

Abstract
Alzheimer’s disease (AD) is the most common cause of dementia worldwide, and its prevalence is rapidly increasing due to extended lifespans. Among the increasing number of genetic risk factors identified, the apolipoprotein E (APOE) gene remains the strongest and most prevalent, impacting more than half of all AD cases. While the ε4 allele of the APOE gene significantly increases AD risk, the ε2 allele is protective relative to the common ε3 allele. These gene alleles encode three apoE protein isoforms that differ at two amino acid positions. The primary physiological function of apoE is to mediate lipid transport in the brain and periphery; however, additional functions of apoE in diverse biological functions have been recognized. Pathogenically, apoE seeds amyloid-β (Aβ) plaques in the brain with apoE4 driving earlier and more abundant amyloids. ApoE isoforms also have differential effects on multiple Aβ-related or Aβ-independent pathways. The complexity of apoE biology and pathobiology presents challenges to designing effective apoE-targeted therapeutic strategies. This review examines the key pathobiological pathways of apoE and related targeting strategies with a specific focus on the latest technological advances and tools.
The pathogenesis of Alzheimer disease
Alzheimer’s disease (AD) is the most common cause of dementia, accounting for approximately 60–80% of all dementia cases [1]. Despite the broad prevalence and rising incidence of AD, there are only six FDA-approved drugs currently used to treat symptoms including cholinesterase inhibitors, NMDA receptor antagonists, and other neuromodulatory agents that are presently prescribed to AD patients to ease the cognitive symptoms [2]. The newest FDA-approved drug, aducanumab, is an antibody-based immunotherapy designed to remove amyloid-β (Aβ) plaques. The clinical efficacy of this new drug is under debate limiting its application to broad AD patients [3, 4]. Overall, there is an urgent need to develop effective therapeutics to reverse or robustly attenuate pathological progression of AD and associated neurodegeneration.
At the cellular level, AD is characterized by synaptic dysfunction and neurodegeneration, neuroinflammation, as well as vascular dysfunction [5, 6]. This combination of neuronal loss surrounded by a heightened inflammatory state is thought to give rise to the eventual decline in cognition seen in AD patients. The key neuropathological hallmarks of AD include extracellular Aβ plaques and intracellular neurofibrillary tangles (NFTs) composed of aggregated tau protein [7, 8]. Much of what we have learned about the etiology of AD comes from studies of the familial form of early-onset AD (FAD), which occurs before 65 years of age. This hereditary form of AD is primarily caused by specific mutations in the genes encoding amyloid precursor protein (APP), presenilin 1 (PSEN1), and presenilin 2 (PSEN2), essential for Aβ production, suggesting a critical role of Aβ in disease development [9]. However, more than 95% of AD cases are sporadic or late-onset AD (LOAD), where the etiology is heavily influenced by environmental and genetic risk factors [10]. With considerable genetic heterogeneity in LOAD cases, it has been difficult to pinpoint specific genes or pathways that directly lead to the onset of clinical pathology. However, as the strongest genetic risk factor for LOAD [11], the apolipoprotein E (APOE) gene clearly impacts the majority of the pathogenic pathways that contribute to AD. As such, targeting apoE offers a unique opportunity to potentially benefit a greater number of AD patients [12].
ApoE isoform is a risk determinant for AD and related dementias
There are three predominant APOE alleles in humans; the ε2 (APOE2), ε3 (APOE3), and ε4 (APOE4) alleles which confer varying levels of disease risk. APOE4 is a major genetic risk factor for AD in a gene dose-dependent manner increasing risk by up to 15 fold in homozygotes [13], whereas APOE2 reduces AD risk by almost half and contributes to longevity [14]. In addition to the relatively common variants, there have been various rare variants of apoE identified, such as apoE3-R136S (apoE3-Christchurch; apoE3-Ch), apoE3-V236E (apoE3-Jacksonville; apoE3-Jac,) and apoE4-R251G [15–19], that are thought to confer some protection against AD pathology. These rare mutations are being investigated in the hope of identifying the molecular mechanisms involved in alleviating disease risks and developing novel therapeutic strategies. Mounting evidence demonstrates that APOE4 increases the risk of developing AD via a combination of gain of toxic effects and loss of protective functions [20]. While APOE2 has been shown to offer protection against AD-related pathology [21, 22], the mechanisms involved remain unclear. Intriguingly, Insel, et al. has reported that carrying an APOE2 in the presence of APOE4 may offer some protection against Aβ accumulation compared to APOE3 [23]. However, others have found that the odds ratio of APOE2/4 individuals is more similar to that of APOE4 carriers than APOE2 carriers [21], suggesting that the increased risk associated with APOE4 is more dominant than the protection offered by APOE2. The multitude of pathological pathways by which apoE impacts AD risk and disease progression makes it an ideal therapeutic target for AD [24].
APOE4 has also been shown to increase the risk for other dementias. For example, APOE4 carriers are more likely to have increased severity of Lewy body pathology, independently of AD pathology [25–27]. Similarly, APOE4 was shown to increase the risk of Parkinson’s disease dementia as well as reduce the age of symptom onset [28]. More recently, animal studies have revealed that apoE4 regulates α-synuclein pathology and exacerbates its toxic effects independent of amyloid pathology [29, 30]. Additionally, patients with vascular dementia carrying APOE4 present with greater cognitive impairment compared to other alleles [31]. Thus, gaining an understanding of the mechanisms of apoE in disease pathogenesis will ultimately shed light on therapeutic strategies for the treatment of AD and related dementias. Here, we summarize relevant biological functions of apoE and potential pathological mechanisms that contribute to disease progression. We will further discuss current therapeutic strategies targeting apoE to improve AD pathology and offer insights on how these strategies could be improved based on recent evidence from novel technologies.
APOE biology: synthesis, structure, and function
ApoE is a 299 amino acid glycoprotein with a molecular weight of 34 kDa. It primarily functions as a lipid transporter responsible for delivering cholesterol and phospholipids throughout the body. In the periphery, apoE is mainly produced by hepatocytes and macrophages in the liver [32]. While apoE does not cross the blood-brain barrier (BBB), it is also abundantly expressed in the central nervous system (CNS) by astrocytes, activated microglia, vascular mural cells, choroid plexus cells, and to a lesser extent in stressed neurons [33–37]. The three major isoforms of human apoE are distinguished by amino acid positions 112 and 158, which vary between a cystine and an arginine (apoE2: Cys112/Cys158; apoE3: Cys112/Arg158; apoE4: Arg112/Arg158). This single amino acid substitution between apoE4 and apoE3, and apoE3 and apoE2 drastically alters the functionality of apoE, resulting in isoform-specific variations in structure that modulate lipid binding, receptor binding, oligomerization propensity, and stability [38–41] (Fig. 1). Thus, understanding key differences in the apoE structure between isoforms is paramount to understanding its function. Although the precise structure of native apoE is still ambiguous due to the protein’s propensity to aggregate, there is a Nuclear Magnetic Resonance (NMR)-based working model generated from a mutated, monomeric apoE3 [42]. ApoE consists of two main structural domains connected by a hinge region. The N-terminal domain (residues 1-167) contains the receptor binding region [43], while the C-terminal domain (residues 206–299) contains the lipid-binding region [44] (Fig. 1). Lipids are loaded onto apoE via interaction with transmembrane ATP-binding cassette (ABC) transporters such as ABCA1 and ABCG1 [45].
Fig. 1.

Structural model of apoE highlighting AD-related amino acid variations. ApoE is a 299 amino acid glycoprotein with a molecular weight of 34 kDa (PDB 2L7B). It is formed of two independently folded domains linked by a hinge region: the N-terminal domain (residues 1-167) contains the receptor-binding region while the C-terminal domain (residues 206–299) includes the lipid-binding region. There are three major apoE isoforms that differ at amino acid positions 112 and 158: apoE2 (C112/C158); apoE3 (C112/R158); and apoE4 (R112/R158). Additional rare apoE variants have been identified: apoE3-Christchurch (R136S), apoE3-Jacksonville (apoE3-V236E), and apoE4-R251G
In the brain, apoE plays an important role in transporting cholesterol and other lipids to neurons through binding to cell-surface apoE receptors involved in lipoprotein metabolism, such as low-density lipoprotein receptor (LDLR), and LDLR-related protein 1 (LRP1) [46]. ApoE also interacts with heparan sulfate proteoglycans (HSPG) [47] and binds heparin through two separate sites [48]. Modulation of these apoE receptors has been shown to affect amyloid and tau pathologies [49–52], further supporting the role of apoE in AD pathogenesis. Receptor binding of apoE is dependent on the isoform, lipidation status, and aggregation status of apoE [5, 42]. However, it remains unclear whether downstream receptor-mediated pathways are modulated by apoE isoforms. Understanding these mechanisms will help to fully elucidate isoform-dependent pathways involved in AD pathogenesis and could provide novel therapeutic targeting strategies.
Physiological function of apoE
ApoE is abundantly expressed in the periphery and in the CNS. However, due to the BBB, apoE in the periphery and in the CNS exist as distinct pools [53]. Therefore, it is critical to consider the independent role that each may play in AD pathogenesis, and the opportunity presented by each for therapeutic intervention. ApoE in the periphery is predominantly produced by the liver [32]. Peripheral apoE maintains lipid homeostasis by participating in the redistribution and metabolism of lipids, such as triglycerides, cholesterol, cholesteryl esters, and phospholipids, through the formation of lipoprotein particles. ApoE isoforms have been shown to be differentially associated with peripheral lipoprotein particles. For instance, apoE4 is mostly found in triglyceride-rich particles like chylomicrons and very low-density lipoproteins (VLDL), whereas apoE2 and apoE3 have preference to the high-density lipoproteins (HDL). Thus, the single Arg to Cys amino-acid substitution at position 112 dictates the differential distribution of apoE isoforms among lipoprotein particles [54]. Single amino acid substitutions in rare apoE isoforms may also modulate their distribution among peripheral lipoproteins. For instance, apoE3-Jac exhibits higher cholesterol efflux capacity compared to apoE3, suggesting that apoE3-Jac may differentially bind lipids and could be differentially distributed among lipoprotein particles compared to the common apoE isoforms. Peripheral apoE is also important for cardiovascular function and immune modulation, both factors that contribute to AD risk [20, 55]. These effects will be discussed in more detail later in this review.
In the CNS, apoE-mediated cholesterol and lipid transport plays a critical role in synapse formation and tissue repair [5]. It also plays a role in neurite outgrowth following injury in an isoform dependent manner with astrocytic apoE3 inducing greater neurite outgrowth than astrocytes secreting apoE4 [56]. Human apoE4-targeted replacement (TR) mice, as well as human APOE4 carriers, show a reduction in dendritic spine density even in the absence of disease pathology [57]. This is consistent with studies which revealed that apoE4 alters structural reorganization of neurons [58], reduces expression of key synaptic proteins [59], and inhibits glutamatergic signaling which are critical for neuronal plasticity and network maintenance [60].
Emerging evidence shows that the function of apoE is largely cell type-specific [61, 62]. While apoE expression in the brain was first described in astrocytes, it has been found to be drastically up regulated in activated microglia and in stressed neurons under pathological conditions and injury [34, 36, 63, 64]. Recent reports also suggest that primary astrocytes and microglia express and secrete apoE with varying sizes likely due to different post-translational modifications and lipid compositions, which might contribute to cell type-specific functions in response to injury [65]. Other cell types, such as pericytes and oligodendrocytes, have also been reported to express apoE more abundantly after liver X receptor (LXR) stimulation and after injury [66–69]. Thus, exploring the structure, lipidation status, and biochemical properties of apoE isoforms expressed by individual brain cell types will be critical for understanding apoE-related effects in the brain.
Pathobiological function of apoE
ApoE and Aβ
It is well established that apoE co-deposits with Aβ in amyloid plaques [70]. The interaction between apoE and pathological Aβ deposition appears to be a central mechanism by which apoE contributes to AD risk. Knocking out endogenous Apoe in amyloid model mice alters the morphology of Aβ plaques from compact to diffused [71, 72], suggesting that apoE may play a major role in Aβ fibrilization and amyloid deposition. Importantly, the effect of apoE on amyloid pathology is shown to be isoform-dependent (apoE4 > apoE3 > apoE2). ApoE4-TR mice show increased Aβ levels in brain tissues and in the CSF compared to apoE3-TR [73, 74]. Additionally, post-mortem studies of human brain tissue have increased Aβ plaque deposition and earlier onset of amyloid pathology in individuals carrying APOE4 [75–77]. Positron emission tomography (PET) imaging revealed that APOE4 carriers show earlier deposition of Aβ [78, 79], as well as greater overall deposition and broader cortical distribution [80–82]. Conversely, APOE2 carriers showed delayed onset of Aβ deposition, less severe pathology, and protected cognitive function [83]. Studies of Aβ kinetics in the presence of apoE suggests that apoE4 stabilizes soluble, cytotoxic, oligomeric Aβ fragments and enhances fibrillogenesis [84]. In addition, apoE4 has been shown to accelerate early seeding of amyloid pathology. For instance, using mouse models in which apoE isoform expression is conditionally induced, astrocytic expression of apoE4 during the seeding stage of Aβ plaque development, but not in the rapid growth period, resulted in a substantial increase in plaque deposition as well as an increased Aβ half-life in the brain [85]. Similar results were found using APP/PS1-21 animal models treated with antisense oligonucleotides (ASOs) against APOE4, where inhibition of apoE4 during the seeding stage leads to larger Aβ plaques with reduced plaque-associated neuritic dystrophy [86]. The findings of earlier Aβ deposition [87], greater Aβ oligomerization, and lower CSF and plasma Aβ levels in human APOE4 carriers [88, 89] strongly support the idea that apoE isoforms differentially regulate Aβ fibril formation via its interaction with Aβ, and that this interaction may represent a potential target for therapeutic intervention at early stages of the disease.
In addition to promoting Aβ plaque formation, apoE is also involved in the clearance of Aβ via various mechanisms, such as receptor-mediated clearance and proteolytic degradation. LRP1 receptor in neurons is shown to mediate Aβ clearance via the uptake of Aβ/apoE complexes [90–92]. Due to the reduced stability of the complex between apoE4 and Aβ [93, 94], this uptake process is impaired in APOE4 carriers. This clearance impairment is further compounded by alterations in receptor binding and the competition of apoE for Aβ receptor binding sites [95], resulting in dramatically reduced receptor-mediated clearance in the presence of apoE4. In addition, soluble Aβ can be cleared by proteolytic enzymes, such as metalloendopeptidases, plasminogen activators, matrix metalloproteinases, and lysosomal peptidases [96]. It has been shown that apoE promotes Aβ degradation within microglia in an isoform-dependent manner through neprilysin (NEP) and insulin-degrading enzyme (IDE) in the extracellular space. Enhanced expression of lipidated apoE was also shown to stimulate proteolytic Aβ degradation through LXRs and ABCA1 [97]. Given that apoE4 is a less-effective lipid transporter compared to apoE2 and apoE3, the apoE4-mediated proteolytic degradation of Aβ is also compromised [98], leading to reduced Aβ clearance.
ApoE and tau
Along with Aβ plaque formation, a major hallmark of AD pathology is the presence of NFTs composed of hyperphosphorylated tau aggregates. ApoE4 has been shown to increase tau phosphorylation compared to apoE2 and apoE3 in the presence of Aβ oligomers [99]. Human studies using PET imaging revealed that APOE4 carriers show an increased tau deposition both in the presence and absence of Aβ plaques [100]. Neuronal apoE4 was shown to promote tau phosphorylation and cell death compared to apoE3 in induced pluripotent stem cell (iPSC) cultures [101]. Additionally, animal models of tauopathies have shown that apoE4 is associated with greater total tau and phospho-tau levels [102], and exacerbates tau-mediated neurodegeneration by modulating microglial activation [103, 104]. A recent study reported that selective deletion of astrocytic apoE4 can reduce tau-related synaptic degeneration, disease-associated gene signatures, and protect against microglial phagocytosis [105]. Moreover, a study using an adeno-associated virus (AAV)-tau delivery approach found that apoE2 may lead to increased tau phosphorylation and aggregation and is associated with the risk of developing primary tauopathy. The increased tau aggregation is potentially due to the formation of tau/apoE complexes, which primarily occur in the presence of non-lipidated apoE2 [106]. A recent GWAS study indicates that apoE2 distinctly regulates protein phosphatase 2 A (PP2A) activity, a major tau phosphatase in the human brain, which could offer protection against AD risk. The authors further suggest that the protective mechanism of APOE2 might be distinct from the deleterious effect of APOE4 on AD risk [107]. These results demonstrate the effects of apoE on the pathogenesis of tauopathies, as well as tau-mediated neurotoxicity, and provide supporting evidence that the role of apoE is isoform-dependent.
The isoform-specific interactions between apoE and tau have become of interest not only in AD research, but also for other tauopathies such as frontotemporal dementia (FTD), chronic traumatic encephalopathy (CTE), and corticobasal degeneration (CBD) [103, 106, 108, 109]. For example, human APOE4 carriers with FTD, a primary tauopathy, are younger in age at the onset of tau pathology, exhibit signs of exacerbated neurodegeneration, and show greater cognitive decline than non-APOE4 carriers [108, 110]. Together these findings suggest that apoE influences tau pathology, even independently of Aβ pathology, which contributes to disease progression. Thus, understanding the molecular mechanisms of apoE in the context of tauopathy might provide critical information on developing strategies for AD and other tau-related neurodegenerative diseases.
Early research showed that apoE binds to the region of tau believed to be responsible for aggregation into pathogenic NFTs. It has been postulated that apoE might bind tau directly and block its phosphorylation sites. Such interaction was also observed to be isoform-specific, with apoE3 showing much stronger binding affinity to the microtubule-binding region of tau than apoE4 [111]. The reduced affinity of apoE4 binding to tau may increase the availability and likelihood of GSK3-mediated tau hyperphosphorylation, which leads to an increased formation of NFTs [112]. Alternatively, it has been postulated that apoE4 inhibits the Wnt signaling pathway through LRP5/6 receptors by increasing GSK3 activity, leading to increased tau phosphorylation [113]. These potential mechanisms continue to be studied to better understand the contributions of apoE to tau pathogenesis.
ApoE and neuroinflammation
A growing body of literature now suggests that inflammation plays a key role in neurodegenerative processes and is subject to modulation by apoE. While apoE contributes to AD pathogenesis in a wide range of pathways, recent data suggests that these different pathways may have a common node in neuroinflammation [114]. Microglia, often observed surrounding plaques in post-mortem brain tissue, have been shown to mediate the inflammatory response and phagocytosis of amyloid plaques [115]. Interestingly, studies have demonstrated that apoE-deficient mice show reduced microglial reactivity to plaques [105, 116, 117], suggesting that apoE may be necessary for the microglial response to amyloid aggregation. Emerging studies show that disease-associated microglia (DAM), or microglial neurodegenerative (MGnD) phenotype, exhibit a conserved transcriptional signature across AD mouse models with Apoe serving as one of the central regulators [116, 118]. Furthermore, the impact of apoE on microglial function is likely isoform-specific [119, 120]. A recent report demonstrates that apoE3 is more efficient at inducing microglial response to injected Aβ compared to apoE4, and such observation may be mediated by triggering receptor expressed on myeloid cells 2 (TREM2) [121]. Expressed specifically by microglia in the brain, TREM2 is shown to interact with apoE with high affinity and modulate microglial responses [122]. There is also evidence that binding of apoE to TREM2 is dependent on apoE isoform [123] and lipidation status [124], which may account for differences in microglial function between isoforms. Specifically, apoE4 may impair homeostatic microglial functions compared to other isoforms [125] potentially due to its reduced lipidation and reduced affinity to TREM2.
In the peripheral immune system, APOE genotype has been shown to modulate the level of C-reactive protein (CRP) in plasma or serum in multiple cohorts [126–128]. CRP is an inflammatory protein primarily produced by hepatocytes in response to injury or inflammation [129]. Interestingly, APOE4 carriers show lower levels of CRP relative to APOE3 individuals and APOE2 carriers [126–128]. CSF proteomics has also shown a reduction of CRP and complement cascade proteins in APOE4 carriers versus APOE2 carriers and APOE3 individuals. Contrary to this trend in genotype, cumulative incidence of AD was shown to markedly increase as levels of serum CRP increase, with the greatest effect in APOE4 carriers [126]. However, APOE but not CRP haplotype was associated with life-long cognitive decline in a longitudinal cohort [130], which does not support a causal effect of CRP on cognitive decline. Therefore, APOE4 carriers may be subjected to aberrant immune responses to pathological development, which can ultimately lead to harmful effects on injury responses and cognitive deficits. Thus, targeting apoE-mediated inflammatory responses may attenuate AD pathologies and neurodegeneration, and is a valid therapeutic approach to be explored.
ApoE in dysregulation of lipid metabolism
As a major lipid transporter in the brain, apoE has been shown to mediate lipid and cholesterol metabolism in an isoform-dependent manner [131]. In rat primary astrocytes and neurons, recombinant apoE2 was more efficient at promoting the efflux of cholesterol and phosphatidylcholine (PC), followed by apoE3, and then apoE4 [132]. Deficiency in facilitating cholesterol efflux may result in its intracellular accumulation, which can lead to cytotoxicity. For instance, APOE4 iPSC-derived astrocytes tend to build up more unsaturated fatty acids and cholesterol compared to APOE3 astrocytes [62, 133], and are more prone to accumulation of esterified cholesterol stored as lipid droplets [134]. In addition to astrocytes, microglia can also accumulate lipid droplets during aging and under disease-associated conditions [135, 136]. Claes et al. have reported that human iPSC-derived xenografted microglia (xMG) transplanted into the brains of an amyloid mouse model accumulate numerous lipid droplets in the vicinity of amyloid plaques, but not in those distant from plaques [136]. In animal studies, no major changes in lipid profiles were reported in the brains of young and middle aged apoE-TR mice [137]. However, apoE2-TR mice were shown to have lower cholesterol levels in the cortex, and higher levels in CSF and plasma compared to apoE3- and apoE4-TR mice [138], suggesting more efficient cholesterol efflux in apoE2-TR mice.
In human observational cohort studies, individuals with different APOE genotypes exhibit distinct lipid profiles in the periphery. APOE2 carriers present with lower total cholesterol, and APOE4 carriers with higher total cholesterol compared to APOE3 carriers [115]. However, both APOE2 and APOE4 carriers have higher plasma triglycerides compared to APOE3 carriers [115]. Lipid dyshomeostasis has also been reported in postmortem AD brains [134] and has been shown to be apoE isoform dependent. For instance, demented APOE4 carriers display decreased levels of several lipid classes, such as phosphatidylethanolamine (PE) and phosphatidic acid (PA) [139]. The association of apoE4 with lower cholesterol transport capacity and increased lipid droplet accumulation may be a major contributing mechanism through which APOE4 presents a strong risk for AD.
