

Abstract
Background and Aims
Acute liver failure (ALF) is associated with high mortality. Gasdermin D (GSDMD) is the executioner of pyroptosis and is involved in the pathophysiology of immune dysregulation This study investigated the role of the GSDMD inhibitor necrosulfonamide (NSA) in ALF.
Methods
An ALF model was established by lipopolysaccharide/D-galactosamine challenge in C57BL/6J mice. Mice were divided into four groups: normal controls (control group), ALF group (ALF group), dimethyl sulfoxide group (DMSO group), and NSA intervention group (NSA group). Survival was monitored, liver damage was determined by hematoxylin and eosin staining, and serum alanine aminotransferase (ALT). Underlying mechanisms were explored by quantitative real-time PCR, western blotting, and enzyme-linked immunosorbent assays.
Results
Pyroptosis was activated in ALF model mice. Mice treated with GSDMD inhibitor NSA developed less severe liver failure. NSA reduced the expression of GSDMD, NLRP3, cleaved caspase-1, cleaved caspase-11, and secretion of interleukin-1 beta in ALF mice model.
Conclusions
Pyroptosis was activated in ALF. NSA alleviated ALF via the pyroptosis pathway.
Keywords: Liver failure, Acute, Pyroptosis, GSDMD, GSDMD-NT, Necrosulfonamide
Introduction
Acute liver failure (ALF) is a severe liver injury,1 there is a lack of effective treatment, and the condition has extremely high short-term mortality.2 Twenty-one-day mortality of ALF patients is as high as 50%.3 In-depth characterization of the pathogenesis of ALF and early intervention are of great significance for improving prognosis.
The core events in ALF are massive death of liver cells, uncontrollable intrahepatic inflammation, and systemic inflammation.4 Release of a large number of pro-inflammatory cytokines in ALF leads to systemic inflammatory response syndrome.5 Pyroptosis is a newly discovered type of pro-inflammatory programmed cell death.6–9 The canonical pathway of pyroptosis is mediated by inflammasomes, including NLRP3.10,11 Activation of caspase induces release of pro-inflammatory factors such as interleukin-1 beta (IL-1β) and interleukin-18 (IL-18), which leads to breakdown of nuclear DNA in target cells and the formation of pores in the cell membrane.12–14 The non-canonical pathway is mediated by caspase-11, which does not require the participation of inflammasomes.15,16
Recent research has revealed the involvement of the canonical pathway of pyroptosis in ALF.17–19 Studies have found that activation of NLRP3 inflammasomes causes hepatocyte pyroptosis, liver inflammation, fibrosis,20 and injury associated with ischemia-reperfusion,21 heat-shock,22 and sepsis.23 Studies conducted in mouse models of ALF have also shown activation of NLRP3 inflammasomes, induced release of pro-inflammatory factors and aggravated liver failure.17 However, most of the studies only focused on the canonical pathway. GSDMD protein is executioner of pyroptosis in both the canonical and non-canonical pathways; inhibition of GSDMD is thus, another potential therapeutic target for ALF. This study investigated the activation of canonical and non-canonical pyroptosis pathways and explored the role of GSDMD inhibitor necrosulfonamide (NSA) in a mouse model of ALF.
Methods
Animals
Seventy male 8-week-old specific pathogen-free (SPF)-grade C57BL/6J mice weighing 20–25 g were purchased from Shanghai SLAC Laboratory Animal Co. Ltd. The mice were raised at the Experimental Animal Center of Fujian Medical University in a controlled environment at 23–25°C, SPF-level environment, 40–60% humidity, and five mice per cage. Before the procedures, mice were adaptively reared in a 12-h light/dark cycle for 1 week, with ad libitum access to food and water. All procedures involving were conducted in accord with the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health, and were approved by the Animal Welfare & Ethics Committee of the Fujian Medical University (no. FJMU IACUC2019-0050).
