

Abstract
Long non-coding RNAs have been implicated in many of the hallmarks of cancer. Herein, we found that the expression of lncRNA152 (lnc152; a.k.a. DRAIC), which we annotated previously, is highly upregulated in luminal breast cancer (LBC) and downregulated in triple-negative breast cancer (TNBC). Knockdown of lnc152 promotes cell migration and invasion in LBC cell lines. In contrast, ectopic expression of lnc152 inhibits growth, migration, invasion, and angiogenesis in TNBC cell lines. In mice, lnc152 inhibited the growth of TNBC cell xenografts, as well as metastasis of TNBC cells in an intracardiac injection model. Transcriptome analysis of the xenografts indicated that lnc152 downregulates genes controlling angiogenesis. Using pull down assays followed by LC-MS/MS, we identified RBM47, a known tumor suppressor in breast cancer, as a lnc152-interacting protein. The effects of lnc152 in TNBC cells are mediated, in part, by regulating the expression of RBM47. Collectively, our results demonstrate that lnc152 is an angiogenesis-inhibiting tumor suppressor that attenuates the aggressive cancer-related phenotypes found in TNBC.
Keywords: Angiogenesis, Cancer, lncRNA152/DRAIC, Long non-coding RNAs (LncRNAs), Luminal breast cancer, Metastasis, RBM47, Triple-negative breast cancer (TNBC), Tumor suppressor
Introduction
Metastasis leads to the spread of cancer from its primary site of origin to a distant site via the circulatory system to establish a secondary tumor. The process begins with invasion and intravasation into the surrounding lymphatics and blood vessels, and culminates with colonization of the disseminated tumor cells in a distal organ, angiogenesis, and growth (1,2). Angiogenesis is an essential component of the metastatic pathway. New blood vessels promote tumor metastasis by providing the route of exit for tumor cells from the primary tumor and colony formation at secondary sites (3,4). Moreover, new blood vessels within the tumor are needed to provide sufficient oxygen and nutrients to support tumor growth (3,4). As such, anti-angiogenic therapies have gained significant attention as a potential therapeutic approach for cancer treatment (5,6). In breast cancers, metastasis is the major cause of breast cancer-related morbidity and mortality. Breast cancer preferentially metastasizes to the lungs, brain, bone, liver, and distal lymph-nodes (7,8). A better understanding of the molecular mechanisms regulating metastasis is crucial for development of novel therapeutic strategies.
Long non-coding RNAs (lncRNAs) are non-protein coding RNA transcripts with lengths greater than 200 nucleotides, with many thousands of lncRNA genes identified in the human genome (9,10). LncRNAs mediate a host of disparate molecular and cellular functions, including gene regulation (9,10). Increasingly, lncRNAs have been implicated in many of the hallmarks of cancer, such as proliferation, growth suppression, motility, immortality, angiogenesis, and viability (11,12). Numerous lncRNAs have been found to be mutated or abnormally expressed in various cancer types (13,14), suggesting that they can potentially be used as prognostic biomarkers and therapeutic targets in cancer.
The biological functions of lncRNAs in cancer are diverse, with connections to many different aspects of cancer biology. LncRNAs may function as tumor suppressors or oncogenes during cancer progression (15). Increasing evidence indicates that lncRNAs can mediate tumor suppressive functions in a wide variety of prevalent cancer types (15–19). LncRNAs have also been shown to play a role in metastasis-related functions, such as angiogenesis (10,15). LncRNAs play a critical role in tumor angiogenesis by regulating the various underlying biological processes that control angiogenesis, including epigenetic mechanisms that control angiogenic gene expression programs (20,21). The diversity of lncRNA expression, sequences, structures, interaction partners, subcellular localization, and functions makes them ideally suited to control a broad array of critical cancer-related biological functions.
In a previous study, we used genomics and bioinformatics to discover and annotate lncRNAs controlling cell cycle gene expression and proliferation in breast cancer cells, including lncRNA152 (lnc152; a.k.a. DRAIC), which is located in a tumor suppressor locus on human chromosome 15q23 (22). In a series of functional assays, we observed key roles for lnc152 in proliferation, cell cycle progression, and regulation of the estrogen signaling pathway in breast cancer cells (22). A contemporaneous study identified lnc152/DRAIC as a tumor suppressor in prostate cancer (23). Subsequent studies have affirmed a key role for lnc152/DRAIC in the biology of an array of cancers (24–30), but the precise mechanisms by which it suppresses a wide range of cancer-related behaviors, such as proliferation, migration, invasion, metastasis, and angiogenesis, remain largely unknown. Here, our studies demonstrate that lnc152 exhibits elevated expression in the luminal subtype of breast cancer, but is downregulated in the triple-negative (TN)/basal subtype of breast cancer. Lnc152 inhibits cancer-related phenotypes, including growth, migration, invasion, metastasis, and angiogenesis by upregulating the expression of the RNA-binding tumor suppressor protein, RBM47, to control a tumor suppressing gene expression program.
Materials and Methods
Additional details about the materials and methods can be found in the Supplementary Materials and Methods under the same headings listed here.
Cell Culture and Treatments
MCF-7, MDA-MB-231, T-47D, HCC1143, 293T, primary human mammary epithelial cells (HMEC), and human umbilical vein endothelial cells (HUVEC) cells were purchased from the American Type Cell Culture (ATCC). Luciferase-expressing MDA-MB-231 cells (31) were from the lab of Dr. Srinivas Malladi. A detailed description of the culture conditions is provided in the Supplementary Materials and Methods. Fresh cell stocks were regularly replenished from the original stocks every few months, verified for cell type identity using the GenePrint 24 system (Promega, B1870), and confirmed as mycoplasma-free every three months using a commercial testing kit. For induction with doxycycline (Dox; Sigma-Aldrich, D9891), the cells were treated with 0.5–1 μg/mL of Dox for the indicated times before collection.
Molecular Cloning to Generate Expression Vectors
cDNA pools were prepared by extraction of total RNA from MCF-7 cells as described previously (32) to amplify lnc152 and RBM47 cDNAs, which were then cloned into multiple vectors such as pcDNA3, pET19b, and pINDUCER20.
Purification of RBM47 Protein Expressed in E. coli.
His-tagged human RBM47 was expressed in E. coli Rosetta (DE3) cells using the pET19b-based bacterial expression vector. The transformed bacteria were grown in LB containing ampicillin and chloramphenicol at 37 °C until the OD595 reached ~0.7. Recombinant protein expression was induced by the addition of 0.5 mM IPTG for 14 h at 20 °C. The cells were collected by centrifugation, and the cell pellets were flash-frozen in liquid N2. RBM47 was purified from the cells as described previously (32). The purified proteins were quantified using a Bradford protein assay (Bio-Rad), aliquoted, flash-frozen in liquid N2, and stored at −80°C.
Generation of Knockdown, Knockout, and Ectopic Expression Cell Lines
siRNA-mediated knockdown in MCF-7 cells.
siRNA oligos targeting human lnc152 [described previously (22)] were transfected into MCF-7 cells at a final concentration of 10 nM using Lipofectamine RNAiMAX reagent (Invitrogen, 13778150) according to the manufacturer’s instructions.
Dox-inducible ectopic expression in MDA-MB-231 and HCC1143 cells.
Lentiviruses were generated by transfection of pINDUCER20-based vectors (Addgene, plasmid no. 44012) for Dox-inducible expression of lnc152, GFP, or RBM47, along with pCMV-VSV-G, pCMV-GAG-Pol-Rev, and pAdVAntage into 293T cells using Lipofectamine 3000 according to the manufacturer’s protocol. Stably transduced cells were isolated under drug selection with 1 μg/mL Geneticin (Life Technologies, 11811031), and were used for Dox-induced ectopic expression of lnc152, GFP, and RBM47 to perform a variety of experiments described herein.
Cell Proliferation Assays
Breast cancer cells expressing lnc152, GFP, or RBM47 were plated and grown in Dox-containing medium (0.5 μg/mL), which was replenished daily, for the times noted before collection. For the cell proliferation assays, the cells were fixed and then stained using crystal violet, as previously described (32). The cells were washed to remove unincorporated stain, and the crystal violet stain was then extracted using 10% glacial acetic acid. The absorbance of the extracted stain was was read at 595 nm.
Preparation of Nuclear Extracts and Western Blotting
Whole cell lysates from cell lines and xenografts were prepared as described previously (32). Western bolting from cell and tumor tissue extracts were performed as described previously (32).
RNA Isolation and Reverse Transcription Quantitative PCR (RT-qPCR)
The cells were collected at the indicated time points and cDNA pools were prepared by extraction of total RNA from cell lines using Trizol (Sigma- Aldrich, T9424), followed by reverse transcription using MMLV reverse transcriptase (Promega, M150B) with random hexamer or oligo(dT) primers (Sigma-Aldrich). The cDNA samples were subjected to qPCR as described previously (32) using gene-specific primers.
RNA-sequencing
Generation of RNA-seq libraries.
Total RNA from seven xenograft tumor tissues of each group was isolated using an RNeasy Plus Kit (Qiagen, 74134). The RNA was assessed for quality and then used to produce strand-specific RNA-seq libraries as described previously (33). The RNA-seq libraries were subjected to QC analyses (i.e., number of PCR amplification cycles, final library yield, and final library DNA fragment size distribution) and sequenced using an Illumina NextSeq 500.
Analysis of RNA-seq data sets.
We performed QC analyses on raw data using FastQC (34). The raw reads were aligned to the human reference genome (hg19/GRCh37) using Tophat (v2.0.12) with default parameters (35). Uniquely mappable reads were converted into bigWig files using BEDTools (36) and visualized in the Integrative Genomics Viewer (37). The transcriptome was assembled using cufflinks v.2.2. (38) with default parameters. To call differential gene expression using cuffdiff, we used a distinct, non-overlapping set of transcripts generated with cuffmerge (38).
Gene set enrichment analysis.
We performed Gene Ontology (GO) analyses using an online tool called DAVID (Database for Annotation, Visualization, and Integrated Discovery) (39).
Mining of Public Databases
The expression of lnc152 and RBM47 in normal and different subtypes of breast cancer tissues was determined using GEPIA based on the RPKM values in the TCGA dataset (40).
Analysis of ChIP-seq Data
FoxA1 ChIP-seq libraries from MCF-7 cells (NCBI GEO accession number GSE59530) were generated and analyzed as described previously (41).
In Vitro Transcription to Generate GFP mRNA and lnc152
The in vitro transcribed GFP mRNA and lnc152 were prepared as described previously (32). To generate biotin-labeled RNAs for pull down assays, an NTP labeling mixture containing biotin-UTP (10 mM each ATP, CTP, and GTP, 6.5 mM UTP, 3.5 mM biotin-UTP; Sigma, 11685597910) was used for in vitro transcription.
In Vitro RNA Pull Down Assays Combined with LC-MS/MS Analysis
In vitro RNA pull down.
