

Abstract
The discovery of the cannabinoid receptors CB1 and CB2 in 1990 and 1993, respectively, as well as of the two main endocannabinoids, anandamide in 1992 and 2-arachidonylglycerol in 1995, was an important step in identifying the strongest homeostatic system in the human body, namely the endocannabinoid system. Ever since, research has highlighted the crucial part played by this system in all the reproduction stages: folliculogenesis, spermatogenesis, oogenesis, fecundation, transport of the egg through the fallopian tubes, blastocyte implantation and pregnancy progression, as well as its implications in the physiopathology of the reproductive system: in endometriosis, ectopic pregnancy, miscarriage, preeclampsia, endometrial cancer, polycystic ovary syndrome, ovarian cancer. A special attention must be paid to the phytocannabinoids, natural components originating especially from the Cannabis plant inflorescences, whose medical effects are well-established nowadays with also acting on the receptors of the endocannabinoid system. The most recent research mainly focuses on the reproductive dysfunctions and disorders of the reproductive tissues, respectively, through its action upon the endocannabinoid system. Medical cannabis is nowadays legalized in more and more countries all over the world. At the same time, recreational cannabis remains one of the most consumed drugs (in Romania the most consumed one by young adults). Therefore, it is mandatory for specialists in obstetrics and gynecology, endocrinology, public health, hygiene or for general practitioners, to permanently update their information on this subject.
Keywords: endocannabinoid system, medical cannabis, reproductive system, public health and hygiene, family medicine
Introduction
In the last 2-3 decades, 50 countries world widely have legalized or decriminalized the medical use of cannabis. In Europe, 23 countries finalized the process of legalizing the use of medical cannabis, while other countries, among which Romania as well, have this problem on their agenda. At the same time, recreational cannabis remains one of the most used drugs. According to the National Report on Drug Situation in 2020 – New evolution and trends (1) the recreational cannabis consumption, although having the lowest rates in Europe, preserves its ever-growing trend, thus continuing to be the most consumed drug in Romania. The most frequent consumption rate is observed in the young population, of fertile age (15-34 years old), with the same sex ratio male/female 2/1, like in any other age groups. In Romania, one in 10 young adults (18-34 years old) has tried to smoke cannabis (10%), one in 20 smoked in the last year (5.8%) and 1 in 40 declared cannabis consumption when questioned (2.5%) (2).
Consequently, it is mandatory for the medical staff to be updated with as much information as possible regarding the endocannabinoid system, as well as with its impact on the reproductive system.
Therefore, the aim of the present editorial was to bring new insights on the endocannabinoid system, its main receptors, the metabolization pathways as well as its impact in women reproduction.
Short description of the endocannabinoid system in relation with the reproduction
Ever since the 15th Century there has been proof of the medical use of cannabis, during birth (3). Still, for many years, the most known side effects of recreational cannabis use on fertility are: it decreases the LH serum levels (4,5), inhibits prolactin secretion (6) increases testosterone serum level, disturbs menstrual cycle (7), causes a low rate of oocyte collection during the in vitro fertilization (IVF) treatment (8,9), negatively influences intrauterine fetal growth (10,11), increases premature birth incidence (12,13), low birth weight and prematurity (VLBW) (7), as well as difficult child delivery (14,15). Nevertheless, the mechanisms through which these produce their effect have not been clearly understood. The discovery of the cannabinoid receptors CB1 and CB2 in 1990 and 1993, respectively, as well as of the two main endocannabinoids, anandamide in 1992 and 2-arachidonylglycerol in 1995, were also the initial steps in identifying the strongest homeostatic system in the human body, namely the endocannabinoid system (ECS). The decades following the discovery clarified the role played by the ECS in all stages of reproduction: spermatogenesis, oogenesis, fecundation, transport of the egg through the fallopian tubes, blastocyte implantation and pregnancy progression. Also, the role of the ECS in the pathologies of the reproductive system was clarified: endometriosis, ectopic pregnancy, miscarriage, preeclampsia, endometrial cancer, polycystic ovary syndrome, ovarian cancer (16).
The endocannabinoid system (ECS) is defined as a group of receptors, ligands (endocannabinoids), enzymes of their synthesis and degradation and also their transmembrane transporters. Endocannabinoids (eCBs) are fatty acid derived lipids (most often arachidonic acid derivates) synthesized by the human body under the action of some specific enzymes, under a nervous or hormonal impulse, or of the cytokines signaling the cells. The best studied endocannabinoids are, anandamide (AEA) or N- arachidonoylethanolamide and 2-arachidonoylglyerol (2-AG).
The main receptors in the endocannabinoid system are:
- CB1 receptors identified in the brain in 1990 (17); they are mainly found in the central nervous system and the spine, being the main psychoactive receptors, and also in the fatty tissue, liver, pancreas, skeletal muscles and immune cells, including most endocrine organs. They are also found in the ovaries, endometrium and testicles (18,19). Intracellularly, CB1 receptors are found on the external membrane of mitochondria (20). Mitochondria play an important role in the control of cellular apoptosis, their dysfunction affecting gametogenesis, embryogenesis and embryonic stem cell production (21). Mitochondrial dysfunction is also associated with oxidative stress in the placenta, thus leading to a vascular remodeling with an impact on the oxygenation and functioning of fetal organs (22,23).
