

Abstract
Background
Although older adults are at a high risk of severe or critical Covid-19, there are many cases of unvaccinated centenarians who had a silent infection or recovered from mild or moderate Covid-19. We studied three Brazilian supercentenarians, older than 110 years, who survived Covid-19 in 2020 before being vaccinated.
Results
Despite their advanced age, humoral immune response analysis showed that these individuals displayed robust levels of IgG and neutralizing antibodies (NAbs) against SARS-CoV-2. Enrichment of plasma proteins and metabolites related to innate immune response and host defense was also observed. None presented autoantibodies (auto-Abs) to type I interferon (IFN). Furthermore, these supercentenarians do not carry rare variants in genes underlying the known inborn errors of immunity, including particular inborn errors of type I IFN.
Conclusion
These observations suggest that their Covid-19 resilience might be a combination of their genetic background and their innate and adaptive immunity.
Supplementary Information
The online version contains supplementary material available at 10.1186/s12979-022-00310-y.
Keywords: Covid-19, Supercentenarians, SARS-CoV-2, Elderly
Background
The emergence of the Covid-19 pandemic resulted in more than six million deaths worldwide, with a higher risk for older adults and people with comorbidities to develop severe cases of the disease [1–4]. Covid-19 deaths of individuals over 60 represented over 70% of total Covid-19-related deaths in Brazil [5]. In the United States, about 80% of Covid-19 deaths have been among people older than 65 years [6]. The risk of dying from Covid-19 for an individual aged 85 years or more is 340 higher than for young adults (< 30 years old) [7].
One of the leading hypotheses for the higher Covid-19 severity in older people is a decrease in the immune response that occurs with aging [8]. The immunosenescence phenomenon is associated with significant changes in cytokine patterns and activation of inflammatory pathways, which result in the dysfunction of innate and adaptive immune responses [9, 10]. The immune cells’ senescence significantly contributes to immunity decline [11]. The thymus degenerates gradually with aging, resulting in a significant loss of diversity of the T cell repertoire, depletion, and/or diminished function of mature lymphocytes in secondary lymphoid tissues [12, 13]. Such a decline in immunity is responsible for higher susceptibility to infectious diseases and a decrease in the effectiveness of vaccinations in elderly cohorts [14, 15].
The chronic physiological stimulation of the immune system during life can establish the inflammaging phenomenon, characterized by a progressive and continuous increase of circulating levels of pro-inflammatory mediators [16, 17]. This pro-inflammatory basal state in the elderly may enhance the release of a large amount of pro-inflammatory cytokines as a response to the SARS-CoV-2 infection, which is directly correlated with lung tissue injury, multi-organ failure, and increased risk of dying from Covid-19 – the cytokine storm phenomenon [10, 18–20]. Also, comorbidities in older individuals, as a consequence of the multiple phenomena associated with organic aging [8], are strongly associated with an increased risk of Covid-19 complications, including sepsis and multiple organ dysfunction [21, 22].
In addition, recent studies reported the presence of pre-existing autoantibodies (auto-Abs) neutralizing type I IFNs in patients with life-threatening Covid-19 pneumonia, which block the antiviral activity of correspondent type I IFNs against SARS-CoV-2 [23]. As these auto-Abs skyrocket after age 65, they may represent an additional risk factor for critical Covid-19, especially in the elderly [24].
Despite that, worldwide reports of unvaccinated centenarians and supercentenarians (105 years or older) recovered from Covid-19 with mild or moderate symptoms called our attention [25–29]. Understanding why some individuals overcome the disease despite aging, such as the nun Ms. Randon, the oldest person already reported to survive Covid-19 at age 116 [30] is of great interest.
Host genotype influences how an individual responds to viral infections. For example, natural resistance to HIV-1 infection has been associated with a specific mutation in the CCR5 gene [31]. In contrast, AIDS progression and many other infectious diseases are associated with specific alleles from the MHC, particularly HLA-B [32, 33]. In the context of Covid-19, the resistance to infection is still being investigated [34]. On the other hand, it has been reported that some genetic variants account for the variability in individuals’ susceptibility to Covid-19 and the severity of the disease. In this sense, a set of genes might explain how supercentenarians (some with comorbidities) overcome the disease without major complications.
Here, we present three cases of Brazilian supercentenarians who recovered from Covid-19 before the vaccination onset, including a 114 years old woman, the second oldest person in the world in this condition. Covid-19 in these three volunteers occurred in 2020 before new SARS-CoV-2 variants were reported in Brazil (especially Gamma variant - P.1). Aiming to enhance our comprehension of the underlying factors contributing to their resistance to the disease, we performed a comprehensive immunogenetic assessment and whole-exome sequencing.
Results
Humoral response against SARS-CoV-2
Serological assays for SARS-CoV-2 RBD IgA, IgG, and IgM were performed through enzyme-linked immunosorbent assay (ELISA) for the Receptor-binding domain (RBD) of the Spike protein, and Nucleocapsid (NP) protein, at least four weeks after Covid-19 initial diagnosis. IgG seroconversion was detected for RBD and NP for all three individuals (Table 1). We must highlight that we performed these immune assays before the volunteers’ vaccination against Covid-19. Neutralization capacity was evaluated, and the asymptomatic individual produced low titers compared to the two who presented moderate symptoms. However, all of them presented titers above 160 (Fig. 1).
Table 1.
Humoral immune-response profile of the presence of binding antibodies and type I autoantibodies of the volunteers.
| BINDING ANTIBODIES AGAINST SARS-CoV-2* | |||||
| Participants | ID 01 | ID 02 | ID 03 | ||
| Covid-19 episode | Symptomatic | Asymptomatic | Symptomatic | ||
|
Specific SARS-CoV-2antibodies (Ratio) |
IgA | NP | - (0.0) | - (0.2) | - (0.9) |
| RBD | - (0.1) | - (0.5) | + (5.4) | ||
| IgM | NP | - (0.3) | - (0.0) | - (0.1) | |
| RBD | - (0.5) | + (1.5) | - (0.7) | ||
| IgG | NP | + (1.3) | + (3.1) | + (9.5) | |
| RBD | + (4.2) | + (3.1) | + (7.8) | ||
| TYPE I IFN AUTOANTIBODIES | |||||
|
Auto-anti IFNS IFN-α2, IFN-β and/or -ω (Ratio) |
- (0.0) | - (0.0) | - (0.0) | ||
Fig. 1.
