragment was at least 3 kbp in length. Candidate crRNAs were manually checked for unique mapping by aligning them to the human genome (Hg38) using BLAST and excluding regions overlapping common single-nucleotide polymorphisms (MAF >0.01, dbSNP database). The final crRNAs were prepared by Integrated DNA Technologies (Coralville, IA): 5′-CCA CCT GAT TCA CTG CGA TA-3′ with genomic coordinates Chr3:129,175,929–129,175,948, and 5′-GGC TTC TCA TTC CAC GAC CA-3′ with genomic coordinates Chr3:129,171,664–129,171,683. The reaction mixture comprised 10 µM of each crRNA, 10 µM of transactivation crRNA (tracrRNA) and 62 µM Alt-R S.p. HiFi Cas9 Nuclease v3 in 1× CutSmart Buffer for the generation of ribonucleoprotein (RNP) complexes (all components from Integrated DNA Technologies) according to the ONT protocol (version ENR_9084_v109_revD_04Dec2018).
The dephosphorylation, Cas9-mediated digestion, and dA-tailing of 1–10 µg input genomic DNA were carried out according to the ONT protocol. The genomic DNA was incubated with the RNPs for 20 min at 37°C and then 2 min at 80°C for enzyme inactivation. ONT sequencing adapters (AMX) were ligated to the cleaved and dA-tailed target ends for 10 min at room temperature before stopping the reaction by adding one volume of 10 mM Tris–EDTA (pH 8.0). Short fragments (<3 kbp) and residual enzymes were removed by adding 0.3× AMPure XP beads (Beckman-Coulter, Brea, CA) and washing twice in long fragment buffer (ONT). The DNA was eluted by incubating for 10 min at room temperature in elution buffer (ONT). Cas9-multiplexing experiments were carried out as described above but EXP-NBD104 native barcodes (ONT) were ligated to the cleaved and dA-tailed target ends using Blunt/TA Ligase Master Mix (New England Biolabs). Samples were quantified and all available nanograms were pooled in a final volume of 65 µl nuclease-free water. ONT sequencing adapters (AMXII from EXP-NBD104) were ligated, followed by purification, washing, and elution as described above for the singleplex experiments. The purified library was mixed with 37.5 µl sequencing buffer (ONT) and 25.5 µl of library loading beads (ONT). The library was loaded onto a FLO-MIN106D (R9.4.1) flow cell and sequenced using MinKNOW v20.06.5 (ONT) until a plateau was reached.
ONT sequence data analysis
Raw fast5 files were base called using Guppy v3.4.5 in high-accuracy mode, with parameters ‘-r -i $FAST5_DIR -s $BASECALLING_DIR --flowcell FLO-MIN106 --kit SQK-LSK109 --pt_scaling TRUE’ (the last parameter was recommended by ONT technical support to achieve the most appropriate scaling of reads with biased sequence composition, as expected for repeat motifs with low complexity). Reads from multiplexed runs were demultiplexed using Guppy v3.4.5 with parameters ‘-i $BASECALLING_DIR -s $DEMulTIPLEXING_DIR --trim_barcodes --barcode_kits EXP-NBD104’. Quality filtering was carried out using NanoFilt v2.7.1 (De Coster et al., 2018), requiring a minimum quality score of 7. Reads spanning the full repeat were identified using a combination of the scripts msa.sh and cutprimers.sh from BBMap suite v38.87 (https://sourceforge.net/projects/bbmap/). In particular, we used in silico PCR with 100 bp primers annealing to the microsatellite flanking regions (minimum alignment identity = 80%), at least 600 bp up-/downstream, to extract only those reads containing the repeat entirely (https://github.com/MaestSi/MosaicViewer_CNBP/blob/main/Figures/CNBP_left_right_alignment.png; Maestri, 2020). These ‘complete sequences’ were extracted, aligned to either the normal or expanded allele based on length, and subsequent analysis was carried out using a k-means clustering method (k = 2). Reads from each allele and each sample were then processed separately. Length variability of normal and expanded alleles was determined using the coefficient of variation, measured by the ratio of the standard deviations to the average of the normal/expanded allele lengths.
An accurate consensus sequence was generated by collapsing reads from the normal allele as previously described, using the medaka ‘r941_min_high_g344’ model (Grosso et al., 2021). Finally, the polished consensus sequence was screened for repeats using Tandem Repeat Finder v4.09 (Benson, 1999). Scripts for the generation of consensus sequences and repeat annotations for the normal allele have been deposited online (https://github.com/MaestSi/CharONT2; Maestri, 2022a).
