

1. RHYTHMS IN COGNITION
Brain activity abounds with rhythmic patterns (Capilla et al., 2021; Groppe et al., 2013; Keitel & Gross, 2016) that reflect ongoing fluctuations of neuronal excitability (Adrian & Matthews, 1934; Bishop, 1932). These rhythms can be described as oscillations that cycle through phases at specific frequencies and with a certain strength or amplitude (Figure 1). Attempts to account for the functional relevance of rhythms have gone from associating cognitive function X with rhythmic activity Y to a more comprehensive understanding of the role of brain rhythms in human everyday experience and behaviour (Buzsáki & Draguhn, 2004; Fries, 2015; Lakatos et al., 2019; Wang, 2010). These accounts support a simple, yet fundamental idea: Cognitive functions themselves may be intrinsically rhythmic.
FIGURE 1.
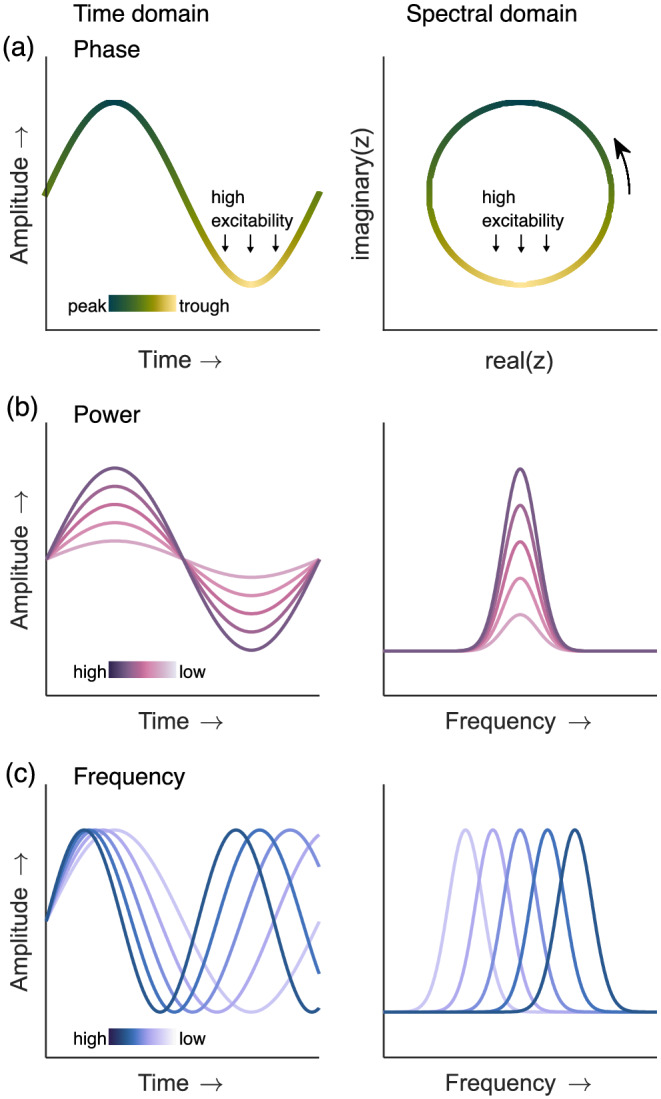
Three main properties of an oscillation used to describe rhythms: (a) instantaneous phase corresponds to the position of an oscillation, for example, the peak or trough in the time domain signal (left), at a specific moment in time. Phase is a circular measure in the spectral domain (right), as computed with a Fourier or wavelet transform. To implement a rhythmic sampling mechanism, certain periods (here the trough) are thought to index higher neuronal excitability than others. (b) The power of an oscillation is a direct measure of its amplitude. It expresses the magnitude of peaks and troughs in the time domain (left), which can be quantified by power spectra (right) indicating signal power for different frequencies. (c) Frequency corresponds to cycles per time unit (left: time domain; right; power spectrum)
For instance, everyday experience may arise from a fundamentally discrete sampling of our sensory environment (a.k.a. ‘rhythmic sampling’, ‘perceptual cycling’), an idea that starkly contrasts with our intuition of a continuous perceptual flow. Its origins can be traced back to the beginning of the 20th century: Inspired by the recently invented cinematograph, the philosopher Henri Bergson proposed that human perception may operate similarly to a motion picture, in which an apparently continuous experience is made up of temporally discrete events (Bergson & Frye, 1911). Further 20th‐century inventions such as the digital computer, television, and early computational neural networks provided further analogies for temporally discrete information processing, and brain rhythms were seen as obvious candidates for their physiological implementation (see Busch & VanRullen, 2014 for a historical review). For instance, Norbert Wiener, one of the founders of cybernetics, in reviewing Pitts and McCulloch's (1947) computational model of object recognition, proposed (Wiener, 2019, p. 141):
“The scanning apparatus should have a certain intrinsic period of operation which should be identifiable in the performance of the brain. [...] In fact, it has the order of frequency appropriate for the alpha rhythm of the brain, as shown in electroencephalograms. We may suspect that this alpha rhythm is associated with form perception, and that it partakes of the nature of a sweep rhythm, like the rhythm shown in the scanning process of a television apparatus.”
Theories derived from these early ideas have been reiterated over the past 90 years. During this time, research has focussed on variations of a framework claiming that cognitive processes depend on brain rhythms and has offered a wealth of empirical evidence (Schroeder et al., 2010; VanRullen, 2016; VanRullen & Koch, 2003). Rhythmic sampling, implemented through brain rhythms, has been proposed as a fundamental process that underlies not only perception but also attention (VanRullen et al., 2007), memory (Staresina et al., 2015), and language (Giraud & Poeppel, 2012).
Linking brain rhythms and cognition through rhythmic sampling is particularly attractive because it allows using global mechanistic principles to bridge two different levels of explanation: normal brain function, which involves rhythmic activity, and human behaviour and experiences. In addition, it leaves open the possibility of actively intervening in the sampling process through external stimulation of brain rhythms, either to advance theory, for example, to close in on the causal role of brain rhythms for cognition and behaviour (see section ‘Can we manipulate brain rhythms to take control of cognition?’), or to develop applications for clinical interventions, such as rehabilitation, and brain‐computer interfaces.
The field, however, is not immune to controversy. The available evidence does not unequivocally support the assumed role of brain rhythms in rhythmic sampling, for example, in visual processing (Benwell et al., 2017; Chaumon & Busch, 2014; Fekete et al., 2018; O'Hare, 1954; Ruzzoli et al., 2019; van Diepen et al., 2015; Van Diepen et al., 2019; Walsh, 1952), and various empirical and theoretical inconsistencies have been highlighted (Fekete et al., 2018; White, 2018). For instance, the exact sampling frequencies implicated in cognitive functions often differ between studies and have been shown to depend on the task and/or stimulus characteristics (Chen et al., 2020; Ho et al., 2017; Merholz et al., 2022; Ronconi et al., 2017; also see table in Ruzzoli et al., 2019), which is difficult to reconcile with any simple model of fixed, discrete temporal ‘frames’ (White, 2018). Additionally, the effect sizes in studies showing periodicity in behavioural or neural measures tend to be small, hence not in line with clear, all‐or‐nothing frame boundaries (Milton & Pleydell‐Pearce, 2016; White, 2018) or utility for real‐life applications (Vigué‐Guix et al., 2020). Furthermore, the literature suffers from a lack of direct replications, pre‐registered studies, data, and code sharing (Garrett‐Ruffin et al., 2021; Niso et al., 2021; Pavlov et al., 2021), as well as low statistical power (Button et al., 2013). Finally, publication bias in favour of statistically significant results may have distorted the evidence and undermined the reliability of the published literature (Szucs & Ioannidis, 2017).
Despite extensive research, the existence and exact nature of rhythmic sampling processes remain intensely debated, and the idea of rhythms in cognition continues to attract vast interest. In this Special Issue, we called for methodologically principled studies, irrespective of their outcome, that would contribute to clarifying if, and under which conditions, a rhythmic sampling of cognition can be observed. In addition to 26 original research articles (including pre‐registered original studies, as well as direct or conceptual replications), we also received literature reviews, methodological advances, and theoretical position papers. Following our aim to re‐visit the evidence for rhythms in cognition, we evaluated the primary outcomes of the 23 research articles that tested the rhythmic sampling hypothesis directly. A split picture emerged with 11 studies reporting supportive evidence and 12 others reporting null or inconclusive findings (see Figure 2).
FIGURE 2.
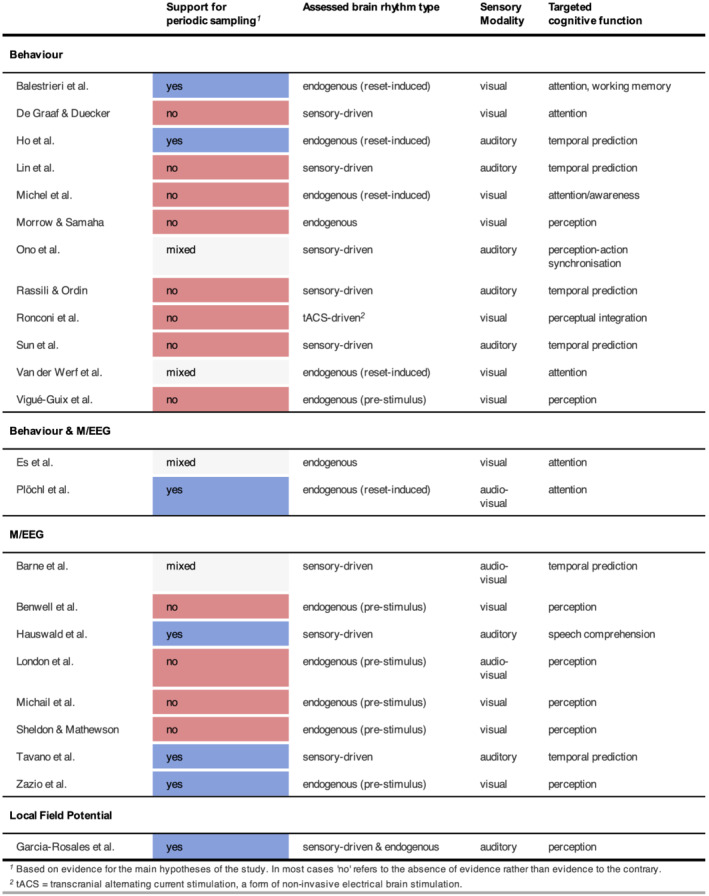
All 23 research articles published in this special issue, in which periodic sampling was assessed directly, ordered by the primary measurement ‐ behaviour, electrophysiological measures (MEG, EEG, and local field potentials), or both. We broadly classified studies as providing supporting (blue), mixed (grey), or no (red) evidence for the periodic sampling idea
Within this context, we here review the 36 publications in our Special Issue with an emphasis on how each contribution informs open questions or controversies in the wider literature on rhythms in cognition. In doing so, we cluster the different contributions around two prevalent topics – Does cognition operate rhythmically? And if so, can we manipulate brain rhythms through rhythmic external stimulation to take control of cognitive processes?
2. DOES COGNITION OPERATE RHYTHMICALLY?
The majority of evidence for rhythms in cognition has come from studies showing rhythmic patterns in behavioural performance or from studies showing an influence of rhythmic neural activity on behavioural outcomes. Below, we summarise previous findings from these two lines of research and discuss the contributions of the relevant studies published in this Special Issue.
2.1. Rhythmicity in behaviour
Numerous studies have demonstrated rhythms in behavioural performance by probing performance at various time intervals relative to a reference event. Provided that the intervals are distributed across a wide‐enough range and sampled densely within that range, the resulting time course of performance across intervals can be submitted to a spectral analysis and tested for rhythmicity. This approach is not new (e.g., Latour, 1967; Purushothaman et al., 2000) but has become increasingly popular within the last decade (e.g., Dugué et al., 2015; Fiebelkorn et al., 2013; Landau & Fries, 2012; Senoussi et al., 2019). We refer the reader to Kienitz et al. (2021) for a comprehensive overview of the experimental paradigms employed to study rhythms in behaviour during attentive vision in both human and non‐human primates.
