

Abstract
Objective
Insulinoma is one of the functional neuroendocrine tumors, which induce hypoglycemia caused by inadequate high secretion of insulin. In rats, two genes coding for insulin, insulin 1 (Ins1) and insulin 2 (Ins2) are found on chromosome 1. Some structural feature studies have shown that Ins1 was generated from a transcript of Ins2 and was inserted into the genome by an RNA-mediated duplication–transposition event.
Methods
In this study, the author has investigated how the expression of insulin genes, Ins 1 and Ins2, are altered by Pdx-1 gene overexpression in the insulinoma cell line, Rin-5F.
Results
Overexpression of the Pdx-1 gene increases Ins1 gene mRNA expression, not Ins2 gene mRNA expression, in the insulinoma cell line. Thus, levels of the rat insulin 1 and insulin 2 peptides may be changed under specific conditions.
Conclusions
This is the first report that Ins1, but not Ins2, is significantly increased by Pdx-1 gene overexpression in the insulinoma cell line. This could indicate more research and analysis of insulinoma tumorigenesis and Pdx-1 gene expression.
Keywords: Ins1, Ins2, Insulinoma, Pdx-1, Rin-5F
Introduction
Insulinoma is one functional neuroendocrine tumor that induces hypoglycemia caused by inadequate high secretion of insulin. It appears as a solitary benign tumor, but can also be part of Multiple endocrine neoplasia (MEN) type 1. Pancreatic and duodenal homeobox-1 (Pdx-1) is a transcription factor that regulates the expression of numerous critical B cell-specific genes and is involved in B cell differentiation and the maintenance of mature B cell function (1-3). In addition, the partial deficiency of Pdx-1 increases B cell apoptosis, resulting in a decrease in B cell mass and diabetes in rodents and humans (4). Similarly, a recent study found that glucotoxicity and lipotoxicity cause B cell death and dysfunction as a result of an oxidative stress-induced decrease in Pdx-1 expression (5).
Insulin is essential for mammalian health, but its function has changed among species. The advent of genome sequencing has identified a wide variety of insulin-coding in the genomes of various mammals, and has identified the number of genes in genomes and unexpected changes in their sequences that have the potential to affect biological function (6). Indeed, in rats, two genes coding for insulin, insulin 1 (Ins1) and insulin 2 (Ins2) are found on chromosome 1. Some structural feature studies have shown that Ins1 was generated from a transcript of Ins2 and was inserted into the genome by an RNA-mediated duplication-transposition event (7).
In insulinoma cell lines, the connection between Pdx-1 overexpression and insulin gene expression was not thoroughly investigated. The Rin-5F cell line has been established as a continuous cell line in rats that preserves numerous pancreatic B cell features, including insulin secretion (8,9). In this report, the author has investigated how the expression of insulin genes, Ins1 and Ins2, is altered by overexpression of the Pdx-1 gene in the insulinoma cell line, Rin-5F.
MATERIALS AND METHODS
Cell culture
Rin-5F cells (#CRL-2058) were derived from the American Type Culture Collection (Manassas, VA, USA). The cell lines were grown in RPMI 1640 medium supplemented with 10% FBS, 100 U/ml penicillin, and 100 µg/ml streptomycin. At 37 °C, cells were cultured in an incubator with humidified air containing 95% air and 5% CO2.
Plasmid construction and cell transfection
The vector of pcDNA 3.4-Pdx-1 was created according to previously reported procedures (2). The Rin-5F cells were seeded in 6-well plates, grown overnight to 80% confluence before transfection, and transfected with pcDNA 3.4-Pdx-1 vector or pcDNA 3.4 control vector. According to the manufacturer’s instructions, all plasmid transactions were carried out with Lipofectamine 3000 (Invitrogen, Carlsbad, CA, USA). Within 5 days of incubation, the transfected Rin-5F cells were respectively harvested for RNA extraction (each group, n = 6).