- CB2 receptors, identified in 1993, are mainly found in the peripheral nervous system and in the immune cells, in the cortex and ovarian medulla area, as well as in the ovarian follicles. CB2 are mainly immunomodulatory and anti-inflammatory receptors (24).
- other identified receptors of ECS are transient receptor potential vanilloid-1 (TRPV1), peroxisome proliferator-activated receptors y (PPARy) and G protein-coupled receptor 18, 55 or 119 (GPR18, GPR55, GPR119) (25).
ECS is involved in multiple physiological processes, like nociception, cognitive function, mood regulation, appetite regulation, lipid metabolism, neurogenesis, neuroprotection, cell growth and proliferation etc. The receptor localization and the ligand type explain the effects of cannabinoids (26). For example, the CB1 receptors in the hypothalamus are involved in appetite regulation, those in the amygdale cause memory effects and emotional reactions, those in the peripheral nerves act upon pain sensation. Anandamide (AEA) has a partially agonist action on CB1 receptor. Endocannabinoid 2-arachidonoylglyerol (2-AG) acts like an agonist for both types of receptors. It should be specified that endocannabinoids act more as neuromodulators in the brain, with highly regulated, spatiotemporal specific patterns, than as neurotransmitters, due to their rapid synthesis, as well as fast degradation (27).
Ligands of the cannabinoid receptors may also be the phytocannabinoids present especially in the cannabis plant, as well as synthetic cannabinoids. The phytocannabinoids found in the cannabis plant are lipophilic terpeno-phenolic compounds, with a pharmacological action similar to that of endocannabinoids, still with a different structure (28).
More enzymes are responsible for the synthesis or catabolism of endocannabinoids, as shown in Figure 1. Endocannabinoids are neurotransmitters produced “on request” by a cell in membrane phospholipids (29).
Figure 1.

Metabolization of endocannabinoids. Adapted after (30).
Fatty acid amide hydrolase (FAAH) is the main enzyme responsible for the anandamide hydrolase into arachidonic acid (AA) and ethanolamine (EA). Anandamide (AEA) is synthesized from N-arachidonoyl-phosphati-dyetanolamine (NAPE) by phospholipase D (PLD) and it is decomposed by fatty acid amide hydrolase (FAAH) and cyclooxygenase 2 (COX-2), and 12-LOX into arachidonic acid, prostaglandin-ethanolamides or prostamides (PM) and 12-hydroxi-AEA (12-HAEA), respectively. NAPE, in its turn, comes from phosphatidylethanolamine under the action of N-acyltransferase (NAT). 2-arachidonoylglycerol (2-AG) is synthesized under the action of diacylglycerol lipase (DAGL) from diacylglycerol, being hydrolase by monoacylglycerol lipase (MAGL), COX-2, 12-LOX into arachidonic acid, prostaglandin glycerol ester (PG) and 12-HETE (12-hydroxyeicosatetraenoic acid) (31). Arachidonic acid is metabolized by COX-2, PGHS-1, PGHS-2 into prostanoids: prostaglandines (PGD2, PGE2, PGF2α), prostacyclin (PGI2) and thromboxane A2 (TxA2). Recently, there were also identified enzymes of the serine hydrolase family as regulators of the intracellular pool of 2-AG (30,32).
ECS involvement in women reproduction
Endocannabinoids, phytocannabinoids and synthetic cannabinoids have an impact during all the reproductive stages in women, from oocyte development to parturition, the cells and tissues of the reproductive organs being equipped with a completely functional ECS (8,32,33). Numerous studies showed the presence of ECS components in the serum, follicular fluid, ovary, uterus, placenta, endometrium, immune cells, as well as in the hypothalamus, having an important impact on fertility, reproduction and endocrine function (34). Apparently, most of them are present in the ovary, oviduct (35) and uterus (36).
ECS is engaged in the central control of the processes involved in reproduction: Gamete’s maturation takes place under the action of hypothalamic-pituitary-ovarian axis (HPO axis). ECS was identified in hypothalamic areas involved in the production of some hormones like gonadotropin releasing hormone (GnRH), acting through the HPO axis for the regulation of various reproductive processes (37). Endocannabinoids, their metabolic enzymes, and target receptors have been shown to respond to endocrine signals (38). The estrogen produced by the ovarian follicles inhibits FAAH, thus resulting high levels of AEA, with a role in follicle development and ovulation, followed by a decrease of FAAH inhibiting after ovulation, due to estrogen level decrease, and AEA degradation (39). The same AEA increases under the influence of estrogens and stimulates CB1 in the hypothalamus, thus leading to the reduction of GnRH release and inhibiting FSH and LH release in the pituitary, gonadal suppression and reduction of estrogen and progesterone levels (40). The levels of endocannabinoids are mildly regulated by the enzymes involved in their synthesis and degradation. High doses of exogenous cannabinoids (phytocannabinoids and synthetic cannabinoids) exert an inhibiting effect on the GnRH release (39).