In the study of rhythms in behaviour, an ongoing debate surrounds potential functional dissociations between rhythms in different frequencies. Michel et al. (2021) addressed this topic and evaluated information processing during up and down phases of both theta (4–7 Hz) and alpha (8–12 Hz) rhythms. The participants performed an orientation judgement task after a non‐informative cue (50% validity) indicated the target location. Michel et al. observed that the precision of the orientation judgement oscillated at alpha frequencies for invalidly cued targets, whilst the guess rate oscillated at theta frequencies for validly and invalidly cued targets. The results suggest distinct roles for the two rhythms, with alpha potentially subserving perception and theta subserving attentional fluctuations. However, the authors suggested that the resulting environmental sampling likely drives fluctuations in spatial resolution, rather than a strict succession of fixed, discrete perceptual snapshots and blind gaps during the ‘up’ and ‘down’ phases of the rhythms, respectively.
In addition to possible functional dissociations between rhythms of different frequencies, the degree to which the rhythms observed in behavioural performance is fixed or rather depends on task demands and/or participant state remains unclear. In this regard, Balestrieri et al. (2021, this issue) showed an interplay between attention and working memory. Rhythms in visual detection performance were observed during the maintenance period of a simultaneous visual working memory task. Interestingly, the rhythmic detection frequency was modulated by the working memory load, oscillating at ~7.5 Hz versus 5 Hz for low versus high load, respectively, suggesting that the sampling frequency might be modulated depending on the allocation of limited attentional resources to the perceptual or the memory task.
In another study in this Special Issue, Morrow and Samaha (2021, this issue) tested whether visual rhythmic sampling occurs at alpha‐band frequencies (Chakravarthi & VanRullen, 2012; Sokoliuk & VanRullen, 2019; VanRullen, 2016), capitalising on two motion‐related visual illusions: the flash‐lag effect and the Fröhlich effect. Indeed, according to a recently proposed model of discrete perceptual sampling (K. A. Schneider, 2018), visual perception might be supported by a series of discrete ‘perceptual moments’, the exact duration of which should account for individual differences in the experience of the two illusions. Morrow and Samaha hypothesised that if the flash‐lag and Fröhlich effects are driven by discrete sampling at the endogenous alpha peak frequency of participants, then the magnitude of each illusion should be correlated across individuals. The authors successfully induced both illusions in all participants but found no correlation in the magnitude of the illusions. A Bayesian analysis favoured their null hypothesis that a single underlying oscillatory mechanism could not account for both the flash‐lag and the Fröhlich illusions.
Plöchl et al. (2021) examined whether attentional sampling also occurs in the auditory modality. Their study included visual, auditory, and bimodal conditions in a detection task, which closely replicated the seminal study from Landau and Fries (2012). Specifically, they capitalised on an attentional reset stimulus (in the visual or auditory modality) to automatically drive attention to one of two (left vs. right) streams of events and then tested perceptual detection at cued versus uncued locations. The results showed that both visual and auditory target detection fluctuated at a theta rhythm that was in counter phase at the two attended locations, confirming that attentional rhythms are supramodal and not specific to visual processing.
Ho et al. (2021, this issue) focussed on rhythmic sampling in the auditory modality entirely and replicated an earlier finding where they showed an alpha‐range (9 Hz) periodicity in response bias in perceptual decision making (Ho et al., 2017). Additionally, they combined the original task with a dichotic oddball paradigm to test the effect of violating sensory expectations and made the novel observation that oddball stimuli played to the non‐anticipated ear would induce subsequent theta‐range (7.6 Hz) fluctuations in response accuracy. Ho et al. suggest that this rhythm may reflect an update mechanism for auditory representations following violations of sensory expectations.
In their review on rhythms in behaviour, Kienitz et al. (2021) concluded that, although rhythmicity in behaviour is corroborated by a (growing) number of studies, special considerations at both methodological and analytical levels are necessary (see also Brookshire, 2021). For instance, appropriate test time intervals need to be chosen, which might require studies with a large number of trials and participants. On that note, van der Werf et al. (2021) were unable to replicate previous findings of rhythmic attentional sampling (Fiebelkorn et al., 2018; Helfrich et al., 2018) in a modified Egly‐Driver task (Egly et al., 1994) This task involves detecting a target that can appear at one end (i.e., the cued location) or at the other end (i.e., space‐based, non‐cued location) of a bar, or at the equidistant end of another bar (object‐based, non‐cued location). No rhythmic pattern of detection accuracy was found at cued locations for any level of predictive cue validity, though an exploratory analysis did reveal a significant ~7–8 Hz behavioural rhythm at non‐cued locations when the cues were moderately informative. In a commentary on Van der Werf et al.'s study, Fiebelkorn (2021, this issue) proposes that, despite being comparable to the original study (Helfrich et al., 2018), the number of trials per condition employed in the replication may have been too low to reliably detect attentional rhythms. Fiebelkorn further suggests that using purely behavioural data to detect rhythms in cognition might be limiting and suggests that combining behavioural data with simultaneously recorded electrophysiology might be more appropriate (but see ten Oever et al., 2022, this issue, for a response to Fiebelkorn's commentary).
2.2. Linking pre‐stimulus neural oscillations to behaviour
Magneto‐ and electroencephalography (M/EEG) allow investigating the rhythmic sampling hypothesis in brain activity. They enable analyses of behavioural performance, for example, stimulus detection, as a function of parameters of neural rhythms (phase and amplitude) at the time of stimulus presentation. Most of the M/EEG analyses published in this Special Issue revolve around the question of whether pre‐stimulus brain oscillations affect behaviour, either by attempting to replicate previous studies or by investigating these effects under specific, previously untested conditions.
For example, EEG data presented by Plöchl et al. (2021, this issue) supported the findings of rhythmic patterns in behavioural performance whilst the participants detected auditory and visual targets at cued and uncued locations. Indeed, alpha‐band power was modulated rhythmically at theta frequencies, in counter‐phase between cued and uncued locations, and predicted behavioural performance in both modalities.
In another study, Benwell et al. (2021, this issue) employed a challenging letter identification task to measure accuracy and subjective perceptual awareness on each trial. They confirmed previous reports that low pre‐stimulus alpha power is associated with subjective awareness but not accuracy (also see Benwell et al., 2017; Samaha et al., 2017). By contrast, pre‐stimulus alpha phase had no effect on either subjective awareness or accuracy, again replicating previous studies that have shown no influence of oscillatory phase on perceptual outcomes (Benwell et al., 2017; O'Hare, 1954; Ruzzoli et al., 2019; van Diepen et al., 2015; Walsh, 1952). Similarly, Michail et al. (2021, this issue) found an effect of low pre‐stimulus alpha power on visual discrimination performance, and no corresponding effect of pre‐stimulus phase. A unique aspect of this experiment was the long duration of the stimuli of up to 2.5 s. Accordingly, the authors speculate that oscillatory phase might be less likely to affect perception of stimuli lasting for several oscillatory cycles.
Sheldon and Mathewson (2021, this issue) set out to test whether pre‐stimulus and/or post‐stimulus oscillations affect the precision of visual perception or the likelihood of guessing. They employed an orientation discrimination task in which the participants reproduced the stimulus orientation as precisely as possible and analysed the resulting error distribution with statistical models that yield separate estimates of perceptual precision and guessing. Conceptually, their study was similar to Michel et al. (2021, this issue) who analysed behavioural rhythms (see section ‘Rhythmicity in behaviour’). Interestingly, Sheldon and Mathewson did not replicate a previously observed combined effect of pre‐stimulus alpha phase and power on performance (Mathewson et al., 2009; also see Fakche et al., 2022), because either pre‐stimulus or post‐stimulus alpha phase or power had an effect on performance. Instead, only the power and phase of 2–7 Hz post‐stimulus activities were associated with performance.
Zazio et al. (2021, this issue), in turn, found clear evidence for pre‐stimulus phase effects on behaviour. They employed a visual target detection task that resembled an earlier study (Busch et al., 2009), which had reported effects of pre‐stimulus alpha phase and power on detection performance. Using MEG source analysis, Zazio et al. demonstrated that target detection and the stimulus‐evoked MEG response were inversely related to pre‐stimulus alpha power in occipito‐temporal brain areas, whereas phase effects on behaviour were localised to both occipital and prefrontal areas.
Whilst the other studies described in this section assessed rhythmic sampling of sensory information by analysing temporal variations in behavioural performance, van Es et al. ( 2020, this issue) used a support vector machine to decode attended stimulus orientation (counter/clockwise) from the MEG signal. They found that classification performance fluctuated along with the phase of MEG theta/alpha oscillations in the frontal eye field and parietal cortex contralateral to the attended grating stimulus. However, these fluctuations were not reflected in behaviour (reaction times), leaving the functional relevance of the rhythmic neural representations unknown. Similarly, Gaillard and Ben Hamed (2020, this issue) review neurophysiological studies and argue that attention seems to operate continuously in prefrontal brain regions but alternates periodically between the center and periphery at various frequencies in parieto‐frontal networks. This would be in line with a two‐stage model positing that a combination of continuous and discrete processes interact to form conscious percepts (Herzog et al., 2020).
Vigué‐Guix et al. (2020, this issue) also attempted to demonstrate an effect of pre‐stimulus alpha phase on behavioural performance. Their experiment had several interesting features that stand out from the other studies described in this section. First, the authors aimed at replicating Callaway and Yeager (1960), one of the early seminal papers looking into the rhythmic sampling idea. Vigué‐Guix et al. used a closed‐loop approach to analyse alpha phase in real‐time and presented stimuli (bright LED flashes) selectively at specific phases of the ongoing alpha rhythm. Second, the participants kept their eyes closed throughout the experiment, which amplified the ongoing alpha rhythm and facilitated extracting its phase. Finally, their study used a comparably small sample of participants because they aimed at demonstrating a phase effect in each individual rather than at the sample level. However, although they showed a reliable real‐time estimation of the phase parameter and successfully managed to present visual stimuli accordingly, the phase at stimulus onset had no effect on response times in any participant. The results suggest that any putative effect of pre‐stimulus phase was not sizable enough to be of practical relevance, for example, as a control signal for brain‐computer interface applications.
Along with the influence of oscillatory power and phase, recent studies have also investigated whether the instantaneous frequency of the EEG signal is predictive of behavioural outcomes. For instance, both within‐ and between‐participant differences in peak alpha frequency have been shown to be predictive of the likelihood of perceptual integration of temporally proximal stimuli both in vision (Samaha & Postle, 2015) and in multisensory (audiovisual) processing (Cecere et al., 2015; Keil & Senkowski, 2017), in line with the theory of perceptual cycles (VanRullen, 2016). London et al. (2022, this issue) investigated whether these results would extend to an audiovisual temporal order judgement task with supra‐threshold stimuli. Whilst low pre‐stimulus alpha power over occipito‐parietal electrodes predicted increased temporal audiovisual sensitivity, no effect of instantaneous alpha frequency was found. These results, along with those of another recent study (Buergers & Noppeney, 2022), cast doubt on the notion that peak alpha frequency is a determining factor in the temporal sensitivity of perception.
2.3. Interim discussion
Does cognition operate rhythmically? The evidence provided by the studies reviewed thus far does not provide a simple answer either way (see also Figure 2). Behavioural studies in this special issue found evidence for perceptual or cognitive rhythms more frequently than M/EEG studies (Figure 2). This is perhaps surprising and contrasts with Fiebelkorn's suggestion (Fiebelkorn, 2021, this issue), given that M/EEG represents a more direct measurement of the rhythms in question. Whilst the effect of pre‐stimulus oscillatory power on behavioural performance across various tasks appears to be quite robust, particularly for alpha rhythms (see, e.g., D. Schneider et al., 2021, this issue), only one study in this Special Issue found evidence for the effects of pre‐stimulus alpha phase on behaviour (Zazio et al., 2021). Therefore, evidence presented in favour of pre‐stimulus phase effects was markedly weaker compared to the power effects. This aligns with a number of previous studies reporting alpha power but not phase effects (Benwell et al., 2017; Benwell, Keitel, et al., 2019; Chaumon & Busch, 2014; O'Hare, 1954; Ruzzoli et al., 2019; Samaha et al., 2017; van Diepen et al., 2015; Walsh, 1952).
Notably, even within the subset of studies that can be considered direct replication attempts of previous findings (Plöchl et al., 2021; van Der Werf et al., 2021), or at least studies with very similar designs (van Der Werf et al., 2021; Zazio et al., 2021), the results are mixed. Replication and extension of key findings in the literature will be critical for the development of the field (Pavlov et al., 2021), and the replication studies included in this Special Issue provide valuable examples. It is of course crucial that methodologies should be optimised for replication attempts (Fiebelkorn, 2021; ten Oever et al., 2022). However, if seminal studies are considered to have methodological flaws (such as low statistical power), then it is all the more important that the results are validated through replications.
