

Abstract
Candida albicans is an endogenous opportunistic pathogenic fungus that is harmless when the host system remains stable. However, C. albicans could seriously threaten human life and health when the body’s immune function declines or the normal flora is out of balance. Due to the increasing resistance of candidiasis to existing drugs, it is important to find new strategies to help treat this type of systemic fungal disease. Biological control is considered as a promising strategy which is more friendly and safer. In this review, we compare the bacteriostatic behavior of different antagonistic microorganisms (bacteria and fungi) against C. albicans. In addition, natural products with unique structures have attracted researchers’ attention. Therefore, the bioactive nature products produced by different microorganisms and their possible inhibitory mechanisms are also reviewed. The application of biological control strategies and the discovery of new compounds with antifungal activity will reduce the resistance of C. albicans, thereby promoting the development of novel diverse antifungal drugs.
Keywords: Candida albicans, antagonistic microorganisms, biocontrol strategy, bioactive compounds
1. Introduction
Fungal infection is a common global problem affecting humans and its incidence is on the rise [1]. Among them, Candida has been a life-threatening pathogen for a long time, accounting for almost 80% of fungal infections. Recently C. albicans infection causes more than 400,000 cases of blood infection each year, with a mortality rate of about 42% [2,3,4]. C. albicans, a small number in the normal body, is a part of healthy flora. It can exist in the oral cavity, intestinal tract, upper respiratory tract, and other parts. When growing in the state of unicellular yeast cells, it does not cause disease. However, when the normal flora interacts with each other out of balance or the body’s immune function and defenses decline, C. albicans proliferates and grows into hyphae, invading cells and causing disease. It has been a major cause of morbidity and mortality in immunocompromised populations [5].
In host, the pathogenicity of C. albicans is caused by the decline of immune function, the change of conventional flora and the destruction of the epithelial protective barrier. During infection, the formation of C. albicans biofilm and the morphological switch from yeast-like to hyphal-like are considered to be two significant pathogenic characteristics of C. albicans. First of all, its morphological plasticity is crucial to the pathogenicity of fungi, as the hyphal form has a key role in the infection process [6,7,8]. In addition, the pathogenicity of Candida is greatly enhanced by the formation of biofilms [9]. Biofilms are microbial communities that irreversibly attach to surfaces. Biofilms behave very differently from planktonic cells, and once formed, they can increase resistance to existing antibiotics and immune responses [10]. Therefore, inhibition of hyphal development and inhibition of biofilm formation are considered to be an effective strategy against C. albicans infection.
Currently, there are very few drugs for the treatment and prevention of Candidiasis in clinic. The polyene antibiotic is the earliest specific drug isolated from Streptomyces nodosus in the 1950s to treat yeast infection. Since then, many antifungal agents have been developed [11,12]. There are four types of antifungal agents for C. albicans infection [13]. The most commonly used antifungal drugs and the mechanism of action include:(1) The widest range and most effective is polyene (Amphotericin B), which can kill most fungi. Polyenes bind to ergosterol in fungal cell membranes, creating stomata and causing cell death [14,15]. (2) Triazole antifungal drugs (fluconazole, voriconazole and itraconazole). Azoles can inhibit lanosterol 14α demethylase, which is an important enzyme in ergosterol biosynthesis [16,17,18,19]. (3) 5-fluorocytosine, it inhibits fungal DNA synthesis by inhibiting thymidylate synthetase [12,20]. (4) There are also some echinocandin antifungal drugs (anidulafungin, micafungin, and caspofungin) [21,22,23,24,25,26,27]. The mechanisms of these bioactive compounds against C. albicans are mainly related to inhibition of biofilm formation, inhibition of virulence factors and destruction of cell wall integrity. With the increasing drug resistance of C. albicans, it is compelling to find new antifungal methods and reagents to solve this complex medical problem. Biological control is considered to be a more effective and safe strategy [1,28,29].
Novel natural compounds produced by microorganisms, due to their complex structures, may exhibit novel antibacterial mechanisms and different modes of action. Moreover, they were considered as candidates to reduce drug resistance. People have been trying to find unique antifungal drugs from nature, which has led to important advances in the development of new antifungal drugs.
In recent years, there have been some reviews on natural products that could inhibit C. albicans [2,30,31,32,33,34,35,36,37]. In this paper, we have reviewed the antagonistic microorganisms against C. albicans considered in recent years and have also reviewed the active natural products produced by microorganisms that inhibit C. albicans. Researchers focus on the study of antagonistic microorganisms in order to use probiotics to inhibit C. albicans. Through the review of secondary metabolites, it can provide a reference for clinical drug development.
2. Antagonistic Microbes against C. albicans
Traditional azoles and their derivatives have poor effect on preventing recurrence of pathogenic fungus. In some patients, fluconazole can cause some side effects such as headache, discomfort, dizziness, gastrointestinal and, rash [38]. Bacteria, yeast, and fungus all can develop resistance to antibiotics and bactericidal chemicals [39]. Biological control of microbial infections is an alternative approach that utilizes antagonistic microorganisms to prevent the growth and infection of harmful microorganisms. Diverse microorganisms, including fungi (such as non-toxic Aspergillus, Trichoderma, Penicillium), yeast strains, and bacteria, have been studied as potential antagonistic organisms for the control of C. albicans. In this review, the microorganisms that inhibit C. albicans and their secondary metabolites are introduced from the perspective of antagonistic microorganisms. The microorganisms that have potential antagonism against C. albicans are listed in Table 1. The main species and inhibition activities of these antagonistic strains are also discussed. We have reviewed the antagonistic microorganisms against C. albicans in recent years with the aim to develop a new natural material, using beneficial bacteria or fungus, that would be useful for inhibiting the growth of pathogenic C. albicans in the human body.
Table 1.
Antagonistic Microbes against C. albicans.
| Antagonists | Species | Activity | References |
|---|---|---|---|
| Bacillus spp. | B. sphaericus A16, B. circulans M142, B. brevis M166, B. brevis T122 | Strains showed extensive inhibition against C. albicans. |
[40] |
| B. subtilis spizizenii DK1-SA11 | Cell-free supernatant had significant inhibitory activity against C. albicans. | [41] | |
| B. velezensis DTU001 | Significantly inhibited the proliferation of C. albicans, and the inhibition ability of the strain was better than that of a single lipopeptide. | [42] | |
| Bifidobacterium |
B. amyloliquefaciens SYBC H47 |
Cell-free supernatant and Cell suspension had obvious inhibition against C. albicans. | [43] |
| B. velezensis 1B-23 | Inhibited C. albicans growth in vitro. | [44] | |
| B. longum BB536 | The supernatant of fermented broccoli could inhibit the growth of C. albicans in vitro. | [45,46] | |
| Lactobacillus genus | L. johnsonii MT4 | Inhibited planktonic growth and biofilm formation of C. albicans | [47] |
| Lactobacillus | Regulated growth and virulence of C. albicans through niche competition. | [48] | |
| Yeast | Metschnikowia pulcherrima | Strong antagonistic activity against C. albicans. | [49] |
| Other strains | Enterococcus | Regulated growth and virulence of C. albicans through niche competition. | [48] |
| Pseudomonas fluorescens | The strain showed extensive inhibition against C. albicans. |
[40] | |
| Salivarius MG242 | The strain had significant inhibitory effect on C. albicans. |
[50] |
As shown in Figure 1, the article reporting Bacillus spp. antagonists were dominant (40%) compared with the article reporting antagonistic Bifidobacterium (20%), antagonistic Lactobacillus genus (13.33%), antagonistic yeast (6.67%) and other antagonistic strains (20%).
Figure 1.

Percentages of different antagonistic microbes of C. albicans.
2.1. Antagonistic Effect of Bacillus spp. against C. albicans
Some beneficial bacteria or fungus are widely used in biocontrol. In particular, it is well known that Bacillus spp. is an excellent source of antifungal drugs, thus Bacillus spp. is widely used as a biological control agent [51,52,53]. Bacillus species are Gram-positive bacteria that can survive in different environments. They could form endospores and produce a large number of metabolites [53].
Researchers isolated four strains of Bacillus A16 (B. sphaericus), M142 (B. circulans), M166 (B. brevis) and T122 (B. brevis) from soil samples. These Bacillus showed extensive inhibitory activity against C. albicans [40]. Among them, B. brevis M166 showed antifungal activity against all tested microorganisms (Sclerotium rolfsii, Rhizoctonia solani, Fusarium oxysporum, Staphylococcus aureus and C. albicans), with a relatively wide antimicrobial spectrum. B. circulans M142 had strong antibacterial activity against C. albicans and S. aureus, while B. brevis T122 only had antibacterial activity against C. albicans. To our knowledge, no specific compounds inhibiting C. albicans had been identified.
In addition to the antifungal activity of Bacillus spp. from soil samples, Bacillus spp. from marine samples was also found to have inhibitory activity against C. albicans. B. subtilis spizizenii DK1-SA11 was isolated from Bay of Yellow Sea in China [41]. The cell-free supernatant had significant inhibitory activity against C. albicans. The inhibitory active ingredient had not been identified but was stable in nature, while the enzymatic hydrolysis of lipase, trypsin and papain made it lose activity. Antimicrobial activity tests against pathogens indicated that this strain could be used as a source of antibiotics, synbiotics, and probiotics.
B. velezensis was widespread in the environments and produced abundant lipopeptides with good bacteriostatic effect. Some researchers have studied on the inhibitory spectrum of B. velezensis DTU001 against 20 different species of human and/or plant pathogenic fungi [42]. The results showed that B. velezensis DTU001 was superior to a single lipopeptide (fengycin and iturin) in inhibiting the selected fungi. Co-culture of B. velezensis DTU001 and C. albicans significantly inhibited C. albicans proliferation, which further supported the biological control properties of B. velezensis DTU001.