Reads from the expanded allele were aligned to sequences flanking the repeat, screened for repeats containing the motifs ‘TG’, ‘CCTG’, and/or ‘TCTG’, and visualized in a genome browser (Grosso et al., 2021). The extracted sequences and repeat summary file were then imported into R and used to generate a simplified version of the reads with a custom script. In particular, read coordinates corresponding to annotated repeats were replaced with a single-nucleotide stretch equal in length to the annotated repeat, with each motif corresponding to a different nucleotide stretch. In more detail, the flanking region was reported as unchanged to simplify the alignment of reads, ‘TG’ was replaced with ‘GG’, ‘CCTG’ with ‘CCCC’, ‘TCTG’ with ‘TTTT’, and remaining nonannotated portions were replaced with stretches of ‘N’ of the same length. Reverse complements were generated for simplified reads originally in the reverse orientation, and all simplified reads were aligned to the flanking sequence using Minimap2 v2.17-r941 and visualized in Integrative Genomics Viewer (IGV) v2.8.3. Scripts for the annotation of repeats and the generation of simplified reads for the expanded allele have been deposited online (https://github.com/MaestSi/MosaicViewer_CNBP; Maestri, 2022b).
Acknowledgements
This research was funded by the Muscular Dystrophy Association (https://www.mda.org/). We also thank the Italian DiMio onlus association of DM patients (https://www.dimio.it/) for funding to support research projects.
Funding Statement
The funders had no role in study design, data collection, and interpretation, or the decision to submit the work for publication.
Contributor Information
Annalisa Botta, Email: botta@med.uniroma2.it.
Marzia Rossato, Email: marzia.rossato@univr.it.
Murim Choi, Seoul National University, Republic of Korea.
Martin R Pollak, Harvard Medical School, United States.
Funding Information
This paper was supported by the following grants:
Muscular Dystrophy Association MDA 876149 to Massimo Delledonne.
Italian DiMio onlus association of DM patients (www.dimio.it) volunteer donation to Annalisa Botta.
Additional information
Competing interests
No competing interests declared.
is a partner of Genartis srl.
is a partner of Genartis srl, Verona.
Author contributions
Investigation, Methodology, Writing - original draft, Writing - review and editing.
Investigation, Writing - original draft.
Validation.
Validation.
Software, Methodology, Writing - original draft.
Formal analysis.
Resources.
Formal analysis.
Supervision.
Conceptualization, Supervision, Funding acquisition, Methodology.
Conceptualization, Supervision, Funding acquisition, Writing - review and editing.
Conceptualization, Supervision, Funding acquisition, Methodology, Writing - original draft, Writing - review and editing.
Ethics
The study was approved by the institutional review board of Policlinico Tor Vergata (document no. 232/19). All experimental procedures were carried out according to The Code of Ethics of the World Medical Association (Declaration of Helsinki). Informed consent was obtained from all nine participants and all samples and clinical information were anonymized immediately after collection using a unique alphanumeric identification code.
Additional files
The table reports the feature of each Cas9-mediated sequencing experiment performed and the sequencing statistics of each ONT run. Average values are also provided for singleplex and multiplex runs separately.
Data availability
The sequencing data generated in this study have been submitted to the NCBI BioProject database under accession number PRJNA818354 (https://www.ncbi.nlm.nih.gov/bioproject/PRJNA818354).
