

Abstract
Abstract—As the population ages, age-related cognitive impairments are becoming an increasingly pressing problem. Currently, the role of polyamines (putrescine, spermidine, and spermine) in the pathogenesis of cognitive impairments of various origin is actively discussed. It was shown that the content of polyamines in the brain tissue decreases with age. Exogenous administration of polyamines makes it possible to avoid cognitive impairment and/or influence the pathogenetic processes associated with disease progression. There are 3 known ways that polyamines can enter the human body: food, synthesis by intestinal bacteria, and biosynthesis in the body. Currently, one of the most promising approaches to the prevention of cognitive impairment is the use of foods with a high content of polyamines, as well as the use of various probiotics that affect intestinal bacteria that synthesize polyamines. Since 2018, in a number of European countries projects have been launched aimed at evaluation of the impact of a diet high in polyamines on cognitive processes. The review, based on analysis of modern scientific literature and the authors' own data, presents material on the effect of polyamines on cognitive processes and the role of polyamines in the regulation of neurotransmitter processes, and discusses the role of polyamines in cognitive disorders in mental and neurological diseases.
Keywords: cognitive impairment, polyamines, spermine, spermidine, putrescine
The purpose of this mini-review is to summarize information on some aspects of polyamine metabolism in neurological and psychiatric diseases and to evaluate the possibilities of using polyamines to correct cognitive impairment.
GENERAL CHARACTERISTICS OF POLYAMINES
Polyamines are a large group of organic polycations. Since the primary and secondary amino groups of polyamines are bound to H+, at physiological pH values, polyamines interact electrostatically with negatively charged molecules such as DNA, RNA, proteins, and phospholipids [1]. In living organisms, polyamines are present in both free (protonated) and bound forms. The interaction of polyamines with nucleic acids and acidic macromolecules is more stable than with inorganic cations (Mg2+ and Ca2+), which is determined by the presence of two or more positively charged groups in polyamine molecules [2].
Polyamines are involved in various important biochemical processes, such as the synthesis, functioning, and maintenance of the stability of nucleic acids (DNA and RNA) and proteins [2]. They play key roles in cellular signaling, DNA binding, transcription, RNA splicing, cytoskeleton functioning, and eukaryotic translation using translation initiation factor 5A (eIF5A) [3–7].
Endogenous polyamines such as putrescine, spermine, and spermidine are organic polycations that are synthesized from L-ornithine during decarboxylation (Fig. 1).
Fig. 1.

The structure of polyamines and some of their acetyl derivatives.
The large group of polyamines includes acetyl derivatives of spermidine and spermine, as well as cadaverine and agmatine, the formation of which is associated with the decarboxylation of lysine and arginine. This article discusses the exchange and role of spermidine and spermine polyamines, the formation of which occurs with the participation of the enzyme ornithine decarboxylase (ODC) (Fig. 2), and for which the ability to increase lifespan was shown [8–14].
Fig. 2.

Scheme of synthesis and breakdown of polyamines (after [14]). List of abbreviations: ODC, ornithine decarboxylase; SpdSy, spermidine synthase; SpmSy, spermine synthase; SSAT, spermidine/spermine N1-acetyltransferase; APAO, acetylpolyamine oxidase; SMO, spermine oxidase.
SPERMIDINE AND SPERMINE INCREASE THE LIFESPAN OF ANIMALS
For polyamines, especially spermidine, an increase in lifespan has been shown in a number of organisms and cell systems, including yeast (S. cerevisiae), Drosophila, nematodes, and human and mouse immune cells [9–14]. It was found that the mechanism of lifespan increase by spermidine is associated with direct inhibition of histone acetyltransferase (HAT) activity and autophagy induction [11]. The effects of spermidine on HAT activity lead to a global scenario for histones, which suppresses transcription due to histone hypoacetylation of many genes [13]. Autophagy activation by spermidine includes inhibition of Atg gene expression, regulation of elF5A and TFEB transcription, and inhibition of EP300, which directly promotes Atg gene acetylation and indirectly stimulates tubulin deacetylation [13]. Obviously, the study of the metabolism of polyamines is an urgent problem in connection with the effects described above.
POLYAMINE METABOLISM IN NORMAL MAMMALS
Synthesis. Putrescine is synthesized from ornithine (Fig. 2) [14]. The rate-limiting factor for biosynthesis of polyamines is the ODC enzyme [15]. Spermidine synthase (SpdSy) transfers the NH2 (CH2)3 fragment from S-adenosylmethionine to the amino group of putrescine, which leads to the formation of spermidine. Spermine is formed by the transfer of this fragment to spermidine with the participation of the enzyme spermine synthase (SPMSy). Spermine and spermidine are converted back to putrescine as a result of catabolism involving the enzymes polyamine oxidase, spermine oxidase (SMO), acetylpolyamine oxidase (APAO), and, in turn, SMO carries out the oxidative conversion of spermine to spermidine [16, 17].
Breakdown. The breakdown of polyamines may be associated with their conversion to lower molecular weight polyamines via two successive enzymatic reactions. First, spermine or spermidine is acetylated by the enzyme spermidine/spermine N1-acetyltransferase 1 (SSAT), which is a key regulatory enzyme that determines the degradation of polyamines [18]. The second is oxidative cleavage between C3 and N4 in acetylated spermine and spermidine to form lower molecular weight polyamines and 3-aminopropanal. It is important to note that both the synthesis of polyamines and their decomposition are carried out with the transfer of the triatomic (CH2)3 fragment.
Oxidation also leads to the degradation of polyamines.
Polyamine oxidation products are the lower molecular weight polyamines, Н2О2, 3-aminopropanal (3-AP), and 3-acetylaminopropanal [2, 18]. These aldehydes are unstable and spontaneously convert to acrolein after deamination. Acrolein, an unsaturated aldehyde, readily reacts with protein lysine residues to form protein-conjugated acrolein (PCAcrolein) [17]. Acrolein, a highly toxic unsaturated aldehyde, when interacting with proteins, lipids, and nucleic acids, has a systemic damaging effect, which leads to cell death. It has been shown that in experimental photo-induced stroke, the content of acrolein in the necrosis zone increases by 28 times, which is associated with the activation of the breakdown of polyamines. Acrolein is a much more toxic compound than H2O2 [17].
Regulation of polyamine metabolism. The activities of the enzymes, ODC and SSAT, for the biosynthesis and degradation of polyamines are regulated by changes in polyamine concentration, as well as by various compounds: growth factors, hormones, etc. [1, 2, 15–17]. At high concentrations of spermidine and spermine in cells, the activities of polyamine synthesis enzymes (ODC and SAMDC) are inhibited, while the activity of degradation enzymes (SSAT) is increased. Conversely, when cellular polyamine content is decreased, ODC and SAMDC are upregulated and SSAT is downregulated. The regulation of the synthesis and degradation of polyamines occurs at the level of transcription and translation of the biosynthesis of these enzymes [16]. With an increase in the content of polyamines inside the cell, ODC activity is inhibited by the induction of a protein, antizyme (AZI), which forms a complex with the ODC monomer, leading to inactivation of its enzymatic activity (Fig. 3) [19].
Fig. 3.

Scheme of regulation of polyamine biosynthesis (after [19]). Abbreviations: ODC, ornithine decarboxylase; AZI, antizyme, AZIN, antizyme inhibitor.