B. amyloliquefaciens SYBC H47 was isolated from honey [43]. The cultured cell-free supernatant had significant inhibitory activity against C. albicans. The main antibacterial substances were surfactin, fengycin and bacillomycin. Three compounds had an inhibitory effect on spore germination of Botryosphaeria dothidea. However, compounds that inhibit C. albicans had not been identified.
Bacillus velezensis 1B-23 had inhibitory effect on the growth of C. albicans in vitro. It had a certain application prospect as a biological agent for biological control of fungal pathogens [44].
2.2. Antagonistic Effect of Bifidobacterium spp. against C. albicans
Bacillus spp. has been used clinically because of its bacteriostatic activity. Another probiotic, Bifidobacterium, can also be used to prevent and treat intestinal flora disorders in clinic. Bifidobacterium is a vital member of the normal human gut microbiota. Some strains of Bifidobacterium can be used as probiotics in food, medicine and feed [54,55]. Bifidobacteria could produce acetic acid and/or lactic acid during metabolism. Moreover, the action of lactic acid would reduce intestinal pH. Thereby, Bifidobacterium could inhibit the proliferation of pathogenic microorganisms [56,57].
Bifidobacterium longum BB536 which was isolated from the feces of healthy infants had been commercially used in various food applications and was considered safe [45,46,58]. The researchers fermented broccoli using B. longum. The supernatant could inhibit the growth of C. albicans and some other pathogenic bacteria in vitro. Researchers used beneficial bacteria such as bifidobacteria and used broccoli as a substrate for the growth of beneficial bacteria to develop substances. Maybe, we can use beneficial microorganisms and their secondary metabolites to develop products that inhibit the growth of pathogenic microorganisms. For example, as a daily oral care preparation, it can prevent the growth of C. albicans in human oral cavity [59].
2.3. Antagonistic Effect of Lactobacillus spp. against C. albicans
Lactobacillus johnsonii is a probiotic with wide antimicrobial characteristics and can be used as an antiallergic drug. Recent studies have shown that L. johnsonii also has inhibitory effects on C. albicans. L. johnsonii MT4 was isolated from the oral cavity of healthy mice. The strain affected the C. albicans growth in both biofilm and planktonic conditions. L. johnsonii MT4 showed an antagonistic effect on C. albicans, thus inhibiting the biofilm formation of C. albicans and planktonic growth. The study on the strain genome had shown that it produced metabolites with anti-C. albicans activity, but no active substances against fungi have been reported so far. The antibacterial mechanism needed to be further explored [47].
In addition to producing secondary metabolites that antagonize C. albicans, the competition for ecological niches of different strains during the growth process would also cause antagonism among strains, such as C. albicans and lactic acid bacteria in the gastrointestinal (GI) tract [48,60]. Non-pathogenic colonization of the human GI tract by C. albicans was common. C. albicans could regulate bacterial community in mice treated with broad-spectrum antibiotics. One of the most striking features was the significant change in the lactic acid bacteria (LAB) levels. C. albicans and Lactobacillus species shared a metabolic niche throughout the GI tract. LAB could antagonize Enterococcus and C. albicans in the GI tract. C. albicans and Lactobacillus could mutually regulate each other’s growth and virulence in the GI tract [48].
2.4. Antagonistic Effect of Yeast against C. albicans
In addition to the bacteria mentioned above, yeast can also be used for biological control. Metschnikowia could accumulate pigments in cells and growth media. It was a highly effective biocontrol yeast. Antagonism of M. pulcherrima against phytopathogens had been demonstrated [49]. The researcher investigated three new strains of Metschnikowia which were isolated from grapes. The strain had strong antagonistic activity against C. albicans. The three strains produced the same amount of nevus pigments, but there were significant differences in antifungal activities against different microorganisms [61,62].
2.5. Antagonistic Effect of Other Strains against C. albicans
Salivarius MG242 isolated from human vagina presented a potential application in the biological control of C. albicans. MG242 had an obvious inhibitory impact on C. albicans, and the strain had the possibility to be developed into a probiotic product for the treatment of C. albicans. In order to develop stable living cell products, it was necessary to maintain anti-Candida activity and preserve cell viability during lyophilization. Lower storage temperature extended shelf life to 8.31 months [50]. Strains of K124 (P. fluorescens) was also isolated from soil samples, e.g., B. sphaericus A16, B. circulans M142, B. brevis M166 and B. brevis T122. P. fluorescens K124 showed extensive inhibitory activity against C. albicans [40]. P. fluorescens K124 only had antifungal activity against C. albicans. At present, no inhibitory compounds produced by the strain have been identified.
2.6. A Conclusion of Antagonistic Microbes
In conclusion, Bacillus, Bifidobacterium, Lactobacillus, and yeast strains can antagonize the growth of C. albicans. In particular, many strains of Bacillus have obvious advantages to exert antagonistic strains. Most of the strains exert antagonistic effects by producing active compounds. Moreover, some inhibit the growth of C. albicans through niche competition. We should intensify research on strains with inhibitory activity, especially probiotics. Research on different strains, especially probiotics, with antifungal activity is helpful to develop the agent for inhibiting C. albicans. Since the effective components of some strains against C. albicans are not clear, the compounds with obvious inhibitory activity should be further analyzed.
3. Inhibitory Nature Metabolites Produced by Diverse Antagonists
Secondary metabolites derived from many plants and microorganisms are valuable natural compounds. Many natural products have significant biological activities, such as anti-tumor activity, antibacterial activity [63,64,65]. The antagonistic effect of the strain is mainly due to the production of natural secondary metabolites, such as antibiotics and antimicrobial peptides [66,67,68]. The antifungal compounds reviewed in this paper are secondary metabolites derived from microorganisms for biological control of C. albicans and have strong inhibition against C. albicans. Table 2 lists the various antagonistic microbial strains, the characteristics of the active compounds produced, and their inhibition mechanism against C. albicans. Table 3 lists the structure and the activity of these inhibitory compounds.
Table 2.
Inhibitory nature metabolites produced by antagonists against C. albicans.
| Sources | Inhibitory Compounds |
Main Characteristics of the Compounds |
Other Inhibitory Actions | References |
|---|---|---|---|---|
| Bacteria | ||||
| Bacillus subtilis | 5HM2F | Inhibit morphological transition | Reduced levels of secreted virulence factors and ergosterol to reduce the main sources of biofilms. | [69] |
|
Pantoea agglomerans C9-1 |
2-amino-3-(oxane-2,3-dicarboxamido) propanoyl-valine | Inhibit growth | None | [70] |
|
Tenacibaculum discolor sv11 |
Dipyrrolepyridines A and B |
Inhibit growth | [71] | |
| Yeast | ||||
|
Saccharomyces
boulardii |
Capric acid | Inhibit hyphal formation, adhesion and biofilm development | Transcriptional levels of HWP1, INO1 and CSH1 genes were decreased. | [72] |
| Eendophytic fungi | ||||
| Biatriospora sp. | Biatriosporin D | Inhibit adhesion, biofilm formation and hyphal morphogenesis | Regulated Ras1-CAMP-Efg1 pathway, disrupted morphological transition and attenuated virulence | [73] |
| Drechmeria sp. | Drechmerin B | Inhibit growth | None | [74] |
| Phoma sp. SYSU-SK-7 | Colletotric A | Inhibit growth | None | [75] |
|
Stachybotrys
chartarum |
Atranone Q | Inhibit growth | None | [76] |
| Xylaria sp. YM 311647 | Sesquiterpenes and Isomatanic diterpenes | Inhibit growth | None | [77] |
| Marine fungi | ||||
| Aspergillus isolates from Waikiki Beach | Waikialoid A and Waikialide A | Inhibit biofilm formation | None | [78] |
|
Penicillium meleagrinum var. viridiflavum |
PF1163A and B | Inhibit growth | None | [79] |
| Penicillium minioluteum ZZ1657 | Purpurides E and F | Inhibit growth | None | [80] |
| Marine actinomycetes | ||||
| Actinoalloteichus cyanogriseus WH1-2216-6 | Caerulomycin A and C | Inhibit growth | None | [81] |
| Streptomyces sp. | Bahamaolides A | Inhibit isocitrate lyase | None | [82] |
| Streptomyces sp. ZZ741 | Streptoglutarimides A-J and Streptovitacin A | Inhibit growth | None | [83] |
| Lichen | ||||
| lichens | Usnic acid | Reduce the thickness of mature biofilms and Inhibit biofilm adhesion. | [84] | |
| lichens | Retigeric acid B | Inhibit hyphal formation | RAB regulated the Ras1-cAMP-Efg1 pathway and inhibited hyphal formation |
[85] |
| Lichens with Talaromyces funiculosu | Funiculosone | Inhibit growth | None | [86] |
| Other strains | ||||
| Acremonium sp. PSU-MA70 | 8-Deoxytrichocin and trichodermol | Inhibit growth | None | [87] |
| Aspergillus micronesiensis | Cyschalasins A and B | Inhibit growth | None | [88] |
| Curvularia hawaiiensis TA26-15 | Moriniafungins B-G | Inhibit growth | None | [89] |
| Fusarium and Gibberella species | Zearalenone | Inhibit biofilm formation of and hyphal morphogenesis | None | [90,91,92] |
| Fusarium spp. | Deoxynivalenol | Inhibit biofilm formation and reduce metabolic activity | DON and its derivatives interplayed with lanosterol 14a-demethylase | [93] |
| Penicillium fuscum and Penicillium camembertii/clavigerum | Berkleyolactone A | Inhibit growth | A new mode of action that had not been resolved | [94] |
| Ustilago maydis | Ustilagic acid B and C | Inhibit growth | None | [95] |
5HM2F: 5-hydroxymethyl-2-furaldehyde.
