

Abstract
Sustainable agriculture in the future will depend on crops that are tolerant to biotic and abiotic stresses, require minimal input of water and nutrients and can be cultivated with a minimal carbon footprint. Wild plants that fulfill these requirements abound in nature but are typically low yielding. Thus, replacing current high-yielding crops with less productive but resilient species will require the intractable trade-off of increasing land area under cultivation to produce the same yield. Cultivating more land reduces natural resources, reduces biodiversity and increases our carbon footprint. Sustainable intensification can be achieved by increasing the yield of underutilized or wild plant species that are already resilient, but achieving this goal by conventional breeding programs may be a long-term prospect. De novo domestication of orphan or crop wild relatives using mutagenesis is an alternative and fast approach to achieve resilient crops with high yields. With new precise molecular techniques, it should be possible to reach economically sustainable yields in a much shorter period of time than ever before in the history of agriculture.
Keywords: Accelerated domestication, Breeding, CRISPR/Cas, Mutagenesis, Orphan crops, Sustainable intensification
Introduction
Several strategies have been devised to meet the increasing demand for food posed by a growing population estimated to surpass 9 billion people by 2050 (Breseghello and Guedes Coelho 2013, Crossa et al. 2021, Gao 2021). One argument is that agricultural production must increase globally to supply the food required (The Government Office for Science 2011; Tilman et al. 2011). Supply can expand either via increasing the yield over the same area (intensification) or via bringing more land into agricultural production (extensification) (Gabriel et al. 2013). Intensification is challenging in highly productive areas, where yield increases have already plateaued (Ray et al. 2012), which leaves us with extensification, a solution that threatens nature and biodiversity.
Still, increasing supply may not be a solution at all. Increasing food supply typically generates increasing demand, creating a vicious circle (Benton and Bailey 2019). For example, providing more food for more people may result in a positive feedback loop, potentially delaying the problem rather than solving it (Rull 2010). Consequently, for future food production to be sustainable, an alternative theory is that demand must decrease, for example, through dietary changes like refraining from certain foods, particularly beef, in parallel with efforts to reduce food waste (Benton et al. 2021). However, even if projected demand is reduced, just maintaining supply is challenging. Ongoing and predicted climate change, the negative impact of agriculture on the environment and shrinking natural resources are likely to exacerbate existing yield gaps (Fisher 2015) and may even decrease yields (Godfray et al. 2010, Foley et al. 2011, FAO et al. 2018).
Sustainable intensification aims to increase agricultural productivity without negatively impacting natural resources or the integrity of associated systems (Cassman and Grassini 2020). This approach depends on high-yielding crops that require minimal input, tolerate abiotic and biotic stresses, have a low climate footprint and are nutritious. However, our current crops have been mainly selected for yield, while traits related to hardiness and stress tolerance, alongside others, either have not been selected for or have been lost (Østerberg et al. 2017). To reintroduce tolerance to unfavorable environments, one suggestion is to restore modern elite varieties with the lost properties that their landraces possessed (Palmgren et al. 2015). Backcrossing with old cultivars or directly introducing specific genes can in some cases furnish a crop with a lost stress tolerance trait, but these approaches are challenging since most traits are multigenic (Wingen et al. 2017).
A possible alternative may be to direct breeding efforts toward plant species that are already adapted to harsh environments or contribute to naturally stable ecosystems (Østerberg et al. 2017). Potential candidates could be hardy orphan crops that have yet to be fully domesticated, for example, perennial grain crops (DeHaan et al. 2020), orphan legumes (e.g. Lathyrus sativus) (Lambein et al. 2019), or even wild plants, including nitrogen-fixing perennials (Table 1). Natural genetic diversity of plants is enormous, with plants found in almost every ecological niche. Thus, nature provides us with a rich source of novel crops, many of which are promising for use in sustainable agriculture. However, yields of wild plants or underutilized orphan crops are typically much lower compared to our current high-yielding crops. Therefore, to facilitate this switch, yields of these new crops must be improved to prevent dramatic reductions in food supply. Such domestication efforts should focus on increasing yield, while maintaining innate stress tolerance. While such a yield increase will not increase the current food supply, the additional resilience could secure food production and limit soil degradation under increasingly erratic climate conditions.
Table 1.
Perennial orphan crops and candidates for domestication from a sustainability point of view.
| Common name | Scientific name | Beneficial traits | Life cycle | Reference |
|---|---|---|---|---|
| Cereals | ||||
| European beachgrass | Ammophila arenaria | Tolerance to abiotic stresses | Perennial | Zhang et al. 2020 |
| Wild barley | Hordeum bulbosum | Tolerance to biotic and abiotic stresses | Perennial | Westerbergh et al. 2018 |
| Intermediate wheatgrass | Thinopyrum intermedium | Tolerance to abiotic stresses | Perennial | DeHaan et al. 2020 |
| Legumes | ||||
| Alfalfa | Medicago sativa | Nitrogen fixing, nutritious, intercropping | Perennial | The Land Institute, USA |
| Kura clover | Trifolium ambiguum | Nitrogen fixing, nutritious, intercropping | Perennial | The Land Institute, USA |
| Sainfoin | Onobrychis viciifolia | Nitrogen fixing, nutritious, intercropping | Perennial | The Land Institute, USA |
| Lupin | Lupinus spp. | Nitrogen fixing, nutritious, intercropping | Perennial | The Land Institute, USA |
| Oil seeds | ||||
| Rosinweed | Silphium integrifolium | Oil, long root | Perennial | The Land Institute, USA |
| Root crops | ||||
| Wild potato | Solanum spp. | Tolerance to biotic and abiotic stresses | Perennial | Cadima et al. 2014 |
| Legume trees | ||||
| Reonja | Acacia leucophloea | Nitrogen fixing, livestock fodder | Perennial | Toensmeier 2016 |
| Gum arabic tree | Acacia nilotica | Nitrogen fixing, livestock fodder | Perennial | Toensmeier 2016 |
| Coojong | Acacia saligna | Nitrogen fixing, livestock fodder | Perennial | Toensmeier 2016 |
| Gum acacia | Acacia senegal | Nitrogen fixing, livestock fodder | Perennial | Toensmeier 2016 |
| Red acacia | Acacia seyal | Nitrogen fixing, livestock fodder | Perennial | Toensmeier 2016 |
| Umbrella thorn acacia | Acacia tortilis | Nitrogen fixing, livestock fodder | Perennial | Toensmeier 2016 |
