

Abstract
Fibrolamellar carcinoma (FLC) is a rare type of liver cancer that affects adolescents and young adults. The most effective treatment for FLC is surgical resection, but no standardized systemic therapy exists for patients with recurrent or unresectable FLC. As a first step to understand the immune microenvironment of FLC, we investigated targetable immune-checkpoint pathways, PD-1, PD-L1, B7-H3, IDO-1, and LAG3, in relation to CD8+ cytotoxic T-lymphocyte density. Thirty-two FLC tumor specimens were analyzed using IHC staining for PD-L1, CD8, PD-1, IDO, LAG3, and B7-H3. Sixty-three percent of FLC cases demonstrated membranous PD-L1 expression on tumor cells, and almost 70% of cases demonstrated PD-L1+ tumor-infiltrating lymphocytes and tumor-associated macrophages (TIL/TAM). Myeloid-derived cells appeared to be a major component of PD-L1+ tumor-infiltrating immune cells. Forty percent of the cases showed B7-H3 expression in the tumor zone, with 91% cases showing B7-H3 expression in TILs and TAMs. IDO and PD-1 expression was highest in the tumor interface zone. B7-H3 or IDO expression on tumor cells significantly correlated with higher CD8+ T-cell density. In conclusion, a high proportion of FLC cases showed robust expression of PD-1, PD-L1, B7-H3, and IDO in an adaptive immune-resistance pattern. Our findings provide further basis for targeting these different immune-checkpoint axes in FLC.
Introduction
Fibrolamellar carcinoma (FLC) is a liver cancer that was formally categorized as a subset of hepatocellular carcinoma (HCC) but now recognized as an entirely different cancer with unique clinicopathologic and molecular features (1, 2). Unlike classic HCC, FLC affects adolescents and young adults without chronic liver disease. FLC is associated with fewer chromosomal changes and less genomic heterogeneity than HCC and is characterized by the presence of a single genetic deletion in chromosome 19 and formation of a fusion of DNAJB1 and PRKACA transcripts in 79% to 100% of the cases (1–3). The most effective treatment for FLC is complete surgical resection. However, outcomes for patients with FLC are poor, with an overall survival of 30% at 5 years (4, 5), and currently, no standardized systemic treatment recommendation exists for patients with recurrent or unresectable disease. Because of this, novel and effective therapies for patients with FLC are needed.
Data on systemic treatment of FLC are limited. One retrospective study demonstrated that the combination of 5-fluorouracil and subcutaneous recombinant interferon alfa (IFNα) has clinical activity for FLC (6). The potential clinical activity of IFNα in FLC suggests that immune evasion may be an important feature of FLC oncogenesis and, therefore, may be an opportunity for therapeutic intervention. Little is known, however, about the immune microenvironment and the mechanisms of immune evasion in FLC.
Immune-checkpoint molecules such as programmed death-1 (PD-1), and its ligand (PD-L1), modulate cellular immunity and can be co-opted by cancers to sustain immune tolerance. The development of immune-checkpoint inhibitors has had a profound impact on the clinical management of many cancers, including liver cancer (7). To date, immunotherapy is not routinely administered in FLC because of the rarity of the disease and the uncertainty in its treatment response. The direct analysis of the cancer immune microenvironment has also emerged as an important step in understanding the relationship between the patients’ immune systems and their cancer. For example, PD-L1 expression on tumor cells has been identified among multiple tumor types that respond to PD-1 pathway inhibition, including melanoma, lung, and renal carcinoma (8, 9). PD-L1 expression on tumor-infiltrating immune cells has also been associated with higher clinical response rate to inhibitors of the PD-1 pathway (10–12).
To date, the expression of immune-checkpoint molecules has not previously been assessed in FLC. The objective of the current study was to define the expression of checkpoint molecules PD-1, PD-L1, IDO, B7-H3, and LAG-3 on tumors and tumor immune infiltrates in a cohort of patients with FLC. The findings provide insights about the immune microenvironment in FLC.
Materials and Methods
Case selection
The study was approved by the Johns Hopkins University Institutional Review Board and conducted in accordance with the ethical standards of the Declaration of Helsinki. All samples were obtained in accordance with the Health Insurance Privacy and Accountability Act. Specimens from 25 unique patients with FLC were obtained between 2000 and 2016 from the Johns Hopkins Hospital surgical pathology archives (n = 22) and the StoreMyTumor tumor bank (http://www.StoreMyTumor.com; n = 3). From these 25 patients, 32 specimens were studied (18 patients with one specimen on file, and 8 patients with multiple specimens). Hematoxylin and eosin–stained slides were reviewed by two pathologists (F. Bhaijee and R. Anders) to confirm the diagnosis of FLC, and a single representative block was chosen for additional analysis. Clinical information such as age, sex, and previous treatment history was extracted from the electronic medical record for the 22 patients from Johns Hopkins Hospital (Table 1).
Table 1.
Patient and tumor characteristics
| Patient/tumor characteristics | N (%) |
|---|---|
| Median age (IQR), years | 23.5 (19.5–29.5) |
| Patient sexa | |
| Male | 9 (41) |
| Female | 13 (59) |
| Patient race (white) | 22 (100.0) |
| Location of primary sampleb | |
| Liver | 20 (63) |
| Lymph node | 2 (6) |
| Other metastasis sites | 10 (31) |
| Median tumor size (IQR), cmb | 10.0 (3.5–11.2) |
| Disease recurrence | |
| No | 4 (18) |
| Yes | 11 (50) |
| Unknown | 7 (32) |
| Died during follow-up | |
| No | 8 (36) |
| Yes | 7 (32) |
| Unknown | 7 (32) |
Clinical characteristics only include patients from Johns Hopkins Hospital (n = 22).
Analysis presented here on tumor samples. The denominator for these analyses is 32.
IHC staining, slide scanning, and annotation
Serial 5-μm sections from formalin-fixed, paraffin-embedded tissue were cut onto glass slides for each specimen. IHC staining for PD-L1, CD8, Foxp3, PD-1, IDO, LAG3, CD11c, CD68, and B7-H3 was performed by the Tumor Microenvironment Center at Johns Hopkins School of Medicine and the IHC laboratory in Johns Hopkins Hospital, using the reagents listed in Supplementary Table S1. For IDO, automated IDO1 staining was performed on the Leica Bond RX (Leica Biosystems) using the Bond Polymer Refine Kit (DS9800, Leica Biosystems). Slides were baked online using a *Bake and Dewax protocol, followed by high pH antigen retrieval using a *HIER 20 min with ER2 protocol. At room temperature, endogenous peroxidase was blocked using Peroxide Block (Leica Biosystems) for 10 minutes, followed by blocking of nonspecific binding using Protein block (X090930, Agilent Technologies) for 15 minutes. Anti-IDO1 (clone SP260; ab228468, Abcam plc.) was applied for 15 minutes at a concentration of 1.19 μg/mL. Detection was performed by applying anti-rabbit polymer-HRP (Leica Biosystems) for 8 minutes followed by DAB Refine (Leica Biosystems) for 10 minutes. Samples were counter-stained using hematoxylin (Leica Biosystems) for 5 minutes. Slides were dehydrated through a graded alcohol series into Histoclear II (508990150, Thermo Fisher Scientific) and cover-slipped using Ecomount (5082832, Biocare Medical). Human tonsil specimens were used as positive controls for each staining batch. An IgG control was performed for each individual case as a control for PD-L1 staining.