The degradation of this complex occurs in the 26S proteasomes. In addition, antizymes are able to inhibit the absorption of polyamines or stimulate their secretion. The existence of 3 forms of AZI antizymes has been shown: AZI 1, AZI 2, and AZI 3 [21]. A more complex regulation of ODC activity is provided by antizyme inhibitor (AZIN) proteins, which are able to interact with AZI with a higher affinity than with ODC. Currently, two forms have been identified: AZIN 1 and AZIN 2 [21].
Transport of polyamines into the cell is carried out by OCT transporters (organic cation transporter) [21].
Thus, normally, the homeostasis of polyamines in the body is maintained by complex feedback mechanisms at the level of biosynthesis, breakdown, intake, and removal from the cell. At the same time, an equally important issue in the regulation of polyamine metabolism is assessment of their entrance from the intestine. There are three ways that polyamines enter the human body: food, synthesis by intestinal bacteria, and biosynthesis in the body. Many works are devoted to the study of the content of polyamines in food products and the assessment of the effect of probiotics on the content of polyamines in the human body [22–24].
POLYAMINE HOMEOSTASIS DISTURBANCES
Disturbances in polyamine metabolism may result in severe neurological diseases, for example, mutations in spermine synthase and subsequent changes in the level of spermine and spermidine are a cause of Snyder–Robinson syndrome [25]. Similarly, mutation in the ODC1 gene leads to severe consequences such as macrosomia, macrocephaly, developmental delay, alopecia, spasticity, hypotension, cutaneous vascular malformations, impaired visual perception, and sensorineural hearing loss [26]. Studies by Jain et al., 2018 showed that the inhibition of polyamine breakdown is associated with spermine-mediated damage to the airway epithelium and provokes the development of asthma [27]. In model experiments on mice with SMOX inactivation and with combined inactivation of SMOX and SSAT1, an increase in the level of spermine and a decrease in the level of spermidine in the brain and cerebellum of Smox-KO and Smox/Sat1-dKO were found compared with intact animals [28]. Experiments with Smox/Sat1-dKO mice have shown that prolonged deficiency of polyamine catabolism causes TGM2 90 activation and increased expression of α-synuclein, polyamination and protein aggregation, followed by activation of a chain of events that lead to cerebellar injury and ataxia in Smox/Sat1- dKO mice [28].
FEATURES OF POLYAMINE METABOLISM IN NERVOUS TISSUE
Putrescine, spermine, and spermidine are present in all brain structures [29, 30]. The ODC enzymatic system is localized in the nuclei of neurons, in dendrites, axons, and nerve terminals. High activity of spermine and spermidine synthases was found in neurons [31–33].
In the brain, all the enzymes for the breakdown of polyamines are present, SSAT, diamino oxidase, and polyamine oxidase. MAO is also involved in the degradation process of polyamines [30]. It is important to note that the modification of the molecular structure of MAO caused by oxidative stress leads to a change in the substrate specificity and activation of MAO with respect to polyamines [30]. There are several other metabolic pathways for the conversion of polyamines within the CNS. Putrescine was shown to be involved in GABA biosynthesis and further conversion into a histidine-containing dipeptide, homocarnosine [34]. The content of polyamines in glia is significantly higher than in neurons [31, 33]. The fact that polyamines are synthesized in neurons but accumulates in glia allowed Skachkov et al. [31] to develop a hypothesis about the role of polyamines as gliotransmitters that regulate neuronal function. Endothelium and capillaries are involved in the transport of polyamines to brain cells [31].
The process of uptake/release of polyamines occurs with the participation of organic cation transporter 1 (OCT 1), through glial gap junctions and connexin-43 hemichannels (Cx43 HCs), and through polyamine-permeable receptors and channels [33].
Polyamines and gap junctions of the brain. In the CNS, in addition to participating in the implementation of basic regulatory functions, such as the regulation of protein, RNA, and DNA biosynthesis, etc., polyamines perform a number of specific functions: they modulate various aspects of the metabolism, synthesis, and breakdown of various neurotransmitters, as well as their interaction with receptors [30, 35]. Due to the ability of polyamines to interact with membranes, they can modulate the activities of membrane-bound enzymes, in particular Na,K-ATPase [36].
It should be noted there is a relationship between polyamines and intercellular gap junctions of the brain in normal conditions and in the development of neuropathologies. Gap junctions (GJ) are a system of densely packed channels penetrating the bilipid layers of the membranes of two contacting cells. Each channel consists of two docked half-channels called connexons, with the subunit of each connexon being the protein connexin. In a normal adult brain, GJs promote the formation of glial syncytia, three-dimensional glial scaffolds that perform a number of important functions, such as controlling the concentrations of extracellular ions and neurotransmitters, as well as the implementation of metabolic processes [37]. To maintain these functions in syncytium, a necessary condition is the opening of the GJ, which is associated with the presence of polyamines [38]. At the same time, in the adult brain, accumulation of polyamines (spermine, spermidine, putrescine, and agmatine) is a distinctive feature of astrocytes but not neurons [39–41, 31]. The unique ability of connexin 43 (Cx43) to maintain the GJ in the syncytium in the open state was shown by comparing Cx43 and Cx40 and demonstrating the phenomenon of polyamine blockade of the amino acid sequence (binding site) at the N-terminus of Cx40 [17–21]. Such specific binding sites for the spermine polyamine were not found in Cx43 [42]. Moreover, in further studies, it was noted that Cx43 is sensitive to polyamines, which open GJ channels, and, most importantly for cell physiology, polyamines eliminate the blockade of these channels by hydrogen [44] and calcium [45] cations. Elimination of the cationic block by polyamines is a critical factor for the functioning of the GJ in glia, since it is the acidification of the cytoplasm and the subsequent release of calcium (the so-called calcium waves in astrocytes) that is a distinctive property of astrocytic syncytium but not neuronal ensembles. By providing the propagation of potassium and calcium waves in the glial syncytium, GJs and hemichannels are directly able to control the activity of both individual neurons and neural networks. At the same time, depolarization of such syncytium with an increase in the concentration of potassium ions during epilepsy, neurotrauma, or ischemia [46] leads to the loss of the ability of astrocytes to eliminate excess potassium from the intercellular space, which leads to inactivation of neuronal activity and neuropathologies.
THE ROLE OF POLYAMINES IN THE FUNCTIONAL ACTIVITY OF THE BRAIN
Gating control. Polyamines are classically regarded as ion channel gating regulators. Spermine regulates ion transport by participating in the rectification of potassium channels. For the first time, A. Lopatin et al. revealed the mechanism of the so-called anomalous rectification, an ability of spermine to rectify currents of potassium ion channels of the KirX.X. family which is potential dependent [47–49].
It is now established that polyamines provide rectification of potassium channels, AMPA receptors and nACh receptors [35, 50]. The mechanism of channel rectification by polyamines is simple: membrane depolarization causes spontaneous attraction of cytosolic polyamines into the channel pore, thereby blocking the flow of positively charged ions into the cell through the receptor pores [50]. Thus, intracellular polyamines control the overexcitation of these receptors. This property of cytoplasmic polyamines is due to the fact that they have a low molecular weight and at physiological pH values they are positively charged.
Regulation of nACh receptor synthesis by polyamines. Spermine was shown not only to gate nACh receptors but also to modulate the activity of the ACh metabolism enzyme, acetylcholinesterase. Another level of regulation of acetylcholine metabolism by polyamines was recently discovered by Dhara et al. [51]. It was demonstrated that polyamines control the assembly of neuronal nicotinic α4β2 and α7 acetylcholine receptors [41]. This ability is unique because polyamines do not modulate the assembly of any other ion channels. It is noteworthy that a decrease in the level of polyamine increases the levels of α4β2 and α7Ach [51] in the brain. The authors showed a clear correlation between acetylcholine-induced action potentials and SSAT activity. α4β2- and α7Ach-receptors are the most common acetylcholine receptors.