| Red lucky seed | Adenanthera pavonina | Nitrogen fixing, livestock fodder | Perennial | Toensmeier 2016 |
| Monkeypod | Albizia saman | Nitrogen fixing, livestock fodder | Perennial | Toensmeier 2016 |
| Camel’s foot | Bauhinia thonningii | Nitrogen fixing, edible | Perennial | Toensmeier 2016 |
| Pink shower tree | Cassia grandis | Livestock fodder, some are nitrogen fixing | Perennial | Toensmeier 2016 |
| Carob | Ceratonia siliqua | Livestock fodder, edible | Perennial | Toensmeier 2016 |
| Guanacaste | Enterolobium cyclocarpum | Nitrogen fixing, livestock fodder | Perennial | Toensmeier 2016 |
| Psonay | Erythrina edulis | Nitrogen fixing, livestock fodder, edible food | Perennial | Toensmeier 2016 |
| Apple-ring acacia | Faidherbia albida | Nitrogen fixing, livestock fodder, edible food | Perennial | Toensmeier 2016 |
| Honey locust | Gleditsia triacanthos | Livestock fodder, edible food | Perennial | Toensmeier 2016 |
| Golden rain | Laburnum anagyroides | Nitrogen fixing, prolific in temperate climate, toxic seeds | Perennial | Zhang et al. 2020 |
| East African newtonia | Newtonia buchananii | Livestock fodder | Perennial | Toensmeier 2016 |
| African locust bean | Parkia biglobosa | Nitrogen fixing, livestock fodder, edible | Perennial | Toensmeier 2016 |
| Palo verde | Parkinsonia aculeata | Livestock fodder, edible | Perennial | Toensmeier 2016 |
| Fara tree | Piliostigma thongii | Nitrogen fixing, livestock fodder, edible | Perennial | Toensmeier 2016 |
| Manila tamarind | Pithecellobium dulce | Nitrogen fixing, livestock fodder | Perennial | Toensmeier 2016 |
| African mesquite | Prosopis africana | Nitrogen fixing, livestock fodder | Perennial | Toensmeier 2016 |
| White carob tree | Prosopis alba | Nitrogen fixing, livestock fodder, edible food | Perennial | Toensmeier 2016 |
| Chilean mesquite | Prosopis chilensis | Nitrogen fixing, livestock fodder | Perennial | Toensmeier 2016 |
| Ghaf | Prosopis cineraria | Nitrogen fixing, livestock fodder | Perennial | Toensmeier 2016 |
| Honey mesquite | Prosopis glandulosa | Nitrogen fixing, livestock fodder, edible | Perennial | Toensmeier 2016 |
| Long-thorn kiawe | Prosopis juliflora | Nitrogen fixing, livestock fodder, edible | Perennial | Toensmeier 2016 |
| American carob | Prosopis pallida | Nitrogen fixing, livestock fodder, edible | Perennial | Toensmeier 2016 |
| Tamarugo | Prosopis tamarugo | Nitrogen fixing, livestock fodder | Perennial | Toensmeier 2016 |
| Rain tree | Samanea (=Albizia) saman | Nitrogen fixing, livestock fodder | Perennial | Toensmeier 2016 |
| Wild Cassia | Senna singueana | Livestock fodder | Perennial | Toensmeier 2016 |
Crop domestication involved phenotypic transformation that rendered species dependent on humans for propagation, whereas the breeding and crop improvement efforts that followed produced high-yielding crops that are adapted to high-input agricultural practices (Hufford et al. 2012). Traits showing clear dimorphisms (e.g. brittle vs. non-brittle rachis in cereals) between domesticated and wild forms represent the domestication process, whereas traits with a continuous distribution (e.g. seed size) between domesticated and wild relatives mostly reflect post-domestication evolution directed by local farmers (Abbo et al. 2014). During domestication, yield and harvestability were simultaneously enhanced; thereafter, crop improvement has focused largely on yield improvement (Abbo et al. 2014, Haas et al. 2019).
Accelerating domestication on a timeline to meet the urgent need for sustainable intensification (Cassman and Grassini 2020) now seems possible with the advent of improved molecular screening methods and new breeding technologies. Here, we review strategies that could be targeted for the domestication of underutilized or even wild plants as well as ways to transform wild-type genes into domestication genes. The result could be a new repertoire of crops for environmentally friendly, sustainable and high-yielding agriculture.
What Is Meant by Yield?
Yield is essentially the amount of edible biomass that can be harvested per area of land. We must consider not only the size of seeds and grains but seed number per plant and planting density, alongside crop harvestability, losses during transport and processing, nutritional value and the presence of toxic defense compounds that reduce edibility. All these elements should be considered in strategies to increase the yield.
Defense Compounds Also Affect Yield
Plants produce secondary metabolites for defense against herbivores and pathogens, which must be considered during breeding (Kaiser et al. 2020). Plants naturally protected this way are beneficial from a sustainability perspective but may be inedible. Toxic secondary metabolites in plants include cyanogenic glucosides, terpenes, glycoalkaloids, quinolizidine alkaloids, saponins and erucic acid (Kaiser et al. 2020). For example, alkaloids and glycoalkaloids, many of which are toxic to humans, are common to Solanaceae, including pepper, tomato, eggplant and potato. The selection against bitter tubers helped to domesticate potato by reducing tuber glycoalkaloids (Kozukue et al. 1987, Friedman et al. 2003). Meanwhile, domestication of wild mustard (Brassica oleracea) to domesticated cabbage resulted in a significant decrease of both constitutive and induced chemical defenses (e.g. aliphatic glucosinolates and indolic glucosinolates) in the leaves (Moreira et al. 2018). Secondary metabolites are common domestication traits (Meyer et al. 2012) originating either through natural evolution (e.g. zoochory) or in response to human appetites (e.g. human-directed cultivation toward increased sugar content, pleasant aroma or favorable colors, while reducing bitterness and acidity) (Alseekh et al. 2021).
The Origin of Agriculture and Plant Breeding
Human agricultural activities date back to the Neolithic Period, about 10,000–15,000 years ago. Agriculture was associated with a major change in human history: a culture of hunting and gathering seeds shifted to one of farming and establishing settlements. The time and place of origin of agriculture have been extensively explored using improved bioarcheological methods. Accumulated data from archaeobotany and plant genomic studies have clarified where this transition happened, identifying several centers of agricultural origin around the world (Lev-Yadun et al. 2000, Price and Bar-Yosef 2011, Gepts et al. 2012, Larson et al. 2014, Jones et al. 2021). However, what prompted our ancestors to change their lifestyle in this way remains debated (Cohen 1977, Jones et al. 2021, Martin-Merino 2021).
During the practice of agriculture, humans accumulated extensive knowledge about plants, allowing them to identify and benefit from important yield-related traits (Harlan 1975) and some plant species were selected based on their increased yield (Fig. 1).
Fig. 1.
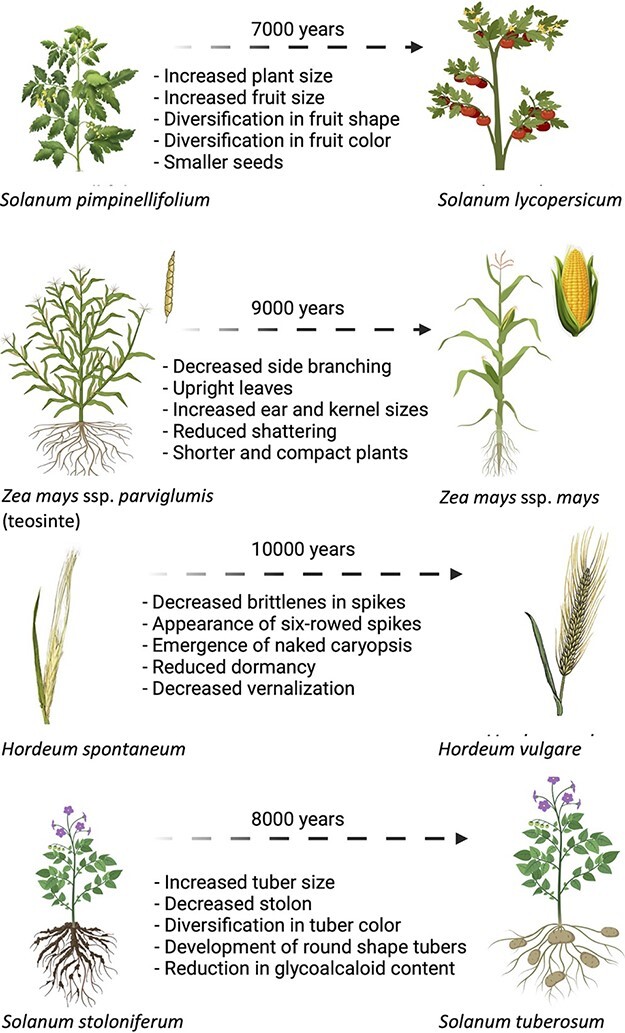
Phenotypic alterations of different plant species during domestication directed by human selection toward increasing yield. Despite common belief, most of the gained traits are the result of mutations that result in the loss of natural functions. See text for details.