IHC scoring for PD-L1, B7-H3, and Lag3
Scoring for PD-L1, B7-H3, and LAG3 was performed by two separate pathologists (R. Anders and A.J. Layman) blinded to the clinicopathologic features of the cases using standard brightfield microscopy (Nikon Eclipse 50i). The percentage of malignant cells demonstrating positive stain expression was scored at 5% increments, and cases demonstrating ≥ 5% tumor cell expression of a given marker were considered positive (8, 13–15). Immune cells, including tumor-infiltrating lymphocytes and tumor-associated macrophages (TIL/TAM), were scored separately as positive (≥ 5% of cells staining). The scoring of the two pathologists was subsequently averaged into a combined final score.
Slide scanning and scoring for CD8, PD-1, Foxp3, and IDO
Slides were scanned at 20 × objective equivalent (0.49 μm/pixel) using the Hamamatsu XR NanoZoomer bright field system (Hamamatsu Photonics, K.K.). For CD8-, Foxp3-, PD-1–, and IDO-stained slides, the tumor–stroma interface was annotated. Specifically, 250 μm from the tumor–stromal border into the tumor center and further into the stroma from this demarcation were considered the “tumor interface” and “nontumor interface” zones, respectively. Additional tumor areas studied included the remaining central aspect of the tumor (referred to here as “tumor” zone), and the areas further into the stroma (away from the tumor) beyond the nontumor interface (referred to here as “nontumor” zone). Greater than 50% of the nontumor area was annotated for analysis (Supplementary Fig. S1). Digitally assisted image analysis (HALO Indica Labs) was used to assess the density (cells/mm2) of cells expressing CD8, PD-1, Foxp3, and IDO in each of the four tumor microenvironment (TME) zones described above.
Statistical analysis
Categorical variables were reported as whole numbers and proportions, whereas continuous variables were summarized as medians with interquartile range (IQR) as appropriate. Pearson χ2 test was used to compare categorical variables (PDL-1 and B7-H3 staining) between patient groups, whereas quintile regression of medians was used to compare continuous data (CD8+, Foxp3+, IDO+, and PD-1+ cell densities). For all analyses, a P value of ≤0.05 was used to define statistical significance. All analyses were performed using STATA 14.0 (Stata Corp.).
Results
The pathologic features of 32 FLC specimens are provided in Table 1. Twenty specimens (63%) were from primary tumor, two (6%) from lymph nodes, and ten (31%) were from other metastatic sites. Clinical characteristics were reported for patients from Johns Hopkins Hospital (n = 22). An even sex distribution between males and females was seen, and median patient age was 24. The majority of the patients did not receive any systemic treatment prior to their surgery. Fifty percent of the patients had disease recurrence after surgery. Other detailed treatment history and clinical outcome at the time of data collection are shown in Supplementary Table S2.
Expression of PD-L1, B7-H3, IDO, and LAG-3 in FLC tumor cells and immune infiltrates
As shown in Fig. 1A, 20 of the 32 cases (63%) demonstrated membranous tumor cell expression of PD-L1 in the tumor zone, and 22 cases (69%) had positive PD-L1 expression present on the tumor-infiltrating immune cells. In contrast, only 16% of cases showed PD-L1+ nonneoplastic hepatocytes, which was significantly lower than the cases with positive tumor cells (P = 0.007). Forty-one percent of the cases had PD-L1 expression on immune cells in the nontumor zone. Of the cases with PD-L1 expression on TILs and TAMs, 46% of the cases had greater than 25% of the cells stained positive.
Figure 1.
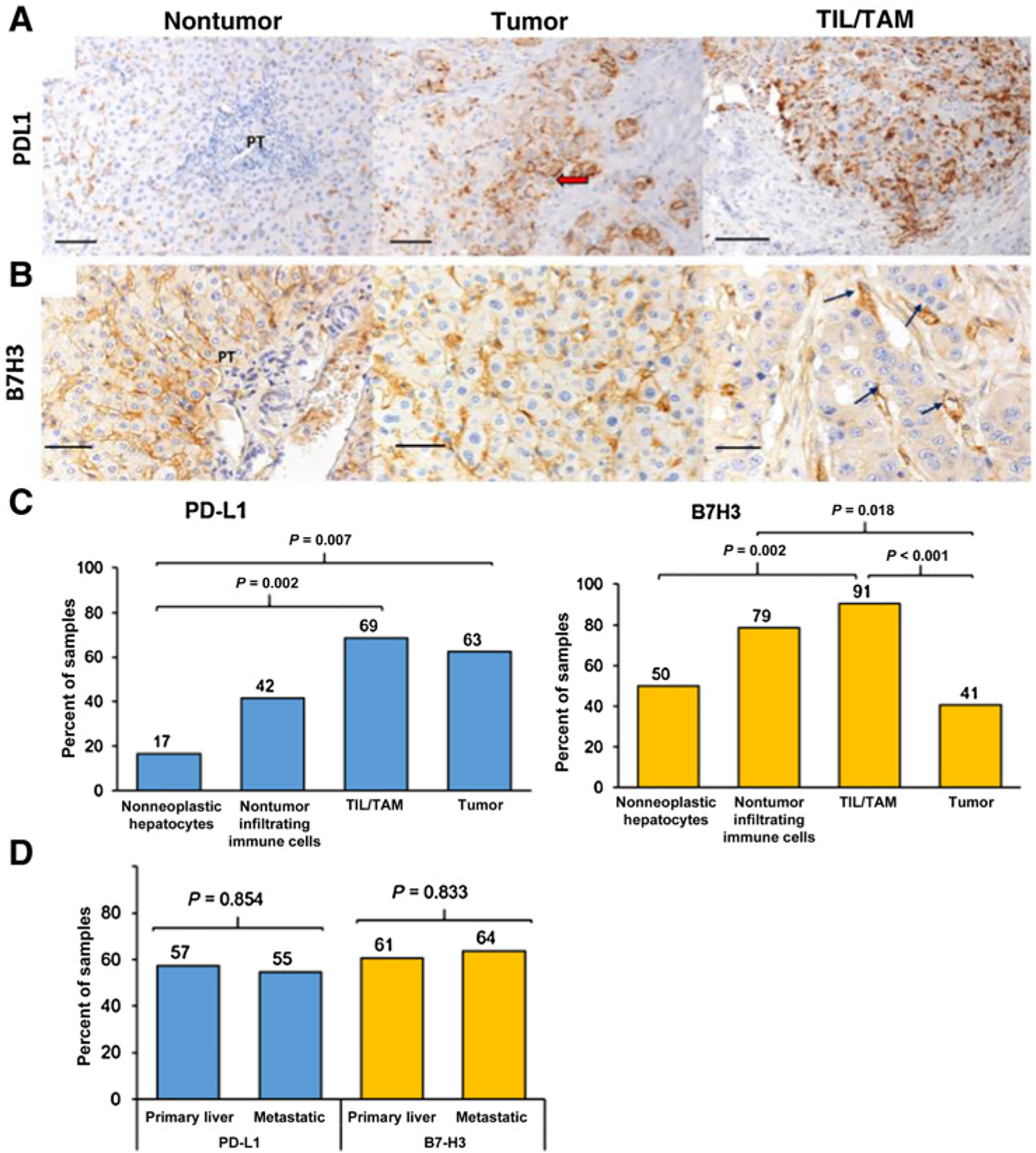
PD-L1 and B7-H3 in the FLC TME. A, Specimens were stained with PD-L1, and its expression in nontumor hepatocytes, tumor cells, and tumor-infiltrating lymphocytes and macrophages (TILs/TAMs) is shown. Red arrow shows membranous expression of PD-L1 in tumor cells. PT, portal tract. B, Specimens were stained with B7-H3, and its expression in nontumor hepatocytes, tumor cells, and TILs/TAMs is shown. Immune cells lining the sinusoids are indicated with black arrows. C, Percentages indicate the proportion of positively stained specimens in each representative region of FLC for PD-L1 and B7-H3 using 32 tumor regions and 15 nontumor regions per patient specimen. D, Comparison of primary tumor (n = 20) and metastatic sites (n = 12) for PD-L1 and B7-H3 expression in tumor and TILs/TAMs. Specimens shown in 20 magnification with a 100-μm scale bar, except for B7-H3 TILs/TAMs (B, right), which is at 40 × with a 50-μm scale bar. Pearson χ2 test was used between patient groups with a P value of ≤ 0.05 to define statistical significance.