Table 3.
The structures and activity of compounds against C. albicans.
| Inhibitory Compounds | Compound Structure | Activity | References |
|---|---|---|---|
| Terpenoids | |||
| Isomatanic diterpenes |
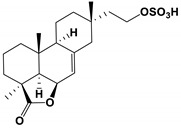
|
The MIC value was 16 μg/mL | [77] |
| Purpurides E and F |
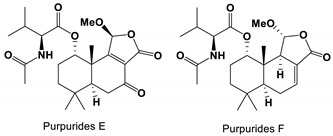
|
The MIC values were 12 and 6 μg/mL, respectively. | [80] |
| Usnic acid |
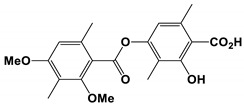
|
The MBIC value was 100 μg/mL. | [84] |
| Moriniafungins E |
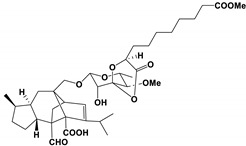
|
The MIC value was 2.9 μM. | [89] |
| Macrolides | |||
| PF1163 A and B |
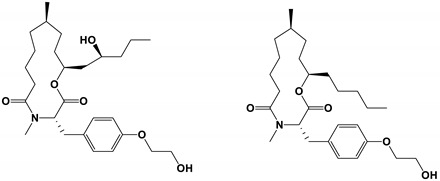
|
The inhibitory concentrations were 1 and 2 μg/mL, respectively. | [79] |
| Bahamaolides A |
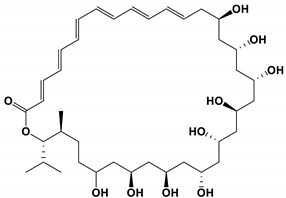
|
The MIC value was 12.5 μg/mL. | [82] |
| Berkleyolactone A |
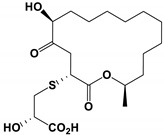
|
The MIC value was 1–2 μg/mL. | [94] |
| Organic acids | |||
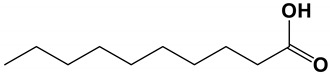
| Capric acid |
|
The inhibitory concentration was 45.3 μg/mL. | [72] |
| Retigeric acid B |
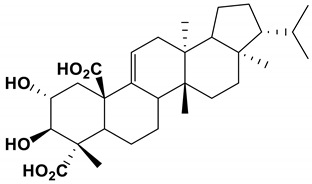
|
The MIC80 value was 8 μg/mL. | [85] |
| Ustilagic acid B and C |
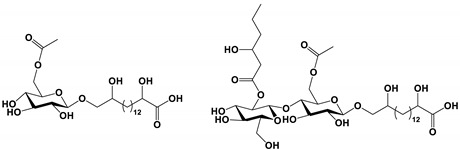
|
The MIC values were 50 and 100 μg/mL, respectively. | [95] |
| Alkaloids | |||
| Ketones | |||
| Colletotric A |
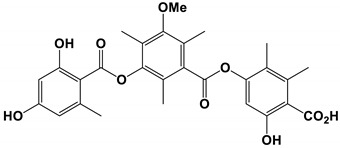
|
The MIC value was 3.27 μg/mL. | [75] |
| Atranone Q |
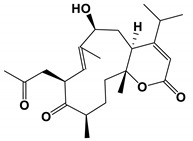
|
The MIC value was 8 μg/mL | [76] |
| Waikialoid A and Waikialide A |
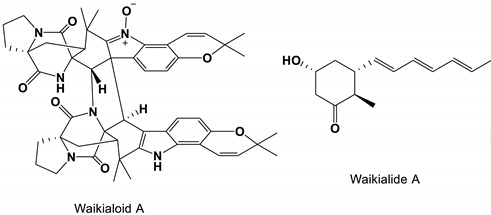
|
The IC50 values were 1.4 and 32.4 μM, respectively. | [78] |
| Caerulomycin A and C |
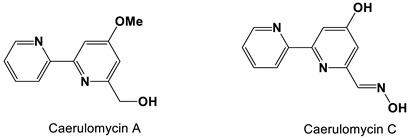
|
The MIC values were 21.8 and 19.3 μM, respectively. | [81] |
| Cyschalasins A and B |
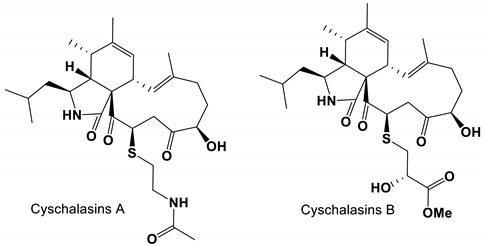
|
The MIC50 values were 43.3 ± 1.5 and 94.7 ± 1.3 μg/mL, respectively. | [88] |
| Zearalenone |
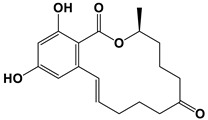
|
The inhibitory concentration was 100 μg/mL | [90,91,92] |
| Alcohols | |||
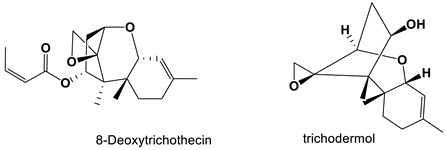
| 8-Deoxytrichothecin and trichodermol |
|
The MIC values were 16 and 64 μg/mL, respectively. | [87] |
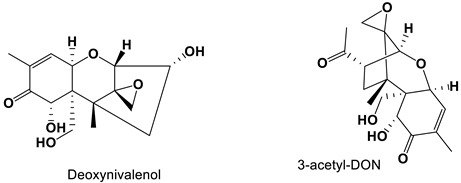
| Deoxynivalenol and 3-acetyl-DON |
|
All inhibitory concentrations were 50 μg/mL. | [93] |
| Other structural compounds | |||
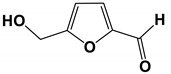
| 5HM2F |
|
The MBIC value was 400 μg/mL. | [69] |
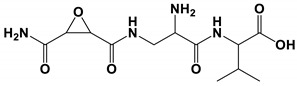
| 2-amino-3-(oxane-2,3-dicarboxamido) propanoyl-valine |
|
The inhibitory concentration was 1.5 μg/mL. | [70] |
| Dipyrrolepyridines A and B |
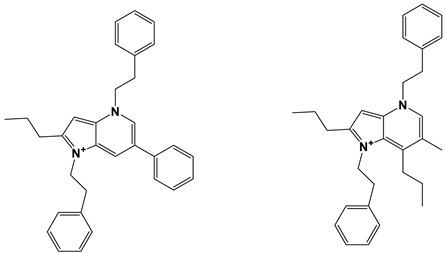
|
Certain antibacterial activity. | [71] |
| Biatriosporin D |
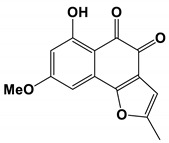
|
The inhibitory concentration was 2 μg/mL | [73] |
| Drechmerin B |
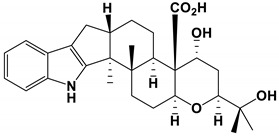
|
The MIC value was 12.5 μg/mL. | [74] |
| Streptoglutarimides D |
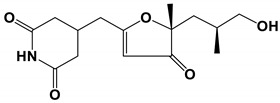
|
The MIC value was 4 μg/mL. | [83] |
| Funiculosone |
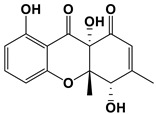
|
The IC50 value was 35 μg/mL. | [86] |
BEC80: 80% of biofilm-eradicating concentration; MBIC: maximum biofilm inhibitory concentration; 5HM2F: 5-hydroxymethyl-2-furaldehyde.
3.1. Nature Products Produced by Bacteria
Bacillus produces diverse active compounds, such as proteases, amylases, surfactants, and antibiotics [66,96,97,98,99]. Due to the high yield of antifungal active substances and the advantage of releasing peptides directly into the extracellular, Bacillus subtilis is a potential strain for the production of antifungal compounds [100,101,102]. The B.subtilis isolated from marine had antifungal membrane effect on C. albicans. It was found that 5-hydroxymethyl-2-furaldehyde (5HM2F) was one of the main components that inhibited C. albicans in the fermentation broth [69]. 5HM2F effectively disrupted the hyphal-like morphological transition of C. albicans and prevented the initial adhesion process. Further studies showed that 5HM2F reduced the main source of biofilms by reducing the levels of secreted virulence factors and ergosterol. In addition, the combination of 5HM2F with azole antifungal drugs effectively enhanced the anti-C. albicans activity of the tested drugs. Transcriptional level studies showed that 5HM2F increased the sensitivity of C. albicans to antifungal drugs by negatively regulating the expression levels of genes related to drug resistance mechanisms. As an antagonist, 5HM2F effectively inhibited the biofilm formation and reduced the resistance of C. albicans to traditional antifungal drugs.
Pantoea agglomerans are widespread in the environment [103,104]. P. agglamerans strain C9-1 was used as a biological control agent (BlightBan C9-1). A peptide antibiotic was isolated. The compound was 2-amino-3-(oxane-2,3-dicarboxamido)propanoyl-valine. This compound showed effectively inhibition on the growth of C. albicans [70].