The presence of CB1 receptors on the mitochondrial membrane allows endo and phytocannabinoids to influence the ovarian function and oogenesis, respectively. Still, it is interesting to observe that, in the ovarian follicles, CB2 receptors prevail comparing to CB1 ones, thus leading to the idea of an immunological part played primarily by the ECS in the ovarian function (19). AEA is produced by the follicular cells so that the concentration correlates with the follicular growth (41). In early stages of folliculogenesis, sustaining a low level of AEA allows the maturation of the oocyte, a maximum level of AEA being reached in the periovulatory phase (32). The AEA action on TRPV1 receptors in the spermatozoon facilitates the fusion with the oocyte (42,43). Multiple studies showed the presence of ECS components in the oviduct, more precisely AEA, NAPE-PLD, CB1, CB2, and FAAH (35,44). The transport through the oviduct of the fertilized egg and its growth into a morula are controlled by an AEA concentration gradient in the oviduct. This increase of the AEA level from the ampulla to the isthmus is maintained high by NAPE-PLD (the enzyme responsible for the synthesis) in the isthmus and low in the ampullar region, where FAAH (the enzyme responsible for degradation) is also high. Simultaneously, the FAAH expression increases in the embryonic cells that need to be protected from the AEA cytotoxicity (18). In the preimplantation embryos, CB1 and CB2 receptors are expressed, thus allowing AEA to influence the embryo growth through their activation (39). The low expression of CB1 receptors in the oviduct and decidua vera is apparently correlated with the ectopic pregnancy (45).
ECS modulates the endometrium plasticity and the uterus receptivity. AEA is considered to be the main factor enabling the synchronization between the embryo development and the endometrium, thus allowing its implantation during the so-called “implantation window” (Fig. 2) (46–51). AEA levels are high in non-receptive uteri and in the interimplantary areas in the receptive uterus, and low in receptive uteri and in implantary areas (low levels of NAPE-PLD and high FAAH activity and expression), thus leading to the idea that a balanced endocannabinoid system is essential for the synchronization between embryo implantation and endometrium preparation for it (37). The trophoblast differentiation is mildly controlled by AEA.
Figure 2.
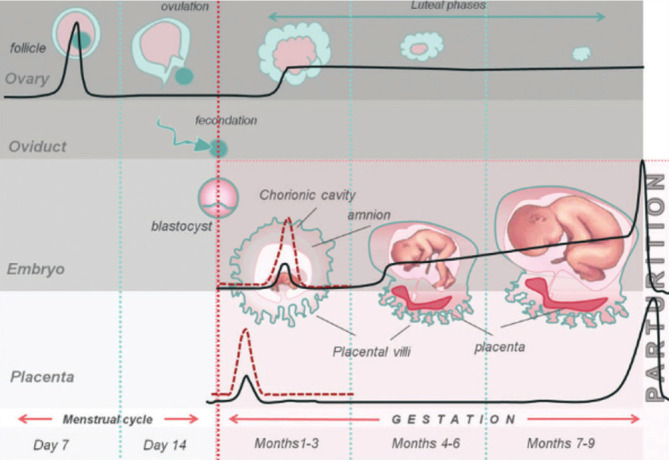
Fluctuation of AEA level during the menstrual cycle and during pregnancy. Under physiological conditions (black bold line), the plasma level of AEA is higher than in the follicular phase and lower than in the luteal phase of the menstrual cycle. In the first stage of pregnancy, for a good uterine receptivity and pregnancy progression, the AEA level should remain low. In the mature placenta and during labor, the AEA level is high. Under pathological conditions, like eclampsia, ectopic or non-viable pregnancies, miscarriage (red dotted line), the AEA level is high (33).
Its survival is favored by the FAAH increase, while this enzyme expression is stimulated by the pro-fertility signaling of leptin, progesterone and cytokines T Helper 2 (Th2) and inhibited by the anti-fertility signaling of cytokines T Helper 1 (Th1) (52). There are studies showing that the plasma and ovary AEA levels have a positive correlation (46). Low FAAH levels and high AEA levels in peripheral blood are associated with a hight risk of miscarriage, or with the failure of in vitro fertilization (53). Placenta formation is accompanied by decrease of CB1 receptors and increase of FAAH, while delivery is accompanied only by CB1 increasing. The AEA plasma level decreases from the first to the second trimester of pregnancy, without any other differences between the AEA plasma levels during the second and third trimesters up to a sudden increase during delivery. Most probably, this increase leads to the prostaglandin level increase through an AEA degradation and arachidonic acid release (54) (Fig. 3).
Figure 3.
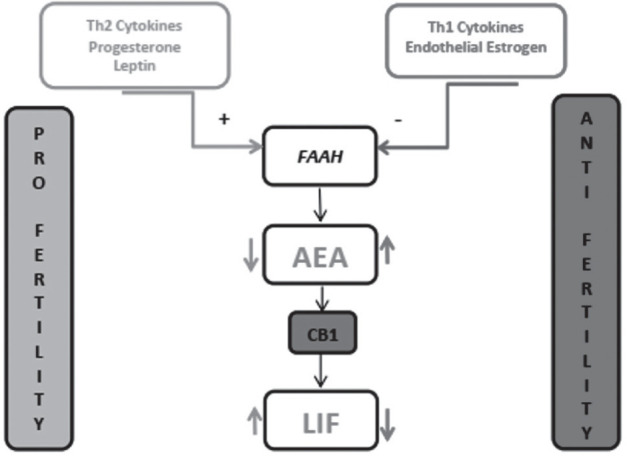
FAAH and AEA as molecular pro- or anti-fertility integrators. Light grey symbolizes the pro-fertility action, while dark grey is used for anti-fertility. Downwards arrow symbolizes the decrease, while the upwards arrow shows increase. Adapted after (33).
