

Abstract
Dynamin superfamily proteins (DSPs) mediate membrane fission and fusion necessary for endocytosis, organelle biogenesis and maintenance, as well as for bacterial cytokinesis. They also function in the innate immune response to pathogens and in organizing the cytoskeleton. In this review, we summarize the current understanding of the molecular mechanism of DSPs, with emphasis on the structural basis of function. Studies from the past decade on the structure and mechanism of DSPs enable comparative analysis of shared mechanisms and unique features of this protein family.
The Dynamin Superfamily
DSPs are present in all organisms and comprise proteins with diverse roles in membrane remodeling, actin bundling, and innate immunity (Figure 1). Dynamin is the founding member of this family, and insights into its function came from observations that Drosophila melanogaster with a temperature-sensitive mutation in the dynamin homolog Shibire had deficient endocytosis [1–4]. Work over the past three decades has determined that the principal function of dynamin is membrane fission leading to the budding of vesicles [5]. Additional DSPs have been identified based on sequence homology and shared structural features. Akin to dynamin, Drp1 (Dnm1p in yeast and ARC5 in plants) mediates membrane fission in mitochondria, peroxisomes, and chloroplasts [6–11]. Intriguingly, DSPs also include proteins involved in membrane fusion. For example, the mitofusins (Fzo1p in yeast) [12–14] and OPA1 (Mgm1p and Msp1p in Saccharomyces cerevisiae and Schizosaccharomyces pombe, respectively) mediate mitochondrial fusion [15,16], while atlastin (Sey1p in yeast and RHD3 in plants) functions in fusion of endoplasmic reticulum (ER) membranes [17–19]. DSPs also include the antimicrobial guanylate-binding proteins (GBPs), MxA and MxB [20–23], yeast Vps1p involved in both fusion and fission [24,25], and bacterial dynamin homologs involved in bacterial cell division and vesicle secretion [26–28]. In this review, we provide background to the molecular mechanisms of DSPs in membrane remodeling, innate immune responses, and cytokinesis, which is timely because it puts the recent structural studies of DSPs into context [29–35].
Figure 1. Overview of Cellular Processes Mediated by Dynamin Superfamily Proteins (DSPs).

Membrane fission: dynamin functions in clathrin-mediated endocytosis and budding of vesicles from the Golgi. Drp1 mediates mitochondrial fission. Membrane fusion: fusion of endoplasmic reticulum (ER) membranes is mediated by atlastin (Sey1p in yeast and RHD3 in plants) and fusion of mitochondrial outer and inner membranes is mediated by mitofusins (Fzo1p in yeast) and OPA1 (Mgm1p and Msp1p in Saccharomyces cerevisiae and Schizosaccharomyces pombe) respectively. Other roles: dynamin bundles actin; MxA, MxB, and guanylate-binding proteins (GBPs) restrict pathogens; Vps1p is specific to yeast and mediates both vacuolar fission and fusion; and bacterial DSPs (bacterial-like dynamin protein, BDLP, and DynA) localize at septa to mediate bacterial cytokinesis.
What Makes a Dynamin Superfamily Protein?
DSPs all have two common domains: a GTPase domain and an elongated α-helical bundle domain that, in most cases, drives self-assembly (Figure 2). The antimicrobial GBP1 protein comprises only these two domains, while other DSPs have additional domains that mediate specialized roles [36]. For example, dynamin has a pleckstrin homology domain (PHD) involved in lipid binding and a proline-rich domain (PRD) that enables binding to SH3 domain-containing proteins [5] (Figure 2). The function and mechanism of domains conserved in dynamin superfamily proteins are described later.
Figure 2. Models of Selected Dynamin Superfamily Proteins (DSPs) Showing Similarity in Structure and Domain Organization.
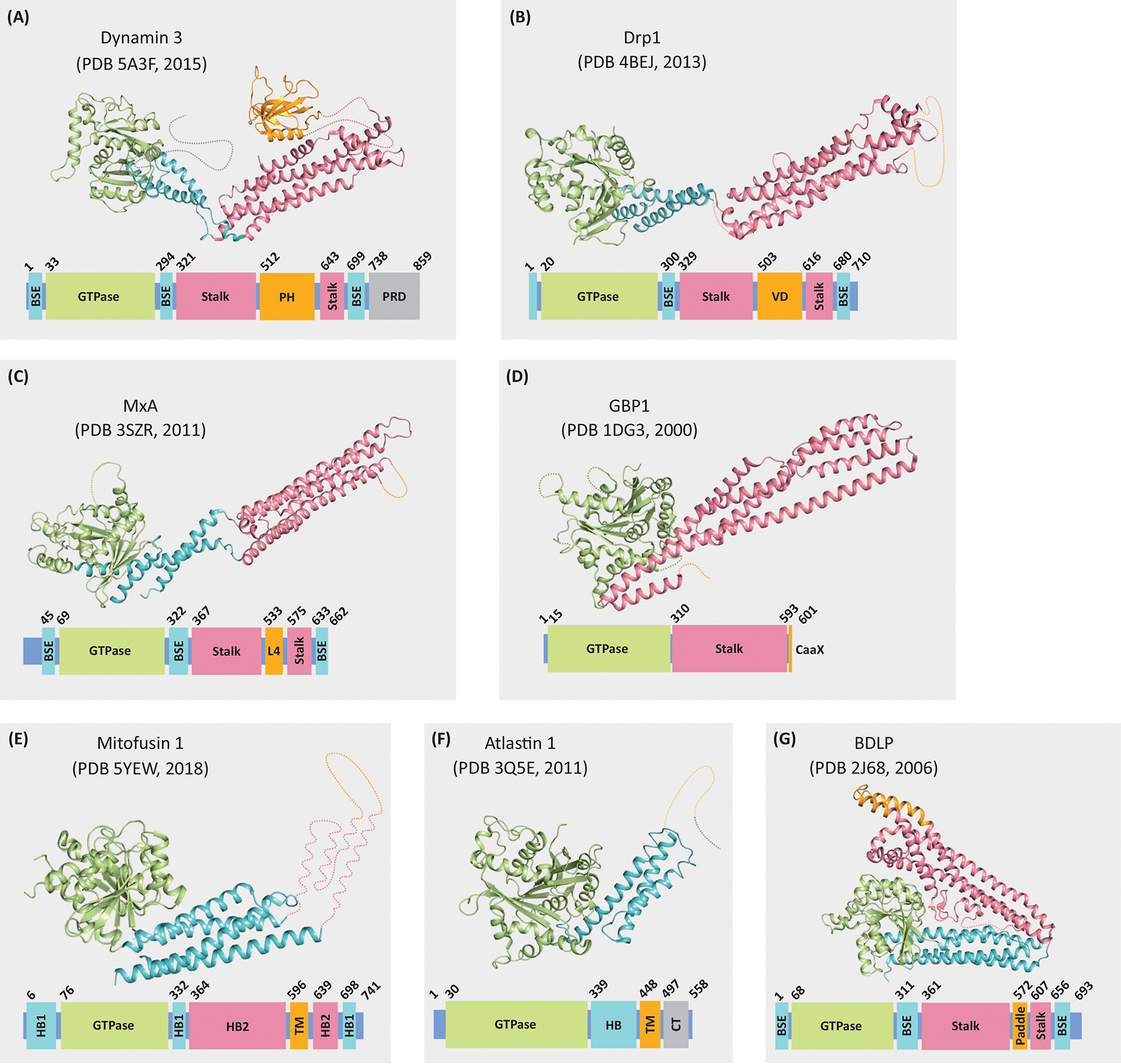
Regions not included in the crystal structures are indicated by dotted lines. Structures are color coded to show the GTPase domain (green), bundle signaling element (BSE; blue), helical bundle domain (HB; blue), stalk (pink), helical bundle domain 2 (HB2; pink), proline-rich domain (PRD; grey) and C-terminal tail domain (CT; gray), and lipid-interacting domain (orange). (A) dynamin 3; (B) Drp1; (C) MxA; (D) guanylate-binding protein 1 (GBP1); (E) mitofusin 1; (F) atlastin 1; and (G) bacterial dynamin-like protein (BDLP) from Nostoc punctiforme.
The GTPase Domain
The most highly conserved region of DSPs is the GTPase domain. Nucleotide binding and hydrolysis is mediated by four consensus elements, G1–G4 (Figure 3A,C). The G1 motif, or P-loop, (GxxxxGKS/T) binds the phosphates. The G2 and switch 1 region includes a threonine that enables GTP hydrolysis by coordinating a Mg2+ ion. The G3 and switch 2 region (DxxG) also coordinates the Mg2+ ion through aspartate, while glycine binds the γ-phosphate. The G4 motif (N/TK/RxD) binds the nucleotide base [37,38]. For dynamin, mutations in the GTPase domain (K44A, T65A, T141D, and K142A) lead to inhibition of endocytosis [39–41].
Figure 3. Biochemical and Structural Features of the GTPase Domain from Dynamin Superfamily Proteins (DSPs).