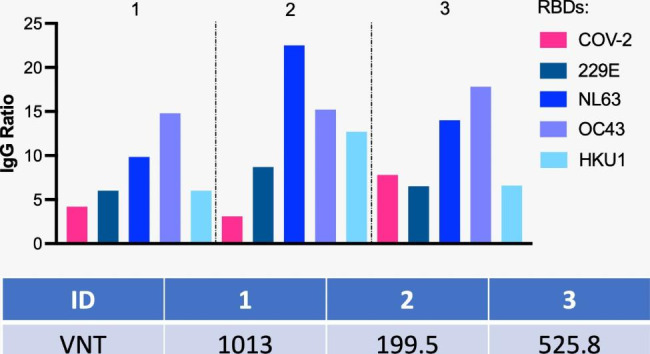
Supercentenarians present higher titers of IgG to RBDs of seasonal coronaviruses than for SARS-CoV-2 and neutralization titers above 160 (the minimum titer initially established by the FDA for convalescent plasma donors) [35]. The numbers represent the 3 individuals. The graph on top indicates IgG ratios for each RBD and the table above shows VNT for SARS-COV-2. VNT - Virus Neutralization Titers. Antibody levels expressed in ratios are shown for SARS-CoV-2 and the four seasonal coronaviruses. Neutralization titers are expressed in dilution. ID 02 was asymptomatic, and IDs 1 and 3 were symptomatic and recovered from Covid-19
We also assessed pre-existing antibodies to RBD of the four common seasonal human coronaviruses (HCoV) α-CoV 229E, α-CoV NL63, β-CoV OC43, and β-CoV HKU1. All supercentenarians displayed IgG antibodies for all four HCoV tested, presenting higher titers for NL63 and OC43 (Fig. 1). Interestingly, IgG normalized values were generally lower for SARS-CoV-2 compared to HCoVs. IgG for SARS-COV-2 RBD did not correlate with neutralization titers among them, meaning that the one who presented the highest titer of RBD was not the one with the highest virus neutralization titer (Fig. 1).
Finally, autoantibodies (auto-Abs) neutralizing type I IFNs assays showed that none of the three supercentenarians had IFN-α, IFN-β, and/or INF-ω auto-Abs, based on negative results of luciferase-based immunoprecipitation (LIPS) assay. Table 1 summarizes the humoral responses assessed, except IgG for RBDs, shown in Fig. 1.
Proteomic and metabolomic plasma analyses
For label-free quantitative proteomics analysis, we compared the 3 supercentenarian’s plasma samples with 3 healthy subjects older than 95 years-old (non-infected). A total of 702 proteins were identified among all the samples analyzed. We performed a t-test to obtain the differentially expressed proteins between these two groups (Fig. 2 A). We found 33 altered proteins, 5 of them in low abundance (IGKV1-6, IGKV2-24, IGKV2-28, GPLD1, IGHV3-49) and 28 were up-regulated. The biological annotation enrichment of proteins which were up-abundant in the 3 supercentenarians showed processes associated with glycolytic pathways (P‑Value = 3.27E-05) and innate immune response like platelet aggregation (P‑Value = 3.75E-05), defense response to fungus (P‑Value = 1.81E-07), antimicrobial response (P‑Value = 5.21E-04).
Fig. 2.
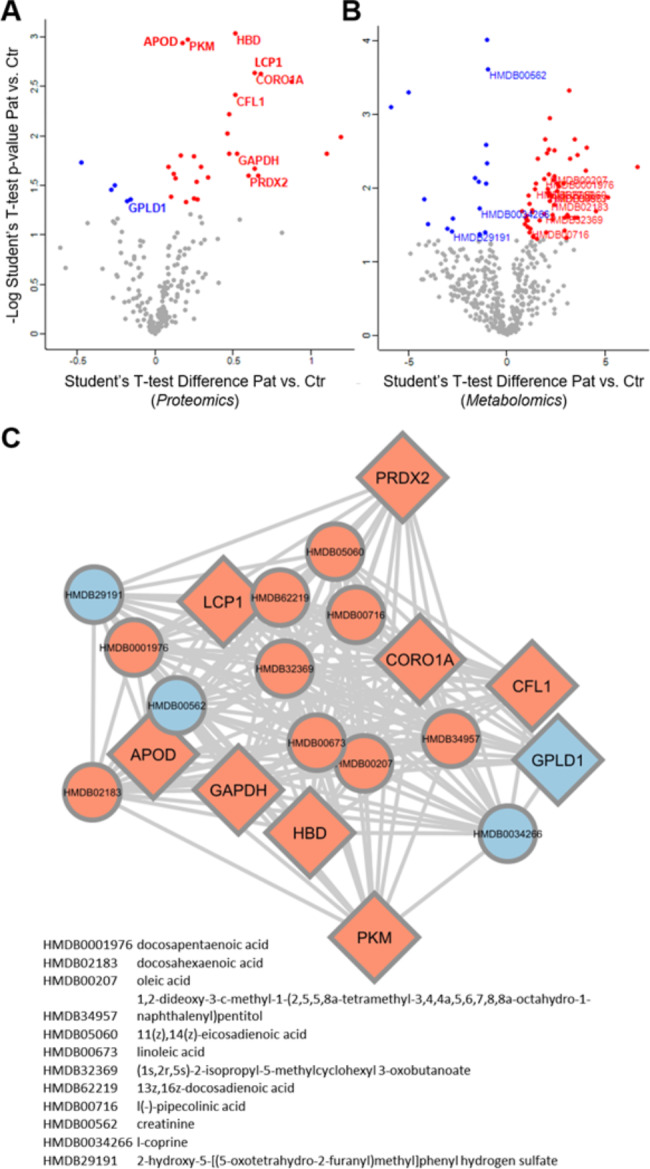
Plasma proteomics and metabolomics analyses of the supercentenarians infected with SARS-CoV-2 compared to healthy elderly subjects (> 95 years-old / non-infected). Volcano plots showing differentially expressed proteins (A) and metabolites (B). Red and blue dots represent the up- and down-regulated proteins/metabolites, respectively. Protein-metabolite interaction network was built with the statistically significant proteins with OmicsAnalyst platform (C). Red and blue nodes denote the up- and down-regulated proteins/metabolites, respectively. Circle and diamond node shapes represent metabolites and proteins, respectively
The untargeted metabolomics approach detected 474 metabolites through positive (291 metabolites) and negative (183 metabolites) ionization modes. The proteomics statistical analysis was also applied to metabolomics (Fig. 2B). In the supercentenarians, 62 metabolites were up-regulated; while 17 metabolites were down-regulated. Enrichment analysis was mainly focused on the up-regulated metabolites, with the lower p-value for the biosynthesis of unsaturated fatty acids (P‑Value = 5.96E-4). Other metabolic pathways such as linoleic acid, purine, and ether lipid metabolisms were also enriched. Down-regulated metabolites presented an enrichment for the primary bile acid biosynthesis pathway. The protein-metabolite network represents the omics dataset’s integration (Fig. 2 C), and their convergence in the main disrupted biological processes and metabolic pathways: glycolysis and the immune system.
Genetic ancestry
The genetic ancestry estimations for these three supercentenarians are shown in Table 2, which corresponds to an average of African ancestry (all volunteers) or Native American (ID 1) much higher than observed in the admixed Brazilian population from the same city [36–38].
Table 2.