Replication studies should ideally aim for increased statistical power both within participants (i.e., trial numbers) and in terms of overall sample size. This could be facilitated by collaborative studies run across several labs (Open Science Collaboration, 2015). Another important future step will be to derive and test hypotheses from existing theories about the conditions, tasks, and states under which rhythmic sampling should be observed (and those in which it should not). For instance, the notion that rhythms reflect fluctuations in neuronal excitability (Adrian & Matthews, 1934; Bishop, 1932) predicts that behaviour should be rhythmically modulated only when behavioural performance is, in fact, limited by excitability. This is arguably the case for short stimulus durations and tasks like contrast detection or simple feature discrimination but might not be the case for longer stimulus durations (Michail et al., 2021; Sheldon & Mathewson, 2021; both this issue).
A related challenge for this field will be to define a benchmark paradigm that ‘just works’, that is, a set of stimuli, tasks, and analyses that can be trusted to yield a behavioural rhythm or effect of neural oscillatory phase on behaviour. This is in contrast to other fields in cognitive neuroscience that have developed conventional paradigms and analyses for eliciting an effect of interest, for example, attentional capture (Theeuwes, 1992), the effect of attentional cueing on contrast sensitivity (Carrasco, 2011), or EEG markers of visual memory (Ngiam et al., 2021). Establishing such a standard will provide a valuable vantage point from which to chart the parameters that determine when neural phase affects cognition.
3. CAN WE MANIPULATE BRAIN RHYTHMS THROUGH RHYTHMIC EXTERNAL STIMULATION?
Manipulating brain rhythms through external periodic stimulation has also been used as a tool to study rhythmic sampling. As Quigley (2021, this issue) reports, this approach dates back to Adrian and Matthews (1934) and Walter (1954), who used periodic light stimulation and compared light‐induced brain responses to endogenous brain rhythms. The process of synchronisation between periodic inputs and endogenous brain rhythms has since been termed ‘entrainment’. The entrainment idea remained pervasive and drew strengths through two major routes: the explanatory power of entrainment as a neural process subserving cognitive functions, and the versatility of the entrainment technique for experimenters and, later on, interventionists.
Regarding its explanatory power, entrainment has been considered the neural mechanism underlying Dynamic Attending Theory, which posits that internal oscillators (i.e., brain rhythms) synchronise with external periodic stimulation (Jones & Boltz, 1989; also see Henry & Herrmann, 2014). Once established, synchronised internal cycling allows phases of optimal cortical excitability to coincide with upcoming stimuli and facilitates their processing (Schroeder & Lakatos, 2009). Similarly, entrainment has widely been promoted as a general mechanism subserving interactions between the brain and sensory input (Lakatos et al., 2019; also see Tavano et al., 2022, this issue). As an emblematic example, entrainment may crucially underlie our ability to tune in to segment and understand speech (Giraud & Poeppel, 2012; Rimmele et al., 2015; also see Hauswald et al., 2020, this issue).
From the perspective of experimenters, entrainment has also been proposed as a versatile method for directly manipulating endogenous brain activity. Next to rhythmic sensory stimulation, rhythmic magnetic and electrical non‐invasive brain stimulation (NIBS) can also be used (Thut, Schyns, & Gross, 2011). The prospect of influencing brain rhythms through sensory stimulation has further inspired research into potential interventions for pathological brain conditions. In this regard, please see Tichko et al. (2020) who reviewed studies that used a gamma‐frequency stimulation (40 Hz) to attempt to counteract pathophysiologies linked with Alzheimer's disease (Iaccarino et al., 2016) and made a strong case for integrating periodic sensory stimulation into music‐based interventions as currently employed in Alzheimer's therapy.
Despite being received enthusiastically overall, it remains unclear, however, whether entrainment is a fundamental neural mechanism implementing rhythmic sampling for cognition in general or whether it only applies in more circumscribed situations.
3.1. Sensory entrainment
One approach to determine the functional significance of entrainment is to test whether it benefits cognition beyond extracting temporal regularities from sensory input. For instance, if entrainment implements selective attention (Calderone et al., 2014; Lakatos et al., 2013), it should selectively facilitate the processing of behaviourally relevant stimulus features in addition to providing optimal periods for processing (also see Barne et al., 2021, this issue). Indeed, Tavano et al. (2022, this issue) present evidence from an EEG experiment that more predictably recurring features of a stimulus (here: tone frequency) lead to higher cortical entrainment than less predictable features, both embedded in regular tone sequences.
In another study, Barne et al. (2021, this issue) presented rhythmically interleaved auditory and visual target stimuli and instructed participants to compare them to a probe after a brief retention interval. Although they did not find a behavioural benefit of rhythmic over non‐rhythmic stimulation (in line with de Graaf & Duecker, 2021; W. M. Lin, Oetringer, et al., 2021; Rassili & Ordin, 2020; Sun et al., 2021; all this issue), nor neural activity correlated with pre‐probe feature templates during the retention period, sensory cortices showed intermittent pre‐activation at the stimulus rhythm, likely in anticipation of processing the probe stimulus. This anticipatory anti‐phasic activation of auditory and visual cortices supports the idea of a periodic process that puts the sensory cortex into a maximally excitable state at the time of (potential) probe delivery. Moreover, it suggests that this process is induced by rhythmic stimulation and can self‐sustain for a number of cycles after the rhythmic stimulation ceases.
In contrast, Lin et al. (2021) present data from three experiments using different versions of an auditory pitch discrimination task. In a total of 181 participants, they found no evidence that targets occurring in phase with a rhythmic cue were discriminated faster or more accurately compared to targets occurring out‐of‐phase. Notably, in their first two experiments, they used low‐frequency auditory stimulation in the range of 1–3 Hz. In a similar range, but using a different paradigm, Hickok et al. (2015) had previously reported that participants detected in‐phase targets more easily, in line with the notion of entrainment.
Similarly, Sun et al. (2021, this issue) attempted to replicate Hickok et al.'s (2015) findings. After a rhythmic cueing period with amplitude‐modulated noise, the participants responded to target tones embedded in constant noise but at different times relative to the phase of the pre‐target noise. Although target detection fluctuated periodically for some participants, these periodicities were not consistently linked with the rhythmic cueing (but see Saberi & Hickok, 2021).
Rassili and Ordin (2020, this issue) compared whether presenting a regular sequence of (linguistic) syllables and (non‐linguistic) natural sounds similarly affected the detection of an intermediately presented target sound. They observed that neither type of stimuli showed an effect that exceeded target detection performance in a control condition with irregular stimulus presentation. Further exploratory analyses hinted at faster reactions to later targets. This effect was exclusively found for linguistic stimuli, leading the authors to suggest a connection with the tuning of the auditory system to process the (quasi‐) regular speech.
In the visual modality, de Graaf and Duecker (2021, this issue) tested whether synchronising alpha rhythms through rhythmic stimulation impacts visual target detection speed. They exploited the known lateralisation of alpha power in occipital cortices when attention focusses covertly on the left or right visual hemifields (e.g., Clayton et al., 2018). De Graaf and Duecker reasoned that rhythmic stimulation in the unattended hemifield should facilitate the inhibitory effect of alpha power if the stimulation synchronised (or entrained) with the cortical alpha generators. However, they found no evidence of such an effect in a large sample of participants (N = 115).
Finally, linking perception and action, entrainment is also considered pivotal for producing complex rhythmic behaviours such as making music or dancing, both of which require constant synchronisation of sensory input and motor output (Egger et al., 2020). Entrainment‐based accounts of these behaviours have received widespread support from studies using beat‐based finger tapping as a model (Repp, 2005; Repp & Su, 2013). However, not all findings fully align with the idea of synchronised internal oscillators. Ono et al. (2021, this issue) tested alternative accounts for the so‐called subdivision benefit, that is, the effect that we tap more synchronously with a beat interval that contains intermediate tones (versus none). Their results favour an explanation that considers the singular intermediate tones as an additional temporal reference over an entrainment account.
3.2. Non‐invasive brain stimulation
Transcranial magnetic (Thut, Veniero, et al., 2011) and electric brain stimulation (Zaehle et al., 2010) can also be used to non‐invasively entrain or modulate specific parameters of oscillatory brain activity. Kasten and Herrmann (2020, this issue) reviewed the main evidence for rhythmic aspects of cognition gathered through NIBS studies. They highlight two possible ways by which NIBS can probe periodic sampling. First, NIBS can be used to control the phase of a spontaneous oscillation, therefore making it easier to present a target at the optimal phase for a particular cognitive process. For example, Helfrich et al. (2014) used concurrent transcranial alternating current stimulation (tACS; 10 Hz) and EEG recordings to prove that parieto‐occipital alpha activity synchronises with tACS and that behavioural detection of visual stimuli is phase dependent. Second, NIBS can be used to modify the spontaneous frequency of an oscillation, therefore changing the rate of any periodic process (e.g., Janssens et al., 2021, this issue). For example, tACS has been applied with a frequency closely matching individual alpha rhythms to study phase‐dependent perceptual integration (Cecere et al., 2015). Following this approach Ronconi et al. (2020, this issue) stimulated parietal cortex using an innovative multi‐channel tACS montage while the participants performed a visual temporal integration/segregation task. The tACS frequency was tuned to the individual alpha frequency (IAF) and delivered at slightly slower and faster frequencies (IAF ± 2 Hz) with the intention to affect temporal integration and segregation in opposite ways. Contrary to the authors' expectations, this tACS protocol did not consistently increase or decrease the IAF and did not affect temporal integration or segregation processes. However, note that this NIBS approach to test rhythmic sampling implies a careful consideration of the stimulation parameters (Riddle & Frohlich, 2021; Woods et al., 2016). In this regard, Janssens et al. (2021, this issue) provide methodological insights for the reliable estimation of IAF that can then be targeted with tACS. In addition, there is a third way in which NIBS can be used to test periodic sampling, which uses single‐pulse TMS to probe brain rhythms at multiple delays (Dugué et al., 2019; Dugué & VanRullen, 2017).
Although Kasten and Herrmann (2020, this issue) propose an overall optimistic view about using NIBS to test for rhythms in cognition, they also discuss failed attempts to replicate previous findings and highlight methodological considerations that may influence experimental outcomes. These include the type of task used (e.g., detection vs. discrimination), the dependent variable collected (e.g., accuracy vs. RT), and the dose and duration of the stimulation (Riddle & Frohlich, 2021; Woods et al., 2016).
3.3. Interim discussion
Can we manipulate brain rhythms through external periodic stimulation to take control of cognitive processes? The studies in this Special Issues that tested the effects of sensory entrainment on behaviour add to a growing body of research that challenges the central role of entrainment in cognitive processes (Duecker et al., 2021; Gray & Emmanouil, 2019; van der Plas et al., 2020; Zou et al., 2021). They suggest that the sensory systems may not always be able to reap benefits from rhythmic input, either because entrainment does not occur invariably or because it does not influence behaviour.
Contributions to this Special Issue that used or advocated NIBS‐based entrainment add to an ongoing debate about the effectiveness of tACS to entrain neural activity. tACS is an appealing approach because it may allow causal inferences about rhythmic brain activity and its function, overcoming correlational approaches typical of EEG and MEG studies (Bergmann & Hartwigsen, 2021). However, it also has several limitations which make straightforward interpretations of experimental results challenging (Asamoah et al., 2019a; Ruhnau et al., 2020). First, tACS cannot readily be applied with high spatial precision (but see Vöröslakos et al., 2018 for a suggested improvement); second, our knowledge regarding the correct dose to induce the desired effects is limited; third, and most important, how and in which way tACS influences the brain is still under debate (Asamoah et al., 2019a; Huang et al., 2021; Vöröslakos et al., 2018). Caution is therefore advisable when using tACS if the goal is to test the role of rhythmic brain activity subserving cognition.
Reconciling findings from this Special Issue with previous positive findings may require using the term entrainment with more rigour. A precise physical definition exists – driving an oscillator, that is, a brain rhythm, with such force that it starts oscillating with the period of the external drive instead of its own, usually close‐by resonant frequency (see, e.g., Herrmann et al., 2016; van Bree et al., 2021) – but the term has repeatedly been used more broadly, also for cases where brain responses merely match the temporal characteristics of a stimulus (as discussed in Alexandrou et al., 2020; Haegens, 2020; Meyer et al., 2020; Obleser & Kayser, 2019). Methodologically, these cases do not require assuming the involvement of endogenous brain rhythms but remain difficult to discern from entrainment proper (Keitel et al., 2021). Going forward, additional criteria may be needed to establish whether entrainment took place. For example, an entrained oscillator should reverberate for a number of cycles after the cessation of its external drive (Barne et al., 2021, this issue; Lakatos et al., 2013; Lin, Shukla, et al., 2021; van Bree et al., 2021).