(A) Nucleotide-binding and hydrolysis motifs are conserved in all DSPs and small GTPases (e.g., HRas). Protein accession codes are as follows: dynamin 1 (GenBank: AAH50279.2), Drp1 (NCBI reference sequence: NP_001317309.1), OPA1 (GenBank: AKI72360.1), mitofusin 1 (GenBank: EAW78406.1), atlastin 1 (GenBank: AAH10708.2), GBP1 (NCBI reference sequence: NP_002044.2), MxA (NCBI reference sequence: NP_001171517.1), Vps1p (UniProtKB: P21576.2), Dnm1p (GenBank: CAA97444.1), Mgm1p (GenBank: ONH78423.1), Fzo1p (GenBank: EWH19560.1), Sey1p (GenBank: AJU15101.1), BDLP (UniProKB: B2IZD3.1), DynA (GenBank: CAB14120.1), and HRas (GenBank: AAH99130.1). Biochemical properties of dynamin GTPases depicting the guanine nucleotide kcat and binding affinities (right columns) for dynamin GTPases compared with the small GTPase, HRas. (Dynamin 1 [39,159–162], Drp1 [32,116,118,163,164], OPA1 [165], Mitofusin 1 [34,35], Atlastin 1 [46,133], GBP1 [36,166], MxA [167], Vps1p [168], Dnm1p [169,170], Mgm1p [171], Fzo1p, Sey1p [127], BDLP [26], DynA, and HRas [172,173]). (B) Nucleotide-mediated dimerization of the GTPase domain of dynamin with a bound GDP/AlF4− (orange) [Protein Data Bank (PDB): 2X2E] [43]. (C) Structure of the nucleotide-binding pocket illustrating conserved motifs. GTPase domain dimerization is stabilized via trans stabilizing residues (e.g., D180) that promote nucleotide binding and hydrolysis [43]. Motifs color-coded as in (A).
The GTPase domain of DSPs belongs to the G proteins activated by nucleotide-dependent dimerization (GAD) family of GTPases [42]. These GTPases do not require guanine exchange factors (GEFs) or GTPase-activating proteins (GAPs), and their activity is enhanced by nucleotide-induced dimerization [42,43]. Dynamin GTPases do not require GEFs due to their low micromolar affinity for nucleotides, unlike Ras GTPases, which have picomolar affinity (Figure 3A). Dynamin GTPases also have a higher basal rate of GTP hydrolysis compared with small GTPases and do not require a GAP [44]. GAPs stabilize the emerging negative charge at the transition state of GTP hydrolysis by providing an arginine finger in trans [45]. By contrast, most DSPs utilize a cation in the active site to serve this function (Figure 3C). The family members GBP1 and atlastin differ slightly in that they accomplish this using an arginine finger in cis within the P loop [43,45–48]. Mitofusin and bacterial dynamin-like protein (BDLP) are unique in having a conserved tryptophan within the G4 motif, which occupies the nucleotide-binding pocket in the apo-state but moves into a hydrophobic groove upon GTP binding [35]. This tryptophan switch is essential for mitofusins because mutating the residue to alanine abolished nucleotide binding and hydrolysis [35]. Binding and hydrolysis of GTP results in conformational changes in the GTPase domain, for example within the P loop and switch 2 regions for mitofusin 1 [34], which result in a swing in the bundle signaling element (BSE), or helical bundle domain, that may be transmitted as mechanical work to power DSPs [26,43,48,49].
The Elongated α-Helical Bundle Domains
Several DSPs have two elongated α-helical bundle domains, referred to as the BSE, and the stalk which are structurally contiguous (Figure 2). The BSE is adjacent to the GTPase domain and comprises noncontiguous sequences from the N- and C-terminal helix of the GTPase domain, as well as the first helix immediately following the stalk domain. A large swing in the BSE during the GTPase cycle is proposed to cause the powerstroke of DSPs that leads to fission and fusion [43,48,50]. Early hints into the role of the BSE in regulating dynamin came from the identification of suppressor of Shibire (Sushi) mutants in this region that rescue temperature-sensitive Shibire mutants, which occur within the GTPase domain and cause defects in GTP binding and endocytosis [51,52]. The stalk links the GTPase-BSE domain to the lipid-interacting domains and also provides interfaces for higher-order assembly.
The mechanochemical function of DSPs, in general, is dependent on the oligomerization of the protein. Whereas the GTPase domain serves as the mechanical switch, the helical bundles are functional scaffolds for dynamin polymerization. In some dynamin superfamily proteins, the nomenclature used for these α-helical bundle domains is different. For mitofusin and atlastin, the helical bundle adjacent to the GTPase domain is referred to as helical bundle 1 (HB1) domain and helical bundle (HB) domain, respectively (Figure 2B,F), and the second α-helical bundle in mitofusins is referred to as HB2 (Figure 2).
Lipid Interaction
DSPs that remodel membranes, and even those involved in innate immunity, interact with membrane lipids. The most extensively studied lipid binding domain is the ~100-amino acid PHD of dynamin [53–56]. Other membrane-remodeling DSPs lack a PHD but retain interactions with lipids. Mitofusin, OPA1, and atlastin have transmembrane domains (TM), while Drp1 utilizes its variable domain (VD) [57]. MxA proteins use the L4 loop in lipid binding [58], BDLPs have the paddle domain [59], and, in GBPs, the CaaX motif is isoprenylated to mediate lipid interaction [60].
Dynamin Superfamily Proteins Involved in Membrane Fission
Dynamin
The most extensively characterized role of dynamin is the scission of budding endocytic vesicles from the plasma membranes during endocytosis (Figure 4Ai) [1,2]. Dynamin also regulates the organization of microtubules and actin to facilitate endocytosis, cell motility, and cytokinesis [4,61–65], and there are emerging roles for dynamin in controlling fusion pore expansion and closure during events such as exocytosis [66–69]. The three mammalian isoforms of dynamin share ~80% sequence identity, with differences concentrated in the PRD. Dynamin isoforms have different tissue expression profiles, with dynamin 1 found predominantly in neurons, dynamin 2 ubiquitously expressed, and dynamin 3 found in the brain, lungs, and testis [70]. The tissue expression profile of dynamin 2 suggests that it mediates house-keeping clathrin-mediated endocytosis (CME), while dynamin 1 and 3 have specialized roles. This is supported by observations that deleting dynamin 2 in mice results in embryonic lethality [65], but there is no clear phenotype in dynamin 3 knockout mice [71]. Deleting dynamin 1 shortens the lifespan of mice, and the phenotype can be exacerbated to perinatal lethality when both dynamin 1 and 3 are deleted [71,72]. Even though dynamin 2 is critical for house-keeping CME in all cells, dynamin 1 also contributes, but at earlier stages [73]. In conclusion, the essential role of dynamin is supported by the recent identification of mutations in dynamin 2 associated with neurological diseases, including Charcot-Marie-Tooth (CMT), centronuclear myopathy, and hereditary spastic paraplegia [74–78].
Figure 4. Mechanism of Dynamin and Drp1-Mediated Membrane Fission and their Associated Assembled Structures.

(A) Model for dynamin-mediated membrane fission during endocytosis (i). Dynamin assembles at the neck of budding endocytic vesicles and constricts to a superconstricted state that leads to scission and release of dynamin as a tetramer in solution. (ii) Crystal structure of the soluble dynamin tetramer. Interfaces 1–3 are highlighted, and one monomer is color-coded similarly to Figure 2 in the main text. (iii) Cryo-electron microscopy (EM) structure of the dynamin tetramer in the lipid-bound state. Arrows indicate the conformational change that occurs upon assembly into a polymer. (iv) Dynamin helical polymers bound to lipid in the presence of GMPPCP (constricted state). Domains are colored as in Figure 2 in the main text. Inner lumens and outer diameters are indicated below each polymer. (v) Dynamin helical polymers bound to lipid in the presence of GTP (superconstricted state). (B) Model of Drp1-mediated mitochondrial fission (i). Drp1 assembles around the mitochondria and constricts the membrane, which leads to fission of mitochondria and the release of Drp1 into the cytoplasm. (ii) Crystal structure of the soluble Drp1 dimer. Interface 2 is highlighted and one monomer is color coded similar to Figure 2 in the main text. (iii) Cryo-EM structure of Drp1 in complex with MiD49 (teal) with the four Drp1/MiD49 interfaces highlighted (circles). (iv) Drp1 helical polymers bound to cardiolipin-containing lipid tubes (narrow lumen). (v) Dnm1p helical polymers bound to lipid tubes (wide lumen). Abbreviation: PE:PIP, phosphatidylethanolamine:phosphatidylinositol 4-phosphate.
Function and Mechanism of Dynamin in Endocytosis
The early observations that dynamin is located at sites of budding endocytic vesicles and that inhibiting dynamin results in the accumulation of endocytic pits suggested a role in membrane fission [1,41]. Furthermore, in vitro studies demonstrated that dynamin self-assembles into helical polymers around tubulated liposomes [79,80] and causes membrane fission when incubated with GTP [80–82]. Dynamin-mediated membrane fission begins with the recruitment of dynamin to sites of CME and subsequent binding to membranes through interactions with negatively charged lipids. The SH3 proteins endophilin, intersectin, amphiphysin, and syndapin have key roles in recruiting and regulating the activity of dynamin during CME [65,70,83–89]. In addition, endophilin and amphiphysin generate membrane curvature at the necks of budding vesicles, which enables dynamin assembly [90–93].