ApoE in vascular dysfunction and BBB integrity
The contribution of the vasculature to AD pathogenesis is gaining an increased interest as research has shown that vascular cognitive impairment and dementia (VCID) and AD share some converging pathology and etiology, including the involvement of apoE [140]. ApoE4 intensifies cholesterol dysregulation, stimulates inflammation, promotes metabolic dyshomeostasis, and enhances BBB breakdown, which promotes cerebrovascular damage and increases the risk for both VCID and AD [140]. Interestingly, both APOE2 and APOE4 have been found to correlate with increased amyloid accumulation in the parenchymal and meningeal cerebrovasculature, classified as cerebral amyloid angiopathy (CAA) [141]. However, APOE2 was found to reduce Aβ positivity in patients with AD-related cognitive decline but increase Aβ positivity in patients with VCID, potentially due to the presence of vascular amyloid which increases the risk of CAA [142]. The progression of CAA alters Aβ clearance mechanisms and leads to vascular pathological changes, including augmented vascular pulsation and reduced vascular smooth muscle cell coverage [143]. The reduction of Aβ clearance may be further exacerbated by apoE4, which causes premature shrinkage of meningeal lymphatic vessels, resulting in abnormal lymphatic function [144]. Conversely, apoE-enriched HDL particles have been shown to drastically diminish CAA in bioengineered vessels in an in vitro system by reducing vascular Aβ deposition [145]. Further investigation is required to elucidate how apoE-containing HDL particles reduce CAA, and whether these mechanisms are isoform-dependent.
Vascular mural cells (VMCs), including smooth muscle cells and pericytes are responsible for the homeostasis and function of the cerebrovasculature. It was recently demonstrated that VMC-derived apoE4 leads to a reduction of arteriole blood flow and behavioral deficits, likely due to increased astrogliosis of the vasculature [146]. In addition, cognitively normal APOE4 carriers have reduced retinal capillary density, which could act as a measurable in vivo model of impaired capillary blood flow in the cerebral vasculature [147]. However, it is interesting to note that APOE4 carriers have a reduced risk of developing glaucoma due to impaired activation of microglia in the retina [148]. Another study shows that APOE4 carriers presenting with increased arteriole stiffness perform worse on memory tasks compared to non-carriers [149]. Overall, APOE4 is associated with reduced blood flow, increased arteriole stiffness, and enhanced sensitivity to hypertension [150]. As such, long-term treatment with angiotensin receptor blockers has been shown to improve memory and neuroinflammation independently of Aβ pathology in female mice expressing human APOE4 [151]. Similarly, research suggests that vascular endothelial growth factor A (VEGFA) is protective against AD in apoE4-TR mice and could be a promising therapeutic target [152]. In addition, systemic treatment with epidermal growth factor (EGF) improved memory performance of apoE4-TR mice independent of Aβ pathology [153]. Neuropilin 1, a regulator of angiogenesis, also modifies the risk for poor cognitive outcomes based on APOE4 status [154]. Together these data provide evidence that apoE contributes to vascular pathology in an isoform-dependent manner and could be a viable target for therapies toward AD and related vascular dementias.
Emerging evidence suggests that the breakdown of the BBB may be one of the earliest pathogenic events of cognitive decline and AD pathology. ApoE has been shown to modulate cerebrovascular tight junction integrity independent of CAA in AD brains [155]. Additionally, a study showed that BBB breakdown contributes to cognitive decline in APOE4 carriers independent of Aβ or tau pathology [156]. A major component of the BBB are vascular endothelial cells, which form tight junctions to create a functional barrier [157]. Interestingly, apoE2 and apoE3 are shown to induce proliferation of vascular endothelial cells while apoE4 decreases their proliferation [158], potentially contributing to the breakdown of the BBB. Studies also indicate that apoE4 disrupts the endothelial BBB integrity by influencing extracellular matrix, cell adhesion machinery, cytoskeleton stability, and translation in brain endothelium. The progressive BBB breakdown in the presence of apoE4 leads to synaptic dysfunction and behavior deficits [159, 160]. BBB breakdown is also shown to be a result of dysfunctional pericytes, which regulate the BBB by controlling gene expression in endothelial cells and inducing polarization of astrocytic end-feet [161]. Pericytes expressing apoE4 have a reduced capacity for supporting endothelial cell function, which leads to impaired BBB integrity and increased susceptibility to cognitive decline [66]. Importantly, accelerated BBB breakdown and degeneration of capillary pericytes can also be recapitulated in amyloid mouse models and human iPSC models [67, 162]. Using a novel iPSC-based 3-D model that recapitulates human BBB in vitro, a recent study showed that dysregulation of calcineurin/NFAT-signaling in pericytes induces apoE4-associated CAA pathology, whereas inhibition of calcineurin reduces apoE expression and vascular amyloid accumulation [67]. Additionally, apoE4 accelerates the degradation of the BBB through activation of the cyclophilin A-matrix metalloproteinase-9 (MMP9) pathway in pericytes, whereas suppression of this pathway improves BBB integrity and prevent apoE4-mediated behavioral deficits in amyloid mouse models [162]. These studies demonstrate that apoE plays a major role in maintaining BBB integrity and that targeting these pathways may present novel therapeutics for AD-related neurodegeneration.
ApoE and other comorbidities
Many AD patients present with comorbidities including cardiovascular disease (CVD) and type 2 diabetes mellitus (T2DM). Not surprisingly, diabetes appears to accelerate cognitive decline and increase vascular pathology [163]. Interestingly, APOE4 has been shown to increase the risk for both CVD (OR = 3.0, p = 0.018) and T2DM (OR = 2.2, p = 0.04) independently of AD pathology [164, 165]. In addition, APOE4 is shown to increase the prevalence and hazard for metabolic syndrome in a cross-sectional study of 4,408 middle-aged men [166]. Evidence suggests that these metabolic deficits can occur before AD pathology becomes evident [167]. Thus, some have proposed research to focus on metabolic dysregulation for novel biomarkers and therapeutic targets for AD pathology. Given the influence of metabolism on modifying AD pathology, Polis and Samson have suggested a new perspective of AD as a complex metabolic disorder, which may offer alternative therapeutic strategies in the future [168].
Studies that inform therapeutic strategies
While APOE has been known to be the strongest genetic risk factor for AD for decades, there are currently no apoE-targeted FDA-approved therapeutics for the treatment of AD. Although apoE has been extensively studied in the context of AD pathogenesis, there is still much more to learn about the mechanisms involved. Recently, a novel “ApoE Cascade Hypothesis” was proposed suggesting that the biochemical and biophysical properties of apoE impact a cascade of events at the cellular and systems levels, which ultimately contribute to the aging-related cognitive decline, pathogenic conditions, and AD disease development [169]. Here, we highlight some of the most recent advances in the field that may help guide novel apoE-targeted therapeutic strategies.
ApoE in omics analysis
Recent advances in technology have allowed for the generation of bioinformatic data from Omics experiments in iPSCs and mouse models, as well as in AD patients. Collectively, these data have shed light on important genetic targets and pathways that are modulated by APOE genotype (Table 1). Recently, RNA-sequencing of isogenic iPSC-derived neurons revealed that apoE4 disrupts pathways related to synaptic formation, which leads to increased synaptic transmission [62]. Additionally, apoE expression is reduced in apoE4 astrocytes compared to apoE3, and the presence of apoE4 was shown to alter cholesterol metabolism and impair the uptake of Aβ [62]. Likewise, apoE4 expression in microglia is associated with less efficient Aβ clearance and activation of inflammatory genes. These data suggest that APOE4 causes global gene expression changes that may alter cellular function and lead to AD pathology [62]. Similar results can be seen from single nucleus RNA-sequencing of post-mortem human samples, providing further evidence that APOE expression plays a major role in AD pathology [170]. In support of these results, transcriptome profiling of microglia during disease progression reveals that apoE may be a key upstream regulator of the transition from homeostatic to DAM [116]. Additionally, transcriptomic analysis of apoE-TR mice found differential regulation of genes involved in energy metabolism in apoE4 mice compared to apoE3 mice, suggesting that apoE4 mice are more vulnerable to bioenergetic deficits which could also induce or exacerbate AD pathology [171]. Transcriptomics analysis of cerebral organoids generated using iPSCs from AD patients reveals genes associated with disrupted RNA metabolism with stress granule formation especially in the presence of APOE4. Generating cerebral organoids from isogenic iPSC lines where apoE4 is converted to apoE3 attenuates the apoE4-related phenotypes [172]. This is also recapitulated in a study using apoE-TR mice which identified distinct serum metabolite profiles and upregulated lipid levels in APOE2 mice compared to APOE3 and APOE4 [173]. Furthermore, proteomic analysis of human brain tissue and CSF has revealed apoE isoform-dependent changes, with a reduction in synaptic and mitochondrial function and increased abundance of neuroimmune signaling [174–176]. These data demonstrate the importance of apoE in the initiation and progression of AD pathology across multiple levels. One question the field has been hoping to address with recent bioinformatic approaches is how various genetic risk factors, such as APOE and TREM2, play a synergistic role and interact through common pathways. Recently, a multi-omics comparison of human ESC-derived microglia-like cell lines found that upregulation of apoE is a converging pathogenic node between SORL1 and TREM2 mutant models of AD. These data suggest that various risk factors for AD may have overlapping mechanisms through the upregulation of apoE, specifically in microglia [177].
Table 1.
APOE isoform effects in aging and AD: Insights from multi-omics and biomarker studies
| Cohort | Findings | Source | DOI |
|---|---|---|---|
| Transcriptomics | |||
| Normal; AD | • APOE2/3 AD brains are enriched in complement pathway genes (C4A, C4B, and HSPA2) whose expressions are associated with an increase of pTau231/tTau [296] | Brain | 10.1038/s41380-021-01266-z |
| AD | • APOE2 is associated with genes involved in protein synthesis, folding and degradation, response to unfolded protein, autophagy, and mitochondrial function [139] | Brain | 10.1186/s13195-019-0558-0 |
| Normal; AD | • APOE is one of the DAM-like signature genes that is significantly up-regulated in human AD brain assessed by single cell- or single nuclei-RNA Sequencing [64, 297–299] | Brain |
10.1038/s41467-020-19737-2 10.1038/s41586-019-1195-2 10.1007/s00401-020-02200-3 10.1007/s00401-021-02263-w |
|
Normal APOE mice |
• APOE4 reduces energy expenditure in young females and decreases glucose oxidation by redirecting flux through aerobic glycolysis [300] |
Brain Plasma |
10.1186/s13024-021-00483-y |
|
APOE mice Amyloid mouse models |
• APOE4 increases the expression of Serpina3 family genes, whereas APOE2 drives distinct blood metabolome profile [173] • DAM/MGnD/ARM exhibit conserved transcriptional signatures across different AD mouse models with Apoe being one of the central regulators [116, 118, 301] |
Brain Plasma |
10.1016/j.neuron.2020.02.034 10.1016/j.cell.2017.05.018 10.1016/j.immuni.2017.08.008 10.1016/j.celrep.2019.03.099 |
| Normal |
• Microglial gene expression modules associated with APOE4 and sex are also enriched with genes involved in cholesterol absorption and lipid digestion [302] • Microglial gene expression modules associated with APOE4 and age suggest perturbations in lipid and carbohydrate metabolism as well as microglial activation [302] |
Brain | 10.1111/acel.13606 |
|
Normal; AD APOE mice |
• APOE4 astrocytes and microglia demonstrate dysregulated lipid metabolism [179] • APOE4 alters the matrisome, ECM, and immune pathways in hiPSC-mixed cortical cultures and AD brains [179] |
Brain hiPSC |
10.1016/j.cell.2022.05.017 |
| Proteomics | |||
| Normal; AD | • AIF1, APP, GDI2, HSP90AA1, METAP2, NACA, NCK1, PRDX1, RPS27A, SFTPD and UFC1 are downregulated in AD versus control among APOE4 carriers. [303] | Plasma, brain | 10.18632/aging.202950 |
| Normal |
• APOE genotype is associated with levels of PSD95 in superior temporal cortex in AD (E4/* > E3/E3 > E2/*) [59] • APOE2 is associated with significantly increased levels of PSD95 in superior temporal cortex [59] |
Brain | 10.1016/j.neurobiolaging.2005.04.008 |
| Control; AsymAD; AD | • The matrisome module (i.e., extracellular matrix associated proteins) is influenced by the APOE4 but is not associated with cognitive decline rate after adjustment for neuropathology [304]. | Brain | 10.1038/s41593-021-00999-y |
| Lipidomics | |||
| Normal; AD |
• APOE4 copy number is positively associated with LysoPE and acylcarnitine species [305] • APOE4 copy number is negatively associated with PE(O), PE(P), ceramides, and triglycerides versus APOE2 carriers [305] |
Plasma | 10.3233/jad-191304 |
| Normal; Aging |
• LDL cholesterol levels are genotype dependent (E4/E4 > E4/E3 > E3/E3 > E2/E3 > E2/E2) [306] • APOE2 is associated with increased levels of most phospholipids (i.e., lysophosphatidylcholines and all PE subclasses) [306] • APOE4 is associated with reduced levels of phosphatidylinositol relative to APOE2 and APOE3 carriers [306] |
Plasma | 10.3233/jad-190524 |
| AD | • APOE4 is associated with reduced levels of CAR, LPC, LPE, PA, PC, PE, PI, PS, SM, and ST [139] | Brain | 10.1186/s13195-019-0558-0 |
| Apoe-KO mice |
• Both APOE3 and APOE4 treatment reduces hyperlipidemia in a dose-dependent manner, lowering both plasma cholesterol and lipoprotein levels [307] • Expression of APOE4 increases VLDL-cholesterol and reduces HDL-cholesterol levels relative to apoE3 [307] |
Plasma | 10.1161/atvbaha.112.301193 |
| Metabolomics | |||
| AD | • APOE4 is associated with reduced LysoPC(18:1), LysoPC(P-18:0), and Cardiolipin [308] | Plasma | 10.1016/j.jpba.2019.113088 |
| Normal; Young |
• APOE4 carriers show higher levels of cholesterol relative to APOE2 carriers [309] • APOE influences GlycA, isoleucine, LDL-TG, VLDL-TG, and M-VLDL (E2 < E3 < E4) [309] • APOE influences LDL particle diameter (E2 > E3 > E4) [309] |
Serum | 10.1038/s41598-018-36450-9 |
| Biomarkers | |||
| Normal |
• APOE4 is associated with increased LDL, IGF-1, SHBG, bilirubin, triphosphate, ApoB, and total cholesterol, and reduced HDL, HbA1C, lipoprotein A, CRP, GGT, vitamin D, creatine, urate, and urea compared to APOE3 [310] • APOE2 is associated with increased HDL, CRP, vitamin D, CysC, ApoA, creatinine, and alkaline phosphatase; and reduced LDL, IGF-1, bilirubin, and ApoB, compared to APOE3 [310] |
Serum blood | 10.3233/jad-200338 |
| Normal; Aging | • APOE2 reduces total cholesterol, LDL, lipoprotein A, and ApoB and increases apoA1 compared to APOE3 [311] | blood | 10.18632/aging.103405 |
| Normal | • APOE4 is associated with increased SNAP-25 in cognitively normal patients [312] | CSF | 10.1016/j.neurobiolaging.2021.02.008 |
| Normal; Aging |
• APOE2 carriers without dementia have reduced Aβ burden, with no differences in tau accumulation [313] • APOE4 carriers without dementia have increased Aβ burden and tau burden [313] |
PET | 10.1007/s00259-021-05192-8 |
| AD | • APOE4 carriers with preclinical AD have increased VILIP-1 [314] | CSF | 10.2147/ndt.s235395 |
| AD | • APOE4 is associated with increased levels of CDH6 and HAGH in AD patients [315] | Plasma | 10.1038/s41598-020-65038-5 |
| AD |
• Levels of CRP are influenced by APOE (E2/E3 > E3/E3 > E3/E4 > E4/E4 > E2/E4) [316] • Levels of ApoB are influenced by APOE (E2/E3 < E2/E4 < E3/E3 < E3/E4 < E4/E4) [316] • Levels of IL-13 are influenced by APOE (E2/E3 < E2/E4 < E3/E3 < E3/E4 < E4/E4) [316] • Levels of CXCL9 are influenced by APOE (E3/E3 > E3/E4 > E4/E4) [316] |
Plasma | 10.1001/archneurol.2012.1070 |
| APOE mice | • APOE4 is associated with increased NP1 levels [317] | Plasma | 10.1016/j.nbd.2018.02.014 |
Abbreviations: pTau: phosphorylated Tau, tTau: total Tau, DAM: disease-associated microglia, MGnD: microglial neurodegenerative phenotype, ARM: activated response microglia, AIF1: Allograft inflammatory factor 1, APP: Amyloid precursor protein, GDI2: Guanosine Diphosphate Dissociation Inhibitor 2, HSP90AA1: Heat Shock Protein 90 Alpha Family Class A Member 1, METAP2: Methionyl Aminopeptidase 2, NACA: Nascent Polypeptide Associated Complex Subunit Alpha, NCK1: Non-catalytic region of tyrosine kinase adaptor protein 1, PRDX1: Peroxiredoxin 1, RPS27A: Ribosomal Protein S27a, SFTPD: Surfactant Protein D, UFC1: Ubiquitin-Fold Modifier Conjugating Enzyme 1, PSD95: Postsynaptic density protein 95, AsymAD: asymptomatic AD, LysoPE: Lysophosphatidylethanolamine, PE: Phosphatidylethanolamine, LDL: Low-density lipoprotein, CAR: Carnitine, LPC: Lysophosphatidylcholine, LPE: Lysophosphatidylethanolamine, PA: Phosphatidic acid, PC: Phosphatidylcholine, PI: Phosphatidylinositol, PS: Phosphatidylserine, SM: Sphingomyelin, ST: Sterol, VLDL: Very low-density lipoprotein, HDL: High-density lipoprotein, LysoPC: Lysophosphatidylcholine, GlycA: Glycoprotein acetylation, TG: Triglyceride, IGF-1: Insulin-like growth factor-1, SHBG: Sex hormone binding globulin, HbA1C: Hemoglobin A1C, CRP: C-reactive protein, GGT: Gamma-glutamyl transferase, CysC: Cystatin C, ApoA: Apolipoprotein A, ApoB: Apolipoprotein B, VILIP-1: Visinin-like protein 1, CDH6: Cadherin 6, HAGH: Hydroxyacylglutathione Hydrolase, IL-13: Interleukin-13, CXCL9: Chemokine ligand 9, NP1: Neuronal pentraxin 1
Collectively, these studies suggest that not only does APOE4 increase the risk of developing various non-amyloid neuropathologies, but this risk may be synergistically heightened by altered immunomodulation and inhibited ability to respond to neuronal injuries on a synaptic level to maintain proper signaling networks. Future research on the complex interplay between apoE, various neuropathologies, immunomodulation, and synaptic function will help to shed light on the pathogenic mechanisms of AD and other neurodegenerative diseases. Although much of the therapeutic research in LOAD has been geared toward targeting Aβ and tau pathologies, the role of apoE as a common mediator of several amyloid-dependent and amyloid-independent pathways suggests that apoE itself could be a powerful target upstream of multiple AD pathologies.
Cell-type specific functions
Studies using iPSC-derived cellular models, animal models, and human brains have revealed cell-type and isoform specific functions of apoE. Using iPSC-derived isogenic lines for APOE, it was shown that APOE4 is associated with altered transcriptomic profiles related to synaptic function in neurons compared to APOE3. In astrocytes, apoE4 expression results in intracellular cholesterol accumulation, as well as impairment of Aβ clearance [62]. Another study also showed that apoE4 is associated with impaired lipid and fatty acid metabolism by disrupting neuron-astrocyte coupling. Fatty acid accumulation in neurons leads to disruption in fatty acid oxidation and lipid dysregulation in astrocytes. Compromised lipid metabolism in astrocytes can then affect several downstream pathways, including Aβ clearance [178]. Astrocytic apoE expression has also been shown to affect Aβ clearance and deposition in vivo [85]. In addition, microglia-like iPSC-derived cells expressing APOE4 exhibit transcriptomic changes associated with immune response, resulting in morphological alterations that correlate with diminished phagocytosis of Aβ, amongst several other downstream effects [62]. Whether apoE isoforms differentially modulate lipid metabolism and neuroinflammation in a cell type-dependent manner warrants further investigation. Interestingly, a very recent study showed that APOE4 drives lipid metabolic dysregulation in astrocytes and microglia that may contribute to increased AD risk [179]. Together, these data demonstrate that restoring apoE4-mediated dysregulation of Aβ and lipid metabolism in astrocytes and microglia could be a cell-type driven therapeutic avenue.
A genotype-dependent role for APOE has also been reported in VMCs, including pericytes. VMC-expressed apoE has been shown to differentially regulate neurobehaviors, gliovascular functions, and transcriptomic profiling depending on isoforms [146]. Transcriptomic profiles in human pericytes isolated from the prefrontal cortex and the hippocampus show an upregulation of apoE4 both at the transcript and at the protein levels compared to apoE3 [67]. Similarly, in a human iPSC-derived BBB model, apoE4-expressing VMCs with pericyte-like properties display elevated apoE levels compared to those expressing apoE3, which could be in part responsible for amyloid accumulation in the vasculature. Thus, APOE is differentially regulated depending on the cell-type it is expressed by, supporting the consideration of cell-type specific targeting of apoE [67].
A study using single-nucleus RNA-sequencing of 48 prefrontal cortex tissues brought to light several new findings relating to both sex-dependent and cell type-dependent mechanisms in AD pathophysiology. Cell-type specific changes mostly occurred in the early stages of AD pathogenesis, with disease-related cell population changes occurring predominantly in females. This study also identified disease-driven changes in myelination-related pathways, pointing to a major role for myelination in AD [64]. Understanding how apoE isoforms modulate myelination-related pathways and transcriptomic changes in different cell types during disease progression may further shed light on the pathogenic mechanisms of AD.
Rare apoE variants associated with AD risk
The APOE3-Christchurch (APOE3-Ch) variant has sparked interest in the AD field since its homozygosity was linked to remarkable resistance against an aggressive form of familial AD driven by the PSEN1-E280A mutation in a single case report [15]. Carrying two copies of this rare allele significantly reduced tau pathology and neurodegeneration in the patient, with preserved glucose metabolism and cognition. The effect on amyloid pathology is unclear since high amyloid burden was still detected by amyloid PET [15]. APOE3-Ch was discovered in 1987 as a susceptibility factor for type III hyperlipoproteinemia in Caucasian populations [180]. It encodes a missense mutation on an APOE3 background which results in an Arg to Ser substitution at position 136, within the receptor binding region of apoE [180, 181] (Fig. 1). In vitro studies using bacterially produced apoE3-Ch have suggested that the R136S mutation leads to reduced binding to the LDL receptor and heparin [15, 181]. How these differences in biochemical properties result in protection against AD is still unclear. It is possible that deficient interaction between apoE3-Ch and HSPG may directly or indirectly impact AD pathologies in a positive manner.
Other rare APOE variants have since been linked to either AD risk or AD protection. The APOE3-Jacksonville (APOE3-Jac) mutation on an APOE3 backbone results in a Val to Glu substitution at position 236 within the C-terminal domain (Fig. 1). APOE3-Jac was linked to reduced risk for AD and dementia with Lewy bodies (DLB) [17, 18]. A recent study showed that apoE3-Jac promotes healthy brain aging and decreases amyloid deposition and associated toxicity by reducing apoE self-association and increasing lipidation [17]. The C-terminal region of apoE is involved in its oligomerization in solution and it also contains a lipid binding region. Given that lipidation of apoE is favored when the protein is in its monomeric form [84], it is possible that the reduced hydrophobicity and/or conformational change in apoE3 caused by the V236E substitution results in its reduced oligomerization and increased lipidation capacity, contributing to its protective role in AD. Other mutations in APOE, such as L28P in an APOE4 backbone (APOE4-Freiburg) or APOE3-R145C (APOE3-Philladelphia) (Fig. 1) have been linked to increase risk for lipid disorders and cardiovascular diseases [182, 183]. However, genetic association of these rare variants to LOAD have not been robustly studied yet [18]. In a more recent report, a new APOE protective variant (APOE4-R251G) was discovered upon analyses of multiple disease cohorts [19]. Interestingly, this mutation is on an APOE4 backbone with a Arg to Gly substitution at position 251 within the C-terminal domain of apoE. How this APOE4-R251G variant protects against AD risk is not clear although changes in apoE structure and/or lipidation capability are among the possibilities. The location of the protective APOE3-Jac and APOE4-R251G variants within the carboxyl-terminal portion of apoE suggests that this region of apoE plays an important role in apoE biology and pathobiology. In summary, understanding how mutations in apoE affect its structure, biochemical properties, and function, and how this may translate in differential effects on AD pathobiology, would allow for the identification of new therapeutic avenues and personalized medicine.