The first scientific works to unveil the principles of trait inheritance and effects of selection in plants and animals were conducted by Charles Darwin (1802–1889) (Darwin 1868) and Gregor Mendel (1822–1884), and a combination of their works founded the basis of genetics, evolution and modern plant breeding. Rediscovery of Mendel’s work in the early 20th century was exploited widely by plant breeders to study traits and improve varieties through controlled crosses in different crop species such as maize and self-pollinating plants (East 1910, Roberts 1919, Brink 1921). Plant breeding has been defined as a deliberate manipulation of plants to improve the quality and quantity of yield in a permanent and heritable way (Acquaah 2007, Kingsbury 2009). Plant breeding depends on genetic variation, and the aim of plant breeding is thus to identify and exploit genetic variations (Holme et al. 2019).
Modern breeding programs consist of several general steps, beginning with objectives, sourcing of germplasm, identification of target traits and environments and trait improvement and assessment, before the ultimate release of new improved cultivars. Design of breeding strategies and objectives are dictated by multiple factors, relating to mating type, trait genetic complexity, heritability and gene action, while the breeders’ equation helps determine both the selection pressure and response (Udall 2003, Mackay et al. 2009). Central to plant breeding activities is the selection of the best individuals for the desired traits from a population. Breeding procedures and techniques have been developed over many years and range from directed crosses and simple bulk selection for a trait to marker-assisted and genomic selection. These latter techniques have been significantly aided by improved phenotyping capabilities and the advent of low-cost genetic markers. Presently, hundreds to thousands of DNA markers can be used to genotype large populations and determine trait associations, identifying both trait genetic associations and individuals with superior breeding values (Meuwissen et al. 2001, Sleper and Poehlman 2006, Breseghello and Guedes Coelho 2013, Crossa et al. 2021).
Diversity is the raw material for selection and assures the productivity, resilience and adaptive capacity of agricultural systems (Renard and Tilman 2019, Khoury et al. 2022). In traditional agroecosystems, genetically heterogeneous orphan crops, selected by local farmers and traded among communities, are cultivated in a mosaic format (polyculture) of different varieties and species (Jarvis et al. 2008, Liebman 2018). Conversely, modern crop cultivars are bred to be genetically homogeneous and are cultivated over large geographic areas as monocultures, resulting in the gradual elimination of locally adapted landraces and their wild relatives. Furthermore, long-term farming and selection for a few desired traits in newly introduced cultivars leads to loss of diversity, termed genetic erosion (van de Wouw et al. 2010).
Over a century ago, concerns were raised about the consequences of the disappearance of traditional landraces and its effects on the future of plant breeding (Baur 1914). However, the Green Revolution of the 1960s accelerated genetic erosion by intensifying directional selection, whereupon elite high-yielding cultivars replaced well-adapted local landraces. Three-quarters of crop diversity is estimated to have been lost in the 20th century (FAO Annual Report 2004), coincidental with repeated reports of the susceptibility of modern cultivars to pests and disease outbreaks and abiotic stresses due to genetic homogeneity (Keneni et al. 2012). For example, the Irish potato famine can be attributed to the large-scale cultivation of the potato cultivar ‘Aran Banner’, which although very productive was susceptible to fungal disease late blight (Phytophthora infestans). The resultant famine had catastrophic socio-economical impacts in Ireland and the rest of Europe during the 1840s (Goodwin et al. 1994).
The term ‘genetic erosion’ is used to denote different degrees of gene pool shrinkage: an absolute loss of cultivars or landraces (Gao 2003, Tsegaye and Berg 2007), reduction in richness (reduction in the total number of crops, varieties or alleles) (Hammer et al. 1996, Dyer et al. 2014) and a reduction of specific rare alleles (Khlestkina et al. 2004). While overwhelming evidence indicates that crop diversity has declined across multiple scales, studies indicate a wide array of trends in particular instances, with uncertainty remaining about the significance of changes observed (Khoury et al. 2022).
How Can Modern Crops be Refurbished with the Traits They Have Lost?
Traditionally, introducing new variation into crops has been performed through backcross breeding (Vogel 2009). Breeders have attempted to tackle the issue of genetic erosion through crossing cultivars with landraces and wild relatives harboring desired traits to introduce novel variation (Breseghello and Guedes Coelho 2013, Sharma et al. 2021). However, this strategy has its limitations. First, it can only be used to introduce traits for which genetic variation already exists. Second, the outcome of a cross is often associated with undesirable traits obtained from the donor parent due to linkage drag, which need to be eliminated through multiple backcrosses over several cycles. Third, the strategy is largely limited to monogenic traits, whereas most agronomically important traits are polygenic and difficult to maintain after a cross to non-elite material (Kushwah et al. 2020). Backcross breeding has been utilized most successfully to introduce single genes that provide new resistance or control other qualitative traits. However, the majority of yield-related traits are polygenic, i.e. composed of multiple genes of small-effect size with accumulative effects (Cooper et al. 2009).
Another more recent approach to improve the hardiness of major crops utilizes transgenesis. This approach is independent of the ability of plants to form crosses with each other; therefore, the possibilities are, at least theoretically, greater. For example, a resistance trait found in a tolerant species can be transferred to an unrelated elite variety of a crop to confer disease resistance (Bailey-Serres et al. 2019). It has been proposed that transgenic technologies will inevitably be deployed for most major crops in the future (Tester and Langridge 2010). However, although transgenesis became possible approximately 40 years ago, there are few examples where it has been commercially viable as a technique to move a trait from one plant to another, with only a few transgenic crops successfully entering the market (Raman 2017). Single stress–responsive genes can be overexpressed to confer salt and drought tolerance, but this comes with a cost as transgenic plants may have reduced fitness in the absence of stress (Wani et al. 2017). Even simple traits are the result of many different genes that work together in a coordinated fashion, and establishing a new trait furthermore requires another layer of regulatory genes. Ultimately, the same problems exist with backcross breeding, as the gene ‘package’ to be transferred increases in size. For example, it is estimated that about 200 genes function together to govern the ability of legumes to utilize atmospheric nitrogen (Roy et al. 2020). Additionally, the limited success of transgenic crops is associated with public resistance against the use of genetically modified organisms (GMOs), which in many countries are considered on a case-by-case basis under strict regulatory systems (Raman 2017).
Prime Candidates for Rapid Domestication Will Meet Urgent Agricultural and Ecological Needs
Rapid domestication may address issues such as free-threshing, seed shattering, lodging, seed size or antinutritional compounds. However, grain, or seed, yield remains a highly polygenic trait likely requiring decades of breeding to deliver improvement, as has been the case for modern grain crops. Therefore, in the near term, we do not expect newly domesticated crops to replace the dominant crops, but de novo domestication of alternative crops could help solve critical human and environmental challenges within a decade.
First, domestication of new crops combined with breeding improvement of orphan crops has the potential to address nutrient deficiencies and stabilize food supply by increasing crop diversity. In Africa, orphan crops have the potential to address vitamin and mineral deficiencies and produce food even under low-input conditions, but these crops often lack full domestication (Jamnadass et al. 2020). For example, fonio millet (Digitaria exilis) has the potential to produce reliable yields of edible grains under harsh conditions, but shattering, lodging and small seed size pose significant agronomic challenges (Abrouk et al. 2020). Similarly, the orphan crop tef (Eragrostis tef) has a beneficial nutritional profile but is limited by lodging, shattering and tiny seeds (Girija et al. 2021). Salinization of land is also dramatically limiting yields in many regions, for which direct domestication of salt-tolerant species has the potential to bring these degraded lands back into production (Razzaq et al. 2021). Consequently, for this strategy to be successful, domestication and yield improvement must go hand in hand, as was the case with the introduction of wheat to Canada in the mid-1800s. Here, wheat breeding efforts trace their origin to the variety ‘Red Fife’, which most likely originated in Ukraine and was introduced by Scottish settlers. Compared to other varieties tested, ‘Red Fife’ tolerated the short season and intense continental climate of the Canadian prairies, was rust resistant and ripened before the onset of autumn frosts (Symko 1999). Similarly, if orphan crops are to be domesticated for cultivation outside of their natural range, breeding efforts should consider how to adapt plants to new climatic zones, soil types and biotic interactions, alongside yield traits.