Upon recruitment of dynamin to sites of CME, it interacts with membrane lipids directly through its PHD. The PHD comprises one helix and two antiparallel β-sheets that are intercalated by three variable loops (Figure 2A) [53–55]. The variable loops are positively charged and contain binding sites for anionic lipids, particularly PI(4,5)P2 [94]. The critical lipid-binding and membrane insertion residues in variable loop 1 in dynamin 1 are K535 and I533, respectively [95–98]. By contrast, variable loop 3 has a role in generating membrane curvature via a tyrosine, Y600 in dynamin 1. This residue is replaced by a leucine (L596) in dynamin 2, causing dynamin 2 to be less efficient in generating curvature [99]. Following membrane binding, the PHD transitions from a closed state in solution, where it is adjacent to the stalk and inhibits oligomerization, to an open conformation that allows dynamin to assemble into large helical polymers necessary for membrane fission [100]. The controlled assembly of dynamin on the membrane may be disrupted in many patients with CMT, where the mutations are in the stalk–PHD interface, which may allow dynamin to assemble in the cytoplasm [75]. In addition to promoting membrane fission by binding the membrane and regulating dynamin assembly, the PHD fluctuates in and out of membranes during GTP hydrolysis, potentially enabling membrane fission [97,101].
Dynamin helices wrap around the neck of budding endocytic vesicles and constrict upon GTP binding and hydrolysis to cause membrane fission (Figure 4A). Structures of dynamin show that dimerization [5,29,49,52,102] depends on interactions between stalks (interface 2), and that tetramerization results when two dimers interact at the ends of the stalk (interfaces 1 and 3) (Figure 4Aii) [52,103]. Mutants, such as K361 in dynamin 3 and G397D in dynamin 1, which disrupt assembly and block endocytosis, have been useful tools for structural studies using X-ray crystallography [52,103,104]. When dynamin assembles on membranes, the GTPase and PH domains extend away from the stalk (Figure 4Aiii, arrows) and a novel bend in one BSE of the dimer is observed [29]. These conformational changes allow the GTPase domains to dimerize between the rungs of the helix, which stabilizes the nucleotide active site and leads to stimulated GTPase hydrolysis (~100-fold increase) [29,43,49] (Figure 3B,C). The asymmetry caused by a novel bend in one BSE is necessary to maintain the GTPase domain dimer while constricting the membrane. Cells expressing a dynamin helix-stabilizing mutant (TLP 292–294, AAA), predicted to be missing this novel bend, were impaired in endocytosis by ~50% [29]. Upon the powerstroke of dynamin, the BSE swings down and transiently stabilizes a hemifission state as well as disrupting the polymer, such that it becomes disordered and the GTPase dimer interface across the rungs of the helix is disrupted [105]. During the GTP cycle, dynamin constricts the underlying membrane from a 7-nm lumen in the GTP-bound state to 3.4 nm in the post-GTP-hydrolysis superconstricted state (Figure 4Aiv,v), which eventually proceeds to a hemifission state [29,49,102,105]. Curiously, the transition to the superconstricted state is achieved by a helical rearrangement from a 1-start to a 2-start helix. The switch from a 1-start to a 2-start helix explains how superconstriction is achieved without major conformational changes in the dynamin tetramer. Following constriction, the final fission event may require additional conformational changes of the dynamin polymer, or it may occur simply by the disassembly of dynamin.
Dynamin Interaction with the Cytoskeleton
Although dynamin was first identified as a microtubule-binding protein, cellular roles for this interaction were identified only recently [4]. Dynamin colocalizes with microtubules during the cell cycle and enhances their dynamic instability [63,106]. In fact, mutations in dynamin 2, which result in CMT, cause defects in dynamin–microtubule interactions [63,77]. Dynamin also interacts with actin filaments at the tail of actin comets, in podosomes and filopodia, and in protrusions that enable myoblast fusion [5,64]. The exact role of dynamin in these actin assemblies is currently being explored, and it may have a similar role as seen with microtubules [106]. A mutation in dynamin 2 linked with centronuclear myopathy causes impaired actin organization, further demonstrating the role of dynamin in regulating actin organization [74].
Drp1
Drp1 (Dnm1p in yeast) is a dynamin-related protein that mediates fission of mitochondria to maintain organelle integrity (Figure 4Bi) [6,7,107–109]. Drp1 exists as a dimer–tetramer equilibrium in solution [31,110–112]. Similar to dynamin, Drp1 forms helices around lipid tubes, but with a larger range of outer diameters (~50–120 nm) that constrict upon GTP hydrolysis (Figure 4Biv,v) [110]. The Drp1 helix is proposed to form a more flexible oligomer than dynamin, which may accommodate fission of the mitochondrial double membrane [32]. After fission, membrane-bound Drp1 (Figure 4Biv,v) potentially transitions to a closed conformation, which has the GTPase domain packed next to the stalk, potentially blocking assembly on membranes [31].
Drp1 is a cytosolic protein that is recruited to the outer mitochondrial membrane through the integral membrane proteins, Fis1, Mff, MiD49, and MiD51 [113]. A recent cryo-electron microscopy (EM) structure identified four interfaces mediating the interaction between MiD49 and Drp1, which begins to reveal the structural basis of Drp1 recruitment to mitochondria (Figure 4B) [30]. Similar to dynamin, the GTPase domains of Drp1 form dimers between rungs of the helix, stabilizing the active site and stimulating GTPase activity [114]. Unlike dynamin, Drp1 does not have a PHD for binding membrane lipids. Instead, the variable domain of Drp1 mediates binding to cardiolipin [32,115,116] and is necessary for the oligomerization of Drp1. This interaction with cardiolipin may also contribute to Drp1-catalyzed membrane fission [32,117], downstream of Drp1 binding to protein adaptors, which are critical for initial Drp1-mitochondrial recruitment [118,119].
Questions remain about the mechanism of Drp1. For example, there are no reported studies of Drp1-mediated fission in vitro. Furthermore, the recent finding that a knockout of dynamin 2 results in elongated mitochondria suggests that dynamin 2 performs the scission step or works in tandem with Drp1 during mitochondrial fission [120]. Based on this work, a new model is emerging where Drp1 constricts mitochondria in mammalian cells, while dynamin 2 is recruited to mediate the final fission step.
Vps1p
Vps1p mediates both fission and fusion of vacuoles and may have a role in endocytosis within yeast [25,121]. It has the essential dynamin domains, the GTPase and α-helical bundle domains, and its cellular localization suggests a role in retrograde traffic between endosomes and the Golgi [24,122]. The bifunctional role of Vps1p was postulated from observations that yeast deficient in this protein had both enlarged and small vacuoles, potentially due to defects in fission and fusion, respectively [25]. The dual function of Vps1p is further supported by observations that the dynamin inhibitor dynasore partially blocks vacuolar fission and fusion [123]. It is speculated that Vps1p functions in membrane fusion by binding VAM3 SNARE and regulating the formation of the trans-SNARE complex [123,124]. By contrast, fission of endosomes by Vps1p is enhanced by Mvp1 localized at endosomes [122]. Future structural studies on Vps1p will help address outstanding questions on the mechanism of membrane fission and fusion, including how Vps1p interacts with lipid and protein partners.
Dynamin Superfamily Proteins Involved in Membrane Fusion
Atlastin
Atlastin (Sey1p in yeast) mediates the fusion of ER membranes to maintain ER morphology [17,18,125–127]. Fusion of ER membranes maintains the interconnected tubuloreticular structure of the ER, providing a large surface area [128]. The critical role of atlastin is supported by studies showing defective ER tubule interconnection in cells deficient in atlastin, and the identification of mutations that cause premature termination of atlastin translation in individuals with the disease hereditary spastic paraplegia [129,130].
Atlastin and Sey1p have a cytosolic N-terminal GTPase domain followed by a HB analogous to the BSE domain of other DSPs (Figure 2A,F). The TM downstream of the HB and a cytoplasmic tail with an amphipathic helix at the C terminus, anchor the protein in the ER (Figure 5A,B). The cytosolic domains mediate membrane tethering, while the TM is required for fusion [131]. The amphipathic C-terminal helix also contributes to membrane fusion by destabilizing the lipid bilayer [132]. Cycling of atlastin through the GTPase cycle regulates membrane tethering and fusion. Upon GTP hydrolysis, atlastin homodimerization occurs either on the same (cis) or different membranes (trans) [131]. X-ray structures of atlastin have captured distinct steps in the fusion mechanism (Figure 5A,B). Atlastin bound to GDP shows two dimer forms: form 1 has a strong interaction between the GTPase domains and swapped HB domains, whereas form 2 has only a dimerization interface between GTPase domains [46,133]. Form 2 is believed to represent a state that can form in trans (atlastin on separate membranes) resulting in membrane fusion. Crystal form 1 is believed to represent a post-fusion state, still bound to GDP, between forms 2 and 3. The nonhydrolyzable GTP analog GMPPNP and the transition state analog GDP.AlF4− (form 3) captures an intermediate that may occur shortly before or after GTP hydrolysis [134]. This atlastin dimer also has swapped HB domains, similar to form 1, but closer together and, thus, capable of bringing membranes in tight contact for fusion. Dissociation of nucleotide leads to the formation of atlastin monomers. It is yet to be determined how fusion occurs via a hemifusion intermediate and how atlastin assembles on membranes to promote fusion.