Real-time PCR analysis
A widely available kit was used to isolate RNA from Rin-5F cell lines (RNA easy mini kit; Qiagen GmbH, Hilden, Germany). The purity of the obtained total RNA was evaluated by electrophoretic separation on an ethidium bromide-containing 1% agarose gel, and RNA was quantified spectrophotometrically at 260/280 nm. A PrimeScript™ RT reagent Kit with gDNA Eraser cDNA was used to reverse-transcribe cDNA from whole RNA (Takara Bio Inc., Kusatsu, Japan). The real-time PCR analysis was carried out using the GeneFields Quick Real-Time PCR system instrument (Kurabo Co., Osaka, Japan). SYBER GREEN Quick PCR Master Mix was used for the experiment (Kurabo Co., Osaka, Japan). The copy counts of Pdx-1, Ins1, Ins2, and Gapdh mRNA were calculated using the melting curves and CT values produced. The PCR primers and matching sequences were designed using the online tool (https://www.takara-bio.co.jp/research/prt/intro.htm ) (Takara Bio Inc., Kusatsu, Japan) (Table 1).
Table 1.
Primer sequences of Pdx-1, Ins1, Ins2, Gapdh genes used in the real-time PCR analysis
| Gene name | Forward | Reverse | Product size (bp) |
|---|---|---|---|
| Pdx-1 | GGATGAAATCCACCAAAGCTCAC | AGCATCACTGCCAGCTCCA | 187 |
| Ins1 | AGCGTGGCATTGTGGATCAG | TCAAAGGCTTTATTCATTGCAGAGG | 117 |
| Ins2 | CAGCACCTTTGTGGTTCTCACTT | CTCCACCCAGCTCCAGTTGT | 130 |
| Gapdh | GGCACAGTCAAGGCTGAGAATG | ATGGTGGTGAAGACGCCAGTA | 143 |
Western blot analysis
RIPA buffer was used to extract total protein from cells (Aproscience, Tokushima, Japan). The protein concentration of the supernatant was determined using the BCA protein assay kit (Takara Bio Inc., Kusatsu, Japan). By using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), equal amounts of protein samples (40 µg) were separated and electrically transferred onto polyvinylidene difluoride (PVDF) membrane (Life Technologies, CA, USA). Incubation with 5% fat-free milk in Tris-buffered saline with 0.1% Tween-20 (TBST) at room temperature for two hours inhibited non-specific binding. After that, the membranes were treated at overnight 4 °C with primary antibodies Pdx-1 (1:1000; Cell Signaling Technology Inc., MA, USA) and Gapdh (1:5000; Abcam Plc, Cambridge, UK). The membranes were washed and incubated for two hours at room temperature with HRP-conjugated secondary antibodies (Cell Signaling Technology Inc, MA, USA), diluted in 1:2000. ECL Prime Western Blotting Detection Reagent (Amersham, Buckinghamshire, UK) was used to view immunoblots, which were subsequently scanned using ImageQuant LAS 4000mini scanner (GE Healthcare, Tokyo, Japan). The relative integrated density values were normalized to that of Gapdh. ImageJ software (https://imagej.nih.gov/ij ) was used to analyze the images.
Statistical analysis
The results are presented as mean ± SEM. The Mann-Whitney U test was used to examine the amounts of mRNA and protein. The statistical analysis was carried out using SPSS statistical program version 11.0 (SPSS Japan Inc., Tokyo, Japan). If the P < 0.05 (2-tailed), the differences between the groups were considered significant.
RESULTS
The Pdx-1 mRNA expressions of pcDNA 3.4-Pdx-1 vector-transfected Rin-5F cells of 2 days were significantly higher than those of other days (Figure 1). Also, at 2 days, the Pdx-1 proteins of pcDNA 3.4-Pdx-1 vector-transfected Rin-5F cells were approximately 3 times higher than that of pcDNA 3.4-Control vector-transfected cells (Fig. 2). Moreover, at 2 days, the Ins1 mRNA expression of these cells increased significantly and then decreased to the initial levels at 5 days (Fig. 3A), whereas in the Ins2 mRNA expression, no significant increases were observed after as long as 5 days (Fig. 3B).
