

Abstract
Coronavirus disease 2019 (Covid-19) is a global diastrophic disease caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). Covid-19 leads to inflammatory, immunological, and oxidative changes, by which SARS-CoV-2 leads to endothelial dysfunction (ED), acute lung injury (ALI), acute respiratory distress syndrome (ARDS), and multi-organ failure (MOF). Despite evidence illustrating that some drugs and vaccines effectively manage and prevent Covid-19, complementary herbal medicines are urgently needed to control this pandemic disease. One of the most used herbal medicines is berberine (BBR), which has anti-inflammatory, antioxidant, antiviral, and immune-regulatory effects; thus, BBR may be a prospective candidate against SARS-CoV-2 infection. This review found that BBR has anti-SARS-CoV-2 effects with mitigation of associated inflammatory changes. BBR also reduces the risk of ALI/ARDS in Covid-19 patients by inhibiting the release of pro-inflammatory cytokines and inflammatory signaling pathways. In conclusion, BBR has potent anti-inflammatory, antioxidant, and antiviral effects. Therefore, it can be utilized as a possible anti-SARS-CoV-2 agent. BBR inhibits the proliferation of SARS-CoV-2 and attenuates the associated inflammatory disorders linked by the activation of inflammatory signaling pathways. Indeed, BBR can alleviate ALI/ARDS in patients with severe Covid-19. In this sense, clinical trials and prospective studies are suggested to illustrate the potential role of BBR in treating Covid-19.
Keywords: Covid-19, Inflammatory signaling pathways, Anti-inflammatory, Antioxidant, Antiviral, Immune-regulatory effects
Background
The current coronavirus disease, SARS-CoV-2, is caused by a novel coronavirus and has global diastrophic effects (Al-Kuraishy et al. 2021a, b, c). Acute respiratory distress syndrome (ARDS) may result from direct SARS-CoV-2 cytopathic damage and the release of pro-inflammatory cytokines (Al-kuraishy et al. 2022). SARS-CoV-2 primarily infects lung alveolar type II pneumocyte cells due to greater expression of angiotensin-converting enzyme 2 (ACE2), a receptor for SARS-CoV-2 binding and entrance (Biswas, Mahmud et al. 2021). Most Covid-19 patients are asymptomatic or have mild respiratory symptoms. However, a small percentage of Covid-19 patients may suffer severe respiratory symptoms because of the development of acute lung injury (ALI) and/or ARDS. SARS-CoV-2 could cause endothelial dysfunction (ED) either directly or indirectly via elevated pro-inflammatory cytokines (Lugnier et al. 2021). Similarly, during SARS-CoV-2 infection, oxidative stress (OS) is generated, leading to ED. Oxidative stress has been found to cause tissue harm during many viral infections, including hepatitis viruses, human immunodeficiency virus-1 (HIV-1), and most RNA viruses, including coronavirus (Ghosh, Mukerjee et al. 2021, Elekhnawy and Negm 2022).
Generally, viral infections increase free radical production and decrease endogenous antioxidant capacity through mitochondrial dysfunction. Covid-19-induced cytokine storms and high levels of pro-inflammatory cytokines have also been linked to the production of reactive oxygen species (ROS) (Al-Kuraishy et al. 2021a, b, c). In addition, pro-inflammatory cytokines activate inducible nitric oxide synthase (iNOS), which produces oxidizing peroxy nitrite by reacting with ROS and superoxide ions (Adedara et al. 2018). Hypoxia, tissue ischemia, hypertension, and diabetes mellitus, all of which are frequently associated with Covid-19, are linked to a decrease in cellular antioxidant capacity (Engwa 2018; Elekhnawy et al. 2022).
In Covid-19 patients with underlying comorbidities, a reduction in antioxidant capacity may promote OS progression (Karkhanei et al. 2021). Despite evidence illustrating that some drugs and vaccines are efficient in managing and preventing Covid-19, complementary herbal medicines are urgently needed for more control of this pandemic disease. One of the most used herbal medicines is berberine (BBR), and Fig. 1 illustrates its chemical structure.
Fig. 1.

Chemical structure of berberine
BBR is quaternary ammonium derived from isoquinoline salts. BBR is an odorless yellow powder with a bitter taste, characterized by high water-soluble with slight solubility in methanol and ethanol (Habtemariam 2020). BBR can easily gain from medicinal plants or by total synthesis. BBR is widely distributed in plants' roots, stems, and rhizomes, including Papaveraceae, Euphorbiaceae, and Ranunculaceae. However, BBR is mainly derived from Rhizoma coptidis, a common Chinese herbal medicine utilized to treat diabetes mellitus (DM) and chronic inflammatory disorders since 2000 A.D (Xu et al. 2017).
Herberger and Buchner early discovered BBR in 1830 (Gao et al. 2020), and Hongjing Tao first reported the beneficial effects of BBR in treating DM (Zhang et al. 2014). Furthermore, Indian researchers in 1960 illustrated that BBR salts are promising in treating severe diarrhea, amoebiasis, and cholera (Subbaiah and Amin 1967).
Pharmacology of berberine
Different studies revealed that BBR has important beneficial effects in managing various cardiovascular, metabolic, hematological, neurological, and immunological diseases [Table 1]. BBR has acute effects in managing DM through increasing insulin secretion, activating glycolysis, and inhibiting adipogenesis mediated by adenosine monophosphate protein kinase (AMPK) pathway glycokinase activity (Cicero and Tartagni 2012). Besides, BBR improves peripheral glucose uptake by increasing the expression of glucose transporter 4 (GLUT-4) (Mi et al. 2019). Likewise, BBR stimulates the release of glucagon-like peptide 1(GLP-1), which through GLP-1, activates AMPK and insulin release (Wang et al. 2021a, b). Indeed, BBR triggers peroxisome proliferator-activated receptor gamma (PPAR-γ), which reduces peripheral insulin resistance (Zhou et al. 2019). Furthermore, BBR has been shown to improve glucose metabolism in the intestine with significant insulin-sensitizing effects (Zhang et al. 2014).
Table 1.
Pharmacological effects of berberine (BBR)
| Ref | Type of the study | Findings |
|---|---|---|
| (Zhang, Ren et al. 2008, Li, Li et al. 2014a, b) |
Experimental study, 40 rats Experimental study |
Administration of 36 mg/kg of BBR effectively attenuated heart failure development BBR prevents the progression of left ventricular hypertrophy caused by pressure overload in rats through inhibition of inflammatory signaling and enhancement of autophagy |
| (Guo, Chen et al. 2021) | Experimental study, 20 rats | BBR reduces consequent complications of heart failure, like cardiac fibrosis, through upregulation of relaxin expression in the cardiomyocytes |
| (Huang et al. 2015) | Experimental study, 20 mice | BBR 40 mg/kg reduces the risk of ischemic-reperfusion injury by direct antioxidant effect, inhibition of post-ischemic inflammation, direct coronary vasodilator effect, anti-apoptotic effect, promoting angiogenesis, and suppression of cardiomyocyte autophagy |
| (Fatehi‐Hassanabad, Jafarzadeh et al. 2005) | Experimental study, 30 rats | BBR 0.4 mg/kg IV has hypotensive effects through different mechanisms, including α-1 adrenoceptor antagonist, enhancement of the hypotensive effect of acetylcholine, inhibition of the angiotensin-converting enzyme, and direct vasodilatation effect through activation of K-channel, Ca + 2-channel blocking effect in the cardiomyocytes |
| (Kong et al. 2004) | In vitro and clinical studies | Oral administration of BBR 1 g/day in 32 hypercholesterolemic patients for 3 months reduced serum cholesterol by 29%, triglycerides by 35%, and LDL cholesterol by 25% |
| (Wei et al. 2016) | Experimental study, 30 mice | Mice with metabolic syndrome treated with BBR 40–60 mg/kg/day for four weeks experienced significant reductions in the metabolic syndrome criteria |
| (Ma et al. 2010) | Experimental study, 20 rats | BBR can effectively treat obesity by modulation lipid dysregulation, weight reduction, and decreasing appetite |
| (Sun et al. 2016) | Experimental study, 30 rats | BBR can reduce body weight by upregulating brain GLP-1, neuropeptide Y, and orexin-A, suppressing the hypothalamic feeding center |
| (Bäckhed et al. 2007) | Experimental study, 20 mice | BBR-induced AMPK pathway is associated with improved blood glucose and oxidation of fatty acids |
On the other hand, BBR has a potential role in managing various cardiovascular diseases and complications. BBR exerts a protective effect against the development of ED and atherosclerosis (Ma, Shi et al. 2020). Of interest, this effect of BBR is mediated by modulation of cellular pro-atherogenic events, reduction of oxidative stress by increasing bioavailability of nitric oxide (NO), and suppression of xanthine oxidase (XO) (Hussienet al. 2019). Therefore, BBR, by regulating NO/ROS, may attenuate oxidative stress-induced ED and atherosclerotic complications. Moreover, BBR attenuates cardiac hypertrophy, remodeling, and heart failure development via mitophagy suppression (Abudureyimu et al. 2020). An experimental study by (Zhang, Ren et al. 2008), revealed that administration of BBR 63 mg/day for 4 weeks reduces the development of heart failure by reducing cardiomyocyte Ca+2 overload in rats (Zhang, Ren et al. 2008). Similarly, BBR prevents the progression of left ventricular hypertrophy caused by pressure overload in rats through inhibition of inflammatory signaling and enhancement of autophagy (Li et al. 2014a, b). Thus, BBR may reduce consequent complications of heart failure like cardiac fibrosis, which is involved in the induction of cardiac arrhythmia. This effect is mediated by the upregulation of relaxin expression in the cardiomyocytes (Gu et al. 2012). As well, BBR reduces the risk of ischemic-reperfusion injury by direct antioxidant effect, inhibition of post-ischemic inflammation, direct coronary vasodilator effect, anti-apoptotic effect, promoting angiogenesis and suppression of cardiomyocyte autophagy (Huang et al. 2015). BBR has hypotensive effects through different mechanisms, including α-1 adrenoceptor antagonist, enhancement of the hypotensive effect of acetylcholine, inhibition of the angiotensin-converting enzyme, and direct vasodilatation effect through activation of K-channel, Ca + 2-channel blocking effect in the cardiomyocytes (Fatehi‐Hassanabad, Jafarzadeh et al. 2005).
Moreover, BBR improves lipid and glucose metabolism and can be used to manage dyslipidemia, so BBR is regarded as a new lipid-lowering agent (Kong et al. 2004). In addition, BBR controls lipid metabolism via the regulation expression of low-density lipoprotein receptors (LDLRs), extracellular signal-regulated kinases (ERKs), and proprotein convertase subtilisin/kexin type 9 (PCSK9) (Cao et al. 2019). The experimental study illustrated that mice with metabolic syndrome treated with BBR 40–60 mg/kg/day for four weeks experienced significant reductions in metabolic syndrome criteria. This effect is mainly mediated through the inhibition of hepatic gluconeogenesis and regulation of lipid metabolism via reduction expression of hepatic miR122 and hepatocyte nuclear factor 4α (Kong et al. 2004; Wei et al. 2016).