Figure 5. Mechanism of Dynamin Superfamily Protein (DSP)-Mediated Membrane Fusion based on Structural Models.

(A) Model of membrane fusion mediated by dynamin family members. Distinct domains are colored; GTPase (green), BSE/HB/HB1 (blue), stalk and HB2 (pink), lipid-interacting domain (orange) and the lipid bilayer (magenta). Dimerization of dynamin proteins tethers two membranes. Upon GTP hydrolysis, further conformational changes lead to membrane docking and fusion. (B) Model of atlastin dimers in the extended and bent forms during endoplasmic reticulum membrane fusion. (C) Mitofusin at various steps of mitochondrial outer membrane fusion. (D) Proposed mechanism of OPA1-mediated fusion of inner mitochondrial membranes. (E) Structures of bacterial dynamin-like protein (BDLP) reveal a potential mechanism for membrane fusion during bacterial cytokinesis. Abbreviations: CL, cardiolipin; CLBD, cardiolipin-binding domain.
Mitofusins
Mitochondrial fusion requires the coordinated fusion of the outer and inner mitochondrial membranes (OMM and IMM, respectively) mediated by mitofusins and OPA1, respectively [14,135,136]. Fusion of mitochondria is a vital cellular process, as illustrated by the essential role of mitofusins during embryonic development of mice [136]. The role of mitofusins in membrane fusion was elegantly demonstrated when targeting it to the ER was sufficient to cause ER membrane fusion [137].
Mitofusins are proposed to mediate membrane fusion by a similar mechanism to atlastin (Figure 5). Conformational changes in these proteins cause docked membranes to move closer together for fusion. Alternatively, destabilization of membranes by fusion proteins could promote the fusion of membranes. Mitofusins have a cytosolic N-terminal GTPase and two helical bundle domains (HB1 and HB2), as well as the transmembrane domain [35] (Figure 5C). As with atlastin, structures of mitofusin in different nucleotide-bound states are beginning to elucidate the mechanism of membrane fusion (Figure 5A,C). Structures of mitofusin show that the GDP.AlF4− bound state is in an extended open conformation [48], which transitions to the closed state when bound to GDP.BeF3− [35,48]. These findings suggest a model where mitofusins from two membranes dimerizes upon GTP binding to tether the membranes. GTP hydrolysis results in a ~70° swing of HB1 to bring the two membranes closer together for fusion [48].
Given that the current structures of mitofusins do not contain both HB domains, it will be informative to determine how these domains are affected during the GTPase cycle. Likewise, obtaining the structures of larger assemblies of mitofusin in complex with membranes will contribute to revealing the mechanism of mitofusins.
OPA1
OPA1 has a key role in the fusion of the IMM. Coordination of IMM and OMM fusion by OPA1 and mitofusins is crucial for mitochondrial network integrity and metabolism [138,139]. Insights into its function originated from studies of the OPA1 ortholog Mgm1p in S. cerevisiae [140]. EM captured impaired mitochondrial fusion in yeast with defective Mgm1p [141]. In humans, OPA1 is also crucial for mitochondrial maintenance. For example, clumping of mitochondria was observed in monocytes obtained from an individual with optic neuropathy due to a deletion in the C terminus of OPA1 [15]. Given the targeting and localization of OPA1 to the IMM, it has been implicated specifically in the fusion of this membrane [16,142].
OPA1 exists as either a long form (OPA1L), which contains a TM, or a soluble short form (OPA1S), which is generated post-translationally after cleavage by the YmeL and OMA1 inner membrane peptidases [142–144]. OPA1S and the bulk of OPA1L reside in the mitochondrial intermembrane space. The current model for fusion involves OPA1L residing in one membrane binding to cardiolipin on the target membrane (Figure 5D) [138,145]. A region between residues 581 and 941 (CLBD) mediates binding to cardiolipin [145]. The OPA1S may work in concert with the long form by dimerizing with OPA1L on one end and interacting with the target membrane on the other (Figure 5D) [145]. GTP hydrolysis triggers a conformational change that brings the two membranes together for fusion, which is enhanced by destabilization of the membranes by OPA1S [146]. Determining the structures of OPA1 in different nucleotide-bound states will help elucidate how GTP hydrolysis is coupled to membrane fusion. In addition, a structure of OPA1 bound to cardiolipin will clarify how this interaction stimulates GTPase activity and contributes to membrane fusion.
Bacterial Dynamin-Like Proteins
BDLPs function in bacterial cytokinesis and vesicle secretion. The best-characterized proteins include BDLPs from Nostoc punctiforme, DynA and DynB from Streptomyces, and Leo A, B, and C from enterotoxigenic E. coli [26–28,59]. The role of BDLP, DynA, and DynB in cytokinesis is supported by their localization at the septal ring of dividing bacteria, and that the knockout of these proteins results in deficient bacterial division [26,27]. Bacterial cytokinesis requires membrane fusion at the septal ring, and this fusion activity has been demonstrated for DynA in vitro [147]. Unlike BDLP and DynA, Leo proteins are localized in the periplasm, not the cytoplasm or septal ring. Knockout of these proteins in enterotoxigenic E. coli results in impaired vesicle secretion, but the mechanism by which Leo proteins mediate vesicle secretion remains unknown [28].
Bacterial dynamin-like proteins comprise the globular GTPase and helical stalk domains that are linked by the BSE (also referred to as the helical neck) [26,28,59]. In the nucleotide-free and GDP state, BDLP is in a V-shaped closed conformation, similar to the closed Drp1 structure (Figure 5E) [26,31]. The cryo-EM structure of BDLP-GMPPNP bound to lipid tubes shows the protein in an extended linear conformation with the stalk domain no longer interacting with the GTPase domain (Figure 5E). In this conformation, lipid binding is mediated by a paddle region, specifically residues L576, L577, and F584 (Figure 5E). For DynA, lipid binding also occurs through a domain that extends from the stalk, referred to as the D1 module [147]. The drastic change in bacterial DSPs from an extended GTP-bound state to a closed GDP-bound state hints at its mechanism in membrane fusion. It is possible that the extended form tethers two membranes that bend upon GTP hydrolysis, drawing membranes together for fusion.
Dynamin Superfamily Proteins Involved in Innate Immune Response to Pathogens
Guanylate-Binding Proteins
The interferon-inducible GBPs are involved in the innate immune response to diverse intracellular pathogens, including viruses, bacteria, and eukaryotic parasites [21,148]. GBPs are targeted to microbe-containing vacuoles, where they may trap or inhibit the microbes [149,150] (Figure 6A). Targeting to vacuoles is proposed to be enabled by the isoprenylation of GBP1 at the CaaX box motif. Additionally, the polybasic residues preceding the CaaX box motif may direct binding to negatively charged phospholipids [60]. The crystal structure of GBP1 reveals that it is a monomer in the nucleotide-free state and transitions to a dimer when bound to nucleotides [36,151] (Figure 6B). It has been proposed that targeting and assembly of GBP to microbe-containing vacuoles restricts pathogens by blocking escape from the vacuole or by activating host immune responses, including the inflammasome and activation of autophagy [150].
Figure 6. Dynamin Superfamily Proteins (DSPs) in Innate Immune Response to Pathogens.

(A) Guanylate-binding proteins (GBP) interact with vacuole membranes and may recruit other immune factors to restrict pathogens, such as viruses (1) or Toxoplasma gondii (2). (B) Structure of GBP1 showing functional domains and proposed dimer assembly. (C) MxA interacts with ribonucleoprotein to restrict transcription and replication of influenza virus (1). MxB interacts directly with viral nucleocapsid to prevent uncoating and import of HIV-1 viral RNA into the nucleus (2). (D) Structure of MxB showing domains and dimer assembly. The bottom figure is the structure of the MxB oligomer determined by cryo-electron microscopy. Interfaces are labeled 1, 2, and 3.
Mx Proteins
The interferon-stimulated MX genes are found in vertebrates and confer resistance mainly to RNA viruses (Figure 6C). For example, MxA inhibits nuclear import of influenza nucleocapsid, and MxB binds HIV-1 capsid to prevent uncoating [152–155]. Both Mx proteins have the GTPase, BSE, and stalk domains that are characteristic of DSPs, as well as a lipid-binding L4 loop (Figure 6D). In humans, MxA and MxB restrict influenza and HIV-1, respectively (Figure 6C) and their antiviral activity depends on GTP binding and hydrolysis [152]. Assembly of Mx proteins is also critical for their antiviral activity. Structural studies reveal that Mx proteins assemble into dimers, tetramers, and higher-order oligomers via interfaces 1, 2, and 3 between stalks (Figure 6D), similar to other dynamin polymers, and an additional interface (4) exists between rungs of the helix [33,153,156,157]. The L4 loop mediates binding to negatively charged lipids via a motif comprising four adjacent lysine residues (residues 554–557) and is proposed to regulate viral specificity [58,158]. Following the remarkable structural studies over the past decade, determining how Mx proteins specifically interact with viral components will be informative.