Supercentenarians’ continental ancestry composition.
| ID | Age | Sex | European | African | Native American | East Asian |
|---|---|---|---|---|---|---|
| 01 | 114 | Female | 0.34 | 0.43 | 0.23 | 0.00 |
| 02 | 111 | Male | 0.61 | 0.34 | 0.05 | 0.00 |
| 03 | 110 | Male | 0.00 | 1.00 | 0.00 | 0.00 |
| Brazilian average genetic ancestry(average ± standard error) (36) | 0.73 ± 0.26 | 0.18 ± 0.21 | 0.07 ± 0.07 | 0.03 ± 0.16 | ||
Inborn errors of type I IFN immunity (IEI) genes
None of the supercentenarians carry rare variants in genes associated with inborn errors of Toll-like receptor 3 (TLR3) and interferon regulatory factor 7 (IRF7) dependent type I IFN immunity, which underlease life-threatening Covid-19 pneumonia [39, 40]. Also, we did not detect any copy number variation (CNV) in IEI genes for the three supercentenarians.
MHC genes
Due to its central role in the adaptive immune response, genes from the MHC, especially those in the Human leukocyte antigen (HLA) system, are likely candidates to influence infection outcomes. Some HLA alleles were previously associated with severe Covid-19. Here we described three centenarians that recovered from Covid-19, which is a rare condition. Therefore, the small sample size does not allow an in-depth analysis of associated polymorphism. Nevertheless, since these data might be important to understand the mechanisms underlying HLA associations, and other groups evaluating centenarians might be interested in this data, we report the HLA alleles observed for all these centenarians as supplementary data (Table S1).
Discussion
Despite the higher mortality risk associated with aging, analysis of exceptionally resilient supercentenarians may help to elucidate possible resistance mechanisms against SARS-CoV-2 infection in such extreme age.
The serological results of the three supercentenarians showed that they achieved seroconversion of IgG with robust levels against NP and RBD viral proteins. Such observations support the role of the humoral response against SARS-CoV-2. Our data corroborate a study by Foley and Colleagues, which reported higher anti-spike IgG antibody titers in nonagenarians and centenarians exposed to SARS-CoV-2 in a long-term care home (n = 15) than in younger individuals living in the same environment [27].
Neutralization analysis revealed that the asymptomatic individual produced lower titers than the two symptomatic ones. At first glance, this seems contradictory, but only a few asymptomatic individuals produce detectable neutralization titers [41] even with a low viral load during infection. On the other hand, Covid-19 patients admitted to a hospital usually display higher neutralizing antibody levels than mild disease or asymptomatic cases [42]. All three centenarians presented titers higher than 160, which is considered the cutoff for high levels [35].
We also assessed pre-existing antibodies to RBD of the four common seasonal HCoVs α-CoV 229E, α-CoV NL63, β-CoV OC43, and β-CoV HKU1. All supercentenarians displayed IgG antibodies for all four HCoV tested, presenting higher titers for NL63 and OC43. Interestingly, normalized values were generally lower for SARS-CoV-2 compared to HCoV. IgG for SARS-COV-2 RBD did not correlate with neutralization titers among them. The one who presented the highest titer of RBD was not the one with the highest virus neutralization titer.
The literature on the influence of pre-existing humoral immunity to HCoVs in SARS-CoV-2 infection is still controversial. Some studies have pointed out that immunity to the HCoVs has a protective effect on Covid-19 [43]. In contrast, others have described that pre-existing HCoV antibodies may hinder effective immunity against SARS-CoV-2 [44]. In our cohort, the supercentenarians displayed seroreactivity against all four HCoVs, as expected for the elderly, with normalized values showing high titers for all four HCoV. Previous results from a cohort of almost 400 Covid-19-infected individuals showed that HCoV immunity might impact disease severity, and patients with high HCoV reactivity are less likely to require hospitalization [45]. People infected with HCoV viruses during their life (and thus imprinted with that set of antigens/epitopes) would be protected later in life against infections with a related virus, as shown for Influenza [46]. Therefore, these high titers for seasonal coronaviruses might have positively impacted specific responses for SARS-CoV-2.
It is very likely that supercentenarians have been exposed to various pathogens in their life even when children and acquired active immunity making them more prone to defeat SARS-CoV-2. In this sense, it is tempting to speculate that the 1918 H1N1 influenza virus immunity could confer some protection against SARS-CoV-2 infection [47]. The hypothesis is that elderly born before 1918 could have developed immune memory cells able to recognize epitopes antigenically related to the H1N1 virus that would persist even one century later [48]. Interestingly, plasma samples of elderly who survived the Spanish flu pandemic revealed that neutralizing antibodies to the strain 1918 H1N1 influenza derived from isolated B cells have lasted a lifetime [49]. Our volunteers were born before 1918 and there is a report in Brazilian local media that ID03, who has lived his whole life in a region that was the most affected by the 1918 influenza virus, was infected by the virus as a child. It is possible that IDs 01 and 02 might have also been exposed to the Spanish flu in their early life, since both lived in regions affected by the virus but official data are lacking [50, 51].
Besides, a specific and unique subset of CD4 T cells has cytotoxic features in supercentenarians. These cells were accumulated during life exposition to pathogens. They might be considered an adaptation to aging since the immune system needs extra support to eliminate abnormal and infected cells. Such observation corroborates the hypothesis that the original antigenic sin phenomenon [52] could play a significant role in the recovery of the three supercentenarians through a mechanism of immunological memory [53].
It has been reported that circulating auto-Abs neutralizing type I IFNs (IFN-α and/or -ω) were distinctly found in elderly patients with severe Covid-19 and rarely detected in asymptomatic, benign infectious, or healthy individuals. They account for about 20% of critical Covid-19 cases in people over the 80s and total fatal Covid-19 cases [23, 24]. Interestingly, none of the three supercentenarians had neutralized auto-Abs against the type I IFNs, suggesting that they were not at a greater risk for complications in Covid-19 despite their advanced age.
Regarding the plasma proteomic and metabolomic analyses, we were able to compare these 3 supercentenarians with a control group of 3 individuals with comparable age, albeit no older than 110-years, whose serology was negative for Covid-19. Proteomics analysis showed that glycolytic proteins were more abundant in the supercentenarians compared to the control group, which is associated with the infection phisiopatology [54–57]. Krishnan and colleagues used targeted proteomics as well as untargeted metabolomics approaches in plasma samples and cell-line models and discovered that glycolysis and glutaminolysis are essential for virus replication [58]. In addition, during Covid-19 and other viral infections [59–61], there is a reprogramming of the glucose metabolism that overexpress glycolitic enzymes as glyceraldehyde 3-phosphate dehydrogenase - GAPDH (enriched in Fig. 2 C) in non-immune and immune cells, specially the ones involved in innate immunity - which are activated [54, 62]. In parallel, we observed that some processes related to innate immunity were found up-regulated in the plasma of these supercentenarians, demonstrating that they also displayed a first-line of defense capable of effectively neutralizing the infection in addition to their robust adaptive immune responses.