We also investigated the expression of B7-H3, another member of the B7-family closely related to PD-L1 and known to modulate T cells in FLC (Fig. 1B). It is established that B7-H3 is expressed in numerous solid tumors including lung, stomach, prostate, and HCC (16–18). Overall, expression of B7-H3 in the entire TME of FLC was observed. Forty percent of the cases showed B7-H3+ tumor cells in the tumor zone, which was similar to the adjacent nontumor tissue (50%). Ninety-one percent of the cases had B7-H3+ TILs and TAMs, and 79% showed B7-H3+ nontumor immune cells (Fig. 1C). As seen with PD-L1 staining on primary FLC samples, almost all immune cells, including TILs and TAMs, were located in the sinusoids of the liver, which are known to be enriched with immune cells such as macrophages (Kupffer cells) and liver-associated natural killer cells (19, 20). Comparison of the primary tumors to metastatic sites did not show any significant difference in PD-L1 and B7-H3 expression in tumor cells or TILs and TAMs (Fig. 1D).
Indoleamine 2,3-dioxygenase (IDO) is another major inhibitor of immune responses (10, 21), and its expression in tumor cells and tumor-associated immune cells is associated with a worse prognosis (22, 23). In the FLC samples, IDO expression was concentrated at the tumor interface (Fig. 2A), and its cell density was almost 3-fold greater (44.8 cells/mm2) compared with other zones (Fig. 2B). However, no significant difference in IDO expression was observed between the tumor versus nontumor interface, as well as the nontumor zones (15.4, 9.7, and 15.9 cells/mm2, respectively).
Figure 2.
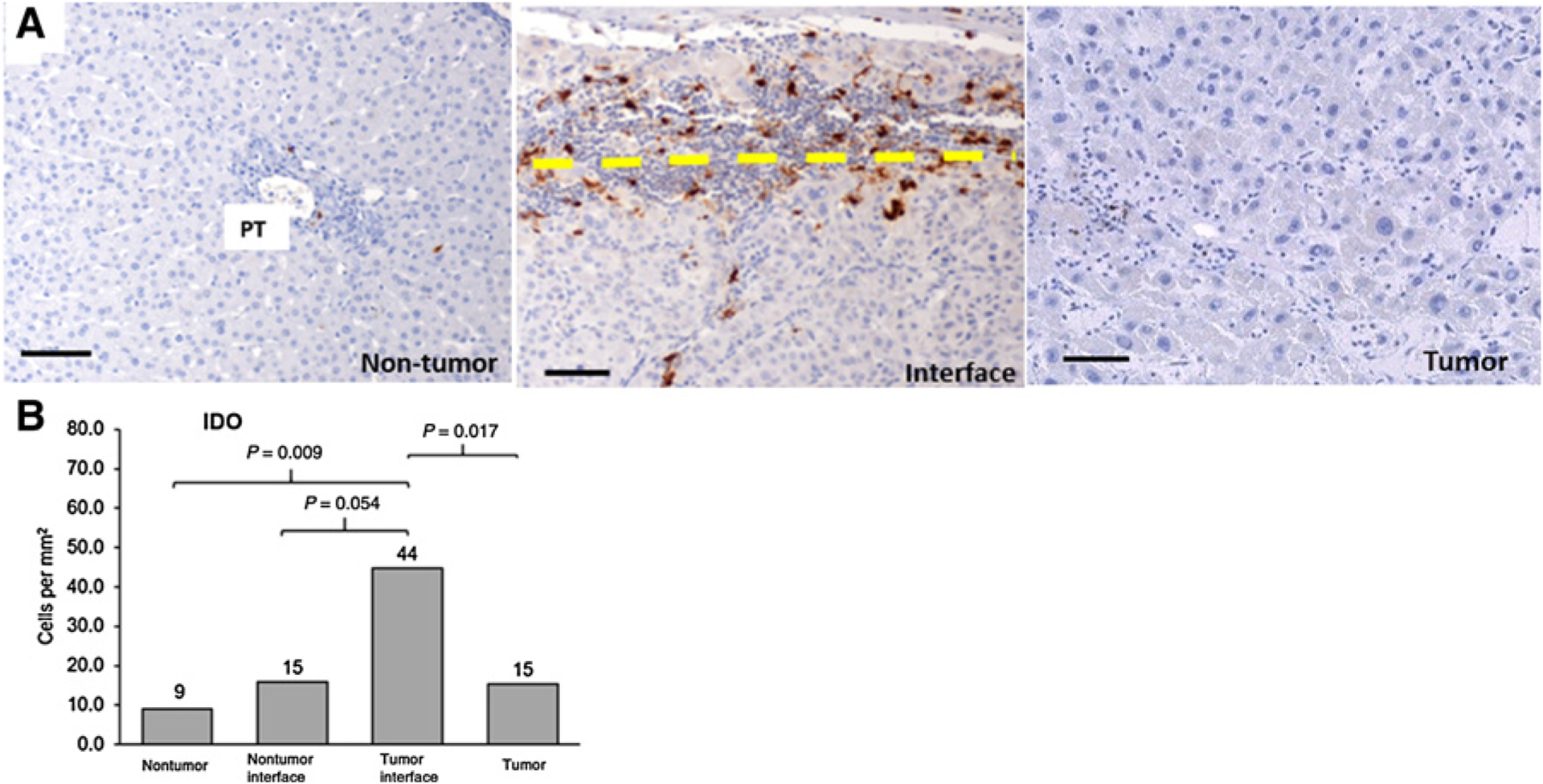
A, IDO expression in the TME. Representative images of IDO expression in tumor and nontumor zones. The interface was further stratified as tumor-side (above the dashed line) and nontumor side (below the dashed line). Dark brown stains indicate IDO expression. PT, portal tract. B, Median cell densities of each region are shown above each bar. Scale bar, 100 μm. 20 × magnification. Thirty-one tumor regions, 10 tumor and nontumor interface regions, and 15 nontumor regions were analyzed. Quintile regression analysis was used with a P value of ≤ 0.05 to define statistical significance.