Concluding Remarks
DSPs are found in all cells, where they primarily mediate membrane-remodeling events, such as endocytosis and organelle maintenance in eukaryotes, and cytokinesis in bacteria. DSPs also have roles in interacting with the cytoskeleton during eukaryotic cell division and migration, and in innate immunity to diverse pathogens. Even though DSPs have diverse functions, they all have a globular GTPase domain and at least one adjoining elongated α-helical bundle domain. This suggests that DSPs function by a common theme of GTP binding and/or hydrolysis that results in conformational changes in the α-helical bundle domains. Dynamin family proteins also have the common theme of self-assembly driving function. Moreover, the unique roles of specific DSPs may be due to targeting to specific sites via lipid-interacting motifs, such as the TM of atlastin and mitofusin, or via protein partners, such as SH3 domain-containing proteins, which recruit dynamin to endocytic sites.
Our understanding of the molecular mechanisms of DSPs during membrane fission and fusion has been advanced largely by structural studies of DSPs in various nucleotide-bound states using X-ray crystallography and cryo-EM. It is essential to further pursue structural studies of these membrane-remodeling proteins in membrane environments, a challenge that is addressable by cryo-EM. Another significant gap in our knowledge is the structural and molecular basis by which the GBPs and Mx proteins recognize pathogen markers for inhibition. Elucidation of these recognition mechanisms may then be exploited to develop therapeutics against pathogens such as HIV, influenza and protozoan parasites (see Outstanding Questions). Structural studies over the past two decades have captured snapshots of DSPs in various states of nucleotide binding and hydrolysis, and begin to reveal how DSPs self-assemble, interact with other proteins or membranes, and function. It is exciting to postulate that continued structural studies may reveal close to a complete picture on the molecular mechanism of how DSPs function in the cell.
Outstanding Questions.
What is the role of dynamin in the final step of membrane fission when membranes undergo scission?
What are the molecular mechanisms by which interacting proteins regulate DSPs?
How do disease-associated mutations in DSPs cause defects in protein function and result in disease? For example, mutations throughout OPA1 lead to optic atrophy and yet the mechanism remains unknown.
What is the structural arrangement of DSP membrane fusion proteins in opposing membranes? Do they form sheets or rings that mediate membrane tethering and subsequently fusion?
What is the mechanism of membrane fusion by DSPs? Is it accomplished by active tethering and fusion of membranes by DSPs or by destabilization of membranes?
How do Mx proteins interact with viral components to restrict viruses?
Highlights.
DSPs are found in all organisms, from bacteria to humans.
DSPs function in membrane fission and fusion, bundling of microtubules, and during the innate immune response to microbes.
Common structural features of DSPs include the presence of a globular GTPase domain with an adjacent α-helical bundle domain.
The mechanochemical properties of DSPs are driven by GTP hydrolysis that results in conformational changes in the α-helical bundle domains.
Different DSPs have unique features, such as distinct lipid-binding motifs and protein–protein interaction domains, which target them to specific sites where they perform specific functions.
Acknowledgments
We are grateful to colleagues in the dynamin field and apologize to those whose studies were not included in this review due to space limitations. We thank Bertram Canagarajah, Paula Flicker, Andrew Kehr, Melissa Mikolaj, Leopold Kong, Shannon Ohlemacher, Abigail Stanton and Huaibin Wang for their discussions on this review article. We are also grateful to Jason Mears for providing us with EM maps of Drp1. This work is supported by the intramural research program at the NIDDK, NIH.
References
- 1.Kosaka T and Ikeda K (1983) Possible temperature-dependent blockage of synaptic vesicle recycling induced by a single gene mutation in Drosophila. J. Neurobiol. 14, 207–225 [DOI] [PubMed] [Google Scholar]
- 2.Chen MS et al. (1991) Multiple forms of dynamin are encoded by shibire, a Drosophila gene involved in endocytosis. Nature 351, 583–586 [DOI] [PubMed] [Google Scholar]
- 3.Obar RA et al. (1990) Molecular cloning of the microtubule-associated mechanochemical enzyme dynamin reveals homology with a new family of GTP-binding proteins. Nature 347, 256–261 [DOI] [PubMed] [Google Scholar]
- 4.Shpetner HS and Vallee RB (1989) Identification of dynamin, a novel mechanochemical enzyme that mediates interactions between microtubules. Cell 59, 421–432 [DOI] [PubMed] [Google Scholar]
- 5.Antonny B et al. (2016) Membrane fission by dynamin: what we know and what we need to know. EMBO J. 35, 2270–2284 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Smirnova E et al. (1998) A human dynamin-related protein controls the distribution of mitochondria. J. Cell Biol. 143, 351–358 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Otsuga D et al. (1998) The dynamin-related GTPase, Dnm1p, controls mitochondrial morphology in yeast. J. Cell Biol. 143, 333–349 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Koch A et al. (2003) Dynamin-like protein 1 is involved in peroxisomal fission. J. Biol. Chem. 278, 8597–8605 [DOI] [PubMed] [Google Scholar]
- 9.Kuravi K et al. (2006) Dynamin-related proteins Vps1p and Dnm1p control peroxisome abundance in Saccharomyces cerevisiae. J. Cell Sci. 119, 3994–4001 [DOI] [PubMed] [Google Scholar]
- 10.Li X and Gould SJ (2003) The dynamin-like GTPase DLP1 is essential for peroxisome division and is recruited to peroxisomes in part by PEX11. J. Biol. Chem. 278, 17012–17020 [DOI] [PubMed] [Google Scholar]
- 11.Osteryoung KW and Pyke KA (2014) Division and dynamic morphology of plastids. Annu. Rev. Plant Biol. 65, 443–472 [DOI] [PubMed] [Google Scholar]
- 12.Santel A and Fuller MT (2001) Control of mitochondrial morphology by a human mitofusin. J. Cell Sci. 114, 867–874 [DOI] [PubMed] [Google Scholar]
- 13.Rapaport D et al. (1998) Fzo1p is a mitochondrial outer membrane protein essential for the biogenesis of functional mitochondria in Saccharomyces cerevisiae. J. Biol. Chem. 273, 20150–20155 [DOI] [PubMed] [Google Scholar]
- 14.Hermann GJ et al. (1998) Mitochondrial fusion in yeast requires the transmembrane GTPase Fzo1p. J. Cell Biol. 143, 359–373 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Delettre C et al. (2000) Nuclear gene OPA1, encoding a mitochondrial dynamin-related protein, is mutated in dominant optic atrophy. Nat. Genet. 26, 207–210 [DOI] [PubMed] [Google Scholar]
- 16.Olichon A et al. (2002) The human dynamin-related protein OPA1 is anchored to the mitochondrial inner membrane facing the inter-membrane space. FEBS Lett. 523, 171–176 [DOI] [PubMed] [Google Scholar]
- 17.Orso G et al. (2009) Homotypic fusion of ER membranes requires the dynamin-like GTPase atlastin. Nature 460, 978–983 [DOI] [PubMed] [Google Scholar]
- 18.Hu J et al. (2009) A class of dynamin-like GTPases involved in the generation of the tubular ER network. Cell 138, 549–561 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zhang M et al. (2013) ROOT HAIR DEFECTIVE3 family of dynamin-like GTPases mediates homotypic endoplasmic reticulum fusion and is essential for Arabidopsis development. Plant Physiol. 163, 713–720 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Haller O et al. (1981) Virus-specific interferon action: Protection of newborn Mx carriers against lethal infection with influenza virus. J. Exp. Med. 154, 199–203 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Anderson SL et al. (1999) Interferon-induced guanylate binding protein-1 (GBP-1) mediates an antiviral effect against vesicular stomatitis virus and encephalomyocarditis virus. Virology 256, 8–14 [DOI] [PubMed] [Google Scholar]
- 22.Degrandi D et al. (2007) Extensive characterization of IFN-induced GTPases mGBP1 to mGBP10 involved in host defense. J. Immunol. 179, 7729–7740 [DOI] [PubMed] [Google Scholar]
- 23.Strehlow I et al. (1994) The interferon-inducible GBP1 gene: structure and mapping to human chromosome 1. Gene 144, 295–299 [DOI] [PubMed] [Google Scholar]
- 24.Rothman JH et al. (1990) A putative GTP binding protein homologous to interferon-inducible Mx proteins performs an essential function in yeast protein sorting. Cell 61, 1063–1074 [DOI] [PubMed] [Google Scholar]
- 25.Peters C et al. (2004) Mutual control of membrane fission and fusion proteins. Cell 119, 667–678 [DOI] [PubMed] [Google Scholar]
- 26.Low HH and Löwe J (2006) A bacterial dynamin-like protein. Nature 444, 766–769 [DOI] [PubMed] [Google Scholar]