Regulation of NMDA receptors by polyamines. Numerous studies have shown multiple effects of extracellular polyamines on glutamate receptors of the NMDA, AMPA, and kainate subtypes [35, 50, 52, 53]. Polyamines have a biphasic effect on glutamate receptors: they either block AMPA and NMDA receptor channels at high doses or activate kainate and NMDA receptors at low concentrations [22].
In a series of works, Williams et al. (1984–1989) showed that the NMDA receptor complex, like connexins, has a polyamine recognition site [54–58, 37]. NMDA receptors are the predominant molecular system for controlling synaptic plasticity and memory function at the cellular level, and polyamines seem to effectively modulate them. The effects of polyamines on NMDA receptors were first discovered by analyzing their binding to spermine and spermidine but not to diamine putrescine. The ability of spermine and spermidine to specifically interact with the polyamine-binding allosteric site of glutamate receptors was shown [35].
Four types of effects of spermine on NMDA receptors in the brain were identified, which differ depending on the membrane potential, pH, and the concentration of the agonist and co-agonist. The first type is stimulation of the flow of Ca ions in the presence of “replacing” concentrations of glycine, the so-called glycine-dependent stimulation [56, 59]. Spermine increases the affinity of NMDA receptors for glycine, regardless of whether these receptors contain GluN2A or GluN2B subunits. Moreover, this effect of spermine was not sensitive to changes in extracellular pH. The second type of NMDA receptor response to spermine is the stimulation of cation flux, which is observed in the presence of saturated concentrations of glutamate and glycine, the so-called glycine-independent stimulation [55, 60]. Moni et al. identified the binding site responsible for spermine-induced, glycine-independent stimulation of the GluN2B-containing subunit in NMDA receptors [61]. It was shown that polyamines activate NMDA receptors by shielding negative charges in the amino acid sequence in the proteins of the GluN1 and GluN2B subunits [61]. The third type of response is as follows: spermine can decrease the sensitivity of glutamate receptors to glutamate, which is associated with a decrease in affinity of the receptor. This effect is observed only in receptors containing GluN1 and GluN2B subunits, and probably involves the spermine binding site responsible for glycine-independent stimulation [61]. Finally, the fourth type of response; spermine can also bind to ion channel pores and induce voltage-dependent blocking of the channel at the site associated with the extracellular binding site for Mg2+ [62]. Moreover, the efficiency of spermine is two orders of magnitude higher than that of Mg2+ [31]. Selective inhibitors of the binding of polyamines to the NR2B NMDA receptor subunits—arcaine, ifenprodil, and traxoprodil—have been synthesized, and their administration to experimental animals causes impairment of the process of memorization and reproduction of a conditioned reflex [64]. After administration of spermine or spermidine into the hippocampus, a significant increase in learning ability is observed. Thus, a direct link was established between the polyamine binding site with the NR2B subunits of NMDA receptors and impaired memory processes [35, 50, 65].
In a model of focal cerebral ischemia in rats, it was found that in memory and learning disorders caused by ischemia, glycine facilitated the formation of but did not affect the retention of an acquired skill, while spermine contributed to the development of a conditioned reflex, which was reproduced throughout the entire observation period [30].
Obviously, polyamines are positive modulators of NMDA receptors and are able to compensate for the dysfunction of glutamate and cholinergic transmission, thereby eliminating memory and learning disorders.
Neurotoxicity of polyamines. It was previously shown that excessive levels of polyamines are neurotoxic [50] and that direct intracerebral infusion of spermine can provoke an impairment of the innate immune response. The addition of polyamines can have a toxic effect on cultured neurons. Upon incubation of a pure neuronal culture with 50 μM spermine, cell death is observed, while in the integral structure of brain sections consisting of neurons and glia (astrocytes), this concentration of spermine did not cause cell death [31].
The neurotoxicity of polyamines may be associated with the development of oxidative stress, which is caused by the induction, first, of H2O2 and, second, of unsaturated aldehydes such as 3-AP and acrolein [16, 17, 30]. The addition of spermine and spermidine to a cell culture containing blood serum inhibits the proliferation of any cell types due to the formation of the polyamine oxidation products, H2O2 and acrolein [16]. In neuronal cell culture, complete inhibition of cell growth is achieved upon incubation with 10 μM acrolein, 100 μM H2O2, and 20 μM OH [16]. Acrolein is a more toxic agent for brain tissue than ROS. The administration of aldehyde dehydrogenase, an enzyme that eliminates acrolein, into a neuronal culture containing toxic doses of spermine prevented cell death [23]. Using a model of ischemia caused by glucose-oxygen deprivation in neuroblastoma culture, M. Nakamura et al. found that it is Ca2+ that induces the toxicity of polyamines, while in the neuron there is a decrease in the content of polyamines and an increase in the formation of their decay products (PCAcrolein) [21]. One of the main pathways for Ca2+ entry into neurons is AMPA and NMDA receptor channels. Animal experiments have shown that the administration of N1, N4, N8 tribenzylspermidine, an NMDA receptor channel blocker, leads to a decrease in the content of Ca2+ and PCAcrolein in neurons, which is associated with a decrease in the size of the brain infarction focus [21]. It is established that glial polyamines are able to protect neurons from death by regulating these neuronal receptor channels [31].
Polyamines as neuroprotectors. The neuroprotective effect of polyamines was shown in animal models of neurodegenerative diseases. These diseases include Huntington’s disease caused by mutations—an increased number of CAG trinucleotide repeats that encode the amino acid glutamine. Expansion of CAG repeats leads to an increase in the length of the polyglutamine region in the huntingtin (Htt) protein. The clinical picture of Huntington’s disease is characterized by motor, mental, and cognitive disorders associated with glutamate receptors [68]. A dose-dependent neuroprotective effect of spermidine was found when modelling Huntington’s disease with intrastriatal administration of quinolic acid [69]. The authors found that a low dose of spermidine increased the activity of NMDA receptors in the striatum, while a high dose decreased the activity of NMDA receptors in this model [68].
Parkinson’s disease is an age-related neurodegenerative disease, the pathological features of which are motor and cognitive impairment associated with the loss of dopaminergic neurons in the substantia nigra, abnormal accumulation of iron in this structure, and the presence of intracellular aggregates of misfolded proteins, consisting mainly of alpha-synuclein fibrils and some other proteins [70]. The neuroprotective effect of spermidine was shown when modeling Parkinson’s disease by the administration of rotenone, which is associated with the antioxidant properties of polyamines [70].
Neurodegenerative diseases, despite the difference in trigger events, are characterized by common mechanisms leading to the death of brain cells. Excessive formation of ROS, deficiency of the endogenous antioxidant system, and excitotoxicity are viewed as such mechanisms. Polyamines, as “traps” for free radicals, can significantly reduce the level of ROS [71, 72] and excitotoxicity [35, 50].