The expression of LAG3 was also investigated in this cohort in the first 22 samples. In contrast to other checkpoint inhibitor expression in our FLC samples, only 10% of the samples (2 of 22 samples) had less than 1% of lymphocyte expression in the tumor area, as shown in Supplementary Table S3.
CD8+, Foxp3+, and PD-1+ cell densities in the TME of FLC
Given that T-cell activation is critical to the development of the adaptive immune resistance associated with PD-L1 expression in tumors (13), we quantified standard T-cell subsets using antibodies to CD8+, as well as Foxp3+ [representative of regulatory T cells (Tregs)] in the tumor, peritumoral interface, and nontumor regions. Overall, the median density of CD8+ T lymphocytes was significantly higher in the tumor interface zone compared with the tumor zone (P = 0.037), and the CD8+ density was similar in the tumor zone versus the nontumor zone (71 cells/mm2 and 111 cells/mm2, respectively; P = 0.557). This was consistent with our previous finding in HCC, where the mean CD8+ T-lymphocyte density was highest at the tumor interface and lowest in the tumor region (24). Foxp3+ cell density was lower compared with CD8+ cells (Fig. 3), but no statistically significant differences in Foxp3+ cell density were observed among the tumor regions.
Figure 3.
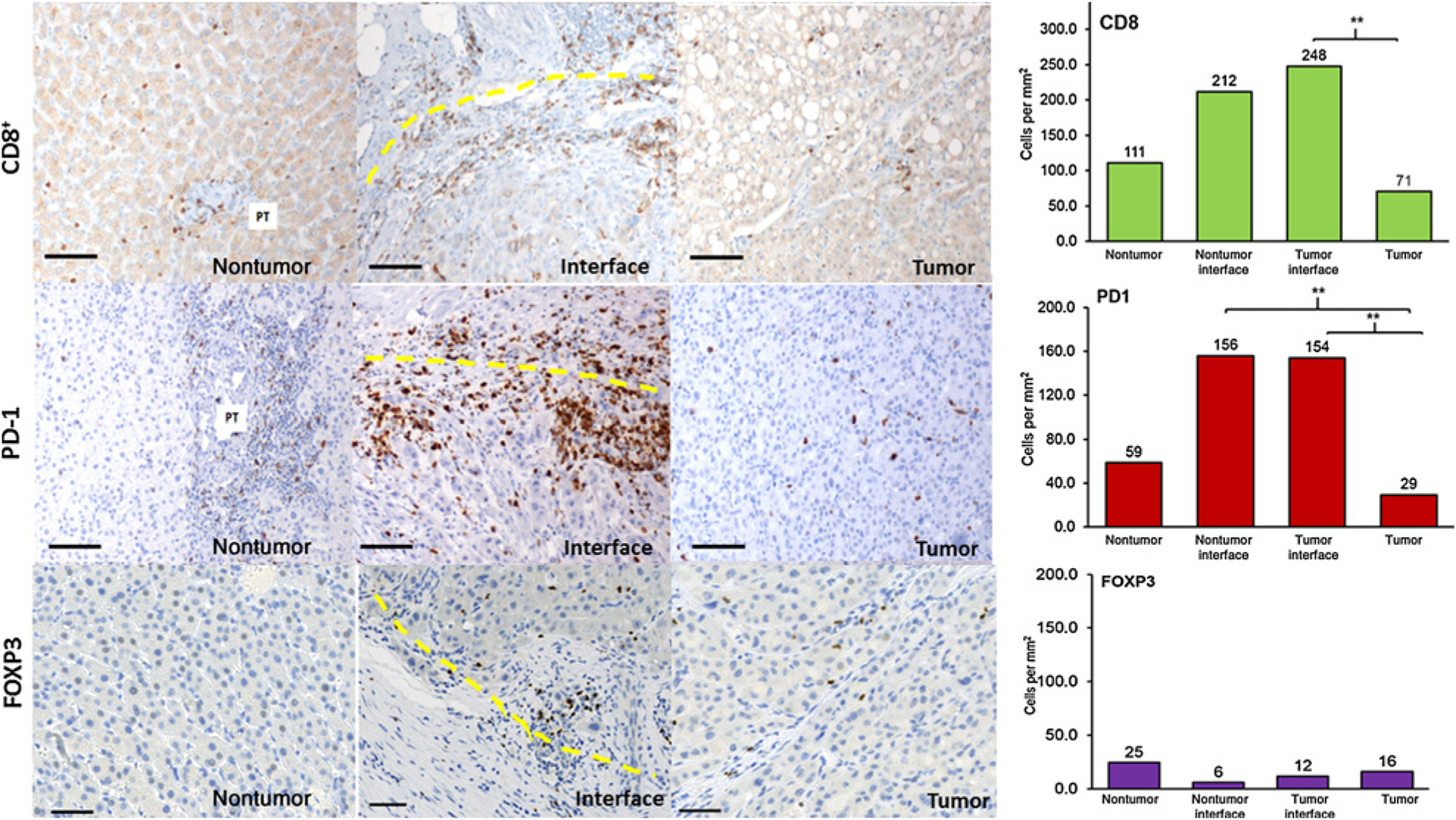
CD8+, PD-1+, and FOXP3+ expression in the TME. Nontumor regions, peritumoral interface, and tumor regions were stained by CD8+ (top row), PD-1+ (middle row), and FOXP3+ (bottom row) markers. Interface zone: yellow line. Scale bar, 100 μm. 20 × magnification. Graph: median cell densities of each region are shown above each bar graph. Thirty-one tumor regions, 10 tumor and nontumor interface regions, and 15 nontumor regions were analyzed. Quintile regression analysis was used with a P value of ≤ 0.05 to define statistical significance, as noted by **.
Similar to the distribution of CD8+ cells, a high concentration of PD-1+ lymphocytes in the interface area was seen compared with the tumor and nontumor areas (Fig. 3), and cell densities were similar for the tumor and nontumor side of the interface for PD-1. No differences in CD8+ and PD-1+ cell densities were observed between tumor and nontumor regions (P = 0.598).
Association of immune-activating molecules and immune-checkpoint marker expression
Given the association of B7-H3 and PD-L1 to T-cell activation and infiltration in tumors (13, 25, 26), CD8+ T-cell densities in B7-H3+ tumor samples were compared with B7-H3− tumor samples. The median CD8+ T-lymphocyte density in B7-H3+ tumor samples was more than 5-fold greater than in B7-H3− samples (245 vs. 33.1 cells/mm2, P < 0.001; Fig. 4A). These data suggested that despite similar B7-H3 expression between tumor and nontumor zones (Fig. 1), there was a correlation between T-cell activation and expression of B7H3 in tumor area. A similar correlation was seen between CD8+ and PD-L1+ tumor samples, although it did not meet statistical significance (Fig. 4A). The median CD8+ T-lymphocyte density was 128.3 cells/mm2 in samples with PD-L1+ tumor cells versus 34.9 cells/mm2 in samples with PD-L1− tumor cells (P = 0.184). A similar pattern was seen in adjacent nontumor regions, where CD8+ T-lymphocyte density (312.2 cells/mm2) was almost 3-fold higher in samples with PD-L1+ nontumor hepatocytes versus the samples with PD-L1− cells (124.2 cells/mm2, P = 0.089). A linear relationship between IDO+ cell density in the tumor zone with CD8+ density (P < 0.001) was noted, with a similar positive trend in the interface region (P = 0.289; Fig. 4B), also suggesting a local interaction between T cells and its ligand, IDO, in the FLC microenvironment.