On the other hand, the metabolomics approach showed some up-regulated metabolites in the supercentenarians’ plasma related to fatty acid metabolism, especially the biosynthesis of unsaturated fatty acids. These molecules have a central role in modulating the immune pathways and inflammatory responses [63]. Some studies have described the importance of highly unsaturated fatty acids controlling both inflammation and thrombosis caused by Covid-19 [64–66]. The unsaturated fatty acid also mediates protein complex formation in lipid rafts and thus modulates SARS-CoV-2 entry gateways [67].
The genetic analysis indicated that all these three supercentenarians do not present variants associated with inborn errors of type I IFN immunity (IEI) genes, which is not surprising considering their advanced age without associated diagnosis. Because of the small sample size, conducting an in-depth analysis of polymorphisms associated with their Covid-19 resistance phenotype is not feasible. Nevertheless, their DNA WES data is available to the community for joint efforts to detect variants related to Covid-19 resistance.
Furthermore, the HLA alleles that each of these centenarians carry were described, being likely candidates to influence infection outcomes [68–70] and longevity [71]. Many studies have reported potential HLA alleles implicated in response to SARS-CoV-2 infection [72], whether they were identified in a specific geographic region or globally. This influence is suggestively related to differential antigen presentation and interaction with the T cell receptor [73].
Conclusion
In the present study, we investigated three Covid-19 recovered supercentenarians (older than 110-years-old) who displayed robust IgG levels and neutralization titers against SARS-CoV-2. An enrichment of plasma proteins and metabolites related to innate immune response and host defense was observed. Despite their advanced age, none of them had neutralized auto-Abs against the type I IFNs. Also, they do not carry variants associated with inborn errors of type I IFN immunity (IEI) genes. They belong to a selected group of individuals with a long lifetime of pathogens’ expositions, immunity training, and genetic factors that lead them to develop mild symptoms not only against Covid-19 but also for several other diseases. Understanding the underlying mechanisms may be important to protect us from future pandemics.
Methods
Participants’ recruitment and Sample Collection
Three Brazilian unvaccinated supercentenarians who recovered from Covid-19 were contacted by our Human Genome and Stem Cell Research Center (HUG-CELL) research group following their report in national media: a 114-years-old woman (ID 01) and two men aged 111 (ID 02) and 110 (ID 03) years-old, respectively, at blood collection time. To our knowledge, these were the longest-lived people who recovered from Covid-19 in South America before the vaccination started and new SARS-CoV-2 variants emerged. RT-PCR tests confirmed the previous diagnosis of Covid-19, and all relevant clinical data related to the disease episode and comorbidities were collected from clinical reports and interviews. Baseline characteristics of the three supercentenarians are shown in Table 3.
Table 3.
Demographic and clinical data of the participants
| GENERAL INFORMATION | |||
|---|---|---|---|
| ID | 01 | 02 | 03 |
| Sex | F | M | M |
| Age | 114 | 111 | 110 |
| Year of birth | 1906 | 1910 | 1911 |
| Life status | Death on Feb 2021, as a consequence of urinary tract infection | Death in Nov 2021 from natural causes | Still alive |
| Comorbidities* | Dementia | Diabetes mellitus | Hypertension |
| Covid-19-RELATED EVENTS | |||
| SARS-CoV-2 exposure/ beginning of symptoms | August 2020 | June 2020 | May 2020 |
| Disease severity** | Moderate | Asymptomatic | Moderate |
| Hospital admission and discharge |
Hospital admission on Aug 31, 2020 ICU admission necessary |
Not applicable |
Hospital admission in June 2020 ICU admission necessary Hospital discharge after 18 days |
| Positive SARS-CoV-2 serology |
Sep 2020 MAGLUMI 2019 nCoV IgM and IgM reagents |
Jan 2021 ELISA IgM and IgG anti-Spike and anti-NP (Table 1) |
Jun 2021 ELISA IgM and IgG anti-Spike and anti-NP (Table 1) |
We collected peripheral blood samples of the volunteers from 30 to 120 days after the reported viral infections. For DNA extraction, samples were taken in vacutainer tubes with ethylenediaminetetraacetic acid - EDTA (BD Biosciences, USA, Catalog #. 360,057). Plasma and Serum were obtained by centrifugation for 10 min at 2000 x g at room temperature within 30 min after venipuncture. Then, the supernatant was transferred in aliquots of 1.5 mL into cryovials (Corning®, USA, Catalog #. 430,487). Samples were transferred to a -80 °C freezer until the moment of use.
Humoral immune response assessment
The humoral immune response was analyzed by ELISA for IgA, IgM, and IgG-binding antibodies against the receptor-binding domain of Spike protein and NP protein of SARS-CoV-2. Besides, RBDs from human seasonal coronaviruses (HCoV) HKU-1, OC43, NL63, and 229E were also tested. RBDs from HCoVs were expressed in HEK293T cells, which plasmids are described in [76]. ELISA was performed using 96-well high-binding half-area polystyrene plates coated overnight at 4oC with 4 µg/mL of SARS-CoV-2 RBD, 0.8 µg/mL of the RBD of HCoVs, and 2 µg/mL NP (Kindly provided by Dr. Ricardo Gazzinelli, UFMG). Volunteers’ plasma samples were incubated at 56 °C for 30 min, diluted at 1:100, and run-in triplicates in ELISA. Results were given as the ratio of participant sample/average of a set of 20 control pre-pandemic samples. An antibody ratio of ≥ 1.2 was considered positive.
The detection of auto-Abs neutralizing type I IFNS (IFN-α2, IFN-β, and/or -ω) in plasma samples of the supercentenarians was assessed by LIPS assay, as described in [23]. Briefly, HEK293 cells transfected with type I IFNS fused to firefly luciferase were lysate (with doses from 0.1 pg/mL to 10 ng/mL of IFN-α2, -β, and/or -ω) and incubated with 10% diluted plasma of the volunteers. The resulting complexes were conjugated with agarose beads to capture the immune complexes. Then, the luciferase substrate furimazine was added to the reaction and the luminescence intensity (LU) was proportional to the presence of anti-Abs.
Neutralization titers were measured in a pseudovirus assay adapted from [77], only changing transfection to use lipofectamine 2000 (Thermo).
Proteomics and metabolomics analyses
Plasma proteomics and metabolomics analyses from the supercentenarians were performed using tandem mass spectrometry. Detailed protocols concerning both analyses and data processing are available in Supplementary Methods. Three individuals older than 95 years-old who were not infected by SARS-CoV-2 and displayed a negative COVID-19 serology were included as the control group.
Genomic assays
Whole-exome sequencing (WES) was performed in peripheral blood DNA with the Illumina NovaSeq platform at HUG-CELL facilities. Sequencing data were analyzed following bwa-mem and GATK Best Practices workflow, quality control, and annotation were performed as previously described [36]. HLA genes were realigned and called using hla-mapper [78], and the pipeline was described elsewhere [79].