DISTURBANCE OF POLYAMINE METABOLISM IN NEUROLOGICAL DISEASES
There is experimental evidence of the involvement of polyamines in the pathogenesis of many of the known neurological diseases in which memory and behavioral disorders are observed. Cognitive impairments, the occurrence of which is associated with impaired metabolism of polyamines, are characteristic of patients with Alzheimer’s disease, Parkinson’s disease, Huntington’s disease, an acute inflammatory process in the nervous system, acute and chronic cerebrovascular diseases and traumatic brain injury [31, 32, 35, 50, 66]. It is especially important to study the role of polyamines in the molecular mechanisms of the pathogenesis of Alzheimer’s disease. It has been shown that Alzheimer’s disease is characterized by the accumulation of tau protein, which forms neurofibrillary tangles, and neurotoxic amyloid beta peptide, which is responsible for the formation of senile plaques [34]. At the same time, patients with Alzheimer’s disease exhibit high ODC activity, an increase in its gene expression, and an increase in the content of spermidine and spermine in the brain, which the authors attribute to the role of polyamines in cognitive deficit and synaptic loss [50]. Modeling Alzheimer’s disease (by administration of amyloid beta-peptide) in animal experiments has shown that administration of the polyamine synthesis inhibitor DFMO (difluoromethylornithine) or blocking the site of spermine binding to NMDA receptors with arcaine restores impaired memory in mice.
Parkinson’s disease is another neurodegenerative disease in which disorders in polyamine metabolism associated with genetic mutations were found [73, 74]. Deficiency of ATP13A2 (PARK9) leads to pronounced changes in the export of polyamines [74]. In Parkinson’s disease, the expression of the SSAT enzyme changes, which leads to an increase in the content of acetyl derivatives of polyamines and disruption of their metabolism [73]. The mechanisms of polyamine involvement in cognitive decline in patients with Parkinson’s disease are associated with modulation of NMDA receptors, as well as with α-synuclein aggregation [75]. We have studied the content of spermidine in the erythrocytes of patients suffering from Parkinson’s disease, which correlated with MMSE. Thus, in the blood of patients with stages 1–2 of the disease (MMSE = 25–27 points), the content of spermidine increased significantly compared to donor blood, whereas in the blood of patients at the later stages (3–4) (MMSE = 12) with marked cognitive impairment, spermidine content was lower compared with the control [75].
POLYAMINES AND MENTAL ILLNESS
There is now evidence for the involvement of polyamines in the pathogenesis of many mental illnesses [76]. Changes in the expression and activity of polyamine degradation enzymes, and in the content of individual polyamines (putrescine, spermidine, and spermine) were shown in schizophrenia, depression, anxiety syndromes, etc. [77–79]. A special place in the neurochemistry of mental illness is occupied by works devoted to the study of the role of polyamines in the pathogenesis of depressive states with subsequent suicidal outcome [79]. In autopsy samples of 22 brain regions of persons who died as a result of suicide, significant differences were found in comparison with the control in the expression of genes responsible for the metabolism of polyamines: ODC, AZI 1, and AZI 2, spermine oxidase, spermine synthetase, and the aldehyde dehydrogenase family (enzymes that remove products of polyamine breakdown—toxic aldehydes) [79]. Moreover, as found in several independent studies, the decrease in the expression of the SSAT enzyme is especially pronounced in autopsy samples of the cerebral cortex in people who died by suicide [80]. A decrease in SSAT expression in the brain of patients who died by suicide leads to a significant increase in the content of spermidine and putrescine in brain structures compared to their content in the brain of mentally healthy individuals.
Thus, apparently, genetically determined disorders in the metabolism of polyamines is one of the pathogenetic mechanisms of mental disorders.
POLYAMINES AND AGE-RELATED CHANGES
With age, the content of polyamines in the brain decreases [81], which, according to some authors, plays a leading role in age-related changes in cognitive functions [81, 82]. It is established that the administration of spermine and/or spermidine has a significant positive effect on cognitive and behavioral responses in old animals [83]. It is important to note that administration of spermidine or spermine has a regulatory effect on autophagy proteins [84]. Spermidine and spermine maintain the energy metabolism of neurons [84–86]. In addition, the results of the analysis of such indices as Bcl-2, Bax, Caspase-3, NLRP3, IL-18, and IL-1β showed that spermidine and spermine prevent apoptosis and inflammation [86]. In 2013 Gupta V.K. et al. in experiments on aging flies found that spermidine dietary supplements attenuate age-related memory impairment through autophagy mechanisms [86]. In 2016, Eisenberg T. et al. reported that oral administration of spermidine to mice reduced myocardial hypertrophy and systemic blood pressure, enhanced cardiac autophagy, and improved the mechanoelastic properties of cardiomyocytes [86].
Given the relevance of the problem, we should dwell on the data on the possibility of using spermidine as an anti-aging compound in counteracting COVID-19 infection [85]. The available literature reflects the growing concern about the impact of coronavirus infection not only on the physical but also on the mental health of a person. The most important features of mental disorders seen with COVID-19 are symptoms of depression and anxiety, as well as cognitive impairment. Attention is drawn to the crosstalk between signs of aging and systems that are affected by coronavirus infection, as well as a higher mortality rate in older patients. One of the signs of age-related changes is the constant shortening of telomeres, which are protective structures at the ends of chromosomes. Shortening of telomeres contributes to the reduction of tissues to regeneration, loss of tissue homeostasis, and diseases. When determining the length of telomeres in peripheral blood lymphocytes of patients with COVID-19, it was found that shorter telomeres are associated with greater disease severity [85]. The property of spermidine to maintain telomeres, and lengthen them when added to the diet, can apparently determine both the antiaging effects of spermidine and its contribution to the antiviral effect [12]. Some authors believe that impaired autophagy, a characteristic feature of aging, may lead to the severity of COVID-19 [85]. Spermidine is an autophagy inducer. However, the mechanisms of interaction between autophagy and COVID-19 remain unclear.
In general, the study of the antiviral effects of anti-aging compounds, in particular polyamines, is of great importance for resolving issues of therapeutic strategy.
THE INFLUENCE OF INTRODUCING POLYAMINES TO THE DIET ON COGNITIVE PROCESSES IN ELDERLY PATIENTS. CLINICAL TRIALS
The basis for clinical trials was the effect of spermidine on autophagy. Spermidine has the ability to trigger an important process of beta-amyloid plaque dissolution by autophagy [50, 86]. A clear correlation was established between blood spermidine levels and cognitive performance in Alzheimer’s patients with dementia [87]. Dietary intake of spermidine correlates with cognitive performance in humans [88]. Studies have been conducted in mice that confirmed the safety and good tolerance of polyamines derived from wheat germ [86]. These data allowed the use of spermidine in clinical trials. It was found that polyamine levels of 13.5 µg/kg body weight per day for spermidine or 3.1 µg/kg body weight per day for spermine are the “no side effect” dose.
In 2018, Wirth M. et al. published the results of one of the first pilot clinical trials evaluating the ability of polyamines to improve cognitive function in elderly patients, in which it was shown that the intake of spermidine 1200 mg per day for 3 months improves cognitive function compared with the control group [88]. Pekar T. et al. reported in 2020 that a spermidine-enriched diet may prevent memory loss with aging. A clear correlation was established between spermidine concentration and improvement in cognitive performance in patients with mild to moderate dementia. Simultaneous dietary administration of 200 µg putrescine, 900 µg spermidine, and 500 µg spermine (total 1600 µg) per day for 12 months in elderly people had the most effective positive effect on cognitive processes [89, 90].
Currently, a number of studies discuss the results of examinations of patients with Alzheimer’s disease who received a diet with polyamines [91, 92].
INFLUENCE OF STRESS ON POLYAMINE METABOLISM IN THE BRAIN
Polyamines are actively involved in the molecular mechanisms of a nonspecific stress response, the so-called polyamine stress response (PSR) [93, 94]. Under a stressful effect on the body as a whole (burn, injury, etc.), ODC is activated in the brain, followed by a significant increase in the content of polyamines on the first day, which triggers the activation of the “early” response genes—c-fos, c-jus, c-myc [95, 96]. We found that in the brain of animals that underwent burn stress, the content of spermine and spermidine decreased on days 5–7 after burn stress compared with the control. The introduction of an adaptogen (DSIP) after stress had a modulatory effect on the metabolism of polyamines in the brain on days 5–7 after the burn: the content of spermidine increased on the 5th day, the intensity of incorporation of the radioactive label from [C14]-ornithine into polyamines increased 3 times for spermine (p ≤ 0.01) on the 7th day after stress compared with the control (Fig. 4).