Whole cell extracts were prepared from MCF-7 cells, followed by incubation with 5 nM of folded RNA for 1 hour at room temperature. Equilibrated Streptavidin-agarose beads (ThermoFisher Scientific, 20377) were then added to the extracts and incubated for 30 minutes at room temperature. The protein-RNA complex-bound beads were washed four times in Binding Buffer containing an additional 200 mM of NaCl. The reactions were terminated by adding 4x SDS-PAGE Loading Buffer and heating to 100°C for 10 minutes to denature the proteins.
LC-MS/MS analysis.
Gel purified samples were analyzed using an Ultimate 3000 RSLC-Nano liquid chromatography system (Dionex) connected an Orbitrap Fusion Lumos mass spectrometer (Thermo Electron). Additional details regarding the sample processing are provided in the Supplementary Materials and Methods.
Cell Migration and Invasion Assays
Boyden chamber assays were used to determine the migration and invasive capacity of cells. Cells that migrated or invaded into the lower side of the membrane were fixed and stained with 0.5% crystal violet in 20% methanol solution for 15 minutes, washed with water, and air-dried.
In Vitro Endothelial Tube Formation Assay
Preparation of conditioned medium (CM).
The serum-free conditioned medium in the presence of 1 μg/mL of Dox was collected and concentrated using Centricon 10 (Millipore) centrifuge filter unit, aliquoted, flash-frozen in liquid N2, and stored at −80 °C.
Endothelial tube formation assay.
HUVECs were resuspended in conditioned medium from MDA-MB-231 cells expressing GFP mRNA or lnc152 and were seeded onto Matrigel Basement Membrane Matrix coated 24-well plates and incubated at 37°C for 4–12 hours. Capillary-like tubes were then stained with cell-permeable dye Calcein AM (ThermoFisher, C1430) and imaged by fluorescence microscopy.
Quantification of tube networks.
The total number of nodes, number of segments, total segment length, and branching were analyzed by the Angiogenesis Analyzer plugin in ImageJ.
Indirect Immunofluorescence
Xenograft tissues were processed for paraffin sectioning using standard protocols. After staining, the immunofluorescent signals were imaged using a Zeiss LSM880 confocal microscope.
Experiments with Mice
All animal experiments were performed in compliance with the Institutional Animal Care and Use Committee (IACUC) at the UT Southwestern Medical Center. Female athymic nude (Foxn1nu; Envigo #069) mice at 6–8 weeks of age were used.
Xenograft experiments.
Breast cancer xenografts using MDA-MB-231 cells engineered for Dox-inducible expression of GFP mRNA or lnc152 were performed as described previously (32).
Metastasis experiments.
MDA-MB-231 cells harboring a luciferase reporter were engineered for Dox-inducible expression of GFP mRNA or lnc152 and injected intracardially into the arterial circulation of 5 weeks old female athymic nude (Foxn1nu) mice (Envigo #069) to mimic hematogenous dissemination (42). Successful injection was confirmed by the detection of luciferase signal in the whole mouse.
Whole-body bioluminescent imaging.
Mice were monitored weekly for metastatic outgrowth by bioluminescence imaging. D-luciferin (150 mg/kg) was injected retro-orbitally, and the mice were imaged using IVIS Spectrum with Living Image 4.4 software (PerkinElmer).
Data and Code Availability
Genomic datasets.
The new RNA-seq data generated for this study can be accessed from the NCBI’s Gene Expression Omnibus (GEO) repository (http://www.ncbi.nlm.nih.gov/geo/) using the superseries accession number GSE193634.
Custom scripts.
Custom R scripts for genomic data analyses are available from the Lead Contact on request.
Mass spectrometry datasets.
The new mass spectrometry datasets generated for this study can be accessed from MassIVE and also are available as supplemental data provided with this manuscript.
Results
Lnc152 is highly expressed in luminal breast cancer and acts to attenuate cell migration and invasion
We previously identified lnc152 in MCF-7 luminal human breast cancer cells (22) (Figure S1, A and B). To further functionally characterize lnc152 and its role in breast cancer, we examined its expression in breast cancer patient samples and cell lines. We observed that lnc152 is highly expressed in breast cancers compared to normal breast tissues using TCGA data (Figure 1A). Moreover, upon further stratification of patient-derived breast cancer samples (TCGA), we found that lnc152 is upregulated in luminal and HER2 subtypes of breast cancer, but downregulated in basal-like breast cancer (Figure 1B). Similar results were observed in breast cancer cell lines (Figure 1C; Figure S2). In perturbation-response experiments, siRNA-mediated knockdown of lnc152 promoted the migration of and invasion by MCF-7 cells in culture-based assays (Figure 1, D and E; Figure S3). siRNA-mediated knockdown of lnc152 also altered gene expression in MCF-7 cells as assessed by RNA-seq, including the upregulation of many genes involved in cancer-related biological processes, such as extracellular matrix organization, and cell migration (Figure 1F).
Figure 1. Lnc152 is differentially expressed across the distinct molecular subtypes of breast cancer and is required for suppression of luminal breast cancer cell invasiveness.
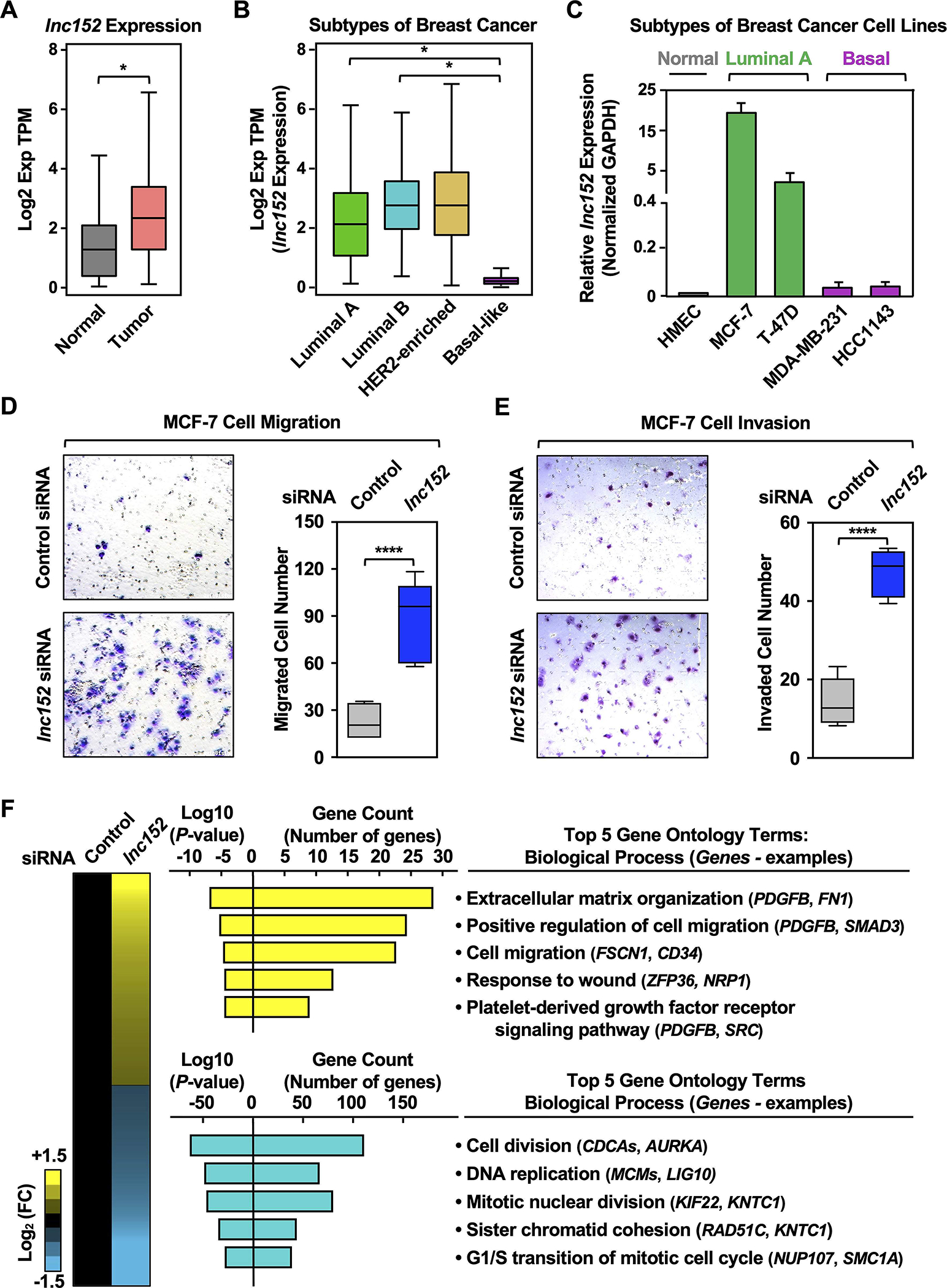
(A) RNA-seq expression data for lnc152 in primary tumors (TCGA breast cancer samples, n = 1085) compared to normal breast tissues (TCGA normal and GTEx data, n = 291). Bars marked with asterisks are statistically different from each other (two-tailed Student’s t test, * p < 0.05).
(B) Box plots comparing the expression of lnc152 in different molecular subtypes of breast cancer derived from patient samples [luminal A (n = 415), luminal B (n = 194), HER2-enriched (n = 66), and basal-like (n = 135); TCGA]. Bars marked with asterisks are statistically different from each other (two-tailed Student’s t test, * p < 0.05).
(C) Bar graphs showing the expression of lnc152 in breast cancer cell lines representing different molecular subtypes.
(D) (Left) siRNA-mediated knockdown of lnc152 promotes the cell migration of MCF-7 cells (luminal A breast cancer subtype) compared to knockdown with a control siRNA. (Right) Quantification of the results from the experiments shown in the left panel. Each box plot represents the mean ± SEM, n = 7. Bars marked with asterisks are statistically different from each other (two-tailed Student’s t test, **** p < 0.0001).
(E) (Left) siRNA-mediated knockdown of lnc152 promotes the cell invasion of MCF-7 cells (luminal A breast cancer subtype) compared to knockdown with a control siRNA. (Right) Quantification of the results from the experiments shown in the left panel. Each box plot represents the mean ± SEM, n = 6. Bars marked with asterisks are statistically different from each other (two-tailed Student’s t test, **** p < 0.0001).
(F) siRNA-mediated knockdown of lnc152 alters gene expression in MCF-7 cells. (Left) Heatmap showing the results of RNA-seq assays from lnc152-depleted MCF-7 cells. (Middle) Gene ontology analysis showing biological processes that are enriched (yellow) and those that are de-enriched (cyan). (Right) GO terms in each cluster indicating the top five biological processes determined by REViGO analysis. Genes of interest are listed in each GO term.