ApoE-Targeted therapeutic strategies
Removal of amyloid plaques is a promising therapeutic avenue in AD and efforts have led to the development of aducanumab, a newly FDA-approved treatment that targets amyloid aggregates [184]. A recent encouraging clinical trial result reported that another anti-Aβ drug, lecanemab, could potentially slow the cognitive decline in people with early onset of AD by 27% over 18 months. However, the long-term safety and effectiveness of this new therapy still need to be explored. Whether APOE genotype further modifies the safety and efficacy of this drug will also require further investigation. It is well-known that anti-amyloid immunotherapy can increase the incidence of amyloid-related imaging abnormalities (ARIA) with brain edema or hemorrhage [185]. Interestingly, APOE4 is greatly associated with ARIA and exhibits a gene dose effect; thus, obtaining APOE genotype status has been recommended to be a prerequisite for an AD therapy to better inform ARIA risk, treatment plan, and clinician vigilance [186, 187]. In addition, since APOE genotype has been shown as a key determinant of AD risk impacting multiple disease pathways, apoE-targeted therapy has become an attractive avenue for consideration of novel therapies. Here, we outline and discuss the major apoE-related properties and mechanisms being examined as potential apoE-targeted therapeutic strategies.
Modulating the levels of apoE
Characterization of apoE isoforms in animal models and clinical studies has shown different protein levels among them [188, 189], which appears to be independent of Aβ levels or AD diagnosis [190]. Since apoE plays a major role in Aβ plaque formation [71, 85], several strategies have focused on modulation of apoE levels to prevent or reduce pathological development of AD. ApoE haploinsufficiency was shown to reduce amyloid deposition in amyloid model mice [191]. Furthermore, astrocyte-specific deletion of the Apoe gene improves cognitive performance [192] and reduces Aβ deposition and apoE accumulation in the brain of AD mouse models [105]. Thus, the dependence of Aβ on apoE levels has given rise to several strategies aimed toward altering apoE levels for prophylactic and therapeutic benefits.
One promising apoE-focused therapeutic avenue is the use of immunotherapies to reduce apoE, in particular apoE4, and consequently alleviate Aβ deposition (Fig. 2). In vivo experiments showed that intraperitoneal injection of HJ6.3, a monoclonal antibody specific against apoE, is effective in reducing amyloid deposition by modulating microglial responses and inflammatory cytokine levels [193]. Administration of HJ6.3 to amyloid model mice reduces Aβ pathology, improves spatial learning performance, and restores functional connectivity without altering plasma cholesterol levels when administered both prior to and after plaque onset [194]. These data demonstrate that reducing apoE4 level may be beneficial in attenuating AD pathology.
Fig. 2.
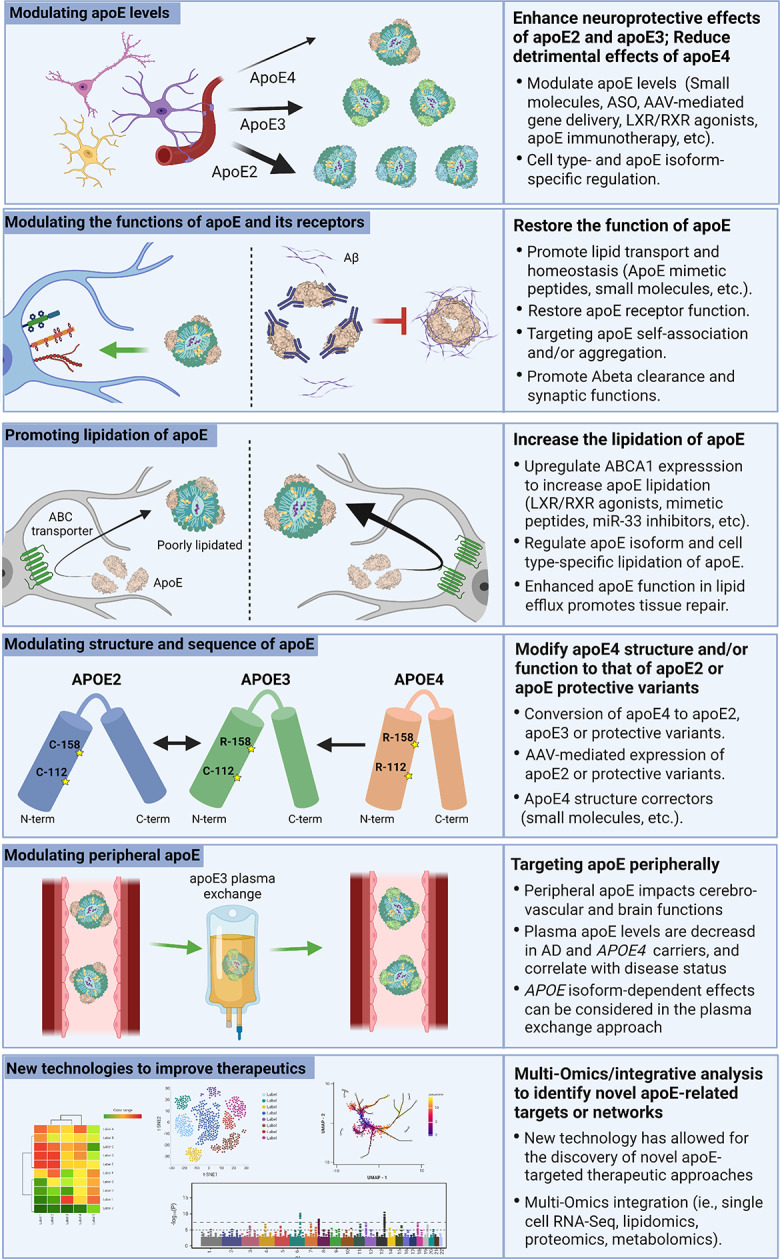
ApoE-targeted therapeutic strategies for AD. One avenue of AD therapy is modulating apoE expression from various cell types. This can be achieved through LXR/RXR agonists which increase apoE levels and lipidation. LXR/RXRs are upstream regulators of apoE expression making them a suitable target for modulating apoE levels. Targeting apoE should also consider the isoform- and cell type-specific effects. Another apoE-targeted therapeutic strategy is the use of small molecules that modulate apoE functions. These include peptides designed to mimic the binding site for apoE on LDL and HDL, which has been shown to increase apoE lipidation and secretion. Additionally, mimetic peptides can increase the function of apoE receptors to improve cholesterol transport. Small molecules or immunotherapies that prevent apoE self-association and/or aggregation may increase the lipid carrying capacity of apoE and reduce Aβ seeding. Similarly, modulating the lipidation of apoE has become an interesting target. Mimetic peptides can be used to increase the activity of ABCA1, which increases the lipidation of apoE4 and improves cognitive function. This can also be achieved through anti-sense oligonucleotide (ASO) inhibition of miR-33. Another promising therapeutic avenue is structural modification of apoE through genetic manipulation or small molecules. The CRISPR/Cas9 system has the potential to directly convert APOE4 to APOE3 or APOE2. This may also be achieved through an AAV system to induce apoE2 expression. A similar approach without genetic manipulation would be small molecule inhibitors to reduce interdomain interactions and structurally modify apoE to alter its function. Lastly, targeting peripheral apoE may be an alternative avenue for AD therapy. For example, plasma exchange by infusing APOE3 young plasma in APOE4 carriers is currently being tested in clinical trials to determine the beneficial effects of young plasma and the isoform-dependent effects. While these various therapeutic approaches have shown some promise in preclinical and clinical settings, they have yet to make a significant impact on the overall prognosis of AD. Research continues to seek alternative approaches to refine the current therapeutic strategies. Presently, many new technologies are being employed to discover new targets and networks such as transcriptomics, proteomics, lipidomics, and metabolomics. These multi-omics and integrative analysis may help better inform future apoE-related disease modifying therapy for AD.
Peptide-based interventions for apoE
Insights into the structure of apoE and its biochemical interactions have opened the door to several mechanisms by which the function of apoE could be modulated. One such mechanism is to inhibit apoE receptor binding through mimetic peptides, short peptide sequences designed to compete for apoE binding sites and thereby reduce its function [24, 195] (Fig. 2). For example, peptides encompassing the apoE receptor binding region (amino acids 130–150) have been shown to reduce microglial immunoreactivity in vitro [196], and improve survival in mouse models of traumatic brain injury [197, 198] and AD [199]. On the other hand, peptide mimetics have also been used to promote apoE function. In one approach, peptides were designed to mimic HDL to bind apoE, thereby increasing apoE secretion and enhancing lipidation status to negate the impacts on Aβ metabolism [200]. These preclinical data support the idea that mimetic peptides might be a useful strategy for therapeutics targeting of apoE function.
Given the role of apoE in amyloid deposition, inhibiting the interaction between apoE and Aβ peptides is another potential strategy for preventing AD pathogenesis (Fig. 2). Peptides binding to residues 12–28 of apoE have been shown to inhibit this interaction [201] and effectively reduce Aβ pathology and synaptic loss in mouse models of AD [202]. Further upstream in the amyloid processing pathway, peptides that prevent apoE binding to APP have been shown to reduce Aβ and tau pathology, improve cognitive performance, and have no effects on lipid profiles in a mouse model [203]. Thus, preventing amyloid deposition by modulating the function of apoE with mimetic peptides demonstrates a promising therapeutic avenue in preclinical studies.
ApoE self-association and aggregation
ApoE protein is deposited within Aβ plaques and CAA. ApoE4 is believed to have a higher propensity to oligomerize, thus reducing its lipidation and capacity for Aβ clearance as well as facilitating Aβ aggregation [20, 39, 46]. A previous study showed that apoE4 potently increases amyloid deposition by promoting Aβ seeding [85], suggesting that apoE4 may nucleate Aβ aggregation or plaque formation through its self-aggregating propensity. Importantly, both central [chronic intracerebroventricular (i.c.v.) infusion] and peripheral [intraperitoneal (i.p.) injection] administration of an anti-human apoE antibody (HAE-4), which selectively targets poorly-lipidated, aggregated forms of apoE present in the plaque, has been shown to remove Aβ plaques via a microglial-mediated clearance mechanism [204]. Of note, i.p. injection of HAE-4 antibody had a slightly greater effect on reducing Aβ levels than those with i.c.v. infusion likely due to relatively higher HAE-4 antibody levels remaining in the brain after 6 weeks of treatment in the i.p. injection paradigm. Of note, HAE-4 treatment can reduce CAA and Aβ parenchymal plaques after the onset of plaque deposition without inducing ARIA, the adverse vascular effects, and inhibit Aβ-associated tau seeding and spreading [205, 206]. Furthermore, a recent study showed that expression of apoE3-Jac variant which exhibits a decreased self-association reduces the risk of AD and amyloid deposition [17]. These findings suggest that targeting aggregated pools of apoE and/or apoE self-aggregation might be an ideal therapeutic strategy for AD.
ApoE lipidation status and lipid metabolism
Given that lipidated apoE is more effective in supporting brain homeostasis and injury repair, as well as mediating Aβ clearance compared to non-lipidated apoE, another therapeutic strategy is to alter the physiological balance between lipidated and non-lipidated apoE (Fig. 2). Peptide mimetics used to upregulate ABCA1 have been shown to increase apoE4 lipidation, and reduce Aβ, tau, and cognitive deficits in a mouse model [207]. Similar results may be accomplished by reducing apoE aggregation to increase its ability to accept lipids [204]. The LXR and retinoid X receptors (RXRs) family are known to promote apoE lipidation and expression [208]. Oral administration of bexarotene, an LXR/RXR agonist and FDA-approved anti-cancer agent, resulted in enhanced clearance of Aβ and improved cognitive performance in animal models [209]. However, Phase 1B studies with APOE3 individuals showed that bexarotene failed to increase CNS apoE or alter Aβ levels, likely due to poor BBB permeability [210]. A different cohort of AD patients, which included diverse APOE genotypes, showed that bexarotene treatment did reduce Aβ deposits in several brain areas. However, there were no signs of cognitive improvements and elevated plasma triglycerides as well as liver toxicity were some of the negative consequences observed in the clinical trial [211]. Although the clinical trials for bexarotene in the treatment of AD showed disappointing results, the studies on LXR/RXR agonists highlighted the importance of considering APOE genotype in modulating the response to clinical trials and identifying potential therapeutic targets. In addition, administration of choline, a soluble phospholipid precursor, is shown to restore the defective lipid homeostasis in APOE4 iPSC-derived astrocytes [133]. These studies suggest that modulating the lipidation status of apoE and/or lipid metabolism in the brain may serve as a therapeutic approach to alleviate the lipid dysregulation associated with apoE4.
ApoE structural correctors
Due to the amino acid substitutions at positions 112 and 158, the amino- and carboxyl-termini of apoE4 have an interdomain interaction which makes the protein more compact compared to apoE2 and apoE3. It has been suggested that this interdomain interaction contributes to the pathogenic effects of apoE4 [212]. Thus, small molecules have been proposed as a method to target and inhibit interdomain interactions [213], essentially acting as a structural corrector of apoE4 (Fig. 2). Human iPSC studies suggest that phthalazinone derivatives may inhibit these domain interactions and thus ameliorate the toxic effects of apoE4 in neurons [214, 215]. This suggests that structural modification of apoE4 may be an alternative approach to reduce the toxic effects of apoE4. Further research on the structure of apoE and its receptors will yield greater mechanistic insight toward this goal.
Gene therapies targeting APOE
The CRISPR/Cas9 genome-editing system has been explored as a therapeutic avenue for several genetically inherited disorders and is currently being tested in clinical trials to treat certain types of cancers [216, 217]. One intriguing idea is that CRISPR/Cas9 could be used to convert the APOE genotype of APOE4 carriers to APOE3 or APOE2, thus ameliorating the toxic effects of apoE4 and conferring the protective benefits of other isoforms (Fig. 2). In human-derived iPSCs, CRISPR/Cas9 was effective in altering the genome to produce isogenic lines homozygous for all three major APOE alleles [218]. In iPSC-derived neurons, CRISPR/Cas9 correction of APOE4/4 to an APOE3/3 genotype reduces susceptibility to cytotoxicity, tau secretion, and tau phosphorylation [101]. Experiments in iPSC-derived organoids show that converting APOE4 to APOE3 seems to attenuate Aβ pathology by improving astrocytic and microglial clearance of Aβ [62]. Although there are challenges with using CRISPR/Cas9 in vivo, including neuroanatomical specificity, potential off-target effects, and timing of treatment, its potential as a precise and efficacious therapeutic is unlimited.
A viral delivery approach, such as AAV, is another method that has been used to safely and accurately introduce recombinant genes into a host genome [219] and has proven safe and effective for several genetic disorders [220, 221]. Harnessing the protective effects of apoE2 by using AAVs could negate the toxic effects of apoE4. Experiments in mouse models show that transduction with AAV vectors encoding the three apoE isoforms differentially alters Aβ deposition and clearance in an isoform-dependent manner [222]. Furthermore, viral mediated overexpression of apoE2 in apoE4-TR mice enhances apoE lipidation and associated cholesterol, whereas overexpression of apoE4 has an opposite effect [223]. These results suggest that introducing apoE2 in APOE4 carriers could be beneficial to treating AD. However, AAV-mediated intracerebroventricular delivery of apoE appears to have no effect on tau burden in models of tauopathy [224]. Although there are certainly hesitations regarding CNS delivery of AAVs in humans, work in non-human primates has proven this method to be safe and effective in promoting widespread expression of apoE2 [225]. Furthermore, a phase 1 clinical trial is currently recruiting patients to evaluate the use of intracisternal administration of AAV-mediated expression of APOE2 in APOE4 homozygous patients (NCT03634007) with expected completion by September 2024. The efficacy of treating APOE4 homozygotes with expression of apoE2 may be dependent on the protective effects of apoE2 being able to compensate for the toxic effects of apoE4 which warrants further investigation.
MicroRNAs
Micro-RNAs (miRNAs) have emerged as important contributors to several pathogenic processes in AD [226], as well as potential biomarkers for disease progression in LOAD [227]. Research has identified miRNAs that contribute to Aβ-induced neurotoxicity [228], synaptic dysfunction [229], immune reactions [230], and tau pathogenesis [231]. One such example is miRNA-33, which has been identified as a key modulator of ABCA1, apoE levels, and subsequently of Aβ metabolism in AD [232]. Targeting of miRNA-33 via ASO results in increased ABCA1 levels, elevated apoE lipidation, and decreased Aβ levels in mouse models [233]. Not only can miRNAs alter apoE function through a variety of mechanisms, but some miRNAs appear to be expressed differentially based on APOE genotype. For instance, miRNA-146a is involved in inhibitory feedback for the brain’s immune responses and is reduced in the brain and plasma of APOE4 mice, thus exacerbating innate immune responses [234]. Further research on miRNAs related to apoE and AD pathogenesis is needed to reveal insights into this mechanistic relationship and the transcriptional networks involved in AD development and progression.
Targeting the periphery
In addition to the role of apoE in the CNS, the contributions of peripheral apoE have also become a topic of interest. Although there are indeed separate pools of apoE in the CNS and periphery due to BBB impermeability [24, 53, 235], research suggests that peripheral apoE can influence cognitive function [236]. In Apoe-deficient animal models, restoration of peripheral, but not CNS apoE reduced synaptic loss and improved cognitive function [237]. However, it seems that reducing the levels of hepatic apoE in the periphery did not influence Aβ pathology in a mouse model in which apoE is present in the CNS [238]. A recent study showed that liver-expressed apoE4 compromises synaptic plasticity and cognitive behaviors likely by impairing cerebrovascular functions [160]. In addition, exposure of young apoE3 plasma ameliorates aging-related BBB damage and promotes endothelial barrier integrity, shedding light on the therapeutic potential of young plasma based on APOE genotype. In humans, these effects are also shown to be isoform-specific, perhaps due to APOE4 carriers having reduced levels of plasma apoE than APOE3 or APOE2 individuals [239]. It has been shown that low plasma apoE levels seen in APOE4 carriers are associated with increased plasma glucose levels, which negatively impacts cerebral glucose metabolism [240]. In addition, researchers have found differences in peripheral extracellular vesicles (pEVs), which may function to promote crosstalk between the brain and periphery. A study reports that pEVs from APOE4 carriers have reduced neurotrophic and inflammatory markers, and these changes may predict AD five years before symptom onset [241]. Interestingly, apoE deficiency can lead to accumulation of pyrrolated serum albumin, which has a higher binding affinity for apoE3 compared to apoE2 and apoE4 and increases the immune response [242]. Though the mechanism remains unclear, these data suggest a relationship between peripheral apoE expression and inflammatory signaling that is isoform-dependent.
Thus, interventions harnessing the protective function of peripheral apoE in an isoform-specific manner are appealing options with numerous potential mechanisms of action (Fig. 2). A clinical trial targeting the periphery with promising results is the Alzheimer Management by Albumin Replacement (AMBAR) study, where patients undergo plasmapheresis with albumin replacement [243]. However, this study has not considered the influence of age, sex, or APOE genotype [244]. To further investigate the potential benefits of peripheral apoE, clinical trials are currently exploring the effects of introducing plasma from young donors to cognitively impaired and AD patients in an isoform-specific manner (NCT02256306, NCT03887741). While these clinical trials are still in early stages, the safety, tolerability, and feasibility appear promising [245]. Alternative approaches could be used to alter levels of apoE in the periphery via genetic interventions, or with the use of structural modifiers to modulate the function of peripheral apoE in an isoform-specific fashion as discussed in earlier sections.
Lifestyle changes
Exercise
Lifestyle changes, such as physical exercise and diets, can potentially attenuate AD pathology. Physical activity has been shown to reduce Aβ accumulation, improve cholesterol levels, reduce neuroinflammation, and enhance cognitive function [246]. Despite some contradictory evidence in the field, it is becoming increasingly apparent that the beneficial effects of exercise and diet are likely dependent on APOE genotype [247]. Following the assessment of physical activity and cognitive function in 806 participants over 6 years, a recent study showed that an active lifestyle has a favorable correlation with cognitive performance in APOE4 non-carriers [248]. Though some studies suggest the effects of the APOE4 allele are mixed and need to be further characterized [249], many reports provide evidence that APOE4 carriers are more responsive to exercise interventions compared to non-carriers [250, 251]. Additionally, recent studies have shown that APOE4 carriers who participated in aerobic exercise show increased hippocampal blood flow, improved verbal memory performance, improved neuropsychiatric symptoms and physical mobility compared to non-carriers and sedentary APOE4 carriers [252, 253]. Another study shows that physical activity was able to reduce functional connectivity, preserve brain structure, and reduce anxiety levels in APOE4 individuals [254]. It has been hypothesized that physical exercise and APOE genotype impact amyloid clearance and the proteasome system through epigenetic mechanisms [255]. Interestingly, exercise has been shown to increase plasma antioxidant capability and influence Aβ accumulation in APOE4 carriers [255]. Although the mechanism by which specific exercise paradigms benefit APOE4 carriers has yet to be established, it remains a valuable therapeutic strategy for reducing or delaying AD symptoms and pathology (Fig. 3).
Fig. 3.

Lifestyle changes can influence the pathogenesis of AD. ApoE4 has been shown to increase Aβ and tau aggregation, inflammation, and lipid dysregulation while reducing glucose metabolism, microbiome diversity, and BBB integrity. Healthy lifestyle changes have been suggested to benefit cognitive function and ameliorate AD pathology even in the presence of APOE4. Studies have demonstrated that ketogenic and Mediterranean diets as well as dietary supplements such as DHA can improve clinical outcomes. Along with diet, exercise has been shown to improve AD prognosis in apoE4 carriers. Chronic sleep disturbance appears to accelerate Aβ and tau pathology and exacerbate cognitive symptoms. The influence of APOE4 on sleep quality may lead to sleep disturbances in people at increased risk for dementia. Thus, improving sleep quality could reduce AD pathology and attenuate the negative impact of APOE4 on AD risk. The communication between gut microbiome and the brain, the microbiota-gut-brain axis, plays an important role in modulating AD pathology. ApoE isoforms have been shown to differentially modulate microbiome diversity. Evidence supports the use of sesamol to reshape the gut microbiome and prevent systemic inflammation. Thus, understanding the link between AD, apoE, and gut microbiota modulated through dietary approaches may offer avenues for identifying novel biomarkers and therapeutic strategies against AD
Diet
Preclinical and clinical studies over the past decade have suggested the ketogenic diet to be neuroprotective against AD by reducing Aβ toxicity, decreasing neuroinflammation, protecting against reactive oxygen, and regulating mitochondrial homeostasis [256, 257]. Since apoE is strongly associated with lipid and glucose metabolism, it is no surprise that APOE genotype affects preferences for cellular energy sources and modifies the response to diet [258]. The low-carbohydrate, high-fat ketogenic diet provides ketones as energy to mitigate the effects of impaired glucose metabolism in APOE4 carriers [259]. Recent case studies have shown that 10 weeks of the ketogenic diet was able to improve cognition along with reducing the blood levels of insulin, triglycerides, and glucose in APOE4 carriers [260, 261]. APOE4 carriers appear to have a delayed response to ketogenic therapy compared to non-carriers, though the outcomes were still overall positive [262]. A recent review highlights the importance of determining the APOE-dependent effects of ketogenic dietary modifications, as well as identifying effective doses of dietary compounds to improve clinical outcomes [263]. Multiple other dietary changes have been proposed to affect AD pathology in an APOE-dependent manner including the Mediterranean diet which consists of extra virgin olive oil, capers, red onions, cruciferous vegetables, and fatty fish. These components have been shown to increase LRP1 and ABCA1 levels, decrease NFκB and MMP9 activity, and increase brain docosahexaenoic acid (DHA) levels [264]. In sum, specific dietary changes may provide beneficial outcomes to those at-risk for developing cognitive impairments (Fig. 3).