The following dataset was generated:
Alfano M. 2022. Characterization of full-length CNBP expanded alleles in myotonic dystrophy type 2 patients by Cas9-mediated enrichment and nanopore sequencing. NCBI BioProject. PRJNA818354
References
- Addis M, Serrenti M, Meloni C, Cau M, Melis MA. Triplet-primed PCR is more sensitive than southern blotting-long PCR for the diagnosis of myotonic dystrophy type1. Genetic Testing and Molecular Biomarkers. 2012;16:1428–1431. doi: 10.1089/gtmb.2012.0218. [DOI] [PubMed] [Google Scholar]
- Bachinski LL, Udd B, Meola G, Sansone V, Bassez G, Eymard B, Thornton CA, Moxley RT, Harper PS, Rogers MT, Jurkat-Rott K, Lehmann-Horn F, Wieser T, Gamez J, Navarro C, Bottani A, Kohler A, Shriver MD, Sallinen R, Wessman M, Zhang S, Wright FA, Krahe R. Confirmation of the type 2 myotonic dystrophy (CCTG)n expansion mutation in patients with proximal myotonic myopathy/proximal myotonic dystrophy of different european origins: A single shared haplotype indicates an ancestral founder effect. American Journal of Human Genetics. 2003;73:835–848. doi: 10.1086/378566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ballester-Lopez A, Koehorst E, Almendrote M, Martínez-Piñeiro A, Lucente G, Linares-Pardo I, Núñez-Manchón J, Guanyabens N, Cano A, Lucia A, Overend G, Cumming SA, Monckton DG, Casadevall T, Isern I, Sánchez-Ojanguren J, Planas A, Rodríguez-Palmero A, Monlleó-Neila L, Pintos-Morell G, Ramos-Fransi A, Coll-Cantí J, Nogales-Gadea G. A DM1 family with interruptions associated with atypical symptoms and late onset but not with A milder phenotype. Human Mutation. 2020;41:420–431. doi: 10.1002/humu.23932. [DOI] [PubMed] [Google Scholar]
- Benson G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Research. 1999;27:573–580. doi: 10.1093/nar/27.2.573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Botta A, Bonifazi E, Vallo L, Gennarelli M, Garrè C, Salehi L, Iraci R, Sansone V, Meola G, Novelli G. Italian guidelines for molecular analysis in myotonic dystrophies. Acta Myologica. 2006;25:23–33. [PubMed] [Google Scholar]
- Botta A, Rossi G, Marcaurelio M, Fontana L, D’Apice MR, Brancati F, Massa R, G Monckton D, Sangiuolo F, Novelli G. Identification and characterization of 5’ CCG interruptions in complex DMPK expanded alleles. European Journal of Human Genetics. 2017;25:257–261. doi: 10.1038/ejhg.2016.148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Botta A, Visconti VV, Fontana L, Bisceglia P, Bengala M, Massa R, Bagni I, Cardani R, Sangiuolo F, Meola G, Antonini G, Petrucci A, Pegoraro E, D’Apice MR, Novelli G. A 14-year italian experience in DM2 genetic testing: frequency and distribution of normal and premutated CNBP alleles. Frontiers in Genetics. 2021;12:668094. doi: 10.3389/fgene.2021.668094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Braida C, Stefanatos RKA, Adam B, Mahajan N, Smeets HJM, Niel F, Goizet C, Arveiler B, Koenig M, Lagier-Tourenne C, Mandel JL, Faber CG, de Die-Smulders CEM, Spaans F, Monckton DG. Variant CCG and GGC repeats within the CTG expansion dramatically modify mutational dynamics and likely contribute toward unusual symptoms in some myotonic dystrophy type 1 patients. Human Molecular Genetics. 2010;19:1399–1412. doi: 10.1093/hmg/ddq015. [DOI] [PubMed] [Google Scholar]
- Catalli C, Morgante A, Iraci R, Rinaldi F, Botta A, Novelli G. Validation of sensitivity and specificity of tetraplet-primed PCR (TP-PCR) in the molecular diagnosis of myotonic dystrophy type 2 (DM2) The Journal of Molecular Diagnostics. 2010;12:601–606. doi: 10.2353/jmoldx.2010.090239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ciosi M, Cumming SA, Chatzi A, Larson E, Tottey W, Lomeikaite V, Hamilton G, Wheeler VC, Pinto RM, Kwak S, Morton AJ, Monckton DG. Approaches to sequence the HTT CAG repeat expansion and quantify repeat length variation. Journal of Huntington’s Disease. 