The enrichment and function of lnc152 in the luminal breast cancer subtype is reflected in its regulation by FoxA1, a key transcription factor in luminal breast cancers (43–46) (Figure S1C). In this regard: (1) FoxA1 binds at the promoter and upstream regions of the lnc152 gene (Figure S1B), (2) the expression of FOXA1 mRNA and lnc152 are correlated in public gene expression data sets (Figure S1D–S1F) and luminal breast cancer cell lines (Figure S1G). Finally, siRNA-mediated knockdown of FoxA1 decreases the expression level of lnc152 in luminal A breast cancer cell lines (Figure S1H). Collectively, these results indicate that lnc152 is highly expressed in luminal breast cancers and downregulated in triple-negative/basal breast cancers. In addition, we demonstrate that lnc152 regulates a gene expression program that promotes cell migration and invasion by luminal breast cancer cells.
Ectopic expression of lnc152 inhibits cancer-related cellular phenotypes in triple-negative breast cancer cells, including angiogenesis
As shown above, lnc152 is significantly downregulated in basal-like breast cancer and triple-negative breast cancer (TNBC) cells, suggesting that the low levels of lnc152 may be associated with more aggressive cancer features. To determine whether lnc152 can modulate the aggressiveness of TNBC cells, we assayed a range of cancer-related cellular phenotypes, such as cell proliferation, migration, invasion, and blood vessel formation, using TNBC cells (i.e., MDA-MB-231 and HCC1143) engineered for Dox-inducible ectopic expression of lnc152, or GFP mRNA as a control (Figure 2A; Figure S4A). Dox-induced ectopic expression of lnc152 inhibited the proliferation of MDA-MB-231 cells (Figure 2B), but not HCC1143 cells (Figure S4B), compared to ectopic expression of GFP mRNA (Figure 2B; Figure S4B). In contrast, Dox-induced ectopic expression of lnc152 inhibited migration and invasion for both cell lines, compared to ectopic expression of GFP mRNA (Figure 2, C and D; Figure S4, C and D). While both HCC1143 and MDA-MB-231 are TNBC cell lines, HCC1143 (derived from ductal carcinoma) and MDA-MB-231 (derived from adenocarcinoma) can be further stratified into different classes with different phenotypes (47), which may account for the different effects of lnc152 expression in these two cell lines.
Figure 2. Ectopic expression of lnc152 inhibits aggressive cancer-related outcomes in MDA-MB-231 TNBC cells.
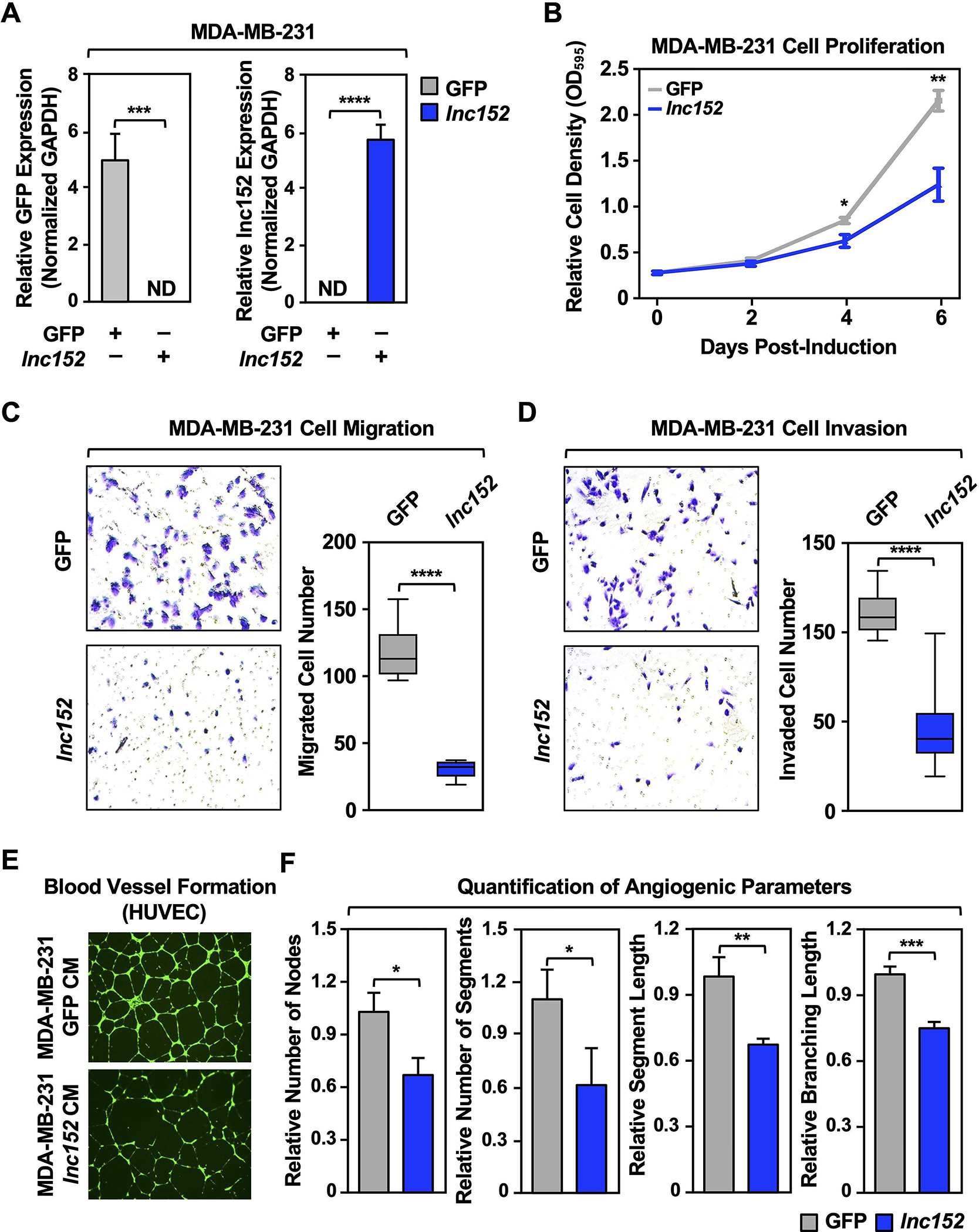
(A) Generation of MDA-MB-231 (basal breast cancer subtype) cell lines for Dox-inducible ectopic expression of GFP or lnc152. GFP mRNA and lnc152 levels were determined by RT-qPCR and normalized to GAPDH mRNA. ND indicates expression level was not detectable. Each bar represents the mean + SEM, n = 3. Bars marked with asterisks are statistically different from each other (two-tailed Student’s t test, *** p < 0.001, **** p < 0.0001).
(B) Dox-induced ectopic expression of lnc152 inhibits MDA-MB-231 cell proliferation compared to ectopic expression of GFP mRNA. Cells were treated daily with 0.5 μg/mL of Dox for six days. Each point represents the mean ± SEM, n = 3. Points marked with asterisks are statistically different from each other (two-tailed Student’s t-test, * p < 0.05, ** p < 0.01).
(C) (Left) Dox-induced ectopic expression of lnc152 inhibits the migration of MDA-MB-231 cells compared to ectopic expression of GFP mRNA. (Right) Quantification of the results from the experiments shown in the left panel. Each bar represents the mean ± SEM, n = 8. Bars marked with asterisks are statistically different from each other (two-tailed Student’s t test, **** p < 0.0001).
(D) (Left) Dox-induced ectopic expression of lnc152 inhibits the invasion of MDA-MB-231 cells compared to ectopic expression of GFP mRNA. (Right) Quantification of the results from the experiments shown in the left panel. Each bar represents the mean ± SEM, n = 8. Bars marked with asterisks are statistically different from each other (two-tailed Student’s t test, **** p < 0.0001).
(E) Dox-induced ectopic expression of lnc152 prevents human umbilical vein endothelial cell (HUVEC) tube formation on Matrigel. Morphological appearance of HUVEC grown on Matrigel with conditioned medium (CM) collected from MDA-MB-231 cells expressing Dox-induced GFP or lnc152, stained with Calcein-AM (green) and detected by fluorescence microscopy.
(F) Quantification of the results from the experiments shown in panel E. Image J software with the Angiogenesis plugin was used to detect the number of nodes, number of segments, segment length, and branching length. Each bar represents the mean + SEM, n = 4. Bars marked with asterisks are statistically different from each other (two-tailed Student’s t test, * p < 0.05, ** p < 0.01, *** p < 0.001).
Importantly, conditioned medium (CM) collected from MDA-MB-231 cells with Dox-induced ectopic expression of lnc152, but not GFP, prevented tube formation by human umbilical vein endothelial cells (HUVECs) on Matrigel (Figure 2, E and F), an in vitro assay of angiogenesis (48). These results show that lnc152 can inhibit cancer-related cellular phenotypes in triple-negative breast cancer cells. This includes inhibition of angiogenic phenotypes in HUVECs by TNBC cells whose proliferation is inhibited by lnc152 expression (e.g., MDA-MB-231 cells, not HCC1143 cells) through production of a putative soluble factor.
Lnc152 acts as a tumor suppressor in TNBC cells
To investigate the inhibition of cancer-related phenotypes by lnc152 in vivo, we monitored the growth and biology of xenograft tumors formed in nude mice from MDA-MB-231 cells engineered for Dox-inducible ectopic expression of lnc152. We observed that ectopic expression of lnc152 caused a significant reduction in the growth of the xenograft tumors compared to GFP-expressing tumors (Figure 3A–C). Moreover, ectopic expression of lnc152 caused a significant reduction in blood vessel development, as determined by positive staining for platelet endothelial cell adhesion molecule (PECAM-1), in the xenograft tumors (Figure 3D–F).
Figure 3. Ectopic expression of lnc152 reduces tumor growth, inhibits angiogenesis, and alters gene expression in xenograft tumors in vivo.