Mice were randomly assigned to four groups using a random number table: (1) A normal control group (n=10) with intraperitoneal injection of the same volume of saline as the ALF model modeling reagent; (2) An ALF group (n=20) with intraperitoneal injection of LPS (Sigma-Aldrich, Saint Louis, Missouri, USA ) 50 µg/kg + D-GalN 800 mg/kg (Macklin, Shanghai, China) to establish an ALF model24,25; (3) A dimethyl sulfoxide (DMSO) intervention group (n=20) with intraperitoneal injection of the same amount of DMSO (Sigma-Aldrich) as the NSA group prior to establishment of the model; and (4) an NSA intervention group (n=20) with NSA (EMD Millipore, Darmstadt, Germany ) dissolved in DMSO. NSA 20 mg/kg was administered by intraperitoneal injection 30 m before the establishment of the animal model.26 A diagram the study design is shown in Supplementary Figure 1.
Survival
Survival was observed for 24 h.27,28 Except for the control group, 10 animals in each group were randomly selected for observation of survival after modeling once every 30 m.
Serum enzyme-linked immunosorbent (ELISA) and ALT assays
Except for those observed to determine the survival rate, mice in each group were sacrificed 6 h after modeling. The eyes were removed to collect blood, which was placed in a 1.5 mL centrifuge tube, centrifuged at 4,000 rpm for 15 m, and the supernatant was removed. Serum levels of IL-1β and IL-18 were determined with ELISA kits (mlbio, Shanghai, China). Serum ALT was assayed with a FUJI NX500iVC automatic dry biochemical analyzer (FUJIFILM, Shanghai, China).
Histological analysis
After the mice were sacrificed, the liver was quickly removed and fixed in 10% formaldehyde solution for pathological sections. After paraffin embedding, hematoxylin-eosin staining (HE) was performed. Liver inflammation was evaluated by light microscopy (DM2500; Leica, Buffalo Grove, IL, USA). Two pathologists assessed liver tissue injury in a double-blind manner according to the following four parameters: structural destruction of liver lobules, hepatocyte necrosis, hepatic sinusoidal hemorrhage, and inflammatory cell infiltration. Five different randomly selected fields (400× magnification) per mouse were used for scoring. The scoring criteria were: 1=0–25%; 2=25–50%; 3=50–75%; 4=75–100%. The liver injury score was the sum of the scores for the above four parameters.21,29 The liver injury score ranged from 4 to 16.
Quantitative real-time polymerase chain reaction (qRT-PCR)
Quick-RNA MicroPrep (ZYMO, Irvine, CA, U.S.A) was used to extract total RNA. Reverse transcription was performed following the instructions of Takara PrimeScript. RT Reagent Kit with gDNA Eraser (Perfect Real Time) (Takara, Beijing, China). qRT-PCR was performed with a LightCycler 96 qRT-PCR instrument (Roche, Mannheim, Germany) following the instructions included with the TB Green Premix Ex Taq™ (Tli RNase H Plus) (Takara) reagent. The qRT-PCR primers are shown in Table 1.
Table 1. qRT-PCR primer sequences.