Figure 4.
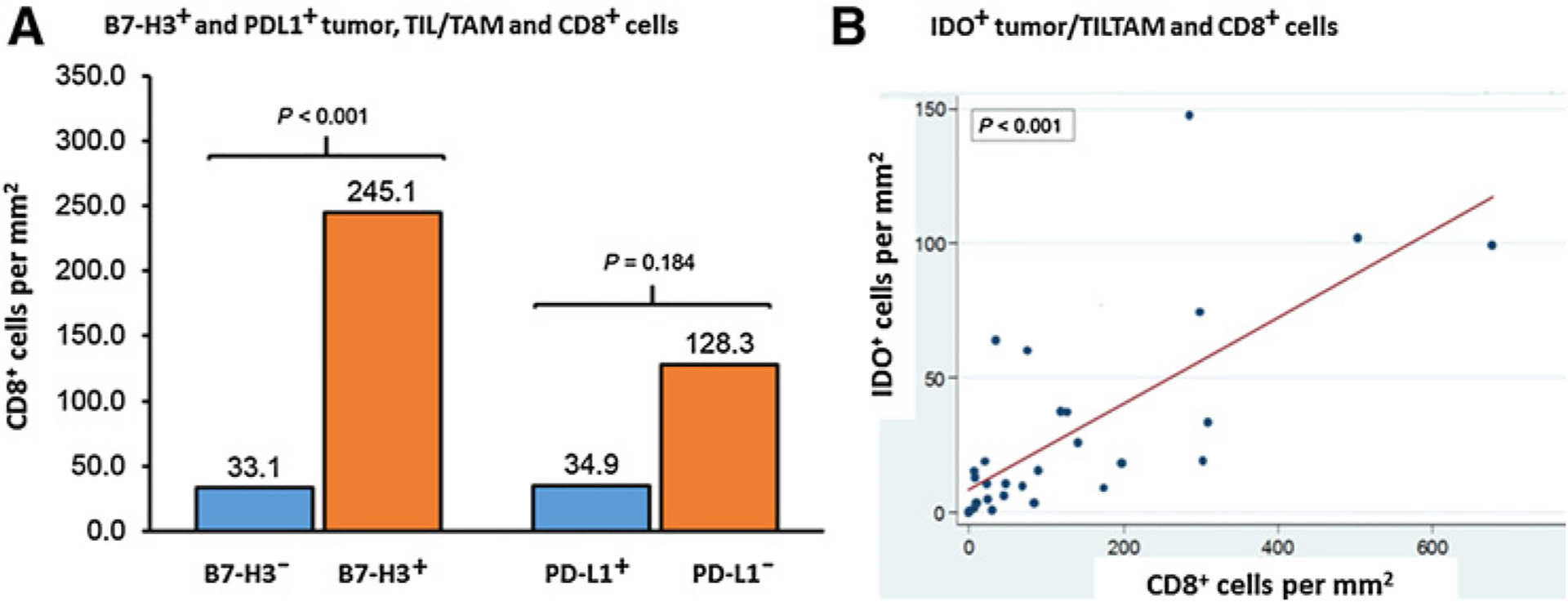
CD8+ T cells correlate with B7H3+, PD-L1+, and IDO+ tumor cells and TILs/TAMs. A, Median CD8+ T-cell density between B7-H3+ tumors, TILs, TAMs (orange bar), and B7-H3− tumors, TILs, TAMs (blue) are compared in the left side of the panel. Mean CD8+ cell density values are indicated above each bar. Median CD8+ T-cell density between PD-L1+ (orange) and PD-L1− (blue) specimens is shown on the right. A total of 32 tumor regions per specimen were used in the analysis. Quintile regression of medians was used with a P value of ≤ 0.05 to define statistical significance. B, Correlation analysis between the cell densities for IDO+ and CD8+ T cells was performed using linear regression and correlation coefficient. TIL/TAM: tumor-infiltrating lymphocyte/tumor-associated macrophage.
In melanoma and ovarian cancer patients, PD-L1 expression on myeloid cells, including myeloid dendritic cells (DC) and macrophages, has a critical function in T-cell immunity and in response to immunotherapy (27, 28). To investigate the role of myeloid cells in FLC, we selected a subset of the FLC specimens with more than >25% PD-L1 expression in TILs and TAMs and stained for DCs and macrophages with CD11c and CD68 antibodies, respectively. Phenotypically, these myeloid-derived cells appeared to be a major component of PD-L1+ nontumor cells (Fig. 5), with more CD11c+ cells within the tumor-infiltrating immune cells than CD68+ cells, suggesting an important role of PD-L1–expressing antigen-presenting cells (APC) in immune microenvironment of FLC.
Figure 5.
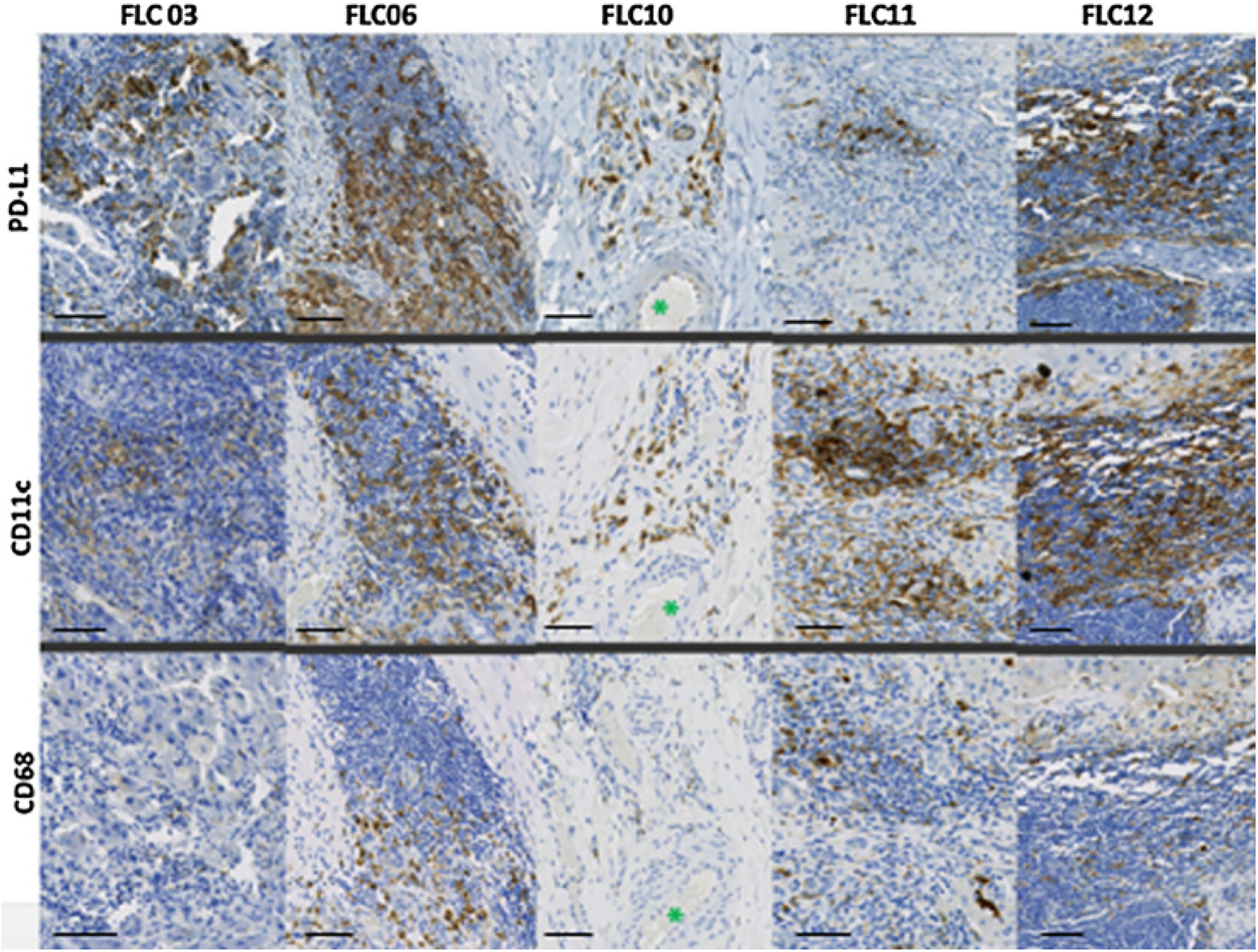
PD-L1+ expression in myeloid-derived cells. Five selected specimens with PD-L1+ expression were stained for myeloid-derived cells by staining for CD11c and CD68. The same location of the tissue is presented for comparison within each sample. Asterisks indicate an artery for location comparison. Specimens shown in 20 × magnification with a 100-μm scale bar.