Fig. 4.
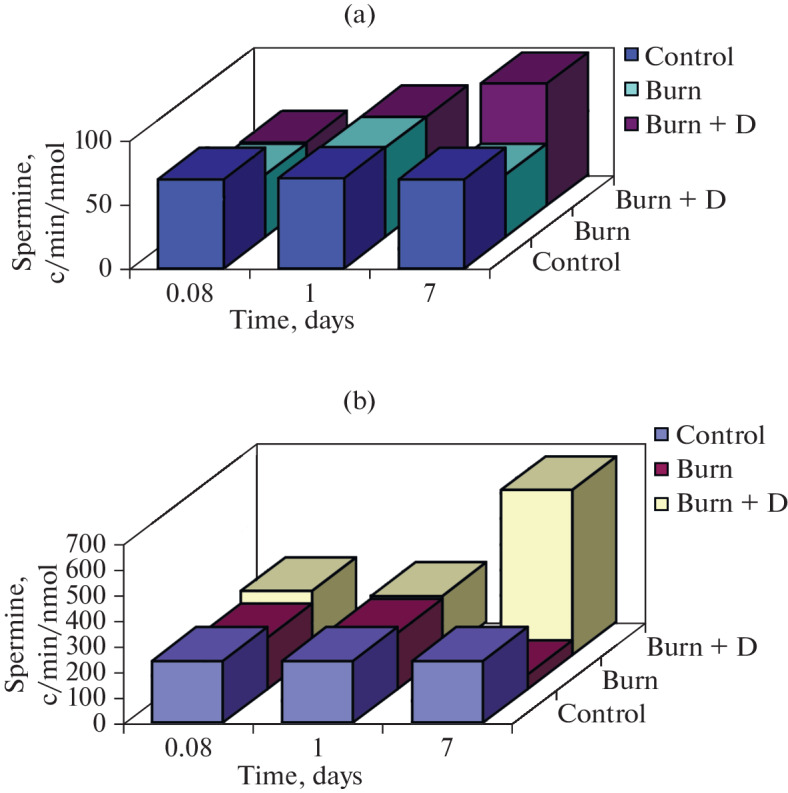
Effect of DSIP administration on the intensity of incorporation of the radioactive label from [C14]-ornithine into spermidine (a) and spermine (b) in the brain of rats after burns.
This stress caused memory and learning disorders. The administration of adaptogen (DSIP) and the subsequent increase in polyamine biosynthesis caused by it facilitated the formation of new skills and also had a positive effect on the retention of the acquired skill before stress [95].
POLYAMINE STRESS RESPONSE AND TAU PATHOLOGIES
Metabolic features of PSR were found in the nervous tissue, which depend on the nature of the stress effect [79]. With psycho-emotional stress, as well as ischemia/hypoxia of the brain, there is a more intense activation of enzymes for the breakdown of polyamines, which exceeds the rate of their synthesis. At the same time, there is a dramatic increase in ODC activity and the content of putrescine with a simultaneous decrease in the content of spermidine and spermine. In model experiments with animals, it was found that the response of the polyamine system to stress factors differs depending on the duration of stress and determines the character of polyamine metabolism in the brain. Data from Sandusky-Beltran et al. in 2019, indicate that an increase in the level of polyamines caused by short-term stress exposure is beneficial for the nervous system [97]. At the same time, spermidine and spermine are able to reduce the content of microtubule-associated tau protein by activating autophagy. However, long-term chronic stress exposure leads to maladaptive dysregulation of polyamine metabolism and has a negative impact.
Sandusky-Beltran et al. in 2021 reported a unique association between polyamine metabolism, behavioral disorders, and tau protein fate [98]. As is known, tau pathologies are characterized by the presence of various cognitive and affective behavioral disorders [99]. It was found that in Alzheimer’s disease there is a dysregulation of transcripts of genes associated with the metabolism of polyamines in the brain, while the level of the antizyme ODC inhibitor AZIN 2 considerably grows, which significantly increases the content of acetyl derivatives of polyamines, enhances the accumulation of tau protein, and contributes to cognitive and affective behavioral disorders [98]. Maintaining homeostasis in polyamine metabolism is a potential way to treat Alzheimer’s disease and tau pathologies [100].
Thus, PSR can cause both positive and negative effects in the CNS, depending on the duration and nature of the stressor. The administration of adaptogens had a modulatory effect on the content of polyamines, which positively influenced the processes of memorization and learning. It is important to note that PSR is currently viewed as a provocative factor in the triggering mechanisms of neurological and mental diseases.
CONCLUSIONS
Currently, disturbances in the metabolism of polyamines and in their intracellular distribution are considered pathogenetic mechanisms for the development of many diseases that are accompanied by cognitive disorders. Spermidine and spermine are able to significantly improve the process of learning and memorization and increase performance due to the modulation of neurotransmitter processes (nAch, the polyamine site of NMDA receptors) and the activation of autophagy in the CNS. The fact that the functional activity of these receptors is associated with the control of synaptic signaling and plasticity mechanisms, as well as with the formation of memory and memorization mechanisms, allows us to view polyamines as positive modulators of cognitive processes. In this regard, polyamines represent a promising direction for the development of therapy for cognitive impairment. The successful practical application of polyamine therapy is directly related to the development of our understanding of the functional significance of polyamines in the pathogenesis of cognitive impairment, as well as a thorough analysis of possible contraindications to its use.
FUNDING
This work was supported by the Russian Science Foundation, project no. 22-25-00795, https://rscf.ru/project/ 22-25-00795/.
COMPLIANCE WITH ETHICAL STANDARDS
Conflict of interests. The authors declare no conflict of interest.
Footnotes
Corresponding author; address: Gagarina square 1, Rostov-on-Don, 344003 Russia; e-mail: mgm52@bk.ru.