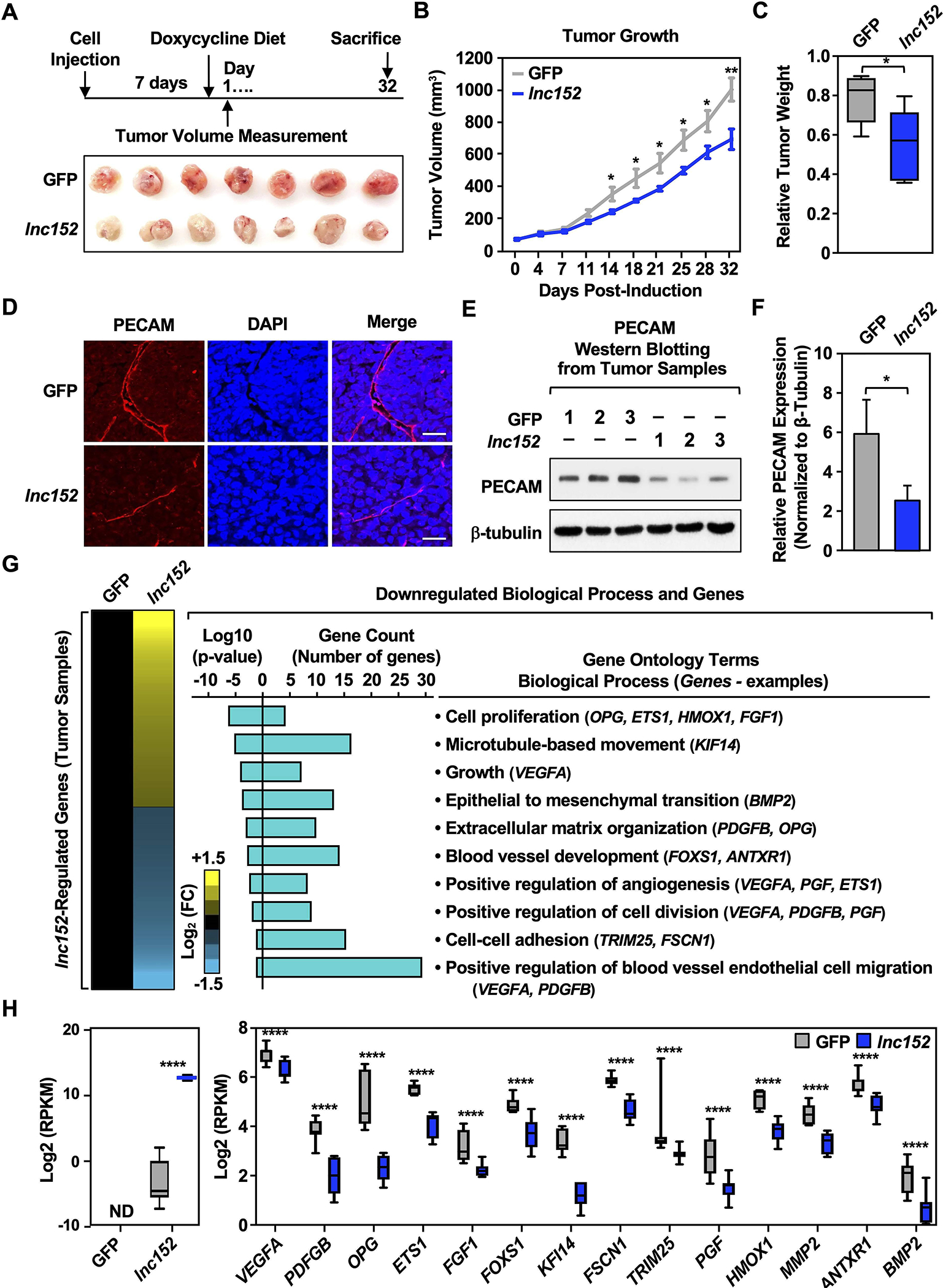
(A) Ectopic expression of lnc152 inhibits the growth of MDA-MB-231 xenograft tumors. (Top) Schematic representation of the experimental timeline for the xenograft tumor model. (Bottom) Images of xenograft tumors formed from MDA-MB-231 cells expressing Dox-induced GFP or lnc152.
(B) Growth curves for MDA-MB-231 xenograft tumors engineered for ectopic expression of GFP or lnc152. Each point represents the mean ± SEM, n = 7. Points marked with asterisks are statistically different from each other (two-tailed Student’s t-test, * p < 0.05, ** p < 0.01).
(C) Box plots showing tumor weights at the end of the experiment. Points marked with asterisks are statistically different from each other (two-tailed Student’s t test, *p < 0.05).
(D-F) Ectopic expression of lnc152 inhibits blood vessel development in MDA-MB-231 xenograft tumors. (D) Representative immunohistochemical staining of PECAM from MDA-MB-231 xenograft tumor tissue from panel A. (E) Three tumor tissues from each group were analyzed by Western blotting for PECAM and β-tubulin. (F) Quantification of protein levels from the experiments shown in panel E. Bars marked with asterisks are statistically different from each other (two-tailed Student’s t test, * p < 0.05).
(G) Gene expression and gene ontology analyses in MDA-MB-231 xenograft tumors. (Left) Heatmap showing the results of the RNA-seq assays from MDA-MB-231 tumors engineered for ectopic expression of GFP or lnc15. (Right) Gene ontology analysis showing terms and genes de-enriched upon ectopic lnc152 expression in MDA-MB-231 xenograft tumors relative to ectopic expression of GFP. Genes of interest are listed in each GO term.
(H) Bar graphs showing the relative expression of a set of genes upon ectopic expression of lnc152 from MDA-MB-231 xenograft tumor tissue (RNA-seq RPKM values, two-tailed Student’s t test, **** p < 0.001).
In RNA-seq assays, ectopic expression of lnc152 altered the expression of hundreds of genes in the MDA-MB-231 xenograft tumors compared to ectopic expression of GFP (Figure 3, G and H; Supplementary Tables S1 and S2). Interestingly, ectopic expression of lnc152 caused downregulation of genes involved in biological processes such as cell proliferation, extracellular matrix organization, and blood vessel development (Figure 3G). The global effects of lnc152 on the downregulation of gene expression were reflected in the expression of individual genes controlling relevant biological processes (Figure 3H; Supplementary Table S1). These effects were confirmed by RT-qPCR from the same xenograft tumor tissues (Figure S5A), as well as in MDA-MB-231 cells (Figure S5B) and HCC1143 cells (Figure S5C). A set of genes were also upregulated upon ectopic expression of lnc152 compared to GFP in the MDA-MB-231 xenografts (Figure S6; Supplementary Table S2). Together, these results suggest that lnc152 acts as a tumor suppressor in TNBC. Ectopic expression of lnc152 reduces tumor growth, inhibits angiogenesis, and modulates the expression of genes that are involved in cell proliferation, blood vessel development, and positive regulation of angiogenesis in MDA-MB-231 xenograft tumors in vivo.
Lnc152 inhibits the metastasis of TNBC cells
The effects of lnc152 on biological processes, such as cell proliferation, extracellular matrix organization, and blood vessel development, suggested a potential role for lnc152 in metastasis. To test this possibility, we generated MDA-MB-231 cell lines stably expressing luciferase (Luc) with Dox-inducible ectopic expression of GFP or lnc152 (Figure S7A) and used them in an intracardiac injection model of cancer metastasis (42) (Figure 4A). These Luc-expressing cell lines exhibited similar lnc152-dependent outcomes as the cell-based models shown above, such as cell proliferation (Figure S7B), effects on the expression of selected genes (Figure S7C), and tube formation in HUVECs on Matrigel (Figure S7, D and E).
Figure 4. Ectopic expression of lnc152 inhibits tumor metastasis in vivo.
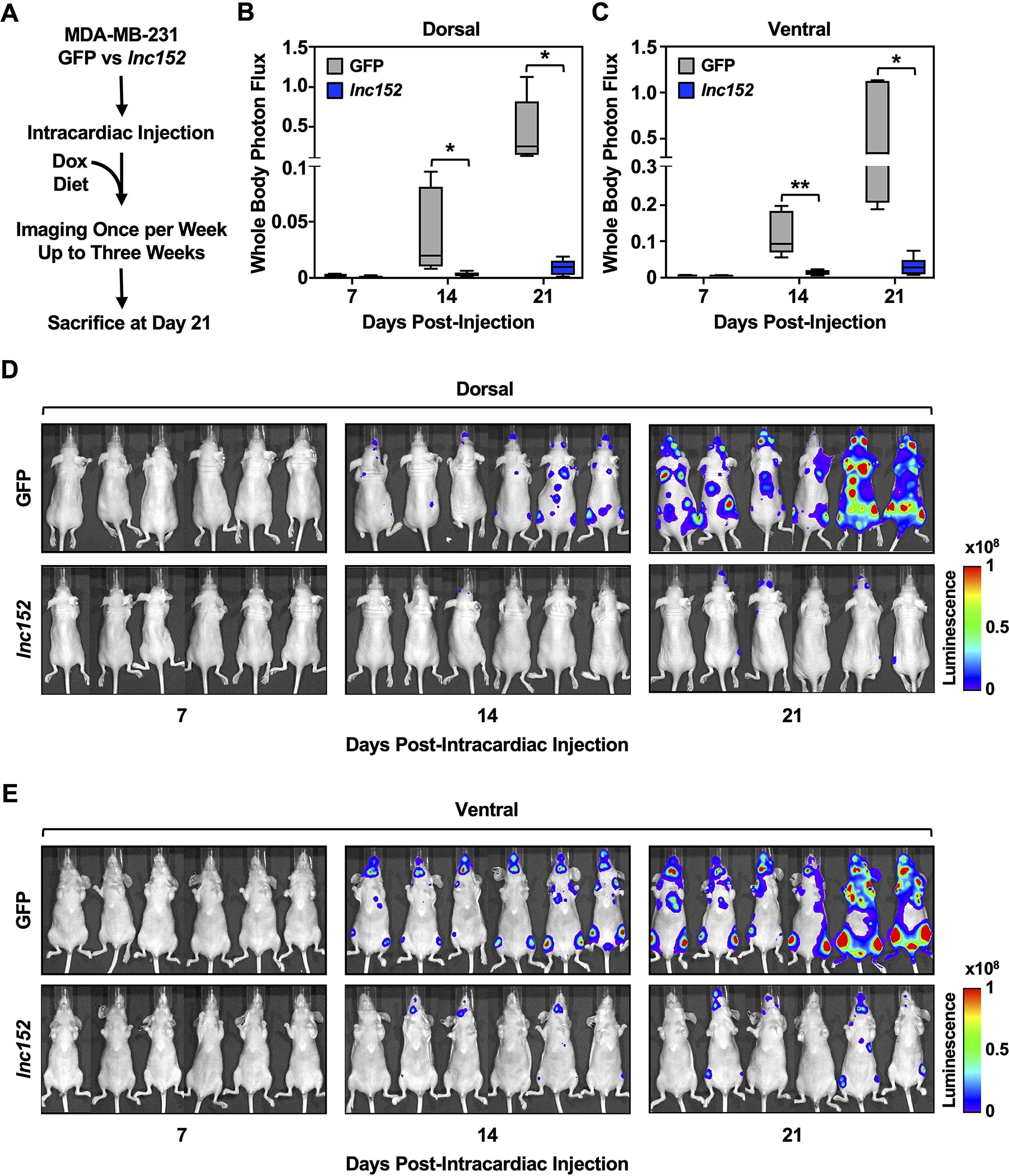
(A) Schematic representation of the experimental design for the intracardiac injection model of cancer metastasis in vivo.
(B-C) Ectopic expression of lnc152 reduces the metastatic burden in an MDA-MB-231 cell metastasis mouse model. Box plots of bioluminescence signals from the dorsal (B) and ventral (C) views of mice at various times after intracardiac injection of tumor cells expressing GFP or lnc152. Each bar represents the mean ± SEM, n = 6. Bars marked with asterisks are statistically different from each other (two-tailed Student’s t-test, * p < 0.05, ** p < 0.01).
(D-E) Bioluminescence tracing of the dorsal (D) and ventral (E) views from each group at various times after tumor cell intracardiac injection shows exponential increases in luciferase activity in mice expressing GFP compared to lnc152.