| Gene | Primer | Sequence (5′–3′) |
|---|---|---|
| Nlrp3 | Forward | CCCTTGGAGACACAGGACTC |
| Reverse | GAGGCTGCAGTTGTCTAATTCC | |
| Casp1 | Forward | ACCCTCAAGTTTTGCCCTTT |
| Reverse | GATCCTCCAGCAGCAACTTC | |
| Gsdmd | Forward | CCATCGGCCTTTGAGAAAGTG |
| Reverse | ACACATGAATAACGGGGTTTCC | |
| Casp11 | Forward | ACAAACACCCTGACAAACCAC |
| Reverse | CACTGAGTTCAGACTTGTTAAA | |
| Il-1β | Forward | GCCACCTTTTGACAGTGATGAG |
| Reverse | CCTGAAGCTCTTGTTGATGTGC | |
| Il-18 | Forward | TCTGACATGGCAGCCATTGT |
| Reverse | CAGGCCTGACATCTTCTGCAA | |
| β-actin | Forward | GAGACCTTCAACACCCCAGC |
| Reverse | ATGTCACGCACGATTTCCC |
Western blotting
Protein concentration was determined with a bicinchoninic acid (BCA) protein quantification kit (ThermoFisher Scientific, Rockford, IL, USA) following to the manufacturer’s instructions. After preparation of the gels, the sample was slowly added to the lane. After electrophoresis, the proteins were transferred and blocked. After blocking, the polyvinylidene difluoride (PVDF) membranes were incubated overnight with the primary antibodies anti-caspase-11 (ab180673; Abcam, Cambridge, UK), anti-GSDMD (AF4012; Affinity, Jiangsu, China), anti-caspase-1 (ab1872; Abcam), anti-NLRP3 (ab214185; Abcam), or anti-IL-1β (ab9722; Abcam). Subsequently, the membranes were incubated with the secondary antibody. Then, the membranes were exposed with a gel imager (Gel Doc2000; Bio-Rad, Hercules, CA, USA).
Statistical analysis
GraphPad Prism 8.0.1 (http://www.graphpad.com/) was used for statistical analysis. Data were reported as means±standard deviations. Between-group differences were assessed with t-tests or Mann-Whitney tests. Comparisons of three or more groups were performed using one-way analysis of variance or non-parametric tests. Pairwise comparisons were tested by the Sidak test, Tamhane’s T2 test, or Dunn’s test. Survival analysis was performed by the Kaplan-Meier method, and between-group differences in survival were assessed using log-rank tests. Two-sided p-values <0.05 were considered significant.
Results
Effect of NSA on ALF survival
Survival curves are shown in Figure 1A. The first death in the ALF group occurred 6.5 h after modeling. Most deaths occurred at 7 h, and all animals in the ALF group died within 8 h. The survival rate in the DMSO group was similar to that in the ALF group. The first death in the NSA group occurred at 7.5 h, which was longer than that in the ALF and DMSO groups. The 24 h survival rate in the NSA group was 60%, which was significantly better than that in the ALF group (p=0.0001).
Fig. 1. (A) Survival and serum, (B) ALT, (C) IL-1β, and (D) IL-18.
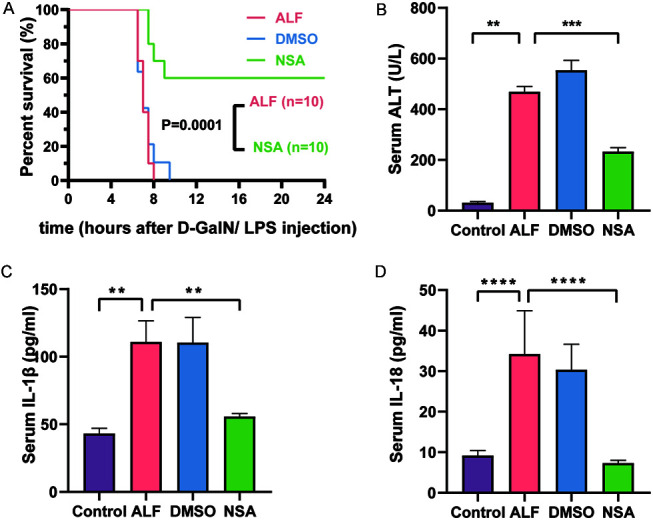
Data are means±standard deviations (ALT, n=3; IL-1β and IL-18, n=5). **p<0.01, ***p<0.001, ****p<0.0001. ALF, acute liver failure; ALT, alanine aminotransferase; D-GalN, D-galactosamine; DMSO, dimethyl sulfoxide; IL-1β, interleukin 1 beta; IL-18, interleukin 18; LPS, lipopolysaccharide; NSA, necrosulfonamide intervention group.