The potential benefits of dietary supplements, such as DHA have also been extensively studied in the context of AD. It has been shown that APOE4 carriers have impaired metabolism of plasma DHA, an omega-3 fatty acid partly responsible for BBB integrity, which is associated with behavioral and cognitive impairments [265]. Preclinical and clinical studies of DHA supplementation have had success in rebalancing DHA levels and improving cognitive outcomes [266], although it seems to be ineffective in APOE4 carriers [267, 268]. Interestingly, it has been proposed that APOE4 carriers have impaired transport of free DHA into the brain, while increased amounts of DHA in phospholipid form (DHA-lyso-PC) from fish intake has led to positive benefits [269]. This idea has been corroborated by evidence suggesting that disrupted DHA absorption from plasma to CSF exists in APOE4 carriers and can reduce markers of AD pathology [270]. Further studies will be required to fully elucidate the effects of DHA supplementation on the cognitive function of individuals with different APOE genotype.
Sleep-wake cycle
Sleep disruptions and nightly restlessness have been a common complaint of AD patients for decades. Disturbed sleep has been shown to result in increased Aβ and tau aggregation, which ultimately leads to sleep disruptions [271]. The potential role of sleep disruption and specific oscillatory patterns are suggested to be useful in diagnosing and tracking the evolution of AD, and as a biomarker for cognitive decline [272]. Mounting evidence suggests sleep disturbances may be worsened or more common in patients who carry APOE4, as they have been shown to have reduced rapid eye movement (REM) sleep and increased fragmented sleep compared to non-carriers [273, 274]. Furthermore, APOE4 exacerbates objective sleep disturbances in individuals, which precedes subjective sleep complaints [275]. This may ultimately lead to longer total sleep duration in APOE4 carriers with cognitive decline [276, 277]. Importantly, it has been reported that for each sleep interruption, Aβ42 clearance is reduced by 5.4%, which is further reduced by the presence of an APOE4 allele [278]. Thus, understanding the molecular mechanisms by which apoE isoforms influence sleep/awake cycles may facilitate the development of therapeutic approaches to alleviate AD-related sleep disturbances (Fig. 3).
The observance of sleep disturbances in AD patients has led researchers to examine the role of melatonin in sleep regulation for relieving several AD-related symptoms. Melatonin has been shown to reduce Aβ toxicity [279], and reverse the pro-aggregatory and neurotoxic properties of apoE4 [280]. In addition, increased levels of melatonin contribute to cholesterol regulation, choline transport, neurogenesis, tau inhibition, insulin-regulation, calcium homeostasis, mitochondrial function, and can delay cellular senescence [281]. However, compared to age-matched controls, melatonin levels in the CSF are decreased in AD patients, and this reduction is exacerbated by the presence of APOE4 [282]. Thus, melatonin supplementation has been explored as a remedy for AD-related sleep disturbances. Recently, a meta-analysis of randomized control trials concluded that while melatonin improves total sleep time and sleep efficacy, cognitive function is not significantly altered [283]. Thus, the potential of melatonin for future treatments of AD merits continued research efforts [284, 285]. Recent studies corroborate the need for earlier intervention, and to consider APOE genotype when testing interventions targeting the sleep-wake cycle in AD patients [273, 275, 277, 286]. These findings suggest that APOE genotype may be one of the critical factors impacting the efficacy of melatonin-based therapies.
Gut microbiome
More recent evidence suggests an emerging role for the gut microbiome in modulating AD pathology through metabolism and immune function [287] (Fig. 3). Interestingly, work in transgenic mouse models demonstrated that gut microbiome species are modulated by both APOE genotype and gender [288–290]. Furthermore, it has been shown that AD patients have altered gut microbiota, which is associated with APOE genotype [291]. Research has also shown ways to modulate the gut microbiome in a beneficial way. In an APOE4 mouse model, insulin supplementation was shown to increase the levels of beneficial microbiota which increases metabolism and reduces neuroinflammation [292]. Additionally, sesamol has been shown to reshape the gut microbiome and prevent systemic inflammation in an APOE-dependent manner [293]. The effects of diet, microbiome, and inflammation on AD pathogenesis further support the notion of a critical communication between the brain and the periphery [294, 295].
Together, understanding the relationship between APOE genotype, exercise, diet, and microbiome is necessary for elucidating the underlying mechanisms that drive AD pathology, which may inform the search for robust biomarkers for successful interventions and therapeutics.
Conclusion
Mounting evidence confirms that apoE plays a key role in the pathogenesis of AD. ApoE4 conveys risk by a combination of gain of toxic functions and loss of protective functions, while apoE2 is protective against AD with overlapping and distinct mechanisms. ApoE impacts amyloid, tau, and additional neuropathologies, while also influencing neurodegeneration and the immune responses to those insults. The role of apoE as an upstream mediator in complex pathways underlying neurodegeneration and cognitive decline makes it an ideal therapeutic target for AD and related dementias. As we gain an enhanced understanding of the structure-function relationship of apoE, as well as the mechanistic relationships between apoE and neuropathologies, we will be able to use therapeutic tools to modulate apoE levels, structure, lipidation, oligomerization, and related outcomes to alter pathological progression. Further studies exploring systematic changes via multi-omics analysis, cell type-specific functions, and novel APOE variants may further yield critical insights in the future of AD therapeutic development. In addition, adopting new techniques for research and therapeutic delivery will prove vital in the development of novel treatment options for AD.
Acknowledgements
We thank Dr. Hongmei Li for proofreading of the manuscript.
Abbreviations
- AD
Alzheimer’s disease
- Aβ
Amyloid-β
- APOE
Apolipoprotein E
- LOAD
late-onset AD
- NFTs
neurofibrillary tangles
- eFAD
early-onset familial AD
- APP
amyloid precursor protein
- PSEN1
presenilin 1
- PSEN2
presenilin 2
- CNS
central nervous system
- NMR
nuclear magnetic resonance
- ABC
ATP-binding cassette
- LDLR
low-density lipoprotein receptor
- HSPG
heparan sulfate proteoglycans
- VLDL
very low-density lipoproteins
- HDL
high-density lipoproteins
- PET
positron emission tomography
- ASOs
antisense oligonucleotides
- iPSC
induced pluripotent stem cell
- AAV
adeno-associated virus
- FTD
Frontotemporal dementia
- DAM
disease-associated microglia
- MGnD
microglial neurodegenerative
- TREM2
triggering receptor expressed on myeloid cells 2
- VCID
impairment and dementia
- CAA
cerebrovascular amyloid angiopathy
- DLB
dementia with Lewy bodies.
Authors’ contributions
A.R., S.D. and Z.T. wrote the manuscript with significant input from T.I. and C.-C.L. A.R., S.D., T.I. and C.-C.L. prepared the figures and table. G.B., and C.-C.L. supervised the writing and edited the manuscript. All authors have read and approved the final manuscript.
Funding
Work in authors’ laboratories related to this topic has been supported by BrightFocus foundation and a NIH grant R01AG062110 (C.-C.L.), a NIH grant U19AG069701 (to G.B., and C.-C.L.), NIH grants R37AG027924, and RF1AG057181 (to G.B.), and Cure Alzheimer’s Fund (to G.B.).
Availability of data and materials
Not applicable.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
GB consults for SciNeuro and Lexeo, has consulted for AbbVie, E-Scape, Eisai, and Vida Ventures, and is on the scientific advisory board for Kisbee Therapeutics. All other authors declare no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Ana-Caroline Raulin and Sydney V. Doss contributed equally.
Contributor Information
Ana-Caroline Raulin, Email: Raulin.Ana-Caroline@mayo.edu.
Sydney V. Doss, Email: Doss.Sydney@mayo.edu
Zachary A. Trottier, Email: Trottier.Zach@gmail.com
Tadafumi C. Ikezu, Email: ikezu.tadafumi@mayo.edu
Guojun Bu, Email: Guojun.Bu@MolecularNeurodegeneration.org.
Chia-Chen Liu, Email: Liu.chiachen@mayo.edu.
References
- 1.Blennow K, de Leon MJ, Zetterberg H. Alzheimer’s disease. Lancet. 2006;368:387–403. doi: 10.1016/S0140-6736(06)69113-7. [DOI] [PubMed] [Google Scholar]
- 2.Kelley BJ, Petersen RC. Alzheimer’s disease and mild cognitive impairment. Neurol Clin. 2007;25:577–609. doi: 10.1016/j.ncl.2007.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Alexander GC, Emerson S, Kesselheim AS. Evaluation of Aducanumab for Alzheimer Disease: Scientific Evidence and Regulatory Review Involving Efficacy, Safety, and Futility. JAMA. 2021;325:1717–8. doi: 10.1001/jama.2021.3854. [DOI] [PubMed] [Google Scholar]
- 4.Gandy S, Knopman DS, Sano M. Talking points for physicians, patients and caregivers considering Aduhelm® infusion and the accelerated pathway for its approval by the FDA. Mol neurodegeneration. 2021;16:74. doi: 10.1186/s13024-021-00490-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bu G. Apolipoprotein E and its receptors in Alzheimer’s disease: pathways, pathogenesis and therapy. Nat Rev Neurosci. 2009;10:333–44. doi: 10.1038/nrn2620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Guo T, Zhang D, Zeng Y, Huang TY, Xu H, Zhao Y. Molecular and cellular mechanisms underlying the pathogenesis of Alzheimer’s disease. Mol neurodegeneration. 2020;15:40. doi: 10.1186/s13024-020-00391-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Serrano-Pozo A, Frosch MP, Masliah E, Hyman BT. Neuropathological alterations in Alzheimer disease. Cold Spring Harb Perspect Med. 2011;1:a006189. doi: 10.1101/cshperspect.a006189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.DeTure MA, Dickson DW. The neuropathological diagnosis of Alzheimer’s disease. Mol neurodegeneration. 2019;14:32. doi: 10.1186/s13024-019-0333-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Cacace R, Sleegers K, Van Broeckhoven C. Molecular genetics of early-onset Alzheimer’s disease revisited. Alzheimers Dement. 2016;12:733–48. doi: 10.1016/j.jalz.2016.01.012. [DOI] [PubMed] [Google Scholar]
- 10.Barber RC. The genetics of Alzheimer’s disease. Scientifica (Cairo) 2012;2012:246210. doi: 10.6064/2012/246210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Wightman DP, Jansen IE, Savage JE, Shadrin AA, Bahrami S, Holland D, Rongve A, Borte S, Winsvold BS, Drange OK, et al. A genome-wide association study with 1,126,563 individuals identifies new risk loci for Alzheimer’s disease. Nat Genet. 2021;53:1276–82. doi: 10.1038/s41588-021-00921-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.McDade E, Llibre-Guerra JJ, Holtzman DM, Morris JC, Bateman RJ. The informed road map to prevention of Alzheimer Disease: A call to arms. Mol neurodegeneration. 2021;16:49. doi: 10.1186/s13024-021-00467-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Strittmatter WJ, Saunders AM, Schmechel D, Pericak-Vance M, Enghild J, Salvesen GS, Roses AD. Apolipoprotein E: high-avidity binding to beta-amyloid and increased frequency of type 4 allele in late-onset familial Alzheimer disease. Proc Natl Acad Sci U S A. 1993;90:1977–81. doi: 10.1073/pnas.90.5.1977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Corder EH, Saunders AM, Risch NJ, Strittmatter WJ, Schmechel DE, Gaskell PC, Jr, Rimmler JB, Locke PA, Conneally PM, Schmader KE, et al. Protective effect of apolipoprotein E type 2 allele for late onset Alzheimer disease. Nat Genet. 1994;7:180–4. doi: 10.1038/ng0694-180. [DOI] [PubMed] [Google Scholar]
- 15.Arboleda-Velasquez JF, Lopera F, O’Hare M, Delgado-Tirado S, Marino C, Chmielewska N, Saez-Torres KL, Amarnani D, Schultz AP, Sperling RA, et al. Resistance to autosomal dominant Alzheimer’s disease in an APOE3 Christchurch homozygote: a case report. Nat Med. 2019;25:1680–3. doi: 10.1038/s41591-019-0611-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Sepulveda-Falla D, Sanchez JS, Almeida MC, Boassa D, Acosta-Uribe J, Vila-Castelar C, Ramirez-Gomez L, Baena A, Aguillon D, Villalba-Moreno ND, et al. Distinct tau neuropathology and cellular profiles of an APOE3 Christchurch homozygote protected against autosomal dominant Alzheimer’s dementia. Acta Neuropathol. 2022;144:589–601. doi: 10.1007/s00401-022-02467-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Liu CC, Murray ME, Li X, Zhao N, Wang N, Heckman MG, Shue F, Martens Y, Li Y, Raulin AC, et al. APOE3-Jacksonville (V236E) variant reduces self-aggregation and risk of dementia. Sci Transl Med. 2021;13:eabc9375. doi: 10.1126/scitranslmed.abc9375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Medway CW, Abdul-Hay S, Mims T, Ma L, Bisceglio G, Zou F, Pankratz S, Sando SB, Aasly JO, Barcikowska M, et al. ApoE variant p.V236E is associated with markedly reduced risk of Alzheimer’s disease. Mol neurodegeneration. 2014;9:11. doi: 10.1186/1750-1326-9-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Le Guen Y, Belloy ME, Grenier-Boley B, de Rojas I, Castillo-Morales A, Jansen I, Nicolas A, Bellenguez C, Dalmasso C, Kucukali F, et al. Association of Rare APOE Missense Variants V236E and R251G With Risk of Alzheimer Disease. JAMA Neurol. 2022;79:652–63. doi: 10.1001/jamaneurol.2022.1166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Liu CC, Liu CC, Kanekiyo T, Xu H, Bu G. Apolipoprotein E and Alzheimer disease: risk, mechanisms and therapy. Nat Rev Neurol. 2013;9:106–18. doi: 10.1038/nrneurol.2012.263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Goldberg TE, Huey ED, Devanand DP. Association of APOE e2 genotype with Alzheimer’s and non-Alzheimer’s neurodegenerative pathologies. Nat Commun. 2020;11:4727. doi: 10.1038/s41467-020-18198-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Li Z, Shue F, Zhao N, Shinohara M, Bu G. APOE2: protective mechanism and therapeutic implications for Alzheimer’s disease. Mol neurodegeneration. 2020;15:63. doi: 10.1186/s13024-020-00413-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Insel PS, Hansson O, Mattsson-Carlgren N. Association Between Apolipoprotein E ε2 vs ε4, Age, and β-Amyloid in Adults Without Cognitive Impairment. JAMA Neurol. 2021;78:229–35. doi: 10.1001/jamaneurol.2020.3780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Williams T, Borchelt DR, Chakrabarty P. Therapeutic approaches targeting Apolipoprotein E function in Alzheimer’s disease. Mol neurodegeneration. 2020;15:8. doi: 10.1186/s13024-020-0358-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Dickson DW, Heckman MG, Murray ME, Soto AI, Walton RL, Diehl NN, van Gerpen JA, Uitti RJ, Wszolek ZK, Ertekin-Taner N, et al. APOE ε4 is associated with severity of Lewy body pathology independent of Alzheimer pathology. Neurology. 2018;91:e1182–95. doi: 10.1212/WNL.0000000000006212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Outeiro TF, Koss DJ, Erskine D, Walker L, Kurzawa-Akanbi M, Burn D, Donaghy P, Morris C, Taylor JP, Thomas A, et al. Dementia with Lewy bodies: an update and outlook. Mol neurodegeneration. 2019;14:5. doi: 10.1186/s13024-019-0306-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.van Steenoven I, Koel-Simmelink MJA, Vergouw LJM, Tijms BM, Piersma SR, Pham TV, Bridel C, Ferri GL, Cocco C, Noli B, et al. Identification of novel cerebrospinal fluid biomarker candidates for dementia with Lewy bodies: a proteomic approach. Mol neurodegeneration. 2020;15:36. doi: 10.1186/s13024-020-00388-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Pankratz N, Byder L, Halter C, Rudolph A, Shults CW, Conneally PM, Foroud T, Nichols WC. Presence of an APOE4 allele results in significantly earlier onset of Parkinson’s disease and a higher risk with dementia. Mov disorders: official J Mov Disorder Soc. 2006;21:45–9. doi: 10.1002/mds.20663. [DOI] [PubMed] [Google Scholar]
- 29.Zhao N, Attrebi ON, Ren Y, Qiao W, Sonustun B, Martens YA, Meneses AD, Li F, Shue F, Zheng J, et al: APOE4 exacerbates α-synuclein pathology and related toxicity independent of amyloid. Science translational medicine 2020, 12. [DOI] [PMC free article] [PubMed]
- 30.Davis AA, Inman CE, Wargel ZM, Dube U, Freeberg BM, Galluppi A, Haines JN, Dhavale DD, Miller R, Choudhury FA, et al: APOE genotype regulates pathology and disease progression in synucleinopathy. Science translational medicine 2020, 12. [DOI] [PMC free article] [PubMed]
- 31.McGuinness B, Barrett SL, Craig D, Lawson J, Passmore AP. Attention deficits in Alzheimer’s disease and vascular dementia. J Neurol Neurosurg Psychiatry. 2010;81:157–9. doi: 10.1136/jnnp.2008.164483. [DOI] [PubMed] [Google Scholar]
- 32.Huang Y, Weisgraber KH, Mucke L, Mahley RW. Apolipoprotein E: diversity of cellular origins, structural and biophysical properties, and effects in Alzheimer’s disease. J Mol Neurosci. 2004;23:189–204. doi: 10.1385/JMN:23:3:189. [DOI] [PubMed] [Google Scholar]
- 33.Boyles JK, Pitas RE, Wilson E, Mahley RW, Taylor JM. Apolipoprotein E associated with astrocytic glia of the central nervous system and with nonmyelinating glia of the peripheral nervous system. J Clin Investig. 1985;76:1501–13. doi: 10.1172/JCI112130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Xu Q, Bernardo A, Walker D, Kanegawa T, Mahley RW, Huang Y. Profile and regulation of apolipoprotein E (ApoE) expression in the CNS in mice with targeting of green fluorescent protein gene to the ApoE locus. J neuroscience: official J Soc Neurosci. 2006;26:4985–94. doi: 10.1523/JNEUROSCI.5476-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Aoki K, Uchihara T, Sanjo N, Nakamura A, Ikeda K, Tsuchiya K, Wakayama Y. Increased expression of neuronal apolipoprotein E in human brain with cerebral infarction. Stroke. 2003;34:875–80. doi: 10.1161/01.STR.0000064320.73388.C6. [DOI] [PubMed] [Google Scholar]
- 36.Mahley RW, Huang Y. Apolipoprotein e sets the stage: response to injury triggers neuropathology. Neuron. 2012;76:871–85. doi: 10.1016/j.neuron.2012.11.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Grubman A, Chew G, Ouyang JF, Sun G, Choo XY, McLean C, Simmons RK, Buckberry S, Vargas-Landin DB, Poppe D, et al. A single-cell atlas of entorhinal cortex from individuals with Alzheimer’s disease reveals cell-type-specific gene expression regulation. Nat Neurosci. 2019;22:2087–97. doi: 10.1038/s41593-019-0539-4. [DOI] [PubMed] [Google Scholar]
- 38.Ruiz J, Kouiavskaia D, Migliorini M, Robinson S, Saenko EL, Gorlatova N, Li D, Lawrence D, Hyman BT, Weisgraber KH, Strickland DK. The apoE isoform binding properties of the VLDL receptor reveal marked differences from LRP and the LDL receptor. J Lipid Res. 2005;46:1721–31. doi: 10.1194/jlr.M500114-JLR200. [DOI] [PubMed] [Google Scholar]
- 39.Hatters DM, Zhong N, Rutenber E, Weisgraber KH. Amino-terminal domain stability mediates apolipoprotein E aggregation into neurotoxic fibrils. J Mol Biol. 2006;361:932–44. doi: 10.1016/j.jmb.2006.06.080. [DOI] [PubMed] [Google Scholar]
- 40.Yamazaki Y, Painter MM, Bu G, Kanekiyo T. Apolipoprotein E as a Therapeutic Target in Alzheimer’s Disease: A Review of Basic Research and Clinical Evidence. CNS Drugs. 2016;30:773–89. doi: 10.1007/s40263-016-0361-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ma Q, Zhao Z, Sagare AP, Wu Y, Wang M, Owens NC, Verghese PB, Herz J, Holtzman DM, Zlokovic BV. Blood-brain barrier-associated pericytes internalize and clear aggregated amyloid-β42 by LRP1-dependent apolipoprotein E isoform-specific mechanism. Mol neurodegeneration. 2018;13:57. doi: 10.1186/s13024-018-0286-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Chen J, Li Q, Wang J. Topology of human apolipoprotein E3 uniquely regulates its diverse biological functions. Proc Natl Acad Sci U S A. 2011;108:14813–8. doi: 10.1073/pnas.1106420108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Wilson C, Wardell MR, Weisgraber KH, Mahley RW, Agard DA. Three-dimensional structure of the LDL receptor-binding domain of human apolipoprotein E. Science. 1991;252:1817–22. doi: 10.1126/science.2063194. [DOI] [PubMed] [Google Scholar]
- 44.Weisgraber KH. Apolipoprotein E: structure-function relationships. Adv Protein Chem. 1994;45:249–302. doi: 10.1016/s0065-3233(08)60642-7. [DOI] [PubMed] [Google Scholar]
- 45.Hirsch-Reinshagen V, Zhou S, Burgess BL, Bernier L, McIsaac SA, Chan JY, Tansley GH, Cohn JS, Hayden MR, Wellington CL. Deficiency of ABCA1 impairs apolipoprotein E metabolism in brain. J Biol Chem. 2004;279:41197–207. doi: 10.1074/jbc.M407962200. [DOI] [PubMed] [Google Scholar]