We used the Luc-expressing cell lines to examine the potential effects of lnc152 on the metastasis of MDA-MB-231 cells. In this model, ectopic expression of lnc152 inhibited metastasis of MDA-MB-231 cells to the brain, lung, and bones (Figure 4, B and C). Images of the bioluminescence signals from the dorsal (Figure 4D) and ventral (Figure 4E) views of the mice at various times after intracardiac injection of tumor cells shows a dramatic reduction in metastatic burden for cells expressing lnc152 compared to GFP. In control experiments, dorsal bioluminescence signals from each group immediately after intracardiac injection show no significant differences in Luc activity in mice expressing lnc152 compared to GFP (Figure S7, F–H). Together, these results demonstrate that expression of lnc152 significantly modulates the metastatic behavior of TNBC cells.
The RNA-binding tumor suppressor RBM47 interacts with lnc152 and mediates similar functions
Interacting proteins are important effectors for lncRNA function. In turn, interactions with lncRNAs can modulate the stability and enzymatic activity of their interacting proteins. Thus, lncRNA-protein interactions can play a crucial role in fundamental cellular processes, including regulation of gene expression. To better understand the mechanism of action of lnc152, we sought to identify its interacting proteins in cells. In this regard, we used in vitro RNA-pull down assays followed by LC-MS/MS analysis to determine the repertoire of lnc152-interacting proteins in MCF-7 breast cancer cells in which lnc152 is naturally highly expressed. Proteomic analysis of the lnc152 interactome identified a set of lnc152-interacting proteins (Figure 5, A–C; Figure S8, A–C; Supplementary Table S3), including RNA binding motif protein 47 (RBM47), which was one of the most highly enriched proteins in the lnc152 interactome (Figure 5D). RBM47 is a known tumor suppressor and emerging evidence has highlighted the role of RBM47 in inhibition of breast cancer progression and metastasis (49,50). Given the evidence that lnc152 can also modulate migration, invasion, and metastasis, we examined the nature of the lnc152-RBM47 interaction in more detail. In vitro RNA-pull down and RNA immunoprecipitation assays confirmed that lnc152 interacts directly with RBM47 (Figure 5, E–G; Figure S9).
Figure 5. Proteomic analysis of the lnc152 interactome reveals an association between lnc152 and RBM47.
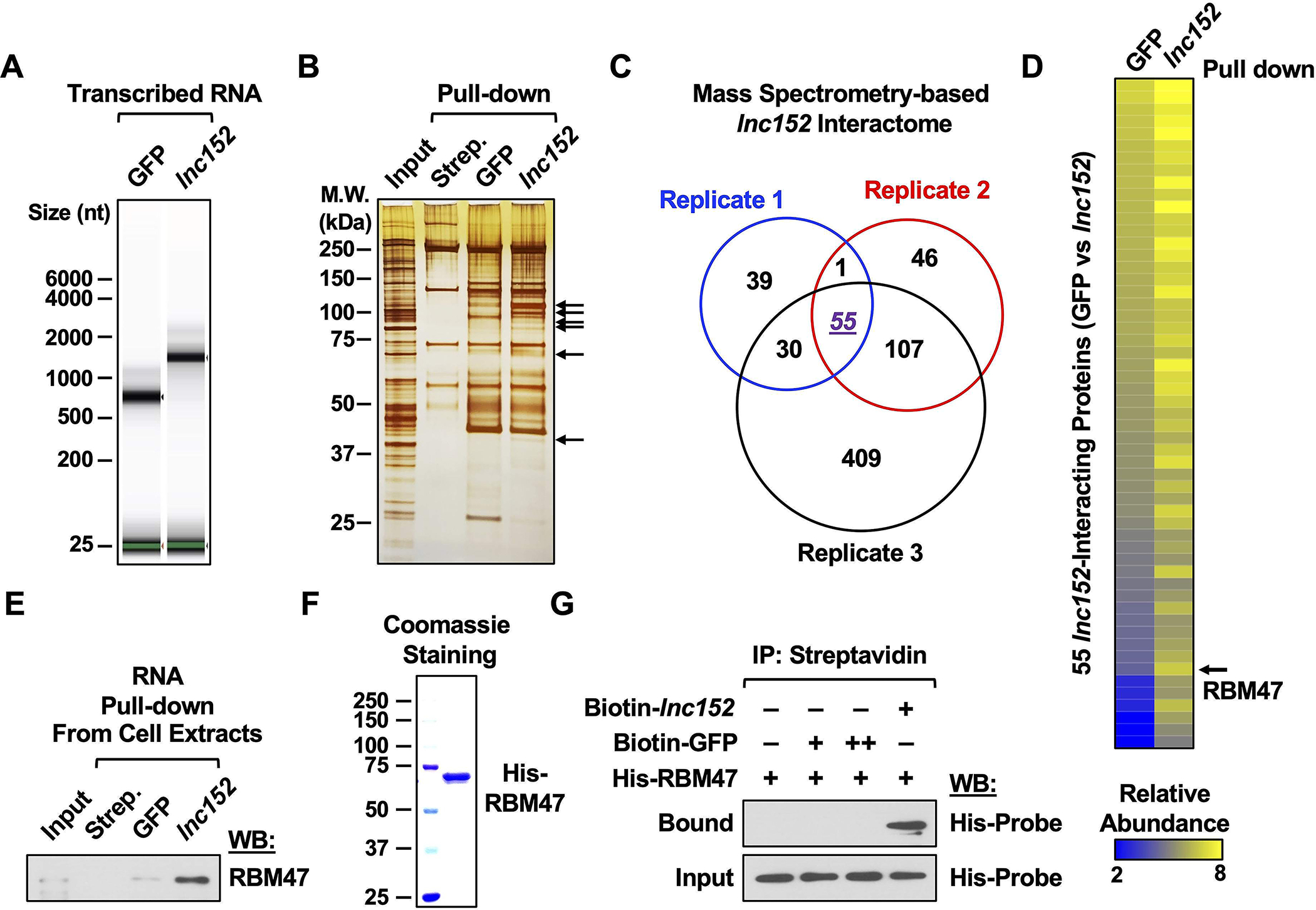
(A) In vitro transcribed and biotinylated GFP or lnc152 RNAs were visualized using an RNA BioAnalyzer.
(B) Silver stained SDS-PAGE gel of proteins pulled down by GFP or lnc152 RNAs from MCF-7 cell extracts. The arrows indicate significantly enriched proteins in the lnc152 RNA pulldown compared to the GFP RNA pulldown. Streptavidin (Strep.) pulldown was used as a negative control.
(C) A diagram depicting the lnc152 interactome from three independent RNA pulldown assays.
(D) Heatmap representing 55 lnc152-interacting proteins shared between three independent RNA pulldown assays (highlighted in purple in panel C). RBM47 is one of the most highly enriched proteins in the lnc152 pulldown.
(E) RNA pulldown assay showing a direct interaction between lnc152 and RBM47. In vitro and biotinylated GFP or lnc152 RNAs were incubated with whole cell extracts from MCF-7 cells, immunoprecipitated using streptavidin agarose beads, and then analyzed by Western blotting for RBM47.
(F) Purified recombinant RBM47 protein was analyzed using SDS-PAGE with Coomassie blue staining.
(G) RNA pulldown assay showing a direct interaction between lnc152 and RBM47. In vitro transcribed and biotinylated GFP or lnc152 RNAs were incubated with purified recombinant RBM47 protein, immunoprecipitated using streptavidin agarose beads, and then analyzed by Western blotting for RBM47.
To determine potential biological outcomes of lnc152-RBM47 interactions, we first determined if the expression of RBM47 correlates with the expression of lnc152 across breast cancer subtypes or cell lines. We found that RBM47 is highly expressed in breast cancer tissues (TCGA) compared to normal breast tissues (GTEx; Figure 6A). RBM47 is upregulated in luminal and HER2-enriched tumors, but downregulated in basal-like tumors, from breast cancer patients (TCGA; Figure 6B). In each case, there was a significant positive correlation in lnc152 and RBM47 RNA expression, strengthening the data on the interaction and biological connection of lnc152 and RBM47 (Figure S10, A–C) .Likewise, RBM47 is upregulated in luminal A, but downregulated in basal, breast cancer cell lines (Figure 6C; Figure S11).
Figure 6. Ectopic expression of RBM47 inhibits aggressive cancer-related outcomes in MDA-MB-231 TNBC cells.
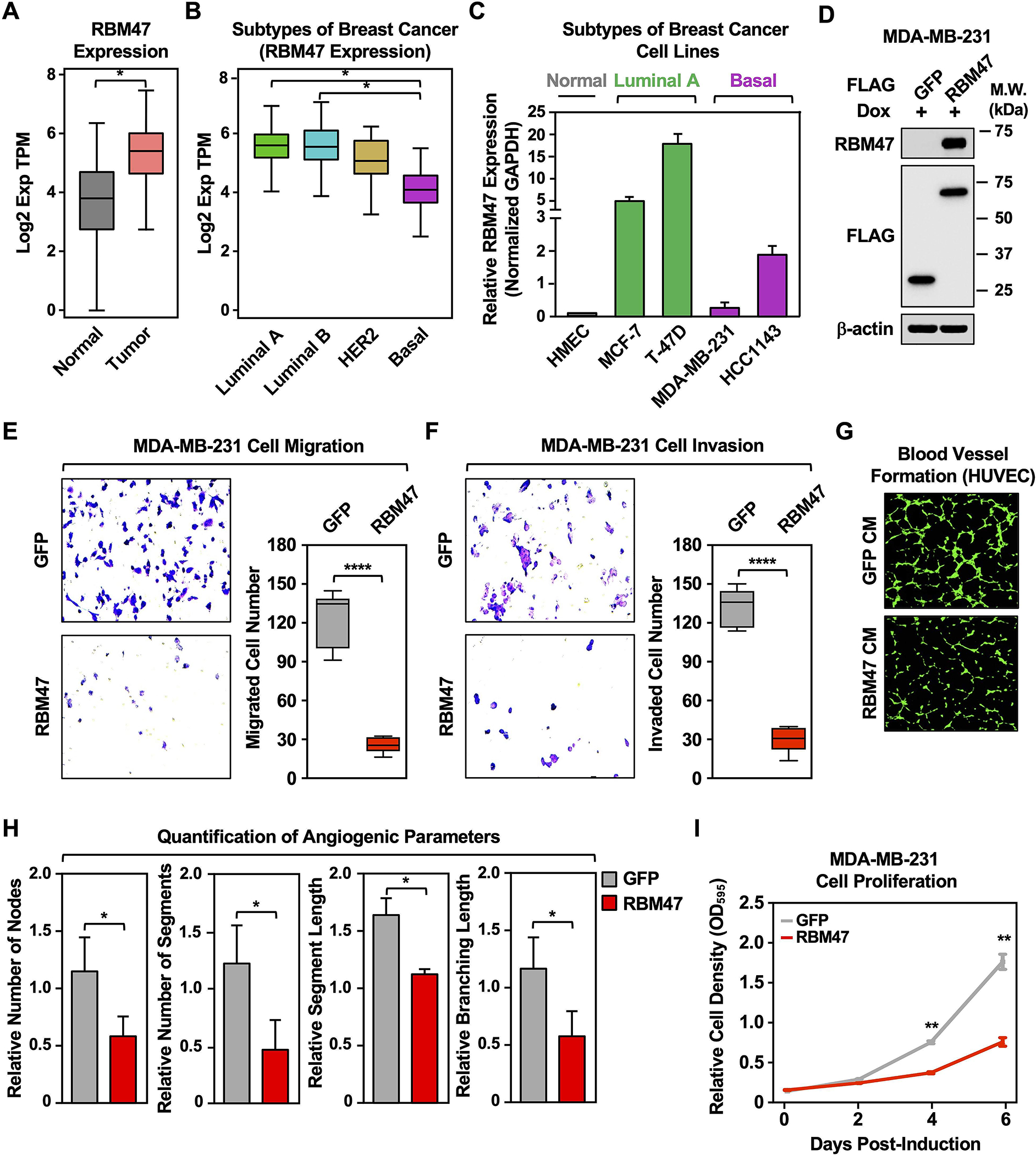
(A) RNA-seq expression data for RBM47 in primary tumors (TCGA breast cancer samples, n = 1085) compared with normal breast tissues (TCGA normal and GTEx data, n = 291). Bars marked with asterisks are statistically different from each other (two-tailed Student’s t test, * p < 0.05).