NSA alleviated liver injury in ALF mice
Six hours after establishing the ALF mice model, the mean serum ALT levels in the ALF, DMSO, and NSA groups were 468.3 U/L, 552.7 U/L, and 232 U/L, respectively. The ALT level in the NSA group was significantly lower than that in the ALF group (p<0.005; Fig. 1B). The gross pathology results, including HE staining and liver injury scores are shown in Figure 2 and Supplementary Figure 2. There were no signs of liver injury in the control group (Fig. 2). On gross examination, livers of the ALF and DMSO groups were dark red, relatively swollen, and had scattered bleeding points on the surface. Histopathologically, massive necrosis and hepatic sinusoidal hemorrhage were found in both the ALF and DMSO groups. Both ALF and DMSO groups showed predominant inflammatory cell infiltration. The NSA intervention significantly reduced the liver damage, scores that were significantly lower than those in the ALF group (p<0.005; Supplementary Fig. 2).
Fig. 2. Effects of NSA on liver tissue pathology in different groups.

Triangles (▲) indicate hepatic structure collapse; asterisks (★) indicate hepatocyte edema and necrosis; arrows (→) indicate hepatic sinusoid hemorrhage and inflammation. ALF, acute liver failure group; Control, normal group; DMSO, dimethyl sulfoxide group; NSA, necrosulfonamide intervention group.
NSA reduced the expression of the inflammatory cytokines in ALF mice
The ELISA results are shown in Figure 1C, D. Serum IL-1β and IL-18 levels in the ALF group were higher than those in the control group (p<0.01 and p<0.001, respectively). Serum of IL-1β and IL-18 levels in the DMSO group were not significantly different from those in the ALF group. Serum IL-1β and IL-18 levels in the NSA group were significantly lower than those in the ALF group (p<0.01 and p<0.001, respectively).
Pyroptosis was activated during ALF
The transcription levels of Casp1, Casp11, and Gsdmd are shown in Figure 3. Casp1 and Casp11 mRNA expression was significantly higher in the ALF and DMSO groups than in the control group (Casp1, p<0.05 and Casp11, p<0.01). Gsdmd transcription was inhibited in the ALF group (p<0.05). The expression of proteins involved in the pyroptosis pathway, NLRP3, cleaved caspase-11, cleaved caspase-1, and IL-1β are shown in Figures 4 and 5. All were upregulated in the ALF group (all p<0.05). The executors of the pyroptosis pathway, GSDMD, and GSDMD-NT, were also significantly upregulated in the ALF group (both p<0.0001).
Fig. 3. Effects of NSA on mRNA levels of key molecules in the pyroptosis pathway.
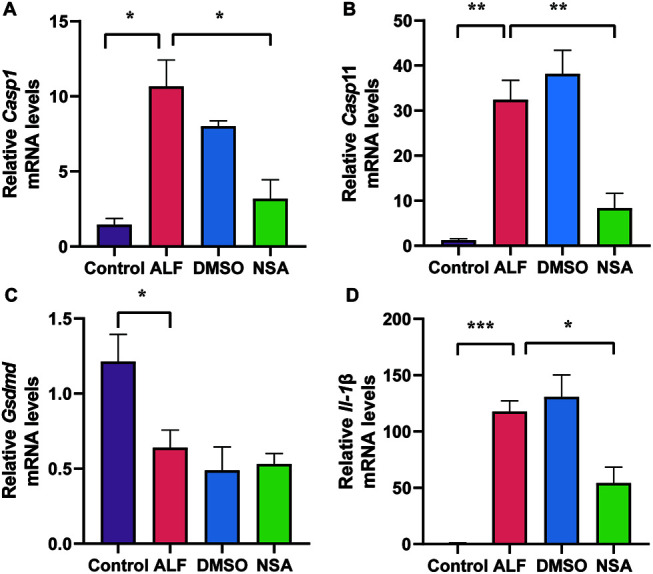
Expression of (A) Casp1 mRNA; (B) Casp11 mRNA. (NSA and DMSO, n=3; Control and ALF, n=4; (C) Gsdmd mRNA, n=4; (D) Il-1β mRNA. (NSA and DMSO, n=3; ALF, n=4; and control, n=5). Data are means±standard deviation. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001. ALF, acute liver failure; Control, normal; DMSO, dimethyl sulfoxide; NSA, necrosulfonamide.