Finally, in line with our conclusion of the section ‘Does cognition operate rhythmically?’, the entrainment approach to testing for effects of rhythmic sampling would also benefit from well‐powered, potentially multi‐lab replications, as well as establishing paradigms that produce entrainment effects reliably, and against which new experimental results can be evaluated.
4. BEYOND OSCILLATORY PHASE
The topic of our Special Issue emphasised rhythmic aspects of cognitive functions, mainly linked to the perceptual sampling idea. These rhythms have been closely tied to the phase property of cortical oscillations. Naturally, oscillations have other properties, such as power and instantaneous frequency, that undergo dynamics linked to cognitive processes (see Figure 1). These properties are interdependent in several ways: For example, estimating oscillatory phase in noisy EEG signals or the time courses of behavioural data depends on oscillatory power (Cohen, 2014; Fakche et al., 2022). Furthermore, from a functional perspective, oscillations of different frequencies can interact by means of phase‐amplitude coupling (Canolty & Knight, 2010; Jensen & Colgin, 2007). Therefore, some contributions to this Special Issue went beyond the role of oscillatory phase to study the wider role of neural rhythms in cognition.
For example, Hauswald et al. (2020, this issue) reported a decrease in the power of alpha oscillations with increased listening effort, in addition to showing phase‐related effects on the cortical tracking of speech. These results suggest that a combination of two neural mechanisms, phase‐dependent cortical tracking and alpha suppression, benefit speech comprehension (also see Tune et al., 2021; Wöstmann et al., 2016).
Modulations of alpha power have also been associated with the quality of encoding of new items into memory (Tuladhar et al., 2007; for reviews see Clayton et al., 2018; Schneider et al., 2021, this issue). Wynn et al. (2021, this issue) employed a memory task in which the participants memorised target images whilst disregarding distractor images. They found that relatively stronger alpha power in the cortical hemisphere processing distractor images compared to the hemisphere processing target images was associated with better memory performance. Interestingly, they compared this effect between younger and older participants, who were comparably more distractible. Moreover, older participants only showed inhibitory alpha power modulation during high distraction.
In another study, Whitmarsh et al. (2021, this issue) investigated how several physiological indicators of somatosensory attention determined the subjective experience of tactile stimulation. The participants performed a somatosensory attention task, in which rare targets were embedded in a sustained, rhythmic tactile stimulation. After each trial, they rated their introspective experience of attention. Higher attention ratings were correlated with reduced alpha power in contralateral somatosensory cortex. Likewise, attention was correlated with increased pupil size – a basal forebrain – and brainstem‐driven neuromodulatory influence on cortical activity (Dahl et al., 2022; Pfeffer et al., 2022). Interestingly, whilst pupil dilation in a given trial was also correlated with attention in the following trial, the correlation between alpha and attention was restricted to the present trial. This finding might indicate that pupil size reflects slow changes in physiological, autonomic arousal, whilst alpha power reflects more dynamic changes in neuronal excitability under cognitive control. In general, this study also showcases how indicators beyond behavioural performance (here subjective ratings and pupillometry) can meaningfully complement investigations into the role of neural rhythms in cognition.
Garcia‐Rosales et al. (2020, this issue) assessed whether the power of high‐frequency gamma‐range cortical activity modulates the phase of low‐frequency delta‐ and theta‐range oscillations in bats, a process that might underlie the segmentation of steady sensory input into behaviourally relevant chunks of information. Frontal areas involved in auditory processing showed different phase‐amplitude coupling profiles than auditory cortices, thus giving an indication of how the different functional roles of these regions express themselves in local circuit dynamics. The authors further issue a note of caution on the potential influence of stimulus rhythmicity, as well as respiration as a physiological rhythm, on their own and similar results.
Finally, Donoghue et al. (2021, this issue) give a comprehensive picture of a range of further parameters of oscillations, including waveform shape, asymmetry, and temporal variability (oscillatory bursts vs. continuous oscillations), to consider when analysing brain rhythms and how to leverage their full complexity.
Taken together, these contributions suggest that we can build a more comprehensive picture of cognitive function by integrating the phase‐based perceptual sampling idea with other signature qualities of oscillatory brain activity and setting it in context with other non‐cortical, physiological rhythms.
5. NEW PERSPECTIVES ON RHYTHMS IN COGNITION
This Special Issue provides an updated cross section of results, opinions, and methodological considerations regarding rhythms in cognition, including 26 original research contributions. A number of submissions also made their data and analysis codes publicly available, in line with our endeavour to foster transparency and open science best practices. About half of the research studies included reported findings that do not directly support the existence of rhythmic sampling in cognition (Figure 2).
Although this Special Issue only presents a snapshot of the current state of the field, interesting patterns emerged when classifying studies based on whether they did or did not provide evidence for rhythms in cognition. For instance, whilst the majority of studies testing for electrophysiological evidence came up empty handed, studies testing for endogenous rhythms in behavioural measures presented mostly positive findings. In turn, however, studies that used the entrainment approach did not find supportive evidence in behavioural data.
Interestingly, the mixed evidence for the effects of pre‐stimulus oscillatory phase on stimulus perception presented in this Special Issue undermines the entrainment approach to studying rhythmic sampling to some extent. Put differently, why make an effort to control rhythmic sampling through entrainment, when the phase of endogenous oscillations does not seem to affect cognition consistently? Which processes can be targeted with entrainment impactfully? Further, absent support for the effects of entrainment on behaviour in this Special Issue contrasts starkly with the studies supporting rhythmic sampling in behaviour without using entrainment (see Figure 2). Therefore, can rhythmic brain activity driven by periodic stimulation generally be considered functionally equivalent to endogenous rhythms?
Further progress in linking rhythms and cognition will come from an overarching theoretical understanding of rhythmic brain activity. Expanding the incomplete (yet rapidly increasing) understanding of how precisely brain rhythms are generated (for alpha, see Halgren et al., 2019; Lörincz et al., 2008, 2009; Nestvogel & McCormick, 2022) will help embedding the rhythmic sampling idea in a complex cortical reality abound with neural rhythms. This will also tackle oversimplifications such as referring to ‘the alpha rhythm’, although the brain likely generates more than one such rhythm at any given time, even in nearby cortical areas (Barzegaran et al., 2017; Benwell, London, et al., 2019; Chaumon & Busch, 2014; Keitel & Gross, 2016; Schaworonkow & Nikulin, 2022; Sokoliuk et al., 2019). Moreover, each rhythm occurs in the context of other periodic and aperiodic activity in the brain (Ibarra Chaoul & Siegel, 2021; Donoghue et al., 2021, this issue) and in brain–body interactions. For example, cardiac (Al et al., 2020), respiratory (Grund et al., 2022; Kluger et al., 2021), and gastric cycles (Richter et al., 2017) have all been proposed to impinge on cognitive processes. Stronger links between rhythms and cognitive function will therefore require more complex theories that stem from principles of large‐scale neuronal interactions, such as communication‐through‐coherence (Fries, 2015), oscillatory phase reset, and divisive normalisation (van Atteveldt et al., 2014). These can be explored in computational modelling (Alamia & VanRullen, 2019; Avramiea et al., 2020; Montijn et al., 2012) to derive stronger hypotheses to test in experimental data. Finally, these advances will need to be complemented with a deeper understanding of predictions of different aspects of periodic sampling to build stronger and more concrete links with rhythmic brain activity (Menétrey et al., 2021, this issue).
A different path forward may be using closed‐loop brain‐computer interfaces (BCIs) to draw stronger associations between brain rhythms and behaviour. Although this approach requires a high level of technical sophistication, it has been used in the past (Callaway & Yeager, 1960) and more recently to study rhythmic sampling (Ramot & Martin, 2022; Zrenner et al., 2016; Vigué‐Guix et al., 2020, this issue). One advantage is that closed‐loop BCIs reduce the degrees of freedom in the signal processing pipeline because the analysis must be conducted in real‐time; therefore, the features of interest and the parameters to extract them have to be set a priori. From a more theoretical perspective, a closed‐loop BCI approach allows researchers to determine the flow of an experiment (e.g., presentation of a visual stimulus) using critical brain states (e.g., alpha phase). This makes closed‐loop BCI experiments a rigorous benchmark for any assumed brain rhythm‐behaviour link, with the added potential of drawing stronger conclusions as to the causal role of neural rhythms in cognition (also see Ramot & Martin, 2022).
This Special Issue contributes to current attempts to counter publication biases (Chambers et al., 2015) and remedy the scarcity of published replications (e.g., the #EEGManyLabs project; Pavlov et al., 2021). For that purpose, we cannot stress enough the critical importance of giving studies with negative findings and replication attempts, irrespective of their outcome, an appropriate outlet in the literature. Another prevailing concern is the influence of analytical decisions on the robustness of published findings (Cohen, 2017). For instance, M/EEG studies typically use complex sequences of analysis steps on multi‐dimensional datasets. Results can therefore be influenced strongly by researchers' degrees of freedom at each step (Clayson et al., 2021). Methods for investigating rhythmic patterns in densely sampled behavioural data have also been criticised (Asamoah et al., 2019b; Brookshire, 2021; Ruhnau et al., 2020). In principle, several spectral analyses or statistical models can be run to identify which produce statistically significant results. Such questionable practices pander to the publication bias by producing findings that may be perceived as being more publishable (Luck & Gaspelin, 2017). In future research, it would therefore also be valuable to ‘revisit the evidence’ not only by obtaining new empirical data but by investigating the impact that analysis parameters have on the existing evidence (Botvinik‐Nezer et al., 2020; Clayson et al., 2021; Silberzahn et al., 2018).
In conclusion, whilst ‘revisiting the evidence’ for rhythms in cognition, and in particular the evidence for the periodic sampling idea, we found that this evidence remains equivocal, with interesting patterns emerging as to where and when to find it. As research follows cyclic patterns, too, we hope that these considerations come at a highly excitable time for new advances in our field.
CONFLICT OF INTERESTS
The authors declare that there were no competing interests with respect to the authorship or publication of this editorial.
AUTHOR CONTRIBUTIONS
All authors have drafted, revised, and have seen and approved the final version of the manuscript.
PEER REVIEW
The peer review history for this article is available at https://publons.com/publon/10.1111/ejn.15740.
ACKNOWLEDGEMENTS
The guest editors of this Special Issue would like to thank every author for submitting their work, all reviewers for spending their valuable time on commenting on submissions, and John J Foxe and Dana Helmreich, as well as the team at the EJN editorial office for their support.
Keitel, C. , Ruzzoli, M. , Dugué, L. , Busch, N. A. , & Benwell, C. S. Y. (2022). Rhythms in cognition: The evidence revisited. European Journal of Neuroscience, 55(11–12), 2991–3009. 10.1111/ejn.15740
Funding information M.R. was supported by Ministerio de Ciencia e Innovación (MICIIN), the Agencia Estatal de Investigación (AEI) under the Ramón y Cajal program (RYC2019‐027538‐I/0.13039/501100011033), and the Basque Foundation for Science (Ikerbasque). L.D. has received funding from the European Research Council (ERC) under the European Union's Horizon 2020 research and innovation program (grant agreement No 852139). L.D. and N.A.B have received funding from the Agence Nationale de la Recherche (ANR) ‐ Deutsche Forschungsgemeinschaft (DFG) program (grant agreement No J18P08ANR00 – L.D.; BU 2400/8‐1 – N.A.B.). C.S.Y.B was supported by the British Academy/Leverhulme Trust (SRG19/191169). All authors are members of the Scottish‐EU Critical Oscillations Network (SCONe), funded by the Royal Society of Edinburgh (RSE Saltire Facilitation Network Award to C.K., Reference Number 1963).
Edited by: Yoland Smith
Funding information Agence Nationale de la Recherche, Grant/Award Number: J18P08ANR00; Deutsche Forschungsgemeinschaft, Grant/Award Number: BU‐2400/8‐1; H2020 European Research Council, Grant/Award Number: 852139; Leverhulme Trust, Grant/Award Number: SRG19/191169; Ministerio de Ciencia e Innovación, Grant/Award Number: RYC2019‐027538‐I/0.13039/ 501100011033; Royal Society of Edinburgh, Grant/Award Number: RSE Saltire Facilitation Network Award to C.K., Reference Number 1963
Contributor Information
Christian Keitel, Email: christian.keitel@stir.ac.uk.