Genetic ancestry inference
The inference of genetic ancestry was performed in ADMIXTURE v1.36 [80], in supervised analysis (k = 4), after filtering the markers for linkage disequilibrium (r2 = 0.1) using a 50Kb sliding window with 10 kb steps, totaling 53,987 SNPs. Samples from both the 1000 Genomes Project [81] and the HGDP-CEPH [82] with over 95% inferred ancestry in a given group were used as parent populations, totaling 602 Africans, 624 Europeans, 630 East Asians, and 118 Native Americans.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Acknowledgements
We would like to dedicate this study to the memory of O. J. (1906–2021) and O. S. (1909–2021). The authors are extremely grateful for the participation and collaboration of the three supercentenarians and their families, the nurses for sample collection, and the HUG-CELL technical team. Special thanks to Brazilian senator Mara Gabrilli for financial support.
Abbreviations
- Auto-Abs
Autoantibodies
- CNV
Copy number variation
- EDTA
Ethylenediamine tetraacetic acid
- ELISA
Enzyme-linked immunosorbent assay
- HCoV
Human coronaviruses
- HLA
Human leukocyte antigen
- HUG-CELL
Human Genome and Stem Cell Research Center
- IEI
Inborn errors of type I IFN immunity
- IFN
Interferon
- IRF7
Interferon regulatory factor 7
- LIPS
Luciferase-based immunoprecipitation
- LU
Luminescence intensity
- MHC
Major Histocompatibility Complex
- NAbs
Neutralizing antibodies
- NP
Nucleocapsid
- RBD
Receptor-binding domain
- TLR3
Toll-like receptor 3
- WES
Whole-exome sequencing
Author contribution
M.V.C., M.V.R.S., E.C.C., and K.S.S. contributed equally to this work as co-first authors. All authors contributed significantly to this work. M.V.C. and M.V.R.S. Investigation, Formal Analysis, Visualization, Data Curation, Writing – Original Draft and Review & Editing. M.O.S., K.N., E.C.C., J.Y.M., F.L.A., V.L.O., A.I.S.M., E.J. M.Q.V., P.S.A., J.S.G., and N.P.A. Methodology, Formal Analysis, Data Curation, and Investigation. M.S.N. Conceptualization, Methodology, Formal Analysis, Data Curation, and Investigation. K.S.S., E.C.N., B.S.S., P.B., J.L.C., J.K., F.C.S.N., and G.B.D. Conceptualization, Supervision and Writing – Review & Editing. M.R.P.B. Resources. M.Z. Writing – Review & Editing, Funding Acquisition, and Project Administration. The authors read and approved the final manuscript.
Funding
This work was supported by the Sao Paulo Research Foundation (FAPESP) [grant numbers 2013/08028-1, 2014/50890-5, 2014/50931-3, and 2020/09702-1], the National Council for Scientific and Technological Development (CNPq) [grant numbers 465434/2014-2 and 465355/2014-5], the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior-Brasil (CAPES) - Finance Code 001 and JBS S.A [grant number 69004]. FAPESP/Brazil (Grant numbers 2013/17084-0 and 2017/19223-0) supported the development of the HLA pipeline. United States National Institutes of Health (NIH) [grant number R01 GM075091] supported the genetic ancestry inferences. The funders were not involved in the study design, collection, analysis, interpretation of data, the writing of this review, or the decision to submit it for publication.
Availability of data and materials
The genomic datasets presented in this study are currently being deposited at the “European Genome-phenome Archive (EGA)”, under accession number EGAS00001006376, a permanent public repository, in compliance with “Immunity and Ageing” recommendations and its open data policies. We will provide the public datasets accession numbers prior to publishing the final version.
Declarations
Ethics approval and consent to participate
The study was approved by the Committee for Ethics in Research of the Institute of Biosciences at the University of São Paulo (CAAE 34786620.2.0000.5464) by the Declaration of Helsinki principles, ICH06 Good Clinical Practices and Brazilian Health Regulatory Agency ANVISA resolution number 466 from 2012 that regulates research with humans and the families or caregivers of all participants provided written consent.
Consent for publication
The families or caregivers of all participants and all the authors consented to the publication of this manuscript.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Mateus V. de Castro, Monize V. R. Silva, Erick C. Castelli and Keity S. Santos contributed equally to this work.
References
- 1.WHO Coronavirus (COVID–19) Dashboard. [cited 2022 Jul 11]. Available from: https://covid19.who.int.
- 2.Chai S, Li Y, Li X, Tan J, Abdelrahim MEA, Xu X. Effect of age of COVID–19 inpatient on the severity of the disease: A meta-analysis. Int J Clin Pract. 2021;75(10):e14640. doi: 10.1111/ijcp.14640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hägg S, Jylhävä J, Wang Y, Xu H, Metzner C, Annetorp M. Age, Frailty, and Comorbidity as Prognostic Factors for Short-Term Outcomes in Patients With Coronavirus Disease 2019 in Geriatric Care. J Am Med Dir Assoc. 2020;21(11):1555–9.e2. doi: 10.1016/j.jamda.2020.08.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Thakur B, Dubey P, Benitez J, Torres JP, Reddy S, Shokar N. A systematic review and meta-analysis of geographic differences in comorbidities and associated severity and mortality among individuals with COVID–19. Sci Rep. 2021;20(1):8562. doi: 10.1038/s41598-021-88130-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Conheça a faixa etáriadosmortospor covid– 19 no Brasileemma is 4 países.[cited 2022 Jul 26]. Available from: https://www.poder360.com.br/coronavirus/conheca-a-faixa-etaria-dos-mortos-por-covid?19-no-brasil-e-em-mais?4-paises?4/.
- 6.Centers for Disease Control. Older Adults Risks and Vaccine Information | cdc. 2021[cited 2021 Oct 23]. Available from: https://www.cdc.gov/aging/covid19/covid19-older-adults.html.
- 7.CDC. Cases, Data, and Surveillance. Centers for Disease Control and Prevention. 2020[cited 2021 Nov 18]. Available from: https://www.cdc.gov/coronavirus/2019-ncov/covid-data/investigations-discovery/hospitalization-death-by-age.html.