Relationship among immune-checkpoint markers
We compared the PD-1 and IDO cell densities in tumors with PD-L1 expression to those without the PD-L1 expression (Fig. 6A). The expression of PD-1 and IDO was higher in PD-L1–positive tumors. However, they did not meet statistical difference (Fig. 6A). In B7-H3+ tumors, statistically higher PD-1+ and IDO+ cell densities were observed in comparison with B7-H3− tumors (87.6 vs. 15.7 cells/mm2 and 35.2 vs. 9.3 cells/mm2 for PD-1+ and IDO+ cells, respectively; Fig. 6B). PD-L1+ tumor samples were more likely to also have B7-H3 expression, with 76% of PD-L1+ tumor specimens having B7-H3 expression versus 45% in PD-L1− tumor samples (P = 0.033; Fig. 6C).
Figure 6.
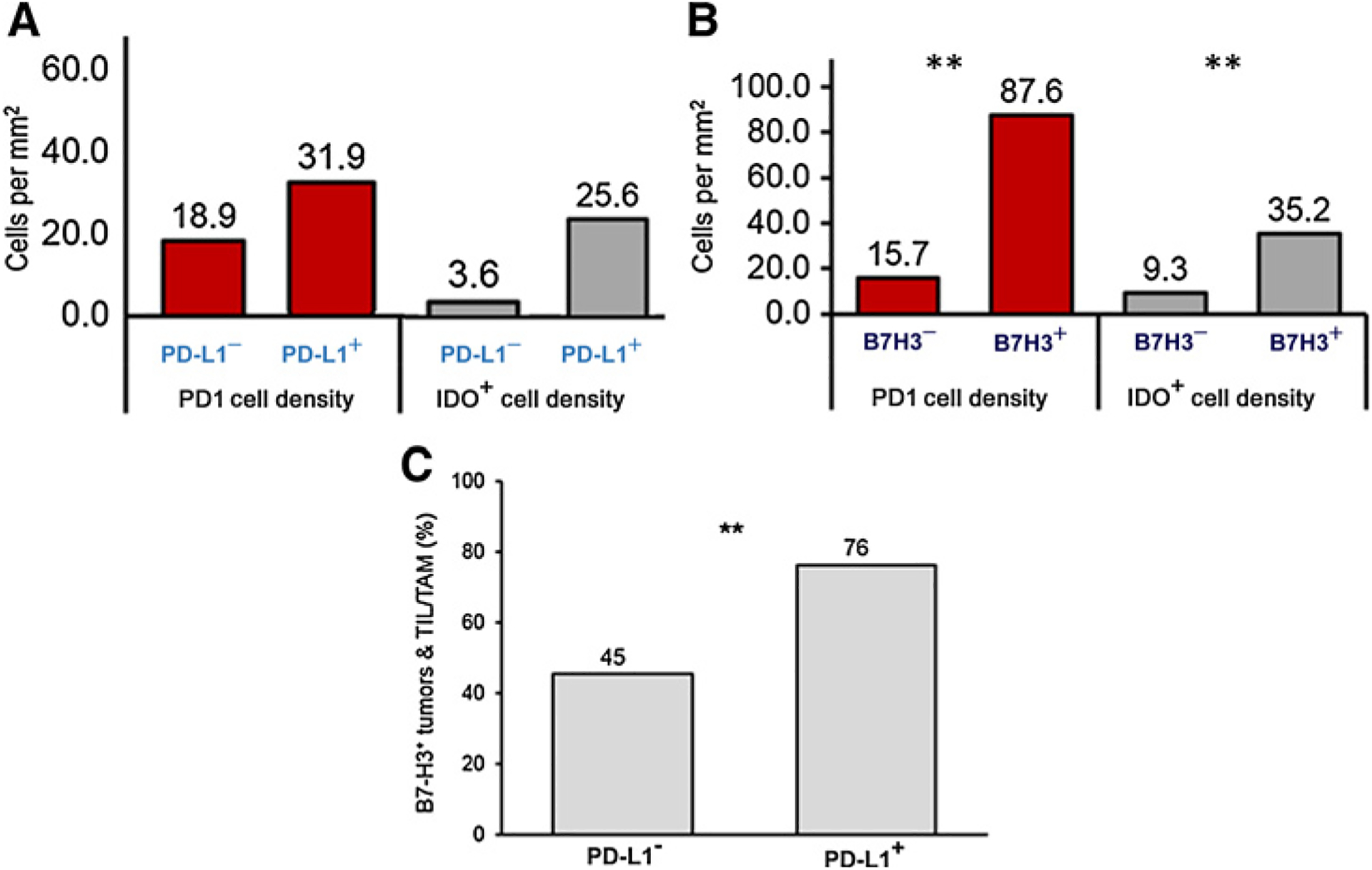
Coexpression of different immune-checkpoint markers. A, PD-1 (red) and IDO (gray) median cell densities in tumors with and without PD-L1 expression and B, in tumors with and without B7H3 expression. 32 tumor regions were analyzed. Mean cell densities are indicated above each bar. Quantile regression was used with a P value of ≤ 0.05 to define statistical significance. C, Percentage of B7-H3 expression in tumors and TILs/TAMs with PD-L1 expression was compared with specimens without PD-L1 expression. Percentage is indicated above each bar. Pearson χ2 test was used with a P value of ≤ 0.05 for significance, as noted by **.
Discussion
In this study, we evaluated the expression of multiple immune-checkpoint molecules in FLC tumor samples, which included PD-1, PD-L1, B7-H3, IDO, and LAG3. We found that FLC is a T-cell inflamed tumor, specifically at the interface region, and has expression of PD-1, PD-L1, B7-H3, and IDO on both tumor cells and immune cells in response to cytotoxic immune responses (i.e., by CD8+ T cells), suggesting an adaptive immune resistance. This upregulation of immune checkpoints in FLC is distinctive from other pediatric tumors, which usually present with an inherently low mutation burden with negative to poor PD-L1 expression (29–32).