- 46.Kanekiyo T, Xu H, Bu G. ApoE and Abeta in Alzheimer’s disease: accidental encounters or partners? Neuron. 2014;81:740–54. doi: 10.1016/j.neuron.2014.01.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Ji ZS, Dichek HL, Miranda RD, Mahley RW. Heparan sulfate proteoglycans participate in hepatic lipaseand apolipoprotein E-mediated binding and uptake of plasma lipoproteins, including high density lipoproteins. J Biol Chem. 1997;272:31285–92. doi: 10.1074/jbc.272.50.31285. [DOI] [PubMed] [Google Scholar]
- 48.Yamauchi Y, Deguchi N, Takagi C, Tanaka M, Dhanasekaran P, Nakano M, Handa T, Phillips MC, Lund-Katz S, Saito H. Role of the N- and C-terminal domains in binding of apolipoprotein E isoforms to heparan sulfate and dermatan sulfate: a surface plasmon resonance study. Biochemistry. 2008;47:6702–10. doi: 10.1021/bi8003999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Liu CC, Zhao N, Yamaguchi Y, Cirrito JR, Kanekiyo T, Holtzman DM, Bu G. Neuronal heparan sulfates promote amyloid pathology by modulating brain amyloid-beta clearance and aggregation in Alzheimer’s disease. Sci Transl Med. 2016;8:332ra344. doi: 10.1126/scitranslmed.aad3650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Liu CC, Hu J, Zhao N, Wang J, Wang N, Cirrito JR, Kanekiyo T, Holtzman DM, Bu G. Astrocytic LRP1 Mediates Brain Abeta Clearance and Impacts Amyloid Deposition. J neuroscience: official J Soc Neurosci. 2017;37:4023–31. doi: 10.1523/JNEUROSCI.3442-16.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Shi Y, Andhey PS, Ising C, Wang K, Snipes LL, Boyer K, Lawson S, Yamada K, Qin W, Manis M, et al. Overexpressing low-density lipoprotein receptor reduces tau-associated neurodegeneration in relation to apoE-linked mechanisms. Neuron. 2021;109:2413–26 e2417. doi: 10.1016/j.neuron.2021.05.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Rauch JN, Luna G, Guzman E, Audouard M, Challis C, Sibih YE, Leshuk C, Hernandez I, Wegmann S, Hyman BT, et al. LRP1 is a master regulator of tau uptake and spread. Nature. 2020;580:381–5. doi: 10.1038/s41586-020-2156-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Linton MF, Gish R, Hubl ST, Bütler E, Esquivel C, Bry WI, Boyles JK, Wardell MR, Young SG. Phenotypes of apolipoprotein B and apolipoprotein E after liver transplantation. J Clin Investig. 1991;88:270–81. doi: 10.1172/JCI115288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Weisgraber KH. Apolipoprotein E distribution among human plasma lipoproteins: role of the cysteine-arginine interchange at residue 112. J Lipid Res. 1990;31:1503–11. [PubMed] [Google Scholar]
- 55.Mahley RW, Weisgraber KH, Huang Y. Apolipoprotein E: structure determines function, from atherosclerosis to Alzheimer’s disease to AIDS. J Lipid Res. 2009;50(Suppl):183–8. doi: 10.1194/jlr.R800069-JLR200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Sun Y, Wu S, Bu G, Onifade MK, Patel SN, LaDu MJ, Fagan AM, Holtzman DM. Glial fibrillary acidic protein-apolipoprotein E (apoE) transgenic mice: astrocyte-specific expression and differing biological effects of astrocyte-secreted apoE3 and apoE4 lipoproteins. J neuroscience: official J Soc Neurosci. 1998;18:3261–72. doi: 10.1523/JNEUROSCI.18-09-03261.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Ji Y, Gong Y, Gan W, Beach T, Holtzman DM, Wisniewski T. Apolipoprotein E isoform-specific regulation of dendritic spine morphology in apolipoprotein E transgenic mice and Alzheimer’s disease patients. Neuroscience. 2003;122:305–15. doi: 10.1016/j.neuroscience.2003.08.007. [DOI] [PubMed] [Google Scholar]
- 58.Arendt T, Schindler C, Bruckner MK, Eschrich K, Bigl V, Zedlick D, Marcova L. Plastic neuronal remodeling is impaired in patients with Alzheimer’s disease carrying apolipoprotein epsilon 4 allele. J neuroscience: official J Soc Neurosci. 1997;17:516–29. doi: 10.1523/JNEUROSCI.17-02-00516.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Love S, Siew LK, Dawbarn D, Wilcock GK, Ben-Shlomo Y, Allen SJ. Premorbid effects of APOE on synaptic proteins in human temporal neocortex. Neurobiol Aging. 2006;27:797–803. doi: 10.1016/j.neurobiolaging.2005.04.008. [DOI] [PubMed] [Google Scholar]
- 60.Sweet RA, MacDonald ML, Kirkwood CM, Ding Y, Schempf T, Jones-Laughner J, Kofler J, Ikonomovic MD, Lopez OL, Garver ME, et al. Apolipoprotein E*4 (APOE*4) Genotype Is Associated with Altered Levels of Glutamate Signaling Proteins and Synaptic Coexpression Networks in the Prefrontal Cortex in Mild to Moderate Alzheimer Disease. Mol Cell Proteomics. 2016;15:2252–62. doi: 10.1074/mcp.M115.056580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kara E, Marks JD, Fan Z, Klickstein JA, Roe AD, Krogh KA, Wegmann S, Maesako M, Luo CC, Mylvaganam R, et al. Isoform- and cell type-specific structure of apolipoprotein E lipoparticles as revealed by a novel Forster resonance energy transfer assay. J Biol Chem. 2017;292:14720–9. doi: 10.1074/jbc.M117.784264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Lin YT, Seo J, Gao F, Feldman HM, Wen HL, Penney J, Cam HP, Gjoneska E, Raja WK, Cheng J, et al. APOE4 Causes Widespread Molecular and Cellular Alterations Associated with Alzheimer’s Disease Phenotypes in Human iPSC-Derived Brain Cell Types. Neuron. 2018;98:1141–54.e1147. doi: 10.1016/j.neuron.2018.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Kang SS, Ebbert MTW, Baker KE, Cook C, Wang X, Sens JP, Kocher JP, Petrucelli L, Fryer JD. Microglial translational profiling reveals a convergent APOE pathway from aging, amyloid, and tau. J Exp Med. 2018;215:2235–45. doi: 10.1084/jem.20180653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Mathys H, Davila-Velderrain J, Peng Z, Gao F, Mohammadi S, Young JZ, Menon M, He L, Abdurrob F, Jiang X, et al. Single-cell transcriptomic analysis of Alzheimer’s disease. Nature. 2019;570:332–7. doi: 10.1038/s41586-019-1195-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Lanfranco MF, Sepulveda J, Kopetsky G, Rebeck GW. Expression and secretion of apoE isoforms in astrocytes and microglia during inflammation. Glia. 2021;69:1478–93. doi: 10.1002/glia.23974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Yamazaki Y, Shinohara M, Yamazaki A, Ren Y, Asmann YW, Kanekiyo T, Bu G. ApoE (Apolipoprotein E) in Brain Pericytes Regulates Endothelial Function in an Isoform-Dependent Manner by Modulating Basement Membrane Components Arteriosclerosis, thrombosis, and vascular biology 2020, 40:128–144. [DOI] [PMC free article] [PubMed]
- 67.Blanchard JW, Bula M, Davila-Velderrain J, Akay LA, Zhu L, Frank A, Victor MB, Bonner JM, Mathys H, Lin YT, et al. Reconstruction of the human blood-brain barrier in vitro reveals a pathogenic mechanism of APOE4 in pericytes. Nat Med. 2020;26:952–63. doi: 10.1038/s41591-020-0886-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Stoll G, Meuller HW, Trapp BD, Griffin JW. Oligodendrocytes but not astrocytes express apolipoprotein E after injury of rat optic nerve. Glia. 1989;2:170–6. doi: 10.1002/glia.440020306. [DOI] [PubMed] [Google Scholar]
- 69.Nelissen K, Mulder M, Smets I, Timmermans S, Smeets K, Ameloot M, Hendriks JJ. Liver X receptors regulate cholesterol homeostasis in oligodendrocytes. J Neurosci Res. 2012;90:60–71. doi: 10.1002/jnr.22743. [DOI] [PubMed] [Google Scholar]
- 70.Namba Y, Tomonaga M, Kawasaki H, Otomo E, Ikeda K. Apolipoprotein E immunoreactivity in cerebral amyloid deposits and neurofibrillary tangles in Alzheimer’s disease and kuru plaque amyloid in Creutzfeldt-Jakob disease. Brain Res. 1991;541:163–6. doi: 10.1016/0006-8993(91)91092-f. [DOI] [PubMed] [Google Scholar]
- 71.Bales KR, Verina T, Dodel RC, Du Y, Altstiel L, Bender M, Hyslop P, Johnstone EM, Little SP, Cummins DJ, et al. Lack of apolipoprotein E dramatically reduces amyloid beta-peptide deposition. Nat Genet. 1997;17:263–4. doi: 10.1038/ng1197-263. [DOI] [PubMed] [Google Scholar]
- 72.Mahan TE, Wang C, Bao X, Choudhury A, Ulrich JD, Holtzman DM. Selective reduction of astrocyte apoE3 and apoE4 strongly reduces Aβ accumulation and plaque-related pathology in a mouse model of amyloidosis. Mol neurodegeneration. 2022;17:13. doi: 10.1186/s13024-022-00516-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Bales KR, Liu F, Wu S, Lin S, Koger D, DeLong C, Hansen JC, Sullivan PM, Paul SM. Human APOE isoform-dependent effects on brain beta-amyloid levels in PDAPP transgenic mice. J neuroscience: official J Soc Neurosci. 2009;29:6771–9. doi: 10.1523/JNEUROSCI.0887-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Holtzman DM, Fagan AM, Mackey B, Tenkova T, Sartorius L, Paul SM, Bales K, Ashe KH, Irizarry MC, Hyman BT. Apolipoprotein E facilitates neuritic and cerebrovascular plaque formation in an Alzheimer’s disease model. Ann Neurol. 2000;47:739–47. [PubMed] [Google Scholar]
- 75.Kok E, Haikonen S, Luoto T, Huhtala H, Goebeler S, Haapasalo H, Karhunen PJ. Apolipoprotein E-dependent accumulation of Alzheimer disease-related lesions begins in middle age. Ann Neurol. 2009;65:650–7. doi: 10.1002/ana.21696. [DOI] [PubMed] [Google Scholar]
- 76.Polvikoski T, Sulkava R, Haltia M, Kainulainen K, Vuorio A, Verkkoniemi A, Niinisto L, Halonen P, Kontula K. Apolipoprotein E, dementia, and cortical deposition of beta-amyloid protein. N Engl J Med. 1995;333:1242–7. doi: 10.1056/NEJM199511093331902. [DOI] [PubMed] [Google Scholar]
- 77.Schmechel DE, Saunders AM, Strittmatter WJ, Crain BJ, Hulette CM, Joo SH, Pericak-Vance MA, Goldgaber D, Roses AD. Increased amyloid beta-peptide deposition in cerebral cortex as a consequence of apolipoprotein E genotype in late-onset Alzheimer disease. Proc Natl Acad Sci U S A. 1993;90:9649–53. doi: 10.1073/pnas.90.20.9649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Kantarci K, Lowe V, Przybelski SA, Weigand SD, Senjem ML, Ivnik RJ, Preboske GM, Roberts R, Geda YE, Boeve BF, et al. APOE modifies the association between Abeta load and cognition in cognitively normal older adults. Neurology. 2012;78:232–40. doi: 10.1212/WNL.0b013e31824365ab. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Murphy KR, Landau SM, Choudhury KR, Hostage CA, Shpanskaya KS, Sair HI, Petrella JR, Wong TZ, Doraiswamy PM. Alzheimer’s Disease Neuroimaging I: Mapping the effects of ApoE4, age and cognitive status on 18F-florbetapir PET measured regional cortical patterns of beta-amyloid density and growth. NeuroImage. 2013;78:474–80. doi: 10.1016/j.neuroimage.2013.04.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Gonneaud J, Arenaza-Urquijo EM, Fouquet M, Perrotin A, Fradin S, de La Sayette V, Eustache F, Chetelat G. Relative effect of APOE epsilon4 on neuroimaging biomarker changes across the lifespan. Neurology. 2016;87:1696–703. doi: 10.1212/WNL.0000000000003234. [DOI] [PubMed] [Google Scholar]
- 81.Mishra S, Blazey TM, Holtzman DM, Cruchaga C, Su Y, Morris JC, Benzinger TLS, Gordon BA. Longitudinal brain imaging in preclinical Alzheimer disease: impact of APOE epsilon4 genotype. Brain. 2018;141:1828–39. doi: 10.1093/brain/awy103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Wang X, Huang W, Su L, Xing Y, Jessen F, Sun Y, Shu N, Han Y. Neuroimaging advances regarding subjective cognitive decline in preclinical Alzheimer’s disease. Mol neurodegeneration. 2020;15:55. doi: 10.1186/s13024-020-00395-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Serrano-Pozo A, Qian J, Monsell SE, Betensky RA, Hyman BT. APOEepsilon2 is associated with milder clinical and pathological Alzheimer disease. Ann Neurol. 2015;77:917–29. doi: 10.1002/ana.24369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Garai K, Verghese PB, Baban B, Holtzman DM, Frieden C. The binding of apolipoprotein E to oligomers and fibrils of amyloid-beta alters the kinetics of amyloid aggregation. Biochemistry. 2014;53:6323–31. doi: 10.1021/bi5008172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Liu CC, Zhao N, Fu Y, Wang N, Linares C, Tsai CW, Bu G. ApoE4 Accelerates Early Seeding of Amyloid Pathology. Neuron. 2017;96:1024–32 e1023. doi: 10.1016/j.neuron.2017.11.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Huynh TV, Liao F, Francis CM, Robinson GO, Serrano JR, Jiang H, Roh J, Finn MB, Sullivan PM, Esparza TJ, et al. Age-Dependent Effects of apoE Reduction Using Antisense Oligonucleotides in a Model of β-amyloidosis. Neuron. 2017;96:1013–23.e1014. doi: 10.1016/j.neuron.2017.11.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Lim YY, Mormino EC, Alzheimer’s Disease Neuroimaging I. APOE genotype and early beta-amyloid accumulation in older adults without dementia. Neurology. 2017;89:1028–34. doi: 10.1212/WNL.0000000000004336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Hashimoto T, Serrano-Pozo A, Hori Y, Adams KW, Takeda S, Banerji AO, Mitani A, Joyner D, Thyssen DH, Bacskai BJ, et al. Apolipoprotein E, especially apolipoprotein E4, increases the oligomerization of amyloid beta peptide. J neuroscience: official J Soc Neurosci. 2012;32:15181–92. doi: 10.1523/JNEUROSCI.1542-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.West T, Kirmess KM, Meyer MR, Holubasch MS, Knapik SS, Hu Y, Contois JH, Jackson EN, Harpstrite SE, Bateman RJ, et al. A blood-based diagnostic test incorporating plasma Aβ42/40 ratio, ApoE proteotype, and age accurately identifies brain amyloid status: findings from a multi cohort validity analysis. Mol neurodegeneration. 2021;16:30. doi: 10.1186/s13024-021-00451-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Fuentealba RA, Liu Q, Zhang J, Kanekiyo T, Hu X, Lee JM, LaDu MJ, Bu G. Low-density lipoprotein receptor-related protein 1 (LRP1) mediates neuronal Abeta42 uptake and lysosomal trafficking. PLoS ONE. 2010;5:e11884. doi: 10.1371/journal.pone.0011884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Kanekiyo T, Cirrito JR, Liu CC, Shinohara M, Li J, Schuler DR, Shinohara M, Holtzman DM, Bu G. Neuronal clearance of amyloid-beta by endocytic receptor LRP1. J neuroscience: official J Soc Neurosci. 2013;33:19276–83. doi: 10.1523/JNEUROSCI.3487-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Van Acker ZP, Bretou M, Annaert W. Endo-lysosomal dysregulations and late-onset Alzheimer’s disease: impact of genetic risk factors. Mol neurodegeneration. 2019;14:20. doi: 10.1186/s13024-019-0323-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Tai LM, Bilousova T, Jungbauer L, Roeske SK, Youmans KL, Yu C, Poon WW, Cornwell LB, Miller CA, Vinters HV, et al. Levels of soluble apolipoprotein E/amyloid-beta (Abeta) complex are reduced and oligomeric Abeta increased with APOE4 and Alzheimer disease in a transgenic mouse model and human samples. J Biol Chem. 2013;288:5914–26. doi: 10.1074/jbc.M112.442103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Tarasoff-Conway JM, Carare RO, Osorio RS, Glodzik L, Butler T, Fieremans E, Axel L, Rusinek H, Nicholson C, Zlokovic BV, et al. Clearance systems in the brain-implications for Alzheimer disease. Nat Rev Neurol. 2015;11:457–70. doi: 10.1038/nrneurol.2015.119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Verghese PB, Castellano JM, Garai K, Wang Y, Jiang H, Shah A, Bu G, Frieden C, Holtzman DM. ApoE influences amyloid-beta (Abeta) clearance despite minimal apoE/Abeta association in physiological conditions. Proc Natl Acad Sci U S A. 2013;110:E1807–16. doi: 10.1073/pnas.1220484110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Ries M, Sastre M. Mechanisms of Aβ Clearance and Degradation by Glial Cells. Front Aging Neurosci. 2016;8:160. doi: 10.3389/fnagi.2016.00160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Jiang Q, Lee CY, Mandrekar S, Wilkinson B, Cramer P, Zelcer N, Mann K, Lamb B, Willson TM, Collins JL, et al. ApoE promotes the proteolytic degradation of Abeta. Neuron. 2008;58:681–93. doi: 10.1016/j.neuron.2008.04.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Wildsmith KR, Holley M, Savage JC, Skerrett R, Landreth GE. Evidence for impaired amyloid β clearance in Alzheimer’s disease. Alzheimers Res Ther. 2013;5:33. doi: 10.1186/alzrt187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Hou TT, Han YD, Cong L, Liu CC, Liang XY, Xue FZ, Du YF. Apolipoprotein E Facilitates Amyloid-β Oligomer-Induced Tau Phosphorylation. J Alzheimer’s disease: JAD. 2020;74:521–34. doi: 10.3233/JAD-190711. [DOI] [PubMed] [Google Scholar]
- 100.Therriault J, Benedet AL, Pascoal TA, Mathotaarachchi S, Chamoun M, Savard M, Thomas E, Kang MS, Lussier F, Tissot C, et al. Association of Apolipoprotein E epsilon4 With Medial Temporal Tau Independent of Amyloid-beta. JAMA Neurol. 2020;77:470–9. doi: 10.1001/jamaneurol.2019.4421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Wadhwani AR, Affaneh A, Van Gulden S, Kessler JA. Neuronal apolipoprotein E4 increases cell death and phosphorylated tau release in alzheimer disease. Ann Neurol. 2019;85:726–39. doi: 10.1002/ana.25455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Harris FM, Brecht WJ, Xu Q, Mahley RW, Huang Y. Increased tau phosphorylation in apolipoprotein E4 transgenic mice is associated with activation of extracellular signal-regulated kinase: modulation by zinc. J Biol Chem. 2004;279:44795–801. doi: 10.1074/jbc.M408127200. [DOI] [PubMed] [Google Scholar]
- 103.Shi Y, Yamada K, Liddelow SA, Smith ST, Zhao L, Luo W, Tsai RM, Spina S, Grinberg LT, Rojas JC, et al. ApoE4 markedly exacerbates tau-mediated neurodegeneration in a mouse model of tauopathy. Nature. 2017;549:523–7. doi: 10.1038/nature24016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Shi Y, Manis M, Long J, Wang K, Sullivan PM, Remolina Serrano J, Hoyle R, Holtzman DM. Microglia drive APOE-dependent neurodegeneration in a tauopathy mouse model. J Exp Med. 2019;216:2546–61. doi: 10.1084/jem.20190980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Wang C, Xiong M, Gratuze M, Bao X, Shi Y, Andhey PS, Manis M, Schroeder C, Yin Z, Madore C, et al. Selective removal of astrocytic APOE4 strongly protects against tau-mediated neurodegeneration and decreases synaptic phagocytosis by microglia. Neuron. 2021;109:1657–74.e1657. doi: 10.1016/j.neuron.2021.03.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Zhao N, Liu CC, Van Ingelgom AJ, Linares C, Kurti A, Knight JA, Heckman MG, Diehl NN, Shinohara M, Martens YA, et al. APOE ε2 is associated with increased tau pathology in primary tauopathy. Nat Commun. 2018;9:4388. doi: 10.1038/s41467-018-06783-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Jun GR, You Y, Zhu C, Meng G, Chung J, Panitch R, Hu J, Xia W, Alzheimer’s Disease Genetics C, Bennett DA, et al: Protein phosphatase 2A and complement component 4 are linked to the protective effect of APOE varepsilon2 for Alzheimer’s disease. Alzheimers Dement 2022. [DOI] [PMC free article] [PubMed]
- 108.Koriath C, Lashley T, Taylor W, Druyeh R, Dimitriadis A, Denning N, Williams J, Warren JD, Fox NC, Schott JM, et al. ApoE4 lowers age at onset in patients with frontotemporal dementia and tauopathy independent of amyloid-beta copathology. Alzheimers Dement (Amst) 2019;11:277–80. doi: 10.1016/j.dadm.2019.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Deng H, Ordaz A, Upadhyayula PS, Gillis-Buck EM, Suen CG, Melhado CG, Mohammed N, Lam T, Yue JK: Apolipoprotein E Epsilon 4 Genotype, Mild Traumatic Brain Injury, and the Development of Chronic Traumatic Encephalopathy. Medical sciences (Basel, Switzerland) 2018, 6. [DOI] [PMC free article] [PubMed]
- 110.Agosta F, Vossel KA, Miller BL, Migliaccio R, Bonasera SJ, Filippi M, Boxer AL, Karydas A, Possin KL, Gorno-Tempini ML. Apolipoprotein E epsilon4 is associated with disease-specific effects on brain atrophy in Alzheimer’s disease and frontotemporal dementia. Proc Natl Acad Sci U S A. 2009;106:2018–22. doi: 10.1073/pnas.0812697106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Strittmatter WJ, Saunders AM, Goedert M, Weisgraber KH, Dong LM, Jakes R, Huang DY, Pericak-Vance M, Schmechel D, Roses AD. Isoform-specific interactions of apolipoprotein E with microtubule-associated protein tau: implications for Alzheimer disease. Proc Natl Acad Sci U S A. 1994;91:11183–6. doi: 10.1073/pnas.91.23.11183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Gibb GM, Pearce J, Betts JC, Lovestone S, Hoffmann MM, Maerz W, Blackstock WP, Anderton BH. Differential effects of apolipoprotein E isoforms on phosphorylation at specific sites on tau by glycogen synthase kinase-3 beta identified by nano-electrospray mass spectrometry. FEBS Lett. 2000;485:99–103. doi: 10.1016/s0014-5793(00)02196-7. [DOI] [PubMed] [Google Scholar]
- 113.Caruso A, Motolese M, Iacovelli L, Caraci F, Copani A, Nicoletti F, Terstappen GC, Gaviraghi G, Caricasole A. Inhibition of the canonical Wnt signaling pathway by apolipoprotein E4 in PC12 cells. J Neurochem. 2006;98:364–71. doi: 10.1111/j.1471-4159.2006.03867.x. [DOI] [PubMed] [Google Scholar]