Manuela Ruzzoli, Email: m.ruzzoli@bcbl.eu.
Laura Dugué, Email: laura.dugue@u-paris.fr.
Niko A. Busch, Email: niko.busch@wwu.de.
Christopher S. Y. Benwell, Email: c.benwell@dundee.ac.uk.
REFERENCES
- Adrian, E. D. , & Matthews, B. H. C. (1934). The Berger rhythm: Potential changes from the occipital lobes in man. Brain: A Journal of Neurology, 57, 355–385. 10.1093/brain/57.4.355 [DOI] [PubMed] [Google Scholar]
- Al, E. , Iliopoulos, F. , Forschack, N. , Nierhaus, T. , Grund, M. , Motyka, P. , Gaebler, M. , Nikulin, V. V. , & Villringer, A. (2020). Heart–brain interactions shape somatosensory perception and evoked potentials. Proceedings of the National Academy of Sciences, 117(19), 10575–10584. 10.1073/pnas.1915629117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alamia, A. , & VanRullen, R. (2019). Alpha oscillations and traveling waves: Signatures of predictive coding? PLoS Biology, 17(10), e3000487. 10.1371/journal.pbio.3000487 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexandrou, A. M. , Saarinen, T. , Kujala, J. , & Salmelin, R. (2020). Cortical entrainment: What we can learn from studying naturalistic speech perception. Language, Cognition and Neuroscience, 35(6), 681–693. 10.1080/23273798.2018.1518534 [DOI] [Google Scholar]
- Asamoah, B. , Khatoun, A. , & Mc Laughlin, M. (2019a). tACS motor system effects can be caused by transcutaneous stimulation of peripheral nerves. Nature Communications, 10(1), 1–16. 10.1038/s41467-018-08183-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- Asamoah, B. , Khatoun, A. , & Mc Laughlin, M. (2019b). Analytical bias accounts for some of the reported effects of tACS on auditory perception. Brain Stimulation, 12(4), 1001–1009. 10.1016/j.brs.2019.03.011 [DOI] [PubMed] [Google Scholar]
- Avramiea, A.‐E. , Hardstone, R. , Lueckmann, J.‐M. , Bím, J. , Mansvelder, H. D. , & Linkenkaer‐Hansen, K. (2020). Pre‐stimulus phase and amplitude regulation of phase‐locked responses are maximized in the critical state. eLife, 9, e53016. 10.7554/eLife.53016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balestrieri, E. , Ronconi, L. , & Melcher, D. (2021). Shared resources between visual attention and visual working memory are allocated through rhythmic sampling. European Journal of Neuroscience, 55(11‐12), 3040–3053. 10.1111/ejn.15264 [DOI] [PubMed] [Google Scholar]
- Barne, L. C. , Cravo, A. M. , de Lange, F. P. , & Spaak, E. (2021). Temporal prediction elicits rhythmic preactivation of relevant sensory cortices. European Journal of Neuroscience, 55(11‐12), 3324–3339. 10.1111/ejn.15405 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barzegaran, E. , Vildavski, V. Y. , & Knyazeva, M. G. (2017). Fine structure of posterior alpha rhythm in human EEG: Frequency components, their cortical sources, and temporal behavior. Scientific Reports, 7(1), 8249. 10.1038/s41598-017-08421-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benwell, C. S. Y. , Coldea, A. , Harvey, M. , & Thut, G. (2021). Low pre‐stimulus EEG alpha power amplifies visual awareness but not visual sensitivity. European Journal of Neuroscience, 55(11‐12), 3125–3140. 10.1111/ejn.15166 [DOI] [PubMed] [Google Scholar]
- Benwell, C. S. Y. , Keitel, C. , Harvey, M. , Gross, J. , & Thut, G. (2019). Trial‐by‐trial co‐variation of pre‐stimulus EEG alpha power and visuospatial bias reflects a mixture of stochastic and deterministic effects. European Journal of Neuroscience, 48, 2566–2584. 10.1111/ejn.13688 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benwell, C. S. Y. , London, R. E. , Tagliabue, C. F. , Veniero, D. , Gross, J. , Keitel, C. , & Thut, G. (2019). Frequency and power of human alpha oscillations drift systematically with time‐on‐task. NeuroImage, 192, 101–114. 10.1016/j.neuroimage.2019.02.067 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benwell, C. S. Y. , Tagliabue, C. F. , Veniero, D. , Cecere, R. , Savazzi, S. , & Thut, G. (2017). Prestimulus EEG power predicts conscious awareness but not objective visual performance. Eneuro, 4(6), ENEURO.0182‐17.2017. 10.1523/ENEURO.0182-17.2017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bergmann, T. O. , & Hartwigsen, G. (2021). Inferring causality from noninvasive brain stimulation in cognitive neuroscience. Journal of Cognitive Neuroscience, 33(2), 195–225. 10.1162/jocn_a_01591 [DOI] [PubMed] [Google Scholar]
- Bergson, H. , & Frye, N. (1911). Creative evolution. Holt. http://archive.org/details/creativeevolu1st00berguoft [Google Scholar]
- Bishop, G. H. (1932). Cyclic changes in excitability of the optic pathway of the rabbit. American Journal of Physiology‐Legacy Content, 103(1), 213–224. 10.1152/ajplegacy.1932.103.1.213 [DOI] [Google Scholar]
- Botvinik‐Nezer, R. , Holzmeister, F. , Camerer, C. F. , Dreber, A. , Huber, J. , Johannesson, M. , Kirchler, M. , Iwanir, R. , Mumford, J. A. , Adcock, R. A. , Avesani, P. , Baczkowski, B. M. , Bajracharya, A. , Bakst, L. , Ball, S. , Barilari, M. , Bault, N. , Beaton, D. , Beitner, J. , … Schonberg, T. (2020). Variability in the analysis of a single neuroimaging dataset by many teams. Nature, 582(7810), 84–88. 10.1038/s41586-020-2314-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brookshire, G. (2021). Re‐evaluating rhythmic attentional switching: Spurious oscillations from shuffling‐in‐time (p. 2021.05.07.443101). bioRxiv. 10.1101/2021.05.07.443101 [DOI]
- Buergers, S. , & Noppeney, U. (2022). The role of alpha oscillations in temporal binding within and across the senses. Nature Human Behaviour, 6, 732–742. 10.1038/s41562-022-01294-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Busch, N. A. , Dubois, J. , & VanRullen, R. (2009). The phase of ongoing EEG oscillations predicts visual perception. Journal of Neuroscience, 29(24), 7869–7876. 10.1523/JNEUROSCI.0113-09.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Busch, N. A. , & VanRullen, R. (2014). Is visual perception like a continuous flow or a series of snapshots? In Subjective time: The philosophy, psychology, and neuroscience of temporality (pp. 161–178). Boston Review. 10.7551/mitpress/8516.001.0001 [DOI] [Google Scholar]
- Button, K. S. , Ioannidis, J. P. A. , Mokrysz, C. , Nosek, B. A. , Flint, J. , Robinson, E. S. J. , & Munafò, M. R. (2013). Power failure: Why small sample size undermines the reliability of neuroscience. Nature Reviews Neuroscience, 14(5), 365–376. 10.1038/nrn3475 [DOI] [PubMed] [Google Scholar]
- Buzsáki, G. , & Draguhn, A. (2004). Neuronal oscillations in cortical networks. Science, 304(5679), 1926–1929. 10.1126/science.1099745 [DOI] [PubMed] [Google Scholar]
- Calderone, D. J. , Lakatos, P. , Butler, P. D. , & Castellanos, F. X. (2014). Entrainment of neural oscillations as a modifiable substrate of attention. Trends in Cognitive Sciences, 18(6), 300–309. 10.1016/j.tics.2014.02.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Callaway, E. , & Yeager, C. L. (1960). Relationship between reaction time and electroencephalographic alpha phase. Science, 132(3441), 1765–1766. 10.1126/science.132.3441.1765 [DOI] [PubMed] [Google Scholar]
- Canolty, R. T. , & Knight, R. T. (2010). The functional role of cross‐frequency coupling. Trends in Cognitive Sciences, 14(11), 506–515. 10.1016/j.tics.2010.09.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Capilla, A. , Arana, L. , García‐Huéscar, M. , Melcón, M. , Gross, J. , & Campo, P. (2021). The natural frequencies of the resting human brain: An MEG‐based atlas (p. 2021.11.17.468973). bioRxiv. 10.1101/2021.11.17.468973 [DOI] [PubMed]
- Carrasco, M. (2011). Visual attention: The past 25 years. Vision Research, 51(13), 1484–1525. 10.1016/j.visres.2011.04.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cecere, R. , Rees, G. , & Romei, V. (2015). Individual differences in alpha frequency drive crossmodal illusory perception. Current Biology, 25(2), 231–235. 10.1016/j.cub.2014.11.034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chakravarthi, R. , & VanRullen, R. (2012). Conscious updating is a rhythmic process. Proceedings of the National Academy of Sciences, 109(26), 10599–10604. 10.1073/pnas.1121622109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chambers, C. D. , Dienes, Z. , McIntosh, R. D. , Rotshtein, P. , & Willmes, K. (2015). Registered Reports: Realigning incentives in scientific publishing. Cortex, 66, A1–A2. 10.1016/j.cortex.2015.03.022 [DOI] [PubMed] [Google Scholar]
- Chaumon, M. , & Busch, N. A. (2014). Prestimulus neural oscillations inhibit visual perception via modulation of response gain. Journal of Cognitive Neuroscience, 26(11), 2514–2529. 10.1162/jocn_a_00653 [DOI] [PubMed] [Google Scholar]
- Chen, A. , Zu, G. , Dong, B. , & Zhang, M. (2020). Cortical distance but not physical distance modulates attentional rhythms. Frontiers in Psychology, 11, 541085. 10.3389/fpsyg.2020.541085 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clayson, P. E. , Baldwin, S. A. , Rocha, H. A. , & Larson, M. J. (2021). The data‐processing multiverse of event‐related potentials (ERPs): A roadmap for the optimization and standardization of ERP processing and reduction pipelines. NeuroImage, 245, 118712. 10.1016/j.neuroimage.2021.118712 [DOI] [PubMed] [Google Scholar]
- Clayton, M. S. , Yeung, N. , & Cohen Kadosh, R. (2018). The many characters of visual alpha oscillations. European Journal of Neuroscience, 48(7), 2498–2508. 10.1111/ejn.13747 [DOI] [PubMed] [Google Scholar]
- Cohen, M. X. (2014). Analyzing neural time series data: Theory and practice. MIT Press. [Google Scholar]
- Cohen, M. X. (2017). Rigor and replication in time‐frequency analyses of cognitive electrophysiology data. International Journal of Psychophysiology, 111, 80–87. 10.1016/j.ijpsycho.2016.02.001 [DOI] [PubMed] [Google Scholar]
- Dahl, M. J. , Mather, M. , & Werkle‐Bergner, M. (2022). Noradrenergic modulation of rhythmic neural activity shapes selective attention. Trends in Cognitive Sciences, 26(1), 38–52. 10.1016/j.tics.2021.10.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Graaf, T. A. , & Duecker, F. (2021). No effects of rhythmic visual stimulation on target discrimination: An online alpha entrainment experiment. European Journal of Neuroscience, 55(11‐12), 3340–3351. 10.1111/ejn.15483 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donoghue, T. , Schaworonkow, N. , & Voytek, B. (2021). Methodological considerations for studying neural oscillations. European Journal of Neuroscience, 55(11‐12), 3502–3527. 10.1111/ejn.15361 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duecker, K. , Gutteling, T. P. , Herrmann, C. S. , & Jensen, O. (2021). No evidence for entrainment: Endogenous gamma oscillations and rhythmic flicker responses coexist in visual cortex. The Journal of Neuroscience, 41(31), 6684–6698. 10.1523/jneurosci.3134-20.2021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dugué, L. , Beck, A.‐A. , Marque, P. , & VanRullen, R. (2019). Contribution of FEF to attentional periodicity during visual search: A TMS study. ENeuro, 6(3), ENEURO.0357–ENEU18.2019. 10.1523/ENEURO.0357-18.2019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dugué, L. , McLelland, D. , Lajous, M. , & VanRullen, R. (2015). Attention searches nonuniformly in space and in time. Proceedings of the National Academy of Sciences, 112(49), 15214–15219. 10.1073/pnas.1511331112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dugué, L. , & VanRullen, R. (2017). Transcranial magnetic stimulation reveals intrinsic perceptual and attentional rhythms. Frontiers in Neuroscience, 11, 154. 10.3389/fnins.2017.00154 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Egger, S. W. , Le, N. M. , & Jazayeri, M. (2020). A neural circuit model for human sensorimotor timing. Nature Communications, 11(1), 3933. 10.1038/s41467-020-16999-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Egly, R. , Driver, J. , & Rafal, R. D. (1994). Shifting visual attention between objects and locations: Evidence from normal and parietal lesion subjects. Journal of Experimental Psychology. General, 123(2), 161–177. 10.1037//0096-3445.123.2.161 [DOI] [PubMed] [Google Scholar]