Depending on the plants chosen, direct domestication has the potential to produce new crops with unprecedented potential to protect against soil erosion, sequester atmospheric carbon dioxide, build soil quality and protect ground and surface waters from agricultural runoff as perennial grain-producing crops are developed (Glover et al. 2010). For example, intermediate wheatgrass (Thinopyrum intermedium) is being produced on a small scale and marketed under the name Kernza™. While the grain yield of intermediate wheatgrass remains much lower than other annual grain crops, the ability of a perennial grain to protect soil and scavenge nitrate from deep in the soil profile makes the crop highly attractive. For example, the crop can be used as part of landscape restoration, with plantings used to protect wellheads and streams from nitrate contamination or surface runoff (Muckey 2019). Intermediate wheatgrass has already undergone some improvements in later breeding cycles, and, as breeding solves lodging and increases seed size, the crop is expected to be grown on expanded acres with widespread environmental benefits (Crain et al. 2021).
Finally, accelerated domestication could expand the role of long-lived woody species in agriculture. Agroforestry has a unique potential to facilitate both climate change mitigation and adaptation. For example, tree crops meet human food and fiber needs while building soil quality, reducing runoff and tapping into water and nutrients beyond the reach of typical crops (Duguma et al. 2014). Unfortunately, due to their extended life cycle, most trees have experienced little attention from plant breeders (Molnar et al. 2013). Hybrid poplar has undergone genetic improvement as a biofuel crop but also has the potential as a fodder crop (Charlton et al. 2003, Davis 2008). With new approaches allowing rapid domestication of tree crops, benefits to sustainable food production could be profound.
What We have Learned: Domestication Generally Results from Loss of Function, not from Gain of Function
Domestication genes are genes that when mutated result in a domestication phenotype. Identification of domestication genes in current crops has led to interesting findings regarding the nature of such mutations. In an analysis of 60 domestication genes, Meyer and Purugganan (2013) found that in 28 cases such mutations resulted in the loss of function (e.g. frameshifts and splicing defects that lead to premature truncations). The second largest class were cis-regulatory mutations (15 of 60 genes). In 10 domestication genes, the domestication phenotype resulted from either loss of function or cis-regulatory mutations. These results suggest that loss-of-function mutations or ones that alter gene expression are the basis for human-driven crop evolution. Additional examples are given by Gross and Olsen (2010) and Østerberg et al. (2017).
Loss-of-function variants of transcription factors regulating the biosynthesis of toxic metabolites may give rise to less toxic plants. In the Andes, domestication of quinoa (Chenopodium quinoa) produced many cultivars with various bitter and sweet grain phenotypes (Bazile et al. 2016). Genomic analysis of bitter and sweet cultivars suggests mutation events in the TSARL1 gene, expressed mainly in seeds, are associated with a reduction in saponin concentration of seed pericarps (Jarvis et al. 2017). TSARL1 encodes a basic helix-loop-helix transcription factor, and in sweet cultivars tested, the expression of several genes in the saponin biosynthesis pathway was downregulated (Jarvis et al. 2017).
Cis-regulatory mutations are in gene regulatory regions of DNA and may affect the amount of expressed protein but not its nature. Although it can be argued that cis-regulatory mutations do not lead to loss of function (Gross and Olsen 2010), they result in loss of natural control, likely important for survival in nature. Loss-of-function alleles can also be dominant, as exemplified in einkorn wheat (Triticum boeoticum, wild ancestor of the domesticated form of einkorn, Triticum monococcum), where seeds are more firmly attached to the mother plant, making them easy to harvest (Fans et al. 2005). The observed trait is due to a single-base mutation in a gene named q, which gives rise to the dominant Q allele (MacKey 1954). From a natural selection perspective, this mutation is not actually beneficial, as it reduces the ability of plants to shatter and readily disperse their seed. However, the early farmers selected and multiplied these seeds, maximizing both threshability and the harvest index. Indeed, selection for this phenotype may be one of the first human plant breeding activities, which eventually led to the domestication of modern einkorn wheat, T. monococcum (Schlegel 2018). The levels of the wild-type q gene product are regulated by a microRNA (microRNA172 or miR172). This binds to the q mRNA, and the resulting double-stranded RNA is subjected to rapid degradation. The single point mutation underpinning the two different alleles, converting q to Q, occurs in the sequence that miR172 anneals to and weakens the interaction between Q mRNA and miR172. As a result, fewer gene transcripts are degraded and more protein is synthesized. In this way, the loss of regulation results in greater accumulation of the Q protein and a dominant domestication trait: loss of seed shattering.
The difference between domestication and breeding is sometimes contentious, for example, in the GPC-1 locus (NAM-1) in wheat. Haudry et al. (2007) and Luo et al. (2007) argued that the GPC-1 locus is under domestication and of monophyletic origin, whereas Feldman and Kislev (2007) and Thanh et al. (2014) argued the opposite. Meanwhile, Dubcovsky and Dvorak (2007) suggested indirect fixation of GPC-B1 occurred due to its pleiotropic effect on increasing grain size. In the majority of hexaploid bread wheat (Triticum aestivum) cultivars, the NAM-B1 homoeolog is nonfunctional, except for a group of Fennoscandian varieties (Hagenblad et al. 2012). Adoption of the functional NAM-B1 copy under these environments is thought to be advantageous, due to accelerating senescence under short growing seasons, alongside grain protein content improvement (Asplund et al. 2013). Meanwhile, haplotype variation has also been reported for the NAM-A1 homoeolog (Cormier et al. 2015) and is present in several Australian cultivars, where it is associated with differences in grain maturation and nitrogen use efficiency (Alhabbar et al. 2018).
A common misconception is that the high-yielding crops we grow today have gained something (i.e. an improved gene) compared to their wild progenitors. However, domestication can be seen as human-driven evolution that selects for the loss of traits or functions that are superior for plant survival and propagation in nature. Upon fixation of these domestication traits, subsequent or parallel breeding efforts have also selected loss-, or alteration-, of-function genes that confer important adaptative, agronomically important or quality traits (Haas et al. 2019). In wheat, this includes the selection of promoter deletions of Ppd-A1a and Ppd-D1a, which are associated with earlier flowering under increasing daylengths (Shaw et al. 2013). Other examples include loss-of-function mutations in the granule-bound starch synthase I (GBSSI) Waxy gene in millet, which results in glutinous grains (Hunt et al. 2010); PsMLO1 in pea, which confers mildew resistance (Humphry et al. 2011); and OsBadh2 in rice, which contributes to fragrance (Chen et al. 2008).
Loss-of-Function Strategies Cannot Establish Traits that Require New Genes
Genes are the basic units of heritable traits. For example, nitrogen-fixing legumes are equipped with multiple genes essential for the symbiosis with Rhizobium that cereal crops do not possess. Thus, it is obvious that for a cereal crop to become nitrogen fixing it requires new genes (Mus et al. 2016). In contrast, even complex traits can disappear if just a single gene in the pathway is mutated. Breeding strategies that only require loss-of-function mutations are much more likely to succeed than those that require gain-of-function genes. However, they are unlikely to provide new beneficial traits that allow the plant to survive environmental challenges.
It thus follows that a breeding strategy for the development of new crops tolerant of adverse environmental conditions must be based on plants already adapted to the different target breeding environments or stresses (Østerberg et al. 2017). Thus, if an extremely drought-resistant crop is needed, efforts could focus on identifying and domesticating an edible desert plant, instead of introgressing novel traits into a previously domesticated species. Even when taking into consideration that resilience has a cost, growth potential and drought tolerance need not be trade-offs (Fernández and Reynolds 2000). However, the potential trade-off between stress tolerance and yield must be considered (Gambino and Vilela 2011, Koziol et al. 2012) due to the associated production of costly proteins, stomatal closure or large storage organs, which may reduce the yield (Fereres et al. 2014).