(B) Box plots comparing the expression of RBM47 in different intrinsic molecular subtypes of breast cancers derived from patient samples [luminal A (n = 415), luminal B (n = 194), HER2-enriched (n = 66), and basal-like (n = 135); TCGA]. Bars marked with asterisks are statistically different from each other (two-tailed Student’s t test, * p < 0.05).
(C) Bar graph showing the expression of RBM47 in breast cancer cell lines representing different molecular subtypes.
(D) Generation of MDA-MB-231 cell lines for Dox-inducible ectopic expression of FLAG-GFP or FLAG-RBM47. Whole cell extracts were analyzed by Western blotting for RBM47, FLAG, and β-actin. Comparable protein expression of GFP and RBM47 levels are shown in the Western blots for FLAG.
(E) (Left) Dox-induced ectopic expression of RBM47 inhibits the migration of MDA-MB-231 cells compared to ectopic expression of GFP mRNA. (Right) Quantification of the results from the experiments shown in the left panel. Each bar represents the mean ± SEM, n = 6. Bars marked with asterisks are statistically different from each other (two-tailed Student’s t test, **** p < 0.0001).
(F) (Left) Dox-induced ectopic expression of RBM47 inhibits the invasion of MDA-MB-231 cells compared to ectopic expression of GFP mRNA. (Right) Quantification of the results from the experiments shown in the left panel. Each bar represents the mean ± SEM, n = 5. Bars marked with asterisks are statistically different from each other (two-tailed Student’s t test, **** p < 0.0001).
(G) Dox-induced ectopic expression of RBM47 prevents human umbilical vein endothelial cell (HUVEC) tube formation on Matrigel. Morphological appearance of HUVECs grown on Matrigel with conditioned medium collected from MDA-MB-231 cells expressing Dox-induced GFP or RBM47, stained with Calcein-AM (green) and detected by fluorescence microscopy.
(H) Quantification of the results from the experiments shown in panel G. Image J software with the Angiogenesis plugin was used to detect the number of nodes, number of segments, segment length, and branching length. Each bar represents the mean + SEM, n = 4. Bars marked with asterisks are statistically different from each other (two-tailed Student’s t test, * p < 0.05).
(I) Dox-induced ectopic expression of RBM47 inhibits MDA-MB-231 cell proliferation more effectively than ectopic expression of GFP. Cells were treated daily with 1 μg/mL of Dox for six days. Each point represents the mean ± SEM, n = 3. Points marked with asterisks are statistically different from each other (two-tailed Student’s t-test, ** p < 0.01).
We also determined if ectopic expression of RBM47 might suppress the aggressive cancer features of TNBC lines, as seen with lnc152 expression. Dox-induced ectopic expression of RBM47 (Figure 6D) inhibited the migration and invasion of MDA-MB-231 cells compared to ectopic expression of GFP (Figure 6, E and F). Moreover, Dox-induced ectopic expression of RBM47 prevented HUVEC tube formation on Matrigel using MDA-MB-231 conditioned medium (Figure 6, G and H). Finally, Dox-induced ectopic expression of RBM47 inhibited MDA-MB-231 cell proliferation compared to ectopic expression of GFP (Figure 6I). Together, these results illustrate similar expression patterns, roles, and functional outcomes for RBM47 and lnc152.
Lnc152 enhances RBM47 expression to drive cancer-related phenotypes
Both lnc152 and RBM47 are downregulated in basal breast cancer cell lines, and ectopic expression of either lnc152 or RBM47 suppressed the aggressive cancer phenotypes of TNBC cells, indicating that they are positively correlated in the regulation of TNBC. Since lncRNAs can promote gene expression and bind to proteins to regulate their stability, we tested the possibility that lnc152 may affect RBM47 mRNA expression or RBM47 protein stability. siRNA-mediated knockdown of lnc152 decreased the levels RBM47 protein in MCF-7 cells (Figure 7A), whereas Dox-induced ectopic expression of lnc152 increased the levels of RBM47 protein in both MDA-MB-231 cells (Figure 7B) and MDA-MB-231 xenograft tumor tissue (Figure 7C). Likewise, siRNA-mediated knockdown of lnc152 decreased the steady-state levels of RBM47 mRNA in MCF-7 cells (Figure 7D), whereas Dox-induced ectopic expression of lnc152 increased the levels of RBM47 mRNA in both MDA-MB-231 cells (Figure 7E) and MDA-MB-231 xenograft tumor tissue (Figure 7F). This regulation is one-directional, as neither knockdown of RBM47 in MCF-7 cells (Figure S12A) or ectopic expression of RBM47 in MDA-MB-231 cells (Figure S12B) had any significant effect on lnc152 RNA levels. We also observed that concomitant knockdown of RBM47 in MDA-MB-231 cells ectopically expressing lnc152 did not rescue the decreased proliferation, indicating that lnc152 mediates its effect through various binding partners, including RBM47 (Figure S13). Furthermore, Dox-induced ectopic expression of lnc152 in MDA-MB-231 xenograft tumor tissue increased the expression of RBM47-upregulated genes, such as CASP7 and HTATIP2, which also act as tumor suppressors (Figure 7G; Figure S14, A and B) and decreased the expression levels of RBM47-downregulated genes, such as SLC29A1, CDK2, and SUPT16H (Figure 7G; Figure S14, A and B). Together, these results indicate that lnc152 enhances RBM47 expression to drive cancer-related phenotypes.
Figure 7. Lnc152 stabilizes RBM47 mRNA and enhances RBM47 protein levels to promote RBM47-mediated gene expression in MDA-MB-231 TNBC cells.
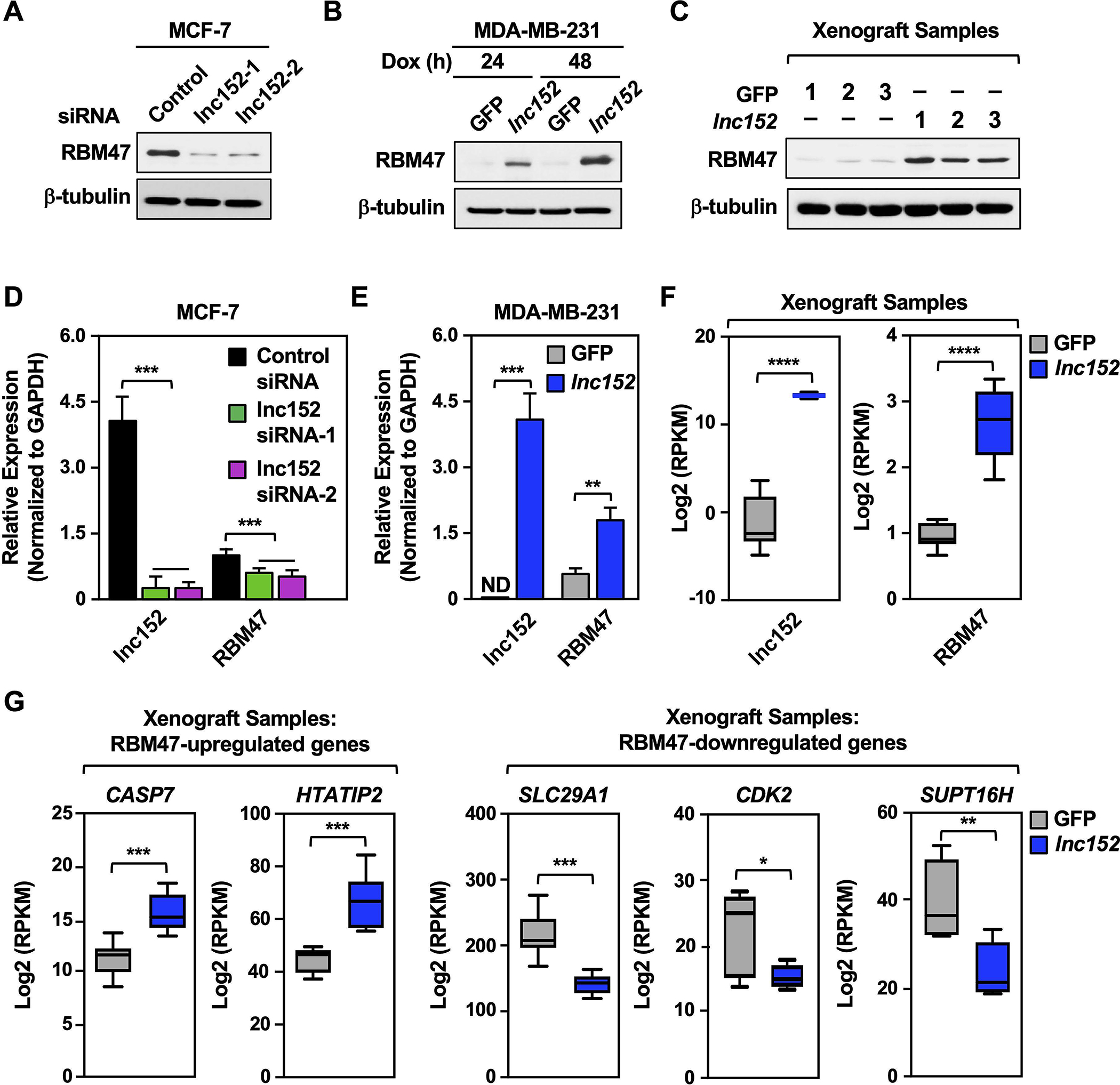
(A) siRNA-mediated knockdown of lnc152 decreases the level of RBM47 in MCF-7 cells. Western blotting for RBM47 and β-tubulin is shown.
(B) Dox-induced ectopic expression of lnc152 increases the level of RBM47 in MDA-MB-231 cells. Western blotting for RBM47 and β-tubulin is shown.
(C) Dox-induced ectopic expression of lnc152 increases the level of RBM47 in MDA-MB-231 xenograft tumor tissue. Western blotting for RBM47 and β-tubulin is shown.