- 27.Schlimpert S et al. (2017) Two dynamin-like proteins stabilize FtsZ rings during. Proc. Natl. Acad. Sci. U. S. A. 114, E6176–E6183 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Michie KA et al. (2014) LeoA, B and C from enterotoxigenic Escherichia coli (ETEC) are bacterial dynamins. PLoS One 9, e107211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kong L et al. (2018) Cryo-EM of the dynamin polymer assembled on lipid membrane. Nature 560, 258–262 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kalia R et al. (2018) Structural basis of mitochondrial receptor binding and constriction by DRP1. Nature 558, 401–405 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Bohuszewicz O and Low HH (2018) Structure of a mitochondrial fission dynamin in the closed conformation. Nat. Struct. Mol. Biol. 25, 722–731 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Francy CA et al. (2017) Cryo-EM studies of Drp1 reveal cardiolipin interactions that activate the helical oligomer. Sci. Rep. 7, 10744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Alvarez FJD et al. (2017) CryoEM structure of MxB reveals a novel oligomerization interface critical for HIV restriction. Sci. Adv. 3, e1701264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Yan L et al. (2018) Structural basis for GTP hydrolysis and conformational change of MFN1 in mediating membrane fusion. Nat. Struct. Mol. Biol. 25, 233–243 [DOI] [PubMed] [Google Scholar]
- 35.Cao YL et al. (2017) MFN1 structures reveal nucleotide-triggered dimerization critical for mitochondrial fusion. Nature 542, 372–376 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Prakash B et al. (2000) Structure of human guanylate-binding protein 1 representing a unique class of GTP-binding proteins. Nature 403, 567–571 [DOI] [PubMed] [Google Scholar]
- 37.Niemann HH et al. (2001) Crystal structure of a dynamin GTPase domain in both nucleotide-free and GDP-bound forms. EMBO J. 20, 5813–5821 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bramkamp M (2012) Structure and function of bacterial dynamin-like proteins. Biol. Chem. 393, 1203–1214 [DOI] [PubMed] [Google Scholar]
- 39.Marks B et al. (2001) GTPase activity of dynamin and resulting conformation change are essential for endocytosis. Nature 410, 231–235 [DOI] [PubMed] [Google Scholar]
- 40.Song BD et al. (2004) Dynamin GTPase domain mutants that differentially affect GTP binding, GTP hydrolysis, and clathrin-mediated endocytosis. J. Biol. Chem. 279, 40431–40436 [DOI] [PubMed] [Google Scholar]
- 41.Damke H et al. (1994) Induction of mutant dynamin specifically blocks endocytic coated vesicle formation. J. Cell Biol. 127, 915–934 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Gasper R et al. (2009) It takes two to tango: regulation of G proteins by dimerization. Nat. Rev. Mol. Cell Biol. 10, 423–429 [DOI] [PubMed] [Google Scholar]
- 43.Chappie JS et al. (2010) G domain dimerization controls dynamin’s assembly-stimulated GTPase activity. Nature 465, 435–440 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Praefcke GJ and McMahon HT (2004) The dynamin superfamily: universal membrane tubulation and fission molecules? Nat. Rev. Mol. Cell Biol. 5, 133–147 [DOI] [PubMed] [Google Scholar]
- 45.Scheffzek K et al. (1997) The Ras-RasGAP complex: structural basis for GTPase activation and its loss in oncogenic Ras mutants. Science 277, 333–338 [DOI] [PubMed] [Google Scholar]
- 46.Bian X et al. (2011) Structures of the atlastin GTPase provide insight into homotypic fusion of endoplasmic reticulum membranes. Proc. Natl. Acad. Sci. U. S. A 108, 3976–3981 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Ghosh A et al. (2006) How guanylate-binding proteins achieve assembly-stimulated processive cleavage of GTP to GMP. Nature 440, 101–104 [DOI] [PubMed] [Google Scholar]
- 48.Yan L et al. (2018) Structural basis for GTP hydrolysis and conformational change of MFN1 in mediating membrane fusion. Nat. Struct. Mol. Biol. 25, 233–243 [DOI] [PubMed] [Google Scholar]
- 49.Chappie JS et al. (2011) A pseudoatomic model of the dynamin polymer identifies a hydrolysis-dependent powerstroke. Cell 147, 209–2222 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Chappie JS et al. (2009) An intramolecular signaling element that modulates dynamin function in vitro and in vivo. Mol. Biol. Cell 20, 3561–3571 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Narayanan R et al. (2005) An internal GAP domain negatively regulates presynaptic dynamin in vivo: a two-step model for dynamin function. J. Cell Biol. 169, 117–126 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Ford MG et al. (2011) The crystal structure of dynamin. Nature 477, 561–566 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Ferguson KM et al. (1994) Crystal structure at 2.2 A resolution of the pleckstrin homology domain from human dynamin. Cell 79, 199–209 [DOI] [PubMed] [Google Scholar]
- 54.Timm D et al. (1994) Crystal structure of the pleckstrin homology domain from dynamin. Nat. Struct. Biol. 1, 782–788 [DOI] [PubMed] [Google Scholar]
- 55.Downing AK et al. (1994) Three-dimensional solution structure of the pleckstrin homology domain from dynamin. Curr. Biol. 4, 884–891 [DOI] [PubMed] [Google Scholar]
- 56.Zhang P and Hinshaw JE (2001) Three-dimensional reconstruction of dynamin in the constricted state. Nat. Cell Biol. 3, 922–926 [DOI] [PubMed] [Google Scholar]
- 57.Clinton RW et al. (2016) Dynamin-related protein 1 oligomerization in solution impairs functional interactions with membrane-anchored mitochondrial fission factor. J. Biol. Chem. 291, 478–492 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.von der Malsburg A et al. (2011) Stalk domain of the dynamin-like MxA GTPase protein mediates membrane binding and liposome tubulation via the unstructured L4 loop. J. Biol. Chem. 286, 37858–37865 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Low HH et al. (2009) Structure of a bacterial dynamin-like protein lipid tube provides a mechanism for assembly and membrane curving. Cell 139, 1342–1352 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Fres JM et al. (2010) Purification of the CaaX-modified, dynamin-related large GTPase hGBP1 by coexpression with farnesyltransferase. J. Lipid Res. 51, 2454–2459 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Mooren OL et al. (2009) Dynamin2 GTPase and cortactin remodel actin filaments. J. Biol. Chem. 284, 23995–24005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Lee E and De Camilli P (2002) Dynamin at actin tails. Proc. Natl. Acad. Sci. U. S. A. 99, 161–166 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ishida N et al. (2011) Dynamin 2 associates with microtubules at mitosis and regulates cell cycle progression. Cell Struct. Funct. 36, 145–154 [DOI] [PubMed] [Google Scholar]
- 64.Ochoa GC et al. (2000) A functional link between dynamin and the actin cytoskeleton at podosomes. J. Cell Biol. 150, 377–389 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Ferguson SM et al. (2009) Coordinated actions of actin and BAR proteins upstream of dynamin at endocytic clathrin-coated pits. Dev. Cell 17, 811–822 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Anantharam A et al. (2011) A new role for the dynamin GTPase in the regulation of fusion pore expansion. Mol. Biol. Cell 22, 1907–1918 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Chiang HC et al. (2014) Post-fusion structural changes and their roles in exocytosis and endocytosis of dense–core vesicles. Nat. Commun. 5, 3356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Zhao WD et al. (2016) Hemi-fused structure mediates and controls fusion and fission in live cells. Nature 534, 548–552 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Shin W et al. (2018) Visualization of membrane pore in live cells reveals a dynamic-pore theory governing fusion and endocytosis. Cell 173, 934–945 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Ferguson SM and De Camilli P (2012) Dynamin, a membrane-remodelling GTPase. Nat. Rev. Mol. Cell Biol. 13, 75–88 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Raimondi A et al. (2011) Overlapping role of dynamin isoforms in synaptic vesicle endocytosis. Neuron 70, 1100–1114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Ferguson SM et al. (2007) A selective activity-dependent requirement for dynamin 1 in synaptic vesicle endocytosis. Science 316, 570–574 [DOI] [PubMed] [Google Scholar]
- 73.Srinivasan S et al. (2018) A noncanonical role for dynamin-1 in regulating early stages of clathrin-mediated endocytosis in non-neuronal cells. PLoS Biol. 16, e2005377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.González-Jamett AM et al. (2017) Dynamin-2 mutations linked to centronuclear myopathy impair actin-dependent trafficking in muscle cells. Sci. Rep. 7, 4580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Faelber K et al. (2013) Oligomerization of dynamin superfamily proteins in health and disease. Prog. Mol. Biol. Transl. Sci. 117, 411–443 [DOI] [PubMed] [Google Scholar]
- 76.Nan J et al. (2017) Molecular regulation of mitochondrial dynamics in cardiac disease. Biochim. Biophys. Acta 1864, 1260–1273 [DOI] [PubMed] [Google Scholar]
- 77.Tanabe K and Takei K (2009) Dynamic instability of microtubules requires dynamin 2 and is impaired in a Charcot-Marie-Tooth mutant. J. Cell Biol. 185, 939–948 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Sambuughin N et al. (2015) Adult-onset autosomal dominant spastic paraplegia linked to a GTPase-effector domain mutation of dynamin 2. BMC Neurol. 15, 223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Hinshaw JE and Schmid SL (1995) Dynamin self-assembles into rings suggesting a mechanism for coated vesicle budding. Nature 374, 190–192 [DOI] [PubMed] [Google Scholar]