How to Identify Orthologs of Domestication Genes in a Wild Plant?
Genetic variants of natural gene orthologs underpin plant domestication events, resulting in novel domestication phenotypes. Thus, the strategy for accelerated domestication is to introduce mutations in such orthologous genes. Genes, which when mutated in progenitors of our major crops, result in domestication phenotypes, as listed by Østerberg et al. (2017). However, how would one identify true orthologs of domestication genes in orphan crops or wild unrelated plants?
Homology search of domestication genes through mining databases is a useful method for the identification of potential target genes but usually produces many candidates. A problem with a strategy that depends on loss-of-function mutagenesis is that even diploid plants may have multiple versions of a gene possessing the same function, termed functional redundancy. This problem is even more significant in polyploid species due to the presence of homoeologous gene copies. We suggest performing functional complementation of already domesticated crops as a screening method to identify true orthologs of domestication genes in polyploid species. Pourkheirandish et al. (2015) demonstrated that introduction of functional Brittle rachis1 (Btr1) and Btr2 from wild Hordeum spontaneum could revert a domestication phenotype to wild type when transferred into domesticated Hordeum vulgare.
An example of an interesting domestication gene is the rice semi-dwarfing gene (sd-1), known as the ‘green revolution gene’. This allele provides rice cultivars with short, thick culms that are associated with increasing harvest index, improving lodging resistance and responsiveness to nitrogen fertilizer (Hedden 2003). The dwarfing gene originated from a Chinese cultivar, Dee-geo-woo-gene, which encodes a 383-bp deletion in the GA20ox gene (OsGA20ox2), leading to deficiency in gibberellin (GA) synthesis (Monna et al. 2002, Spielmeyer et al. 2002). If the introduction of an sd-1 homolog from a wild grass in a semi-dwarf rice can revert the domesticated phenotype, this gene would be a true ortholog of sd-1, confirming it as a domestication gene. Such a complementation strategy for the identification of true orthologs of domestication genes should take advantage of manageable diploid domesticated crops, such as rice and barley, which are readily transformable (Nishimura et al. 2007, Harwood and Smedley 2009, Sahoo et al. 2011, Marthe et al. 2015) and for which extensive genomic resources are available (Matsumoto et al. 2005, Mayer et al. 2012).
Approaches for Achieving Genetic Variation by Mutagenesis
Two major techniques are available that create genetic variation through mutagenesis: random mutagenesis (mutation breeding) and precision mutagenesis (genome editing) (Chen et al. 2019, Gao 2021) (Fig. 2).
Fig. 2.
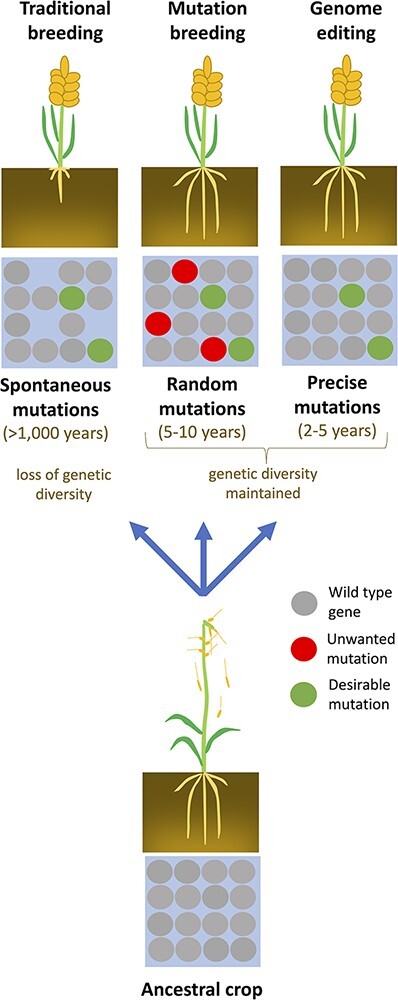
Overview of different strategies for domestication of plants. Traditional breeding depends on spontaneous mutations and is associated with the loss of genetic diversity due to inbreeding. Genetic diversity can be gained by mutagenesis techniques. Random mutagenesis may result in many unwanted mutations. These can be removed by repeated rounds of backcrossing. Genome editing techniques can induce specific mutations without the loss of genetic diversity.
Random mutagenesis
Until the last century, spontaneous mutations were the only source of genetic diversity. However, since the discovery in the late 1920s that experimentally induced mutagenesis had the potential to accelerate plant breeding, induced mutagenesis has been widely used as a tool in practical breeding programs (Oladosu et al. 2015). Mutation breeding relies on mutagens such as chemicals or radiation that randomly induce mutations in the genome (Holme et al. 2019, Ma et al. 2021). Each affected genome may carry hundreds or thousands of mutations, mainly with unknown consequences (Holme et al. 2019). Nevertheless, mutagens can increase genetic variation, and new varieties of crops resulting from random mutagenesis are readily released (Halford 2019).
Application of random mutagenesis was adopted by the Joint Food and Agriculture Organization (FAO)/International Atomic Energy Agency (IAEA) after the 1960s (Oladosu et al. 2015). Crop varieties originating from mutagenesis are reported to the Joint FAO/IAEA program and recorded in the searchable Mutant Variety Database (MVD; available from: http://mvd.iaea.org/, Fig. 3). Physical and chemical mutagenesis leads to different types of mutations. Physical mutagenesis, induced by X-rays, UV and gamma radiation, mainly causes large chromosomal deletions, rearrangements and point mutations (single-base substitutions or deletions) (Shu et al. 2012). Most of the gene mutations induced by physical mutagenesis effectively generate either knockouts or nonfunctional genes, although the majority are recessive (90–100%) (Rakszegi et al. 2010, Mba et al. 2012).
Fig. 3.
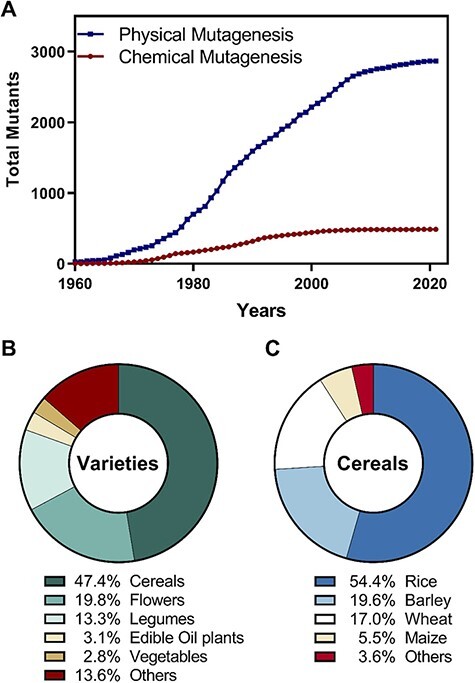
Mutant varieties officially registered in the Mutant Variety Database (MVD; available from: http://mvd.iaea.org/). (A) Evolution of the release of mutant varieties produced by physical or chemical mutagenesis, from 1960 to the present. (B) Relative proportion of mutants released in the principal groups of crops. (C) Relative proportion of cereal mutant varieties.
Chemical mutagens [e.g. N-methyl-N-nitrosourea (MNU), sodium azide (NaN3), hydrogen fluoride, methyl methanesulfonate, ethyl methanesulfonate (EMS)] almost exclusively cause single-nucleotide polymorphisms, mostly GC-to-AT base pair transitions. Currently, EMS and MNU are the most used mutagens due to their high efficiency and ease of application without the need for specialized equipment. Notably, they are being used to create large, highly mutagenized populations to screen for phenotypically relevant changes and specific genomic sequences (Jankowicz-Cieslak and Till 2016) and are widely used in many crops, including rice and barley (Till et al. 2007, Talamè et al. 2008, Uauy et al. 2009, Krasileva et al. 2017).