Furthermore, BBR can effectively treat obesity by modulation lipid dysregulation, weight reduction, and decreasing appetite by controlling central and peripheral AMPK pathways and serotonin neurotransmission (Ma et al. 2010). BBR can reduce body weight by upregulating brain GLP-1, neuropeptide Y, and orexin-A, suppressing the hypothalamic feeding center (Sun et al. 2016). The anti-obesity effects of BBR are linked with the anti-diabetic effects, as the BBR-induced AMPK pathway is associated with improving blood glucose and oxidation of fatty acids. Prolonged treatment with BBR increases the diversity of gut microbiota. It shifts the bacteria to produce fasting-induced adipose factor, which inhibits lipoprotein lipase activity, a potent regulator of fat mobilization (Bäckhed et al. 2007).
These findings revealed that BBR has wide-spectrum pharmacological effects on the cardiovascular system and substantial modulating effects on the cardio-metabolic profile (Fig. 2).
Fig. 2.

Pharmacological effects of berberine(BBR)
Anti-inflammatory and antioxidant effects of berberine
BBR has an essential role in controlling different autoimmune disorders due to its anti-inflammatory and immunomodulatory effects (Adeyemi, Afolabi et al. 2022). Herbal medicines, particularly BBR, can potentially target inflammatory processes in DM, cancer, and neurological disorders with anti-inflammatory, immunomodulatory, and antioxidant effects. BBR inhibits the inflammatory process by suppressing the release of tumor necrosis factor-alpha (TNF-α) and IL-6, matrix metalloproteinases (MMPs), and monocyte chemoattractant protein 1(MCP-1). BBR also suppresses the expression of cyclooxygenase-2 (COX-2), nuclear factor kappa B (NF-κB), and mitogen-activated protein kinase (MAPK) (Chen et al. 2008). Based on these findings, BBR might be an effective agent in managing autoimmune diseases and associated inflammatory disorders (Li, Li et al. 2014a, b).
It has been reported that the abnormal autoreactive response of T helper cells is linked with abnormal production of IL-17 and interferon-gamma (INF-γ) connected with the progression and severity of autoimmune diseases (Liu et al. 2016). Indeed, Th1 and Th2 also recruit other inflammatory cells into injured and inflamed tissues by producing pro-inflammatory cytokines. Various studies showed that BBR could inhibit experimental nephritis by inhibiting proliferation and differentiation of CD4 T cells and down-regulation of Th1. Therefore, in experimental studies, BBR regulates Th1/Th17 immune response with subsequent modulation of INF-γ/Th17 cytokines. The inhibitory effects of BBR on Th1/Th17 cell differentiation are mediated by suppression of transducer and activator of transcription (STAT-3) and Janus kinase (JAK) signaling pathways (Li, Li et al. 2014a, b).
On the other hand, regulatory T cells (Treg) are anti-inflammatory cells that secret anti-inflammatory cytokines such as IL-10, IL-35, IL-10, and tumor growth factor-beta (TGF-β), thereby inhibiting Th1/Th17 immune response and counterbalance of the inflammatory response (Li et al. 2020a, b). Treg cells are often defective in autoimmune disorders and viral infections (Shimokawa et al. 2020; Wan et al. 2020). BBR improves Treg/Th17 imbalance and attenuates inflammatory response in autoimmune diseases (Shimokawa et al. 2020). Several studies demonstrated that BBR could shift the differentiation of naïve T cells into Treg instead of Th17 cells (Dinesh and Rasool 2019).
In addition, activated macrophages produce several pro-inflammatory cytokines, essential for T cells differentiation into Th1 and Th17 cells (Amoah et al. 2015). Yan et al. found that BBR reduces macrophage activity and releases pro-inflammatory cytokines in experimental colitis mice (Yan, Wang et al. 2012). Similarly, BBR suppresses colonic macrophage infiltration by promoting macrophage apoptosis and inhibition of MAPK and NF-κB, which are responsible for the activation of macrophages and the release of pro-inflammatory cytokines. The macrophages are differentiated into two types during the immunological response, classical M1 and alternative M2 phenotypes. Classical M1 produces pro-inflammatory cytokines, while alternative M2 produces anti-inflammatory cytokines (Mantovani et al. 2013). It has been shown that BBR increases the polarization of macrophages toward alternative M2 phenotype rather than classical M1 phenotype through inhibition of TLR4 and myeloid differentiation primary response 88 (MyD88) (Gong et al. 2019).
Additionally, BBR has potent anti-allergic effects by inhibiting IgE generation, release, and interaction. Therefore, BBR is effective against different allergic diseases like asthma (Yang, Wang et al. 2014). Immunotherapeutic BBR nanomedicine was developed to overcome the low oral bioavailability of BBR. It has been reported that immunotherapeutic BBR has 98–100% protection against peanut-induced severe anaphylaxis in mice through inhibition of histamine from mast cells with induction release of INF-γ (Li, Liu et al. 2022). The net anti-inflammatory effect of BBR is illustrated in (Fig. 3).
Fig. 3.

Anti-inflammatory and immunological effects of berberine
BBR has potent antioxidant activity by increasing the antioxidant superoxide dismutase (SOD) enzyme and the AMPK pathway. BBR can also increase mitochondrial uncoupling protein 2 (UCP2), which uncouples and separates ATP from oxidative phosphorylation through modulation of mitochondrial proton outflow (Xu, Shen et al. 2021). BBR attenuates ED by regulating NO/ROS balance in rats with erectile dysfunction (Alorabi et al. 2022). Pei et al. showed that BBR was adept at decreasing oxidized low-density lipoprotein (ox-LDL) and ox-LDL-induced monocyte stimulation by inhibiting the activity of intercellular adhesion molecule 1 (ICAM) and vascular cell adhesion molecule 1 (VCAM-1) (Pei et al. 2019). BBR inhibits vascular inflammation by inhibiting angiotensin II (AngII)-induced monocytes adhesion to endothelial cells with suppression of expression of MCP-1 and reduction of ROS. Besides, BBR inhibits ROS production by suppressing xanthine oxidase (XO) activity in endothelial cells (Ivanov et al. 2006). Indeed, BBR can scavenge hydroxyl radicals in a dose and concentration-dependent manner. The scavenging ability of BBR is increased significantly at 300–1000 µg/mL, close to the effect of Vitamin C (Aoxue and Yijun 2011). Liu et al. demonstrated that BBR derivatives also have antioxidant activity through augmentation of body antioxidant capacity. From these findings, BBR has antioxidant activities that can modulate inflammatory and oxidative stress in different inflammatory and metabolic disorders (Fig. 4).
Fig. 4.
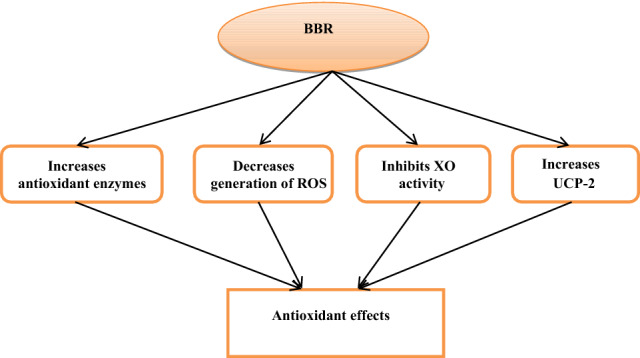
Antioxidant effects of berberine
Antiviral effects of berberine
Different empirical evidence showed that alkaloids from BBR have potent antiviral activity against various viruses [Table 2], including herpes, influenza, respiratory syncytial virus (RSV), and coronaviruses (Warowicka et al. 2020). Hung and colleagues suggested that BBR may inhibit the hepatitis C virus (HCV) entry by targeting viral glycoprotein E2, which is responsible for HCV entry (Hung et al. 2019). Molecular docking confirmed that BBR interacts with HCV glycoprotein E2, so it prevents binding and entry of HCV (Cecil et al. 2011). In addition, BBR displays antiviral effects against dengue and Zika viruses by blocking the activity of NS5 and NS3 of dengue and Zika viruses, respectively (Srivastava 2018). These verdicts indicated that BBR is regarded as a novel viral entry inhibitor, mainly against dengue and Zika viruses.
Table 2.
Antiviral effects of berberine (BBR)
| Ref | Type of the study | Findings |
|---|---|---|
| (Warowicka et al. 2020) | Review study | BBR inhibits replications of DNA and RNA viruses |
| (Hung et al. 2019) | In vitro study | BBR may inhibit the hepatitis C virus (HCV) entry by targeting viral glycoprotein E2, which is responsible for HCV entry |
| (Srivastava 2018) | In silico study | BBR has antiviral effects against dengue and Zika viruses by blocking the activity of NS5 and NS3 of dengue and Zika viruses, respectively |
| (Suryavanshi and Kulkarni 2017) | In vitro study | BBR inhibits RNA synthesis and protein assembly and release of SARS-CoV virions. BBR attenuates SARS-CoV-induced NF-κB activation and release of pro-inflammatory cytokines |
| (Kim et al. 2016) | In vitro study | BBR inhibits the pathogenesis of the Venezuelan equine encephalitis virus (VEEV) and associated inflammatory reactions |
| (Mahata et al. 2011) | In vitro study | BBR reduces the proliferation of human papillomavirus (HPV), mainly HPV 16 and HPV18, by inhibiting viral E2, E6, and E7 oncoproteins |
| (Luganini et al. 2019) | In vitro study | BBR inhibits the replication of herpes viruses by suppressing DNA-dependent RNA polymerase |
| (Wang et al. 2018) | In vitro study | BBR blocks the replication of picorona viruses like enterovirus 71 and associated inflammatory reactions |
| (Zha et al. 2010) | In vitro study | Using 20 µg of BBR resulted in 94% suppression of the reverse transcriptase enzyme in HIV |
Furthermore, BBR is effectual against positive sense, enveloped RNA viruses like severe acute respiratory syndrome coronavirus (SARS-CoV) (Marchant et al. 2010). BBR inhibits RNA synthesis and protein assembly and release of SARS-CoV virions. BBR attenuates SARS-CoV-induced NF-κB activation and release of pro-inflammatory cytokines (Kim et al. 2008; Suryavanshi and Kulkarni 2017). Similarly, BBR reduces the replication of RSV (a negative sense, non-segmental RNA virus) by inhibiting TLR4/ p38MAPK, which is triggered in the early stage of RSV infection. Recently, Tew et al. illustrated that BBR inhibits abnormal immunological reactions in RSV infection through suppression release of IL-6 and NF-κB/ERK/p38MAPK axis (Tew et al. 2020).