Six novel alkaloids containing phenethylamine (PEA) were isolated from the culture medium of Tenacibaculum discolor sv11. Among them, Dipyrrolepyridines A and B had certain inhibitory activity against C. albicans FH2173 [71].
3.2. Nature Products Produced by Yeast
The researchers found that S. boulardii had inhibitory activity on C. albicans. The fermentation broth extracts inhibited hyphae formation, adhesion and biofilm development of C. albicans [72]. Further analysis showed that the fermentation broth contained 2-phenylethanol, capric, caprylic and caproic acid. The fermentation broth and the isolated pure compounds were tested for biological activity against C. albicans. Capric acid inhibited hyphae formation in C. albicans and also reduced adhesion and biofilm formation. However, compared with S. boulardii extract, the inhibitory effect on C. albicans was reduced by three times in the case of capric acid alone, so other compounds were contained to inhibit the adhesion of C. albicans. The transcriptional levels of CSH1, INO1, and HWP1 genes were decreased in C. albicans treated with B. boulardii extract and capric acid.
3.3. Nature Products Produced by Endophytic Fungi
Biatriosporin D (BD), A phenolic compound, was isolated from Biatriospora spp. [73]. The compound inhibited biofilm formation, hyphal morphogenesis and adhesion of C. albicans. Notably, BD efficiently inhibited hyphal formation at doses lower than MIC value. Further studies showed that BD regulated the Ras1-cAMP-Efg1 pathway through reducing the cAMP level. As a prodrug, BD showed potential action against C. albicans. This provided possible application prospects for BD against clinically opportunistic fungi by targeting fungal virulence.
A fungus Drechmeria sp. was isolated from the roots of Panax notoginseng. Four known analogs and seven new indole diterpenoids, drechmerins A-G, were isolated from the fermentation broth. The MIC value of Drechmerin B against C. albicans was 12.5 μg/mL [74].
Five new polyketides and four known analogs were isolated from the Phoma sp. SYSU-SK-7 [75]. Among them, the polyketide colletotric B had strong antifungal activity against C. albicans, and the MIC value of colletotric A was 3.27 μg/mL. The MIC value of 3-hydroxy-5-methoxy-2, 4, 6-trimethylbenzoic acid was 2.62 μg/mL, and the MIC value of orsellinic acid was 2.10 μg/mL.
Three new monomers were isolated from the marine strain Stachybotrys Chartarum. The MIC value of compound Atranone Q was 8 μg/mL [76].
Nine sesquiterpenes and three diterpenes were isolated from the fermentation broth of the Xylaria sp. YM 311647 [77]. The MIC values of nine sesquiterpenes against C. albicans were different, while the activity of diterpenes was higher. One of the sesquiterpenes had the highest antibacterial activity against C. albicans, with an MIC value 16 μg/mL.
3.4. Nature Products Produced by Marine Fungi
One of the prenylated indole alkaloids, waikialoid A, was isolated from a metabolite-rich Aspergillus strain near Waikiki Beach. IC50 value of the natural product was 1.4 μM in inhibiting biofilm formation. Another compound, waikialide A, could inhibit the formation of C. albicans biofilm with a weaker IC50 value of 32.4 μM [78].
Two new 13-membered macrolide compounds and known PF1163A, B, D, H and F were isolated from penicillium strain. All of them had inhibitory activity against C. albicans when used in conjunction with fluconazole [79].
Three drimane sesquiterpene purpurides E-G were isolated from P. minioluteum ZZ1657. Purpurides E exhibited inhibitory activity against C. albicans with MIC values of 6–12 μg/mL, and Purpurides F was 3–6 μg/mL [80].
3.5. Nature Products Produced by Marine Source Actinomycetes
One new phenylpyridinealkaloid, five known analogues and five new bipyridine alkaloids were isolated from Actinoalloteichus cyanogriseus WH1-2216-6. The MICs of caerulomycin A and C against C. albicans were 21.8 and 19.3 μM, respectively [81].
Two new 36-membered macrolides, Bahamaolides A and B, were isolated from sediments of marine actinomycetes (Streptomyces sp.) on the North Cat Reef, Bahamas. Bahamaolides A obviously inhibited isocitrate lyase of C. albicans [82].
Streptovitacin A and new Streptoglutarimides A-J were isolated from marine actinomycetes Streptomyces sp. ZZ741. The MIC values of the obtained compounds against C. albicans were 8–20 μg/mL, and Streptoglutarimides D had a better inhibitory effect with 8 μg/mL [83].
3.6. Nature Products Produced by Lichen
Usnic acid, a secondary metabolite of lichens, effectively inhibited the hyphal switching of C. albicans. Usnic acid significantly reduced the thickness of mature biofilms and prevented the adhesion of biofilms. At the biofilm inhibitory concentration (BIC), the inhibitory effect of usnic acid on C. albicans biofilm could reach 65% [84].
As an inhibitor, Retigeric acid B (RAB) derived from lichen significantly inhibited the hyphae formation of C. albicans [105,106,107]. RAB prolonged the survival time of nematodes infected by C. albicans. RAB regulated the Ras1-CAMP-Efg1 pathway by reducing cAMP levels and inhibitd hyphal formation. By inhibiting the interruption of yeast-hyphal morphological transition and weakening the virulence of C. albicans, it provided a potential application for the treatment of C. albicans infection [85].
Funiculosone, a substituted dihydroxanthene-1, 9-dione, was isolated from the lichens of the Trichosporaceae fungus T. funiculosus. The IC50 value of T. funiculosus was 35 μg/mL [86].
3.7. Nature Products Produced by Other Fungal Sources
8-deoxytrichothecin and trichodermol, isolated from the Acremonium sp. PSU-MA70, exhibited moderate antifungal activity against C. albicans [87]. Two compounds cyschalasins A and B were isolated from Aspergillus Micronesiensis and showed antifungal activity against C. albicans [88]. Moriniafungins B-G, a new tetracyclic diterpene glycoside of Sordarincin, was isolated from Curvularia hawaiiensis TA26-15. Moriniafungins B-G had antifungal activity against C. albicans with an MIC value of 2.9 μM [89].
The F2 mycotoxin zearalenone (ZEN) produced by Fusarium and Gibberella species exhibited in vitro inhibitory effects on different microbial strains [90,91]. 100 μg/mL ZEN treatment significantly inhibited C. albicans hyphal morphogenesis and biofilm formation. Similarly, ZEN effectively destroyed established C. albicans biofilms without disturbing the planktonic cells. In vivo, ZEN prominently inhibited C. albicans infection in Caenorhabditis elegans [92].
Deoxynivalenol (DON), produced by Fusarium spp., was an epoxide sesquiterpene compound [93,108,109,110]. DON and 3-acetyl-DON exhibited a dose-dependent inhibitory effect on C. albicans in vitro. DON obviously reduced C. albicans metabolic activity, disrupted pre-formed biofilms, inhibited biofilm formation and inhibited hyphal that embedded in free-living planktonic cells and colonies. DON and 3-acetyl-DON mimicked the mechanism of through interplaying with lanosterol 14α-demethylase that was like the action of azole drugs. DON exhibited antifungal filament and antifungal membrane potential against C. albicans [111].
A carefully scheduled fermentation of P. camembertii/clavigerum and P. fuscum yielded eight novel 16-membered ring macrolides, Berkelilactone A exhibited the most potent antifungal activity in the macrolide series. It had low micromolar inhibitory activity against C. albicans (MIC = 1–2 μg/mL). Berkelilactone A did not inhibit protein synthesis and did not target ribosomes, suggesting a new mode of mechanism for its antibiotic activity, but the specific mechanism had not yet been elucidated [94].
U. maydis secreted a large amount of the glycolipid biosurfactant ustilagic acid. The new glycolipid ustilagic acid C and B were induced under special culture conditions. And the two compounds showed weak antifungal activity against C. albicans [95].
3.8. A Conclusion of Inhibitory Compounds Produced by Antagonistic Microbes
Many natural products that obtained from diverse microbial sources have been successfully applied in many fields. To overcome the increasing drug resistance of C. albicans, the discovery of new natural antifungal compounds is necessary. This review summarizes about 30 different compounds produced by microorganisms that have been found to have inhibitory effects on C. albicans. These compounds are derived from different bacteria and fungi, including bacteria such as Bacillus, T. discolor sv11 and P. agglomerans; yeast such as S. bombicola and S. boulardii; Phoma spp. SYSU-SK-7, Biatriospora sp.; marine-derived fungi such as Aspergillus, P. minioluteum ZZ1657; Streptomyces sp.; A. cyanogriseus WH1-2216-6; Streptomyces sp. ZZ741 and Actinomycetes of marine origin; other fungal sources: Fusarium, Gibberella species, P. brown, P. camembertii/clavigerum, C. Hawaiian ensis TA26-15, U. maydis; A. micronesiensis, Acremonium sp. PSU-MA70 and other fungi. It can be seen from Table 2 that the antifungal mechanisms of most isolated known or unknown compounds have not been clearly analyzed. Only a few compounds have been studied at the transcriptional level. These microorganisms produce compounds with different structures to inhibit C. albicans in different ways, such as inhibiting biofilm formation and hyphal morphological transformation.
4. Conclusions
With the emergence of C. albicans resistance against conventional antifungal therapies, new strategies to treat C. albicans infection are important [112]. Considering that C. albicans could threaten human life and health when the body’s immune function declines or the normal flora is out of balance. both Bacillus licheniformis and Bifidobacterium can be used in clinic to prevent and treat intestinal microbiota disorders. This article reviews the different antagonistic microorganisms of C. albicans and various bioactive secondary metabolites produced by microorganisms, which are expected to achieve biological control of human pathogenic fungus C. albicans.