Almost two thirds of the FLC specimens demonstrated membranous PD-L1 expression on tumor cells and PD-L1 expression on TILs and TAMs. PD-L1+ TILs/TAMs were also more robust and concentrated compared with PD-L1− tumors, with close to half of all samples encompassing ≥25% PD-L1+ TILs and TAMs. Considering that other cancers with PD-L1+ tumor-infiltrating immune cells show high response rates to immunotherapy (11, 12), this may suggest an important prognostic feature of FLC. We found a subset of PD-L1+ samples with CD11c and CD68 expression, suggesting the possible role of PD-L1+ APCs in antitumor immunity and that there are multiple cell types expressing PD-L1 within the TME that create a PD-L1 immunoprotective barrier.
We also found a high proportion of the samples with B7-H3 expression in both the tumor and adjacent nontumor regions. Although it is still unclear whether tumor-associated B7-H3 is considered to directly augment CD8+ T-cell effector function or play inhibitory roles in T-cell activation (25), we also found that the samples with B7-H3+ tumor cells had much higher CD8+ T-cell density compared with those without B7-H3 expression. This is in contrast to our previous study where B7-H3 expression in other solid tumors did not show any association with an adaptive-type response (33). A similar correlation was seen in samples with PD-L1+ tumor cells and CD8+ T-cell density. We also found that B7-H3+ tumors are more likely to present with PD-L1/PD-1 expression, as well as IDO expression and immune cell infiltration.
Lastly, we also demonstrated the importance of the peritumoral interface area in IDO expression, as this area contained the highest concentration of IDO+ tumor and immune cells, which correlated with CD8+ T-cell density. Similar to the PD-L1 and PD-1 axis (34), IDO is also driven by IFNg secreted by tumor antigen–specific T cells. It is possible that IDO+ tumor cells at the interface between the tumor periphery and the surrounding tissue may form an IDO immune-protective barrier around the tumor to counteract the activated tumor-specific T-cell response. Further investigation is needed to evaluate the immune signatures and identify the critical players that recruit or drive IDO expression at this region of the TME.
The current study had several limitations. Specifically, this was a single-center study that included a small sample size of patients with limited follow-up data. As such, we were unable to correlate the immune-checkpoint expression with survival and, therefore, cannot comment on the prognostic value of these markers. However, considering the rarity of FLC, the current study represents a relatively large cohort of FLC patients. Also, we did not have the multiplex capability in this study to perform multiple color immunofluorescence for coexpression analysis for the immune-specific markers.
There is a long-standing, unmet need for effective systemic therapy in patients with FLC. Data from the study provide further basis for targeting these different immune-checkpoint axes in FLC, including combinational treatments. High frequency of PD-L1, IDO, and B7-H3 expression on TILs and TAMs may play an important role in immunosuppression in the TME, which could be targeted for inhibition. Combination of PD-L1 inhibition with either IDO inhibitors or B7-H3–targeted therapy is already in early clinical trials for other solid cancers (35–37). Although it will be important to interrogate larger tumor cohorts to confirm our findings, the upregulation of multiple immune checkpoints in our cohort provides a basis for investigating immune-checkpoint blockade as a novel therapeutic strategy for patients with FLC.
Supplementary Material
Acknowledgments
This study was supported by Cancer Research Institute/Fibrolamellar Cancer Foundation Grant.
The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Disclosure of Potential Conflicts of Interest
N.J. Llosa reports receiving a commercial research grant from and is a consultant/advisory board member for Bristol-Myers Squibb. M. Yarchoan reports receiving other commercial research support from Bristol-Myers Squibb, Merck, and Exelixis, Inc., and is a consultant/advisory board member for Exelixis, Inc. E.M. Jaffee reports receiving commercial research grants from AduroBiotech, Bristol-Myers Squibb, Amgen, and Corvus; has ownership interest in Aduro Biotech; and is a consultant/advisory board member for Genocea, CSTONE, DragonFly, and Adaptive Biotech. J.M. Taube reports receiving a commercial research grant from Bristol-Myers Squibb and is a consultant/advisory board member for Bristol-Myers Squibb, AstraZeneca, Amgen, and Merck. R. Anders is a consultant/advisory board member for Bristol-Myers Squibb and Merck. No potential conflicts of interest were disclosed by the other authors.
Footnotes
Note: Supplementary data for this article are available at Cancer Immunology Research Online (http://cancerimmunolres.aacrjournals.org/).
References
- 1.Cornella H, Alsinet C, Sayols S, Zhang Z, Hao K, Cabellos L, et al. Unique genomic profile of fibrolamellar hepatocellular carcinoma. Gastroenterology 2015;148:806–18.e10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Graham RP, Yeh MM, Lam-Himlin D, Roberts LR, Terracciano L, Cruise MW, et al. Molecular testing for the clinical diagnosis of fibrolamellar carcinoma. Mod Pathol 2018;31:141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Honeyman JN, Simon EP, Robine N, Chiaroni-Clarke R, Darcy DG, Lim IIP, et al. Detection of a recurrent DNAJB1-PRKACA chimeric transcript in fibrolamellar hepatocellular carcinoma. Science 2014;343:1010–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.El-Serag HB, Davila JA. Is fibrolamellar carcinoma different from hepatocellular carcinoma? A US population-based study. Hepatology 2004;39: 798–803. [DOI] [PubMed] [Google Scholar]
- 5.Darcy DG, Malek MM, Kobos R, Klimstra DS, DeMatteo R, La Quaglia MP. Prognostic factors in fibrolamellar hepatocellular carcinoma in young people. J Pediatr Surg 2015;50:153–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Patt YZ, Hassan MM, Lozano RD, Brown TD, Vauthey JN, Curley SA, et al. Phase II trial of systemic continuous fluorouracil and subcutaneous recombinant interferon Alfa-2b for treatment of hepatocellular carcinoma. J Clin Oncol 2003;21:421–7. [DOI] [PubMed] [Google Scholar]