- 80.Sweitzer SM and Hinshaw JE (1998) Dynamin undergoes a GTP-dependent conformational change causing vesiculation. Cell 93, 1021–1029 [DOI] [PubMed] [Google Scholar]
- 81.Pucadyil TJ and Schmid SL (2008) Real-time visualization of dynamin-catalyzed membrane fission and vesicle release. Cell 135, 1263–1275 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Roux A et al. (2006) GTP-dependent twisting of dynamin implicates constriction and tension in membrane fission. Nature 441, 528–531 [DOI] [PubMed] [Google Scholar]
- 83.Shpetner HS et al. (1996) A binding site for SH3 domains targets dynamin to coated pits. J. Biol. Chem. 271, 13–16 [DOI] [PubMed] [Google Scholar]
- 84.Sundborger A et al. (2011) An endophilin-dynamin complex promotes budding of clathrin-coated vesicles during synaptic vesicle recycling. J. Cell Sci. 124, 133–143 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Hohendahl A et al. (2017) Structural inhibition of dynamin-mediated membrane fission by endophilin. eLife 6, e26856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Evergren E et al. (2007) Intersectin is a negative regulator of dynamin recruitment to the synaptic endocytic zone in the central synapse. J. Neurosci. 27, 379–390 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Anggono V et al. (2006) Syndapin I is the phosphorylation-regulated dynamin I partner in synaptic vesicle endocytosis. Nat. Neurosci. 9, 752–760 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Cowling BS et al. (2017) Amphiphysin (BIN1) negatively regulates dynamin 2 for normal muscle maturation. J. Clin. Invest. 127, 4477–4487 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.McMahon HT et al. (1997) Clathrin interacts specifically with amphiphysin and is displaced by dynamin. FEBS Lett. 413, 319–322 [DOI] [PubMed] [Google Scholar]
- 90.Takei K et al. (1999) Functional partnership between amphiphysin and dynamin in clathrin-mediated endocytosis. Nat. Cell Biol. 1, 33–39 [DOI] [PubMed] [Google Scholar]
- 91.Gallop JL et al. (2006) Mechanism of endophilin N-BAR domain-mediated membrane curvature. EMBO J. 25, 2898–2910 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Ross JA et al. (2011) Dimeric endophilin A2 stimulates assembly and GTPase activity of dynamin 2. Biophys. J. 100, 729–737 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Roux A et al. (2010) Membrane curvature controls dynamin polymerization. Proc. Natl. Acad. Sci. U. S. A. 107, 4141–4146 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Lemmon MA and Ferguson KM (2000) Signal-dependent membrane targeting by pleckstrin homology (PH) domains. Biochem. J. 350 Pt 1, 1–18 [PMC free article] [PubMed] [Google Scholar]
- 95.Vallis Y et al. (1999) Importance of the pleckstrin homology domain of dynamin in clathrin-mediated endocytosis. Curr. Biol. 9, 257–260 [DOI] [PubMed] [Google Scholar]
- 96.Achiriloaie M et al. (1999) Essential role of the dynamin pleckstrin homology domain in receptor-mediated endocytosis. Mol. Cell. Biol. 19, 1410–1415 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Ramachandran R and Schmid SL (2008) Real-time detection reveals that effectors couple dynamin’s GTP-dependent conformational changes to the membrane. EMBO J. 27, 27–37 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Ramachandran R et al. (2009) Membrane insertion of the pleckstrin homology domain variable loop 1 is critical for dynamin-catalyzed vesicle scission. Mol. Biol. Cell 20, 4630–4639 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Liu YW et al. (2011) Differential curvature sensing and generating activities of dynamin isoforms provide opportunities for tissue-specific regulation. Proc. Natl. Acad. Sci. U. S. A. 108, E234–E242 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Srinivasan S et al. (2016) Identification and function of conformational dynamics in the multidomain GTPase dynamin. EMBO J. 35, 443–457 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Mehrotra N et al. (2014) Alternate pleckstrin homology domain orientations regulate dynamin-catalyzed membrane fission. Mol. Biol. Cell 25, 879–890 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Sundborger AC et al. (2014) A dynamin mutant defines a superconstricted prefission state. Cell Rep. 8, 734–742 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Reubold TF et al. (2015) Crystal structure of the dynamin tetramer. Nature 525, 404–408 [DOI] [PubMed] [Google Scholar]
- 104.Faelber K et al. (2011) Crystal structure of nucleotide-free dynamin. Nature 477, 556–560 [DOI] [PubMed] [Google Scholar]
- 105.Mattila JP et al. (2015) A hemi-fission intermediate links two mechanistically distinct stages of membrane fission. Nature 524, 109–113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Maeda K et al. (1992) Interaction of dynamin with microtubules: its structure and GTPase activity investigated by using highly purified dynamin. Mol. Biol. Cell 3, 1181–1194 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Bethoney KA et al. (2009) A possible effector role for the pleckstrin homology (PH) domain of dynamin. Proc. Natl. Acad. Sci. U. S. A. 106, 13359–13364 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Twig G et al. (2008) Fission and selective fusion govern mitochondrial segregation and elimination by autophagy. EMBO J. 27, 433–446 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Bleazard W et al. (1999) The dynamin-related GTPase Dnm1 regulates mitochondrial fission in yeast. Nat. Cell Biol. 1, 298–304 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Mears JA et al. (2011) Conformational changes in Dnm1 support a contractile mechanism for mitochondrial fission. Nat. Struct. Mol. Biol. 18, 20–26 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Fröhlich C et al. (2013) Structural insights into oligomerization and mitochondrial remodelling of dynamin 1-like protein. EMBO J. 32, 1280–1292 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Macdonald PJ et al. (2014) A dimeric equilibrium intermediate nucleates Drp1 reassembly on mitochondrial membranes for fission. Mol. Biol. Cell 25, 1905–1915 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Losón OC et al. (2013) Fis1, Mff, MiD49, and MiD51 mediate Drp1 recruitment in mitochondrial fission. Mol. Biol. Cell 24, 659–667 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Wenger J et al. (2013) Functional mapping of human dynamin-1-like GTPase domain based on x-ray structure analyses. PLoS One 8, e71835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Lu B et al. (2018) Steric interference from intrinsically disordered regions controls dynamin-related protein 1 self-assembly during mitochondrial fission. Sci. Rep. 8, 10879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Bustillo-Zabalbeitia I et al. (2014) Specific interaction with cardiolipin triggers functional activation of Dynamin-Related Protein 1. PLoS One 9, e102738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Stepanyants N et al. (2015) Cardiolipin’s propensity for phase transition and its reorganization by dynamin-related protein 1 form a basis for mitochondrial membrane fission. Mol. Biol. Cell 26, 3104–3116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Osellame LD et al. (2016) Cooperative and independent roles of the Drp1 adaptors Mff, MiD49 and MiD51 in mitochondrial fission. J. Cell Sci. 129, 2170–2181 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Otera H et al. (2016) Drp1-dependent mitochondrial fission via MiD49/51 is essential for apoptotic cristae remodeling. J. Cell Biol. 212, 531–544 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Lee JE et al. (2016) Multiple dynamin family members collaborate to drive mitochondrial division. Nature 540, 139–143 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Smaczynska-de Rooij II et al. (2010) A role for the dynamin-like protein Vps1 during endocytosis in yeast. J. Cell Sci. 123, 3496–3506 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Chi RJ et al. (2014) Fission of SNX-BAR-coated endosomal retrograde transport carriers is promoted by the dynamin-related protein Vps1. J. Cell Biol. 204, 793–806 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Alpadi K et al. (2013) Dynamin-SNARE interactions control trans-SNARE formation in intracellular membrane fusion. Nat. Commun. 4, 1704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Kulkarni A et al. (2014) A dynamin homolog promotes the transition from hemifusion to content mixing in intracellular membrane fusion. Traffic 15, 558–571 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Muriel MP et al. (2009) Atlastin-1, the dynamin-like GTPase responsible for spastic paraplegia SPG3A, remodels lipid membranes and may form tubules and vesicles in the endoplasmic reticulum. J. Neurochem. 110, 1607–1616 [DOI] [PubMed] [Google Scholar]