(D) siRNA-mediated knockdown of lnc152 decreases the steady-state levels of RBM47 mRNA in MCF-7 cells. Lnc152 and RBM47 mRNA levels were determined by RT-qPCR and normalized to GAPDH mRNA. Each bar represents the mean + SEM, n = 3. Bars marked with asterisks are statistically different from each other (two-tailed Student’s t test, *** p < 0.001).
(E) Dox-induced ectopic expression of lnc152 increases the steady-state levels of RBM47 mRNA in MDA-MB-231 cells. Lnc152 and RBM47 mRNA levels were determined by RT-qPCR and normalized to GAPDH mRNA. ND indicates that the expression level was not detectable. Each bar represents the mean + SEM, n = 3. Bars marked with asterisks are statistically different from each other (two-tailed Student’s t test, **p < 0.01, *** p < 0.001).
(F) Bar graphs showing the relative expression of lnc152 and RBM47 mRNA upon ectopic expression of lnc152 in MDA-MB-231 xenograft tumor tissues, RNA-seq RPKM values (two-tailed Student’s t test, **** p < 0.0001).
(G) Dox-induced ectopic expression of lnc152 in MDA-MB-231 xenograft tumor tissue alters the expression levels of RBM47-regulated genes. Bar graphs showing the relative expression of RBM47-regulated genes, RNA-seq RPKM values (two-tailed Student’s t test, *p < 0.05, **p < 0.01, *** p < 0.001).
Discussion
In the studies herein, we explored the cancer-related molecular functions of lnc152 (a.k.a. DRAIC). Our results revealed some key features of the biology of lnc152 in breast cancer. We found that lnc152 is highly expressed in luminal breast cancer cells, where it acts to attenuate cell migration and invasion. Likewise, ectopic expression of lnc152 inhibits cancer-related cellular phenotypes in TNBC cells, including angiogenesis and metastasis. Molecularly, lnc152 interacts with RNA-binding tumor suppressor RBM47 and enhances RBM47 expression to mediate its inhibitory functions. Collectively, our results demonstrate that lnc152 is an angiogenesis-inhibiting tumor suppressor that attenuates the aggressive cancer-related phenotypes found in TNBC (Figure S14C).
Lnc152/DRAIC is emerging as an important regulator of cancer phenotypes
In a previous study, we annotated lnc152 and characterized its functions in the control of cell cycle gene expression and proliferation in breast cancer cells (22). We observed effects of lnc152 in proliferation, cell cycle progression, and regulation of the estrogen signaling pathway (22). A contemporaneous study identified lnc152/DRAIC as a tumor suppressor in prostate cancer, which is downregulated as the cells progress from an androgen-dependent to a castration-resistant state (23). Moreover, higher levels of lnc152/DRAIC in prostate cancers were found to be associated with longer disease-free survival (23). Subsequent studies have affirmed a key role for lnc152/DRAIC in the biology of an array of cancers, including breast, prostate, esophageal, and gastric cancers, as well as gliomas (24–30).
A wide range of molecular and cellular regulatory functions have been ascribed to lnc152/DRAIC, including gene regulation, inhibition of NF-κB activation, interference with NFRKB deubiquitylation, sponging miR-432–5p, inhibition of protein translation, and autophagy (24–30). These regulatory functions underlie the tumor suppressor functions of lnc152/DRAIC, as well as its role in regulating a range of cancer-related phenotypes, such as proliferation, migration, invasion, metastasis, and angiogenesis. Since the expression of lnc152/DRAIC varies dramatically in different cancer subtypes derived from the same tissue, it may have value as a biomarker or prognostic indicator.
Functional interactions between lnc152 and RBM47
One mechanism by which lncRNAs exert their regulatory effects is through specific interacting proteins. Using in vitro RNA-pull down assays followed by LC-MS/MS, we identified the RNA-binding tumor suppressor RBM47 as a lnc152-interacting protein. Previous studies have shown that RBM47 suppresses cancer phenotypes, including proliferation, migration, invasion, and metastasis, in breast cancer and non-small-cell lung cancer (NSCLC) by regulating the splicing and stabilization of its target mRNAs (49,50). For example, RBM47 binds to the 3′-UTR of DKK1 mRNA, which encodes DKK1 (Dickkopf Wnt signaling pathway inhibitor 1), to protect it from destabilizing factors, thus increasing DKK1 secretion in TNBC (50). RBM47 also binds the AXIN1 mRNA, which encodes a pro-oncogenic β-catenin signaling pathway regulator, thereby stabilizing it and enhancing suppression of Wnt/β-catenin signaling in NSCLC (49). We speculate that RBM47 uses the same features that allow it to bind to mRNAs to bind to lnc152.
Although we have not yet elucidated the precise molecular underpinnings of the functional interactions between lnc152 and RBM47, we have shown that the effects of lnc152 in TNBC cells are mediated, in part, by regulating the expression of RBM47. Lnc152 elevates the expression of RBM47 mRNA and protein, which impacts a wide range of cancer-related outcomes. Interestingly, lnc152 and RBM47 are both highly expressed in the luminal subtype of breast cancer, and significantly downregulated in the triple-negative subtype, suggesting that lnc152 and RBM47 levels may serve as biomarkers to predict aggressive features of breast cancer.
Lnc152 inhibits angiogenesis
Angiogenesis is an essential component of the metastatic pathway. The process begins with invasion and intravasation into the surrounding lymphatics and blood vessels, and culminates with colonization of the disseminated tumor cells in a distal organ, angiogenesis, and growth (1,2). Our transcriptome analysis of TNBC xenografts indicated that lnc152 downregulates genes controlling angiogenesis. Moreover, we found that ectopic expression of lnc152 inhibits angiogenic phenotypes in HUVECs by TNBC cells whose proliferation is inhibited by lnc152 expression (e.g., MDA-MB-231 cells) likely through the production of a soluble factor. Such an inhibitory effect of lnc152 on angiogenesis could explain the potent inhibition of TNBC cell metastasis that we observed in mice. In this regard, new blood vessels promote tumor metastasis by providing the route of exit for tumor cells from the primary tumor and colony formation at secondary sites (3,4). Moreover, new blood vessels within the tumor are needed to provide sufficient oxygen and nutrients to support tumor growth (3,4).
Collectively, our results identify lncRNA152 as an angiogenesis-inhibiting tumor suppressor that attenuates the aggressive cancer-related phenotypes found in TNBC by upregulating the expression of tumor the suppressor RBM47. As such, lncRNA152 may serve as a biomarker to track aggressiveness of breast cancer, as well as therapeutic target for treating TNBC.
Supplementary Material
Implications.
This study identifies lncRNA152 as an angiogenesis-inhibiting tumor suppressor that attenuates the aggressive cancer-related phenotypes found in TNBC by upregulating the expression of the tumor suppressor RBM47. As such, lncRNA152 may serve as a biomarker to track aggressiveness of breast cancer, as well as therapeutic target for treating TNBC.
Acknowledgements
We thank members of the Kraus lab for intellectual input and critical comments on this manuscript, Anusha Nagari for assistance with analysis of sequencing data, and Dr. Philipp Scherer for cDNA from various human cell lines. We acknowledge and thank the following UT Southwestern core facilities: Live Cell Imaging Core for microscopy (Dr. Katherine Luby-Phelps; NIH S10 OD021684), Next Generation Sequencing Core for deep sequencing (Dr. Ralf Kittler), Proteomics Core Facility for mass spectrometry (Dr. Andrew Lemoff), Histo Pathology Core for histology (Dr. Bret Evers), and the Small Animal Imaging Shared Resource, which is supported in part by the Harold C. Simmons Cancer Center through an NCI Cancer Center Support Grant (P30 CA142543). This work was supported by grants from the Cancer Prevention and Research Institute of Texas (RP130607, RP160318, RP190235) and funds from the Cecil H. and Ida Green Center for Reproductive Biology Sciences Endowment to W.L.K.; a postdoctoral fellowship from the U.S. Department of Defense Breast Cancer Research Program (BCRP) to D.S.K. (BC134066); and a Komen for the Cure Foundation postdoctoral fellowship to S.S.G. (PDF230441).
Funding:
This work was supported by grants from the Cancer Prevention and Research Institute of Texas (RP130607, RP160318, RP190235) and funds from the Cecil H. and Ida Green Center for Reproductive Biology Sciences Endowment to W.L.K.; a postdoctoral fellowship from the U.S. Department of Defense Breast Cancer Research Program (BCRP) to D.S.K. (BC134066); and a Komen for the Cure Foundation postdoctoral fellowship to S.S.G. (PDF230441).
Footnotes
Disclosures: The authors have no competing interests to declare for this work.
Supplementary Information
This paper includes supplementary tables, figures, materials and methods, and references.