- 8.López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. The hallmarks of aging. Cell. 2013;153(6):1194–217. [DOI] [PMC free article] [PubMed]
- 9.Fuentes E, Fuentes M, Alarcón M, Palomo I. Immune System Dysfunction in the Elderly. An Acad Bras Ciênc. 2017;89:285–99. doi: 10.1590/0001-3765201720160487. [DOI] [PubMed] [Google Scholar]
- 10.Amore S, Puppo E, Melara J, Terracciano E, Gentili S, Liotta G. Impact of COVID–19 on older adults and role of long-term care facilities during early stages of epidemic in Italy. Sci Rep. 2021;15(1):12530. doi: 10.1038/s41598-021-91992-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Aiello A, Farzaneh F, Candore G, Caruso C, Davinelli S, Gambino CM,etal.Immunosenescence and Its Hallmarks: How to Oppose Aging Strategically? A Review of Potential Options for Therapeutic Intervention. Front Immunol. 2019 [cited 2022 Jul 26];10. Available from: https://www.frontiersin.org/articles/10.3389/fimmu.2019.0224710.3389/fimmu.2019.02247. [DOI] [PMC free article] [PubMed]
- 12.Lian J, Yue Y, Yu W, Zhang Y. Immunosenescence: a key player in cancer development. J Hematol OncolJ Hematol Oncol. 2020;10(1):151. doi: 10.1186/s13045-020-00986-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ongrádi J, Kövesdi V. Factors that may impact on immunosenescence: an appraisal. Immun Ageing A. 2010;7:7. [DOI] [PMC free article] [PubMed]
- 14.Haynes L. Aging of the Immune System: Research Challenges to Enhance the Health Span of Older Adults. Front Aging. 2020 [cited 2022 Jul 26];1.Available from: https://www.frontiersin.org/articles/10.3389/fragi.2020.602108. [DOI] [PMC free article] [PubMed]
- 15.Cunha LL, Perazzio SF, Azzi J, Cravedi P, Riella LV. Remodeling of the Immune Response With Aging: Immunosenescence and Its Potential Impact on COVID–19 Immune Response. Front Immunol. 2020;7:1748. doi: 10.3389/fimmu.2020.01748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Dugué PA, Hodge AM, Ulvik A, Ueland PM, Midttun Ø, Rinaldi S, etal. Association of Markers of Inflammation,the Kynurenine Pathway and B Vitamins with Age and Mortality, and a Signature of Inflammaging. J Gerontol A Biol Sci Med Sci. 2022;77(4):826–36. [DOI] [PubMed]
- 17.Franceschi C, Garagnani P, Parini P, Giuliani C, Santoro A. Inflammaging: a new immune–metabolic viewpoint for age-related diseases. Nat Rev Endocrinol. 2018;14(10):576–90. doi: 10.1038/s41574-018-0059-4. [DOI] [PubMed] [Google Scholar]
- 18.Fajgenbaum DC, June CH, Cytokine Storm N Engl J Med. 2020;3(23):2255–73. doi: 10.1056/NEJMra2026131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Cron RQ.COVID–19 cytokine storm: targeting the appropriate cytokine. Lancet Rheumatol. 2021;3(4):e236–7. [DOI] [PMC free article] [PubMed]
- 20.Peron JPS, Nakaya H. Susceptibility of the Elderly to SARS-CoV–2 Infection: ACE–2 Overexpression, Shedding, and Antibody-dependent Enhancement(ADE). Clinics 2020 [cited 2022 Jul 26];75. Available from: http://www.scielo.br/j/clin/a/QDXYJBQk6YyLpfGNgBKtxHQ/?lang=en. [DOI] [PMC free article] [PubMed]
- 21.Yuki K, Fujiogi M, Koutsogiannaki S. COVID–19 pathophysiology: A review. Clin Immunol Orlando Fla. 2020;215:108427. doi: 10.1016/j.clim.2020.108427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Parasher A. COVID–19: Current understanding of its Pathophysiology, Clinical presentation and Treatment. Postgrad Med J. 2021;97(1147):312–20. doi: 10.1136/postgradmedj-2020-138577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Bastard P, Rosen LB, Zhang Q, Michailidis E, Hoffmann HH, Zhang Y, etal. Auto antibodies against type IIFNs in patients with life-threatening COVID–19. Science. 2020;370(6515):eabd4585. [DOI] [PMC free article] [PubMed]
- 24.Bastard P, Gervais A, Le Voyer T, Rosain J, Philippot Q, Manry J, etal. Auto antibodies neutralizing type IIFNs are present in~ 4%of uninfected individuals over 70 years old and account for~ 20% of COVID– 19 deaths. SciImmunol. 2021;6(62): eabl4340. [DOI] [PMC free article] [PubMed]
- 25.Kordowitzki P.Centenarians and COVID–19: Is There a Link between Longevity and Better Immune Defense? Gerontology. 2021;1–2. [DOI] [PMC free article] [PubMed]
- 26.Guerini FR, Cesari M, Arosio B. Hypothetical COVID–19 protection mechanism: hints from centenarians. Immun Ageing. 2021;30(1):15. doi: 10.1186/s12979-021-00226-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Foley MK, Searle SD, Toloue A, Booth R, Falkenham A, Falzarano D. Centenarians and extremely old people living with frailty can elicit durable SARS-CoV–2 spike specific IgG antibodies with virus neutralization functions following virus infection as determined by serological study. EClinicalMedicine. 2021;37:100975. doi: 10.1016/j.eclinm.2021.100975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kong Y, Cai C, Ling L, Zeng L, Wu M, Wu Y. Successful treatment of a centenarian with coronavirus disease 2019 (COVID–19) using convalescent plasma. Transfus Apher Sci. 2020;59(5):102820. doi: 10.1016/j.transci.2020.102820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Toppi E, De Molfetta V, Zarletti G, Tiberi M, Bossù P, Scapigliati G. The Anti-SARS-CoV–2 Antibody Response in a Centenarian Woman: A Case of Long-Term Memory? Viruses. 2021;13(9):1704. [DOI] [PMC free article] [PubMed]
- 30.Siopis G. Supercentenarians that Survived COVID–19. Aging Dis. 2021;12(7):1539–40. [DOI] [PMC free article] [PubMed]
- 31.Huang Y, Paxton WA, Wolinsky SM, Neumann AU, Zhang L, He T. The role of a mutant CCR5 allele in HIV–1 transmission and disease progression. Nat Med. 1996;2(11):1240–3. doi: 10.1038/nm1196-1240. [DOI] [PubMed] [Google Scholar]
- 32.Kiepiela P, Leslie AJ, Honeyborne I, Ramduth D, Thobakgale C, Chetty S,etal.Dominant influence of HLA-B in mediating the potential co-evolution of HIV and HLA. Nature. 2004;432(7018):769–75. [DOI] [PubMed]
- 33.Fellay J, Shianna KV, Ge D, Colombo S, Ledergerber B, Weale M. A whole-genome association study of major determinants for host control of HIV–1. Science. 2007;17(5840):944–7. doi: 10.1126/science.1143767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Andreakos E, Abel L, Vinh DC, Kaja E, Drolet BA, Zhang Q. A global effort to dissect the human genetic basis of resistance to SARS-CoV–2 infection. Nat Immunol. 2022;23(2):159–64. doi: 10.1038/s41590-021-01030-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Investigational COVID–19 Convalescent Plasma; GuidanceforIndustry; Availability. Federal Register. 2020[cited2022Jul26]. Available from: https://www.federalregister.gov/documents/2020/09/21/2020–20800/investigational-covid–19-convalescent-plasma-guidance-for-industry-availability.