Indeed, BBR has potent antiviral effects against the influenza virus, mainly against H1N1, by inhibiting the neuraminidase cytopathogenic effect and associated inflammatory disorders induced by activated NF-κB/TLR4 (Kim et al. 2016). The inhibitory effects of BBR on the TLR4 pathway may limit intestinal mucosal injury during H1N1 (Li et al. 2015). The anti-inflammatory effects of BBR during viral infections are related to the suppression of activated protein 1 (AP-1) and NF-κB signaling pathways. NF-κB and AP-1 are the key transcription factor for regulating gene expression linked with inflammation (Zou et al. 2017). Also, BBR can inhibit MAPK, TLR7 expression, and the release of pro-inflammatory cytokines (Yan et al. 2018).
Furthermore, BBR can inhibit the pathogenesis of the Venezuelan equine encephalitis virus (VEEV) and associated inflammatory reactions by inhibiting signaling pathways and pro-inflammatory cytokines (Kim et al. 2016). Certainly, BBR reduces the proliferation of human papillomavirus (HPV), mainly HPV 16 and HPV18, by inhibiting viral E2, E6, and E7 oncoproteins and host AP-1 (Mahata et al. 2011). BBR inhibits the replication of herpes viruses by suppressing DNA-dependent RNA polymerase and expression of protein E2 with significant down-regulation of the herpes gene (Luganini et al. 2019). Likewise, BBR blocks the replication of corona viruses like enterovirus 71 and links inflammatory reactions by inhibiting autophagy and the Akt signaling pathway (Wang et al. 2018).
BBR and its derivatives have potential effects against human immune deficiency 1(HIV-1) replication and linked inflammatory disorders through inhibition of reverse transcriptase enzyme and modulation expression of IL-6 and ERK signaling. Zha et al. observed that using 20 µg of BBR resulted in 94% suppression of the reverse transcriptase enzyme (Zha et al. 2010). Therefore, BBR has a wide spectrum of antiviral effects through viral entry and proliferation inhibition and suppression of associated inflammatory reactions (Fig. 5).
Fig. 5.
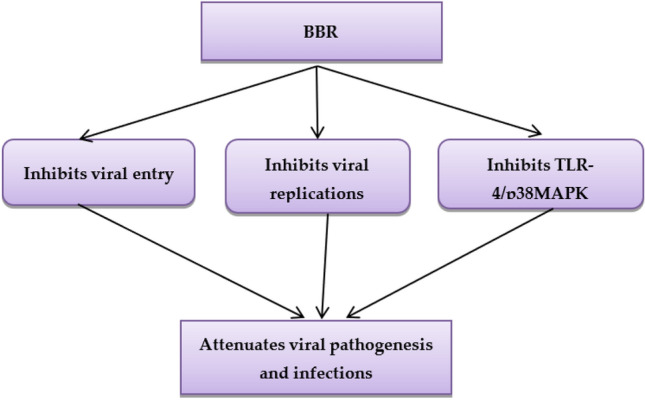
Antiviral effects of berberine
Antiviral mechanisms of berberine
Viral infection can result in cellular changes and induction of inflammatory signaling pathways, including MAPK and NF-κB pathways (Zha et al. 2010). Song et al. illustrated that BBR could block virus-induced activation of the NF-κB pathway with a triggering effect on the IκBα, an endogenous inhibitor of NF-κB (Song et al. 2014). Likewise, BBR down-regulates MAPK, which is involved in various viruses' replication and life cycle. MAPK is mainly activated by HCV, Epstein-Barr virus (EBV), RSV, and SARS-CoV-2 infection (Al-Kuraishy et al. 2021a, b, c), leading to noteworthy inflammatory changes via alteration of protein phosphorylation. HIV-1 protein 6 is phosphorylated by MAPK, resulting in the maturation and release of viral particles (Hemonnot et al. 2004; Attallah et al. 2021). Similarly, the chikungunya virus activates MAPK, which is necessary to release progeny virions. Chikungunya virus non-structural protein 2 (nsP2) activates p38 and JNK in host macrophages (Nayak et al. 2019). It has been reported that BBR reduces the pathogenesis of Chikungunya virus infection by inhibiting p38 and JNK, which are concerned with autophagy and the maturation of viral particles (Varghese et al. 2016). Furthermore, different in vitro studies observed that BBR inhibits hepatitis B virus (HBV) infection by inhibiting p38MAPK, which is necessary to maintain this infection. In addition, BBR, through inhibition of p38MAPK, reduces the proliferation and infectivity of HBV infection (Kim et al. 2017). Thus, p38MAPK inhibitors like biphenyl amide might be a potential target in treating HBV infection, inhibiting HBsAg secretion (Kim et al. 2017).
Moreover, BBR can attenuate the severity of viral infection by inhibiting autophagy, an important mechanism engaged with viral replication. Huang et al. observed that EV71 induces autophagy during its replication (Huang et al. 2009). BBR suppresses virus-induced autophagy by inhibiting viral RNA and protein synthesis with inhibition of JNK/MER/ERK signaling, which engages with the activation of autophagy (Jin et al. 2017).
Taken together, BBR has potential activity against a broad spectrum of viral infections via inhibition of viral binding and entry with a significant reduction of associated inflammatory disorders. From these findings, BBR could be a novel herbal medicine in managing the recent Covid-19 pandemic. The net antiviral mechanism of BBR is illustrated in Fig. 6.
Fig. 6.
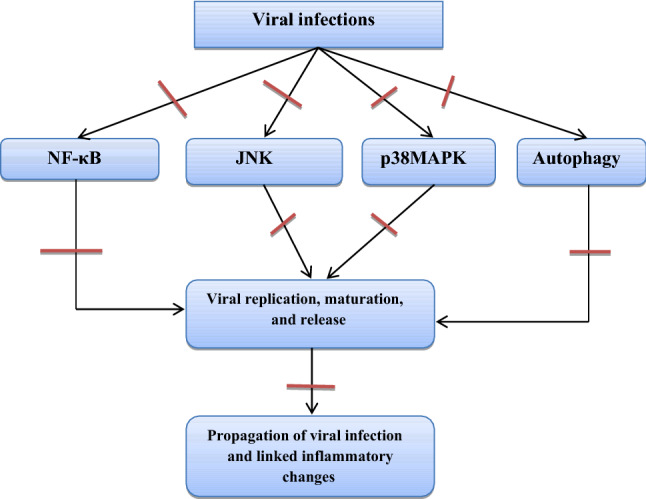
Antiviral mechanisms of berberine
The potential role of berberine in Covid-19
The potential antiviral and anti-inflammatory properties of BBR make it a possible and novel candidate against SARS-CoV-2 infection (Wang et al. 2021a, b). SARS-CoV-2 infection is linked with the development of hyper-inflammation and hypercytokinemia due to the higher release of pro-inflammatory cytokines. The pathway for this inflammatory condition has been confirmed to be mediated by activation and upregulation of the NF-κB signaling pathway (Al-Kuraishy et al. 2021a, b, c). A prospective study of 48 Covid-19 patients revealed that the NF-κB gene and TLR4 are upregulated along with hyper-inflammation (Sohn, Lee et al. 2020). NF-κB is normally sequestered in the cytoplasm in an inactive form by an endogenous inhibitory protein IκB. Phosphorylation with subsequent inhibition of IκB by inflammation induces activation and release of NF-κB, a hallmark of inflammation (Liu et al. 2017; Alotaibi et al. 2021). BBR has potent inhibitory effects on NF-κB with activation of IκB (Song et al. 2014). Thus, BBR may inhibit NF-κB during SARS-CoV-2 infection and reduce the risk of hyper-inflammation and cytokine storm-induced multi-organ failure (MOF) in patients with severe Covid-19 (Wang et al. 2021a, b).
In Covid-19, SARS-CoV-2 directly activates TLR4, which increases the expression of ACE2 in lung alveolar type II (Aboudounya and Heads 2021). Therefore, activated TLR4 is involved in the pathogenesis of SARS-CoV-2 and associated hyper-inflammation via stimulation of innate immune response. So, TLR4 seems to be a promising target in Covid-19 and using TLR4 antagonists may mitigate SARS-CoV-2-induced ALI/ARDS. BBR inhibits TLR4 in different viral infections (Xu et al. 2017), so it may attenuate SARS-CoV-2-mediated hyper-inflammation and development of ALI/ARDS in Covid-19. Liu et al. illustrated that BBR has protective effects against radiation-induced ALI by inhibiting TGFβ-1 and releasing ICAM-1 (Liu et al. 2008). Besides, BBR, through its anti-inflammatory mechanism, reduces the risk of cigarette smoking-mediated ALI (Lin et al. 2013). These findings pointed out the pulmoprotective of BBR and may minimize the risk of ALI in Covid-19 patients.
Indeed, nod-like receptor pyrin 3 (NLRP3) inflammasome is highly activated by SARS-CoV-2, with subsequent release of IL-6 and IL-1β. NLRP3 inflammasome is either triggered by SARS-CoV-2 or damage-associated molecule patterns (DAMPs) from injured cells. NLRP3 inflammasome is also activated by TLR4 signaling and releases TNF-α. Stimulated NLRP3 inflammasome in Covid-19 and released IL-6 increase the risk for cytokine storm development and MOF progression (Meng et al. 2019; Freeman and Swartz 2020). It has been shown that BBR inhibits NLRP3 inflammasome and pyroptosis in non-alcoholic fatty liver disease (NAFLD) and palmitate-induced NLRP3 inflammasome activation (Zhou et al. 2017; Mai et al. 2020). Thus, BBR, inhibiting NLRP3 inflammasome, may reduce Covid-19 severity.
In addition, p38MAPK is a pro-inflammatory pathway concerned with ALI, and myocardial injury is triggered during SARS-CoV-2 infection due to the down-regulation of ACE2 and elevated AngII. Activation of p38MAPK may occur through direct activation by SARS-CoV-2, resulting in the development of ED, vasoconstriction, and thrombosis (Grimes and Grimes 2020). Sharif-Askari et al. illustrated that SARS-CoV-2 infection reduces corticosteroid sensitivity by stimulating the p38MAPK signaling pathway (Sharif-Askari et al. 2021a, b). BBR inhibits the p38MAPK pro-inflammatory signaling pathway in certain viral infections (Kim et al. 2017), thereby reducing the propagation of associated inflammatory reactions. Cui et al. experimental study revealed that BBR attenuates activation of the p38MAPK pathway in mice with type 1 diabetes mellitus (Cui et al. 2009).
Furthermore, SARS-CoV-2 infection may lead to ED and thrombosis via activation of the p38MAPK pro-inflammatory signaling pathway (Aid, Busman-Sahay et al. 2020). A multicenter prospective cohort study involving 150 Covid-19 from a French tertiary hospital revealed that ARDS in Covid-19 is due to the development of thrombosis. However, despite anticoagulant use, many Covid-19 patients with ARDS develop critical thrombotic complications. Therefore, higher doses of anticoagulants are properly suggested. It has been reported that BBR may prevent thrombosis and assist thrombolysis by streptokinase and urokinase (Aid, Busman-Sahay et al. 2020). A recent in vitro study demonstrated that BBR and its derivative 13-Cys-BBR inhibit inflammation-induced thrombosis (Li et al. 2020a, b). Gao et al. observed that BBR decreases lipopolysaccharide (LPS)-induced activation of tissue factor procoagulant activity (Gao et al. 2014). While Wang et al. showed that BBR directs thrombin inhibitory activity (Wang et al. 2017). These verdicts indicate that BBR may reduce the risk of thrombosis in Covid-19.