2021;10:53–74. doi: 10.3233/JHD-200433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Consortium. The Telomere-to-telomere T T Implications of the first complete human genome assembly. Genome Research. 2022;32:595–598. doi: 10.1101/gr.276723.122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cumming SA, Hamilton MJ, Robb Y, Gregory H, McWilliam C, Cooper A, Adam B, McGhie J, Hamilton G, Herzyk P, Tschannen MR, Worthey E, Petty R, Ballantyne B, Warner J, Farrugia ME, Longman C, Monckton DG, Scottish Myotonic Dystrophy Consortium De novo repeat interruptions are associated with reduced somatic instability and mild or absent clinical features in myotonic dystrophy type 1. European Journal of Human Genetics. 2018;26:1635–1647. doi: 10.1038/s41431-018-0156-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cumming SA, Jimenez-Moreno C, Okkersen K, Wenninger S, Daidj F, Hogarth F, Littleford R, Gorman G, Bassez G, Schoser B, Lochmüller H, van Engelen BGM, Monckton DG, OPTIMISTIC Consortium Genetic determinants of disease severity in the myotonic dystrophy type 1 OPTIMISTIC cohort. Neurology. 2019;93:e995–e1009. doi: 10.1212/WNL.0000000000008056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Day JW, Ricker K, Jacobsen JF, Rasmussen LJ, Dick KA, Kress W, Schneider C, Koch MC, Beilman GJ, Harrison AR, Dalton JC, Ranum LPW. Myotonic dystrophy type 2: molecular, diagnostic and clinical spectrum. Neurology. 2003;60:657–664. doi: 10.1212/01.wnl.0000054481.84978.f9. [DOI] [PubMed] [Google Scholar]
- De Coster W, D’Hert S, Schultz DT, Cruts M, Van Broeckhoven C. NanoPack: visualizing and processing long-read sequencing data. Bioinformatics. 2018;34:2666–2669. doi: 10.1093/bioinformatics/bty149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeJesus-Hernandez M, Aleff RA, Jackson JL, Finch NA, Baker MC, Gendron TF, Murray ME, McLaughlin IJ, Harting JR, Graff-Radford NR, Oskarsson B, Knopman DS, Josephs KA, Boeve BF, Petersen RC, Fryer JD, Petrucelli L, Dickson DW, Rademakers R, Ebbert MTW, Wieben ED, van Blitterswijk M. Long-read targeted sequencing uncovers clinicopathological associations for c9orf72-linked diseases. Brain: A Journal of Neurology. 2021;144:1082–1088. doi: 10.1093/brain/awab006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Depienne C, Mandel JL. 30 years of repeat expansion disorders: what have we learned and what are the remaining challenges? American Journal of Human Genetics. 2021;108:764–785. doi: 10.1016/j.ajhg.2021.03.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ebbert MTW, Farrugia S, Sens J, Jansen-West K, Gendron TF, Prudencio M, McLaughlin lanJ, Bowman B, Seetin M, DeJesus-Hernandez M, Jackson J, Brown PH, Dickson DW, van Blitterswijk M, Rademakers R, Petrucelli L, Fryer JD. Long-Read Sequencing across the C9orf72 ‘GGGGCC’ Repeat Expansion: Implications for Clinical Use and Genetic Discovery Efforts in Human Disease. bioRxiv. 2018 doi: 10.1101/176651. [DOI] [PMC free article] [PubMed]
- Fontana L, Santoro M, D’Apice MR, Peluso F, Gori G, Morrone A, Novelli G, Dosa L, Botta A. Identification, molecular characterization and segregation analysis of a variant dmpk pre-mutation allele in a three-generation italian family. Acta Myologica. 2020;39:13–18. doi: 10.36185/2532-1900-002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fukuda H, Yamaguchi D, Nyquist K, Yabuki Y, Miyatake S, Uchiyama Y, Hamanaka K, Saida K, Koshimizu E, Tsuchida N, Fujita A, Mitsuhashi S, Ohbo K, Satake Y, Sone J, Doi H, Morihara K, Okamoto T, Takahashi Y, Wenger AM, Shioda N, Tanaka F, Matsumoto N, Mizuguchi T. Father-to-offspring transmission of extremely long NOTCH2NLC repeat expansions with contractions: genetic and epigenetic profiling with long-read sequencing. Clinical Epigenetics. 2021;13:1–17. doi: 10.1186/s13148-021-01192-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giesselmann P, Brändl B, Raimondeau E, Bowen R, Rohrandt C, Tandon R, Kretzmer H, Assum G, Galonska C, Siebert R, Ammerpohl O, Heron A, Schneider SA, Ladewig J, Koch P, Schuldt BM, Graham JE, Meissner A, Müller FJ. Analysis of short tandem repeat expansions and their methylation state with nanopore sequencing. Nature Biotechnology. 2019;37:1478–1481. doi: 10.1038/s41587-019-0293-x. [DOI] [PubMed] [Google Scholar]