- 114.Kloske CM, Wilcock DM. The Important Interface Between Apolipoprotein E and Neuroinflammation in Alzheimer’s Disease. Front Immunol. 2020;11:754. doi: 10.3389/fimmu.2020.00754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Song WM, Colonna M. The identity and function of microglia in neurodegeneration. Nat Immunol. 2018;19:1048–58. doi: 10.1038/s41590-018-0212-1. [DOI] [PubMed] [Google Scholar]
- 116.Krasemann S, Madore C, Cialic R, Baufeld C, Calcagno N, El Fatimy R, Beckers L, O’Loughlin E, Xu Y, Fanek Z, et al. The TREM2-APOE Pathway Drives the Transcriptional Phenotype of Dysfunctional Microglia in Neurodegenerative Diseases. Immunity. 2017;47:566–81.e569. doi: 10.1016/j.immuni.2017.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Ulrich JD, Ulland TK, Mahan TE, Nystrom S, Nilsson KP, Song WM, Zhou Y, Reinartz M, Choi S, Jiang H, et al. ApoE facilitates the microglial response to amyloid plaque pathology. J Exp Med. 2018;215:1047–58. doi: 10.1084/jem.20171265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Keren-Shaul H, Spinrad A, Weiner A, Matcovitch-Natan O, Dvir-Szternfeld R, Ulland TK, David E, Baruch K, Lara-Astaiso D, Toth B, et al. A Unique Microglia Type Associated with Restricting Development of Alzheimer’s Disease. Cell. 2017;169:1276–90.e1217. doi: 10.1016/j.cell.2017.05.018. [DOI] [PubMed] [Google Scholar]
- 119.Rodriguez GA, Tai LM, LaDu MJ, Rebeck GW. Human APOE4 increases microglia reactivity at Abeta plaques in a mouse model of Abeta deposition. J Neuroinflammation. 2014;11:111. doi: 10.1186/1742-2094-11-111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Fitz NF, Wolfe CM, Playso BE, Biedrzycki RJ, Lu Y, Nam KN, Lefterov I, Koldamova R. Trem2 deficiency differentially affects phenotype and transcriptome of human APOE3 and APOE4 mice. Mol neurodegeneration. 2020;15:41. doi: 10.1186/s13024-020-00394-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Fitz NF, Nam KN, Wolfe CM, Letronne F, Playso BE, Iordanova BE, Kozai TDY, Biedrzycki RJ, Kagan VE, Tyurina YY, et al. Phospholipids of APOE lipoproteins activate microglia in an isoform-specific manner in preclinical models of Alzheimer’s disease. Nat Commun. 2021;12:3416. doi: 10.1038/s41467-021-23762-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Atagi Y, Liu CC, Painter MM, Chen XF, Verbeeck C, Zheng H, Li X, Rademakers R, Kang SS, Xu H, et al. Apolipoprotein E Is a Ligand for Triggering Receptor Expressed on Myeloid Cells 2 (TREM2) J Biol Chem. 2015;290:26043–50. doi: 10.1074/jbc.M115.679043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Jendresen C, Arskog V, Daws MR, Nilsson LN. The Alzheimer’s disease risk factors apolipoprotein E and TREM2 are linked in a receptor signaling pathway. J Neuroinflammation. 2017;14:59. doi: 10.1186/s12974-017-0835-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Yeh FL, Wang Y, Tom I, Gonzalez LC, Sheng M. TREM2 Binds to Apolipoproteins, Including APOE and CLU/APOJ, and Thereby Facilitates Uptake of Amyloid-Beta by Microglia. Neuron. 2016;91:328–40. doi: 10.1016/j.neuron.2016.06.015. [DOI] [PubMed] [Google Scholar]
- 125.Fernandez CG, Hamby ME, McReynolds ML, Ray WJ. The Role of APOE4 in Disrupting the Homeostatic Functions of Astrocytes and Microglia in Aging and Alzheimer’s Disease. Front Aging Neurosci. 2019;11:14. doi: 10.3389/fnagi.2019.00014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Tao Q, Ang TFA, DeCarli C, Auerbach SH, Devine S, Stein TD, Zhang X, Massaro J, Au R, Qiu WQ. Association of Chronic Low-grade Inflammation With Risk of Alzheimer Disease in ApoE4 Carriers. JAMA Netw open. 2018;1:e183597. doi: 10.1001/jamanetworkopen.2018.3597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Martiskainen H, Takalo M, Solomon A, Stančáková A, Marttinen M, Natunen T, Haapasalo A, Herukka SK, Kuusisto J, Soininen H, et al. Decreased plasma C-reactive protein levels in APOE ε4 allele carriers. Ann Clin Transl Neurol. 2018;5:1229–40. doi: 10.1002/acn3.639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Wu S, Hsu LA, Teng MS, Lin JF, Chou HH, Lee MC, Wu YM, Su CW, Ko YL. Interactive effects of C-reactive protein levels on the association between APOE variants and triglyceride levels in a Taiwanese population. Lipids Health Dis. 2016;15:94. doi: 10.1186/s12944-016-0262-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Sproston NR, Ashworth JJ. Role of C-Reactive Protein at Sites of Inflammation and Infection. Front Immunol. 2018;9:754. doi: 10.3389/fimmu.2018.00754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Mooijaart SP, Sattar N, Trompet S, Polisecki E, de Craen AJ, Schaefer EJ, Jahn SE, van Himbergen T, Welsh P, Ford I, et al. C-reactive protein and genetic variants and cognitive decline in old age: the PROSPER study. PLoS ONE. 2011;6:e23890. doi: 10.1371/journal.pone.0023890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Raulin AC, Martens YA, Bu G. Lipoproteins in the Central Nervous System: From Biology to Pathobiology. Annu Rev Biochem. 2022;91:731–59. doi: 10.1146/annurev-biochem-032620-104801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Michikawa M, Fan QW, Isobe I, Yanagisawa K. Apolipoprotein E exhibits isoform-specific promotion of lipid efflux from astrocytes and neurons in culture. J Neurochem. 2000;74:1008–16. doi: 10.1046/j.1471-4159.2000.0741008.x. [DOI] [PubMed] [Google Scholar]
- 133.Sienski G, Narayan P, Bonner JM, Kory N, Boland S, Arczewska AA, Ralvenius WT, Akay L, Lockshin E, He L, et al: APOE4 disrupts intracellular lipid homeostasis in human iPSC-derived glia. Science translational medicine 2021, 13. [DOI] [PMC free article] [PubMed]
- 134.Farmer BC, Kluemper J, Johnson LA. Apolipoprotein E4 Alters Astrocyte Fatty Acid Metabolism and Lipid Droplet Formation. Cells 2019, 8. [DOI] [PMC free article] [PubMed]
- 135.Marschallinger J, Iram T, Zardeneta M, Lee SE, Lehallier B, Haney MS, Pluvinage JV, Mathur V, Hahn O, Morgens DW, et al. Lipid-droplet-accumulating microglia represent a dysfunctional and proinflammatory state in the aging brain. Nat Neurosci. 2020;23:194–208. doi: 10.1038/s41593-019-0566-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Claes C, Danhash EP, Hasselmann J, Chadarevian JP, Shabestari SK, England WE, Lim TE, Hidalgo JLS, Spitale RC, Davtyan H, Blurton-Jones M. Plaque-associated human microglia accumulate lipid droplets in a chimeric model of Alzheimer’s disease. Mol neurodegeneration. 2021;16:50. doi: 10.1186/s13024-021-00473-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Sharman MJ, Morici M, Hone E, Berger T, Taddei K, Martins IJ, Lim WL, Singh S, Wenk MR, Ghiso J, et al. APOE genotype results in differential effects on the peripheral clearance of amyloid-beta42 in APOE knock-in and knock-out mice. J Alzheimer’s disease: JAD. 2010;21:403–9. doi: 10.3233/JAD-2010-100141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Shinohara M, Kanekiyo T, Yang L, Linthicum D, Shinohara M, Fu Y, Price L, Frisch-Daiello JL, Han X, Fryer JD, Bu G. APOE2 eases cognitive decline during Aging: Clinical and preclinical evaluations. Ann Neurol. 2016;79:758–74. doi: 10.1002/ana.24628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Lefterov I, Wolfe CM, Fitz NF, Nam KN, Letronne F, Biedrzycki RJ, Kofler J, Han X, Wang J, Schug J, Koldamova R. APOE2 orchestrated differences in transcriptomic and lipidomic profiles of postmortem AD brain. Alzheimers Res Ther. 2019;11:113. doi: 10.1186/s13195-019-0558-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Duong MT, Nasrallah IM, Wolk DA, Chang CCY, Chang TY. Cholesterol, Atherosclerosis, and APOE in Vascular Contributions to Cognitive Impairment and Dementia (VCID): Potential Mechanisms and Therapy. Front Aging Neurosci. 2021;13:647990. doi: 10.3389/fnagi.2021.647990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Nelson PT, Pious NM, Jicha GA, Wilcock DM, Fardo DW, Estus S, Rebeck GW. APOE-ε2 and APOE-ε4 correlate with increased amyloid accumulation in cerebral vasculature. J Neuropathol Exp Neurol. 2013;72:708–15. doi: 10.1097/NEN.0b013e31829a25b9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Lee JS, Lee H, Park S, Choe Y, Park YH, Cheon BK, Hahn A, Ossenkoppele R, Kim HJ, Kim S, et al. Association between APOE ε2 and Aβ burden in patients with Alzheimer- and vascular-type cognitive impairment. Neurology. 2020;95:e2354–65. doi: 10.1212/WNL.0000000000010811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Kim SH, Ahn JH, Yang H, Lee P, Koh GY, Jeong Y. Cerebral amyloid angiopathy aggravates perivascular clearance impairment in an Alzheimer’s disease mouse model. Acta Neuropathol Commun. 2020;8:181. doi: 10.1186/s40478-020-01042-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Mentis AA, Dardiotis E, Chrousos GP. Apolipoprotein E4 and meningeal lymphatics in Alzheimer disease: a conceptual framework. Mol Psychiatry. 2021;26:1075–97. doi: 10.1038/s41380-020-0731-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Robert J, Button EB, Martin EM, McAlary L, Gidden Z, Gilmour M, Boyce G, Caffrey TM, Agbay A, Clark A, et al. Cerebrovascular amyloid Angiopathy in bioengineered vessels is reduced by high-density lipoprotein particles enriched in Apolipoprotein E. Mol neurodegeneration. 2020;15:23. doi: 10.1186/s13024-020-00366-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Yamazaki Y, Liu CC, Yamazaki A, Shue F, Martens YA, Chen Y, Qiao W, Kurti A, Oue H, Ren Y, et al. Vascular ApoE4 Impairs Behavior by Modulating Gliovascular Function. Neuron. 2021;109:438–47.e436. doi: 10.1016/j.neuron.2020.11.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Elahi FM, Ashimatey SB, Bennett DJ, Walters SM, La Joie R, Jiang X, Wolf A, Cobigo Y, Staffaroni AM, Rosen HJ, et al. Retinal imaging demonstrates reduced capillary density in clinically unimpaired APOE ε4 gene carriers. Alzheimers Dement (Amst) 2021;13:e12181. doi: 10.1002/dad2.12181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Margeta MA, Yin Z, Madore C, Pitts KM, Letcher SM, Tang J, Jiang S, Gauthier CD, Silveira SR, Schroeder CM, et al: Apolipoprotein E4 impairs the response of neurodegenerative retinal microglia and prevents neuronal loss in glaucoma. Immunity 2022. [DOI] [PMC free article] [PubMed]
- 149.Bangen KJ, Smirnov DS, Delano-Wood L, Wierenga CE, Bondi MW, Salmon DP, Galasko D. Arterial stiffening acts synergistically with APOE genotype and AD biomarker status to influence memory in older adults without dementia. Alzheimers Res Ther. 2021;13:121. doi: 10.1186/s13195-021-00851-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Zhou R, Chen H, Ye F, Huang S, Zhang J. Influence of Hypertension on Longitudinal Changes in Brain Glucose Metabolism Was Modified by the APOE4 Allele Among Cognitively Normal Older Individuals. Front Aging Neurosci. 2020;12:85. doi: 10.3389/fnagi.2020.00085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Scheinman SB, Zaldua S, Dada A, Krochmaliuk K, Dye K, Marottoli FM, Thatcher GRJ, Tai LM. Systemic Candesartan Treatment Modulates Behavior, Synaptic Protein Levels, and Neuroinflammation in Female Mice That Express Human APOE4. Front NeuroSci. 2021;15:628403. doi: 10.3389/fnins.2021.628403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Salomon-Zimri S, Glat MJ, Barhum Y, Luz I, Boehm-Cagan A, Liraz O, Ben-Zur T, Offen D, Michaelson DM. Reversal of ApoE4-Driven Brain Pathology by Vascular Endothelial Growth Factor Treatment. J Alzheimer’s disease: JAD. 2016;53:1443–58. doi: 10.3233/JAD-160182. [DOI] [PubMed] [Google Scholar]
- 153.Zaldua S, Damen FC, Pisharody R, Thomas R, Fan KD, Ekkurthi GK, Scheinman SB, Alahmadi S, Marottoli FM, Alford S, et al. Epidermal growth factor treatment of female mice that express APOE4 at an age of advanced pathology mitigates behavioral and cerebrovascular dysfunction. Heliyon. 2020;6:e03919. doi: 10.1016/j.heliyon.2020.e03919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Moore AM, Mahoney E, Dumitrescu L, De Jager PL, Koran MEI, Petyuk VA, Robinson RA, Ruderfer DM, Cox NJ, Schneider JA, et al. APOE ε4-specific associations of VEGF gene family expression with cognitive aging and Alzheimer’s disease. Neurobiol Aging. 2020;87:18–25. doi: 10.1016/j.neurobiolaging.2019.10.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Liu CC, Yamazaki Y, Heckman MG, Martens YA, Jia L, Yamazaki A, Diehl NN, Zhao J, Zhao N, DeTure M, et al. Tau and apolipoprotein E modulate cerebrovascular tight junction integrity independent of cerebral amyloid angiopathy in Alzheimer’s disease. Alzheimers Dement. 2020;16:1372–83. doi: 10.1002/alz.12104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Montagne A, Nation DA, Sagare AP, Barisano G, Sweeney MD, Chakhoyan A, Pachicano M, Joe E, Nelson AR, D’Orazio LM, et al. APOE4 leads to blood-brain barrier dysfunction predicting cognitive decline. Nature. 2020;581:71–6. doi: 10.1038/s41586-020-2247-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Kadry H, Noorani B, Cucullo L. A blood-brain barrier overview on structure, function, impairment, and biomarkers of integrity. Fluids and barriers of the CNS. 2020;17:69. doi: 10.1186/s12987-020-00230-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Masuda T, Shimazawa M, Hashimoto Y, Kojima A, Nakamura S, Suemori S, Mochizuki K, Kawakami H, Kawase K, Hara H. Apolipoprotein E2 and E3, but Not E4, Promote Retinal Pathologic Neovascularization. Investig Ophthalmol Vis Sci. 2017;58:1208–17. doi: 10.1167/iovs.16-20539. [DOI] [PubMed] [Google Scholar]
- 159.Barisano G, Kisler K, Wilkinson B, Nikolakopoulou AM, Sagare AP, Wang Y, Gilliam W, Huuskonen MT, Hung ST, Ichida JK, et al: A “multi-omics” analysis of blood-brain barrier and synaptic dysfunction in APOE4 mice. J Exp Med 2022, 219. [DOI] [PMC free article] [PubMed]
- 160.Liu CC, Zhao J, Fu Y, Inoue Y, Ren Y, Chen Y, Doss SV, Shue F, Jeevaratnam S, Bastea L, et al. Peripheral apoE4 enhances Alzheimer’s pathology and impairs cognition by compromising cerebrovascular function. Nat Neurosci. 2022;25:1020–33. doi: 10.1038/s41593-022-01127-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Armulik A, Genové G, Mäe M, Nisancioglu MH, Wallgard E, Niaudet C, He L, Norlin J, Lindblom P, Strittmatter K, et al. Pericytes regulate the blood-brain barrier. Nature. 2010;468:557–61. doi: 10.1038/nature09522. [DOI] [PubMed] [Google Scholar]
- 162.Montagne A, Nikolakopoulou AM, Huuskonen MT, Sagare AP, Lawson EJ, Lazic D, Rege SV, Grond A, Zuniga E, Barnes SR, et al. APOE4 accelerates advanced-stage vascular and neurodegenerative disorder in old Alzheimer’s mice via cyclophilin A independently of amyloid-β. Nat Aging. 2021;1:506–20. doi: 10.1038/s43587-021-00073-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Shinohara M, Tashiro Y, Suzuki K, Fukumori A, Bu G, Sato N. Interaction between APOE genotype and diabetes in cognitive decline. Alzheimers Dement (Amst) 2020;12:e12006. doi: 10.1002/dad2.12006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.El-Lebedy D, Raslan HM, Mohammed AM. Apolipoprotein E gene polymorphism and risk of type 2 diabetes and cardiovascular disease. Cardiovasc Diabetol. 2016;15:12. doi: 10.1186/s12933-016-0329-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Liu S, Liu J, Weng R, Gu X, Zhong Z. Apolipoprotein E gene polymorphism and the risk of cardiovascular disease and type 2 diabetes. BMC Cardiovasc Disord. 2019;19:213. doi: 10.1186/s12872-019-1194-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Torres-Perez E, Ledesma M, Garcia-Sobreviela MP, Leon-Latre M, Arbones-Mainar JM. Apolipoprotein E4 association with metabolic syndrome depends on body fatness. Atherosclerosis. 2016;245:35–42. doi: 10.1016/j.atherosclerosis.2015.11.029. [DOI] [PubMed] [Google Scholar]
- 167.Peng Y, Gao P, Shi L, Chen L, Liu J, Long J. Central and Peripheral Metabolic Defects Contribute to the Pathogenesis of Alzheimer’s Disease: Targeting Mitochondria for Diagnosis and Prevention. Antioxid Redox Signal. 2020;32:1188–236. doi: 10.1089/ars.2019.7763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Polis B, Samson AO: A New Perspective on Alzheimer’s Disease as a Brain Expression of a Complex Metabolic Disorder. In Alzheimer’s Disease Edited by Wisniewski T. Brisbane (AU): Codon Publications Copyright: The Authors.; 2019. [PubMed]
- 169.Martens YA, Zhao N, Liu CC, Kanekiyo T, Yang AJ, Goate AM, Holtzman DM, Bu G. ApoE Cascade Hypothesis in the pathogenesis of Alzheimer’s disease and related dementias. Neuron 2022. [DOI] [PMC free article] [PubMed]
- 170.He L, Davila-Velderrain J, Sumida TS, Hafler DA, Kellis M, Kulminski AM. NEBULA is a fast negative binomial mixed model for differential or co-expression analysis of large-scale multi-subject single-cell data. Commun biology. 2021;4:629. doi: 10.1038/s42003-021-02146-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Area-Gomez E, Larrea D, Pera M, Agrawal RR, Guilfoyle DN, Pirhaji L, Shannon K, Arain HA, Ashok A, Chen Q, et al. APOE4 is Associated with Differential Regional Vulnerability to Bioenergetic Deficits in Aged APOE Mice. Sci Rep. 2020;10:4277. doi: 10.1038/s41598-020-61142-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Zhao J, Fu Y, Yamazaki Y, Ren Y, Davis MD, Liu CC, Lu W, Wang X, Chen K, Cherukuri Y, et al. APOE4 exacerbates synapse loss and neurodegeneration in Alzheimer’s disease patient iPSC-derived cerebral organoids. Nat Commun. 2020;11:5540. doi: 10.1038/s41467-020-19264-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Zhao N, Ren Y, Yamazaki Y, Qiao W, Li F, Felton LM, Mahmoudiandehkordi S, Kueider-Paisley A, Sonoustoun B, Arnold M, et al. Alzheimer’s Risk Factors Age, APOE Genotype, and Sex Drive Distinct Molecular Pathways. Neuron. 2020;106:727–42 e726. doi: 10.1016/j.neuron.2020.02.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Konijnenberg E, Tijms BM, Gobom J, Dobricic V, Bos I, Vos S, Tsolaki M, Verhey F, Popp J, Martinez-Lage P, et al. APOE epsilon4 genotype-dependent cerebrospinal fluid proteomic signatures in Alzheimer’s disease. Alzheimers Res Ther. 2020;12:65. doi: 10.1186/s13195-020-00628-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Hesse R, Hurtado ML, Jackson RJ, Eaton SL, Herrmann AG, Colom-Cadena M, Tzioras M, King D, Rose J, Tulloch J, et al. Comparative profiling of the synaptic proteome from Alzheimer’s disease patients with focus on the APOE genotype. Acta Neuropathol Commun. 2019;7:214. doi: 10.1186/s40478-019-0847-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Dai J, Johnson ECB, Dammer EB, Duong DM, Gearing M, Lah JJ, Levey AI, Wingo TS, Seyfried NT. Effects of APOE Genotype on Brain Proteomic Network and Cell Type Changes in Alzheimer’s Disease. Front Mol Neurosci. 2018;11:454. doi: 10.3389/fnmol.2018.00454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Liu T, Zhu B, Liu Y, Zhang X, Yin J, Li X, Jiang L, Hodges AP, Rosenthal SB, Zhou L, et al: Multi-omic comparison of Alzheimer’s variants in human ESC-derived microglia reveals convergence at APOE. J Exp Med 2020, 217. [DOI] [PMC free article] [PubMed]
- 178.Qi G, Mi Y, Shi X, Gu H, Brinton RD, Yin F. ApoE4 Impairs Neuron-Astrocyte Coupling of Fatty Acid Metabolism. Cell Rep. 2021;34:108572. doi: 10.1016/j.celrep.2020.108572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Tcw J, Qian L, Pipalia NH, Chao MJ, Liang SA, Shi Y, Jain BR, Bertelsen SE, Kapoor M, Marcora E, et al. Cholesterol and matrisome pathways dysregulated in astrocytes and microglia. Cell. 2022;185:2213–33 e2225. doi: 10.1016/j.cell.2022.05.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Wardell MR, Brennan SO, Janus ED, Fraser R, Carrell RW. Apolipoprotein E2-Christchurch (136 Arg----Ser). New variant of human apolipoprotein E in a patient with type III hyperlipoproteinemia. J Clin Investig. 1987;80:483–90. doi: 10.1172/JCI113096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Lalazar A, Weisgraber KH, Rall SC, Jr, Giladi H, Innerarity TL, Levanon AZ, Boyles JK, Amit B, Gorecki M, Mahley RW, et al. Site-specific mutagenesis of human apolipoprotein E. Receptor binding activity of variants with single amino acid substitutions. J Biol Chem. 1988;263:3542–5. [PubMed] [Google Scholar]
- 182.Orth M, Weng W, Funke H, Steinmetz A, Assmann G, Nauck M, Dierkes J, Ambrosch A, Weisgraber KH, Mahley RW, et al: Effects of a frequent apolipoprotein E isoform, ApoE4Freiburg (Leu28–>Pro), on lipoproteins and the prevalence of coronary artery disease in whites Arteriosclerosis, thrombosis, and vascular biology 1999, 19:1306–1315. [DOI] [PubMed]