References
- 1.Fares J, Fares MY, Khachfe HH, Salhab HA, Fares Y. Molecular principles of metastasis: a hallmark of cancer revisited. Signal Transduct Target Ther 2020;5(1):28 doi 10.1038/s41392-020-0134-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Welch DR, Hurst DR. Defining the hallmarks of metastasis. Cancer Res 2019;79(12):3011–27 doi 10.1158/0008-5472.CAN-19-0458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bielenberg DR, Zetter BR. The contribution of angiogenesis to the process of metastasis. Cancer J 2015;21(4):267–73 doi 10.1097/PPO.0000000000000138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Zuazo-Gaztelu I, Casanovas O. Unraveling the role of angiogenesis in cancer ecosystems. Front Oncol 2018;8:248 doi 10.3389/fonc.2018.00248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Maj E, Papiernik D, Wietrzyk J. Antiangiogenic cancer treatment: The great discovery and greater complexity (Review). Int J Oncol 2016;49(5):1773–84 doi 10.3892/ijo.2016.3709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Oguntade AS, Al-Amodi F, Alrumayh A, Alobaida M, Bwalya M. Anti-angiogenesis in cancer therapeutics: the magic bullet. J Egypt Natl Canc Inst 2021;33(1):15 doi 10.1186/s43046-021-00072-6. [DOI] [PubMed] [Google Scholar]
- 7.Al-Mahmood S, Sapiezynski J, Garbuzenko OB, Minko T. Metastatic and triple-negative breast cancer: challenges and treatment options. Drug Deliv Transl Res 2018;8(5):1483–507 doi 10.1007/s13346-018-0551-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kimbung S, Loman N, Hedenfalk I. Clinical and molecular complexity of breast cancer metastases. Semin Cancer Biol 2015;35:85–95 doi 10.1016/j.semcancer.2015.08.009. [DOI] [PubMed] [Google Scholar]
- 9.Statello L, Guo CJ, Chen LL, Huarte M. Gene regulation by long non-coding RNAs and its biological functions. Nat Rev Mol Cell Biol 2021;22(2):96–118 doi 10.1038/s41580-020-00315-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Sun M, Kraus WL. From discovery to function: the expanding roles of long noncoding RNAs in physiology and disease. Endocr Rev 2015;36(1):25–64 doi 10.1210/er.2014-1034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Bhan A, Soleimani M, Mandal SS. Long noncoding RNA and cancer: a new paradigm. Cancer Res 2017;77(15):3965–81 doi 10.1158/0008-5472.CAN-16-2634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Schmitt AM, Chang HY. Long noncoding RNAs in cancer pathways. Cancer Cell 2016;29(4):452–63 doi 10.1016/j.ccell.2016.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Aznaourova M, Schmerer N, Schmeck B, Schulte LN. Disease-causing mutations and rearrangements in long non-coding RNA gene loci. Front Genet 2020;11:527484 doi 10.3389/fgene.2020.527484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Carlevaro-Fita J, Lanzos A, Feuerbach L, Hong C, Mas-Ponte D, Pedersen JS, et al. Cancer LncRNA Census reveals evidence for deep functional conservation of long noncoding RNAs in tumorigenesis. Commun Biol 2020;3(1):56 doi 10.1038/s42003-019-0741-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Aprile M, Katopodi V, Leucci E, Costa V. LncRNAs in cancer: From garbage to junk. Cancers (Basel) 2020;12(11) doi 10.3390/cancers12113220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kim J, Piao HL, Kim BJ, Yao F, Han Z, Wang Y, et al. Long noncoding RNA MALAT1 suppresses breast cancer metastasis. Nat Genet 2018;50(12):1705–15 doi 10.1038/s41588-018-0252-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Liu Q, Huang J, Zhou N, Zhang Z, Zhang A, Lu Z, et al. LncRNA loc285194 is a p53-regulated tumor suppressor. Nucleic Acids Res 2013;41(9):4976–87 doi 10.1093/nar/gkt182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Shahabi S, Kumaran V, Castillo J, Cong Z, Nandagopal G, Mullen DJ, et al. LINC00261 Is an epigenetically regulated tumor suppressor essential for activation of the DNA damage response. Cancer Res 2019;79(12):3050–62 doi 10.1158/0008-5472.CAN-18-2034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wang C, Tan C, Wen Y, Zhang D, Li G, Chang L, et al. FOXP1-induced lncRNA CLRN1-AS1 acts as a tumor suppressor in pituitary prolactinoma by repressing the autophagy via inactivating Wnt/beta-catenin signaling pathway. Cell Death Dis 2019;10(7):499 doi 10.1038/s41419-019-1694-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hernandez-Romero IA, Guerra-Calderas L, Salgado-Albarran M, Maldonado-Huerta T, Soto-Reyes E. The regulatory roles of non-coding RNAs in angiogenesis and neovascularization from an epigenetic perspective. Front Oncol 2019;9:1091 doi 10.3389/fonc.2019.01091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Yu B, Wang S. Angio-LncRs: LncRNAs that regulate angiogenesis and vascular disease. Theranostics 2018;8(13):3654–75 doi 10.7150/thno.26024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Sun M, Gadad SS, Kim DS, Kraus WL. Discovery, annotation, and functional analysis of long noncoding RNAs controlling cell-cycle gene expression and proliferation in breast cancer cells. Mol Cell 2015;59(4):698–711 doi 10.1016/j.molcel.2015.06.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Sakurai K, Reon BJ, Anaya J, Dutta A. The lncRNA DRAIC/PCAT29 locus constitutes a tumor-suppressive nexus. Mol Cancer Res 2015;13(5):828–38 doi 10.1158/1541-7786.MCR-15-0016-T. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Tiessen I, Abildgaard MH, Lubas M, Gylling HM, Steinhauer C, Pietras EJ, et al. A high-throughput screen identifies the long non-coding RNA DRAIC as a regulator of autophagy. Oncogene 2019;38(26):5127–41 doi 10.1038/s41388-019-0783-9. [DOI] [PubMed] [Google Scholar]
- 25.Li C, Feng SY, Chen L. SET7/9 promotes H3K4me3 at lncRNA DRAIC promoter to modulate growth and metastasis of glioma. Eur Rev Med Pharmacol Sci 2020;24(23):12241–50 doi 10.26355/eurrev_202012_24016. [DOI] [PubMed] [Google Scholar]
- 26.Li F, Zhou X, Chen M, Fan W. Regulatory effect of LncRNA DRAIC/miR-149–5p/NFIB molecular network on autophagy of esophageal cancer cells and its biological behavior. Exp Mol Pathol 2020;116:104491 doi 10.1016/j.yexmp.2020.104491. [DOI] [PubMed] [Google Scholar]
- 27.Saha S, Kiran M, Kuscu C, Chatrath A, Wotton D, Mayo MW, et al. Long noncoding RNA DRAIC inhibits prostate cancer progression by interacting with IKK to inhibit NF-kappaB activation. Cancer Res 2020;80(5):950–63 doi 10.1158/0008-5472.CAN-19-3460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zhang Z, Hu X, Kuang J, Liao J, Yuan Q. LncRNA DRAIC inhibits proliferation and metastasis of gastric cancer cells through interfering with NFRKB deubiquitination mediated by UCHL5. Cell Mol Biol Lett 2020;25:29 doi 10.1186/s11658-020-00221-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Li S, Jia H, Zhang Z, Wu D. DRAIC promotes growth of breast cancer by sponging miR-432–5p to upregulate SLBP. Cancer Gene Ther 2021. doi 10.1038/s41417-021-00388-4. [DOI] [PubMed] [Google Scholar]
- 30.Saha S, Zhang Y, Wilson B, Abounader R, Dutta A. The tumor-suppressive long noncoding RNA DRAIC inhibits protein translation and induces autophagy by activating AMPK. J Cell Sci 2021;134(24) doi 10.1242/jcs.259306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Bos PD, Zhang XH, Nadal C, Shu W, Gomis RR, Nguyen DX, et al. Genes that mediate breast cancer metastasis to the brain. Nature 2009;459(7249):1005–9 doi 10.1038/nature08021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kim DS, Camacho CV, Nagari A, Malladi VS, Challa S, Kraus WL. Activation of PARP-1 by snoRNAs controls ribosome biogenesis and cell growth via the RNA helicase DDX21. Mol Cell 2019;75(6):1270–85 e14 doi 10.1016/j.molcel.2019.06.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Zhong S, Joung JG, Zheng Y, Chen YR, Liu B, Shao Y, et al. High-throughput Illumina strand-specific RNA sequencing library preparation. Cold Spring Harb Protoc 2011;2011(8):940–9 doi 10.1101/pdb.prot5652. [DOI] [PubMed] [Google Scholar]
- 34.Andrews S FASTQC. a Quality Control Tool for High Throughput Sequence Data, version GPL v3 Babraham Bioinformatics 2010. [Google Scholar]
- 35.Kim D, Pertea G, Trapnell C, Pimentel H, Kelley R, Salzberg SL. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol 2013;14(4):R36 doi 10.1186/gb-2013-14-4-r36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Quinlan AR, Hall IM. BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics 2010;26(6):841–2 doi 10.1093/bioinformatics/btq033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Robinson JT, Thorvaldsdottir H, Winckler W, Guttman M, Lander ES, Getz G, et al. Integrative genomics viewer. Nat Biotechnol 2011;29(1):24–6 doi 10.1038/nbt.1754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Trapnell C, Williams BA, Pertea G, Mortazavi A, Kwan G, van Baren MJ, et al. Transcript assembly and quantification by RNA-seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat Biotechnol 2010;28(5):511–5 doi 10.1038/nbt.1621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Dennis G Jr., Sherman BT, Hosack DA, Yang J, Gao W, Lane HC, et al. DAVID: Database for annotation, visualization, and integrated discovery. Genome Biol 2003;4(5):P3. [PubMed] [Google Scholar]
- 40.Tang Z, Li C, Kang B, Gao G, Li C, Zhang Z. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res 2017;45(W1):W98–W102 doi 10.1093/nar/gkx247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Franco HL, Nagari A, Kraus WL. TNFalpha signaling exposes latent estrogen receptor binding sites to alter the breast cancer cell transcriptome. Mol Cell 2015;58(1):21–34 doi 10.1016/j.molcel.2015.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Malladi S, Macalinao DG, Jin X, He L, Basnet H, Zou Y, et al. Metastatic latency and immune evasion through autocrine inhibition of WNT. Cell 2016;165(1):45–60 doi 10.1016/j.cell.2016.02.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Badve S, Turbin D, Thorat MA, Morimiya A, Nielsen TO, Perou CM, et al. FOXA1 expression in breast cancer--correlation with luminal subtype A and survival. Clin Cancer Res 2007;13(15 Pt 1):4415–21 doi 10.1158/1078-0432.CCR-07-0122. [DOI] [PubMed] [Google Scholar]
- 44.Franco HL, Nagari A, Malladi VS, Li W, Xi Y, Richardson D, et al. Enhancer transcription reveals subtype-specific gene expression programs controlling breast cancer pathogenesis. Genome Res 2018;28(2):159–70 doi 10.1101/gr.226019.117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Mehta RJ, Jain RK, Leung S, Choo J, Nielsen T, Huntsman D, et al. FOXA1 is an independent prognostic marker for ER-positive breast cancer. Breast Cancer Res Treat 2012;131(3):881–90 doi 10.1007/s10549-011-1482-6. [DOI] [PubMed] [Google Scholar]
- 46.Perou CM, Sorlie T, Eisen MB, van de Rijn M, Jeffrey SS, Rees CA, et al. Molecular portraits of human breast tumours. Nature 2000;406(6797):747–52 doi 10.1038/35021093. [DOI] [PubMed] [Google Scholar]
- 47.Dai X, Cheng H, Bai Z, Li J. Breast cancer cell line classification and its relevance with breast tumor subtyping. J Cancer 2017;8(16):3131–41 doi 10.7150/jca.18457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Arnaoutova I, Kleinman HK. In vitro angiogenesis: endothelial cell tube formation on gelled basement membrane extract. Nat Protoc 2010;5(4):628–35 doi 10.1038/nprot.2010.6. [DOI] [PubMed] [Google Scholar]
- 49.Shen DJ, Jiang YH, Li JQ, Xu LW, Tao KY. The RNA-binding protein RBM47 inhibits non-small cell lung carcinoma metastasis through modulation of AXIN1 mRNA stability and Wnt/beta-catentin signaling. Surg Oncol 2020;34:31–9 doi 10.1016/j.suronc.2020.02.011. [DOI] [PubMed] [Google Scholar]
- 50.Vanharanta S, Marney CB, Shu W, Valiente M, Zou Y, Mele A, et al. Loss of the multifunctional RNA-binding protein RBM47 as a source of selectable metastatic traits in breast cancer. Elife 2014;3 doi 10.7554/eLife.02734. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Genomic datasets.
The new RNA-seq data generated for this study can be accessed from the NCBI’s Gene Expression Omnibus (GEO) repository (http://www.ncbi.nlm.nih.gov/geo/) using the superseries accession number GSE193634.
Custom scripts.
Custom R scripts for genomic data analyses are available from the Lead Contact on request.
Mass spectrometry datasets.
The new mass spectrometry datasets generated for this study can be accessed from MassIVE and also are available as supplemental data provided with this manuscript.