Due to promotion by the FAO/IAEA agency, some important cultivars have been developed by utilizing physical mutagenesis. Among them are the stiffed-strawed barley varieties (Diamond and Golden Promise) carrying mutant dwarfing genes (sdw1 and DEP1, respectively) and the rice semi-dwarf varieties (Calrose 76, Reimei), carrying a mutation in the gene sd1, previously selected during the Green Revolution (Forster et al. 2001, Hedden 2003, Wendt et al. 2016). In the minor crop lentil (Lens culinaris), chemical mutagenesis has been used to generate tolerance to imidazolinone herbicides (Canadian Food Inspection Agency 2004, Slinkard et al. 2007). Additionally, chemically induced mutations in the barley Mlo gene, rendering the gene nonfunctional and the plant less susceptible to powdery mildew (Blumeria graminis sp. hordei) infection, have been implemented in barley breeding programs (Jørgensen 1992). Varieties developed from the use of random mutagenesis have been used in breeding programs and subsequent variety release.
Notwithstanding the high number of mutant varieties registered in MVD (Fig. 3A) and some commercial success, most of the random mutagenesis methodologies were used to improve already domesticated crops (Fig. 3B-C), including the four major cereal crops (rice, barley, wheat and maize) (Ahloowalia et al. 2004). Applications include tailoring barley for industrial needs, such as for malting and brewing (Knudsen et al. 2020). However, the use of random mutagenesis in orphan or crop wild relatives has been neglected.
Precision mutagenesis
Methods of genome editing techniques have been developed to introduce precise and predictable genome modifications in plants, giving rise to precision breeding (Chen et al. 2019, Gao 2021). To date, the most precise method of mutagenesis in plants is gene editing via site-directed nucleases. Among different techniques, clustered regularly interspaced short palindromic repeats/CRISPR-associated protein9 (CRISPR/Cas9) has become a widely used tool due to its higher efficiency, reduced number of off-target mutations and methodological simplicity (Voytas 2013, Chen et al. 2019). CRISPR/Cas systems provide an unprecedented possibility for precise targeting of homologs of domestication genes in virtually any plant, providing it is amenable to transformation (Wolter et al. 2019). Thus, CRISPR/Cas technology offers an unmatched opportunity to stimulate domestication of neglected, semi-domesticated or wild plants. However, despite the advantages of CRISPR/Cas, new varieties produced by precision mutagenesis are considered GMOs in many countries, making them subject to strong regulatory regimes.
Genome Editing as a Promising Tool to Accelerate Orphan Crop Domestication
Multiplex genome editing and trait stacking
Numerous orphan crops and neodomesticates, such as intermediate wheatgrass (T. intermedium), quinoa (C. quinoa) and wild potato (Solanum stoloniferum), are polyploids (DeHaan et al. 2020, López-Marqués et al. 2020, Del Mar Martínez-Prada et al. 2021). In polyploids, each gene has multiple copies associated with their multiple genomes (homoeologs). Homoeologous genes often demonstrate functional redundancy, and depending on their expression patterns, mutations in multiple homoeologs are often required to obtain the expected phenotype (Adamski et al. 2020). Moreover, when genome data from 16 fully sequenced plant genomes were analyzed, 72% of protein-coding genes belonged to paralogous gene families (Hyams et al. 2018). Many paralogs share similar structures and functions, requiring the simultaneous mutation of numerable paralogs to acquire the desirable phenotype. Genome editing strategies can be multiplexed for simultaneously editing several homoeologous or paralogous copies through allowing multiplex single guide RNAs (sgRNAs) to be transcribed in a single cell (Minkenberg et al. 2017). Using this strategy, Wang and colleagues edited all three MLO homoeologs in bread wheat to create powdery mildew-resistant mutants (Wang et al. 2014). Similarly, using a CRISPR/Cas9 approach, up to 35 α-gliadin paralogs were knocked out in a bread wheat mutant to reduce celiac disease immunoreactivity (Sánchez-León et al. 2018).
Editing of quantitative trait variation
A trait may result from the combined effect of several genes, each of which may have a minor effect and may interact with the environment. Quantitative variation may also be the result of different alleles of the same gene. A domestication quantitative trait was created in tomato via CRISPR/Cas9 targeting of a promoter in tomato (Rodríguez-Leal et al. 2017). Thus, multiplex editing of the cis-regulatory motif in the SlCLV3 gene generated an array of locule number variations in the fruit.
Editing of QTLs
One of the challenges in orphan crop domestication is the improvement of a trait controlled by a quantitative trait locus (QTL), which refers to a specific genetic region to which a mutation(s) causing a quantitative trait has been mapped. However, due to the varying degree of accuracy associated with QTL mapping, a single mapped QTL may cover hundreds of genes that might also have been mutated but are not responsible for the phenotype under question. Thus, the actual genes and mutations responsible for most QTLs are not known, making it extremely difficult to modify these loci by gene editing. QTLs were targeted in maize via multiplex CRISPR/Cas9, which allowed for high-throughput targeting of over 1,000 candidate genes derived from genetic mapping and comparative genomic analysis (Liu et al. 2020).
Haploid induction and artificial apomixis
Domestication of orphan crops via traditional breeding takes six to seven generations of self-pollination to produce homozygotes. To accelerate domestication of orphan crops, doubled haploid technology, which rapidly fixes recombinant haploid genomes within two generations, is promising. Haploid inducer lines can be created via the editing of endogenous genes. For example, one Arabidopsis haploid inducer line was successfully produced using CRISPR/Cas9 to induce a deletion in the N terminus of CENH3 (Kuppu et al. 2020). Similarly, a maternal haploid was induced in maize, rice and wheat via knockout of the ZmPHOSPHOLIPASE-A1 (ZmPLA1) gene, also known in other species as MATRILINEAL (OsMTL) and NOT LIKE DAD (TaNLD) (Zhong et al. 2019).
Another strategy to accelerate the breeding of orphan crops is artificial apomixis, which rapidly produces genetically identical seeds (Sailer et al. 2016). Apomixis seldom occurs naturally in major crops (Willmann 2019) but can be induced by CRISPR/Cas9. For example, apomeiosis was induced in rice via CRISPR/Cas9-mediated knockout of the meiotic genes REC8, PAIR1 and OSD1 (Khanday et al. 2019, Wang et al. 2019). Genome editing of these genes in orphan crops might induce apomixis as these genes are highly conserved.
Large-scale screening for trait discovery
Elucidation of the genetic regulation of agronomically important traits is the prerequisite of orphan crop domestication. CRISPR/Cas9 screening is a promising reverse genetic approach to characterize the relationships between genotypes and phenotypes at the genome scale (Gaillochet et al. 2020). During CRISPR/Cas9 screening, a library of sgRNAs capable of targeting almost all genes in a single plant genome are co-expressed with Cas9. Subsequently, edited plants are screened for the trait of interest, with the genes controlling a trait of interest further identified based on the sgRNA corresponding sequences. CRISPR/Cas9 screening has been successfully applied to identify genes controlling agronomically important traits genome-wide in maize, rice, tomato and soybean (Jacobs et al. 2017, Lu et al. 2017, Meng et al. 2017, Bai et al. 2020, Liu et al. 2020). With the advent of the genome era, increasing numbers of orphan crops have been genome sequenced, which makes CRISPR/Cas9 screening possible as sgRNAs targeting all genes in the genome can be designed based on the genomic information.