There is enough evidence regarding the effect of progesterone and estrogen on the ECS involvement in women fertility. During the menstrual cycle, the progesterone level is well-correlated with FAAH, and also with a the AEA plasma level, with which it has no correlation in early pregnancy (54). Progesterone has the following effects: during the implantation, FAAH decreases and reduces AEA; it plays a part in sustaining pregnancy through the stimulation of the synthesis of pro-fertility cytokines produced by type 2 helper (Th2), inhibiting the anti-fertility ones produced by type 1 helper (Th1) (55); it stimulates the production of leukemia inhibitor factor (LIF) mediated by IL-4, which sustains nidation and pregnancy (56). The estrogen level is positively correlated with the plasmatic AEA level during the menstrual cycle and in non-gestational women after in vitro fertilization and embryo transfer (57). A better predictability of AEA levels is difficult to attain due to the opposite effects of estrogen on NAPE-PLD in various tissues. 17β-Estradiol has the following effects: during folliculogenesis, it stimulates the NAPE-PLD activity and increases the AEA level in the endothelial cells, while, during implantation, it inhibits the FAAH activity and increases the AEA level in the endothelial cells and also down-regulates NAPE-PLD activity and inhibits the FAAH activity in the uterus epithelium (58).
For sustaining pregnancy, AEA low levels and consecutively high levels of leukemia inhibitor factor (LIF) via CB1 receptors are essential (59). LIF is an IL-6 with a role in the proliferation, differentiation and survival of cells, being important in the immune, hematopoiesis and reproduction processes. In humans, the LIF production was detected in the endometrium, reaching a peak during nidation, followed by a decrease towards the end of the luteal phase. The LIF production also exists in the fallopian tubes, being involved in the development and differentiation of the blastocyte. In women with idiopathic infertility or recurrent abortions there was observed a low production of LIF (60).
Also, the immunity adaptation through peripheral T lymphocytes represents a sine-qua-non production for a normal pregnancy. Progesterone induces the pro-fertility cytokines produced by type 2 helper (Th2), involving the CB2 receptor as well. The response of Th1 anti-fertility cytokines is inhibited by Th2 cytokines. AEA acts upon T cells through the CB1 receptor, thus reducing the LIF release in these cells (61).
Another cytokine, leptin, produced by the obese gene (ob) expression in the fatty tissue, ovary and placenta, has an effect on the ECS involvement in pregnancy by stimulating the FAAH activity and consecutive decrease of AEA level (62). Leptin level varies during the menstrual cycle, being lower during the follicular phase than in the secretory one (63). Pregnant women have higher concentrations of serum leptin than non-pregnant. Leptin was identified in the placental syncytiotrophoblast and the endometrium, as well. It plays an important part in reproduction, modulating the growth and development of conception product, with a possible involvement in the angiogenetic processes during early pregnancy (64). Also, it plays a role in the nidation process, the blastocyte presence stimulating its endometrial secretion, and in sustaining pregnancy, with serum levels higher in pregnant women and lower in women undergoing miscarriage during the first trimester (65,66).
Phytocannabinoids mimic the endocannabinoid action, the former being natural components from the inflorescence of Cannabis plant. At present, phytocannabinoids are also attributed medical effects, explained by their action on the human cannabinoid receptors, the medical use of cannabis being completely different from its recreational use. These facts bring back to our attention the Cannabis plant, also explaining the continuous global actions for legalizing medical Cannabis products.
In conclusion, up to the present time, the involvement of endocannabinoids in the regulation of food intake and energy homeostasis of the body, as well as their impact on the endocrine system, including the activity of the pituitary, adrenal cortex, thyroid, pancreas, and gonads (67) were highlighted.
Research performed in the last 20 years showed a major involvement of the ECS in the central and local control of processes related to reproduction. It is well-known the fact that disbalances of the ECS may lead to disorders related to the reproductive system, such as ectopic pregnancy, miscarriage, preeclampsia or conditions caused by the alteration of hormonal homeostasis, like endometriosis or gynecological cancers (45,47,61,68–72).
In the literature, AEA is already proposed as a potential biomarker for pregnancy progression, while the FAAH concentration in peripheral blood is considered a biomarker for infertility (69). Most recent studies focus on the treatment of reproductive dysfunctions, through the manipulation of the endocannabinoid system; nevertheless, there are numerous challenges to overcome in this direction (26), just like it is still difficult to talk about elements of the ECS as biomarkers for various physiological and pathological processes, due to their lack of specificity, with small exceptions, like the heterogeneity and complexity of the ECS.
The connection between the endocannabinoid system and the reproductive system represents an area of great interest that should definitely be known, in the context of legalizing medical use of cannabis in more and more countries all over the world, and whose thorough research may lead to new solutions for women with problems of the reproductive system.
Conflict of interest
The authors declare that they have no conflict of interest.