Biological control of microbial infections is an alternative approach that utilizes antagonistic microorganisms to prevent the growth and infection of harmful microorganisms. Antagonistic microbes, such as bacteria, yeast, and fungus, have been studied as potential antagonistic organisms for the control of C. albicans. Through the study on diverse strains with antifungal activity, it is helpful to develop the agent for inhibiting C. albicans. This is a potential strategy for biological control of C. albicans. On the other hand, secondary metabolites derived from microorganisms are valuable natural compounds. Many natural products have diverse structures and can exhibit significant biological activities. The structures of these compounds include: macrolides, terpenoids, alkaloids, organic acids, and other heterocyclic compounds. The secondary metabolites introduced in Table 2 and Table 3 can significantly inhibit C. albicans. They are produced by diverse microorganisms. However no identified compounds are currently used as a drug against C. albicans. There are still four types of antifungal agents for C. albicans infection: polyene, triazole, 5-fluorocytosine, and echinocandin antifungal drugs [13]. Through the study of these active compounds, it is expected to obtain new drugs for the treatment and prevention of C. albicans infection, thereby maintaining human health.
Author Contributions
Writing—original draft preparation, H.L. (Honghua Li); writing—review and editing, F.S., X.Z., J.Y., X.X. and H.L. (Hehe Li); supervision, F.S. and H.L. (Hehe Li); funding acquisition, H.L. (Honghua Li) and F.S. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable for studies not involving humans or animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Funding Statement
This research was funded by grants from the General Projects of Science and Technology Program of Beijing Municipal Education Commission (KM202210011008), the Key Lab of Marine Bioactive Substance and Modern Analytical Technique, SOA (MBSMAT-2019-06), Research Foundation for Youth Scholars of Beijing Technology and Business University (QNJJ2022-21), and Research Foundation for Advanced Talents of Beijing Technology and Business University (19008021176).
Footnotes
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Ramchandran R., Ramesh S., Thakur R., Chakrabarti A., Roy U. Improved Production of Two Anti-Candida Lipopeptide Homologues Co-Produced by the Wild-Type Bacillus subtilis RLID 12.1 under Optimized Conditions. Curr. Pharm. Biotechnol. 2020;21:438–450. doi: 10.2174/1389201020666191205115008. [DOI] [PubMed] [Google Scholar]
- 2.Karpiński T.M., Ożarowski M., Seremak-Mrozikiewicz A., Wolski H., Adamczak A. Plant Preparations and Compounds with Activities against Biofilms Formed by Candida spp. J. Fungi. 2021;7:360. doi: 10.3390/jof7050360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Brown G.D., Denning D.W., Gow N.A., Levitz S.M., Netea M.G., White T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012;4:165rv13. doi: 10.1126/scitranslmed.3004404. [DOI] [PubMed] [Google Scholar]
- 4.Moran C., Grussemeyer C.A., Spalding J.R., Benjamin D.K., Jr., Reed S.D. Comparison of costs, length of stay, and mortality associated with Candida glabrata and Candida albicans bloodstream infections. Am. J. Infect Control. 2010;38:78–80. doi: 10.1016/j.ajic.2009.06.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Vila T., Sultan A.S., Montelongo-Jauregui D., Jabra-Rizk M.A. Oral Candidiasis: A Disease of Opportunity. J. Fungi. 2020;6:15. doi: 10.3390/jof6010015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ruben S., Garbe E., Mogavero S., Albrecht-Eckardt D., Hellwig D., Häder A., Krüger T., Gerth K., Jacobsen I.D., Elshafee O., et al. Ahr1 and Tup1 Contribute to the Transcriptional Control of Virulence-Associated Genes in Candida albicans. mBio. 2020;11:e00206-20. doi: 10.1128/mBio.00206-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Zakikhany K., Naglik J.R., Schmidt-Westhausen A., Holland G., Schaller M., Hube B. In vivo transcript profiling of Candida albicans identifies a gene essential for interepithelial dissemination. Cell. Microbiol. 2007;9:2938–2954. doi: 10.1111/j.1462-5822.2007.01009.x. [DOI] [PubMed] [Google Scholar]
- 8.Mavor A.L., Thewes S., Hube B. Systemic fungal infections caused by Candida species: Epidemiology, infection process and virulence attributes. Curr. Drug Targets. 2005;6:863–874. doi: 10.2174/138945005774912735. [DOI] [PubMed] [Google Scholar]
- 9.Nobile C.J., Johnson A.D. Candida albicans Biofilms and Human Disease. Annu. Rev. Microbiol. 2015;69:71–92. doi: 10.1146/annurev-micro-091014-104330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Chandra J., Kuhn D.M., Mukherjee P.K., Hoyer L.L., McCormick T., Ghannoum M.A. Biofilm formation by the fungal pathogen Candida albicans: Development, architecture, and drug resistance. J. Bacteriol. 2001;183:5385–5394. doi: 10.1128/JB.183.18.5385-5394.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Nett J.E., Andes D.R. Antifungal Agents: Spectrum of Activity, Pharmacology, and Clinical Indications. Infect. Dis. Clin. N. Am. 2016;30:51–83. doi: 10.1016/j.idc.2015.10.012. [DOI] [PubMed] [Google Scholar]
- 12.Peyclit L., Yousfi H., Rolain J.M., Bittar F. Drug Repurposing in Medical Mycology: Identification of Compounds as Potential Antifungals to Overcome the Emergence of Multidrug-Resistant Fungi. Pharmaceuticals. 2021;14:488. doi: 10.3390/ph14050488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Barantsevich N., Barantsevich E. Diagnosis and Treatment of Invasive Candidiasis. Antibiotics. 2022;11:718. doi: 10.3390/antibiotics11060718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Anderson T.M., Clay M.C., Cioffi A.G., Diaz K.A., Hisao G.S., Tuttle M.D., Nieuwkoop A.J., Comellas G., Maryum N., Wang S., et al. Amphotericin forms an extramembranous and fungicidal sterol sponge. Nat. Chem. Biol. 2014;10:400–406. doi: 10.1038/nchembio.1496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Robbins N., Caplan T., Cowen L.E. Molecular Evolution of Antifungal Drug Resistance. Annu. Rev. Microbiol. 2017;71:753–775. doi: 10.1146/annurev-micro-030117-020345. [DOI] [PubMed] [Google Scholar]
- 16.Hassanmoghadam F., Shokohi T., Hedayati M.T., Aslani N., Haghani I., Nabili M., Lotfali E., Davari A., Moazeni M. High prevalence of itraconazole resistance among Candida parapsilosis isolated from Iran. Curr. Med. Mycol. 2019;5:43–46. doi: 10.18502/cmm.5.3.1746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Galia L., Pezzani M.D., Compri M., Callegari A., Rajendran N.B., Carrara E., Tacconelli E. The Combacte Magnet Epi-Net Network. Surveillance of Antifungal Resistance in Candidemia Fails to Inform Antifungal Stewardship in European Countries. J. Fungi. 2022;8:249. doi: 10.3390/jof8030249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Berkow E.L., Lockhart S.R. Fluconazole resistance in Candida species: A current perspective. Infect. Drug Resist. 2017;10:237–245. doi: 10.2147/IDR.S118892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Whaley S.G., Berkow E.L., Rybak J.M., Nishimoto A.T., Barker K.S., Rogers P.D. Azole Antifungal Resistance in Candida albicans and Emerging Non-albicans Candida Species. Front. Microbiol. 2017;7:2173. doi: 10.3389/fmicb.2016.02173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Delma F.Z., Al-Hatmi A.M.S., Brüggemann R.J.M., Melchers W.J.G., de Hoog S., Verweij P.E., Buil J.B. Molecular Mechanisms of 5-Fluorocytosine Resistance in Yeasts and Filamentous Fungi. J. Fungi. 2021;7:909. doi: 10.3390/jof7110909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ham Y.Y., Lewis J.S., Thompson G.R. Rezafungin: A novel antifungal for the treatment of invasive candidiasis. Future Microbiol. 2021;16:27–36. doi: 10.2217/fmb-2020-0217. [DOI] [PubMed] [Google Scholar]