REFERENCES
- 1.Pegg A.E. J. Biol. Chem. 2016;291:14904–14912. doi: 10.1074/jbc.R116.731661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Park M.H., Igarashi K. Biomol. Ther. (Seoul) 2013;21:1–9. doi: 10.4062/biomolther.2012.097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ramani D., De Bandt J.P., Cynober L. Clin. Nutr. 2014;3:14–22. doi: 10.1016/j.clnu.2013.09.019. [DOI] [PubMed] [Google Scholar]
- 4.Landau G., Ran A., Bercovich Z., Feldmesser E., Horn-Saban S., Korkotian E., Jacob-Hirsh J., Rechavi G., Ron D., Kahana C. J. Biol. Chem. 2012;287:35825–35837. doi: 10.1074/jbc.M112.381335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Handa A.K., Fatima T., Mattoo A. Front. Chem. 2018;6:10. doi: 10.3389/fchem.2018.00010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Zwighaft Z., Aviram R., Shalev M., Rousso-Noori L., Kraut-Cohen J., Golik M., Brandis A., Reinke H., Aharoni A., Kahana C., Asher G. Cell Metab. 2015;22:874–885. doi: 10.1016/j.cmet.2015.09.011. [DOI] [PubMed] [Google Scholar]
- 7.Thomas E., Dever T.E., Ivanov I.P. J. Biol. Chem. 2018;293:18719–18729. doi: 10.1074/jbc.TM118.003338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Minois, N., Carmona-Gutierrez, D., and Madeo, F., Aging (Albany), 2011, vol. 3. [DOI] [PMC free article] [PubMed]
- 9.Eisenberg T., Abdellatif M., Schroeder S., Primessnig U., Stekovic S., Pendl T., Harger A., Schipke J., Zimmermann A., Schmidt A., Tong M., Ruckenstuhl C., Dammbrueck C., Gross A.S., Herbst V., Magnes C., Trausinger G., Narath S., Meinitzer A., Hu Z., Kirsch A., Eller K., Carmona-Gutierrez D., Buttner S., Pietrocola F., Knittelfelder O., Schrepfer E., Rockenfeller P., Simonini C., Rahn A., Horsch M., Moreth K., Beckers J., Fuchs H., Gailus-Durner V., Neff F., Janik D., Rathkolb B., Rozman J., de Angelis M.H., Moustafa T., Haemmerle G., Mayr M., Willeit P., von Frieling-Salewsky M., Pieske B., Scorrano L., Pieber T., Pechlaner R., Willeit J., Sigrist S.J., Linke W.A., Muhlfeld C., Sadoshima J., Dengjel J., Kiechl S., Kroemer G., Sedej S., Madeo F. Nat. Med. 2016;22:1428–1438. doi: 10.1038/nm.4222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Eisenberg T., Knaue H., Schauer A., Buttner S., Ruckenstuhl C., Carmona-Gutierrez D., Ring J., Schroeder S., Magnes C., Antonacci L., Fussi H., Deszcz L., Hartl R., Schraml E., Criollo A., Megalou E., Weiskof D., Laun P., Heeren G., Breitenbach M., Grubeck-Loebenstein B., Herker E., Fahrenkrog B., Frohlich K-U., Sinner F., Tavernarakis N., Minois N., Kroemer G., Madeo F. Nat. Cell. Biol. 2009;11:1305–1314. doi: 10.1038/ncb1975. [DOI] [PubMed] [Google Scholar]
- 11.Ghosh, I., Sankhe, R., Mudgal, J., Arora, D., and Nampoothiri, M., Neuropeptides, p. 83. [DOI] [PubMed]
- 12.Wirth A., Wolf B., Huang C.K., Glage S., Hofer S.J., Bankstahl M., Bar C., Thum T., Kahl K.G., Sigrist S.J., Madeo F., Bankstahl J.P., Ponimaskin E. GeroScience. 2021;43:673–690. doi: 10.1007/s11357-020-00310-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Nakanishi S., Cleveland J.L. Med. Sci. 2021;9:28. doi: 10.3390/medsci9020028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Berezov T.T., Korovkin V.F. Biokhimiya. Moscow: Meditsina; 1997. [Google Scholar]
- 15.Pegg A.E. J. Biol. Chem. 2006;281:14529–14532. doi: 10.1074/jbc.R500031200. [DOI] [PubMed] [Google Scholar]
- 16.Pegg A.E. Chem. Res. Toxicol. 2013;26:1782–1800. doi: 10.1021/tx400316s. [DOI] [PubMed] [Google Scholar]
- 17.Igarashi K., Kashiwagi K. Mol. Nutr. Food Res. 2011;55:1332–1341. doi: 10.1002/mnfr.201100068. [DOI] [PubMed] [Google Scholar]
- 18.Seller, N., Neurochemical Researc, vol. 25, no. 4, pp. 471–490. [DOI] [PubMed]
- 19.Silachev D.N. Tsitologiya. 2018;60:164–172. doi: 10.31116/tsitol.2018.03.02. [DOI] [Google Scholar]
- 20.Kahana, C., J. Biol. Chem., 2018, no. 48, pp. 18730–18735. [DOI] [PMC free article] [PubMed]
- 21.Soda K. Cells. 2022;11:164. doi: 10.3390/cells11010164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Sabater-Molina M., Zamora S. Nutrition. 2007;23:87–95. doi: 10.1016/j.nut.2006.09.006. [DOI] [PubMed] [Google Scholar]
- 23.Ramos-Molina B., Queipo-Ortur M.I., Lambertos A., Tinahones F.J., Penafiel R. Front. Nutr. 2019;6:1–15. doi: 10.3389/fnut.2019.00024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Rika HiranoR. Shirasawa, H., and Kurihara, S. Med. Sci. 2021;9:8. doi: 10.3390/medsci9010008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Murray-Stewart T., Dunworth M., Foley J.R., Schwartz C.E., Casero R.A. Med. Sci. 2018;6:12. doi: 10.3390/medsci6040112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Bupp C.P., Schultz C.R., Uhl K.L., Rajasekaran S., Bachmann A.S. AJMG. 2018;176:2548–2553. doi: 10.1002/ajmg.a.40523. [DOI] [PubMed] [Google Scholar]
- 27.Jain V. Med. Sci. 2018;6:4. [Google Scholar]
- 28.Marcoli M., Cervetto C., Amato S., Fiorucci C., Maura G., Mariottini P., Cervelli M. Biomolecules. 2022;12:204. doi: 10.3390/biom12020204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Berezov T.T., Makletsova M.G., Fedorova T.N. Ann. Klin. Eksperiment. Nevrol. 2012;6:38–43. [Google Scholar]
- 30.Berezov T.T., Makletsova M.G., Syatkin S.P., Rikhireva G.T., Kulikova O.I., Konovalova E.V., Fedorova T.N. Zhurnal Nevrologii i Psikhiatrii im. . S.S. Korsakova. 2013;113:65–70. [PubMed] [Google Scholar]
- 31.Skachkov S.N., Antonov S.M., Iton M. Dzh. Biol. Memb. 2016;33:3–31. doi: 10.7868/S0233475516010114. [DOI] [Google Scholar]
- 32.Nakanishi S., Cleveland J.L. Medical Sciences. 2021;9:28. doi: 10.3390/medsci9020028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kovacs Z., Skatchkov S.N., Veh R.W., Szabo Z., Nemeth K., Szabo P.T., Kardos J., Heja L. Front. Cell. Neurosci. 2022;15:787319. doi: 10.3389/fncel.2021.787319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Makletsova M.G., Uskova N.I., Bondarenko T.I. Biokhimiia. 1992;57:1348–1351. [PubMed] [Google Scholar]
- 35.Guerra G.P., Rubin M.A., Mello C.F. Pharmacol Res. 2016;112:99–118. doi: 10.1016/j.phrs.2016.03.023. [DOI] [PubMed] [Google Scholar]
- 36.Moinard C., Cynober L., Bandt J.-P. Clinical Nutrition. 2005;24:184–197. doi: 10.1016/j.clnu.2004.11.001. [DOI] [PubMed] [Google Scholar]