References
- 1.European Monitoring Center for Drugs and Drug Addiction & ANA Raport National privind situatia drogurilor, 2020, Romania. Noi evolutii si tendinte [Internet] 2020. (Accessed January 04, 2022, at site http://ana.gov.ro/raportul-national-privind-situatia-drogurilor-in-romania-2020/)
- 2.European Monitoring Center for Drugs and Drug Addiction & ANA Raport National privind situatia drogurilor, 2019, Romania. Noi evolutii si tendinte [Internet] 2019. (Accessed January 04, 2022, at site https://www.emcdda.europa.eu/system/files/publications/11364/20191724_TDAT19001RON_PDF.pdf)
- 3.Du Toit BM. Rotterdam: AA Bolkema; 1980. Cannabis in Africa: a survey of its distribution in Africa, and a study of cannabis use and users in multi-ethnic South Africa. [Google Scholar]
- 4.Mendelson JH, Mello NK, Ellingboe J, Skupny AS, Lex BW, Griffin M. Marihuana smoking suppresses luteinizing hormone in women. J Pharmacol Exp Ther. 1986;237(3):862–866. [PubMed] [Google Scholar]
- 5.Mendelson JH, Mello NK. Effects of Marijuana on. Marijuana Eff Endocr Reprod Syst. 1984;44:97. [PubMed] [Google Scholar]
- 6.Mendelson JH, Mello NK, Ellingboe J. Acute effects of marihuana smoking on prolactin levels in human females. J Pharmacol Exp Ther. 1985;232(1):220–222. [PubMed] [Google Scholar]
- 7.Bauman J. Washington: DC US Gov Print Off; 1980. Marijuana and the female reproductive system. [Google Scholar]
- 8.Walker OS, Holloway AC, Raha S. The role of the endocannabinoid system in female reproductive tissues. J Ovarian Res. 2019;12(1):1–10. doi: 10.1186/s13048-018-0478-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Klonoff-Cohen HS, Natarajan L, Chen RV. A prospective study of the effects of female and male marijuana use on in vitro fertilization (IVF) and gamete intrafallopian transfer (GIFT) outcomes. Am J Obstet Gynecol. 2006;194(2):369–376. doi: 10.1016/j.ajog.2005.08.020. [DOI] [PubMed] [Google Scholar]
- 10.Qazi QH, Mariano E, Milman DH, Beller E, Crombleholme W. Abnormalities in offspring associated with prenatal marihuana exposure. Dev Pharmacol Ther. 1985;8:141–148. doi: 10.1159/000457031. [DOI] [PubMed] [Google Scholar]
- 11.Frank DA, Bauchner H, Parker S, Huber AM, Kyei-Aboagye K, Cabral H, Zuckerman B. Neonatal body proportionality and body composition after in utero exposure to cocaine and marijuana. J Pediatr. 1990;117(4):622–626. doi: 10.1016/s0022-3476(05)80702-4. [DOI] [PubMed] [Google Scholar]
- 12.Sherwood RA, Keating J, Kavvadia V, Greenough A, Peters TJ. Substance misuse in early pregnancy and relationship to fetal outcome. Eur J Pediatr. 1999;158(6):488–492. doi: 10.1007/s004310051126. [DOI] [PubMed] [Google Scholar]
- 13.Fried PA, Watkinson B, Willan A. Marijuana use during pregnancy and decreased length of gestation. Am J Obstet Gynecol. 1984;150(1):23–27. doi: 10.1016/s0002-9378(84)80103-9. [DOI] [PubMed] [Google Scholar]
- 14.Greenland S, Staisch KJ, Brown N, Gross SJ. Effects of marijuana on human pregnancy, labor, and delivery. Neurobehav Toxicol Teratol. 1982;4(4):447–450. [PubMed] [Google Scholar]
- 15.Greenland S, Richwald GA, Honda GD. The effects of marijuana use during pregnancy. II. A study in a low-risk home-delivery population. Drug Alcohol Depend. 1983;11(3–4):359–366. doi: 10.1016/0376-8716(83)90026-1. [DOI] [PubMed] [Google Scholar]
- 16.Maia J, Fonseca BM, Teixeira N, Correia-da-Silva G. The fundamental role of the endocannabinoid system in endometrium and placenta: implications in pathophysiological aspects of uterine and pregnancy disorders. Hum Reprod Update. 2020;26(4):586–602. doi: 10.1093/humupd/dmaa005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Trezza V, Cuomo V, Vanderschuren LJMJ. Cannabis and the developing brain: insights from behavior. Eur J Pharmacol. 2008;585(2–3):441–452. doi: 10.1016/j.ejphar.2008.01.058. [DOI] [PubMed] [Google Scholar]
- 18.El-Talatini MR, Taylor AH, Elson JC, Brown L, Davidson AC, Konje JC. Localisation and function of the endocannabinoid system in the human ovary. PLoS One. 2009;4(2):e4579. doi: 10.1371/journal.pone.0004579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Pertwee RG. Pharmacology of cannabinoid CB1 and CB2 receptors. Pharmacol Ther. 1997;74(2):129–180. doi: 10.1016/s0163-7258(97)82001-3. [DOI] [PubMed] [Google Scholar]