- 22.Miesel L., Lin K.Y., Ong V. Rezafungin treatment in mouse models of invasive candidiasis and aspergillosis: Insights on the PK/PD pharmacometrics of rezafungin efficacy. Pharmacol. Res. Perspect. 2019;7:e00546. doi: 10.1002/prp2.546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Miesel L., Cushion M.T., Ashbaugh A., Lopez S.R., Ong V. Efficacy of Rezafungin in Prophylactic Mouse Models of Invasive Candidiasis, Aspergillosis, and Pneumocystis Pneumonia. Antimicrob. Agents Chemother. 2021;65:e01992-20. doi: 10.1128/AAC.01992-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lepak A.J., Zhao M., Andes D.R. Determination of Pharmacodynamic Target Exposures for Rezafungin against Candida tropicalis and Candida dubliniensis in the Neutropenic Mouse Disseminated Candidiasis Model. Antimicrob. Agents Chemother. 2019;63:e01556-19. doi: 10.1128/AAC.01556-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Pfaller M.A., Carvalhaes C., Messer S.A., Rhomberg P.R., Castanheira M. Activity of a Long-Acting Echinocandin, Rezafungin, and Comparator Antifungal Agents Tested against Contemporary Invasive Fungal Isolates (SENTRY Program, 2016 to 2018) Antimicrob. Agents Chemother. 2020;64:e00099-20. doi: 10.1128/AAC.00099-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Farhadi Z., Farhadi T., Hashemian S.M. Virtual screening for potential inhibitors of β(1,3)-D-glucan synthase as drug candidates against fungal cell wall. J. Drug Assess. 2020;9:52–59. doi: 10.1080/21556660.2020.1734010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Szymański M., Chmielewska S., Czyżewska U., Malinowska M., Tylicki A. Echinocandins-structure, mechanism of action and use in antifungal therapy. J. Enzyme Inhib. Med. Chem. 2022;37:876–894. doi: 10.1080/14756366.2022.2050224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sandrin C., Peypoux F., Michel G. Coproduction of surfactin and iturin A, lipopeptides with surfactant and antifungal properties, by Bacillus subtilis. Biotechnol. Appl. Biochem. 1990;12:370–375. [PubMed] [Google Scholar]
- 29.Ahimou F., Jacques P., Deleu M. Surfactin and iturin A effects on Bacillus subtilis surface hydrophobicity. Enzyme Microb. Technol. 2000;27:749–754. doi: 10.1016/S0141-0229(00)00295-7. [DOI] [PubMed] [Google Scholar]
- 30.Newman D.J., Cragg G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020;83:770–803. doi: 10.1021/acs.jnatprod.9b01285. [DOI] [PubMed] [Google Scholar]
- 31.Sun F.J., Li M., Gu L., Wang M.L., Yang M.H. Recent progress on anti-Candida natural products. Chin. J. Nat. Med. 2021;19:561–579. doi: 10.1016/S1875-5364(21)60057-2. [DOI] [PubMed] [Google Scholar]
- 32.Guimarães R., Milho C., Liberal Â., Silva J., Fonseca C., Barbosa A., Ferreira I.C.F.R., Alves M.J., Barros L. Antibiofilm Potential of Medicinal Plants against Candida spp. Oral Biofilms: A Review. Antibiotics. 2021;10:1142. doi: 10.3390/antibiotics10091142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Espinel-Ingroff A. Commercial Methods for Antifungal Susceptibility Testing of yeasts: Strengths and Limitations as Predictors of Resistance. J. Fungi. 2022;8:309. doi: 10.3390/jof8030309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Murphy S.E., Bicanic T. Drug Resistance and Novel Therapeutic Approaches in Invasive Candidiasis. Front. Cell. Infect. Microbiol. 2021;11:759408. doi: 10.3389/fcimb.2021.759408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Li B., Pan L., Zhang H., Xie L., Wang X., Shou J., Qi Y., Yan X. Recent Developments on Using Nanomaterials to Combat Candida albicans. Front. Chem. 2021;9:813973. doi: 10.3389/fchem.2021.813973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Khan F., Bamunuarachchi N.I., Tabassum N., Jo D.M., Khan M.M., Kim Y.M. Suppression of hyphal formation and virulence of Candida albicans by natural and synthetic compounds. Biofouling. 2021;37:626–655. doi: 10.1080/08927014.2021.1948538. [DOI] [PubMed] [Google Scholar]
- 37.Owen M.K., Clenney T.L. Management of vaginitis. Am. Fam. Physician. 2004;70:2125–2132. [PubMed] [Google Scholar]
- 38.Siikala E., Rautemaa R., Richardson M., Saxen H., Bowyer P., Sanglard D. Persistent Candida albicans colonization and molecular mechanisms of azole resistance in autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophy (APECED) patients. J. Antimicrob. Chemother. 2010;65:2505–2513. doi: 10.1093/jac/dkq354. [DOI] [PubMed] [Google Scholar]
- 39.Zida A., Bamba S., Yacouba A., Ouedraogo-Traore R., Guiguemdé R.T. Anti-Candida albicans natural products, sources of new antifungal drugs: A review. J. Mycol. Med. 2017;27:1–19. doi: 10.1016/j.mycmed.2016.10.002. [DOI] [PubMed] [Google Scholar]
- 40.Ghai S., Sood S.S., Jain R.K. Antagonistic and antimicrobial activities of some bacterial isolates collected from soil samples. Indian J. Microbiol. 2007;47:77–80. doi: 10.1007/s12088-007-0014-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Khan M.N., Lin H., Li M., Wang J., Mirani Z.A., Khan S.I., Buzdar M.A., Ali I., Jamil K. Identification and growth optimization of a Marine Bacillus DK1-SA11 having potential of producing broad spectrum antimicrobial compounds. Pak. J. Pharm. Sci. 2017;30:839–853. [PubMed] [Google Scholar]
- 42.Devi S., Kiesewalter H.T., Kovács R., Frisvad J.C., Weber T., Larsen T.O., Kovács Á.T., Ding L. Depiction of secondary metabolites and antifungal activity of Bacillus velezensis DTU001. Synth. Syst. Biotechnol. 2019;4:142–149. doi: 10.1016/j.synbio.2019.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Li X., Zhang Y., Wei Z., Guan Z., Cai Y., Liao X. Antifungal Activity of Isolated Bacillus amyloliquefaciens SYBC H47 for the Biocontrol of Peach Gummosis. PLoS ONE. 2016;11:e0162125. doi: 10.1371/journal.pone.0162125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Li M.S.M., Piccoli D.A., McDowell T., MacDonald J., Renaud J., Yuan Z.C. Evaluating the biocontrol potential of Canadian strain Bacillus velezensis 1B-23 via its surfactin production at various pHs and temperatures. BMC Biotechnol. 2021;21:31. doi: 10.1186/s12896-021-00690-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Bonfrate L., Di Palo D.M., Celano G., Albert A., Vitellio P., De Angelis M., Gobbetti M., Portincasa P. Effects of Bifidobacterium longum BB536 and Lactobacillus rhamnosus HN001 in IBS patients. Eur. J. Clin. Invest. 2020;50:e13201. doi: 10.1111/eci.13201. [DOI] [PubMed] [Google Scholar]
- 46.Lau A.S., Yanagisawa N., Hor Y.Y., Lew L.C., Ong J.S., Chuah L.O., Lee Y.Y., Choi S.B., Rashid F., Wahid N., et al. Bifidobacterium longum BB536 alleviated upper respiratory illnesses and modulated gut microbiota profiles in Malaysian pre-school children. Benef. Microbes. 2018;9:61–70. doi: 10.3920/BM2017.0063. [DOI] [PubMed] [Google Scholar]
- 47.Vazquez-Munoz R., Thompson A., Russell J.T., Sobue T., Zhou Y., Dongari-Bagtzoglou A. Insights From the Lactobacillus johnsonii Genome Suggest the Production of Metabolites With Antibiofilm Activity Against the Pathobiont Candida albicans. Front. Microbiol. 2022;13:853762. doi: 10.3389/fmicb.2022.853762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Zeise K.D., Woods R.J., Huffnagle G.B. Interplay between Candida albicans and Lactic Acid Bacteria in the Gastrointestinal Tract: Impact on Colonization Resistance, Microbial Carriage, Opportunistic Infection, and Host Immunity. Clin. Microbiol. Rev. 2021;34:e0032320. doi: 10.1128/CMR.00323-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Sipiczki M. Metschnikowia strains isolated from botrytized grapes antagonize fungal and bacterial growth by iron depletion. Appl. Environ. Microbiol. 2006;72:6716–6724. doi: 10.1128/AEM.01275-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kang C.H., Han S.H., Kim Y., Paek N.S., So J.S. In Vitro Probiotic Properties of Lactobacillus salivarius MG242 Isolated from Human Vagina. Probiotics Antimicrob. Proteins. 2018;10:343–349. doi: 10.1007/s12602-017-9323-5. [DOI] [PubMed] [Google Scholar]
- 51.Chae G.P., Shoda M., Kubota H. Suppressive effect of Bacillus subtilis and it’s products on phytopathogenic microorganisms. J. Ferment. Bio. Eng. 1990;69:1–7. doi: 10.1016/0922-338X(90)90155-P. [DOI] [Google Scholar]
- 52.Ongena M., Jacques P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008;16:115–125. doi: 10.1016/j.tim.2007.12.009. [DOI] [PubMed] [Google Scholar]
- 53.Abriouel H., Franz C.M., Ben Omar N., Gálvez A. Diversity and applications of Bacillus bacteriocins. FEMS Microbiol. Rev. 2011;35:201–232. doi: 10.1111/j.1574-6976.2010.00244.x. [DOI] [PubMed] [Google Scholar]