- 37.Kirichenko E.Y., Churyumova G.A., Logvinov A.K. Biochem. Mosc. Suppl. Ser. Membr. Cell Biol. 2016;10:207–217. [Google Scholar]
- 38.Benedikt J., Inyushin M., Kucheryavykh Y.V., Rivera Y., Kucheryavykh L.Y., Nichols C.G., Eaton M.J., Skatchkov S.N. Neuroreport. 2012;23:1021–1025. doi: 10.1097/WNR.0b013e32835aa04b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Laube G., Bernstein H.-G. Biochem. J. 2017;474:2619–2640. doi: 10.1042/BCJ20170007. [DOI] [PubMed] [Google Scholar]
- 40.Laube G., Veh R.W. Glia. 1997;19:171–179. doi: 10.1002/(SICI)1098-1136(199702)19:2<171::AID-GLIA8>3.0.CO;2-3. [DOI] [PubMed] [Google Scholar]
- 41.Hamon L., Savarin P., Pastre D. BioEssays. 2016;38:498–507. doi: 10.1002/bies.201500195. [DOI] [PubMed] [Google Scholar]
- 42.Musa H., Fenn E., Crye M., Gemel J., Beyer E.C., Veenstra R.D. J. Physiol. 2004;557:863–878. doi: 10.1113/jphysiol.2003.059386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Gemel J., Lin X., Veenstra R.D., Beyer E.C. J. Cell Sci. 2006;119:2258–2268. doi: 10.1242/jcs.02953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Skatchkov S.N., Bukauskas F.F., Benedikt J., Inyushin M., Kucheryavykh Y.V. Neuroreport. 2015;26:528–532. doi: 10.1097/WNR.0000000000000385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kucheryavykh L.Y., Benedikt J., Cubano L.A., Skatchkov S.N., Bukauskas F.F., Kucheryavykh Y.V. Neuroreport. 2017;28:208–213. doi: 10.1097/WNR.0000000000000746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Wang Q., Wang W., Aten S., Kiyoshi C.M., Du Y., Zhou M. Brain Sci. 2020;10:208. doi: 10.3390/brainsci10040208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Lopatin A.N., Makhina E.N., Nichols C.G. Nature. 1994;372:366–369. doi: 10.1038/372366a0. [DOI] [PubMed] [Google Scholar]
- 48.Lopatin, A.N. and Nichols, C.G., Inward rectifiers in the heart: an update on I, no. (K1). [DOI] [PubMed]
- 49.Nichols C.G., Lee S.J. J. Biol. Chem. 2018;293:18779–18788. doi: 10.1074/jbc.TM118.003344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Polis B., Karasik D., Samson A.O. Aging (Albany) 2021;13:10770–10795. doi: 10.18632/aging.202928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Dhara M., Matta J.A., Lei M., Knowland D., Yu H., Gu S. Nat. Commun. 2020;11:2799. doi: 10.1038/s41467-020-16629-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Girardi B.A., Fabbrin S., Wendel A.L., Mello C.F., Rubin M.A. Psychopharmacology (Berl) 2020;237:681–693. doi: 10.1007/s00213-019-05403-z. [DOI] [PubMed] [Google Scholar]
- 53.Mony L. J Physiology. 2022;600:233–259. doi: 10.1113/JP280875. [DOI] [PubMed] [Google Scholar]
- 54.Williams K., Romano C., Molinoff P.B. Mol. Pharmacol. 1989;36:575–581. [PubMed] [Google Scholar]
- 55.Williams K., Dawson V.L., Romano C., Dichter M.A., Molinoff P.B. Neuron. 1990;5:199–208. doi: 10.1016/0896-6273(90)90309-4. [DOI] [PubMed] [Google Scholar]
- 56.Williams K., Zappia A.M., Pritchett D.B., Shen Y.M., Molinoff P.B. Mol. Pharmacol. 1994;45:803–809. [PubMed] [Google Scholar]
- 57.Williams K. Neurosci. Let. 1995;184:181–184. doi: 10.1016/0304-3940(94)11201-S. [DOI] [PubMed] [Google Scholar]
- 58.Williams K. Biochem. J. 1997;325:289–97. doi: 10.1042/bj3250289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Bowie D., Mayer M.L. Neuron. 1995;15:453–62. doi: 10.1016/0896-6273(95)90049-7. [DOI] [PubMed] [Google Scholar]
- 60.Benveniste M., Mayer M.L. J. Physiol.-Lond. 1994;464:131–163. doi: 10.1113/jphysiol.1993.sp019627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Mony L. Zhu, S., Carvalho, S., and Paoletti, P. EMBO J. 2011;30:3134–3146. doi: 10.1038/emboj.2011.203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Rock D.M., Macdonald R.L. Mol. Pharmacol. 1992;41:83–88. [PubMed] [Google Scholar]
- 63.Yadav, M., Parle, M., Jindal, D.K., and Sharma, N., Biomed. Pharmacother., 2018, vol. 98. [DOI] [PubMed]
- 64.Sharma S., Kumar P., Deshmukh R. Neurochem. Int. 2018;116:104–11. doi: 10.1016/j.neuint.2018.02.010. [DOI] [PubMed] [Google Scholar]
- 65.Ferchmin P.A., Perez D., Biello M. Brain Res. 2000;859:273–79. doi: 10.1016/S0006-8993(00)01973-9. [DOI] [PubMed] [Google Scholar]
- 66.http://www.jbks.ru/archive/issue-17/article-2.
- 67.Nakamura, M., T. Uemura, R. Saiki et al., Atheroscleros, vol. 244, no. is. 2016, pp. 131–137.
- 68.Klyushnikov S.A. Nevrologicheskii zhurnal im. . L.O. Badalyana. 2020;1:139–158. doi: 10.17816/2686-8997-2020-1-3-139-158. [DOI] [Google Scholar]
- 69.Velloso N.A., Dalmolin G.D., Gomes G.M., Rubin M.A., Canas P.M., Cunha R.A., Mello C.F. Neurobiology of learning and memory. 2009;92:574–580. doi: 10.1016/j.nlm.2009.07.006. [DOI] [PubMed] [Google Scholar]
- 70.Buttner S., Broeskamp F., Sommer C., Markaki M., Habernig L., Alavian-Ghavanini A., Carmona-Gutierrez D., Eisenberg T., Michael E., Kroemer G., Tavernarakis N., Sigrist S.J., Madeo F. Cell Cycle. 2014;13:3903–3908. doi: 10.4161/15384101.2014.973309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Limon A., Mamdani F., Hjelm B.E., Vawter M.P., Sequeira A. Neurosci. Biobehav. Rev. 2016;66:80. doi: 10.1016/j.neubiorev.2016.04.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Cao W., Jia G., Zhao H., Chen X., Wu C., Tang J., Wang J., Liu G. Animal Nutrition. 2017;3:85–90. doi: 10.1016/j.aninu.2016.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Saiki S., Sasazawa Y., Fujimaki M., Kamagata K., Kaga N., Taka H., Li Y., Souma S., Hatano T., Imamichi Y., Furuya N., Mori A., Oji Y., Ueno S.I., Nojiri S., Miura Y., Ueno T., Funayama M., Aoki S., Hattori N. Ann. Neurol. 2019;86:251–263. doi: 10.1002/ana.25516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Vrijsen S., Besora-Casals L., van Veen S., Zielich J., Haute C., Hamouda N.N., Fischer C., Ghesquiere B., Tournev I., Agostinis P., Baekelandt V., Eggermont J., Lambie E., Martin S., Vangheluwe P. Proc. Natl. Acad. Sci. USA. 2020;117:31198–31207. doi: 10.1073/pnas.1922342117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Makletsova, M.G., Syatkin, S.P., Poleshchuk, V.V., Urazgildeeva, G.R., Chigaleychik, L.A., Sungrapova, C.Y., and Illarioshkin, S.N., J. Neurology Res., vol. 9, nos. 1–2, pp. 1–7.