Figure 1.

Time course of changes in pdx-1 mRNA levels after pcDNA 3.4-Pdx-1 vector transfection or pcDNA 3.4-control vector transfection. Each n = 6 samples. * P < 0.05 compared with the mRNA levels after the pcDNA 3.4-control vector transfection on the same day.
Figure 2.
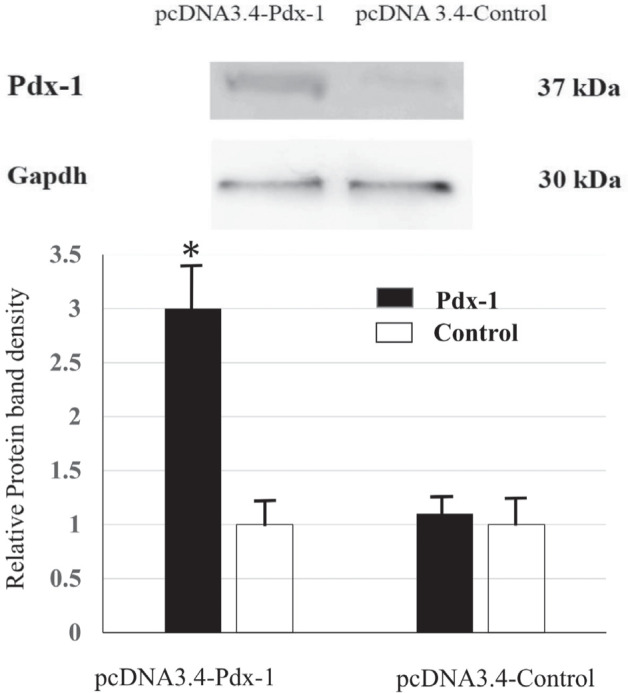
Differences of pdx-1 protein levels between pcDNA 3.4-Pdx-1 vector transfection and pcDNA 3.4-control vector at 2 days after transfection. Each n = 6 samples. * P < 0.05 compared with the protein levels of the pcDNA 3.4-control vector transfection.
Figure 3.

Time course of changes in Ins1 (A) and Ins2 (B) mRNA levels after pcDNA 3.4-Pdx-1 vector transfection or pcDNA 3.4-control vector transfection. Each n = 6 samples. * P < 0.05 compared with the mRNA levels of pcDNA 3.4-control vector transfection at the same days.
DISCUSSION
In the present study, overexpression of the Pdx-1 gene has increased Ins1 gene mRNA expression, but not Ins2 gene mRNA expression. The pcDNA 3.4 TOPO® vector (Life Technologies, CA, USA) is a constitutive mammalian expression vector that may achieve extremely high transgene expression levels. Higher expression levels are often achieved with the improved, full-length CMV promoter and other expression components than with pcDNA 3.1-based expression constructs. In insulin-producing cells and non-B cells, Pdx-1 would have contradictory effects on insulin mRNA expression and insulin promoter activity (10). The promotion of insulin production or the transactivation of insulin promoters is linked to the ectopic expression of Pdx-1 in non-B cells (10). Overexpression of Pdx-1 was previously shown to not increase, but rather decrease, insulin mRNA levels in INS cell line, a rat insulinoma cell line, and the findings showed that Pdx-1 might also influence insulin production or the synthesis or maturation of insulin granules (11). However, in this study, approximately 30 times overexpression of the Pdx-1 gene increased Ins1 gene mRNA expression. Consistent with this, in comparison to non-induced cells, doxycycline-induced Pdx-1 expression in embryonic stem cells elevated Ins1 and Ins2 mRNA by 140% compared to the insulinoma cell line βTC6 (12). In this study, approximately 30 times overexpression of the Pdx-1 gene upregulated Ins1 mRNA by approximately 300% compared to the insulinoma cell line Rin-5F.