- 36.Naslavsky MS, Scliar MO, Yamamoto GL, Wang JYT, Zverinova S, Karp T,etal.Whole-genome sequencing of 1,171 elderly admixed individuals from São Paulo, Brazil. Nat Commun. 2022;13(1):1004. [DOI] [PMC free article] [PubMed]
- 37.Naslavsky MS, Yamamoto GL, de Almeida TF, Ezquina SAM, Sunaga DY, Pho N. Exomic variants of an elderly cohort of Brazilians in the ABraOM database. Hum Mutat. 2017;38(7):751–63. doi: 10.1002/humu.23220. [DOI] [PubMed] [Google Scholar]
- 38.Souza AM de, Resende SS, Sousa TN de, Brito CFA de. Asystematic scoping review of the genetic ancestry of the Brazilian population. Genet Mol Biol. 2019;42:495–508. [DOI] [PMC free article] [PubMed]
- 39.Zhang Q, Bastard P, Liu Z, Le Pen J, Moncada-Velez M, Chen J,etal. In bornerrors of type IIFN immunity in patients with life-threatening COVID–19. Science. 2020;370(6515):eabd4570. [DOI] [PMC free article] [PubMed]
- 40.Zhang Q, Bastard P, Cobat A, Casanova JL. Human genetic and immunological determinants of critical COVID–19 pneumonia. Nature. 2022;603(7902):587–98. doi: 10.1038/s41586-022-04447-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Marchi S, Viviani S, Remarque EJ, Ruello A, Bombardieri E, Bollati V. Characterization of antibody response in asymptomatic and symptomatic SARS-CoV–2 infection. PLoS ONE. 2021;16(7):e0253977. doi: 10.1371/journal.pone.0253977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Maciola AK, La Raja M, Pacenti M, Salata C, De Silvestro G, Rosato A. Neutralizing Antibody Responses to SARS-CoV–2 in Recovered COVID–19 Patients Are Variable and Correlate With Disease Severity and Receptor-Binding Domain Recognition. Front Immunol. 2022;13:830710. doi: 10.3389/fimmu.2022.830710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Dugas M, Grote-Westrick T, Merle U, Fontenay M, Kremer AE, Hanses F. Lack of antibodies against seasonal coronavirus OC43 nucleocapsid protein identifies patients at risk of critical COVID–19. J Clin Virol Off Publ Pan Am Soc Clin Virol. 2021;139:104847. doi: 10.1016/j.jcv.2021.104847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Lin CY, Wolf J, Brice DC, Sun Y, Locke M, Cherry S, etal. Pre-existing humoral immunity to human common cold coronaviruses negatively impacts the protective SARS-CoV–2 antibody response. Cell Host Microbe. 2022;30(1):83–96.e4. [DOI] [PMC free article] [PubMed]
- 45.Abela IA, Pasin C, Schwarzmüller M, Epp S, Sickmann ME, Schanz MM, etal. Multifactorial seroprofiling dissects the contribution of pre-existing human coronaviruses responses to SARS-CoV–2 immunity. Nat Commun. 2021;12(1):6703. [DOI] [PMC free article] [PubMed]
- 46.Zhang A, Stacey HD, Mullarkey CE, Miller MS. Original Antigenic Sin: How First Exposure Shapes Life long Anti-Influenza Virus Immune Responses. J Immunol Baltim Md 1950. 2019;202(2):335–40. [DOI] [PubMed]
- 47.Debisarun PA, Gössling KL, Bulut O, Kilic G, Zoodsma M, Liu Z. Induction of trained immunity by influenza vaccination - impact on COVID–19. PLOS Pathog. 2021;25(10):e1009928. doi: 10.1371/journal.ppat.1009928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Poulain M, Chambre D, Pes GM. Centenarians exposed to the Spanish flu in their early life better survived to COVID–19. Aging. 2021;13(18):21855–65. [DOI] [PMC free article] [PubMed]
- 49.Yu X, Tsibane T, McGraw PA, House FS, Keefer CJ, Hicar MD, etal. Neutralizing antibodies derived from the B cells of 1918 influenza pandemic survivors. Nature. 2008;455(7212):532–6. [DOI] [PMC free article] [PubMed]
- 50.Junior S, Santos Ados. Corpos hígidos: o limpo e o sujo na Paraíba(1912–1924). Universidade Federal da Paraíba; 2011[cited 2022 Jul 26]. Available from: https://repositorio.ufpb.br.
- 51.Gripe Espanhola em MOC fechou. lojas efábricas e transformou escola em hospital, revela pesquisador da Unimontes. Universidade Estadual de Montes Claros-Unimontes. 2020 [cited 2022 Jul 26]. Available from: https://unimontes.br/gripe-espanhola-em-moc-fechou-lojas-e-fabricas-e-transformou-escola-em-hospital-revela-pesquisador-da-unimontes/.
- 52.Rijkers GT, van Overveld FJ.The “original antigenic sin” and its relevance for SARS-CoV–2 (COVID–19) vaccination. Clin Immunol Commun. 2021;1:13–6. [DOI] [PMC free article] [PubMed]
- 53.Hashimoto K, Kouno T, Ikawa T, Hayatsu N, Miyajima Y, Yabukami H,etal. Single-cell transcriptomics reveals expansion of cytotoxic CD4 T cells in supercentenarians. Proc Natl Acad Sci. 2019;116(48):24242–51. [DOI] [PMC free article] [PubMed]
- 54.Santos AF, Póvoa P, Paixão P, Mendonça A, Taborda-Barata L. Changes in Glycolytic Pathway in SARS-COV 2 Infection and Their Importance in Understanding the Severity of COVID–19. Front Chem. 2021;10:685196. doi: 10.3389/fchem.2021.685196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Codo AC, Davanzo GG, MonteiroLdeB,deSouzaGF, Muraro SP, Virgilio-da-Silva JV,etal. Elevated Glucose Levels Favor SARS-CoV–2 Infection and Monocyte Response through a HIF–1α/Glycolysis-Dependent Axis. Cell Metab. 2020;32(3):437–446.e5. [DOI] [PMC free article] [PubMed]
- 56.Alomar FA, Alshakhs MN, Abohelaika S, Almarzouk HM, Almualim M, Al-Ali AK. Elevated plasma level of the glycolysis byproduct methylglyoxal on admission is an independent biomarker of mortality in ICU COVID–19 patients. Sci Rep. 2022;9(1):9510. doi: 10.1038/s41598-022-12751-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Medini H, Zirman A, Mishmar D. Immune system cells from COVID–19 patients display compromised mitochondrial-nuclear expression co-regulation and rewiring toward glycolysis. iScience. 2021;17(12):103471. doi: 10.1016/j.isci.2021.103471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Krishnan S, Nordqvist H, Ambikan AT, Gupta S, Sperk M, Svensson-Akusjärvi S. Metabolic Perturbation Associated With COVID–19 Disease Severity and SARS-CoV–2 Replication. Mol Cell Proteomics MCP. 2021;20:100159. doi: 10.1016/j.mcpro.2021.100159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Sanchez EL, Lagunoff M. Viral activation of cellular metabolism. Virology. 2015;1:609–18. doi: 10.1016/j.virol.2015.02.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Kishimoto N, Yamamoto K, Abe T, Yasuoka N, Takamune N, Misumi S. Glucose-dependent aerobic glycolysis contributes to recruiting viral components into HIV–1 particles to maintain infectivity. Biochem Biophys Res Commun. 2021;549:187–93. [DOI] [PubMed]