Fig. 4. Effects of NSA on key molecules in the pyroptosis pathway.

(A) Western blotting of NLRP3, pro caspase-11, cleaved caspase-11, pro-caspase-1, cleaved caspase-1, and IL-1β. Relative expression of (B) NLRP3; (C) pro-caspase-11; (D) cleaved caspase-11; (E) pro-caspase-1; (F) cleaved caspase-1; and (G) IL-1β. Data are means±standard deviations (n=3). *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001. ALF, acute liver failure; Control, normal; DMSO, dimethyl sulfoxide; NSA, necrosulfonamide.
Fig. 5. Effect of NSA on GSDMD in ALF.

(A) Western blotting of GSDMD and GSDMD-NT; (B) Relative expression of GSDMD and (C) GSDMD-NT. Data are means±standard deviations (n=3). **p<0.01, ***p<0.001, ****p<0.0001. ALF, acute liver failure; Control, normal; DMSO, dimethyl sulfoxide; NSA, necrosulfonamide.
NSA attenuated ALF via inhibition of the pyroptosis pathway
Casp1 and Casp11 mRNA expression was lower in the NSA group than in the ALF group (both p<0.05; Fig. 3A, B). Gsdmd transcription was inhibited in the ALF group, and was not changed by NSA intervention (Fig. 3C). After NSA intervention, the expression of pyroptosis pathway molecules, including GSDMD, GSDMD-NT, NLRP3, cleaved caspase-1, cleaved caspase-11, and L-1β, were significantly lower than those in the ALF group (Figs. 4, 5) (GSDMD, p<0.01; GSDMD-NT, p<0.001; NLRP3, p<0.05; cleaved caspase-1, p<0.05; cleaved caspase-11, p<0.005; and IL-1β, p<0.01).
Discussion
Given that GSDMD is the common executor protein of the two pyroptosis pathways, direct inhibition of GSDMD may be useful for treating pyroptosis-related diseases. In this animal study, we observed activation of pyroptosis in ALF and revealed the potential role of NSA in ALF mice. The pyroptosis pathway was activated in the ALF mice model, and NSA inhibited the pyroptosis pathway and improved survival of ALF model mice.
NSA is a recently identified direct inhibitor of GSDMD. Rathkey et al.26 were the first to report the direct binding of NSA and GSDMD in cells and mouse models of sepsis, resulting in the inhibition of GSDMD-NT oligomerization and inhibition of the formation of pyroptotic pores. Although NSA acts on human mixed lineage kinase domain-like (MLKL) protein, mouse MLKL protein has no Cys86 site, and the formation of disulfide bonds in MLKL protein is limited, resulting in NSA being unable to inhibit necrosis in mice.26 After NSA intervention, there was significant improvement in survival time and liver injury, as shown by the serum indices and pathological findings, which suggests a potential therapeutic role of NSA in ALF.