- 20.Bénard G, Massa F, Puente N, Lourenço J, Bellocchio L, Soria-Gómez E, Matias I, Delamarre A, Metna-Laurent M, Cannich A, Hebert-Chatelain E, Mulle C, Ortega-Gutiérrez S, Martín-Fontecha M, Klugmann M, Guggenhuber S, Lutz B, Gertsch J, Chaouloff F, López-Rodríguez ML, Grandes P, Rossignol R, Marsicano G. Mitochondrial CB1 receptors regulate neuronal energy metabolism. Nat Neurosci. 2012;15(4):558–564. doi: 10.1038/nn.3053. [DOI] [PubMed] [Google Scholar]
- 21.Ramalho-Santos J, Varum S, Amaral S, Mota PC, Sousa AP, Amaral A. Mitochondrial functionality in reproduction: from gonads and gametes to embryos and embryonic stem cells. Hum Reprod Update. 2009;15(5):553–572. doi: 10.1093/humupd/dmp016. [DOI] [PubMed] [Google Scholar]
- 22.Myatt L, Cui X. Oxidative stress in the placenta. Histochem Cell Biol. 2004;122(4):369–382. doi: 10.1007/s00418-004-0677-x. [DOI] [PubMed] [Google Scholar]
- 23.Abán C, Martinez N, Carou C, Albamonte I, Toro A, Seyahian A, Franchi A, Leguizamón G, Trigubo D, Damiano A, Farina M. Endocannabinoids participate in placental apoptosis induced by hypoxia inducible factor-1. Apoptosis. 2016;21(10):1094–1105. doi: 10.1007/s10495-016-1274-x. [DOI] [PubMed] [Google Scholar]
- 24.Capozzi A, Caissutti D, Mattei V, Gado F, Martellucci S, Longo A, Recalchi S, Manganelli V, Riitano G, Garofalo T, Sorice M, Manera C, Misasi R. Anti-Inflammatory Activity of a CB2 Selective Cannabinoid Receptor Agonist: Signaling and Cytokines Release in Blood Mononuclear Cells. Molecules. 2022;27(1):64. doi: 10.3390/molecules27010064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Fonseca BM, Teixeira NA, Almada M, Taylor AH, Konje JC, Correia-da-Silva G. Modulation of the novel cannabinoid receptor-GPR55-during rat fetoplacental development. Placenta. 2011;32(6):462–469. doi: 10.1016/j.placenta.2011.03.007. [DOI] [PubMed] [Google Scholar]
- 26.Stasiulewicz A, Znajdek K, Grudzień M, Pawiński T, Sulkowska JI. A guide to targeting the endocannabinoid system in drug design. Int J Mol Sci. 2020;21(8):2778. doi: 10.3390/ijms21082778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Katona I, Freund TF. Multiple functions of endocannabinoid signaling in the brain. Annu Rev Neurosci. 2012;35:529–558. doi: 10.1146/annurev-neuro-062111-150420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Pertwee RG, Ross RA. Cannabinoid receptors and their ligands. Prostaglandins. Leukot Essent Fat Acids. 2002;66(2–3):101–121. doi: 10.1054/plef.2001.0341. [DOI] [PubMed] [Google Scholar]
- 29.Pacher P, Kunos G. Modulating the endocannabinoid system in human health and disease–successes and failures. FEBS J. 2013;280(9):1918–1943. doi: 10.1111/febs.12260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Chan H-W, McKirdy NC, Peiris HN, Rice GE, Mitchell MD. The role of endocannabinoids in pregnancy. Reproduction. 2013;146(3):R101–109. doi: 10.1530/REP-12-0508. [DOI] [PubMed] [Google Scholar]
- 31.Keppel Hesselink JM, Kopsky DJ, Sajben NL. Vulvodynia and proctodynia treated with topical baclofen 5% and palmitoylethanolamide. Arch Gynecol Obstet. 2014;290(2):389–393. doi: 10.1007/s00404-014-3218-4. [DOI] [PubMed] [Google Scholar]
- 32.Cecconi S, Rossi G, Castellucci A, D’Andrea G, Maccarrone M. Endocannabinoid signalling in mammalian ovary. Eur J Obstet Gynecol Reprod Biol. 2014;178:6–11. doi: 10.1016/j.ejogrb.2014.04.011. [DOI] [PubMed] [Google Scholar]
- 33.Rapino C, Battista N, Bari M, Maccarrone M. Endocannabinoids as biomarkers of human reproduction. Hum Reprod Update. 2014;20(4):501–516. doi: 10.1093/humupd/dmu004. [DOI] [PubMed] [Google Scholar]
- 34.Schuel H. Tuning the oviduct to the anandamide tone. J Clin Invest. 2006;116(8):2087–2090. doi: 10.1172/JCI29424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wang H, Xie H, Guo Y, Zhang H, Takahashi T, Kingsley PJ, Marnett LJ, Das SK, Cravatt BF, Dey SK. Fatty acid amide hydrolase deficiency limits early pregnancy events. J Clin Invest. 2006;116(8):2122–2131. doi: 10.1172/JCI28621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Scotchie JG, Savaris RF, Martin CE, Young SL. Endocannabinoid regulation in human endometrium across the menstrual cycle. Reprod Sci. 2015;22(1):113–123. doi: 10.1177/1933719114533730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Gammon CM, Freeman GM, Xie W, Petersen SL, Wetsel WC. Regulation of gonadotropin-releasing hormone secretion by cannabinoids. Endocrinology. 2005;146(10):4491–4499. doi: 10.1210/en.2004-1672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Maccarrone M. Endocannabinoids: friends and foes of reproduction. Prog Lipid Res. 2009;48(6):344–354. doi: 10.1016/j.plipres.2009.07.001. [DOI] [PubMed] [Google Scholar]