- Fakche, C. , VanRullen, R. , Marque, P. , & Dugué, L. (2022). α phase‐amplitude tradeoffs predict visual perception. ENeuro, 9(1), ENEURO.0244–ENEU21.2022. 10.1523/ENEURO.0244-21.2022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fekete, T. , Van de Cruys, S. , Ekroll, V. , & van Leeuwen, C. (2018). In the interest of saving time: A critique of discrete perception. Neuroscience of Consciousness, 2018(1), niy003. 10.1093/nc/niy003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fiebelkorn, I. C. (2021). Detecting attention‐related rhythms: When is behavior not enough? (Commentary on van der Werf et al. 2021). European Journal of Neuroscience, 55(11‐12), 3117–3120. 10.1111/ejn.15322 [DOI] [PubMed] [Google Scholar]
- Fiebelkorn, I. C. , Pinsk, M. A. , & Kastner, S. (2018). A dynamic interplay within the frontoparietal network underlies rhythmic spatial attention. Neuron, 99(4), 842–853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fiebelkorn, I. C. , Saalmann, Y. B. , & Kastner, S. (2013). Rhythmic sampling within and between objects despite sustained attention at a cued location. Current Biology, 23(24), 2553–2558. 10.1016/j.cub.2013.10.063 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fries, P. (2015). Rhythms for cognition: Communication through coherence. Neuron, 88(1), 220–235. 10.1016/j.neuron.2015.09.034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaillard, C. , & Ben Hamed, S. (2020). The neural bases of spatial attention and perceptual rhythms. European Journal of Neuroscience, 55(11‐12), 3209–3223. 10.1111/ejn.15044 [DOI] [PubMed] [Google Scholar]
- García‐Rosales, F. , López‐Jury, L. , González‐Palomares, E. , Cabral‐Calderín, Y. , Kössl, M. , & Hechavarria, J. C. (2020). Phase‐amplitude coupling profiles differ in frontal and auditory cortices of bats. European Journal of Neuroscience, 55(11‐12), 3483–3501. 10.1111/ejn.14986 [DOI] [PubMed] [Google Scholar]
- Garrett‐Ruffin, S. , Hindash, A. C. , Kaczkurkin, A. N. , Mears, R. P. , Morales, S. , Paul, K. , Pavlov, Y. G. , & Keil, A. (2021). Open science in psychophysiology: An overview of challenges and emerging solutions. International Journal of Psychophysiology, 162, 69–78. 10.1016/j.ijpsycho.2021.02.005 [DOI] [PubMed] [Google Scholar]
- Giraud, A.‐L. , & Poeppel, D. (2012). Cortical oscillations and speech processing: Emerging computational principles and operations. Nature Neuroscience, 15(4), 511–517. 10.1038/nn.3063 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gray, M. J. , & Emmanouil, T. A. (2019). Individual alpha frequency increases during a task but is unchanged by alpha‐band flicker. Psychophysiology, 57(2), e13480. 10.1111/psyp.13480 [DOI] [PubMed] [Google Scholar]
- Groppe, D. M. , Bickel, S. , Keller, C. J. , Jain, S. K. , Hwang, S. T. , Harden, C. , & Mehta, A. D. (2013). Dominant frequencies of resting human brain activity as measured by the electrocorticogram. NeuroImage, 79, 223–233. 10.1016/j.neuroimage.2013.04.044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grund, M. , Al, E. , Pabst, M. , Dabbagh, A. , Stephani, T. , Nierhaus, T. , Gaebler, M. , & Villringer, A. (2022). Respiration, heartbeat, and conscious tactile perception. Journal of Neuroscience, 42(4), 643–656. 10.1523/JNEUROSCI.0592-21.2021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haegens, S. (2020). Entrainment revisited: A commentary on Meyer, Sun, and Martin (2020). Language, Cognition and Neuroscience, 35(9), 1119–1123. 10.1080/23273798.2020.1758335 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halgren, M. , Ulbert, I. , Bastuji, H. , Fabó, D. , Erőss, L. , Rey, M. , Devinsky, O. , Doyle, W. K. , Mak‐McCully, R. , Halgren, E. , Wittner, L. , Chauvel, P. , Heit, G. , Eskandar, E. , Mandell, A. , & Cash, S. S. (2019). The generation and propagation of the human alpha rhythm. Proceedings of the National Academy of Sciences, 116(47), 23772–23782. 10.1073/pnas.1913092116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hauswald, A. , Keitel, A. , Chen, Y.‐P. , Rösch, S. , & Weisz, N. (2020). Degradation levels of continuous speech affect neural speech tracking and alpha power differently. European Journal of Neuroscience, 55(11‐12), 3288–3302. 10.1111/ejn.14912 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Helfrich, R. F. , Fiebelkorn, I. C. , Szczepanski, S. M. , Lin, J. J. , Parvizi, J. , Knight, R. T. , & Kastner, S. (2018). Neural mechanisms of sustained attention are rhythmic. Neuron, 99(4), 854–865.e5. 10.1016/j.neuron.2018.07.032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Helfrich, R. F. , Schneider, T. R. , Rach, S. , Trautmann‐Lengsfeld, S. A. , Engel, A. K. , & Herrmann, C. S. (2014). Entrainment of brain oscillations by transcranial alternating current stimulation. Current Biology, 24(3), 333–339. 10.1016/j.cub.2013.12.041 [DOI] [PubMed] [Google Scholar]
- Henry, M. J. , & Herrmann, B. (2014). Low‐frequency neural oscillations support dynamic attending in temporal context. Timing & Time Perception, 2(1), 62–86. 10.1163/22134468-00002011 [DOI] [Google Scholar]
- Herrmann, C. S. , Murray, M. M. , Ionta, S. , Hutt, A. , & Lefebvre, J. (2016). Shaping intrinsic neural oscillations with periodic stimulation. Journal of Neuroscience, 36(19), 5328–5337. 10.1523/JNEUROSCI.0236-16.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herzog, M. H. , Drissi‐Daoudi, L. , & Doerig, A. (2020). All in good time: Long‐lasting postdictive effects reveal discrete perception. Trends in Cognitive Sciences, 24(10), 826–837. 10.1016/j.tics.2020.07.001 [DOI] [PubMed] [Google Scholar]
- Hickok, G. , Farahbod, H. , & Saberi, K. (2015). The rhythm of perception: Entrainment to acoustic rhythms induces subsequent perceptual oscillation. Psychological Science, 26(7), 1006–1013. 10.1177/0956797615576533 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ho, H. T. , Burr, D. C. , Alais, D. , & Morrone, M. C. (2021). Propagation and update of auditory perceptual priors through alpha and theta rhythms. European Journal of Neuroscience, 55(11‐12), 3083–3099. 10.1111/ejn.15141 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ho, H. T. , Leung, J. , Burr, D. C. , Alais, D. , & Morrone, M. C. (2017). Auditory sensitivity and decision criteria oscillate at different frequencies separately for the two ears. Current Biology, 27(23), 3643–3649.e3. 10.1016/j.cub.2017.10.017 [DOI] [PubMed] [Google Scholar]
- Huang, W. A. , Stitt, I. M. , Negahbani, E. , Passey, D. J. , Ahn, S. , Davey, M. , Dannhauer, M. , Doan, T. T. , Hoover, A. C. , Peterchev, A. V. , Radtke‐Schuller, S. , & Fröhlich, F. (2021). Transcranial alternating current stimulation entrains alpha oscillations by preferential phase synchronization of fast‐spiking cortical neurons to stimulation waveform. Nature Communications, 12(1), 1–20. 10.1038/s41467-021-23021-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iaccarino, H. F. , Singer, A. C. , Martorell, A. J. , Rudenko, A. , Gao, F. , Gillingham, T. Z. , Mathys, H. , Seo, J. , Kritskiy, O. , Abdurrob, F. , Adaikkan, C. , Canter, R. G. , Rueda, R. , Brown, E. N. , Boyden, E. S. , & Tsai, L.‐H. (2016). Gamma frequency entrainment attenuates amyloid load and modifies microglia. Nature, 540(7632), 230–235. 10.1038/nature20587 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ibarra Chaoul, A. , & Siegel, M. (2021). Cortical correlation structure of aperiodic neuronal population activity. NeuroImage, 245, 118672. 10.1016/j.neuroimage.2021.118672 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janssens, S. E. W. , Sack, A. T. , Oever, S. T. , & de Graaf, T. A. (2021). Calibrating rhythmic stimulation parameters to individual electroencephalography markers: The consistency of individual alpha frequency in practical lab settings. European Journal of Neuroscience, 55(11‐12), 3418–3437. 10.1111/ejn.15418 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jensen, O. , & Colgin, L. L. (2007). Cross‐frequency coupling between neuronal oscillations. Trends in Cognitive Sciences, 11(7), 267–269. 10.1016/j.tics.2007.05.003 [DOI] [PubMed] [Google Scholar]
- Jones, M. R. , & Boltz, M. (1989). Dynamic attending and responses to time. Psychological Review, 96(3), 459–491. 10.1037/0033-295x.96.3.459 [DOI] [PubMed] [Google Scholar]
- Kasten, F. H. , & Herrmann, C. S. (2020). Discrete sampling in perception via neuronal oscillations—Evidence from rhythmic, non‐invasive brain stimulation. European Journal of Neuroscience, 55(11‐12), 3402–3417. 10.1111/ejn.15006 [DOI] [PubMed] [Google Scholar]
- Keil, J. , & Senkowski, D. (2017). Individual alpha frequency relates to the sound‐induced flash illusion. Multisensory Research, 30(6), 565–578. [DOI] [PubMed] [Google Scholar]
- Keitel, A. , & Gross, J. (2016). Individual human brain areas can be identified from their characteristic spectral activation fingerprints. PLoS Biology, 14(6), e1002498. 10.1371/journal.pbio.1002498 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keitel, C. , Obleser, J. , Jessen, S. , & Henry, M. J. (2021). Frequency‐specific effects in infant electroencephalograms do not require entrained neural oscillations: A commentary on Köster et al. (2019). Psychological Science, 32(6), 966–971. 10.1177/09567976211001317 [DOI] [PubMed] [Google Scholar]
- Kienitz, R. , Schmid, M. C. , & Dugué, L. (2021). Rhythmic sampling revisited: Experimental paradigms and neural mechanisms. European Journal of Neuroscience, 55(11‐12), 3010–3024. 10.1111/ejn.15489 [DOI] [PubMed] [Google Scholar]
- Kluger, D. S. , Balestrieri, E. , Busch, N. A. , & Gross, J. (2021). Respiration aligns perception with neural excitability. eLife, 10, e70907. 10.7554/eLife.70907 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lakatos, P. , Gross, J. , & Thut, G. (2019). A new unifying account of the roles of neuronal entrainment. Current Biology, 29(18), R890–R905. 10.1016/j.cub.2019.07.075 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lakatos, P. , Musacchia, G. , O9Connel, M. N. , Falchier, A. Y. , Javitt, D. C. , & Schroeder, C. E. (2013). The spectrotemporal filter mechanism of auditory selective attention. Neuron, 77(4), 750–761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Landau, A. N. , & Fries, P. (2012). Attention samples stimuli rhythmically. Current Biology, 22(11), 1000–1004. 10.1016/j.cub.2012.03.054 [DOI] [PubMed] [Google Scholar]
- Latour, P. L. (1967). Evidence of internal clocks in the human operator. Acta Psychologica, 27, 341–348. 10.1016/0001-6918(67)90078-9 [DOI] [PubMed] [Google Scholar]