- Gilpatrick T, Lee I, Graham JE, Raimondeau E, Bowen R, Heron A, Downs B, Sukumar S, Sedlazeck FJ, Timp W. Targeted nanopore sequencing with cas9-guided adapter ligation. Nature Biotechnology. 2020;38:433–438. doi: 10.1038/s41587-020-0407-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grosso V, Marcolungo L, Maestri S, Alfano M, Lavezzari D, Iadarola B, Salviati A, Mariotti B, Botta A, D’Apice MR, Novelli G, Delledonne M, Rossato M. Characterization of FMR1 repeat expansion and intragenic variants by indirect sequence capture. Frontiers in Genetics. 2021;12:743230. doi: 10.3389/fgene.2021.743230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo P, Lam SL. Unusual structures of CCTG repeats and their participation in repeat expansion. Biomolecular Concepts. 2016;7:331–340. doi: 10.1515/bmc-2016-0024. [DOI] [PubMed] [Google Scholar]
- Hafford-Tear NJ, Tsai YC, Sadan AN, Sanchez-Pintado B, Zarouchlioti C, Maher GJ, Liskova P, Tuft SJ, Hardcastle AJ, Clark TA, Davidson AE. CRISPR/cas9-targeted enrichment and long-read sequencing of the fuchs endothelial corneal dystrophy-associated TCF4 triplet repeat. Genetics in Medicine. 2019;21:2092–2102. doi: 10.1038/s41436-019-0453-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Höijer I, Tsai YC, Clark TA, Kotturi P, Dahl N, Stattin EL, Bondeson ML, Feuk L, Gyllensten U, Ameur A. Detailed analysis of HTT repeat elements in human blood using targeted amplification-free long-read sequencing. Human Mutation. 2018;39:1262–1272. doi: 10.1002/humu.23580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holgersen EM, Gandhi S, Zhou Y, Kim J, Vaz B, Bogojeski J, Bugno M, Shalev Z, Cheung-Ong K, Gonçalves J, O’Hara M, Kron K, Verby M, Sun M, Kakaradov B, Delong A, Merico D, Deshwar AG. Transcriptome-wide off-target effects of steric-blocking oligonucleotides. Nucleic Acid Therapeutics. 2021;31:392–403. doi: 10.1089/nat.2020.0921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hommelsheim CM, Frantzeskakis L, Huang M, Ülker B. PCR amplification of repetitive DNA: A limitation to genome editing technologies and many other applications. Scientific Reports. 2014;4:5052. doi: 10.1038/srep05052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iyer SV, Goodwin S, Kramer M, McCombie WR. Abstract 1360: understanding genetic variation in cancer using targeted nanopore long read sequencing. Cancer Research. 2020;80:1360. doi: 10.1158/1538-7445.AM2020-1360. [DOI] [Google Scholar]
- Iyer SV, Kramer M, Goodwin S, McCombie WR. ACME: An Affinity-Based Cas9 Mediated Enrichment Method for Targeted Nanopore Sequencing. bioRxiv. 2022 doi: 10.1101/2022.02.03.478550. [DOI]
- Kamsteeg EJ, Kress W, Catalli C, Hertz JM, Witsch-Baumgartner M, Buckley MF, van Engelen BGM, Schwartz M, Scheffer H. Best practice guidelines and recommendations on the molecular diagnosis of myotonic dystrophy types 1 and 2. European Journal of Human Genetics. 2012;20:1203–1208. doi: 10.1038/ejhg.2012.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Labun K, Montague TG, Krause M, Cleuren YNT, Tjeldnes H, Valen E. CHOPCHOP v3: expanding the CRISPR web toolbox beyond genome editing. Nucleic Acids Research. 2019;47:W171–W174. doi: 10.1093/nar/gkz365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leeflang EP, Arnheim N. A novel repeat structure at the myotonic dystrophy locus in A 37 repeat allele with unexpectedly high stability. Human Molecular Genetics. 1995;4:135–136. doi: 10.1093/hmg/4.1.135. [DOI] [PubMed] [Google Scholar]