In the bargain, SARS-CoV-2 infection increases the risk of thrombosis via augmentation of platelet activation Al-Kuraishy et al., 2022b). SARS-CoV-2 spike protein binds platelet ACE2, and through activation of p38MAPK, SARS-CoV-2 enhances platelet aggregation with subsequent release of coagulation factors and induction of thrombosis (Zhang et al. 2020) It has been demonstrated that BBR has the anti-platelet effect and can reduce platelet hyper-aggregability in healthy subjects and patients with ischemic stroke via modulation of aldose reductase and NADPH oxidase pathways (Paul et al. 2019). In vitro study revealed that BBR has the ability to inhibit platelet aggregations by inhibiting platelet Ca + 2 ionophore, adenosine diphosphate (ADP), and arachidonic acid. BBR blocks α2-adrenoceptors on the platelet membrane, inhibiting ADP release from activated platelets (Huang et al. 2002). In addition, BBR inhibits platelet activations by decreasing P-selectin and thromboxane A2 levels with attenuating fibrinogen binding to platelet glycoprotein receptor (GP IIb/IIIa) and collagen-induced platelet stimulation (Allijn et al. 2016). Chiang and colleagues illustrated that SARS-CoV-2 infection-induced thrombosis is related to the activation of platelets thromboxane A2 and P-selectin (Chiang, Reddy et al. 2020). So, BBR could reduce platelet activation and the risk of thrombosis in Covid-19. Zhang et al. revealed that BBR, in combination with ligustrazine, reduces the risk of coronary micro-embolization through inhibition of platelet activation, von Willebrand factor, endothelin-1 (ET-1), and ICAM-1(Zhang et al. 2016). Both ICAM-1 and ET-1 are activated during SARS-CoV-2 infection leading to endothelial injury and thrombosis (Sanghavi et al. 2021). In addition, BBR reduces the risk of pulmonary hypertension and pulmonary micro-thrombosis through modulation of the NO signaling pathway (Qiu et al. 2017). These results and findings support the potential role of BBR in preventing and treating SARS-CoV-2-induced thrombosis in critically ill Covid-19 patients.
Furthermore, SARS-CoV-2-induced inflammatory disorders are linked with the activation of COX and LOX inflammatory pathways (Archambault, Zaid et al. 2020). Fatty acid metabolites and activity of both COX and LOX are increased in the lung of Covid-19 patients compared to the healthy controls leading to a lipid storm (Archambault, Zaid et al. 2020). Therefore, inhibition of COX and LOX pathways in Covid-19 may reduce the risk for the development of cytokine and lipid storms (Al-Kuraishy et al. 2022). Recently, BBR inhibits both COX and LOX inflammatory pathways by inhibiting the activity of phospholipase A2 (Jenny and B Kumar 2021). BBR could be a potential candidate against SARS-CoV-2-induced ALI, which is mediated by the activation of COX and LOX inflammatory pathways. Besides, histamine and histamine receptors are upregulated during SARS-CoV-2 infection and linked with the development of cytokine storms (Malone et al. 2021). Therefore, H2-blockers like famotidine potently manage Covid-19 by mitigating histamine-mediated inflammation (Ennis and Tiligada 2021). Fu et al. observed that BBR inhibits mast cells and histamine release by inhibiting MAPK signaling pathway (Fu et al. 2019). Therefore, BBR could play an important role in mitigating inflammatory disorders in Covid-19 patients via regulating mast cells and histamine release.
Interestingly, autophagy and STAT-3 pathways are activated during Covid-19 leading to apoptosis and hyper-inflammation induction through triggering the release of Th1/Th17 cytokines (Fakhri, Nouri et al. 2020). Autophagy is induced by SARS-CoV-2, while the STAT-3 pathway is activated either directly by SARS-CoV-2 or indirectly by associated pro-inflammatory cytokines. The activated STAT-3 pathway promotes lymphopenia, thrombosis, ED, and lung fibrosis (Jafarzadeh et al. 2021). BBR attenuates both autophagy and STAT-3 pathways in different viral infections (Jin et al. 2017). BBR inhibits methotrexate-induced intestinal inflammation by blocking the JAK/STAT-3 signaling pathway (Hassanein et al. 2021).
Remarkably, severe SARS-CoV-2 infection may induce oxidative stress (OS) through ROS generation and the reduction of endogenous antioxidant capacity (Suhail et al. 2020). High AngII due to down-regulation of ACE2 by SARS-CoV-2 may trigger ROS generation and OS induction, which is linked with the pathogenesis of SARS-CoV-2 infection and Covid-19 severity and mortality (Cecchini and Cecchini 2020). It has been shown that SARS-CoV-2 provokes the expression of oxidative genes in a higher manner than other respiratory viruses. Therefore, biomarkers of OS like S100A8 and S100A9 are increased in patients with severe Covid-19 (Sharif-Askari et al. 2021a, b). In this sense, BBR is regarded as potent antioxidant herbal medicine in different OS-related diseases. BBR inhibits ROS generation and increases body antioxidant capacity by activating the AMPK pathway (Eissa et al. 2018). Besides, BBR reduces the severity of OS by inhibiting the xanthine oxidase (XO) enzyme, which is linked with the generation of ROS in diabetic patients (Pagliaro et al. 2015). XO is upregulated during SARS-CoV-2 infection and might explain the induction of OS in Covid-19, so inhibition of XO by allopurinol may reduce Covid-19 severity (Pratomo, Ariane et al. 2021). Therefore, BBR, by blocking XO activity, may reduce OS injury and Covid-19 severity.
Moreover, the advance glycation end-product and receptor (AGE–RAGE) exerts multiple intracellular signaling via PKC, NADPH oxidase, and MAPK, resulting in the activation of NF-κB (Serveaux-Dancer, Jabaudon et al. 2019). An AGE–RAGE signaling pathway is activated during SARS-CoV-2 infection; however, soluble RAGE (sRAGE) has a protective role against Covid-19 severity (Yalcin Kehribar et al. 2021). A study involving 23 Covid-19 patients compared to 35 healthy controls illustrated a high sRAGE level in asymptomatic Covid-19 patients (Yalcin Kehribar et al. 2021). The study revealed that BBR therapy reduces AGEs in diabetic patients (Zhang et al. 2019). Thus, BBR, modulation of the AGE–RAGE signaling pathway, may reduce Covid-19 severity.
Overall, BBR may have a protective effect against Covid-19 and related complications. A prospective study involved 18 Covid-19 patients on BBR, and standard treatment compared with 17 Covid-19 controls treated by standard treatment only. This study illustrated that Covid-19 patients on BBR treatment experience low IL-6, TNF-α, and CRP, suggesting the anti-inflammatory role of BBR in Covid-19. Besides, BBR reduces gastrointestinal inflammation and the release of pro-inflammatory cytokines from the injured mucosal epithelium in Covid-19 (Zhang et al. 2021). The optimal BBR dose for anti-inflammation related to SARS-CoV-2 remains unclear, although we based our dose of BBR 900 mg/daily on the effective anti-inflammation dose according to other literature (Zhang et al. 2021). BBR has recently been produced in nanomolecular form with greater oral availability and proposed as Covid-19 therapy. An orally available immunotherapeutic-berberine nanomedicine, named NIT-X, has been developed and has shown significantly increased oral bioavailability of BBR, increased IFN-γ production by CD8 + T cells, and inhibition of mast cell histamine release in vivo, suggesting a protective immune response.
Moreover, the expression of targets, including ACE2, TMPRSS2, IL-1α, IL-8, IL-6, and CCL-2 in SARS-CoV-2 infected cells, were significantly suppressed by NIT-X. By supporting protective immunity while inhibiting pro-inflammatory cytokines, inhibiting viral infection and replication, inducing apoptosis, and protecting against tissue damage, BBR is a promising candidate for preventing and treating Covid-19 and SARS. Given the high oral bioavailability and safety of BBR nanomedicine, it may lead to the development of BBR as an orally active therapeutic against Covid-19 and SARS (Wang et al., 2021a, b).
Many clinical trials have been conducted that suggest a wide range of therapeutic applications for BBR. These clinical trials confirmed that using BBR in a dose of 0.6–1.6 g/day did not produce any toxic effects. The effective dose of BBR was 1–2 g/day for different diseases, mainly inflammatory ones (Imenshahidi and Hosseinzadeh 2019, Guo, Chen et al. 2021). Depending on these findings, 1.5 g/day could be effective and safe in Covid-19 patients.
BBR has been shown to inhibit SARS-CoV-2 infectivity by interfering with the binding of SARS-CoV-2 to the ACE2 rather than inhibiting ACE2-driven viral entry (Wu et al. 2020). Similarly, a molecular docking study demonstrated that BBR could inhibit host transmembrane protein serine-type 2 (TMPRSS2), which is implicated in activating SARS-CoV-2 spike protein (Wang et al. 2021a, b). As well, BBR has been proven to block SARS-CoV-2 RNA-dependent RNA polymerase (RdRp), three chymotrypsin protease (3CLpro), and main protease (Mpro); suggesting BBR may inhibit the proliferation of SARS-CoV-2 (Narkhede et al. 2020). These findings indicate that BBR may decrease SARS-CoV-2 pathogenicity by inhibiting viral entry via suppression of ACE2 and TMPRSS2. In addition, BBR may lessen SARS-CoV-2 infectivity by inhibiting the proliferation of SARS-CoV-2. The potential role of BBR in Covid-19 is related to improving the immune response in the early phase of SARS-CoV-2 infection, with suppression of SARS-CoV-2-induced hyper-inflammation in the late phase (Wang et al. 2021a, b). Therefore, BBR could be a preventive and therapeutic agent in Covid-19 by targeting inflammatory signaling pathways with suppression of entry/infectivity of SARS-CoV-2 (Fig. 7). Moreover, BBR can be combined with other anti-SARS-CoV-2 drugs like remdesivir (Pizzorno et al. 2020). The mechanism of action of BBR in the treatment of Covid-19 pneumonia and pulmonary fibrosis is through TNF-α inhibited inflammatory reaction and reduced the activation of fibroblasts. Also, BBR inhibited the synergistic effect between IL-6 and STAT-3, and reduced the inflammatory response. Lastly, BBR inhibits the chemotaxis of CCL-2 to fibroblasts and reduces inflammation (Cao et al. 2019). Further clinical trials and prospective studies are recommended in this field to confirm the potential therapeutic effects of BBR in managing Covid-19.
Fig. 7.