- 76.Syatkin S.P., Svinarev V.I. Vestnik RUDN. 2010;1:36–41. [Google Scholar]
- 77.Baroli, G., Sanchez, J.R., Agostinelli, E., Mariottini, P., and Cervelli, M., IJMM, 2019, pp. 3–9. [DOI] [PubMed]
- 78.Chen G., Fiori L., Moquin L., Gratton A., Mamer O., Mechawar N., Turecki G. Neuropsychopharmacology. 2010;35:1477–1484. doi: 10.1038/npp.2010.17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Fiori L.M., Turecki G. J. Psychiatr. Res. 2011;45:1229–1235. doi: 10.1016/j.jpsychires.2011.03.015. [DOI] [PubMed] [Google Scholar]
- 80.Keilhoff G., Laube G., Dobrowolny H., Steiner J. World J. Psychiatr. 2021;11:1177–1190. doi: 10.5498/wjp.v11.i12.1177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Liu P., Jing Y., Zhang H. Neuroscience. 2008;155:789–796. doi: 10.1016/j.neuroscience.2008.06.033. [DOI] [PubMed] [Google Scholar]
- 82.Nishimura K., Shiina R., Kashiwagi K., Igarashi K. J. Biochem. 2006;139:81–90. doi: 10.1093/jb/mvj003. [DOI] [PubMed] [Google Scholar]
- 83.Schroeder S., Hofer S.J., Zimmermann A., Pechlaner R., Dammbrueck C., Pendl T., Marcello G.M., Pogatschnigg V., Bergmann M., Muller M., Gschiel V., Ristic S., Tadic J., Iwata K., Richter G., Farzi A., Ucal M., Schafer U., Poglitsch M., Royer P., Mekis R., Agreiter M., Tolle R.C., Sotonyi P., Willeit J., Mairhofer B., Niederkofler H., Pallhuber I., Rungger G., Tilg H., Defrancesco M., Marksteiner J., Sinner F., Magnes C., Pieber T.R., Holzer P., Kroemer G., Carmona-Gutierrez D., Scorrano L., Dengjel J., Madl T., Sedej S., Sigrist S.J., Racz B., Kiechl S., Eisenberg T. Cell Repots. 2021;35:108985. doi: 10.1016/j.celrep.2021.108985. [DOI] [PubMed] [Google Scholar]
- 84.Liang Y.T., Piao C., Beuschel C.B., Toppe D., Kollipara L., Bogdanow B., Maglione M., Lutzkendorf J., See J.C.K., Huang S., Conrad T.O.F., Kintscher U., Madeo F., Liu F., Sickmann A., Sigrist S.J. Cell Reports. 2021;35:108941. doi: 10.1016/j.celrep.2021.108941. [DOI] [PubMed] [Google Scholar]
- 85.Sharma P., and Sharma R., Saudi J. Biol. Sci. 2021;28:7046–7053. doi: 10.1016/j.sjbs.2021.07.087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Eisenberg T., Abdellatif M., Schroeder S., Primessnig U., Stekovic S., Pendl T., Harger A., Schipke J., Zimmermann A., Schmidt A., Tong M., Ruckenstuhl C., Dammbrueck C., Gross A.S., Herbst V., Magnes C., Trausinger G., Narath S., Meinitzer A., Hu Z., Kirsch A., Eller K., Carmona-Gutierrez D., Buttner S., Pietrocola F., Knittelfelder O., Schrepfer E., Rockenfeller P., Simonini C., Rahn A., Horsch M., Moreth K., Beckers J., Fuchs H., Gailus-Durner V., Neff F., Janik D., Rathkolb B., Rozman J., de Angelis M.H., Moustafa T., Haemmerle G., Mayr M., Willeit P., von Frieling-Salewsky M., Pieske B., Scorrano L., Pieber T., Pechlaner R., Willeit J., Sigrist S.J., Linke W.A., Muhlfeld C., Sadoshima J., Dengjel J., Kiechl S., Kroemer G., Sedej S., Madeo F. Nat. Med. 2016;22:1428–1438. doi: 10.1038/nm.4222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Wirth M., Benson G., Schwarz C., Kobe T., Grittner U., Schmitz D., Sigrist S.J., Bohlken J., Stekovic S., Madeo F., Floel A. Cortex. 2018;109:181–188. doi: 10.1016/j.cortex.2018.09.014. [DOI] [PubMed] [Google Scholar]
- 88.Wirth M., Schwarz C., Benson G., Horn N., Buchert R., Lange C., Kobe T., Hetzer S., Maglione M., Michael E., Marschenz S., Mai K., Kopp U., Schmitz D., Grittner U, Sigrist S.J., Stekovic S., Madeo F., Floel A. Alzheimers Res. Ther. 2019;11:36. doi: 10.1186/s13195-019-0484-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Pekar T., Wendzel A., Flak W., Kremer A., Pauschenwein-Frantsich S., Gschaider A., Wantke F., Jarisch R. Wien Klin. Wochenschr. 2020;132:42–46. doi: 10.1007/s00508-019-01588-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Pekar T., Bruckner K., Pauschenwein-Frantsich S., Gschaider A., Oppliger M., Willesberger J., Ungersback P., Wendzel A., Kremer A., Flak W., Wantke F., Jarisch R. Wien Klin Wochensch.r. 2021;133:484–491. doi: 10.1007/s00508-020-01758-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Hirano R., Shirasawa H., Kurihara S. Med. Sci. 2021;9:8. doi: 10.3390/medsci9010008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Soda K., Uemura T., Sanayama H., Igarashi K., Fukui T. Med. Sci. 2021;9:22. doi: 10.3390/medsci9020022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Gilad G.M., Gilad V. Cell Mol. Neurobiol. 2003;23:637–649. doi: 10.1023/A:1025036532672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Gilad G.M., Gilad V.H. Brain Res. 2002;5:23–29. doi: 10.1016/S0006-8993(02)02479-4. [DOI] [PubMed] [Google Scholar]
- 95.Makletsova M.G., Prokof’ev V.N., Rikhireva G.T., Mikhaleva I.I., Markvo L.I., Kuraev E.G. Novye Lekarstvennye Preparaty. 2007;3:33–43. [Google Scholar]
- 96.Randi A.S., Hernandez S., Sanchez M., Schwarcz M., Kleiman D., Pisarev D.L. Toxicol. Sci. 2003;76:291–298. doi: 10.1093/toxsci/kfg253. [DOI] [PubMed] [Google Scholar]
- 97.Sandusky-Beltran L.A., Kovalenko A., Ma C., Calahatian J. I.T., Placides D.S., Watler M.D., Hunt J.B., Darling A.L., Baker J.D., Blair L.J., Martin M.D., Fontaine S.N., Dickey C.A., Lussier A.L., Weeber E.J., Selenica M.B., Nash K.R., Gordon M.N., Morgan D., Lee D.C. Alzheimers Res. Ther. 2019;11:58. doi: 10.1186/s13195-019-0507-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Sandusky-Beltran, L.A., Kovalenko, A., Placides, D.S., Ratnasamy, K., Ma, C., Hunt, J.B.Jr., Liang, H., Calahatian, J.I.T., Michalski, C., Fahnestock, M., Blair, L.J., Darling, A.L., Baker, J.D., Fontaine, S.N., Dickey, C.A., Gamsby, J.J., Nash, K.R., Abner, E., Selenica, M.B., and Lee, D.C., J. Clin. Invest., 2021, vol. 131, no. 4. [DOI] [PMC free article] [PubMed]
- 99.Prokop J.W., Bupp C.P., Frisch A., Bilinovich S.M., Campbell D.B., Vogt D., Schultz C.R., Uhl K.L., Vansickle E., Rajasekara S., Bachmann A.S. Genes. 2021;12:470. doi: 10.3390/genes12040470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Signor C., Mello C.F., Porto G.P., Ribeiro D.A., Rubin M.A. Eur. J. Pharmacol. 2014;730:72–76. doi: 10.1016/j.ejphar.2014.02.035. [DOI] [PubMed] [Google Scholar]