- Lian M, Law HY, Lee CG, Chong SS. Defining the performance parameters of a rapid screening tool for myotonic dystrophy type 1 based on triplet-primed PCR and melt curve analysis. Expert Review of Molecular Diagnostics. 2016;16:1221–1232. doi: 10.1080/14737159.2016.1241145. [DOI] [PubMed] [Google Scholar]
- Liquori CL, Ricker K, Moseley ML, Jacobsen JF, Kress W, Naylor SL, Day JW, Ranum LPW. Myotonic dystrophy type 2 caused by a CCTG expansion in intron 1 of ZNF9. Science. 2001;293:864–867. doi: 10.1126/science.1062125. [DOI] [PubMed] [Google Scholar]
- Loomis EW, Eid JS, Peluso P, Yin J, Hickey L, Rank D, McCalmon S, Hagerman RJ, Tassone F, Hagerman PJ. Sequencing the unsequenceable: expanded CGG-repeat alleles of the fragile X gene. Genome Research. 2013;23:121–128. doi: 10.1101/gr.141705.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maestri S. MosaicViewer_CNBP. 95ff074GitHub. 2020 https://github.com/MaestSi/MosaicViewer_CNBP/blob/main/Figures/CNBP_left_right_alignment.png
- Maestri S. CharONT2. 8d27df9GitHub. 2022a https://github.com/MaestSi/CharONT2
- Maestri S. MosaicViewer_CNBP. f6fb8f2GitHub. 2022b https://github.com/MaestSi/MosaicViewer_CNBP
- Mahyera AS, Schneider T, Halliger-Keller B, Schrooten K, Hörner EM, Rost S, Kress W. Distribution and structure of DM2 repeat tract alleles in the german population. Frontiers in Neurology. 2018;9:463. doi: 10.3389/fneur.2018.00463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mangin A, de Pontual L, Tsai YC, Monteil L, Nizon M, Boisseau P, Mercier S, Ziegle J, Harting J, Heiner C, Gourdon G, Tomé S. Robust detection of somatic mosaicism and repeat interruptions by long-read targeted sequencing in myotonic dystrophy type 1. International Journal of Molecular Sciences. 2021;22:1–24. doi: 10.3390/ijms22052616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meola G, Cardani R. Myotonic dystrophy type 2: an update on clinical aspects, genetic and pathomolecular mechanism. Journal of Neuromuscular Diseases. 2015;2:S59–S71. doi: 10.3233/JND-150088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller JN, van der Plas E, Hamilton M, Koscik TR, Gutmann L, Cumming SA, Monckton DG, Nopoulos PC. Variant repeats within the DMPK CTG expansion protect function in myotonic dystrophy type 1. Neurology. Genetics. 2020;6:e504. doi: 10.1212/NXG.0000000000000504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitsuhashi S, Matsumoto N. Long-read sequencing for rare human genetic diseases. Journal of Human Genetics. 2020;65:11–19. doi: 10.1038/s10038-019-0671-8. [DOI] [PubMed] [Google Scholar]
- Mizuguchi T, Toyota T, Miyatake S, Mitsuhashi S, Doi H, Kudo Y, Kishida H, Hayashi N, Tsuburaya RS, Kinoshita M, Fukuyama T, Fukuda H, Koshimizu E, Tsuchida N, Uchiyama Y, Fujita A, Takata A, Miyake N, Kato M, Tanaka F, Adachi H, Matsumoto N. Complete sequencing of expanded SAMD12 repeats by long-read sequencing and cas9-mediated enrichment. Brain: A Journal of Neurology. 2021;144:1103–1117. doi: 10.1093/brain/awab021. [DOI] [PubMed] [Google Scholar]
- Mohammad NS, Nazli R, Zafar H, Fatima S. Effects of lipid based multiple micronutrients supplement on the birth outcome of underweight pre-eclamptic women: A randomized clinical trial. Pakistan Journal of Medical Sciences. 2022;38:219–226. doi: 10.12669/pjms.38.1.4396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monckton DG, Wong LJ, Ashizawa T, Caskey CT. Somatic mosaicism, germline expansions, germline reversions and intergenerational reductions in myotonic dystrophy males: small pool PCR analyses. Human Molecular Genetics. 1995;4:1–8. doi: 10.1093/hmg/4.1.1. [DOI] [PubMed] [Google Scholar]
- Montagnese F, Mondello S, Wenninger S, Kress W, Schoser B. Assessing the influence of age and gender on the phenotype of myotonic dystrophy type 2. Journal of Neurology. 2017;264:2472–2480. doi: 10.1007/s00415-017-8653-2. [DOI] [PubMed] [Google Scholar]