- Lin, W. M. , Oetringer, D. A. , Bakker‐Marshall, I. , Emmerzaal, J. , Wilsch, A. , ElShafei, H. A. , Rassi, E. , & Haegens, S. (2021). No behavioural evidence for rhythmic facilitation of perceptual discrimination. European Journal of Neuroscience, 55(11‐12), 3352–3364. 10.1111/ejn.15208 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin, Y.‐J. , Shukla, L. , Dugué, L. , Valero‐Cabré, A. , & Carrasco, M. (2021). Transcranial magnetic stimulation entrains alpha oscillatory activity in occipital cortex. Scientific Reports, 11(1), 18562. 10.1038/s41598-021-96849-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- London, R. E. , Benwell, C. S. Y. , Cecere, R. , Quak, M. , Thut, G. , & Talsma, D. (2022). EEG Alpha power predicts the temporal sensitivity of multisensory perception. European Journal of Neuroscience, 55(11‐12), 3241–3255. 10.1111/ejn.15719 [DOI] [PubMed] [Google Scholar]
- Lörincz, M. L. , Crunelli, V. , & Hughes, S. W. (2008). Cellular dynamics of cholinergically induced α (8–13 Hz) rhythms in sensory thalamic nuclei in vitro. Journal of Neuroscience, 28(3), 660–671. 10.1523/JNEUROSCI.4468-07.2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lörincz, M. L. , Kékesi, K. A. , Juhász, G. , Crunelli, V. , & Hughes, S. W. (2009). Temporal framing of thalamic relay‐mode firing by phasic inhibition during the alpha rhythm. Neuron, 63(5), 683–696. 10.1016/j.neuron.2009.08.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luck, S. J. , & Gaspelin, N. (2017). How to get statistically significant effects in any ERP experiment (and why you shouldn't). Psychophysiology, 54(1), 146–157. 10.1111/psyp.12639 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mathewson, K. E. , Gratton, G. , Fabiani, M. , Beck, D. M. , & Ro, T. (2009). To see or not to see: Prestimulus α phase predicts visual awareness. Journal of Neuroscience, 29(9), 2725–2732. 10.1523/JNEUROSCI.3963-08.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menétrey, M. Q. , Vogelsang, L. , & Herzog, M. H. (2021). A guideline for linking brain wave findings to the various aspects of discrete perception. European Journal of Neuroscience, 55(11‐12), 3528–3537. 10.1111/ejn.15349 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Merholz, G. , Grabot, L. , VanRullen, R. , & Dugué, L. (2022). Periodic attention operates faster during more complex visual search. Scientific Reports, 12(1), 6688. 10.1038/s41598-022-10647-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer, L. , Sun, Y. , & Martin, A. E. (2020). Synchronous, but not entrained: Exogenous and endogenous cortical rhythms of speech and language processing. Language, Cognition and Neuroscience, 35(9), 1089–1099. 10.1080/23273798.2019.1693050 [DOI] [Google Scholar]
- Michail, G. , Toran Jenner, L. , & Keil, J. (2021). Prestimulus alpha power but not phase influences visual discrimination of long‐duration visual stimuli. European Journal of Neuroscience, 55(11‐12), 3141–3153. 10.1111/ejn.15169 [DOI] [PubMed] [Google Scholar]
- Michel, R. , Dugué, L. , & Busch, N. A. (2021). Distinct contributions of alpha and theta rhythms to perceptual and attentional sampling. European Journal of Neuroscience, 55(11‐12), 3025–3039. 10.1111/ejn.15154 [DOI] [PubMed] [Google Scholar]
- Milton, A. , & Pleydell‐Pearce, C. W. (2016). The phase of pre‐stimulus alpha oscillations influences the visual perception of stimulus timing. NeuroImage, 133, 53–61. 10.1016/j.neuroimage.2016.02.065 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Montijn, J. , Klink, P. , & Van Wezel, R. (2012). Divisive normalization and neuronal oscillations in a single hierarchical framework of selective visual attention. Frontiers in Neural Circuits, 6, 22. 10.3389/fncir.2012.00022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morrow, A. , & Samaha, J. (2021). No evidence for a single oscillator underlying discrete visual percepts. European Journal of Neuroscience, 55(11‐12), 3054–3066. 10.1111/ejn.15362 [DOI] [PubMed] [Google Scholar]
- Nestvogel, D. B. , & McCormick, D. A. (2022). Visual thalamocortical mechanisms of waking state‐dependent activity and alpha oscillations. Neuron, 110(1), 120–138.e4. 10.1016/j.neuron.2021.10.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ngiam, W. X. Q. , Adam, K. C. S. , Quirk, C. , Vogel, E. K. , & Awh, E. (2021). Estimating the statistical power to detect set‐size effects in contralateral delay activity. Psychophysiology, 58(5), e13791. 10.1111/psyp.13791 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niso, G. , Krol, L. R. , Combrisson, E. , Dubarry, A.‐S. , Elliott, M. A. , François, C. , Héjja‐Brichard, Y. , Herbst, S. , Jerbi, K. , Kovic, V. , Lehongre, K. , Luck, S. J. , Mercier, M. , Mosher, J. C. , Pavlov, Y. G. , Puce, A. , Schettino, A. , Daniele, S. , Sinnott‐Armstrong, W. , … Chaumon, M. (2021). Good scientific practice in MEEG research: Progress and perspectives. OSF Preprints. 10.31219/osf.io/n2ryp [DOI] [PMC free article] [PubMed]
- O'Hare, J. (1954). The variability of auditory and visual reaction time with change in amplitude and phase of alpha rhythm. American Psychologist, 9, 444. [Google Scholar]
- Obleser, J. , & Kayser, C. (2019). Neural entrainment and attentional selection in the listening brain. Trends in Cognitive Sciences, 23, 913–926. 10.1016/j.tics.2019.08.004 [DOI] [PubMed] [Google Scholar]
- Ono, K. , Hashimoto, J. , & Sasaoka, T. (2021). Intertap interval dependence of the subdivision effect in auditory‐synchronised tapping. European Journal of Neuroscience, 55(11‐12), 3391–3401. 10.1111/ejn.15529 [DOI] [PubMed] [Google Scholar]
- Open Science Collaboration . (2015). Estimating the reproducibility of psychological science. Science, 349(6251), aac4716. 10.1126/science.aac4716 [DOI] [PubMed] [Google Scholar]
- Pavlov, Y. G. , Adamian, N. , Appelhoff, S. , Arvaneh, M. , Benwell, C. S. Y. , Beste, C. , Bland, A. R. , Bradford, D. E. , Bublatzky, F. , Busch, N. A. , Clayson, P. E. , Cruse, D. , Czeszumski, A. , Dreber, A. , Dumas, G. , Ehinger, B. , Ganis, G. , He, X. , Hinojosa, J. A. , … Mushtaq, F. (2021). #EEGManyLabs: Investigating the replicability of influential EEG experiments. Cortex, 144, 213–229. 10.1016/j.cortex.2021.03.013 [DOI] [PubMed] [Google Scholar]
- Pfeffer, T. , Keitel, C. , Kluger, D. S. , Keitel, A. , Russmann, A. , Thut, G. , Donner, T. H. , & Gross, J. (2022). Coupling of pupil‐ and neuronal population dynamics reveals diverse influences of arousal on cortical processing. eLife, 11, e71890. 10.7554/eLife.71890 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pitts, W. , & McCulloch, W. S. (1947). How we know universals the perception of auditory and visual forms. The Bulletin of Mathematical Biophysics, 9(3), 127–147. 10.1007/BF02478291 [DOI] [PubMed] [Google Scholar]
- Plöchl, M. , Fiebelkorn, I. , Kastner, S. , & Obleser, J. (2021). Attentional sampling of visual and auditory objects is captured by theta‐modulated neural activity. European Journal of Neuroscience, 55(11‐12), 3067–3082. 10.1111/ejn.15514 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Purushothaman, G. , Oğmen, H. , & Bedell, H. E. (2000). Gamma‐range oscillations in backward‐masking functions and their putative neural correlates. Psychological Review, 107(3), 556–577. 10.1037/0033-295x.107.3.556 [DOI] [PubMed] [Google Scholar]
- Quigley, C. (2021). Forgotten rhythms? Revisiting the first evidence for rhythms in cognition. European Journal of Neuroscience, 55(11‐12), 3266–3276. 10.1111/ejn.15450 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramot, M. , & Martin, A. (2022). Closed‐loop neuromodulation for studying spontaneous activity and causality. Trends in Cognitive Sciences, 26(4), 290–299. 10.1016/j.tics.2022.01.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rassili, O. , & Ordin, M. (2020). The effect of regular rhythm on the perception of linguistic and non‐linguistic auditory input. European Journal of Neuroscience, 55(11‐12), 3365–3372. 10.1111/ejn.15029 [DOI] [PubMed] [Google Scholar]
- Repp, B. H. (2005). Sensorimotor synchronization: A review of the tapping literature. Psychonomic Bulletin & Review, 12(6), 969–992. 10.3758/BF03206433 [DOI] [PubMed] [Google Scholar]
- Repp, B. H. , & Su, Y.‐H. (2013). Sensorimotor synchronization: A review of recent research (2006–2012). Psychonomic Bulletin & Review, 20(3), 403–452. 10.3758/s13423-012-0371-2 [DOI] [PubMed] [Google Scholar]
- Richter, C. G. , Babo‐Rebelo, M. , Schwartz, D. , & Tallon‐Baudry, C. (2017). Phase‐amplitude coupling at the organism level: The amplitude of spontaneous alpha rhythm fluctuations varies with the phase of the infra‐slow gastric basal rhythm. NeuroImage, 146, 951–958. 10.1016/j.neuroimage.2016.08.043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riddle, J. , & Frohlich, F. (2021). Targeting neural oscillations with transcranial alternating current stimulation. Brain Research, 1765, 147491. 10.1016/j.brainres.2021.147491 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rimmele, J. M. , Zion Golumbic, E. , Schröger, E. , & Poeppel, D. (2015). The effects of selective attention and speech acoustics on neural speech‐tracking in a multi‐talker scene. Cortex, 68, 144–154. 10.1016/j.cortex.2014.12.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ronconi, L. , Melcher, D. , Junghöfer, M. , Wolters, C. H. , & Busch, N. A. (2020). Testing the effect of tACS over parietal cortex in modulating endogenous alpha rhythm and temporal integration windows in visual perception. European Journal of Neuroscience, 55(11‐12), 3438–3450. 10.1111/ejn.15017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ronconi, L. , Oosterhof, N. N. , Bonmassar, C. , & Melcher, D. (2017). Multiple oscillatory rhythms determine the temporal organization of perception. Proceedings of the National Academy of Sciences of the United States of America, 114(51), 13435–13440. 10.1073/pnas.1714522114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruhnau, P. , Rufener, K. S. , Heinze, H.‐J. , & Zaehle, T. (2020). Pulsed transcranial electric brain stimulation enhances speech comprehension. Brain Stimulation, 13(5), 1402–1411. 10.1016/j.brs.2020.07.011 [DOI] [PubMed] [Google Scholar]
- Ruzzoli, M. , Torralba, M. , Morís Fernández, L. , & Soto‐Faraco, S. (2019). The relevance of alpha phase in human perception. Cortex, 120, 249–268. 10.1016/j.cortex.2019.05.012 [DOI] [PubMed] [Google Scholar]
- Saberi, K. , & Hickok, G. (2021). Forward entrainment: Evidence, controversies, constraints, and mechanisms (p. 2021.07.06.451373). bioRxiv. 10.1101/2021.07.06.451373 [DOI]
- Samaha, J. , Iemi, L. , & Postle, B. R. (2017). Prestimulus alpha‐band power biases visual discrimination confidence, but not accuracy. Consciousness and Cognition, 54, 47–55. 10.1016/j.concog.2017.02.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Samaha, J. , & Postle, B. R. (2015). The speed of alpha‐band oscillations predicts the temporal resolution of visual perception. Current Biology, 25(22), 2985–2990. 10.1016/j.cub.2015.10.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schaworonkow, N. , & Nikulin, V. V. (2022). Is sensor space analysis good enough? Spatial patterns as a tool for assessing spatial mixing of EEG/MEG rhythms. NeuroImage, 253, 119093. 10.1016/j.neuroimage.2022.119093 [DOI] [PubMed] [Google Scholar]
- Schneider, D. , Herbst, S. K. , Klatt, L.‐I. , & Wöstmann, M. (2021). Target enhancement or distractor suppression? Functionally distinct alpha oscillations form the basis of attention. European Journal of Neuroscience, 55(11‐12), 3256–3265. 10.1111/ejn.15309 [DOI] [PubMed] [Google Scholar]