- 39.Karasu T, Marczylo TH, Maccarrone M, Konje JC. The role of sex steroid hormones, cytokines and the endocannabinoid system in female fertility. Hum Reprod Update. 2011;17(3):347–361. doi: 10.1093/humupd/dmq058. [DOI] [PubMed] [Google Scholar]
- 40.Sun X, Dey SK. Endocannabinoid signaling in female reproduction. ACS Chem Neurosci. 2012;3(5):349–355. doi: 10.1021/cn300014e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Brents LK. Focus: sex and gender health: Marijuana, the Endocannabinoid System and the female reproductive system. Yale J Biol Med. 2016;89(2):175. [PMC free article] [PubMed] [Google Scholar]
- 42.Maccarrone M, Barboni B, Paradisi A, Bernabò N, Gasperi V, Pistilli MG, Fezza F, Lucidi P, Mattioli M. Characterization of the endocannabinoid system in boar spermatozoa and implications for sperm capacitation and acrosome reaction. J Cell Sci. 2005;118(19):4393–4404. doi: 10.1242/jcs.02536. [DOI] [PubMed] [Google Scholar]
- 43.Francavilla F, Battista N, Barbonetti A, Vassallo MR, Rapino C, Antonangelo C, Pasquariello N, Catanzaro G, Barboni B, Maccarrone M. Characterization of the endocannabinoid system in human spermatozoa and involvement of transient receptor potential vanilloid 1 receptor in their fertilizing ability. Endocrinology. 2009;150(10):4692–4700. doi: 10.1210/en.2009-0057. [DOI] [PubMed] [Google Scholar]
- 44.Wang H, Xie H, Dey SK. Endocannabinoid signaling directs periimplantation events. AAPS J. 2006;8(2):E425–432. doi: 10.1007/BF02854916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Horne AW, Phillips JA, 3rd, Kane N, Lourenco PC, McDonald SE, Williams AR, Simon C, Dey SK, Critchley HO. CB1 expression is attenuated in Fallopian tube and decidua of women with ectopic pregnancy. PLoS One. 2008;3(12):e3969. doi: 10.1371/journal.pone.0003969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.El-Talatini MR, Taylor AH, Konje JC. Fluctuation in anandamide levels from ovulation to early pregnancy in in-vitro fertilization-embryo transfer women, and its hormonal regulation. Hum Reprod. 2009;24(8):1989–1998. doi: 10.1093/humrep/dep065. [DOI] [PubMed] [Google Scholar]
- 47.Trabucco E, Acone G, Marenna A, Pierantoni R, Cacciola G, Chioccarelli T, Mackie K, Fasano S, Colacurci N, Meccariello R, Cobellis G, Cobellis L. Endocannabinoid system in first trimester placenta: low FAAH and high CB1 expression characterize spontaneous miscarriage. Placenta. 2009;30(6):516–522. doi: 10.1016/j.placenta.2009.03.015. [DOI] [PubMed] [Google Scholar]
- 48.Taylor AH, Amoako AA, Bambang K, Karasu T, Gebeh A, Lam PM, Marzcylo TH, Konje JC. Endocannabinoids and pregnancy. Clin Chim acta. 2010;411(13–14):921–930. doi: 10.1016/j.cca.2010.03.012. [DOI] [PubMed] [Google Scholar]
- 49.Fonseca BM, Correia-da-Silva G, Taylor AH, Lam PM, Marczylo TH, Konje JC, Teixeira NA. Characterisation of the endocannabinoid system in rat haemochorial placenta. Reprod Toxicol. 2012;34(3):347–356. doi: 10.1016/j.reprotox.2012.05.036. [DOI] [PubMed] [Google Scholar]
- 50.Ding J, Luo X-T, Yao Y-R, Xiao H-M, Guo M-Q. Investigation of changes in endocannabinoids and N-acylethanolamides in biofluids, and their correlations with female infertility. J Chromatogr A. 2017;1509:16–25. doi: 10.1016/j.chroma.2017.06.029. [DOI] [PubMed] [Google Scholar]
- 51.Maia J, Almada M, Silva A, Correia-da-Silva G, Teixeira N, Sá SI, Fonseca BM. The endocannabinoid system expression in the female reproductive tract is modulated by estrogen. J Steroid Biochem Mol Biol. 2017;174:40–47. doi: 10.1016/j.jsbmb.2017.07.023. [DOI] [PubMed] [Google Scholar]
- 52.Sun X, Cappelletti M, Li Y, Karp CL, Divanovic S, Dey SK. Cnr2 deficiency confers resistance to inflammation-induced preterm birth in mice. Endocrinology. 2014;155(10):4006–4014. doi: 10.1210/en.2014-1387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Maccarrone M, Bisogno T, Valensise H, Lazzarin N, Fezza F, Manna C, Di Marzo V, Finazzi-Agrò A. Low fatty acid amide hydrolase and high anandamide levels are associated with failure to achieve an ongoing pregnancy after IVF and embryo transfer. Mol Hum Reprod. 2002;8(2):188–195. doi: 10.1093/molehr/8.2.188. [DOI] [PubMed] [Google Scholar]
- 54.Habayeb OM, Taylor AH, Evans MD, Cooke MS, Taylor DJ, Bell SC, Konje JC. Plasma levels of the endocannabinoid anandamide in women—a potential role in pregnancy maintenance and labor? J Clin Endocrinol Metab. 2004;89(11):5482–5487. doi: 10.1210/jc.2004-0681. [DOI] [PubMed] [Google Scholar]