- Musova Z, Mazanec R, Krepelova A, Ehler E, Vales J, Jaklova R, Prochazka T, Koukal P, Marikova T, Kraus J, Havlovicova M, Sedlacek Z. Highly unstable sequence interruptions of the CTG repeat in the myotonic dystrophy gene. American Journal of Medical Genetics. Part A. 2009;149A:1365–1374. doi: 10.1002/ajmg.a.32987. [DOI] [PubMed] [Google Scholar]
- Nakamori M, Sobczak K, Moxley RT, Thornton CA. Scaled-down genetic analysis of myotonic dystrophy type 1 and type 2. Neuromuscular Disorders. 2009;19:759–762. doi: 10.1016/j.nmd.2009.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oxford Nanopore Community Oxford Nanopore Technology. 2021. [May 4, 2022]. https://community.nanoporetech.com/posts/can-ffpe-repair-be-avoided
- Oxford Nanopore Community Oxford Nanopore Technology. 2022. [October 31, 2020]. https://community.nanoporetech.com/posts/high-of-unclassified-rea
- Payne A, Holmes N, Rakyan V, Loose M. BulkVis: A graphical viewer for oxford nanopore bulk FAST5 files. Bioinformatics. 2019;35:2193–2198. doi: 10.1093/bioinformatics/bty841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pešović J, Perić S, Brkušanin M, Brajušković G, Rakočević-Stojanović V, Savić-Pavićević D. Repeat interruptions modify age at onset in myotonic dystrophy type 1 by stabilizing dmpk expansions in somatic cells. Frontiers in Genetics. 2018;9:601. doi: 10.3389/fgene.2018.00601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Radvansky J, Ficek A, Minarik G, Palffy R, Kadasi L. Effect of unexpected sequence interruptions to conventional PCR and repeat primed PCR in myotonic dystrophy type 1 testing. Diagnostic Molecular Pathology. 2011;20:48–51. doi: 10.1097/PDM.0b013e3181efe290. [DOI] [PubMed] [Google Scholar]
- Radvanszky J, Surovy M, Polak E, Kadasi L. Uninterrupted CCTG tracts in the myotonic dystrophy type 2 associated locus. Neuromuscular Disorders. 2013;23:591–598. doi: 10.1016/j.nmd.2013.02.013. [DOI] [PubMed] [Google Scholar]
- Santoro M, Masciullo M, Pietrobono R, Conte G, Modoni A, Bianchi MLE, Rizzo V, Pomponi MG, Tasca G, Neri G, Silvestri G. Molecular, clinical, and muscle studies in myotonic dystrophy type 1 (DM1) associated with novel variant CCG expansions. Journal of Neurology. 2013;260:1245–1257. doi: 10.1007/s00415-012-6779-9. [DOI] [PubMed] [Google Scholar]
- Santoro M, Fontana L, Masciullo M, Bianchi MLE, Rossi S, Leoncini E, Novelli G, Botta A, Silvestri G. Expansion size and presence of CCG/CTC/CGG sequence interruptions in the expanded CTG array are independently associated to hypermethylation at the DMPK locus in myotonic dystrophy type 1 (DM1) Biochimica et Biophysica Acta. 2015;1852:2645–2652. doi: 10.1016/j.bbadis.2015.09.007. [DOI] [PubMed] [Google Scholar]
- Santoro M, Masciullo M, Silvestri G, Novelli G, Botta A. Myotonic dystrophy type 1: role of CCG, CTC and CGG interruptions within DMPK alleles in the pathogenesis and molecular diagnosis. Clinical Genetics. 2017;92:355–364. doi: 10.1111/cge.12954. [DOI] [PubMed] [Google Scholar]
- Santoro M, Fontana L, Maiorca F, Centofanti F, Massa R, Silvestri G, Novelli G, Botta A. Expanded [cctg]n repetitions are not associated with abnormal methylation at the cnbp locus in myotonic dystrophy type 2 (dm2) patients. Biochimica et Biophysica Acta. Molecular Basis of Disease. 2018;1864:917–924. doi: 10.1016/j.bbadis.2017.12.037. [DOI] [PubMed] [Google Scholar]
- Sergey N, Koren S, Rhie A, Rautiainen M, Bzikadze AV, Mikheenko A, Vollger MR, Altemose N, Uralsky L, Gershman A, Aganezov S, Hoyt SJ, Diekhans M, Logsdon GA, Alonge M, Antonarakis SE, Borchers M, Bouffard GG, Brooks SY, Caldas GV, Chen NC, Cheng H, Chin CS, Chow W, de Lima LG, Dishuck PC, Durbin R, Dvorkina T, Fiddes IT, Formenti G, Fulton RS, Fungtammasan A, Garrison E, Grady PGS, Graves-Lindsay TA, Hall IM, Hansen NF, Hartley GA, Haukness M, Howe K, Hunkapiller MW, Jain C, Jain M, Jarvis ED, Kerpedjiev P, Kirsche M, Kolmogorov M, Korlach J, Kremitzki M, Li H, Maduro VV, Marschall T, McCartney AM, McDaniel J, Miller DE, Mullikin JC, Myers EW, Olson ND, Paten B, Peluso P, Pevzner PA, Porubsky D, Potapova T, Rogaev EI, Rosenfeld JA, Salzberg SL, Schneider VA, Sedlazeck FJ, Shafin K, Shew CJ, Shumate A, Sims Y, Smit AFA, Soto DC, Sović I, Storer JM, Streets A, Sullivan BA, Thibaud-Nissen F, Torrance J, Wagner J, Walenz BP, Wenger A, Wood JMD, Xiao C, Yan SM, Young AC, Zarate S, Surti U, McCoy RC, Dennis MY, Alexandrov IA, Gerton JL, O’Neill RJ, Timp W, Zook JM, Schatz MC, Eichler EE, Miga KH, Phillippy AM. The complete sequence of a human genome. Science. 