- Schneider, K. A. (2018). The flash‐lag, Fröhlich and related motion illusions are natural consequences of discrete sampling in the visual system. Frontiers in Psychology, 9, 1227. 10.3389/fpsyg.2018.01227 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schroeder, C. E. , & Lakatos, P. (2009). Low‐frequency neuronal oscillations as instruments of sensory selection. Trends in Neurosciences, 32(1), 9–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schroeder, C. E. , Wilson, D. A. , Radman, T. , Scharfman, H. , & Lakatos, P. (2010). Dynamics of active sensing and perceptual selection. Current Opinion in Neurobiology, 20(2), 172–176. 10.1016/j.conb.2010.02.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Senoussi, M. , Moreland, J. C. , Busch, N. A. , & Dugué, L. (2019). Attention explores space periodically at the theta frequency. Journal of Vision, 19(5), 22. 10.1167/19.5.22 [DOI] [PubMed] [Google Scholar]
- Sheldon, S. S. , & Mathewson, K. E. (2021). To see, not to see, or to see poorly: Perceptual quality and guess rate as a function of electroencephalography (EEG) brain activity in an orientation perception task. European Journal of Neuroscience, 55(11‐12), 3154–3177. 10.1111/ejn.15445 [DOI] [PubMed] [Google Scholar]
- Silberzahn, R. , Uhlmann, E. L. , Martin, D. P. , Anselmi, P. , Aust, F. , Awtrey, E. , Bahník, Š. , Bai, F. , Bannard, C. , Bonnier, E. , Carlsson, R. , Cheung, F. , Christensen, G. , Clay, R. , Craig, M. A. , Dalla Rosa, A. , Dam, L. , Evans, M. H. , Flores Cervantes, I. , … Nosek, B. A. (2018). Many analysts, one data set: Making transparent how variations in analytic choices affect results. Advances in Methods and Practices in Psychological Science, 1(3), 337–356. 10.1177/2515245917747646 [DOI] [Google Scholar]
- Sokoliuk, R. , Mayhew, S. D. , Aquino, K. M. , Wilson, R. , Brookes, M. J. , Francis, S. T. , Hanslmayr, S. , & Mullinger, K. J. (2019). Two spatially distinct posterior alpha sources fulfill different functional roles in attention. Journal of Neuroscience, 39(36), 7183–7194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sokoliuk, R. , & VanRullen, R. (2019). Perceptual illusions caused by discrete sampling. In Arstila V., Bardon A., Power S. E., & Vatakis A. (Eds.), The illusions of time: Philosophical and psychological essays on timing and time perception (pp. 315–338). Springer International Publishing. 10.1007/978-3-030-22048-8_17 [DOI] [Google Scholar]
- Staresina, B. P. , Bergmann, T. O. , Bonnefond, M. , van der Meij, R. , Jensen, O. , Deuker, L. , Elger, C. E. , Axmacher, N. , & Fell, J. (2015). Hierarchical nesting of slow oscillations, spindles and ripples in the human hippocampus during sleep. Nature Neuroscience, 18(11), 1679–1686. 10.1038/nn.4119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun, Y. , Michalareas, G. , & Poeppel, D. (2021). The impact of phase entrainment on auditory detection is highly variable: Revisiting a key finding. European Journal of Neuroscience, 55(11‐12), 3373–3390. 10.1111/ejn.15367 [DOI] [PubMed] [Google Scholar]
- Szucs, D. , & Ioannidis, J. P. A. (2017). Empirical assessment of published effect sizes and power in the recent cognitive neuroscience and psychology literature. PLoS Biology, 15(3), e2000797. 10.1371/journal.pbio.2000797 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tavano, A. , Maess, B. , Poeppel, D. , & Schröger, E. (2022). Neural entrainment via perceptual inferences. European Journal of Neuroscience, 55(11‐12), 3277–3287. 10.1111/ejn.15630 [DOI] [PubMed] [Google Scholar]
- ten Oever, S. , van der Werf, O. J. , Schuhmann, T. , & Sack, A. T. (2022). Absence of behavioural rhythms: Noise or unexplained neuronal mechanisms? (response to Fiebelkorn, 2021). European Journal of Neuroscience, 55(11‐12), 3121–3124. 10.1111/ejn.15628 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Theeuwes, J. (1992). Perceptual selectivity for color and form. Perception & Psychophysics, 51(6), 599–606. 10.3758/BF03211656 [DOI] [PubMed] [Google Scholar]
- Thut, G. , Schyns, P. , & Gross, J. (2011). Entrainment of perceptually relevant brain oscillations by non‐invasive rhythmic stimulation of the human brain. Frontiers in Psychology, 2, 170. 10.3389/fpsyg.2011.00170 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thut, G. , Veniero, D. , Romei, V. , Miniussi, C. , Schyns, P. , & Gross, J. (2011). Rhythmic TMS causes local entrainment of natural oscillatory signatures. Current Biology, 21(14), 1176–1185. 10.1016/j.cub.2011.05.049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tichko, P. , Kim, J. C. , Large, E. , & Loui, P. (2020). Integrating music‐based interventions with gamma‐frequency stimulation: Implications for healthy ageing. European Journal of Neuroscience, 55(11‐12), 3303–3323. 10.1111/ejn.15059 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tuladhar, A. M. , ter Huurne, N. , Schoffelen, J.‐M. , Maris, E. , Oostenveld, R. , & Jensen, O. (2007). Parieto‐occipital sources account for the increase in alpha activity with working memory load. Human Brain Mapping, 28(8), 785–792. 10.1002/hbm.20306 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tune, S. , Alavash, M. , Fiedler, L. , & Obleser, J. (2021). Neural attentional‐filter mechanisms of listening success in middle‐aged and older individuals. Nature Communications, 12(1), 4533. 10.1038/s41467-021-24771-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Atteveldt, N. , Murray, M. M. , Thut, G. , & Schroeder, C. E. (2014). Multisensory Integration: Flexible use of general operations. Neuron, 81(6), 1240–1253. 10.1016/j.neuron.2014.02.044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Bree, S. , Sohoglu, E. , Davis, M. H. , & Zoefel, B. (2021). Sustained neural rhythms reveal endogenous oscillations supporting speech perception. PLoS Biology, 19(2), e3001142. 10.1371/journal.pbio.3001142 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Plas, M. , Wang, D. , Brittain, J.‐S. , & Hanslmayr, S. (2020). Investigating the role of phase‐synchrony during encoding of episodic memories using electrical stimulation. Cortex, 133, 37–47. 10.1016/j.cortex.2020.09.006 [DOI] [PubMed] [Google Scholar]
- van Der Werf, O. J. , Ten Oever, S. , Schuhmann, T. , & Sack, A. T. (2021). No evidence of rhythmic visuospatial attention at cued locations in a spatial cuing paradigm, regardless of their behavioural relevance. European Journal of Neuroscience, 55(11‐12), 3100–3116. 10.1111/ejn.15353 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Diepen, R. M. , Cohen, M. X. , Denys, D. , & Mazaheri, A. (2015). Attention and temporal expectations modulate power, not phase, of ongoing alpha oscillations. Journal of Cognitive Neuroscience, 27(8), 1573–1586. 10.1162/jocn_a_00803 [DOI] [PubMed] [Google Scholar]
- Van Diepen, R. M. , Foxe, J. J. , & Mazaheri, A. (2019). The functional role of alpha‐band activity in attentional processing: The current zeitgeist and future outlook. Current Opinion in Psychology, 29, 229–238. 10.1016/j.copsyc.2019.03.015 [DOI] [PubMed] [Google Scholar]
- van Es, M. W. J. , Marshall, T. R. , Spaak, E. , Jensen, O. , & Schoffelen, J.‐M. (2020). Phasic modulation of visual representations during sustained attention. European Journal of Neuroscience, 55(11‐12), 3191–3208. 10.1111/ejn.15084 [DOI] [PMC free article] [PubMed] [Google Scholar]
- VanRullen, R. (2016). Perceptual cycles. Trends in Cognitive Sciences, 20(10), 723–735. 10.1016/j.tics.2016.07.006 [DOI] [PubMed] [Google Scholar]
- VanRullen, R. , Carlson, T. , & Cavanagh, P. (2007). The blinking spotlight of attention. Proceedings of the National Academy of Sciences, 104(49), 19204–19209. 10.1073/pnas.0707316104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- VanRullen, R. , & Koch, C. (2003). Is perception discrete or continuous? Trends in Cognitive Sciences, 7(5), 207–213. 10.1016/S1364-6613(03)00095-0 [DOI] [PubMed] [Google Scholar]
- Vigué‐Guix, I. , Fernández, L. M. , Cuello, M. T. , Ruzzoli, M. , & Soto‐Faraco, S. (2020). Can the occipital alpha‐phase speed up visual detection through a real‐time EEG‐based brain–computer interface (BCI)? European Journal of Neuroscience, 55(11‐12), 3224–3240. 10.1111/ejn.14931 [DOI] [PubMed] [Google Scholar]
- Vöröslakos, M. , Takeuchi, Y. , Brinyiczki, K. , Zombori, T. , Oliva, A. , Fernández‐Ruiz, A. , Kozák, G. , Kincses, Z. T. , Iványi, B. , Buzsáki, G. , & Berényi, A. (2018). Direct effects of transcranial electric stimulation on brain circuits in rats and humans. Nature Communications, 9(1), 483. 10.1038/s41467-018-02928-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walsh, E. G. (1952). Visual reaction time and the α‐rhythm, an investigation of a scanning hypothesis. The Journal of Physiology, 118(4), 500–508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walter, W. G. (1954). The electrical activity of the brain. Scientific American, 190(6), 54–63. [Google Scholar]
- Wang, X.‐J. (2010). Neurophysiological and computational principles of cortical rhythms in cognition. Physiological Reviews, 90(3), 1195–1268. 10.1152/physrev.00035.2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- White, P. A. (2018). Is conscious perception a series of discrete temporal frames? Consciousness and Cognition, 60, 98–126. 10.1016/j.concog.2018.02.012 [DOI] [PubMed] [Google Scholar]
- Whitmarsh, S. , Gitton, C. , Jousmäki, V. , Sackur, J. , & Tallon‐Baudry, C. (2021). Neuronal correlates of the subjective experience of attention. European Journal of Neuroscience, 55(11‐12), 3465–3482. 10.1111/ejn.15395 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiener, N. (2019). Cybernetics or control and communication in the animal and the machine, reissue of the 1961 (second ed.). MIT Press. [Google Scholar]
- Woods, A. J. , Antal, A. , Bikson, M. , Boggio, P. S. , Brunoni, A. R. , Celnik, P. , Cohen, L. G. , Fregni, F. , Herrmann, C. S. , & Kappenman, E. S. (2016). A technical guide to tDCS, and related non‐invasive brain stimulation tools. Clinical Neurophysiology, 127(2), 1031–1048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wöstmann, M. , Herrmann, B. , Maess, B. , & Obleser, J. (2016). Spatiotemporal dynamics of auditory attention synchronize with speech. Proceedings of the National Academy of Sciences, 113(14), 3873–3878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wynn, S. C. , Nyhus, E. , & Jensen, O. (2021). Alpha modulation in younger and older adults during distracted encoding. European Journal of Neuroscience, 55(11‐12), 3451–3464. 10.1111/ejn.15086 [DOI] [PubMed] [Google Scholar]
- Zaehle, T. , Rach, S. , & Herrmann, C. S. (2010). Transcranial alternating current stimulation enhances individual alpha activity in human EEG. PLoS ONE, 5(11), e13766. 10.1371/journal.pone.0013766 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zazio, A. , Ruhnau, P. , Weisz, N. , & Wutz, A. (2021). Pre‐stimulus alpha‐band power and phase fluctuations originate from different neural sources and exert distinct impact on stimulus‐evoked responses. European Journal of Neuroscience, 55(11‐12), 3178–3190. 10.1111/ejn.15138 [DOI] [PubMed] [Google Scholar]
- Zou, J. , Xu, C. , Luo, C. , Jin, P. , Gao, J. , Li, J. , Gao, J. , Ding, N. , & Luo, B. (2021). θ‐band cortical tracking of the speech envelope shows the linear phase property. Eneuro, 8(4), ENEURO.0058‐21.2021. 10.1523/eneuro.0058-21.2021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zrenner, C. , Belardinelli, P. , Müller‐Dahlhaus, F. , & Ziemann, U. (2016). Closed‐loop neuroscience and non‐invasive brain stimulation: A tale of two loops. Frontiers in Cellular Neuroscience, 10, 92. [DOI] [PMC free article] [PubMed] [Google Scholar]