- 126.Rismanchi N et al. (2008) Atlastin GTPases are required for Golgi apparatus and ER morphogenesis. Hum. Mol. Genet. 17, 1591–1604 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Anwar K et al. (2012) The dynamin-like GTPase Sey1p mediates homotypic ER fusion in S. cerevisiae. J. Cell Biol. 197, 209–217 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Lee C and Chen LB (1988) Dynamic behavior of endoplasmic reticulum in living cells. Cell 54, 37–46 [DOI] [PubMed] [Google Scholar]
- 129.Tessa A et al. (2002) SPG3A: an additional family carrying a new atlastin mutation. Neurology 59, 2002–2005 [DOI] [PubMed] [Google Scholar]
- 130.Khan TN et al. (2014) Evidence for autosomal recessive inheritance in SPG3A caused by homozygosity for a novel ATL1 missense mutation. Eur. J. Hum. Genet. 22, 1180–1184 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Liu TY et al. (2015) Cis and trans interactions between atlastin molecules during membrane fusion. Proc. Natl. Acad. Sci. U. S. A. 112, E1851–E1860 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Faust JE et al. (2015) The Atlastin C-terminal tail is an amphipathic helix that perturbs the bilayer structure during endoplasmic reticulum homotypic fusion. J. Biol. Chem. 290, 4772–4783 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Byrnes LJ and Sondermann H (2011) Structural basis for the nucleotide-dependent dimerization of the large G protein atlastin-1/SPG3A. Proc. Natl. Acad. Sci. U. S. A. 108, 2216–2221 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Byrnes LJ et al. (2013) Structural basis for conformational switching and GTP loading of the large G protein atlastin. EMBO J. 32, 369–384 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Hales KG and Fuller MT (1997) Developmentally regulated mitochondrial fusion mediated by a conserved, novel, predicted GTPase. Cell 90, 121–129 [DOI] [PubMed] [Google Scholar]
- 136.Chen H et al. (2003) Mitofusins Mfn1 and Mfn2 coordinately regulate mitochondrial fusion and are essential for embryonic development. J. Cell Biol. 160, 189–200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Huang X et al. (2017) Sequences flanking the transmembrane segments facilitate mitochondrial localization and membrane fusion by mitofusin. Proc. Natl. Acad. Sci. U. S. A. 114, E9863–E9872 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Song Z et al. (2009) Mitofusins and OPA1 mediate sequential steps in mitochondrial membrane fusion. Mol. Biol. Cell 20, 3525–3532 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Detmer SA and Chan DC (2007) Functions and dysfunctions of mitochondrial dynamics. Nat. Rev. Mol. Cell Biol. 8, 870–879 [DOI] [PubMed] [Google Scholar]
- 140.Wong ED et al. (2003) The intramitochondrial dynamin-related GTPase, Mgm1p, is a component of a protein complex that mediates mitochondrial fusion. J. Cell Biol. 160, 303–311 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Meeusen S et al. (2006) Mitochondrial inner-membrane fusion and crista maintenance requires the dynamin-related GTPase Mgm1. Cell 127, 383–395 [DOI] [PubMed] [Google Scholar]
- 142.Song Z et al. (2007) OPA1 processing controls mitochondrial fusion and is regulated by mRNA splicing, membrane potential, and Yme1L. J. Cell Biol. 178, 749–755 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Anand R et al. (2014) The i-AAA protease YME1L and OMA1 cleave OPA1 to balance mitochondrial fusion and fission. J. Cell Biol. 204, 919–929 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Kaser M et al. (2003) Oma1, a novel membrane-bound metallopeptidase in mitochondria with activities overlapping with the m-AAA protease. J. Biol. Chem. 278, 46414–46423 [DOI] [PubMed] [Google Scholar]
- 145.Ban T et al. (2017) Molecular basis of selective mitochondrial fusion by heterotypic action between OPA1 and cardiolipin. Nat. Cell Biol. 19, 856–863 [DOI] [PubMed] [Google Scholar]
- 146.DeVay RM et al. (2009) Coassembly of Mgm1 isoforms requires cardiolipin and mediates mitochondrial inner membrane fusion. J. Cell Biol. 186, 793–803 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Bürmann F et al. (2011) A bacterial dynamin-like protein mediating nucleotide-independent membrane fusion. Mol. Microbiol. 79, 1294–1304 [DOI] [PubMed] [Google Scholar]
- 148.Selleck EM et al. (2013) Guanylate-binding protein 1 (Gbp1) contributes to cell-autonomous immunity against Toxoplasma gondii. PLoS Pathog. 9, e1003320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Kim BH et al. (2012) IFN-inducible GTPases in host cell defense. Cell Host Microbe 12, 432–444 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Praefcke GJK (2017) Regulation of innate immune functions by guanylate-binding proteins. Int. J. Med. Microbiol. Published online November 2, 2017. 10.1016/j.ijmm.2017.10.013 [DOI] [PubMed] [Google Scholar]
- 151.Prakash B et al. (2000) Triphosphate structure of guanylate-binding protein 1 and implications for nucleotide binding and GTPase mechanism. EMBO J. 19, 4555–4564 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Haller O et al. (2015) Mx GTPases: dynamin-like antiviral machines of innate immunity. Trends Microbiol. 23, 154–163 [DOI] [PubMed] [Google Scholar]
- 153.Fribourgh JL et al. (2014) Structural insight into HIV-1 restriction by MxB. Cell Host Microbe 16, 627–638 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Kochs G and Haller O (1999) Interferon-induced human MxA GTPase blocks nuclear import of Thogoto virus nucleocapsids. Proc. Natl. Acad. Sci. U. S. A. 96, 2082–2086 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Fricke T et al. (2014) MxB binds to the HIV-1 core and prevents the uncoating process of HIV-1. Retrovirology 11, 68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Gao S et al. (2010) Structural basis of oligomerization in the stalk region of dynamin-like MxA. Nature 465, 502–506 [DOI] [PubMed] [Google Scholar]
- 157.Gao S et al. (2011) Structure of myxovirus resistance protein a reveals intra- and intermolecular domain interactions required for the antiviral function. Immunity 35, 514–525 [DOI] [PubMed] [Google Scholar]
- 158.Mitchell PS et al. (2012) Evolution-guided identification of antiviral specificity determinants in the broadly acting interferon-induced innate immunity factor MxA. Cell Host Microbe 12, 598–604 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Stowell MH et al. (1999) Nucleotide-dependent conformational changes in dynamin: evidence for a mechanochemical molecular spring. Nat. Cell Biol. 1, 27–32 [DOI] [PubMed] [Google Scholar]
- 160.Barylko B et al. (1998) Synergistic activation of dynamin GTPase by Grb2 and phosphoinositides. J. Biol. Chem. 273, 3791–3797 [DOI] [PubMed] [Google Scholar]
- 161.Binns DD et al. (2000) The mechanism of GTP hydrolysis by dynamin II: a transient kinetic study. Biochemistry 39, 7188–7196 [DOI] [PubMed] [Google Scholar]
- 162.Warnock DE et al. (1996) Dynamin self-assembly stimulates its GTPase activity. J. Biol. Chem. 271, 22310–22314 [DOI] [PubMed] [Google Scholar]
- 163.Francy CA et al. (2015) The mechanoenzymatic core of dynamin-related protein 1 comprises the minimal machinery required for membrane constriction. J. Biol. Chem. 290, 11692–11703 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Macdonald PJ et al. (2016) Distinct splice variants of dynamin-related protein 1 differentially utilize mitochondrial fission factor as an effector of cooperative GTPase activity. J. Biol. Chem. 291, 493–507 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Ban T et al. (2010) OPA1 disease alleles causing dominant optic atrophy have defects in cardiolipin-stimulated GTP hydrolysis and membrane tubulation. Hum. Mol. Genet. 19, 2113–2122 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Praefcke GJ et al. (1999) Nucleotide-binding characteristics of human guanylate-binding protein 1 (hGBP1) and identification of the third GTP-binding motif. J. Mol. Biol. 292, 321–332 [DOI] [PubMed] [Google Scholar]
- 167.Richter MF et al. (1995) Interferon-induced MxA protein. GTP binding and GTP hydrolysis properties. J. Biol. Chem. 270, 13512–13517 [PubMed] [Google Scholar]
- 168.Vater CA et al. (1992) The VPS1 protein, a homolog of dynamin required for vacuolar protein sorting in Saccharomyces cerevisiae, is a GTPase with two functionally separable domains. J. Cell Biol. 119, 773–786 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Fukushima NH et al. (2001) The GTPase effector domain sequence of the Dnm1p GTPase regulates self-assembly and controls a rate-limiting step in mitochondrial fission. Mol. Biol. Cell 12, 2756–2766 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Lackner LL et al. (2009) Mechanistic analysis of a dynamin effector. Science 325, 874–877 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Meglei G and McQuibban GA (2009) The dynamin-related protein Mgm1p assembles into oligomers and hydrolyzes GTP to function in mitochondrial membrane fusion. Biochemistry 48, 1774–1784 [DOI] [PubMed] [Google Scholar]
- 172.Schweins T et al. (1995) Substrate-assisted catalysis as a mechanism for GTP hydrolysis of p21ras and other GTP-binding proteins. Nat. Struct. Biol. 2, 36–44 [DOI] [PubMed] [Google Scholar]
- 173.Hall BE et al. (2002) The structural basis for the transition from Ras-GTP to Ras-GDP. Proc. Natl. Acad. Sci. U. S. A 99, 12138. [DOI] [PMC free article] [PubMed] [Google Scholar]