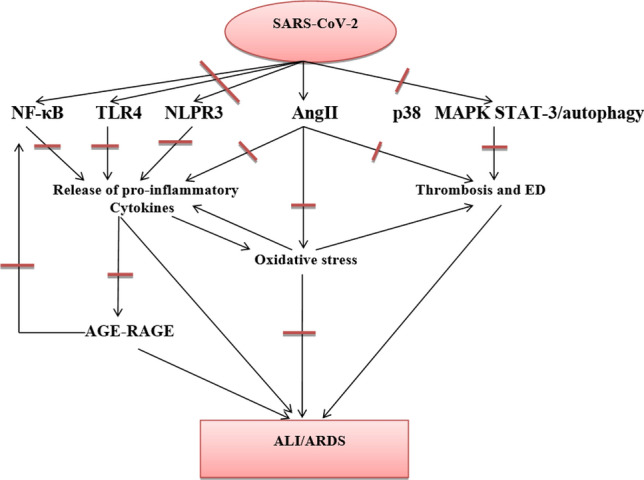
Anti-SARS-CoV-2 mechanisms of berberine
Conclusion
BBR has potent anti-inflammatory, antioxidant, and antiviral effects. Therefore, it can be used as a possible anti-SARS-CoV-2 agent. BBR inhibits the proliferation of SARS-CoV-2 and attenuates the associated inflammatory disorders linked by the activation of inflammatory signaling pathways. BBR has the ability to inhibit the release of pro-inflammatory cytokines through the inhibition of NF-κB and p38MAPK signaling pathways, which are highly activated during SARS-CoV-2 infection. Indeed, BBR can mitigate the risk of ALI/ARDS in patients with severe Covid-19. In this sense, clinical trials and prospective studies are suggested to illustrate the potential role of BBR in treating Covid-19.
Acknowledgements
M.D.W. thanks the French Agence Nationale de la Recherche and the Région Pays de la Loire for financial support on Covid-19 research (ANR Flash COVID 19 call—name: CoV2-E-TARGET—grant number: 2020 07132).
Author contributions
Writing—review and editing; All. All authors have read and agreed to the published version of the manuscript.
Data availability
The authors confirm that the data supporting this study are available within the article.
Declarations
Conflict of interest
The authors declare no conflict of interest.
Informed consent
Not applicable.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Ahmad O. Babalghith, Email: aobabalghith@uqu.edu.sa
Hayder M. Al-kuraishy, Email: Hayderm36@yahoo.com
Ali I. Al-Gareeb, Email: Dr.alialgareeb78@yahoo.com
Michel De Waard, Email: michel.dewaard@univ-nantes.fr.
Sabatier Jean-Marc, Email: sabatier.jm1@gmail.com.
Walaa A. Negm, Email: walaa.negm@pharm.tanta.edu.eg
Gaber El-Saber Batiha, Email: gaberbatiha@gmail.com, Email: dr_gaber_batiha@vetmed.dmu.edu.eg.
References
- Aboudounya MM, Heads RJ. COVID-19 and toll-like receptor 4 (TLR4): SARS-CoV-2 may bind and activate TLR4 to increase ACE2 expression, facilitating entry and causing hyperinflammation. Mediat Inflamm. 2021;2021:1–18. doi: 10.1155/2021/8874339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abudureyimu M, Yu W, Cao RY, Zhang Y, Liu H, Zheng H. Berberine promotes cardiac function by upregulating PINK1/Parkin-mediated mitophagy in heart failure. Front Physiol. 2020;11:565751. doi: 10.3389/fphys.2020.565751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adedara IA, Anao OO, Forcados GE, Awogbindin IO, Agbowo A, Ola-Davies OE, Patlolla AK, Tchounwou PB, Farombi EO. Low doses of multi-walled carbon nanotubes elicit hepatotoxicity in rats with markers of oxidative stress and induction of pro-inflammatory cytokines. Biochem Biophys Res Commun. 2018;503(4):3167–3173. doi: 10.1016/j.bbrc.2018.08.112. [DOI] [PubMed] [Google Scholar]
- Adeyemi OS, Afolabi LB, Rotimi DE, Atanu FO, Ofume DS, Ogunleye T, Oluwalana DO, Oluwole TP, Youssef A, Mostafa-Hedeab G. Differential immunomodulatory potential of silver nanoparticles and effect on the kynurenine pathway in male wistar rats. J Nanomater. 2022;2022:1–9. doi: 10.1155/2022/9883142. [DOI] [Google Scholar]
- Aid M, Busman-Sahay K, Vidal SJ, Maliga Z, Bondoc S, Starke C, Terry M, Jacobson CA, Wrijil L, Ducat S. Vascular disease and thrombosis in SARS-CoV-2-infected rhesus macaques. Cell. 2020;183(5):1354–1366. doi: 10.1016/j.cell.2020.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Al-Kuraishy HM, Al-Gareeb AI, Alkazmi L, Alexiou A, Batiha GE-S. Levamisole therapy in COVID-19. Viral Immunol. 2021;34(10):722–725. doi: 10.1089/vim.2021.0042. [DOI] [PubMed] [Google Scholar]
- Al-Kuraishy HM, Al-Gareeb AI, Alqarni M, Cruz-Martins N, El-Saber Batiha G. Pleiotropic effects of tetracyclines in the management of COVID-19: emerging perspectives. Front Pharmacol. 2021;12:642822. doi: 10.3389/fphar.2021.642822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Al-Kuraishy HM, Al-Gareeb AI, Alzahrani KJ, Cruz-Martins N, Batiha GE-S. The potential role of neopterin in Covid-19: a new perspective. Mol Cell Biochem. 2021;476(11):4161–4166. doi: 10.1007/s11010-021-04232-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Al-kuraishy HM, Al-Gareeb AI, Negm WA, Alexiou A, Batiha GE-S. Ursolic acid and SARS-CoV-2 infection: a new horizon and perspective. Inflammopharmacology. 2022;30:1493–1501. doi: 10.1007/s10787-022-01038-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allijn IE, Vaessen SF, Quarles van Ufford LC, Beukelman KJ, De Winther MP, Storm G, Schiffelers RM. Head-to-head comparison of anti-inflammatory performance of known natural products in vitro. PLoS One. 2016;11(5):e0155325. doi: 10.1371/journal.pone.0155325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alorabi M, Cavalu S, Al-Kuraishy HM, Al-Gareeb AI, Mostafa-Hedeab G, Negm WA, Youssef A, El-Kadem AH, Saad HM, Batiha GE-S. Pentoxifylline and berberine mitigate diclofenac-induced acute nephrotoxicity in male rats via modulation of inflammation and oxidative stress. Biomed Pharmacother. 2022;152:113225. doi: 10.1016/j.biopha.2022.113225. [DOI] [PubMed] [Google Scholar]
- Alotaibi B, Negm WA, Elekhnawy E, El-Masry TA, Elseady WS, Saleh A, Alotaibi KN, El-Sherbeni SA. Antibacterial, Immunomodulatory, and Lung Protective Effects of Boswelliadalzielii Oleoresin Ethanol Extract in Pulmonary Diseases. In Vitro and In Vivo Studies. Antibiotics. 2021;10(12):1444. doi: 10.3390/antibiotics10121444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amoah BP, Yang H, Zhang P, Su Z, Xu H. Immunopathogenesis of myocarditis: the interplay between cardiac fibroblast cells, dendritic cells, macrophages and CD 4+ T cells. Scand J Immunol. 2015;82(1):1–9. doi: 10.1111/sji.12298. [DOI] [PubMed] [Google Scholar]
- Aoxue L, Yijun F. Antioxidant activities of berberine hydrochloride. J Med Plants Res. 2011;5(16):3702–3707. [Google Scholar]
- Archambault A-S, Zaid Y, Rakotoarivelo V, Doré É, Dubuc I, Martin C, Amar Y, Cheikh A, Fares H, El Hassani A. Lipid storm within the lungs of severe COVID-19 patients: extensive levels of cyclooxygenase and lipoxygenase-derived inflammatory metabolites. MedRxiv. 2020;11:1708. [Google Scholar]
- Attallah NG, El-Kadem AH, Negm WA, Elekhnawy E, El-Masry TA, Elmongy EI, Altwaijry N, Alanazi AS, Al-Hamoud GA, Ragab AE. Promising antiviral activity of agrimonia pilosa phytochemicals against severe acute respiratory syndrome Coronavirus 2 Supported with In Vivo Mice Study. Pharmaceuticals. 2021;14(12):1313. doi: 10.3390/ph14121313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bäckhed F, Manchester JK, Semenkovich CF, Gordon JI. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc Natl Acad Sci. 2007;104(3):979–984. doi: 10.1073/pnas.0605374104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biswas, S., S. Mahmud, M. A. Mita, S. Afrose, M. R. Hasan, M. S. S. Shimu, M. A. Saleh, G. Mostafa-Hedeab, M. Alqarni and A. J. Obaidullah (2021). molecular docking and dynamics studies to explore effective inhibitory peptides against the spike receptor binding domain of SARS-CoV-2. Frontiers in Molecular Biosciences 8 [DOI] [PMC free article] [PubMed]
- Cao S, Xu P, Yan J, Liu H, Liu L, Cheng L, Qiu F, Kang N. Berberrubine and its analog, hydroxypropyl-berberrubine, regulate LDLR and PCSK9 expression via the ERK signal pathway to exert cholesterol-lowering effects in human hepatoma HepG2 cells. J Cell Biochem. 2019;120(2):1340–1349. doi: 10.1002/jcb.27102. [DOI] [PubMed] [Google Scholar]
- Cecchini R, Cecchini AL. SARS-CoV-2 infection pathogenesis is related to oxidative stress as a response to aggression. Med Hypotheses. 2020;143:110102. doi: 10.1016/j.mehy.2020.110102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cecil CE, Davis JM, Cech NB, Laster SM. Inhibition of H1N1 influenza a virus growth and induction of inflammatory mediators by the isoquinoline alkaloid berberine and extracts of goldenseal (Hydrastis canadensis) Int Immunopharmacol. 2011;11(11):1706–1714. doi: 10.1016/j.intimp.2011.06.002. [DOI] [PubMed] [Google Scholar]
- Chen F, Yang Z, Liu Y, Li L, Liang W, Wang X, Zhou W, Yang Y, Hu R-M. Berberine inhibits the expression of TNFα, MCP-1, and IL-6 in AcLDL-stimulated macrophages through PPARγ pathway. Endocrine. 2008;33(3):331–337. doi: 10.1007/s12020-008-9089-3. [DOI] [PubMed] [Google Scholar]
- Chiang K, Reddy S, Gupta A. Are Aspirin and Apixaban Sufficient to Prevent Immunothrombosis in COVID-19? SSRN. 2020 doi: 10.2139/ssrn.3755292. [DOI] [Google Scholar]
- Cicero AF, Tartagni E. Antidiabetic properties of berberine: from cellular pharmacology to clinical effects. Hosp Pract. 2012;40(2):56–63. doi: 10.3810/hp.2012.04.970. [DOI] [PubMed] [Google Scholar]
- Cui G, Qin X, Zhang Y, Gong Z, Ge B, Zang YQ. Berberine differentially modulates the activities of ERK, p38 MAPK, and JNK to suppress Th17 and Th1 T cell differentiation in type 1 diabetic mice. J Biol Chem. 2009;284(41):28420–28429. doi: 10.1074/jbc.M109.012674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dinesh P, Rasool M. Berberine mitigates IL-21/IL-21R mediated autophagic influx in fibroblast-like synoviocytes and regulates Th17/Treg imbalance in rheumatoid arthritis. Apoptosis. 2019;24(7):644–661. doi: 10.1007/s10495-019-01548-6. [DOI] [PubMed] [Google Scholar]
- Eissa LA, Kenawy HI, El-Karef A, Elsherbiny NM, El-Mihi KA. Antioxidant and anti-inflammatory activities of berberine attenuate hepatic fibrosis induced by thioacetamide injection in rats. Chem Biol Interact. 2018;294:91–100. doi: 10.1016/j.cbi.2018.08.016. [DOI] [PubMed] [Google Scholar]
- Elekhnawy E, Negm WA. The potential application of probiotics for the prevention and treatment of COVID-19. Egypt J Med Human Genet. 2022;23(1):1–9. doi: 10.1186/s43042-022-00252-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elekhnawy E, Negm WA, El-Aasr M, Kamer AA, Alqarni M, Batiha GE-S, Obaidullah AJ, Fawzy HM. Histological assessment, anti-quorum sensing, and anti-biofilm activities of Dioon spinulosum extract: in vitro and in vivo approach. Sci Rep. 2022;12(1):1–15. doi: 10.1038/s41598-021-03953-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Engwa GA. Free radicals and the role of plant phytochemicals as antioxidants against oxidative stress-related diseases. Phytochem Source Antioxid Role Dis Prev BoDBooks Demand. 2018;7:49–74. [Google Scholar]