Transport engineering
Accelerating de novo domestication of wild toxin–produced plants by genome editing requires the identification of several target genes. These can be genes encoding biosynthetic enzymes or transcription factors, which regulate the expression of such enzymes in target organs, or transport proteins involved in translating the synthesized metabolites to target organs. Genomic sequences of genes of interest in wild plants first need to be established by sequencing tools and bioinformatics analysis. Sequencing and expression analysis of mRNA transcripts could specify the genes expressed, mainly, in the target organs or tissues in the wild plants. Large-scale functional screens can be employed to identify the transport proteins of specific metabolites (Jørgensen et al. 2017).
The challenge is to breed for plants that have edible grains (or other plant parts) but are otherwise naturally protected. In this way, some parts of the domesticated plant are palatable and safe for human consumption, while the rest of the plant is protected against biotic and abiotic factors. If the biosynthetic pathway of a secondary metabolite is disrupted, this secondary metabolite will disappear in all parts of the plant. However, a plant unable to generate its own protective metabolites will need to be protected by other means, e.g. by application of pesticides, which can be environmentally damaging. In this context, it is noteworthy that secondary metabolites are often synthesized in specialized cells whereupon they are transported to other plant parts (Jørgensen et al. 2015). For example, in Nicotiana species, nicotine is synthesized in the root and then transferred to leaves (Hashimoto and Yamada 2003). In contrast, in cassava (Manihot esculenta), cyanogenic glucosides are synthesized in leaves before being transferred to roots (Gleadow and Møller 2014), while seeds of Brassica napus import glucosinolates from maternal tissues (Nour-Eldin et al. 2017). An alternative strategy would be to leave the defense compound biosynthesis pathway intact, enabling the plant to be protected, while neutralizing specific transporters involved in their allocation to harvested tissues. Therefore, genes encoding metabolite transporters could be promising targets for domestication of wild plants in the future.
Considering the advances in plant genome editing, many countries are discussing easier regulatory pathways for approving edited crops. Present regulatory systems remain a critical limitation for applying gene editing to improve major commodity crops and specialty crops and to neodomesticate new species. These regulatory systems limit the potential of edited crops to secure not only food but also sustainability of cropping systems.
Challenges for Genome Editing of Orphan Crops Include Improving Their Regeneration Efficiency
Genome editing has been used successfully to accelerate the domestication of orphan crops. For example, wild tomato (Solanum pimpinellifolium) was partially domesticated using multiplex editing of several domestication-related genes (Li et al. 2018; Zsögön et al. 2018). Similarly, allotetraploid wild rice Oryza alta was de novo domesticated using genome editing (Yu et al. 2021). Below, we focus on some of the challenges associated with genome editing of orphan crops.
Transformation and regeneration of plants
Transformation is the prerequisite for genome editing in plants. However, most orphan crops are transformation recalcitrant, with low regeneration efficiency remaining the bottleneck of genome editing in plants. In particular, legumes, which are key to perennial agricultural systems built around neodomesticates, are notoriously recalcitrant to both regeneration and transformation (Choudhury and Rajam 2021). To solve this problem, boosters, which are developmental regulators, are used in plant transformation. For example, WUSCHEL (WUS) and BABY BOOM (BBM) improved the regeneration frequencies in a variety of transformation recalcitrant crops including maize, sorghum (Sorghum bicolor), sugarcane (Saccharum officinarum) and indica rice (Oryza sativa ssp. indica) (Lowe et al. 2016). The use of tissue-specific promoters to drive the spatiotemporal expression of WUS and BBM can circumvent their adverse effects (Lowe et al. 2018). In addition, growth regulating factors (GRFs), GRF-interacting factors (GIFs) and GRF–GIF chimeras were also used to boost the regeneration of numerous monocot and dicot plant species without any side effects (Debernardi et al. 2020, Kong et al. 2020; Luo and Palmgren 2021). TRA1 is a candidate gene determining Agrobacterium-mediated transformability of barley (Orman-Ligeza et al. 2020). Recently, TaWOX5 was proven to improve wheat regeneration and hence bypassed genotype dependency in wheat transformation (Wang et al. 2022). Combined with de novo meristem induction, boosters can also help regenerate dicots, which bypasses tissue culture (Maher et al. 2020). Collectively, the identification and application of booster homologs are important for facilitating genome editing in orphan crops.
An additional challenge of transformation and regeneration is the number of unintended mutations introduced during tissue culture, known as somaclonal variation (Neelakandan and Wang 2012). Therefore, although genome editing techniques result in a low number of off-target effects themselves, the transformation system often needed for their application can introduce mutations in the produced plants (Graham et al. 2020). Furthermore, even when transformation protocols for difficult-to-transform crops have been developed, they are largely confined to specific lines (Do et al. 2018, Che et al. 2021), limiting the capacity of genome editing in grower-preferred varieties.
Poor sequence information
Unannotated or partly annotated genomes complicate the identification of all relevant target genes. Furthermore, the lack of complete genome sequence information complicates the design of specific sgRNAs, which increases the off-target activity of CRISPR/Cas.
Complicated genetics
As mentioned above, orphan crops are often polyploids, making genome editing difficult. Allopolyploid plants have only one or a maximum of two alleles per plant for each homoeologous gene, which simplifies genotyping and inheritance to some degree, although it is still complicated by the presence of homoeologous gene copies. Additionally, polyploids can have more than two copies of a given gene (paralogs), which further complicates genotyping and inheritance. Most perennial species, such as intermediate wheatgrass, are self-incompatible, meaning creation of inbred (true breeding) lines, as is commonly performed in other domesticated cereals such as maize, rice or wheat, is not an option. Additionally, many, if not most, genes are heterozygous with at least two different alleles for each gene in each plant. In self-compatible plants, low-frequency alleles are usually found in a heterozygous state, meaning recessive traits are usually hidden by dominant alleles unless present at a relatively high frequency in the population. If the frequency of a recessive allele is 0.01 (one in 100), then the frequency of the recessive homozygote (recessive phenotype) is 0.01 × 0.01 = 0.0001 (one in 10,000) in a randomly mating diploid population. The situation is even more complicated in self-incompatible polyploid plants.
Screening for Mutations in Large Populations
As regulatory regimes for genome-edited plants are uncertain, and because the technology may not be applicable in countries hit hardest by climate change, there is an urgent need for alternative approaches for innovative plant breeding (Anders et al. 2020). Previously, mutation breeding depended on selection for visible, or readily quantifiable, traits observable during forward screens of large mutant populations. However, if the domestication gene is recessive, which is the norm, the domestication phenotype is only exhibited in plants that are homozygous for the domestication gene, and such plants are rare. However, this problem was circumvented with the development of TILLING (targeting-induced local lesions in genomes) techniques (McCallum et al. 2000, Henikoff et al. 2004) (Fig. 4). TILLING made it possible to move from screening for a phenotype (forward genetics) to screening for a genotype (reverse genetics) (Holme et al. 2019). Thus, the technique allows for the detection of mutations in a specific, known gene in a mutant population, even though it is not associated with a phenotype. The only requirements are that the DNA sequence of the target gene is known and the mutant population is suitably large (Jankowicz-Cieslak et al. 2017, Holme et al. 2019). A variant of TILLING called EcoTILLING allows for the detection of natural polymorphisms in natural populations (Comai et al. 2004).
Fig. 4.