In this study, overexpression of the Pdx-1 gene has increased Ins1 gene mRNA expression, but not Ins2 gene mRNA expression. Two non-allelic insulin genes are found in rats on a rodent-specific Ins1 gene, which most likely evolved through the transposition of a reverse-transcribed, partially processed mRNA of the ancestral Ins2 (7). However, Ins1 and Ins2 have different promoter elements, tissue- and temporal-specific expression patterns, and imprinting status (13). Furthermore, the translation or processing rates of the two murine preproinsulins were shown to be different (14). Moreover, the Ins1-knockout prevented most cases of autoimmune type 1 diabetes, but the Ins2-knockout caused type 1 diabetes to develop (15). Thus, it is possible that the expression levels of insulin 1 and insulin 2 peptides may be changed under specific conditions, and that the Pdx-1 gene may play a different role in Ins1 and Ins2 gene expression during insulin production. Also, it was reported that targeting Pdx-1 in mouse models of insulinoma prevented death from hyperinsulinemia and hypoglycemia (16). Further studies are needed to clarify whether or not this phenomenon may play a difference in Ins1 and Ins 2 gene expression.
In conclusion, this is the first report that Ins1, but not Ins2, is significantly increased by Pdx-1 gene overexpression in the insulinoma cell line, Rin-5F. Further studies are needed for the reason that overexpression of the Pdx-1 gene increases Ins1 gene mRNA expression, but not Ins2 gene mRNA expression. This may indicate the need for further documents and analysis of insulinoma tumorigenesis and Pdx-1 gene expression.
Conflict of interest
The author declares no conflict of interest.
Ethics approval and consent to participate
All experimental techniques were carried out according to the Guidelines for Recombinant DNA Experiments at Okayama University of Science’s Faculty of Science. The Protocols were accepted by the Okayama University of Science’s Faculty of Science’s Safety Committee for Recombinant DNA Experiments (Approval Number 1714).
References
- 1.Kiba T. Relationships between the autonomic nervous system and the pancreas including regulation of regeneration and apoptosis: recent developments. Pancreas. 2004;29(2):e51–e58. doi: 10.1097/00006676-200408000-00019. [DOI] [PubMed] [Google Scholar]
- 2.Kiba T. Novel codons in rat Pdx-1 complementary DNA. BMC Res Notes. 2018;11(1):726. doi: 10.1186/s13104-018-3837-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Glavas MM, Hui Q, Tudurí E, Erener S, Kasteel NL, Johnson JD, Kieffer TJ. Early overnutrition reduces Pdx1 expression and induces β cell failure in Swiss Webster mice. Sci Rep. 2019;9(1):3619. doi: 10.1038/s41598-019-39177-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Fujimoto K, Chen Y, Polonsky KS, Dorn GW. Targeting cyclophilin D and the mitochondrial permeability transition enhances beta-cell survival and prevents diabetes in Pdx1 deficiency. Proc Natl Acad Sci USA. 2010;107(22):10214–10219. doi: 10.1073/pnas.0914209107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Zhang M, Yang C, Zhu M, Qian L, Luo Y, Cheng H, Geng R, Xu X, Qian C, Liu Y. Saturated fatty acids entrap PDX1 in stress granules and impede islet beta cell function. Diabetologia. 2021;64(5):1144–1157. doi: 10.1007/s00125-021-05389-4. [DOI] [PubMed] [Google Scholar]