- 183.Abou Ziki MD, Strulovici-Barel Y, Hackett NR, Rodriguez-Flores JL, Mezey JG, Salit J, Radisch S, Hollmann C, Chouchane L, Malek J, et al. Prevalence of the apolipoprotein E Arg145Cys dyslipidemia at-risk polymorphism in African-derived populations. Am J Cardiol. 2014;113:302–8. doi: 10.1016/j.amjcard.2013.09.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Sevigny J, Chiao P, Bussiere T, Weinreb PH, Williams L, Maier M, Dunstan R, Salloway S, Chen T, Ling Y, et al. The antibody aducanumab reduces Abeta plaques in Alzheimer’s disease. Nature. 2016;537:50–6. doi: 10.1038/nature19323. [DOI] [PubMed] [Google Scholar]
- 185.Sperling RA, Jack CR, Jr, Black SE, Frosch MP, Greenberg SM, Hyman BT, Scheltens P, Carrillo MC, Thies W, Bednar MM, et al. Amyloid-related imaging abnormalities in amyloid-modifying therapeutic trials: recommendations from the Alzheimer’s Association Research Roundtable Workgroup. Alzheimers Dement. 2011;7:367–85. doi: 10.1016/j.jalz.2011.05.2351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Cummings J, Aisen P, Apostolova LG, Atri A, Salloway S, Weiner M. Aducanumab: Appropriate Use Recommendations. J Prev Alzheimers Dis. 2021;8:398–410. doi: 10.14283/jpad.2021.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Cummings J, Rabinovici GD, Atri A, Aisen P, Apostolova LG, Hendrix S, Sabbagh M, Selkoe D, Weiner M, Salloway S. Aducanumab: Appropriate Use Recommendations Update. J Prev Alzheimers Dis. 2022;9:221–30. doi: 10.14283/jpad.2022.34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Sullivan PM, Han B, Liu F, Mace BE, Ervin JF, Wu S, Koger D, Paul S, Bales KR. Reduced levels of human apoE4 protein in an animal model of cognitive impairment. Neurobiol Aging. 2011;32:791–801. doi: 10.1016/j.neurobiolaging.2009.05.011. [DOI] [PubMed] [Google Scholar]
- 189.Rezeli M, Zetterberg H, Blennow K, Brinkmalm A, Laurell T, Hansson O, Marko-Varga G. Quantification of total apolipoprotein E and its specific isoforms in cerebrospinal fluid and blood in Alzheimer’s disease and other neurodegenerative diseases. EuPA Open Proteomics. 2015;8:6. [Google Scholar]
- 190.Minta K, Brinkmalm G, Janelidze S, Sjödin S, Portelius E, Stomrud E, Zetterberg H, Blennow K, Hansson O, Andreasson U. Quantification of total apolipoprotein E and its isoforms in cerebrospinal fluid from patients with neurodegenerative diseases. Alzheimers Res Ther. 2020;12:19. doi: 10.1186/s13195-020-00585-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Kim J, Jiang H, Park S, Eltorai AE, Stewart FR, Yoon H, Basak JM, Finn MB, Holtzman DM. Haploinsufficiency of human APOE reduces amyloid deposition in a mouse model of amyloid-beta amyloidosis. J neuroscience: official J Soc Neurosci. 2011;31:18007–12. doi: 10.1523/JNEUROSCI.3773-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Zheng JY, Sun J, Ji CM, Shen L, Chen ZJ, Xie P, Sun YZ, Yu RT. Selective deletion of apolipoprotein E in astrocytes ameliorates the spatial learning and memory deficits in Alzheimer’s disease (APP/PS1) mice by inhibiting TGF-β/Smad2/STAT3 signaling. Neurobiol Aging. 2017;54:112–32. doi: 10.1016/j.neurobiolaging.2017.03.002. [DOI] [PubMed] [Google Scholar]
- 193.Kim J, Eltorai AE, Jiang H, Liao F, Verghese PB, Kim J, Stewart FR, Basak JM, Holtzman DM. Anti-apoE immunotherapy inhibits amyloid accumulation in a transgenic mouse model of Abeta amyloidosis. J Exp Med. 2012;209:2149–56. doi: 10.1084/jem.20121274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Liao F, Hori Y, Hudry E, Bauer AQ, Jiang H, Mahan TE, Lefton KB, Zhang TJ, Dearborn JT, Kim J, et al. Anti-ApoE antibody given after plaque onset decreases Abeta accumulation and improves brain function in a mouse model of Abeta amyloidosis. J neuroscience: official J Soc Neurosci. 2014;34:7281–92. doi: 10.1523/JNEUROSCI.0646-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Wolska A, Reimund M, Sviridov DO, Amar MJ, Remaley AT. Apolipoprotein Mimetic Peptides: Potential New Therapies for Cardiovascular Diseases. Cells 2021, 10. [DOI] [PMC free article] [PubMed]
- 196.Christensen DJ, Ohkubo N, Oddo J, Van Kanegan MJ, Neil J, Li F, Colton CA, Vitek MP. Apolipoprotein E and peptide mimetics modulate inflammation by binding the SET protein and activating protein phosphatase 2A. J Immunol. 2011;186:2535–42. doi: 10.4049/jimmunol.1002847. [DOI] [PubMed] [Google Scholar]
- 197.Laskowitz DT, Wang H, Chen T, Lubkin DT, Cantillana V, Tu TM, Kernagis D, Zhou G, Macy G, Kolls BJ, Dawson HN. Neuroprotective pentapeptide CN-105 is associated with reduced sterile inflammation and improved functional outcomes in a traumatic brain injury murine model. Sci Rep. 2017;7:46461. doi: 10.1038/srep46461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Tu TM, Kolls BJ, Soderblom EJ, Cantillana V, Ferrell PD, Moseley MA, Wang H, Dawson HN, Laskowitz DT. Apolipoprotein E mimetic peptide, CN-105, improves outcomes in ischemic stroke. Ann Clin Transl Neurol. 2017;4:246–65. doi: 10.1002/acn3.399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Krishnamurthy K, Cantillana V, Wang H, Sullivan PM, Kolls BJ, Ge X, Lin Y, Mace B, Laskowitz DT. ApoE mimetic improves pathology and memory in a model of Alzheimer’s disease. Brain Res. 2020;1733:146685. doi: 10.1016/j.brainres.2020.146685. [DOI] [PubMed] [Google Scholar]
- 200.Chernick D, Ortiz-Valle S, Jeong A, Swaminathan SK, Kandimalla KK, Rebeck GW, Li L. High-density lipoprotein mimetic peptide 4F mitigates amyloid-beta-induced inhibition of apolipoprotein E secretion and lipidation in primary astrocytes and microglia. J Neurochem. 2018;147:647–62. doi: 10.1111/jnc.14554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Sadowski MJ, Pankiewicz J, Scholtzova H, Mehta PD, Prelli F, Quartermain D, Wisniewski T. Blocking the apolipoprotein E/amyloid-beta interaction as a potential therapeutic approach for Alzheimer’s disease. Proc Natl Acad Sci U S A. 2006;103:18787–92. doi: 10.1073/pnas.0604011103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Kuszczyk MA, Sanchez S, Pankiewicz J, Kim J, Duszczyk M, Guridi M, Asuni AA, Sullivan PM, Holtzman DM, Sadowski MJ. Blocking the interaction between apolipoprotein E and Abeta reduces intraneuronal accumulation of Abeta and inhibits synaptic degeneration. Am J Pathol. 2013;182:1750–68. doi: 10.1016/j.ajpath.2013.01.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Sawmiller D, Habib A, Hou H, Mori T, Fan A, Tian J, Zeng J, Giunta B, Sanberg PR, Mattson MP, Tan J. A Novel Apolipoprotein E Antagonist Functionally Blocks Apolipoprotein E Interaction With N-terminal Amyloid Precursor Protein, Reduces beta-Amyloid-Associated Pathology, and Improves Cognition. Biol Psychiatry. 2019;86:208–20. doi: 10.1016/j.biopsych.2019.04.026. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 204.Liao F, Li A, Xiong M, Bien-Ly N, Jiang H, Zhang Y, Finn MB, Hoyle R, Keyser J, Lefton KB, et al. Targeting of nonlipidated, aggregated apoE with antibodies inhibits amyloid accumulation. J Clin Investig. 2018;128:2144–55. doi: 10.1172/JCI96429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Xiong M, Jiang H, Serrano JR, Gonzales ER, Wang C, Gratuze M, Hoyle R, Bien-Ly N, Silverman AP, Sullivan PM, et al: APOE immunotherapy reduces cerebral amyloid angiopathy and amyloid plaques while improving cerebrovascular function. Science translational medicine 2021, 13. [DOI] [PMC free article] [PubMed]
- 206.Gratuze M, Jiang H, Wang C, Xiong M, Bao X, Holtzman DM. APOE antibody inhibits Abeta-associated tau seeding and spreading in a mouse model. Ann Neurol 2022. [DOI] [PMC free article] [PubMed]
- 207.Boehm-Cagan A, Bar R, Liraz O, Bielicki JK, Johansson JO, Michaelson DM. ABCA1 Agonist Reverses the ApoE4-Driven Cognitive and Brain Pathologies. J Alzheimer’s disease: JAD. 2016;54:1219–33. doi: 10.3233/JAD-160467. [DOI] [PubMed] [Google Scholar]
- 208.Laffitte BA, Repa JJ, Joseph SB, Wilpitz DC, Kast HR, Mangelsdorf DJ, Tontonoz P. LXRs control lipid-inducible expression of the apolipoprotein E gene in macrophages and adipocytes. Proc Natl Acad Sci U S A. 2001;98:507–12. doi: 10.1073/pnas.021488798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Cramer PE, Cirrito JR, Wesson DW, Lee CY, Karlo JC, Zinn AE, Casali BT, Restivo JL, Goebel WD, James MJ, et al. ApoE-directed therapeutics rapidly clear beta-amyloid and reverse deficits in AD mouse models. Science. 2012;335:1503–6. doi: 10.1126/science.1217697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Ghosal K, Haag M, Verghese PB, West T, Veenstra T, Braunstein JB, Bateman RJ, Holtzman DM, Landreth GE. A randomized controlled study to evaluate the effect of bexarotene on amyloid-beta and apolipoprotein E metabolism in healthy subjects. Alzheimers Dement (N Y) 2016;2:110–20. doi: 10.1016/j.trci.2016.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Cummings JL, Zhong K, Kinney JW, Heaney C, Moll-Tudla J, Joshi A, Pontecorvo M, Devous M, Tang A, Bena J. Double-blind, placebo-controlled, proof-of-concept trial of bexarotene Xin moderate Alzheimer’s disease. Alzheimers Res Ther. 2016;8:4. doi: 10.1186/s13195-016-0173-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Ramaswamy G, Xu Q, Huang Y, Weisgraber KH. Effect of domain interaction on apolipoprotein E levels in mouse brain. J neuroscience: official J Soc Neurosci. 2005;25:10658–63. doi: 10.1523/JNEUROSCI.1922-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Chen HK, Ji ZS, Dodson SE, Miranda RD, Rosenblum CI, Reynolds IJ, Freedman SB, Weisgraber KH, Huang Y, Mahley RW. Apolipoprotein E4 domain interaction mediates detrimental effects on mitochondria and is a potential therapeutic target for Alzheimer disease. J Biol Chem. 2011;286:5215–21. doi: 10.1074/jbc.M110.151084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Chen HK, Liu Z, Meyer-Franke A, Brodbeck J, Miranda RD, McGuire JG, Pleiss MA, Ji ZS, Balestra ME, Walker DW, et al. Small molecule structure correctors abolish detrimental effects of apolipoprotein E4 in cultured neurons. J Biol Chem. 2012;287:5253–66. doi: 10.1074/jbc.M111.276162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Wang C, Najm R, Xu Q, Jeong DE, Walker D, Balestra ME, Yoon SY, Yuan H, Li G, Miller ZA, et al. Gain of toxic apolipoprotein E4 effects in human iPSC-derived neurons is ameliorated by a small-molecule structure corrector. Nat Med. 2018;24:647–57. doi: 10.1038/s41591-018-0004-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Lu Y, Xue J, Deng T, Zhou X, Yu K, Deng L, Huang M, Yi X, Liang M, Wang Y, et al. Safety and feasibility of CRISPR-edited T cells in patients with refractory non-small-cell lung cancer. Nat Med. 2020;26:732–40. doi: 10.1038/s41591-020-0840-5. [DOI] [PubMed] [Google Scholar]
- 217.Yu M, Liu X, Cheng H, Kuang L, Zhang S, Yan X: Latest progress in the study of nanoparticle-based delivery of the CRISPR/Cas9 system. Methods (San Diego, Calif) 2021. [DOI] [PubMed]
- 218.Schmid B, Prehn KR, Nimsanor N, Garcia BIA, Poulsen U, Jorring I, Rasmussen MA, Clausen C, Mau-Holzmann UA, Ramakrishna S, et al. Generation of a set of isogenic, gene-edited iPSC lines homozygous for all main APOE variants and an APOE knock-out line. Stem Cell Res. 2019;34:101349. doi: 10.1016/j.scr.2018.11.010. [DOI] [PubMed] [Google Scholar]
- 219.Wang D, Tai PWL, Gao G. Adeno-associated virus vector as a platform for gene therapy delivery. Nat Rev Drug Discov. 2019;18:358–78. doi: 10.1038/s41573-019-0012-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Smalley E. First AAV gene therapy poised for landmark approval. Nat Biotechnol. 2017;35:998–9. doi: 10.1038/nbt1117-998. [DOI] [PubMed] [Google Scholar]
- 221.Dunbar CE, High KA, Joung JK, Kohn DB, Ozawa K, Sadelain M. Gene therapy comes of age. Science 2018, 359. [DOI] [PubMed]
- 222.Hudry E, Dashkoff J, Roe AD, Takeda S, Koffie RM, Hashimoto T, Scheel M, Spires-Jones T, Arbel-Ornath M, Betensky R, et al. Gene transfer of human Apoe isoforms results in differential modulation of amyloid deposition and neurotoxicity in mouse brain. Sci Transl Med. 2013;5:212ra161. doi: 10.1126/scitranslmed.3007000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223.Hu J, Liu CC, Chen XF, Zhang YW, Xu H, Bu G. Opposing effects of viral mediated brain expression of apolipoprotein E2 (apoE2) and apoE4 on apoE lipidation and Abeta metabolism in apoE4-targeted replacement mice. Mol neurodegeneration. 2015;10:6. doi: 10.1186/s13024-015-0001-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Koller EJ, De La Cruz G, Weinrich E, Williams M, Cruz T, Ryu PE, Golde D, Sullivan TE, Lewis PM, Borchelt J, Chakrabarty DR. Intracerebral Expression of AAV-APOE4 Is Not Sufficient to Alter Tau Burden in Two Distinct Models of Tauopathy. Mol Neurobiol. 2020;57:1986–2001. doi: 10.1007/s12035-019-01859-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Rosenberg JB, Kaplitt MG, De BP, Chen A, Flagiello T, Salami C, Pey E, Zhao L, Ricart Arbona RJ, Monette S, et al. AAVrh.10-Mediated APOE2 Central Nervous System Gene Therapy for APOE4-Associated Alzheimer’s Disease. Hum Gene Ther Clin Dev. 2018;29:24–47. doi: 10.1089/humc.2017.231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Ridolfi B, Abdel-Haq H. Neurodegenerative Disorders Treatment: The MicroRNA Role. Curr Gene Ther. 2017;17:327–63. doi: 10.2174/1566523218666180119120726. [DOI] [PubMed] [Google Scholar]
- 227.Walgrave H, Zhou L, De Strooper B, Salta E. The promise of microRNA-based therapies in Alzheimer’s disease: challenges and perspectives. Mol neurodegeneration. 2021;16:76. doi: 10.1186/s13024-021-00496-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.He D, Tan J, Zhang J. miR-137 attenuates Abeta-induced neurotoxicity through inactivation of NF-kappaB pathway by targeting TNFAIP1 in Neuro2a cells. Biochem Biophys Res Commun. 2017;490:941–7. doi: 10.1016/j.bbrc.2017.06.144. [DOI] [PubMed] [Google Scholar]
- 229.Wu BW, Wu MS, Guo JD. Effects of microRNA-10a on synapse remodeling in hippocampal neurons and neuronal cell proliferation and apoptosis through the BDNF-TrkB signaling pathway in a rat model of Alzheimer’s disease. J Cell Physiol. 2018;233:5281–92. doi: 10.1002/jcp.26328. [DOI] [PubMed] [Google Scholar]
- 230.Guedes JR, Santana I, Cunha C, Duro D, Almeida MR, Cardoso AM, de Lima MC, Cardoso AL. MicroRNA deregulation and chemotaxis and phagocytosis impairment in Alzheimer’s disease. Alzheimers Dement (Amst) 2016;3:7–17. doi: 10.1016/j.dadm.2015.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.Smith PY, Hernandez-Rapp J, Jolivette F, Lecours C, Bisht K, Goupil C, Dorval V, Parsi S, Morin F, Planel E, et al. miR-132/212 deficiency impairs tau metabolism and promotes pathological aggregation in vivo. Hum Mol Genet. 2015;24:6721–35. doi: 10.1093/hmg/ddv377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Jan A, Karasinska JM, Kang MH, de Haan W, Ruddle P, Kaur A, Connolly C, Leavitt BR, Sorensen PH, Hayden MR. Direct intracerebral delivery of a miR-33 antisense oligonucleotide into mouse brain increases brain ABCA1 expression. [Corrected] Neurosci Lett. 2015;598:66–72. doi: 10.1016/j.neulet.2015.05.007. [DOI] [PubMed] [Google Scholar]
- 233.Kim J, Yoon H, Horie T, Burchett JM, Restivo JL, Rotllan N, Ramirez CM, Verghese PB, Ihara M, Hoe HS, et al. microRNA-33 Regulates ApoE Lipidation and Amyloid-beta Metabolism in the Brain. J neuroscience: official J Soc Neurosci. 2015;35:14717–26. doi: 10.1523/JNEUROSCI.2053-15.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Teter B, LaDu MJ, Sullivan PM, Frautschy SA, Cole GM. Apolipoprotein E isotype-dependent modulation of microRNA-146a in plasma and brain. NeuroReport. 2016;27:791–5. doi: 10.1097/WNR.0000000000000608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.Chernick D, Ortiz-Valle S, Jeong A, Qu W, Li L. Peripheral versus central nervous system APOE in Alzheimer’s disease: Interplay across the blood-brain barrier. Neurosci Lett. 2019;708:134306. doi: 10.1016/j.neulet.2019.134306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Nielsen HM, Chen K, Lee W, Chen Y, Bauer RJ, Reiman E, Caselli R, Bu G. Peripheral apoE isoform levels in cognitively normal APOE ε3/ε4 individuals are associated with regional gray matter volume and cerebral glucose metabolism. Alzheimers Res Ther. 2017;9:5. doi: 10.1186/s13195-016-0231-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Lane-Donovan C, Wong WM, Durakoglugil MS, Wasser CR, Jiang S, Xian X, Herz J. Genetic Restoration of Plasma ApoE Improves Cognition and Partially Restores Synaptic Defects in ApoE-Deficient Mice. J neuroscience: official J Soc Neurosci. 2016;36:10141–50. doi: 10.1523/JNEUROSCI.1054-16.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Huynh TV, Wang C, Tran AC, Tabor GT, Mahan TE, Francis CM, Finn MB, Spellman R, Manis M, Tanzi RE, et al. Lack of hepatic apoE does not influence early Abeta deposition: observations from a new APOE knock-in model. Mol neurodegeneration. 2019;14:37. doi: 10.1186/s13024-019-0337-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 239.Baker-Nigh AT, Mawuenyega KG, Bollinger JG, Ovod V, Kasten T, Franklin EE, Liao F, Jiang H, Holtzman D, Cairns NJ, et al. Human Central Nervous System (CNS) ApoE Isoforms Are Increased by Age, Differentially Altered by Amyloidosis, and Relative Amounts Reversed in the CNS Compared with Plasma. J Biol Chem. 2016;291:27204–18. doi: 10.1074/jbc.M116.721779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.Edlund AK, Chen K, Lee W, Protas H, Su Y, Reiman E, Caselli R, Nielsen HM. Plasma Apolipoprotein E3 and Glucose Levels Are Associated in APOE ɛ3/ɛ4 Carriers. J Alzheimer’s disease: JAD. 2021;81:339–54. doi: 10.3233/JAD-210065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Ben Khedher MR, Haddad M, Laurin D, Ramassamy C. Apolipoprotein E4-driven effects on inflammatory and neurotrophic factors in peripheral extracellular vesicles from cognitively impaired, no dementia participants who converted to Alzheimer’s disease. Alzheimers Dement (N Y) 2021;7:e12124. doi: 10.1002/trc2.12124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242.Hirose S, Hioki Y, Miyashita H, Hirade N, Yoshitake J, Shibata T, Kikuchi R, Matsushita T, Chikazawa M, Itakura M, et al. Apolipoprotein E binds to and reduces serum levels of DNA-mimicking, pyrrolated proteins. J Biol Chem. 2019;294:11035–45. doi: 10.1074/jbc.RA118.006629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.Boada M, López OL, Olazarán J, Núñez L, Pfeffer M, Paricio M, Lorites J, Piñol-Ripoll G, Gámez JE, Anaya F, et al. A randomized, controlled clinical trial of plasma exchange with albumin replacement for Alzheimer’s disease: Primary results of the AMBAR Study. Alzheimers Dement. 2020;16:1412–25. doi: 10.1002/alz.12137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244.Loeffler DA. AMBAR, an Encouraging Alzheimer’s Trial That Raises Questions. Front Neurol. 2020;11:459. doi: 10.3389/fneur.2020.00459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Sha SJ, Deutsch GK, Tian L, Richardson K, Coburn M, Gaudioso JL, Marcal T, Solomon E, Boumis A, Bet A, et al. Safety, Tolerability, and Feasibility of Young Plasma Infusion in the Plasma for Alzheimer Symptom Amelioration Study: A Randomized Clinical Trial. JAMA Neurol. 2019;76:35–40. doi: 10.1001/jamaneurol.2018.3288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246.Tokgöz S, Claassen J. Exercise as Potential Therapeutic Target to Modulate Alzheimer’s Disease Pathology in APOE ε4 Carriers: A Systematic Review. Cardiol therapy. 2021;10:67–88. doi: 10.1007/s40119-020-00209-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Cancela-Carral JM, López-Rodríguez A, Mollinedo-Cardalda I. Effect of physical exercise on cognitive function in older adults’ carriers versus noncarriers of apolipoprotein E4: systematic review and meta-analysis. J Exerc rehabilitation. 2021;17:69–80. doi: 10.12965/jer.2142130.065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248.Fernández-Matarrubia M, Goni L, Rognoni T, Razquin C, Fernández-Lázaro CI, Bes-Rastrollo M, Martínez-González M, Toledo E. An Active Lifestyle Is Associated with Better Cognitive Function Over Time in APOE ɛ4 Non-Carriers. J Alzheimer’s disease: JAD. 2021;79:1257–68. doi: 10.3233/JAD-201090. [DOI] [PubMed] [Google Scholar]