- Ennis M, Tiligada K. Histamine receptors and COVID-19. Inflamm Res. 2021;70(1):67–75. doi: 10.1007/s00011-020-01422-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fakhri S, Nouri Z, Moradi SZ, Farzaei MH. Astaxanthin, COVID-19 and immune response: focus on oxidative stress, apoptosis and autophagy. Phytother Res. 2020;34(2790):2792. doi: 10.1002/ptr.6797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fatehi-Hassanabad Z, Jafarzadeh M, Tarhini A, Fatehi M. The antihypertensive and vasodilator effects of aqueous extract from Berberis vulgaris fruit on hypertensive rats. Phytother Res Int J Devoted Pharmacol Toxicol Eval Nat Product Deriv. 2005;19(3):222–225. doi: 10.1002/ptr.1661. [DOI] [PubMed] [Google Scholar]
- Freeman TL, Swartz TH. Targeting the NLRP3 inflammasome in severe COVID-19. Front Immunol. 2020;11:1518. doi: 10.3389/fimmu.2020.01518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fu S, Ni S, Wang D, Fu M, Hong T. Berberine suppresses mast cell-mediated allergic responses via regulating FcɛRI-mediated and MAPK signaling. Int Immunopharmacol. 2019;71:1–6. doi: 10.1016/j.intimp.2019.02.041. [DOI] [PubMed] [Google Scholar]
- Gao M-Y, Chen L, Yang L, Yu X, Kou J-P, Yu B-Y. Berberine inhibits LPS-induced TF procoagulant activity and expression through NF-κB/p65, Akt and MAPK pathway in THP-1 cells. Pharmacol Rep. 2014;66(3):480–484. doi: 10.1016/j.pharep.2013.12.004. [DOI] [PubMed] [Google Scholar]
- Gao Y, Wang F, Song Y, Liu H. The status of and trends in the pharmacology of berberine: a bibliometric review [1985–2018] Chinese Medicine. 2020;15(1):1–13. doi: 10.1186/s13020-020-0288-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghosh, A., N. Mukerjee, B. Sharma, A. Pant, Y. K. Mohanta, R. D. Jawarkar, R. L. Bakal, E. M. Terefe, G. E.-S. Batiha and G. Mostafa-Hedeab (2021). Target Specific inhibition of Protein Tyrosine Kinase in conjunction with Cancer and SARS-COV-2 by Olive nutraceuticals. Frontiers in pharmacology 12. [DOI] [PMC free article] [PubMed]
- Gong J, Li J, Dong H, Chen G, Qin X, Hu M, Yuan F, Fang K, Wang D, Jiang S. Inhibitory effects of berberine on proinflammatory M1 macrophage polarization through interfering with the interaction between TLR4 and MyD88. BMC Complement Altern Med. 2019;19(1):1–12. doi: 10.1186/s12906-019-2710-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grimes JM, Grimes KV. p38 MAPK inhibition: A promising therapeutic approach for COVID-19. J Mol Cell Cardiol. 2020;144:63–65. doi: 10.1016/j.yjmcc.2020.05.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gu H-P, Lin S, Xu M, Yu H-Y, Du X-J, Zhang Y-Y, Yuan G, Gao W. Up-regulating relaxin expression by G-quadruplex interactive ligand to achieve antifibrotic action. Endocrinology. 2012;153(8):3692–3700. doi: 10.1210/en.2012-1114. [DOI] [PubMed] [Google Scholar]
- Guo J, Chen H, Zhang X, Lou W, Zhang P, Qiu Y, Zhang C, Wang Y, Liu WJ. The effect of berberine on metabolic profiles in type 2 diabetic patients: a systematic review and meta-analysis of randomized controlled trials. Oxid Med Cell Longev. 2021;2021:1–22. doi: 10.1155/2021/2074610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Habtemariam S. Berberine pharmacology and the gut microbiota: a hidden therapeutic link. Pharmacol Res. 2020;155:104722. doi: 10.1016/j.phrs.2020.104722. [DOI] [PubMed] [Google Scholar]
- Hassanein EH, Kamel EO, Ali FE, Ahmed MA-R. Berberine and/or zinc protect against methotrexate-induced intestinal damage: Role of GSK-3β/NRF2 and JAK1/STAT-3 signaling pathways. Life Sci. 2021;281:119754. doi: 10.1016/j.lfs.2021.119754. [DOI] [PubMed] [Google Scholar]
- Hemonnot B, Cartier C, Gay B, Rebuffat S, Bardy M, Devaux C, Boyer V, Briant L. The host cell MAP kinase ERK-2 regulates viral assembly and release by phosphorylating the p6gag protein of HIV-1. J Biol Chem. 2004;279(31):32426–32434. doi: 10.1074/jbc.M313137200. [DOI] [PubMed] [Google Scholar]
- Huang CG, Chu ZL, Wei SJ, Jiang H, Jiao BH. Effect of berberine on arachidonic acid metabolism in rabbit platelets and endothelial cells. Thromb Res. 2002;106(4–5):223–227. doi: 10.1016/S0049-3848(02)00133-0. [DOI] [PubMed] [Google Scholar]
- Huang SC, Chang CL, Wang PS, Tsai Y, Liu HS. Enterovirus 71-induced autophagy detected in vitro and in vivo promotes viral replication. J Med Virol. 2009;81(7):1241–1252. doi: 10.1002/jmv.21502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang Z, Han Z, Ye B, Dai Z, Shan P, Lu Z, Dai K, Wang C, Huang W. Berberine alleviates cardiac ischemia/reperfusion injury by inhibiting excessive autophagy in cardiomyocytes. Eur J Pharmacol. 2015;762:1–10. doi: 10.1016/j.ejphar.2015.05.028. [DOI] [PubMed] [Google Scholar]
- Hung T-C, Jassey A, Liu C-H, Lin C-J, Lin C-C, Wong SH, Wang JY, Yen M-H, Lin L-T. Berberine inhibits hepatitis C virus entry by targeting the viral E2 glycoprotein. Phytomedicine. 2019;53:62–69. doi: 10.1016/j.phymed.2018.09.025. [DOI] [PubMed] [Google Scholar]
- Hussien NR, Al-Kuraishy HM, Al-Gareeb AI (2019) Reno-protective effect of berberine. J Pak Med Assoc 69 [PubMed]
- Imenshahidi M, Hosseinzadeh H. Berberine and barberry (Berberis vulgaris): a clinical review. Phytother Res. 2019;33(3):504–523. doi: 10.1002/ptr.6252. [DOI] [PubMed] [Google Scholar]
- Ivanov II, McKenzie BS, Zhou L, Tadokoro CE, Lepelley A, Lafaille JJ, Cua DJ, Littman DR. The orphan nuclear receptor RORγt directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell. 2006;126(6):1121–1133. doi: 10.1016/j.cell.2006.07.035. [DOI] [PubMed] [Google Scholar]
- Jafarzadeh A, Nemati M, Jafarzadeh S. Contribution of STAT3 to the pathogenesis of COVID-19. Microb Pathog. 2021;154:104836. doi: 10.1016/j.micpath.2021.104836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jenny J, Kumar PB. Dihydroxy berberine from Tinospora cordifolia: in silico evidences for the mechanism of anti-inflammatory action through dual inhibition of Lipoxygenase and Cyclooxygenase. Indian J Biochem Biophys (IJBB) 2021;58(3):244–252. [Google Scholar]
- Jin Y, Zhang C, Zhang R, Ren J, Chen S, Sui M, Zhou G, Dang D, Zhu J, Feng H. Pulmonary edema following central nervous system lesions induced by a non-mouse-adapted EV71 strain in neonatal BALB/c mice. Virol J. 2017;14(1):1–12. doi: 10.1186/s12985-017-0911-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karkhanei B, Ghane ET, Mehri F. Evaluation of oxidative stress level: total antioxidant capacity, total oxidant status and glutathione activity in patients with Covid-19. New Microbes New Infect. 2021;42:100897. doi: 10.1016/j.nmni.2021.100897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim H-Y, Shin H-S, Park H, Kim Y-C, Yun YG, Park S, Shin H-J, Kim K. In vitro inhibition of coronavirus replications by the traditionally used medicinal herbal extracts, Cimicifuga rhizoma, Meliae cortex, Coptidis rhizoma, and Phellodendron cortex. J Clin Virol. 2008;41(2):122–128. doi: 10.1016/j.jcv.2007.10.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim J-H, Weeratunga P, Kim MS, Nikapitiya C, Lee B-H, Uddin MB, Kim T-H, Yoon J-E, Park C, Ma JY. Inhibitory effects of an aqueous extract from Cortex Phellodendri on the growth and replication of broad-spectrum of viruses in vitro and in vivo. BMC Complement Altern Med. 2016;16(1):1–13. doi: 10.1186/s12906-016-1206-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim S-Y, Kim H, Kim S-W, Lee N-R, Yi C-M, Heo J, Kim B-J, Kim N-J, Inn K-S. An effective antiviral approach targeting hepatitis B virus with NJK14047, a novel and selective biphenyl amide p38 mitogen-activated protein kinase inhibitor. Antimicrob Agents Chemother. 2017;61(8):e00214–00217. doi: 10.1128/AAC.00214-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kong W, Wei J, Abidi P, Lin M, Inaba S, Li C, Wang Y, Wang Z, Si S, Pan H. Berberine is a novel cholesterol-lowering drug working through a unique mechanism distinct from statins. Nat Med. 2004;10(12):1344–1351. doi: 10.1038/nm1135. [DOI] [PubMed] [Google Scholar]
- Li H, Li XL, Zhang M, Xu H, Wang CC, Wang S, Duan RS. Berberine ameliorates experimental autoimmune neuritis by suppressing both cellular and humoral immunity. Scand J Immunol. 2014;79(1):12–19. doi: 10.1111/sji.12123. [DOI] [PubMed] [Google Scholar]
- Li M-H, Zhang Y-J, Yu Y-H, Yang S-H, Iqbal J, Mi Q-Y, Li B, Wang Z-M, Mao W-X, Xie H-G. Berberine improves pressure overload-induced cardiac hypertrophy and dysfunction through enhanced autophagy. Eur J Pharmacol. 2014;728:67–76. doi: 10.1016/j.ejphar.2014.01.061. [DOI] [PubMed] [Google Scholar]
- Li G-X, Wang X-M, Jiang T, Gong J-F, Niu L-Y, Li N. Berberine prevents intestinal mucosal barrier damage during early phase of sepsis in rat through the toll-like receptors signaling pathway. Korean J Physiol Pharmacol off J Korean Physiol Soc Korean Soc Pharmacol. 2015;19(1):1. doi: 10.4196/kjpp.2015.19.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li C, Jiang P, Wei S, Xu X, Wang J. Regulatory T cells in tumor microenvironment: new mechanisms, potential therapeutic strategies and future prospects. Mol Cancer. 2020;19(1):1–23. doi: 10.1186/s12943-019-1085-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li G, Wang T, Zhang X, Zhao S, Wang Y, Wu J, Peng S, Zhao M. Development of 13-Cys-BBR as an agent having dual action of anti-thrombosis and anti-inflammation. Drug Des Dev Ther. 2020;14:2187. doi: 10.2147/DDDT.S249228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, X.-M., C. Liu, K. Srivatava, H. Yu and S. N. Parker (2022). Treatment of immunological disease using berberine nanoparticles, Google Patents.