The study results revealed activation of both canonical and non-canonical pyroptosis pathways in ALF. After NSA intervention, GSDMD, GSDMD-NT, caspase-1, caspase-11, and inflammatory cytokines were inhibited, indicating that NSA functioned via the pyroptosis pathway. We also found that an upstream molecule of the pyroptosis pathway, NLRP3, was decreased after NSA intervention. The reason may be that NSA inhibited the cascade effect of pyroptosis. Pyroptosis can be activated by damage-associated molecular patterns (DAMPs) released by injured or dead cells.30 After NSA inhibited the GSDMD channel formation and prevented cell death, fewer DAMPs were released, which reduced the activation of NLRP3. That led to reduced potassium outflow, causing inhibition of NLRP3 activation.31 Previous studies have also shown that NSA not only inhibits GSDMD-NT oligomerization, but also inhibits the activation of NLRP3, caspase-1, and IL-1β.8
Some study limitations should be acknowledged. Previous studies have demonstrated the participation of both hepatocytes and immune cells in the pyroptotic pathway.18 Our study did not identify the functional cells in this process. Single-cell sequencing, cell coculture, and specific gene knockout may help to answer this question. Second, there may be cross-talk between different cell death pathways in ALF.32,33 The interaction between pyroptosis and other programmed cell death was not clarified in this study, and it is worthy of further attention. Third, NSA was administered 30 m before ALF modeling. However, in clinical practice, medication is typically initiated when the patient has already developed liver failure. Whether NSA is effective in patients with liver failure needs further investigation. Moreover, the side effects of NSA are not yet known, and whether the benefits outweigh the risks in clinical application is still unknown. There is still a long way to go from bench to bedside. In summary, canonical and non-canonical pyroptosis pathways were activated in ALF mice induced by LPS/D-GalN. NSA alleviated ALF by inhibiting the pyroptosis pathway. The results provide evidence in support of possible intervention for the clinical treatment of ALF.
Supporting information
Data are means±standard deviations (NSA, necrosulfonamide, n=4; other groups, n=5). *p<0.05, ***p<0.001.
Abbreviations
- ALF
acute liver failure
- ALT
alanine aminotransferase
- BCA
bicinchoninic acid
- DAMPs
damage-associated molecular patterns
- DMSO
dimethyl sulfoxide
- ELISA
enzyme-linked immunosorbent
- GSDMD
gasdermin D
- HE
hematoxylin-eosin staining
- IL-18
interleukin-18
- IL-1β
interleukin-1 beta
- MLKL
mixed lineage kinase domain-like
- NSA
necrosulfonamide
- PVDF
polyvinylidene difluoride
- qRT-PCR
quantitative real-time polymerase chain reaction
Data sharing statement
The data used in support of the findings of this study are included in the article.
References
- 1.European Association for the Study of the Liver EASL Clinical Practical Guidelines on the management of acute (fulminant) liver failure. J Hepatol. 2017;66(5):1047–1081. doi: 10.1016/j.jhep.2016.12.003. [DOI] [PubMed] [Google Scholar]
- 2.Lin S, Chen J, Wang M, Han L, Zhang H, Dong J, et al. Prognostic nomogram for acute-on-chronic hepatitis B liver failure. Oncotarget. 2017;8(65):109772–109782. doi: 10.18632/oncotarget.21012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Reuben A, Tillman H, Fontana RJ, Davern T, McGuire B, Stravitz RT, et al. Outcomes in Adults With Acute Liver Failure Between 1998 and 2013: An Observational Cohort Study. Ann Intern Med. 2016;164(11):724–732. doi: 10.7326/M15-2211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Clària J, Stauber RE, Coenraad MJ, Moreau R, Jalan R, Pavesi M, et al. Systemic inflammation in decompensated cirrhosis: characterization and role in acute-on-chronic liver failure. Hepatology. 2016;64(4):1249–1264. doi: 10.1002/hep.28740. [DOI] [PubMed] [Google Scholar]