- 249.Martin-Willett R, Morris B, Wilcox R, Giordano G, Andrews-Hanna J, Banich M, Bryan AB. The influence of a 16-week exercise program, APOE status, and age on executive function task performance: A randomized trial. Exp Gerontol. 2021;152:111431. doi: 10.1016/j.exger.2021.111431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250.Deeny SP, Poeppel D, Zimmerman JB, Roth SM, Brandauer J, Witkowski S, Hearn JW, Ludlow AT, Contreras-Vidal JL, Brandt J, Hatfield BD. Exercise, APOE, and working memory: MEG and behavioral evidence for benefit of exercise in epsilon4 carriers. Biol Psychol. 2008;78:179–87. doi: 10.1016/j.biopsycho.2008.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251.Colovati MES, Novais IP, Zampol M, Mendes GD, Cernach MCS, Zanesco A. Interaction between physical exercise and APOE gene polymorphism on cognitive function in older people. Brazilian J Med Biol research = Revista brasileira de pesquisas medicas e biologicas. 2020;54:e10098. doi: 10.1590/1414-431X202010098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 252.Jensen CS, Simonsen AH, Siersma V, Beyer N, Frederiksen KS, Gottrup H, Hoffman K, Høgh P, Frikke-Schmidt R, Sobol NA, et al. Patients with Alzheimer’s disease who carry the APOE ε4 allele benefit more from physical exercise. Alzheimers Dement (N Y) 2019;5:99–106. doi: 10.1016/j.trci.2019.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Kaufman CS, Honea RA, Pleen J, Lepping RJ, Watts A, Morris JK, Billinger SA, Burns JM, Vidoni ED. Aerobic exercise improves hippocampal blood flow for hypertensive Apolipoprotein E4 carriers. Journal of cerebral blood flow and metabolism: official journal of the International Society of Cerebral Blood Flow and Metabolism 2021:271678x21990342. [DOI] [PMC free article] [PubMed]
- 254.de Frutos-Lucas J, Cuesta P, López-Sanz D, Peral-Suárez Á, Cuadrado-Soto E, Ramírez-Toraño F, Brown BM, Serrano JM, Laws SM, Rodríguez-Rojo IC, et al. The relationship between physical activity, apolipoprotein E ε4 carriage, and brain health. Alzheimers Res Ther. 2020;12:48. doi: 10.1186/s13195-020-00608-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255.Piccarducci R, Daniele S, Polini B, Carpi S, Chico L, Fusi J, Baldacci F, Siciliano G, Bonuccelli U, Nieri P, et al: Apolipoprotein E Polymorphism and Oxidative Stress in Human Peripheral Blood Cells: Can Physical Activity Reactivate the Proteasome System through Epigenetic Mechanisms? Oxidative medicine and cellular longevity 2021, 2021:8869849. [DOI] [PMC free article] [PubMed]
- 256.Rusek M, Pluta R, Ułamek-Kozioł M, Czuczwar SJ. Ketogenic Diet in Alzheimer’s Disease. International journal of molecular sciences 2019, 20. [DOI] [PMC free article] [PubMed]
- 257.Neth BJ, Mintz A, Whitlow C, Jung Y, Solingapuram Sai K, Register TC, Kellar D, Lockhart SN, Hoscheidt S, Maldjian J, et al. Modified ketogenic diet is associated with improved cerebrospinal fluid biomarker profile, cerebral perfusion, and cerebral ketone body uptake in older adults at risk for Alzheimer’s disease: a pilot study. Neurobiol Aging. 2020;86:54–63. doi: 10.1016/j.neurobiolaging.2019.09.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258.Yassine HN, Finch CE. APOE Alleles and Diet in Brain Aging and Alzheimer’s Disease. Front Aging Neurosci. 2020;12:150. doi: 10.3389/fnagi.2020.00150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 259.Broom GM, Shaw IC, Rucklidge JJ. The ketogenic diet as a potential treatment and prevention strategy for Alzheimer’s disease. Nutr (Burbank Los Angeles Cty Calif) 2019;60:118–21. doi: 10.1016/j.nut.2018.10.003. [DOI] [PubMed] [Google Scholar]
- 260.Morrill SJ, Gibas KJ. Ketogenic diet rescues cognition in ApoE4 + patient with mild Alzheimer’s disease: A case study. Diabetes & metabolic syndrome. 2019;13:1187–91. doi: 10.1016/j.dsx.2019.01.035. [DOI] [PubMed] [Google Scholar]
- 261.Stoykovich S, Gibas K. APOE ε4, the door to insulin-resistant dyslipidemia and brain fog? A case study. Alzheimers Dement (Amst) 2019;11:264–9. doi: 10.1016/j.dadm.2019.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262.Grammatikopoulou MG, Goulis DG, Gkiouras K, Theodoridis X, Gkouskou KK, Evangeliou A, Dardiotis E, Bogdanos DP. To Keto or Not to Keto? A Systematic Review of Randomized Controlled Trials Assessing the Effects of Ketogenic Therapy on Alzheimer Disease. Adv Nutr (Bethesda Md) 2020;11:1583–602. doi: 10.1093/advances/nmaa073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263.Uddin MS, Kabir MT, Tewari D, Al Mamun A, Barreto GE, Bungau SG, Bin-Jumah MN, Abdel-Daim MM, Ashraf GM. Emerging Therapeutic Promise of Ketogenic Diet to Attenuate Neuropathological Alterations in Alzheimer’s Disease. Mol Neurobiol. 2020;57:4961–77. doi: 10.1007/s12035-020-02065-3. [DOI] [PubMed] [Google Scholar]
- 264.Norwitz NG, Saif N, Ariza IE, Isaacson RS. Precision Nutrition for Alzheimer’s Prevention in ApoE4 Carriers. Nutrients 2021, 13. [DOI] [PMC free article] [PubMed]
- 265.Chouinard-Watkins R, Rioux-Perreault C, Fortier M, Tremblay-Mercier J, Zhang Y, Lawrence P, Vohl MC, Perron P, Lorrain D, Brenna JT, et al. Disturbance in uniformly 13 C-labelled DHA metabolism in elderly human subjects carrying the apoE ε4 allele. Br J Nutr. 2013;110:1751–9. doi: 10.1017/S0007114513001268. [DOI] [PubMed] [Google Scholar]
- 266.Nock TG, Chouinard-Watkins R, Plourde M. Carriers of an apolipoprotein E epsilon 4 allele are more vulnerable to a dietary deficiency in omega-3 fatty acids and cognitive decline. Biochim et Biophys acta Mol cell biology lipids. 2017;1862:1068–78. doi: 10.1016/j.bbalip.2017.07.004. [DOI] [PubMed] [Google Scholar]
- 267.Tomaszewski N, He X, Solomon V, Lee M, Mack WJ, Quinn JF, Braskie MN, Yassine HN. Effect of APOE Genotype on Plasma Docosahexaenoic Acid (DHA), Eicosapentaenoic Acid, Arachidonic Acid, and Hippocampal Volume in the Alzheimer’s Disease Cooperative Study-Sponsored DHA Clinical Trial. J Alzheimer’s disease: JAD. 2020;74:975–90. doi: 10.3233/JAD-191017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 268.Arellanes IC, Choe N, Solomon V, He X, Kavin B, Martinez AE, Kono N, Buennagel DP, Hazra N, Kim G, et al. Brain delivery of supplemental docosahexaenoic acid (DHA): A randomized placebo-controlled clinical trial. EBioMedicine. 2020;59:102883. doi: 10.1016/j.ebiom.2020.102883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 269.Patrick RP. Role of phosphatidylcholine-DHA in preventing APOE4-associated Alzheimer’s disease. FASEB journal: official publication of the Federation of American Societies for Experimental Biology. 2019;33:1554–64. doi: 10.1096/fj.201801412R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 270.Coughlan G, Larsen R, Kim M, White D, Gillings R, Irvine M, Scholey A, Cohen N, Legido-Quigley C, Hornberger M, Minihane AM. APOE ε4 alters associations between docosahexaenoic acid and preclinical markers of Alzheimer’s disease. Brain Commun. 2021;3:fcab085. doi: 10.1093/braincomms/fcab085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 271.Wang C, Holtzman DM. Bidirectional relationship between sleep and Alzheimer’s disease: role of amyloid, tau, and other factors. Neuropsychopharmacology: official publication of the American College of Neuropsychopharmacology. 2020;45:104–20. doi: 10.1038/s41386-019-0478-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 272.Lloret MA, Cervera-Ferri A, Nepomuceno M, Monllor P, Esteve D, Lloret A. Is Sleep Disruption a Cause or Consequence of Alzheimer’s Disease? Reviewing Its Possible Role as a Biomarker. International journal of molecular sciences 2020, 21. [DOI] [PMC free article] [PubMed]
- 273.Koo KYG, Schweizer TA, Fischer CE, Munoz DG. Abnormal Sleep Behaviours Across the Spectrum of Alzheimer’s Disease Severity: Influence of APOE Genotypes and Lewy Bodies. Curr Alzheimer Res. 2019;16:243–50. doi: 10.2174/1567205016666190103161034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 274.Hita-Yañez E, Atienza M, Gil-Neciga E, Cantero JL. Disturbed sleep patterns in elders with mild cognitive impairment: the role of memory decline and ApoE ε4 genotype. Curr Alzheimer Res. 2012;9:290–7. doi: 10.2174/156720512800107609. [DOI] [PubMed] [Google Scholar]
- 275.Drogos LL, Gill SJ, Tyndall AV, Raneri JK, Parboosingh JS, Naef A, Guild KD, Eskes G, Hanly PJ, Poulin MJ. Evidence of association between sleep quality and APOE ε4 in healthy older adults: A pilot study. Neurology. 2016;87:1836–42. doi: 10.1212/WNL.0000000000003255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 276.Basta M, Zaganas I, Simos P, Koutentaki E, Dimovasili C, Mathioudakis L, Bourbouli M, Panagiotakis S, Kapetanaki S, Vgontzas A, Apolipoprotein E. ɛ4 (APOE ɛ4) Allele is Associated with Long Sleep Duration Among Elderly with Cognitive Impairment. J Alzheimer’s disease: JAD. 2021;79:763–71. doi: 10.3233/JAD-200958. [DOI] [PubMed] [Google Scholar]
- 277.Tranah GJ, Yaffe K, Nievergelt CM, Parimi N, Glymour MM, Ensrud KE, Cauley JA, Ancoli-Israel S, Mariani S, Redline S, Stone KL. APOEε4 and slow wave sleep in older adults. PLoS ONE. 2018;13:e0191281. doi: 10.1371/journal.pone.0191281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 278.Grimmer T, Laub T, Hapfelmeier A, Eisele T, Fatke B, Hölzle P, Lüscher S, Parchmann AM, Rentrop M, Schwerthöffer D, et al. The overnight reduction of amyloid β 1–42 plasma levels is diminished by the extent of sleep fragmentation, sAPP-β, and APOE ε4 in psychiatrists on call. Alzheimers Dement. 2020;16:759–69. doi: 10.1002/alz.12072. [DOI] [PubMed] [Google Scholar]
- 279.Pappolla M, Bozner P, Soto C, Shao H, Robakis NK, Zagorski M, Frangione B, Ghiso J. Inhibition of Alzheimer beta-fibrillogenesis by melatonin. J Biol Chem. 1998;273:7185–8. doi: 10.1074/jbc.273.13.7185. [DOI] [PubMed] [Google Scholar]
- 280.Poeggeler B, Miravalle L, Zagorski MG, Wisniewski T, Chyan YJ, Zhang Y, Shao H, Bryant-Thomas T, Vidal R, Frangione B, et al. Melatonin reverses the profibrillogenic activity of apolipoprotein E4 on the Alzheimer amyloid Abeta peptide. Biochemistry. 2001;40:14995–5001. doi: 10.1021/bi0114269. [DOI] [PubMed] [Google Scholar]
- 281.Shukla M, Govitrapong P, Boontem P, Reiter RJ, Satayavivad J. Mechanisms of Melatonin in Alleviating Alzheimer’s Disease. Curr Neuropharmacol. 2017;15:1010–31. doi: 10.2174/1570159X15666170313123454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 282.Liu RY, Zhou JN, van Heerikhuize J, Hofman MA, Swaab DF. Decreased melatonin levels in postmortem cerebrospinal fluid in relation to aging, Alzheimer’s disease, and apolipoprotein E-epsilon4/4 genotype. J Clin Endocrinol Metab. 1999;84:323–7. doi: 10.1210/jcem.84.1.5394. [DOI] [PubMed] [Google Scholar]
- 283.Xu J, Wang LL, Dammer EB, Li CB, Xu G, Chen SD, Wang G. Melatonin for sleep disorders and cognition in dementia: a meta-analysis of randomized controlled trials. Am J Alzheimers Dis Other Demen. 2015;30:439–47. doi: 10.1177/1533317514568005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 284.Li Y, Zhang J, Wan J, Liu A, Sun J. Melatonin regulates Aβ production/clearance balance and Aβ neurotoxicity: A potential therapeutic molecule for Alzheimer’s disease. Biomed pharmacotherapy = Biomedecine pharmacotherapie. 2020;132:110887. doi: 10.1016/j.biopha.2020.110887. [DOI] [PubMed] [Google Scholar]
- 285.Nous A, Engelborghs S, Smolders I. Melatonin levels in the Alzheimer’s disease continuum: a systematic review. Alzheimers Res Ther. 2021;13:52. doi: 10.1186/s13195-021-00788-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 286.Hwang JY, Byun MS, Choe YM, Lee JH, Yi D, Choi JW, Hwang SH, Lee YJ, Lee DY. Moderating effect of APOE ε4 on the relationship between sleep-wake cycle and brain β-amyloid. Neurology. 2018;90:e1167–73. doi: 10.1212/WNL.0000000000005193. [DOI] [PubMed] [Google Scholar]
- 287.Bulgart HR, Neczypor EW, Wold LE, Mackos AR. Microbial involvement in Alzheimer disease development and progression. Mol neurodegeneration. 2020;15:42. doi: 10.1186/s13024-020-00378-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 288.Parikh IJ, Estus JL, Zajac DJ, Malik M, Maldonado Weng J, Tai LM, Chlipala GE, LaDu MJ, Green SJ, Estus S. Murine Gut Microbiome Association With APOE Alleles. Front Immunol. 2020;11:200. doi: 10.3389/fimmu.2020.00200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 289.Weng YJ, Gan HY, Li X, Huang Y, Li ZC, Deng HM, Chen SZ, Zhou Y, Wang LS, Han YP, et al. Correlation of diet, microbiota and metabolite networks in inflammatory bowel disease. J Dig Dis. 2019;20:447–59. doi: 10.1111/1751-2980.12795. [DOI] [PubMed] [Google Scholar]
- 290.Maldonado Weng J, Parikh I, Naqib A, York J, Green SJ, Estus S, LaDu MJ. Synergistic effects of APOE and sex on the gut microbiome of young EFAD transgenic mice. Mol neurodegeneration. 2019;14:47. doi: 10.1186/s13024-019-0352-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 291.Hou M, Xu G, Ran M, Luo W, Wang H. APOE-ε4 Carrier Status and Gut Microbiota Dysbiosis in Patients With Alzheimer Disease. Front NeuroSci. 2021;15:619051. doi: 10.3389/fnins.2021.619051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 292.Hoffman JD, Yanckello LM, Chlipala G, Hammond TC, McCulloch SD, Parikh I, Sun S, Morganti JM, Green SJ, Lin AL. Dietary inulin alters the gut microbiome, enhances systemic metabolism and reduces neuroinflammation in an APOE4 mouse model. PLoS ONE. 2019;14:e0221828. doi: 10.1371/journal.pone.0221828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 293.Yuan T, Chu C, Shi R, Cui T, Zhang X, Zhao Y, Shi X, Hui Y, Pan J, Qian R, et al. ApoE-Dependent Protective Effects of Sesamol on High-Fat Diet-Induced Behavioral Disorders: Regulation of the Microbiome-Gut-Brain Axis. J Agric Food Chem. 2019;67:6190–201. doi: 10.1021/acs.jafc.9b01436. [DOI] [PubMed] [Google Scholar]
- 294.Nagpal R, Neth BJ, Wang S, Mishra SP, Craft S, Yadav H. Gut mycobiome and its interaction with diet, gut bacteria and alzheimer’s disease markers in subjects with mild cognitive impairment: A pilot study. EBioMedicine. 2020;59:102950. doi: 10.1016/j.ebiom.2020.102950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 295.Carranza-Naval MJ, Vargas-Soria M, Hierro-Bujalance C, Baena-Nieto G, Garcia-Alloza M, Infante-Garcia C, Del Marco A. Alzheimer’s Disease and Diabetes: Role of Diet, Microbiota and Inflammation in Preclinical Models. Biomolecules 2021, 11. [DOI] [PMC free article] [PubMed]
- 296.Panitch R, Hu J, Chung J, Zhu C, Meng G, Xia W, Bennett DA, Lunetta KL, Ikezu T, Au R, et al: Integrative brain transcriptome analysis links complement component 4 and HSPA2 to the APOE ε2 protective effect in Alzheimer disease. Molecular psychiatry 2021. [DOI] [PMC free article] [PubMed]
- 297.Olah M, Menon V, Habib N, Taga MF, Ma Y, Yung CJ, Cimpean M, Khairallah A, Coronas-Samano G, Sankowski R, et al. Single cell RNA sequencing of human microglia uncovers a subset associated with Alzheimer’s disease. Nat Commun. 2020;11:6129. doi: 10.1038/s41467-020-19737-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 298.Nguyen AT, Wang K, Hu G, Wang X, Miao Z, Azevedo JA, Suh E, Van Deerlin VM, Choi D, Roeder K, et al. APOE and TREM2 regulate amyloid-responsive microglia in Alzheimer’s disease. Acta Neuropathol. 2020;140:477–93. doi: 10.1007/s00401-020-02200-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 299.Gerrits E, Brouwer N, Kooistra SM, Woodbury ME, Vermeiren Y, Lambourne M, Mulder J, Kummer M, Möller T, Biber K, et al. Distinct amyloid-β and tau-associated microglia profiles in Alzheimer’s disease. Acta Neuropathol. 2021;141:681–96. doi: 10.1007/s00401-021-02263-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 300.Farmer BC, Williams HC, Devanney NA, Piron MA, Nation GK, Carter DJ, Walsh AE, Khanal R, Young LEA, Kluemper JC, et al. APOΕ4 lowers energy expenditure in females and impairs glucose oxidation by increasing flux through aerobic glycolysis. Mol neurodegeneration. 2021;16:62. doi: 10.1186/s13024-021-00483-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 301.Sala Frigerio C, Wolfs L, Fattorelli N, Thrupp N, Voytyuk I, Schmidt I, Mancuso R, Chen WT, Woodbury ME, Srivastava G, et al. The Major Risk Factors for Alzheimer’s Disease: Age, Sex, and Genes Modulate the Microglia Response to Aβ Plaques. Cell Rep. 2019;27:1293–306.e1296. doi: 10.1016/j.celrep.2019.03.099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 302.Patel T, Carnwath TP, Wang X, Allen M, Lincoln SJ, Lewis-Tuffin LJ, Quicksall ZS, Lin S, Tutor-New FQ, Ho CCG, et al. Transcriptional landscape of human microglia implicates age, sex, and APOE-related immunometabolic pathway perturbations. Aging Cell. 2022;21:e13606. doi: 10.1111/acel.13606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 303.Madrid L, Moreno-Grau S, Ahmad S, González-Pérez A, de Rojas I, Xia R, Martino Adami PV, García-González P, Kleineidam L, Yang Q, et al. Multiomics integrative analysis identifies APOE allele-specific blood biomarkers associated to Alzheimer’s disease etiopathogenesis. Aging. 2021;13:9277–329. doi: 10.18632/aging.202950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 304.Johnson ECB, Carter EK, Dammer EB, Duong DM, Gerasimov ES, Liu Y, Liu J, Betarbet R, Ping L, Yin L, et al. Large-scale deep multi-layer analysis of Alzheimer’s disease brain reveals strong proteomic disease-related changes not observed at the RNA level. Nat Neurosci. 2022;25:213–25. doi: 10.1038/s41593-021-00999-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 305.Lim WLF, Huynh K, Chatterjee P, Martins I, Jayawardana KS, Giles C, Mellett NA, Laws SM, Bush AI, Rowe CC, et al. Relationships Between Plasma Lipids Species, Gender, Risk Factors, and Alzheimer’s Disease. J Alzheimer’s disease: JAD. 2020;76:303–15. doi: 10.3233/JAD-191304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 306.Wong MWK, Braidy N, Crawford J, Pickford R, Song F, Mather KA, Attia J, Brodaty H, Sachdev P, Poljak A. APOE Genotype Differentially Modulates Plasma Lipids in Healthy Older Individuals, with Relevance to Brain Health. J Alzheimer’s disease: JAD. 2019;72:703–16. doi: 10.3233/JAD-190524. [DOI] [PubMed] [Google Scholar]
- 307.Li H, Dhanasekaran P, Alexander ET, Rader DJ, Phillips MC, Lund-Katz S. Molecular mechanisms responsible for the differential effects of apoE3 and apoE4 on plasma lipoprotein-cholesterol levels. Arterioscler Thromb Vasc Biol. 2013;33:687–93. doi: 10.1161/ATVBAHA.112.301193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 308.Peña-Bautista C, Roca M, López-Cuevas R, Baquero M, Vento M, Cháfer-Pericás C. Metabolomics study to identify plasma biomarkers in alzheimer disease: ApoE genotype effect. J Pharm Biomed Anal. 2020;180:113088. doi: 10.1016/j.jpba.2019.113088. [DOI] [PubMed] [Google Scholar]
- 309.Karjalainen JP, Mononen N, Hutri-Kähönen N, Lehtimäki M, Juonala M, Ala-Korpela M, Kähönen M, Raitakari O, Lehtimäki T. The effect of apolipoprotein E polymorphism on serum metabolome - a population-based 10-year follow-up study. Sci Rep. 2019;9:458. doi: 10.1038/s41598-018-36450-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 310.Ferguson AC, Tank R, Lyall LM, Ward J, Celis-Morales C, Strawbridge R, Ho F, Whelan CD, Gill J, Welsh P, et al. Alzheimer’s Disease Susceptibility Gene Apolipoprotein E (APOE) and Blood Biomarkers in UK Biobank (N=395,769) J Alzheimer’s disease: JAD. 2020;76:1541–51. doi: 10.3233/JAD-200338. [DOI] [PubMed] [Google Scholar]
- 311.Kuo CL, Pilling LC, Atkins JL, Kuchel GA, Melzer D. ApoE e2 and aging-related outcomes in 379,000 UK Biobank participants. Aging. 2020;12:12222–33. doi: 10.18632/aging.103405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 312.Butt OH, Long JM, Henson RL, Herries E, Sutphen CL, Fagan AM, Cruchaga C, Ladenson JH, Holtzman DM, Morris JC, et al. Cognitively normal APOE ε4 carriers have specific elevation of CSF SNAP-25. Neurobiol Aging. 2021;102:64–72. doi: 10.1016/j.neurobiolaging.2021.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 313.Salvadó G, Grothe MJ, Groot C, Moscoso A, Schöll M, Gispert JD, Ossenkoppele R. Differential associations of APOE-ε2 and APOE-ε4 alleles with PET-measured amyloid-β and tau deposition in older individuals without dementia. Eur J Nucl Med Mol Imaging. 2021;48:2212–24. doi: 10.1007/s00259-021-05192-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 314.Wang L, Zhang M, Wang Q, Jiang X, Li K, Liu J. APOE ε4 Allele Is Associated with Elevated Levels of CSF VILIP-1 in Preclinical Alzheimer’s Disease. Neuropsychiatr Dis Treat. 2020;16:923–31. doi: 10.2147/NDT.S235395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 315.Ahmad S, Milan MDC, Hansson O, Demirkan A, Agustin R, Sáez ME, Giagtzoglou N, Cabrera-Socorro A, Bakker MHM, Ramirez A, et al. CDH6 and HAGH protein levels in plasma associate with Alzheimer’s disease in APOE ε4 carriers. Sci Rep. 2020;10:8233. doi: 10.1038/s41598-020-65038-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 316.Soares HD, Potter WZ, Pickering E, Kuhn M, Immermann FW, Shera DM, Ferm M, Dean RA, Simon AJ, Swenson F, et al. Plasma biomarkers associated with the apolipoprotein E genotype and Alzheimer disease. Arch Neurol. 2012;69:1310–7. doi: 10.1001/archneurol.2012.1070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 317.Ma QL, Teng E, Zuo X, Jones M, Teter B, Zhao EY, Zhu C, Bilousova T, Gylys KH, Apostolova LG, et al. Neuronal pentraxin 1: A synaptic-derived plasma biomarker in Alzheimer’s disease. Neurobiol Dis. 2018;114:120–8. doi: 10.1016/j.nbd.2018.02.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.