- 6.Irwin DM. Evolution of the Insulin Gene: Changes in Gene Number, Sequence, and Processing. Front Endocrinol (Lausanne) 2021;12:649255. doi: 10.3389/fendo.2021.649255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Soares MB, Schon E, Henderson A, Karathanasis SK, Cate R, Zeitlin S, Chirgwin J, Efstratiadis A. RNA-mediated gene duplication: the rat preproinsulin I gene is a functional retroposon. Mol Cell Biol. 1985;5(8):2090–2103. doi: 10.1128/mcb.5.8.2090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kiba T. Mutation Status in Yin Yang 1 Gene in X-ray-Induced Insulinoma Cell Line Rin-5F. Pancreas. 2020;49(6):e49–e50. doi: 10.1097/MPA.0000000000001565. [DOI] [PubMed] [Google Scholar]
- 9.Kiba T, Oka N. Mutation Status in Phosphatase and Tensin Homolog Deleted on Chromosome 10 Gene in X-ray-Induced Insulinoma Cell Line Rin-5F. Pancreas. 2021;50(8):e66–e67. doi: 10.1097/MPA.0000000000001884. [DOI] [PubMed] [Google Scholar]
- 10.Ferber S, Halkin A, Cohen H, Ber I, Einav Y, Goldberg I, Barshack I, Seijffers R, Kopolovic J, Kaiser N, Karasik A. Pancreatic and duodenal homeobox gene 1 induces expression of insulin genes in liver and ameliorates streptozotocin-induced hyperglycemia. Nat Med. 2000;6(5):568–572. doi: 10.1038/75050. [DOI] [PubMed] [Google Scholar]
- 11.Wang H, Maechler P, Ritz-Laser B, Hagenfeldt KA, Ishihara H, Philippe J, Wollheim CB. Pdx1 level defines pancreatic gene expression pattern and cell lineage differentiation. J Biol Chem. 2001;276(27):25279–25286. doi: 10.1074/jbc.M101233200. [DOI] [PubMed] [Google Scholar]
- 12.Kubo A, Stull R, Takeuchi M, Bonham K, Gouon-Evans V, Sho M, Iwano M, Saito Y, Keller G, Snodgrass R. Pdx1 and Ngn3 Overexpression Enhances Pancreatic Differentiation of Mouse Es Cell-Derived Endoderm Population. PLoS One. 2011;6(9):e24058. doi: 10.1371/journal.pone.0024058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Mehran AE, Templeman NM, Brigidi GS, Lim GE, Chu KY, Hu X, Botezelli JD, Asadi A, Hoffman BG, Kieffer TJ, Bamji SX, Clee SM, Johnson JD. Hyperinsulinemia drives diet-induced obesity independently of brain insulin production. Cell Metab. 2012;16(6):723–737. doi: 10.1016/j.cmet.2012.10.019. [DOI] [PubMed] [Google Scholar]
- 14.Wentworth BM, Rhodes C, Schnetzler B, Gross DJ, Halban PA, Villa-Komaroff L. The ratio of mouse insulin I: insulin II does not reflect that of the corresponding preproinsulin mRNAs. Mol Cell Endocrinol. 1992;86(3):177–186. doi: 10.1016/0303-7207(92)90142-s. [DOI] [PubMed] [Google Scholar]
- 15.Moriyama H, Abiru N, Paronen J, Sikora K, Liu E, Miao D, Devendra D, Beilke J, Gianani R, Gill RG, Eisenbarth GS. Evidence for a primary islet autoantigen (preproinsulin 1) for insulitis and diabetes in the nonobese diabetic mouse. Proc Natl Acad Sci USA. 2003;100(18):10376–10381. doi: 10.1073/pnas.1834450100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Liu S-H, Rao DD, Nemunaitis J, Senzer N, Zhou G, Dawson D, Gingras M-C, Wang Z, Gibbs R, Norman M, Templeton NS, Demayo FJ, O’Malley B, Sanchez R, Fisher WE, Brunicardi FC. PDX-1 is a therapeutic target for pancreatic cancer, insulinoma and islet neoplasia using a novel RNA interference platform. PLoS One. 2012;7(8):e40452. doi: 10.1371/journal.pone.0040452. [DOI] [PMC free article] [PubMed] [Google Scholar]