- 5.Triantafyllou E, Woollard KJ, McPhail MJW, Antoniades CG, Possamai LA. The Role of Monocytes and Macrophages in Acute and Acute-on-Chronic Liver Failure. Front Immunol. 2018;9(2948):2948. doi: 10.3389/fimmu.2018.02948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Xu B, Jiang M, Chu Y, Wang W, Chen D, Li X, et al. Gasdermin D plays a key role as a pyroptosis executor of non-alcoholic steatohepatitis in humans and mice. J Hepatol. 2018;68(4):773–782. doi: 10.1016/j.jhep.2017.11.040. [DOI] [PubMed] [Google Scholar]
- 7.Tonnus W, Linkermann A. Gasdermin D and pyroptosis in acute kidney injury. Kidney Int. 2019;96(5):1061–1063. doi: 10.1016/j.kint.2019.07.002. [DOI] [PubMed] [Google Scholar]
- 8.Rashidi M, Simpson DS, Hempel A, Frank D, Petrie E, Vince A, et al. The Pyroptotic Cell Death Effector Gasdermin D Is Activated by Gout-Associated Uric Acid Crystals but Is Dispensable for Cell Death and IL-1beta Release. J Immunol. 2019;203(3):736–748. doi: 10.4049/jimmunol.1900228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Orning P, Lien E, Fitzgerald KA. Gasdermins and their role in immunity and inflammation. J Exp Med. 2019;216(11):2453–2465. doi: 10.1084/jem.20190545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Martinon F, Burns K, Tschopp J. The inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol Cell. 2002;10(2):417–426. doi: 10.1016/s1097-2765(02)00599-3. [DOI] [PubMed] [Google Scholar]
- 11.Swanson KV, Deng M, Ting JP. The NLRP3 inflammasome: molecular activation and regulation to therapeutics. Nat Rev Immunol. 2019;19(8):477–489. doi: 10.1038/s41577-019-0165-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lieberman J, Wu H, Kagan JC. Gasdermin D activity in inflammation and host defense. Sci Immunol. 2019;4(39):eaav1447. doi: 10.1126/sciimmunol.aav1447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Aglietti RA, Dueber EC. Recent Insights into the Molecular Mechanisms Underlying Pyroptosis and Gasdermin Family Functions. Trends Immunol. 2017;38(4):261–271. doi: 10.1016/j.it.2017.01.003. [DOI] [PubMed] [Google Scholar]
- 14.Mantovani A, Dinarello CA, Molgora M, Garlanda C. Interleukin-1 and Related Cytokines in the Regulation of Inflammation and Immunity. Immunity. 2019;50(4):778–795. doi: 10.1016/j.immuni.2019.03.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Jon A H, Daniel A P, Youssef A, Robert K E, Edward A M. Cytoplasmic LPS activates caspase-11: implications in TLR4-independent endotoxic shock. Science. 2013;341(6151):1250–1253. doi: 10.1126/science.1240988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Martinon F, Tschopp J. Inflammatory caspases: linking an intracellular innate immune system to autoinflammatory diseases. Cell. 2004;117(5):561–574. doi: 10.1016/j.cell.2004.05.004. [DOI] [PubMed] [Google Scholar]
- 17.Wang J, Ren H, Yuan X, Ma H, Shi X, Ding Y. Interleukin-10 secreted by mesenchymal stem cells attenuates acute liver failure through inhibiting pyroptosis. Hepatol Res. 2018;48(3):E194–E202. doi: 10.1111/hepr.12969. [DOI] [PubMed] [Google Scholar]
- 18.Wu J, Lin S, Wan B, Velani B, Zhu Y. Pyroptosis in Liver Disease: New Insights into Disease Mechanisms. Aging Dis. 2019;10(5):1094–1108. doi: 10.14336/AD.2019.0116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Thurston TL, Matthews SA, Jennings E, Alix E, Shao F, Shenoy AR, et al. Growth inhibition of cytosolic Salmonella by caspase-1 and caspase-11 precedes host cell death. Nat Commun. 2016;7(1):13292. doi: 10.1038/ncomms13292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wree A, Eguchi A, McGeough MD, Pena CA, Johnson CD, Canbay A, et al. NLRP3 inflammasome activation results in hepatocyte pyroptosis, liver inflammation, and fibrosis in mice. Hepatology. 