- Lin K, Liu S, Shen Y, Li Q. Berberine attenuates cigarette smoke-induced acute lung inflammation. Inflammation. 2013;36(5):1079–1086. doi: 10.1007/s10753-013-9640-0. [DOI] [PubMed] [Google Scholar]
- Liu Y, Yu H, Zhang C, Cheng Y, Hu L, Meng X, Zhao Y. Protective effects of berberine on radiation-induced lung injury via intercellular adhesion molecular-1 and transforming growth factor-beta-1 in patients with lung cancer. Eur J Cancer. 2008;44(16):2425–2432. doi: 10.1016/j.ejca.2008.07.040. [DOI] [PubMed] [Google Scholar]
- Liu X, Zhang X, Ye L, Yuan H. Protective mechanisms of berberine against experimental autoimmune myocarditis in a rat model. Biomed Pharmacother. 2016;79:222–230. doi: 10.1016/j.biopha.2016.02.015. [DOI] [PubMed] [Google Scholar]
- Liu T, Zhang L, Joo D, Sun S-C. NF-κB signaling in inflammation. Signal Transduct Target Ther. 2017;2(1):1–9. doi: 10.1038/sigtrans.2017.23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luganini A, Mercorelli B, Messa L, Palù G, Gribaudo G, Loregian A. The isoquinoline alkaloid berberine inhibits human cytomegalovirus replication by interfering with the viral Immediate Early-2 (IE2) protein transactivating activity. Antiviral Res. 2019;164:52–60. doi: 10.1016/j.antiviral.2019.02.006. [DOI] [PubMed] [Google Scholar]
- Lugnier C, Al-Kuraishy HM, Rousseau E. PDE4 inhibition as a therapeutic strategy for improvement of pulmonary dysfunctions in Covid-19 and cigarette smoking. Biochem Pharmacol. 2021;185:114431. doi: 10.1016/j.bcp.2021.114431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma X, Egawa T, Kimura H, Karaike K, Masuda S, Iwanaka N, Hayashi T. Berberine-induced activation of 5′-adenosine monophosphate–activated protein kinase and glucose transport in rat skeletal muscles. Metabolism. 2010;59(11):1619–1627. doi: 10.1016/j.metabol.2010.03.009. [DOI] [PubMed] [Google Scholar]
- Ma, C.-Y., X.-Y. Shi, Y.-R. Wu, Y. Zhang, H.-L. Qu, Y.-L. Guo, Y.-D. Tang and J.-J. Li (2020). "Berberine Improves High-Fat Diet Induced Atherosclerosis and Hepatic Steatosis in Apoe-/-Mice by Down-Regulating PCSK9 via ERK1/2 Pathway
- Mahata S, Bharti AC, Shukla S, Tyagi A, Husain SA, Das BC. Berberine modulates AP-1 activity to suppress HPV transcription and downstream signaling to induce growth arrest and apoptosis in cervical cancer cells. Mol Cancer. 2011;10(1):1–14. doi: 10.1186/1476-4598-10-39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mai W, Xu Y, Xu J, Zhao D, Ye L, Yu G, Wang Z, Lu Q, Lin J, Yang T. Berberine inhibits nod-like receptor family pyrin domain containing 3 inflammasome activation and pyroptosis in nonalcoholic steatohepatitis via the ROS/TXNIP axis. Front Pharmacol. 2020;11:185. doi: 10.3389/fphar.2020.00185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malone RW, Tisdall P, Fremont-Smith P, Liu Y, Huang X-P, White KM, Miorin L, Moreno E, Alon A, Delaforge E. COVID-19: famotidine, histamine, mast cells, and mechanisms. Front Pharmacol. 2021;12:633680. doi: 10.3389/fphar.2021.633680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mantovani A, Biswas SK, Galdiero MR, Sica A, Locati M. Macrophage plasticity and polarization in tissue repair and remodelling. J Pathol. 2013;229(2):176–185. doi: 10.1002/path.4133. [DOI] [PubMed] [Google Scholar]
- Marchant D, Singhera GK, Utokaparch S, Hackett TL, Boyd JH, Luo Z, Si X, Dorscheid DR, McManus BM, Hegele RG. Toll-like receptor 4-mediated activation of p38 mitogen-activated protein kinase is a determinant of respiratory virus entry and tropism. J Virol. 2010;84(21):11359–11373. doi: 10.1128/JVI.00804-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meng Y, Pan M, Zheng B, Chen Y, Li W, Yang Q, Zheng Z, Sun N, Zhang Y, Li X. Autophagy attenuates angiotensin II-induced pulmonary fibrosis by inhibiting redox imbalance-mediated NOD-like receptor family pyrin domain containing 3 inflammasome activation. Antioxid Redox Signal. 2019;30(4):520–541. doi: 10.1089/ars.2017.7261. [DOI] [PubMed] [Google Scholar]
- Mi J, He W, Lv J, Zhuang K, Huang H, Quan S. Effect of berberine on the HPA-axis pathway and skeletal muscle GLUT4 in type 2 diabetes mellitus rats. Diabetes Metab Syndr Obes Targets Ther. 2019;12:1717. doi: 10.2147/DMSO.S211188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Narkhede RR, Pise AV, Cheke RS, Shinde SD. Recognition of natural products as potential inhibitors of COVID-19 main protease (Mpro): In-silico evidences. Nat Prod Bioprospect. 2020;10(5):297–306. doi: 10.1007/s13659-020-00253-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nayak TK, Mamidi P, Sahoo SS, Kumar PS, Mahish C, Chatterjee S, Subudhi BB, Chattopadhyay S, Chattopadhyay S. P38 and JNK mitogen-activated protein kinases interact with chikungunya virus non-structural protein-2 and regulate TNF induction during viral infection in macrophages. Front Immunol. 2019;10:786. doi: 10.3389/fimmu.2019.00786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pagliaro B, Santolamazza C, Simonelli F, Rubattu S. Phytochemical compounds and protection from cardiovascular diseases: a state of the art. BioMed Res Int. 2015;2015(1):17. doi: 10.1155/2015/918069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paul M, Hemshekhar M, Kemparaju K, Girish KS. Berberine mitigates high glucose-potentiated platelet aggregation and apoptosis by modulating aldose reductase and NADPH oxidase activity. Free Radical Biol Med. 2019;130:196–205. doi: 10.1016/j.freeradbiomed.2018.10.453. [DOI] [PubMed] [Google Scholar]
- Pei C, Zhang Y, Wang P, Zhang B, Fang L, Liu B, Meng S. Berberine alleviates oxidized low-density lipoprotein-induced macrophage activation by downregulating galectin-3 via the NF-κB and AMPK signaling pathways. Phytother Res. 2019;33(2):294–308. doi: 10.1002/ptr.6217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pizzorno A, Padey B, Dubois J, Julien T, Traversier A, Dulière V, Brun P, Lina B, Rosa-Calatrava M, Terrier O. In vitro evaluation of antiviral activity of single and combined repurposable drugs against SARS-CoV-2. Antiviral Res. 2020;181:104878. doi: 10.1016/j.antiviral.2020.104878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pratomo IP, Ariane A, Tedjo A, Heryanto R, Paramita RI. Xanthine oxidase inhibition in SARS-CoV-2 infection: the mechanism and potency of allopurinol and febuxostat in COVID-19 management. Med J Indones. 2021;30(1):75–80. [Google Scholar]
- Qiu H, Wu Y, Wang Q, Liu C, Xue L, Wang H, Wu Q, Jiang Q. Effect of berberine on PPARα-NO signalling pathway in vascular smooth muscle cell proliferation induced by angiotensin IV. Pharm Biol. 2017;55(1):227–232. doi: 10.1080/13880209.2016.1257642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanghavi DK, Titus A, Caulfield TR, Freeman WD. Endotheliitis, endothelin, and endothelin receptor blockers in COVID-19. Med Hypotheses. 2021;150:110564. doi: 10.1016/j.mehy.2021.110564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Serveaux-Dancer M, Jabaudon M, Creveaux I, Belville C, Blondonnet R, Gross C, Constantin J-M, Blanchon L, Sapin V. Pathological implications of receptor for advanced glycation end-product (AGER) gene polymorphism. Dis Markers. 2019;2019:1–17. doi: 10.1155/2019/2067353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharif-Askari FS, Sharif-Askari NS, Goel S, Hafezi S, Assiri R, Al-Muhsen S, Hamid Q, Halwani R. SARS-CoV-2 attenuates corticosteroid sensitivity by suppressing DUSP1 expression and activating p38 MAPK pathway. Eur J Pharmacol. 2021;908:174374. doi: 10.1016/j.ejphar.2021.174374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharif-Askari NS, Sharif-Askari FS, Mdkhana B, Alsayed HAH, Alsafar H, Alrais ZF, Hamid Q, Halwani R. Upregulation of oxidative stress gene markers during SARS-COV-2 viral infection. Free Radical Biol Med. 2021;172:688–698. doi: 10.1016/j.freeradbiomed.2021.06.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shimokawa C, Kato T, Takeuchi T, Ohshima N, Furuki T, Ohtsu Y, Suzue K, Imai T, Obi S, Olia A. CD8+ regulatory T cells are critical in prevention of autoimmune-mediated diabetes. Nat Commun. 2020;11(1):1–9. doi: 10.1038/s41467-020-15857-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sohn KM, Lee S-G, Kim HJ, Cheon S, Jeong H, Lee J, Kim IS, Silwal P, Kim YJ, Paik S. COVID-19 patients upregulate toll-like receptor 4-mediated inflammatory signaling that mimics bacterial sepsis. J Korean Med Sci. 2020 doi: 10.3346/jkms.2020.35.e343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song S, Qiu M, Chu Y, Chen D, Wang X, Su A, Wu Z. Downregulation of cellular c-Jun N-terminal protein kinase and NF-κB activation by berberine may result in inhibition of herpes simplex virus replication. Antimicrob Agents Chemother. 2014;58(9):5068–5078. doi: 10.1128/AAC.02427-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Srivastava V. Quinacrine and berberine as antiviral agents against dengue and Zika fever: in silico approach. Biostat Bioinform. 2018;1:12. [Google Scholar]