- 7.El-Khoueiry AB, Sangro B, Yau T, Crocenzi TS, Kudo M, Hsu C, et al. Nivolumab in patients with advanced hepatocellular carcinoma (Check-Mate 040): an open-label, non-comparative, phase 1/2 dose escalation and expansion trial. Lancet 2017;389:2492–502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Topalian SL, Hodi FS, Brahmer JR, Gettinger SN, Smith DC, McDermott DF, et al. Safety, activity, and immune correlates of anti-PD-1 antibody in cancer. N Engl J Med 2012;366:2443–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Taube JM, Klein A, Brahmer JR, Xu H, Pan X, Kim JH, et al. Association of PD-1, PD-1 ligands, and other features of the tumor immune microenvironment with response to anti-PD-1 therapy. Clin Cancer Res 2014;20: 5064–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Herbst RS, Soria J-C, Kowanetz M, Fine GD, Hamid O, Gordon MS, et al. Predictive correlates of response to the anti-PD-L1 antibody MPDL3280A in cancer patients. Nature 2014;515:563–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Powles T, Eder JP, Fine GD, Braiteh FS, Loriot Y, Cruz C, et al. MPDL3280A (anti-PD-L1) treatment leads to clinical activity in metastatic bladder cancer. Nature 2014;515:558–62. [DOI] [PubMed] [Google Scholar]
- 12.Bellmunt J, Mullane SA, Werner L, Fay AP, Callea M, Leow JJ, et al. Association of PD-L1 expression on tumor-infiltrating mononuclear cells and overall survival in patients with urothelial carcinoma. Ann Oncol 2015;26:812–7. [DOI] [PubMed] [Google Scholar]
- 13.Taube JM, Anders RA, Young GD, Xu H, Sharma R, McMiller TL, et al. Colocalization of inflammatory response with B7-h1 expression in human melanocytic lesions supports an adaptive resistance mechanism of immune escape. Sci Transl Med 2012;4:127ra37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Dong H, Strome SE, Salomao DR, Tamura H, Hirano F, Flies DB, et al. Tumor-associated B7-H1 promotes T-cell apoptosis: a potential mechanism of immune evasion. Nat Med 2002;8:793–800. [DOI] [PubMed] [Google Scholar]
- 15.Chen BJ, Chapuy B, Ouyang J, Sun HH, Roemer MGM, Xu ML, et al. PD-L1 expression is characteristic of a subset of Aggressive B-cell lymphomas and virus-associated malignancies. Clin Cancer Res 2013;19:3462–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wu C-P, Jiang J-T, Tan M, Zhu Y-B, Ji M, Xu K-F, et al. Relationship between co-stimulatory molecule B7-H3 expression and gastric carcinoma histology and prognosis. World J Gastroenterol 2006;12:457–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Roth TJ, Sheinin Y, Lohse CM, Kuntz SM, Frigola X, Inman BA, et al. B7-H3 ligand expression by prostate cancer: a novel marker of prognosis and potential target for therapy. Cancer Res 2007;67:7893–900. [DOI] [PubMed] [Google Scholar]
- 18.Maeda N, Yoshimura K, Yamamoto S, Kuramasu A, Inoue M, Suzuki N, et al. Expression of B7-H3, a potential factor of tumor immune evasion in combination with the number of regulatory T cells, affects against recurrence-free survival in breast cancer patients. Ann Surg Oncol 2014;21: S546–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bilzer M, Roggel F, Gerbes AL. Role of Kupffer cells in host defense and liver disease. Liver Int 2006;26:1175–86. [DOI] [PubMed] [Google Scholar]
- 20.Wisse E, Braet F, Luo D, De Zanger R, Jans D, Crabbé E, et al. Structure and function of sinusoidal lining cells in the liver. Toxicol Pathol 1996;24: 100–11. [DOI] [PubMed] [Google Scholar]
- 21.Munn DH, Zhou M, Attwood JT, Bondarev I, Conway SJ, Marshall B, et al. Prevention of allogeneic fetal rejection by tryptophan catabolism. Science 1998;281:1191–3. [DOI] [PubMed] [Google Scholar]
- 22.Uyttenhove C, Pilotte L, Théate I, Stroobant V, Colau D, Parmentier N, et al. Evidence for a tumoral immune resistance mechanism based on tryptophan degradation by indoleamine 2,3-dioxygenase. Nat Med 2003;9: 1269–74. [DOI] [PubMed] [Google Scholar]
- 23.Weinlich G, Murr C, Richardsen L, Winkler C, Fuchs D. Decreased serum tryptophan concentration predicts poor prognosis in malignant melanoma patients. Dermatol Basel Switz 2007;214:8–14. [DOI] [PubMed] [Google Scholar]
- 24.Yarchoan M, Xing D, Luan L, Xu H, Sharma RB, Popovic A, et al. Characterization of the Immune Microenvironment in Hepatocellular Carcinoma. Clin Cancer Res Off J Am Assoc Cancer Res 2017;23:7333–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Chapoval AI, Ni J, Lau JS, Wilcox RA, Flies DB, Liu D, et al. B7-H3: a costimulatory molecule for T cell activation and IFN-gamma production. Nat Immunol 2001;2:269–74. [DOI] [PubMed] [Google Scholar]
- 26.Thompson ED, Zahurak M, Murphy A, Cornish T, Cuka N, Abdelfatah E, et al. Patterns of PD-L1 expression and CD8 T cell infiltration in gastric adenocarcinomas and associated immune stroma. Gut 2017; 66:794–801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Curiel TJ, Wei S, Dong H, Alvarez X, Cheng P, Mottram P, et al. Blockade of B7-H1 improves myeloid dendritic cell-mediated antitumor immunity. Nat Med 2003;9:562–7. [DOI] [PubMed] [Google Scholar]
- 28.Lin H, Wei S, Hurt EM, Green MD, Zhao L, Vatan L, et al. Host expression of PD-L1 determines efficacy of PD-L1 pathway blockade-mediated tumor regression. J Clin Invest 2018;128:805–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Chalmers ZR, Connelly CF, Fabrizio D, Gay L, Ali SM, Ennis R, et al. Analysis of 100,000 human cancer genomes reveals the landscape of tumor mutational burden. Genome Med 2017;9:34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Geoerger B, Kang HJ, Yalon-Oren M, Marshall LV, Vezina C, Pappo AS, et al. Phase 1/2 KEYNOTE-051 study of pembrolizumab (pembro) in pediatric patients (pts) with advanced melanoma or a PD-L1+ advanced, relapsed, or refractory solid tumor or lymphoma. J Clin Oncol 2017;35: 10525–10525. [DOI] [PubMed] [Google Scholar]
- 31.Davis KL, Fox E, Reid JM, Liu X, Minard CG, Weigel B, et al. ADVL1412: Initial results of a phase I/II study of nivolumab and ipilimumab in pediatric patients with relapsed/refractory solid tumors—A COG study. J Clin Oncol 2017;35:10526. [Google Scholar]
- 32.Majzner RG, Simon JS, Grosso JF, Martinez D, Pawel BR, Santi M, et al. Assessment of programmed death-ligand 1 expression and tumor-associated immune cells in pediatric cancer tissues. Cancer 2017;123: 3807–15. [DOI] [PubMed] [Google Scholar]
- 33.Danilova L, Wang H, Sunshine J, Kaunitz GJ, Cottrell TR, Xu H, et al. Association of PD-1/PD-L axis expression with cytolytic activity, mutational load, and prognosis in melanoma and other solid tumors. Proc Natl Acad Sci 2016;113:E7769–77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Topalian SL, Taube JM, Anders RA, Pardoll DM. Mechanism-driven bio-markers to guide immune checkpoint blockade in cancer therapy. Nat Rev Cancer 2016;16:275–87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Study of IDO Inhibitor in Combination With Checkpoint Inhibitors for Adult Patients with Metastatic Melanoma —Full Text View—ClinicalTrials.gov [Internet]. [cited 2018 Feb 12]. Available from: https://clinicaltrials.gov/ct2/show/NCT02073123.
- 36.Rizvi NA, Loo D, Baughman JE, Yun S, Chen F, Moore PA, et al. A phase 1 study of enoblituzumab in combination with pembrolizumab in patients with advanced B7-H3-expressing cancers. J Clin Oncol 2016; 34:TPS3104. [Google Scholar]
- 37.Enoblituzumab (MGA271) in Children With B7-H3-expressing Solid Tumors - Full Text View - ClinicalTrials.gov [Internet]. [cited 2018 Feb 12]. Available from: https://clinicaltrials.gov/ct2/show/NCT02982941.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.