2022;376:44–53. doi: 10.1126/science.abj6987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siena Cd, Cardani R, Brigonzi E, Bosè F, Fossati B, Meola G, Costa E, Valaperta R. Incidence of amplification failure in DMPK allele due to allelic dropout event in a diagnostic laboratory. Clinica Chimica Acta; International Journal of Clinical Chemistry. 2018;484:111–116. doi: 10.1016/j.cca.2018.05.040. [DOI] [PubMed] [Google Scholar]
- Sone J, Mitsuhashi S, Fujita A, Mizuguchi T, Hamanaka K, Mori K, Koike H, Hashiguchi A, Takashima H, Sugiyama H, Kohno Y, Takiyama Y, Maeda K, Doi H, Koyano S, Takeuchi H, Kawamoto M, Kohara N, Ando T, Ieda T, Kita Y, Kokubun N, Tsuboi Y, Katoh K, Kino Y, Katsuno M, Iwasaki Y, Yoshida M, Tanaka F, Suzuki IK, Frith MC, Matsumoto N, Sobue G. Long-read sequencing identifies GGC repeat expansions in NOTCH2NLC associated with neuronal intranuclear inclusion disease. Nature Genetics. 2019;51:1215–1221. doi: 10.1038/s41588-019-0459-y. [DOI] [PubMed] [Google Scholar]
- Stevanovski I, Chintalaphani SR, Gamaarachchi H, Ferguson JM, Pineda SS, Scriba CK, Tchan M, Fung V, Ng K, Cortese A, Houlden H, Dobson-Stone C, Fitzpatrick L, Halliday G, Ravenscroft G, Davis MR, Laing NG, Fellner A, Kennerson M, Kumar KR, Deveson IW. Comprehensive genetic diagnosis of tandem repeat expansion disorders with programmable targeted nanopore sequencing. Science Advances. 2022;8:eabm5386. doi: 10.1126/sciadv.abm5386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thornton CA. Myotonic dystrophy. Neurologic Clinics. 2014;32:705–719. doi: 10.1016/j.ncl.2014.04.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomé S, Dandelot E, Dogan C, Bertrand A, Geneviève D, Péréon Y, Simon M, Bonnefont JP, Bassez G, Gourdon G, DM contraction study group Unusual association of a unique CAG interruption in 5’ of DM1 CTG repeats with intergenerational contractions and low somatic mosaicism. Human Mutation. 2018;39:970–982. doi: 10.1002/humu.23531. [DOI] [PubMed] [Google Scholar]
- Tsai YC, Greenberg D, Powell J, Höijer I, Ameur A, Strahl M, Ellis E, Jonasson I, Mouro Pinto R, Wheeler VC, Smith ML, Gyllensten U, Sebra R, Korlach J, Clark TA. Amplification-Free, CRISPR-Cas9 Targeted Enrichment and SMRT Sequencing of Repeat-Expansion Disease Causative Genomic Regions. bioRxiv. 2017 doi: 10.1101/203919. [DOI]
- Udd B, Meola G, Krahe R, Thornton C, Ranum L, Day J, Bassez G, Ricker K. Report of the 115th ENMC workshop: DM2/PROMM and other myotonic dystrophies. 3rd workshop, 14-16 february 2003, naarden, the netherlands. Neuromuscular Disorders. 2003;13:589–596. doi: 10.1016/s0960-8966(03)00092-0. [DOI] [PubMed] [Google Scholar]
- Wallace AD, Sasani TA, Swanier J, Gates BL, Greenland J, Pedersen BS, Varley KE, Quinlan AR. CaBagE: A cas9-based background elimination strategy for targeted, long-read DNA sequencing. PLOS ONE. 2021;16:e0241253. doi: 10.1371/journal.pone.0241253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wieben ED, Aleff RA, Basu S, Sarangi V, Bowman B, McLaughlin IJ, Mills JR, Butz ML, Highsmith EW, Ida CM, Ekholm JM, Baratz KH, Fautsch MP. Amplification-free long-read sequencing of TCF4 expanded trinucleotide repeats in fuchs endothelial corneal dystrophy. PLOS ONE. 2019;14:e0219446. doi: 10.1371/journal.pone.0219446. [DOI] [PMC free article] [PubMed] [Google Scholar]