2014;59(3):898–910. doi: 10.1002/hep.26592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Dai C, Xiao X, Li D, Tun S, Wang Y, Velkov T, et al. Chloroquine ameliorates carbon tetrachloride-induced acute liver injury in mice via the concomitant inhibition of inflammation and induction of apoptosis. Cell Death Dis. 2018;9(12):1164. doi: 10.1038/s41419-018-1136-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Geng Y, Ma Q, Liu YN, Peng N, Yuan FF, Li XG, et al. Heatstroke induces liver injury via IL-1beta and HMGB1-induced pyroptosis. J Hepatol. 2015;63(3):622–633. doi: 10.1016/j.jhep.2015.04.010. [DOI] [PubMed] [Google Scholar]
- 23.Chen YL, Xu G, Liang X, Wei J, Luo J, Chen GN, et al. Inhibition of hepatic cells pyroptosis attenuates CLP-induced acute liver injury. Am J Transl Res. 2016;8(12):5685–5695. [PMC free article] [PubMed] [Google Scholar]
- 24.Pourcet B, Zecchin M, Ferri L, Beauchamp J, Sitaula S, Billon C, et al. Nuclear Receptor Subfamily 1 Group D Member 1 Regulates Circadian Activity of NLRP3 Inflammasome to Reduce the Severity of Fulminant Hepatitis in Mice. Gastroenterology. 2018;154(5):1449–1464.e20. doi: 10.1053/j.gastro.2017.12.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wang T, Wang Z, Yang P, Xia L, Zhou M, Wang S, et al. PER1 prevents excessive innate immune response during endotoxin-induced liver injury through regulation of macrophage recruitment in mice. Cell Death Dis. 2016;7(4):e2176. doi: 10.1038/cddis.2016.9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Rathkey JK, Zhao J, Liu Z, Chen Y, Yang J, Kondolf HC, et al. Chemical disruption of the pyroptotic pore-forming protein gasdermin D inhibits inflammatory cell death and sepsis. Sci Immunol. 2018;3(26):eaat2738. doi: 10.1126/sciimmunol.aat2738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Xing WW, Zou MJ, Liu S, Xu T, Gao J, Wang JX, et al. Hepatoprotective effects of IL-22 on fulminant hepatic failure induced by d-galactosamine and lipopolysaccharide in mice. Cytokine. 2011;56(2):174–179. doi: 10.1016/j.cyto.2011.07.022. [DOI] [PubMed] [Google Scholar]
- 28.Kim SJ, Lee SM. NLRP3 inflammasome activation in D-galactosamine and lipopolysaccharide-induced acute liver failure: role of heme oxygenase-1. Free Radic Biol Med. 2013;65:997–1004. doi: 10.1016/j.freeradbiomed.2013.08.178. [DOI] [PubMed] [Google Scholar]
- 29.Baranova IN, Souza AC, Bocharov AV, Vishnyakova TG, Hu X, Vaisman BL, et al. Human SR-BI and SR-BII Potentiate Lipopolysaccharide-Induced Inflammation and Acute Liver and Kidney Injury in Mice. J Immunol. 2016;196(7):3135–3147. doi: 10.4049/jimmunol.1501709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Anderton H, Wicks IP, Silke J. Cell death in chronic inflammation: breaking the cycle to treat rheumatic disease. Nat Rev Rheumatol. 2020;16(9):496–513. doi: 10.1038/s41584-020-0455-8. [DOI] [PubMed] [Google Scholar]
- 31.Xue Y, Enosi Tuipulotu D, Tan WH, Kay C, Man SM. Emerging Activators and Regulators of Inflammasomes and Pyroptosis. Trends Immunol. 2019;40(11):1035–1052. doi: 10.1016/j.it.2019.09.005. [DOI] [PubMed] [Google Scholar]
- 32.Doerflinger M, Deng Y, Whitney P, Salvamoser R, Engel S, Kueh AJ, et al. Flexible Usage and Interconnectivity of Diverse Cell Death Pathways Protect against Intracellular Infection. Immunity. 2020;53(3):533–547.e537. doi: 10.1016/j.immuni.2020.07.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kesavardhana S, Malireddi RKS, Kanneganti TD. Caspases in Cell Death, Inflammation, and Pyroptosis. Annu Rev Immunol. 2020;38:567–595. doi: 10.1146/annurev-immunol-073119-095439. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data are means±standard deviations (NSA, necrosulfonamide, n=4; other groups, n=5). *p<0.05, ***p<0.001.