- Subbaiah T, Amin A. Effect of berberine sulphate on Entamoeba histolytica. Nature. 1967;215(5100):527–528. doi: 10.1038/215527a0. [DOI] [PubMed] [Google Scholar]
- Suhail S, Zajac J, Fossum C, Lowater H, McCracken C, Severson N, Laatsch B, Narkiewicz-Jodko A, Johnson B, Liebau J. Role of oxidative stress on SARS-CoV (SARS) and SARS-CoV-2 (COVID-19) infection: a review. Protein J. 2020;39(6):644–656. doi: 10.1007/s10930-020-09935-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun H, Wang N, Cang Z, Zhu C, Zhao L, Nie X, Cheng J, Xia F, Zhai H, Lu Y. Modulation of microbiota-gut-brain axis by berberine resulting in improved metabolic status in high-fat diet-fed rats. Obes Facts. 2016;9(6):365–378. doi: 10.1159/000449507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suryavanshi SV, Kulkarni YA. NF-κβ: a potential target in the management of vascular complications of diabetes. Front Pharmacol. 2017;8:798. doi: 10.3389/fphar.2017.00798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tew XN, Lau NJX, Chellappan DK, Madheswaran T, Zeeshan F, Tambuwala MM, Aljabali AA, Balusamy SR, Perumalsamy H, Gupta G. Immunological axis of berberine in managing inflammation underlying chronic respiratory inflammatory diseases. Chem Biol Interact. 2020;317:108947. doi: 10.1016/j.cbi.2020.108947. [DOI] [PubMed] [Google Scholar]
- Varghese FS, Thaa B, Amrun SN, Simarmata D, Rausalu K, Nyman TA, Merits A, McInerney GM, Ng LF, Ahola T. The antiviral alkaloid berberine reduces chikungunya virus-induced mitogen-activated protein kinase signaling. J Virol. 2016;90(21):9743–9757. doi: 10.1128/JVI.01382-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wan Z, Zhou Z, Liu Y, Lai Y, Luo Y, Peng X, Zou W. Regulatory T cells and T helper 17 cells in viral infection. Scand J Immunol. 2020;91(5):e12873. doi: 10.1111/sji.12873. [DOI] [PubMed] [Google Scholar]
- Wang X, Zhang Y, Yang Y, Wu X, Fan H, Qiao Y. Identification of berberine as a direct thrombin inhibitor from traditional Chinese medicine through structural, functional and binding studies. Sci Rep. 2017;7(1):1–12. doi: 10.1038/srep44040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Y-X, Yang L, Wang H-Q, Zhao X-Q, Liu T, Li Y-H, Zeng Q-X, Li Y-H, Song D-Q. Synthesis and evolution of berberine derivatives as a new class of antiviral agents against enterovirus 71 through the MEK/ERK pathway and autophagy. Molecules. 2018;23(8):2084. doi: 10.3390/molecules23082084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Y, Liu H, Zheng M, Yang Y, Ren H, Kong Y, Wang S, Wang J, Jiang Y, Yang J. Berberine slows the progression of prediabetes to diabetes in Zucker diabetic fatty rats by enhancing intestinal secretion of glucagon-like peptide-2 and improving the gut microbiota. Front Endocrinol. 2021;12:609134. doi: 10.3389/fendo.2021.609134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang ZZ, Li K, Maskey AR, Huang W, Toutov AA, Yang N, Srivastava K, Geliebter J, Tiwari R, Miao M. A small molecule compound berberine as an orally active therapeutic candidate against COVID-19 and SARS: A computational and mechanistic study. FASEB J. 2021;35(4):e21360. doi: 10.1096/fj.202001792R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warowicka A, Nawrot R, Goździcka-Józefiak A. Antiviral activity of berberine. Adv Virol. 2020;165(9):1935–1945. doi: 10.1007/s00705-020-04706-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wei S, Zhang M, Yu Y, Lan X, Yao F, Yan X, Chen L, Hatch GM. Berberine attenuates development of the hepatic gluconeogenesis and lipid metabolism disorder in type 2 diabetic mice and in palmitate-incubated HepG2 cells through suppression of the HNF-4α miR122 pathway. PLoS ONE. 2016;11(3):e0152097. doi: 10.1371/journal.pone.0152097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu C, Liu Y, Yang Y, Zhang P, Zhong W, Wang Y, Wang Q, Xu Y, Li M, Li X. Analysis of therapeutic targets for SARS-CoV-2 and discovery of potential drugs by computational methods. Acta Pharmaceutica Sinica B. 2020;10(5):766–788. doi: 10.1016/j.apsb.2020.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu Z, Feng W, Shen Q, Yu N, Yu K, Wang S, Chen Z, Shioda S, Guo Y. Rhizoma coptidis and berberine as a natural drug to combat aging and aging-related diseases via anti-oxidation and AMPK activation. Aging Dis. 2017;8(6):760. doi: 10.14336/AD.2016.0620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu J, Shen J, Yuan R, Jia B, Zhang Y, Wang S, Zhang Y, Liu M, Wang T. Mitochondrial targeting therapeutics: promising role of natural products in non-alcoholic fatty liver disease. Front Pharmacol. 2021 doi: 10.3389/fphar.2021.796207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yalcin Kehribar D, Cihangiroglu M, Sehmen E, Avci B, Capraz A, Yildirim Bilgin A, Gunaydin C, Ozgen M. The receptor for advanced glycation end product (RAGE) pathway in COVID-19. Biomarkers. 2021;26(2):114–118. doi: 10.1080/1354750X.2020.1861099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan F, Wang L, Shi Y, Cao H, Liu L, Washington MK, Chaturvedi R, Israel DA, Cao H, Wang B. Berberine promotes recovery of colitis and inhibits inflammatory responses in colonic macrophages and epithelial cells in DSS-treated mice. Am J Physiol Gastrointest Liver Physiol. 2012;302(5):G504–G514. doi: 10.1152/ajpgi.00312.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan YQ, Fu YJ, Wu S, Qin HQ, Zhen X, Song BM, Weng YS, Wang PC, Chen XY, Jiang ZY. Anti-influenza activity of berberine improves prognosis by reducing viral replication in mice. Phytother Res. 2018;32(12):2560–2567. doi: 10.1002/ptr.6196. [DOI] [PubMed] [Google Scholar]
- Yang N, Wang J, Liu C, Song Y, Zhang S, Zi J, Zhan J, Masilamani M, Cox A, Nowak-Wegrzyn A. Berberine and limonin suppress IgE production by human B cells and peripheral blood mononuclear cells from food-allergic patients. Ann Allerg Asthma Immunol. 2014;113(5):556–564. doi: 10.1016/j.anai.2014.07.021. [DOI] [PubMed] [Google Scholar]
- Zha W, Liang G, Xiao J, Studer EJ, Hylemon PB, Pandak WM, Jr, Wang G, Li X, Zhou H. Berberine inhibits HIV protease inhibitor-induced inflammatory response by modulating ER stress signaling pathways in murine macrophages. PLoS One. 2010;5(2):e9069. doi: 10.1371/journal.pone.0009069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang X-D, Ren H-M, Liu L. Effects of different dose berberine on hemodynamic parameters and [Ca2+] i of cardiac myocytes of diastolic heart failure rat model. Zhongguo Zhong Yao Za Zhi Zhongguo Zhongyao Zazhi China J Chinese Mater Med. 2008;33(7):818–821. [PubMed] [Google Scholar]
- Zhang Q, Xiao X, Li M, Li W, Yu M, Zhang H, Ping F, Wang Z, Zheng J. Berberine moderates glucose metabolism through the GnRH-GLP-1 and MAPK pathways in the intestine. BMC Complement Altern Med. 2014;14(1):1–10. doi: 10.1186/1472-6882-14-188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y, Ma X-J, Guo C-Y, Wang M-M, Kou N, Qu H, Mao H-M, Shi D-Z. Pretreatment with a combination of ligustrazine and berberine improves cardiac function in rats with coronary microembolization. Acta Pharmacol Sin. 2016;37(4):463–472. doi: 10.1038/aps.2015.147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang LN, Wang XX, Wang Z, Li KY, Xu BH, Zhang J. Berberine improves advanced glycation end products-induced osteogenic differentiation responses in human periodontal ligament stem cells through the canonical Wnt/β-catenin pathway. Mol Med Rep. 2019;19(6):5440–5452. doi: 10.3892/mmr.2019.10193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang S, Liu Y, Wang X, Yang L, Li H, Wang Y, Liu M, Zhao X, Xie Y, Yang Y. SARS-CoV-2 binds platelet ACE2 to enhance thrombosis in COVID-19. J Hematol Oncol. 2020;13(1):1–22. doi: 10.1186/s13045-020-00954-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang B, Chen M, Chen X, Cao K, You Y, Qian Y, Yu W. Berberine reduces circulating inflammatory mediators in patients with severe COVID-19. Br J Surg. 2021;108(1):e9–e11. doi: 10.1093/bjs/znaa021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou H, Feng L, Xu F, Sun Y, Ma Y, Zhang X, Liu H, Xu G, Wu X, Shen Y. "Berberine inhibits palmitate-induced NLRP3 inflammasome activation by triggering autophagy in macrophages: A new mechanism linking berberine to insulin resistance improvement. Biomed Pharmacother. 2017;89:864–874. doi: 10.1016/j.biopha.2017.03.003. [DOI] [PubMed] [Google Scholar]
- Zhou G, Yan M, Guo G, Tong N. Ameliorative effect of berberine on neonatally induced type 2 diabetic neuropathy via modulation of BDNF, IGF-1, PPAR-γ, and AMPK expressions. Dose-Response. 2019;17(3):1559325819862449. doi: 10.1177/1559325819862449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zou K, Li Z, Zhang Y, Zhang H, Li B, Zhu W, Shi J, Jia Q, Li Y. Advances in the study of berberine and its derivatives: a focus on anti-inflammatory and anti-tumor effects in the digestive system. Acta Pharmacol Sin. 2017;38(2):157–167. doi: 10.1038/aps.2016.125. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The authors confirm that the data supporting this study are available within the article.