- 54.Abou-Kassem D.E., Elsadek M.F., Abdel-Moneim A.E., Mahgoub S.A., Elaraby G.M., Taha A.E., Elshafie M.M., Alkhawtani D.M., Abd El-Hack M.E., Ashour E.A. Growth, carcass characteristics, meat quality, and microbial aspects of growing quail fed diets enriched with two different types of probiotics (Bacillus toyonensis and Bifidobacterium bifidum) Poult. Sci. 2021;100:84–93. doi: 10.1016/j.psj.2020.04.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Kadja L., Dib A.L., Lakhdara N., Bouaziz A., Espigares E., Gagaoua M. Influence of Three Probiotics Strains, Lactobacillus rhamnosus GG, Bifidobacterium animalis subsp. Lactis BB-12 and Saccharomyces boulardii CNCM I-745 on the Biochemical and Haematological Profiles and Body Weight of Healthy Rabbits. Biology. 2021;10:1194. doi: 10.3390/biology10111194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Lau A.S., Liong M.T. Lactic Acid Bacteria and Bifidobacteria-Inhibited Staphylococcus epidermidis. Wounds. 2014;26:121–131. [PubMed] [Google Scholar]
- 57.Fukuda S., Toh H., Hase K., Oshima K., Nakanishi Y., Yoshimura K., Tobe T., Clarke J.M., Topping D.L., Suzuki T., et al. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature. 2011;469:543–547. doi: 10.1038/nature09646. [DOI] [PubMed] [Google Scholar]
- 58.El-Zahar K.M., Hassan M.F.Y., Al-Qaba S.F. Protective Effect of Fermented Camel Milk Containing Bifidobacterium longum BB536 on Blood Lipid Profile in Hypercholesterolemic Rats. J. Nutr. Metab. 2021;2021:1557945. doi: 10.1155/2021/1557945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Suido H., Miyao M. Bifidobacterium longum-fermented broccoli supernatant inhibited the growth of Candida albicans and some pathogenic bacteria in vitro. Biocontrol. Sci. 2008;13:41–48. doi: 10.4265/bio.13.41. [DOI] [PubMed] [Google Scholar]
- 60.Chevalier M., Ranque S., Prêcheur I. Oral fungal-bacterial biofilm models in vitro: A review. Med Mycol. 2018;56:653–667. doi: 10.1093/mmy/myx111. [DOI] [PubMed] [Google Scholar]
- 61.Kluyver A.J., van der Walt J.P., van Triet A.J. Pulcherrimin, The Pigment of Candida pulcherrima. Proc. Natl. Acad. Sci. USA. 1953;39:583–593. doi: 10.1073/pnas.39.7.583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Türkel S., Ener B. Isolation and characterization of new Metschnikowia pulcherrima strains as producers of the antimicrobial pigment pulcherrimin. Zeitschrift für Naturforschung. 2009;64:405–410. doi: 10.1515/znc-2009-5-618. [DOI] [PubMed] [Google Scholar]
- 63.Renda G., Kadıoğlu M., Kılıç M., Korkmaz B., Kırmızıbekmez H. Anti-inflammatory secondary metabolites from Scrophularia kotschyana. Hum. Exp. Toxicol. 2021;40:S676–S683. doi: 10.1177/09603271211058889. [DOI] [PubMed] [Google Scholar]
- 64.Muhaj F.F., George S.J., Nguyen C.D., Tyring S.K. Antimicrobials and resistance part II: Antifungals, antivirals, and antiparasitics. J. Am. Acad. Dermatol. 2022;86:1207–1226. doi: 10.1016/j.jaad.2021.11.065. [DOI] [PubMed] [Google Scholar]
- 65.Shala A., Singh S., Hameed S., Khurana S.M.P. Essential Oils as Alternative Promising Anti-Candidal Agents: Progress and Prospects. Curr. Pharm. Des. 2022;28:58–70. doi: 10.2174/1381612827666210914124344. [DOI] [PubMed] [Google Scholar]
- 66.Dehghanifar S., Keyhanfar M., Emtiazi G. Production and partial purification of thermostable bacteriocins from Bacillus pumilus ZED17 and DFAR8 strains with antifungal activity. Mol. Biol. Res. Commun. 2019;8:41–49. doi: 10.22099/mbrc.2019.31563.1367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Boparai J.K., Sharma P.K. Mini Review on Antimicrobial Peptides, Sources, Mechanism and Recent Applications. Protein Pept. Lett. 2020;27:4–16. doi: 10.2174/18755305MTAwENDE80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Bin Hafeez A., Jiang X., Bergen P.J., Zhu Y. Antimicrobial Peptides: An Update on Classifications and Databases. Int. J. Mol. Sci. 2021;22:11691. doi: 10.3390/ijms222111691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Subramenium G.A., Swetha T.K., Iyer P.M., Balamurugan K., Pandian S.K. 5-hydroxymethyl-2-furaldehyde from marine bacterium Bacillus subtilis inhibits biofilm and virulence of Candida albicans. Microbiol. Res. 2018;207:19–32. doi: 10.1016/j.micres.2017.11.002. [DOI] [PubMed] [Google Scholar]
- 70.Sammer U.F., Völksch B., Möllmann U., Schmidtke M., Spiteller P., Spiteller M., Spiteller D. 2-amino-3-(oxirane-2,3-dicarboxamido)-propanoyl-valine, an effective peptide antibiotic from the epiphyte Pantoea agglomerans 48b/90. Appl. Environ. Microbiol. 2009;75:7710–7717. doi: 10.1128/AEM.01244-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Wang L., Linares-Otoya V., Liu Y., Mettal U., Marner M., Armas-Mantilla L., Willbold S., Kurtán T., Linares-Otoya L., Schäberle T.F. Discovery and Biosynthesis of Antimicrobial Phenethylamine Alkaloids from the Marine Flavobacterium Tenacibaculum discolor sv11. J. Nat. Prod. 2022;85:1039–1051. doi: 10.1021/acs.jnatprod.1c01173. [DOI] [PubMed] [Google Scholar]
- 72.Murzyn A., Krasowska A., Stefanowicz P., Dziadkowiec D., Łukaszewicz M. Capric acid secreted by S. boulardii inhibits C. albicans filamentous growth, adhesion and biofilm formation. PLoS ONE. 2010;5:e12050. doi: 10.1371/journal.pone.0012050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Zhang M., Chang W., Shi H., Zhou Y., Zheng S., Li Y., Li L., Lou H. Biatriosporin D displays anti-virulence activity through decreasing the intracellular cAMP levels. Toxicol. Appl. Pharmacol. 2017;322:104–112. doi: 10.1016/j.taap.2017.03.004. [DOI] [PubMed] [Google Scholar]
- 74.Zhao J.C., Wang Y.L., Zhang T.Y., Chen Z.J., Yang T.M., Wu Y.Y., Sun C.P., Ma X.C., Zhang Y.X. Indole diterpenoids from the endophytic fungus Drechmeria sp. as natural antimicrobial agents. Phytochemistry. 2018;148:21–28. doi: 10.1016/j.phytochem.2018.01.010. [DOI] [PubMed] [Google Scholar]
- 75.Chen Y., Yang W., Zou G., Chen S., Pang J., She Z. Bioactive polyketides from the mangrove endophytic fungi Phoma sp. SYSU-SK-7. Fitoterapia. 2019;139:104369. doi: 10.1016/j.fitote.2019.104369. [DOI] [PubMed] [Google Scholar]
- 76.Yang B., He Y., Lin S., Zhang J., Li H., Wang J., Hu Z., Zhang Y. Antimicrobial Dolabellanes and Atranones from a Marine-Derived Strain of the Toxigenic Fungus Stachybotrys chartarum. J. Nat. Prod. 2019;82:1923–1929. doi: 10.1021/acs.jnatprod.9b00305. [DOI] [PubMed] [Google Scholar]
- 77.Wu S.H., He J., Li X.N., Huang R., Song F., Chen Y.W., Miao C.P. Guaiane sesquiterpenes and isopimarane diterpenes from an endophytic fungus Xylaria sp. Phytochemistry. 2014;105:197–204. doi: 10.1016/j.phytochem.2014.04.016. [DOI] [PubMed] [Google Scholar]
- 78.Wang X., You J., King J.B., Powell D.R., Cichewicz R.H. Waikialoid A suppresses hyphal morphogenesis and inhibits biofilm development in pathogenic Candida albicans. J. Nat. Prod. 2012;75:707–715. doi: 10.1021/np2009994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Okabe M., Sugita T., Kinoshita K., Koyama K. Macrolides from a Marine-Derived Fungus, Penicillium meleagrinum var. viridiflavum, Showing Synergistic Effects with Fluconazole against Azole-Resistant Candida albicans. J. Nat. Prod. 2016;79:1208–1212. doi: 10.1021/acs.jnatprod.6b00019. [DOI] [PubMed] [Google Scholar]
- 80.Ma M.Z., Ge H.J., Yi W.W., Wu B., Zhang Z.Z. Bioactive drimane sesquiterpenoids and isocoumarins from the marine-derived fungus Penicillium minioluteum ZZ1657. Tetrahedron Lett. 2019;7:13. doi: 10.1016/j.tetlet.2019.151504. [DOI] [Google Scholar]
- 81.Fu P., Wang S., Hong K., Li X., Liu P., Wang Y., Zhu W. Cytotoxic bipyridines from the marine-derived actinomycete Actinoalloteichus cyanogriseus WH1-2216-6. J. Nat. Prod. 2011;74:1751–1756. doi: 10.1021/np200258h. [DOI] [PubMed] [Google Scholar]
- 82.Kim D.G., Moon K., Kim S.H., Park S.H., Park S., Lee S.K., Oh K.B., Shin J., Oh D.C. Bahamaolides A and B, antifungal polyene polyol macrolides from the marine actinomycete Streptomyces sp. J. Nat. Prod. 2012;75:959–967. doi: 10.1021/np3001915. [DOI] [PubMed] [Google Scholar]
- 83.Zhang D., Yi W., Ge H., Zhang Z., Wu B. Bioactive Streptoglutarimides A-J from the Marine-Derived Streptomyces sp. ZZ741. J. Nat. Prod. 2019;82:2800–2808. doi: 10.1021/acs.jnatprod.9b00481. [DOI] [PubMed] [Google Scholar]