- 61.Ren L, Zhang W, Zhang J, Zhang J, Zhang H, Zhu Y. Influenza A Virus (H1N1) Infection Induces Glycolysis to Facilitate Viral Replication. Virol Sin. 2021;36(6):1532–42. doi: 10.1007/s12250-021-00433-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Yu Q, Wang Y, Dong L, He Y, Liu R, Yang Q,etal. Regulations of Glycolytic Activities on Macrophages Functions in Tumor and Infectious Inflammation. Front Cell Infect Microbiol. 2020 [cited 2022 Aug 18];10. Available from: https://www.frontiersin.org/articles/10.3389/fcimb.2020.00287 [DOI] [PMC free article] [PubMed]
- 63.Calder PC. Eicosanoids. Essays Biochem. 2020;64(3):423–41. [DOI] [PubMed]
- 64.Kothapalli KSD, Park HG, Brenna JT. Polyunsaturated fatty acid biosynthesis pathway and genetics. implications for interindividual variability in prothrombotic, inflammatory conditions such as COVID–19☆,☆☆,★,★★. Prostaglandins Leukot Essent Fatty Acids. 2020;162:102183. doi: 10.1016/j.plefa.2020.102183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Adili R, Hawley M, Holinstat M. Regulation of platelet function and thrombosis by omega–3 and omega–6 polyunsaturated fatty acids. Prostaglandins Other Lipid Mediat. 2018;139:10–8. doi: 10.1016/j.prostaglandins.2018.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Asher A, Tintle NL, Myers M, Lockshon L, Bacareza H, Harris WS. Blood omega–3 fatty acids and death from COVID–19: A pilot study. Prostaglandins Leukot Essent Fatty Acids. 2021;166:102250. doi: 10.1016/j.plefa.2021.102250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Baral PK, Amin MT, Rashid MMO, Hossain MS. Assessment of Polyunsaturated Fatty Acids on COVID–19-Associated Risk Reduction. Rev Bras Farmacogn Orgao Of Soc Bras Farmacogn. 2022;32(1):50–64. doi: 10.1007/s43450-021-00213-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.White CF, Pellis L, Keeling MJ, Penman BS. Detecting HLA-infectious disease associations for multi-strain pathogens. Infect Genet Evol. 2020;1:104344. doi: 10.1016/j.meegid.2020.104344. [DOI] [PubMed] [Google Scholar]
- 69.Sanchez-Mazas A. A review of HLA allele and SNP associations with highly prevalent infectious diseases in human populations. Swiss Med Wkly. 2020;6:w20214. doi: 10.4414/smw.2020.20214. [DOI] [PubMed] [Google Scholar]
- 70.Blackwell JM, Jamieson SE, Burgner D. HLA and infectious diseases. Clin Microbiol Rev. 2009;22(2):370–85. doi: 10.1128/CMR.00048-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Naumova E, Mihaylova A, Ivanova M, Mihailova S. Impact of KIR/HLA ligand combinations on immune responses in malignant melanoma. Cancer Immunol Immunother CII. 2007;56(1):95–100. doi: 10.1007/s00262-006-0151-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Aguiar VRC, Augusto DG, Castelli EC, Hollenbach JA, Meyer D, Nunes K,etal. An immunogenetic view of COVID–19. Genet Mol Biol. 2021[cited2022Jul26];44. Available from: http://www.scielo.br/j/gmb/a/KJzT4HNJmhnLHjTTMBY8QqK/?lang=en. [DOI] [PMC free article] [PubMed]
- 73.Klein J, Sato A. The HLA system. First of two parts. N Engl J Med. 2000;7(10):702–9. doi: 10.1056/NEJM200009073431006. [DOI] [PubMed] [Google Scholar]
- 74.CDC. Coronavirus Disease 2019(COVID–19). Centers for Disease Control and Prevention. 2020 [cited2021Dec5]. Available from: https://www.cdc.gov/coronavirus/2019-ncov/variants/delta-variant.html.
- 75.Living guidance for clinical management of COVID–19. [cited2022Jul26]. Available from: https://www.who.int/publications-detail-redirect/WHO?2019-nCoV-clinical?2021?2.
- 76.Premkumar L, Segovia-Chumbez B, Jadi R, Martinez DR, Raut R, Markmann AJ, et al. The receptor-binding domain of the viral spike protein is an immunodominant and highly specific target of antibodies in SARS-CoV–2 patients. Sci Immunol. 2020;5(48):eabc8413. [DOI] [PMC free article] [PubMed]
- 77.Schmidt F, Weisblum Y, Muecksch F, Hoffmann HH, Michailidis E, Lorenzi JCC. Measuring SARS-CoV–2 neutralizing antibody activity using pseudotyped and chimeric viruses. J Exp Med. 2020;2(11):e20201181. doi: 10.1084/jem.20201181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Castelli EC, Paz MA, Souza AS, Ramalho J, Mendes-Junior CT. Hla-mapper: An application to optimize the mapping of HLA sequences produced by massively parallel sequencing procedures. Hum Immunol. 2018;79(9):678–84. doi: 10.1016/j.humimm.2018.06.010. [DOI] [PubMed] [Google Scholar]
- 79.Castelli EC, de Castro MV, Naslavsky MS, Scliar MO, Silva NSB, Andrade HS, et al. MHC Variants Associated With Symptomatic Versus Asymptomatic SARS-CoV–2 Infection in Highly Exposed Individuals. Front Immunol. 2021 [cited 2022 May 23];12. Available from: https://www.frontiersin.org/article/10.3389/fimmu.2021.74288110.3389/fimmu.2021.742881. [DOI] [PMC free article] [PubMed]
- 80.Alexander DH, Novembre J, Lange K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009;19(9):1655–64. doi: 10.1101/gr.094052.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Auton A, Abecasis GR, Altshuler DM, Durbin RM, Abecasis GR, Bentley DR. A global reference for human genetic variation. Nature. 2015;526(7571):68–74. doi: 10.1038/nature15393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Bergström A, McCarthy SA, Hui R, Almarri MA, Ayub Q, Danecek P. Insights into human genetic variation and population history from 929 diverse genomes. Science. 2020;20(6484):eaay5012. doi: 10.1126/science.aay5012. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The genomic datasets presented in this study are currently being deposited at the “European Genome-phenome Archive (EGA)”, under accession number EGAS00001006376, a permanent public repository, in compliance with “Immunity and Ageing” recommendations and its open data policies. We will provide the public datasets accession numbers prior to publishing the final version.