- 55.Piccinni M-P. T cell tolerance towards the fetal allograft. J Reprod Immunol. 2010;85(1):71–75. doi: 10.1016/j.jri.2010.01.006. [DOI] [PubMed] [Google Scholar]
- 56.Aghajanova L. Leukemia inhibitory factor and human embryo implantation. Ann N Y Acad Sci. 2004;1034(1):176–183. doi: 10.1196/annals.1335.020. [DOI] [PubMed] [Google Scholar]
- 57.Marczylo TH, Lam PMW, Amoako AA, Konje JC. Anandamide levels in human female reproductive tissues: solid-phase extraction and measurement by ultraperformance liquid chromatography tandem mass spectrometry. Anal Biochem. 2010;400(2):155–162. doi: 10.1016/j.ab.2009.12.025. [DOI] [PubMed] [Google Scholar]
- 58.Wang H, Dey SK, Maccarrone M. Jekyll and Hyde: two faces of cannabinoid signaling in male and female fertility. Endocr Rev. 2006;27(5):427–448. doi: 10.1210/er.2006-0006. [DOI] [PubMed] [Google Scholar]
- 59.Habayeb OMH, Taylor AH, Bell SC, Taylor DJ, Konje JC. Expression of the endocannabinoid system in human first trimester placenta and its role in trophoblast proliferation. Endocrinology. 2008;149(10):5052–5060. doi: 10.1210/en.2007-1799. [DOI] [PubMed] [Google Scholar]
- 60.Maccarrone M, Bari M, Battista N, Finazzi-Agrò A. Estrogen stimulates arachidonoylethanolamide release from human endothelial cells and platelet activation. Blood. J Am Soc Hematol. 2002;100(12):4040–4048. doi: 10.1182/blood-2002-05-1444. [DOI] [PubMed] [Google Scholar]
- 61.Shen X, Duan H, Wang S, Hong W, Wang Y-Y, Lin S-L. Expression of cannabinoid receptors in myometrium and its correlation with dysmenorrhea in adenomyosis. Reprod Sci. 2019;26(12):1618–1625. doi: 10.1177/1933719119833483. [DOI] [PubMed] [Google Scholar]
- 62.Brocato B, Zoerner AA, Janjetovic Z, Skobowiat C, Gupta S, Moore BM, 2nd, Slominski A, Zhang J, Schenone M, Phinehas R, Ferry RJ, Jr, Dick E, Jr, Hubbard GB, Mari G, Schlabritz-Loutsevitch N. Endocannabinoid crosstalk between placenta and maternal fat in a baboon model (Papio spp.) of obesity. Placenta. 2013;34(11):983–989. doi: 10.1016/j.placenta.2013.08.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ajala OM, Ogunro PS, Elusanmi GF, Ogunyemi OE, Bolarinde AA. Changes in serum leptin during phases of menstrual cycle of fertile women: relationship to age groups and fertility. Int J Endocrinol Metab. 2013;11(1):27. doi: 10.5812/ijem.6872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Metwally M, Ledger WL, Li TC. Reproductive endocrinology and clinical aspects of obesity in women. Ann N Y Acad Sci. 2008;1127(1):140–146. doi: 10.1196/annals.1434.000. [DOI] [PubMed] [Google Scholar]
- 65.Henson MC, Castracane VD. Leptin in pregnancy: an update. Biol Reprod. 2006;74(2):218–229. doi: 10.1095/biolreprod.105.045120. [DOI] [PubMed] [Google Scholar]
- 66.Linne Y. Effects of obesity on women’s reproduction and complications during pregnancy. Obes Rev. 2004;5(3):137–143. doi: 10.1111/j.1467-789X.2004.00147.x. [DOI] [PubMed] [Google Scholar]
- 67.Borowska M, Czarnywojtek A, Sawicka-Gutaj N, Woliński K, Płazińska MT, Mikołajczak P, Ruchała M. The effects of cannabinoids on the endocrine system. Endokrynol Pol. 2018;69(6):705–719. doi: 10.5603/EP.a2018.0072. [DOI] [PubMed] [Google Scholar]
- 68.Sanchez AM, Cioffi R, Viganò P, Candiani M, Verde R, Piscitelli F, Di Marzo V, Garavaglia E, Panina-Bordignon P. Elevated systemic levels of endocannabinoids and related mediators across the menstrual cycle in women with endometriosis. Reprod Sci. 2016;23(8):1071–1079. doi: 10.1177/1933719116630414. [DOI] [PubMed] [Google Scholar]
- 69.Gebeh AK, Willets JM, Marczylo EL, Taylor AH, Konje JC. Ectopic pregnancy is associated with high anandamide levels and aberrant expression of FAAH and CB1 in fallopian tubes. J Clin Endocrinol Metab. 2012;97(8):2827–2835. doi: 10.1210/jc.2012-1780. [DOI] [PubMed] [Google Scholar]
- 70.Maccarrone M, Valensise H, Bari M, Lazzarin N, Romanini C, Finazzi-Agrò A. Relation between decreased anandamide hydrolase concentrations in human lymphocytes and miscarriage. Lancet. 2000;355(9212):1326–1329. doi: 10.1016/S0140-6736(00)02115-2. [DOI] [PubMed] [Google Scholar]
- 71.Fügedi G, Molnár M, Rigó J, Schönléber J, Kovalszky I, Molvarec A. Increased placental expression of cannabinoid receptor 1 in preeclampsia: an observational study. BMC Pregnancy Childbirth. 2014;14(1):1–8. doi: 10.1186/s12884-014-0395-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Ayakannu T, Taylor AH, Marczylo TH, Willets JM, Brown L, Davies Q. Association of cannabinoid receptor expression with anandamide concentrations in endometrial cancer. Lancet. 2014;383:S23. [Google Scholar]