- 84.Nithyanand P., Beema Shafreen R.M., Muthamil S., Karutha Pandian S. Usnic acid inhibits biofilm formation and virulent morphological traits of Candida albicans. Microbiol. Res. 2015;179:20–28. doi: 10.1016/j.micres.2015.06.009. [DOI] [PubMed] [Google Scholar]
- 85.Chang W., Li Y., Zhang L., Cheng A., Lou H. Retigeric acid B attenuates the virulence of Candida albicans via inhibiting adenylyl cyclase activity targeted by enhanced farnesol production. PLoS ONE. 2012;7:e41624. doi: 10.1371/journal.pone.0041624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Padhi S., Masi M., Cimmino A., Tuzi A., Jena S., Tayung K., Evidente A. Funiculosone, a substituted dihydroxanthene-1,9-dione with two of its analogues produced by an endolichenic fungus Talaromyces funiculosus and their antimicrobial activity. Phytochemistry. 2019;157:175–183. doi: 10.1016/j.phytochem.2018.10.031. [DOI] [PubMed] [Google Scholar]
- 87.Rukachaisirikul V., Rodglin A., Sukpondma Y., Phongpaichit S., Buatong J., Sakayaroj J. Phthalide and isocoumarin derivatives produced by an Acremonium sp. isolated from a mangrove Rhizophora apiculata. J. Nat. Prod. 2012;75:853–858. doi: 10.1021/np200885e. [DOI] [PubMed] [Google Scholar]
- 88.Wu Z., Zhang X., Anbari W.H.A., Zhou Q., Zhou P., Zhang M., Zeng F., Chen C., Tong Q., Wang J., et al. Cysteine Residue Containing Merocytochalasans and 17,18-seco-Aspochalasins from Aspergillus micronesiensis. J. Nat. Prod. 2019;82:2653–2658. doi: 10.1021/acs.jnatprod.9b00016. [DOI] [PubMed] [Google Scholar]
- 89.Zhang M.Q., Xu K.X., Xue Y., Cao F., Yang L.J., Hou X.M., Wang C.Y., Shao C.L. Sordarin Diterpene Glycosides with an Unusual 1,3-Dioxolan-4-one Ring from the Zoanthid-Derived Fungus Curvularia hawaiiensis TA26-15. J. Nat. Prod. 2019;82:2477–2482. doi: 10.1021/acs.jnatprod.9b00164. [DOI] [PubMed] [Google Scholar]
- 90.Geng Z., Zhu W., Su H., Zhao Y., Zhang K.Q., Yang J. Recent advances in genes involved in secondary metabolite synthesis, hyphal development, energy metabolism and pathogenicity in Fusarium graminearum (teleomorph Gibberella zeae) Biotechnol. Adv. 2014;32:390–402. doi: 10.1016/j.biotechadv.2013.12.007. [DOI] [PubMed] [Google Scholar]
- 91.Hidy P.H., Baldwin R.S., Greasham R.L., Keith C.L., McMullen J.R. Zearalenone and some derivatives: Production and biological activities. Adv. Appl. Microbiol. 1977;22:59–82. doi: 10.1016/s0065-2164(08)70160-6. [DOI] [PubMed] [Google Scholar]
- 92.Rajasekharan S.K., Lee J.H., Zhao Y., Lee J. The Mycotoxin Zearalenone Hinders Candida albicans Biofilm Formation and Hyphal Morphogenesis. Indian J. Microbiol. 2018;58:19–27. doi: 10.1007/s12088-017-0690-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Rajasekharan S.K., Byun J., Lee J. Inhibitory effects of deoxynivalenol on pathogenesis of Candida albicans. J. Appl. Microbiol. 2018;125:1266–1275. doi: 10.1111/jam.14032. [DOI] [PubMed] [Google Scholar]
- 94.Stierle A.A., Stierle D.B., Decato D., Priestley N.D., Alverson J.B., Hoody J., McGrath K., Klepacki D. The Berkeleylactones, Antibiotic Macrolides from Fungal Coculture. J. Nat Prod. 2017;80:1150–1160. doi: 10.1021/acs.jnatprod.7b00133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Yang X.L., Takayoshi A., Toshiyuki W., Ikuro A. Induced production of the novel glycolipid ustilagic acid C in the plant pathogen Ustilago maydis. Tetrahedron Lett. 2013;54:3655–3657. doi: 10.1016/j.tetlet.2013.04.131. [DOI] [Google Scholar]
- 96.Caetano T., Krawczyk J.M., Mösker E., Süssmuth R.D., Mendo S. Heterologous expression, biosynthesis, and mutagenesis of type II lantibiotics from Bacillus licheniformis in Escherichia coli. Chem. Biol. 2011;18:90–100. doi: 10.1016/j.chembiol.2010.11.010. [DOI] [PubMed] [Google Scholar]
- 97.Fira D., Dimkić I., Berić T., Lozo J., Stanković S. Biological control of plant pathogens by Bacillus species. J. Biotechnol. 2018;285:44–55. doi: 10.1016/j.jbiotec.2018.07.044. [DOI] [PubMed] [Google Scholar]
- 98.Chen W., Wang J., Huang D., Cheng W., Shao Z., Cai M., Zheng L., Yu Z., Zhang J. Volatile Organic Compounds from Bacillus aryabhattai MCCC 1K02966 with Multiple Modes against Meloidogyne incognita. Molecules. 2021;27:103. doi: 10.3390/molecules27010103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Chu J., Wang Y., Zhao B., Zhang X.M., Liu K., Mao L., Kalamiyets E. Isolation and identification of new antibacterial compounds from Bacillus pumilus. Appl. Microbiol. Biotechnol. 2019;103:8375–8381. doi: 10.1007/s00253-019-10083-y. [DOI] [PubMed] [Google Scholar]
- 100.Fu L., Wang Y., Ju J., Cheng L., Xu Y., Yu B., Wang L. Extracellular production of active-form Streptomyces mobaraensis transglutaminase in Bacillus subtilis. Appl. Microbiol. Biotechnol. 2020;104:623–631. doi: 10.1007/s00253-019-10256-9. [DOI] [PubMed] [Google Scholar]
- 101.Zhang K., Su L., Wu J. Enhancing Extracellular Pullulanase Production in Bacillus subtilis through dltB Disruption and Signal Peptide Optimization. Appl. Biochem. Biotechnol. 2022;194:1206–1220. doi: 10.1007/s12010-021-03617-6. [DOI] [PubMed] [Google Scholar]
- 102.Xia Y., Zhao J., Chen H., Liu X., Wang Y., Tian F., Zhang H.P., Zhang H., Chen W. Extracellular secretion in Bacillus subtilis of a cytoplasmic thermostable beta-galactosidase from Geobacillus stearothermophilus. J. Dairy Sci. 2010;93:2838–2845. doi: 10.3168/jds.2009-2864. [DOI] [PubMed] [Google Scholar]
- 103.Nissan G., Gershovits M., Morozov M., Chalupowicz L., Sessa G., Manulis-Sasson S., Barash I., Pupko T. Revealing the inventory of type III effectors in Pantoea agglomerans gall-forming pathovars using draft genome sequences and a machine-learning approach. Mol. Plant Pathol. 2018;19:381–392. doi: 10.1111/mpp.12528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Nissan G., Chalupowicz L., Sessa G., Manulis-Sasson S., Barash I. Two Pantoea agglomerans type III effectors can transform nonpathogenic and phytopathogenic bacteria into host-specific gall-forming pathogens. Mol. Plant Pathol. 2019;20:1582–1587. doi: 10.1111/mpp.12860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Sun L.M., Cheng A.X., Wu X.Z., Zhang H.J., Lou H.X. Synergistic mechanisms of retigeric acid B and azoles against Candida albicans. J. Appl. Microbiol. 2010;108:341–348. doi: 10.1111/j.1365-2672.2009.04429.x. [DOI] [PubMed] [Google Scholar]
- 106.Chang W.Q., Wu X.Z., Cheng A.X., Zhang L., Ji M., Lou H.X. Retigeric acid B exerts antifungal effect through enhanced reactive oxygen species and decreased cAMP. Biochim. Biophys. Acta. 2011;1810:569–576. doi: 10.1016/j.bbagen.2011.02.001. [DOI] [PubMed] [Google Scholar]
- 107.Chang W., Li Y., Zhang L., Cheng A., Liu Y., Lou H. Retigeric acid B enhances the efficacy of azoles combating the virulence and biofilm formation of Candida albicans. Biol Pharm Bull. 2012;35:1794–1801. doi: 10.1248/bpb.b12-00511. [DOI] [PubMed] [Google Scholar]
- 108.Sobrova P., Adam V., Vasatkova A., Beklova M., Zeman L., Kizek R. Deoxynivalenol and its toxicity. Interdiscip. Toxicol. 2010;3:94–99. doi: 10.2478/v10102-010-0019-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Audenaert K., Vanheule A., Höfte M., Haesaert G. Deoxynivalenol: A major player in the multifaceted response of Fusarium to its environment. Toxins. 2013;6:1–19. doi: 10.3390/toxins6010001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.May H.D., Wu Q., Blake C.K. Effects of the Fusarium spp. mycotoxins fusaric acid and deoxynivalenol on the growth of Ruminococcus albus and Methanobrevibacter ruminantium. Can. J. Microbiol. 2000;46:692–699. doi: 10.1139/w00-045. [DOI] [PubMed] [Google Scholar]
- 111.Berthiller F., Dall’Asta C., Schuhmacher R., Lemmens M., Adam G., Krska R. Masked mycotoxins: Determination of a deoxynivalenol glucoside in artificially and naturally contaminated wheat by liquid chromatography-tandem mass spectrometry. J. Agric Food Chem. 2005;53:3421–3425. doi: 10.1021/jf047798g. [DOI] [PubMed] [Google Scholar]
- 112.Ellepola A.N., Samaranayake L.P., Khan Z.U. Extracellular phospholipase production of oral Candida albicans isolates from smokers, diabetics, asthmatics, denture wearers and healthy individuals following brief exposure to polyene, echinocandin and azole antimycotics. Braz. J. Microbiol. 2016;47:911–916. doi: 10.1016/j.bjm.2016.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.