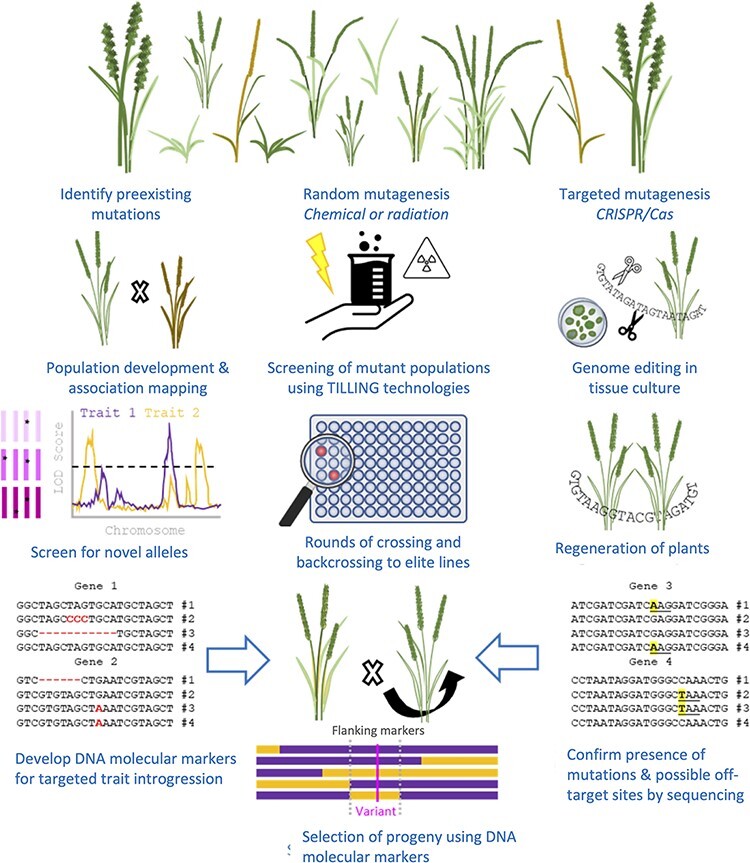
Outline of the procedure for mutagenesis and subsequent identification of plants with mutations in genes of interest. See text for details.
Although TILLING and EcoTILLING enable genome-wide scanning of mutations, they can only screen population sizes of fewer than 10,000 individuals because of technical obstacles and low detection sensitivity. To improve the screening efficiency, the Carlsberg Research Laboratory in Denmark has established the ‘Fast Identification of Nucleotide variants by DigITal PCR’ (FIND-IT) method (Knudsen et al. 2021). Due to the combination of systematic sample pooling-and-splitting and high sensitivity of droplet digital polymerase chain reaction (PCR), FIND-IT can screen a mutant population containing 500,000 individuals and isolate desired variants within 2 weeks. FIND-IT has been proven to identify knockout mutants, non-synonymous codon mutations and mutations in promoter regions and miRNA binding sites with a single-nucleotide resolution. This method has been tested for mutant screening in barley and wheat but has also been successful in other organisms such as yeast (Saccharomyces cerevisiae) and bacteria (Lactobacillus pasteurii). Considering the above advantages, FIND-IT is a promising technique to screen chemically induced mutant populations of orphan crops and wild relatives to create non-GMO crops for the future.
New varieties of plants that result from conventional methods of random mutagenesis are easy to release to the market and, in some regions, such as the European Union, are much easier to release than those resulting from precise targeted mutagenesis (Halford 2019). In these regions, random mutagenesis combined with TILLING techniques or FIND-IT may be preferable to accelerate domestication (Holme et al. 2019, Knudsen et al. 2021).
Conclusion
Nature provides a plethora of plants that have the traits required for crops in a future sustainable agriculture. The challenge now is to increase the yields of these orphan crops so they can compete with present-day elite cultivars. By traditional breeding, this would take an unacceptably long period of time, but with new breeding techniques, it has become possible to accelerate their domestication. The future of green solutions to our future food supply therefore looks bright.
Contributor Information
Guangbin Luo, Department of Plant and Environmental Sciences, University of Copenhagen, Thorvaldsensvej 40, Frederiksberg C DK-1871, Denmark.
Javad Najafi, Department of Plant and Environmental Sciences, University of Copenhagen, Thorvaldsensvej 40, Frederiksberg C DK-1871, Denmark.
Pedro M P Correia, Department of Plant and Environmental Sciences, University of Copenhagen, Thorvaldsensvej 40, Frederiksberg C DK-1871, Denmark.
Mai Duy Luu Trinh, Department of Plant and Environmental Sciences, University of Copenhagen, Thorvaldsensvej 40, Frederiksberg C DK-1871, Denmark.
Elizabeth A Chapman, Carlsberg Research Laboratory, J.C. Jacobsens Gade 4, Copenhagen V DK-1799, Denmark.
Jeppe Thulin Østerberg, Carlsberg Research Laboratory, J.C. Jacobsens Gade 4, Copenhagen V DK-1799, Denmark.
Hanne Cecilie Thomsen, Carlsberg Research Laboratory, J.C. Jacobsens Gade 4, Copenhagen V DK-1799, Denmark.
Pai Rosager Pedas, Carlsberg Research Laboratory, J.C. Jacobsens Gade 4, Copenhagen V DK-1799, Denmark.
Steve Larson, US Department of Agriculture (USDA), USDA–ARS Forage & Range Research Lab, Utah State University Logan, Logan, UT 84322, USA.
Caixia Gao, Center for Genome Editing, Institute of Genetics and Developmental Biology, Innovation Academy for Seed Design, Chinese Academy of Sciences, Beijing 100101, China.
Jesse Poland, Center for Desert Agriculture, King Abdullah University of Science and Technology, Thuwal, Makkah 23955, Saudi Arabia.
Søren Knudsen, Carlsberg Research Laboratory, J.C. Jacobsens Gade 4, Copenhagen V DK-1799, Denmark.
Lee DeHaan, The Land Institute, Salina, KS 67401, USA.
Michael Palmgren, Department of Plant and Environmental Sciences, University of Copenhagen, Thorvaldsensvej 40, Frederiksberg C DK-1871, Denmark.
Data Availability
No new datasets were generated or analyzed in this study.
Funding
NovoCrops (Novo Nordisk Foundation; project number 2019OC53580 to M.P.); the Innovation Fund Denmark (LESSISMORE to M.P.; DEEPROOTS to M.P. and S.K.); the Carlsberg Foundation (RaisingQuinoa, project number CF18-1113 to M.P.; ‘Crops for the future—tackling the challenges of changing climates’, project number CF20-0352 to S.K.).
Disclosures
The authors have no conflicts of interest to declare.
References
- Abbo S., Van-oss R.P., Gopher A., Saranga Y., Ofner I. and Peleg Z. (2014) Plant domestication versus crop evolution: a conceptual framework for cereals and grain legumes. Trends Plant Sci. 19: 351–360. [DOI] [PubMed] [Google Scholar]
- Abrouk M., Ahmed H.I., Cubry P., Šimoníková D., Cauet S., et al. (2020) Fonio millet genome unlocks African orphan crop diversity for agriculture in a changing climate. Nat. Commun. 11: 4488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Acquaah G. (2007) Principles of Plant Genetics and Breeding. Blackwell, Oxford, UK. [Google Scholar]
- Adamski N.M., Borrill P., Brinton J., Harrington S.A., Marchal C., Bentley A.R., et al. (2020) A roadmap for gene functional characterisation in crops with large genomes: lessons from polyploid wheat. Elife 9: e55646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ahloowalia B.S., Maluszynski M. and Nichterlein K. (2004) Global impact of mutation-derived varieties. Euphytica 135: 187–204. [Google Scholar]
- Alhabbar Z., Yang R., Juhasz A., Xin H., She M., Anwar M., et al. (2018) NAM gene allelic composition and its relation to grain-filling duration and nitrogen utilisation efficiency of Australian wheat. PLoS One 13: e0205448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alseekh S., Scossa F., Wen W., Luo J., Yan J., Beleggia R., et al. (2021) Domestication of crop metabolomes: desired and unintended consequences. Trends Plant Sci. 26: 650–661. [DOI] [PubMed] [Google Scholar]
- Anders S., Pareek A., Singla-Pareek S.L., Gupta K.J. and Foyer C.H. (2020) Innovative plant breeding could deliver crop revolution. Nature 577: 622. [DOI] [PubMed] [Google Scholar]
- Asplund L., Bergkvist G., Meino M.W., Westerbergh A. and Weih M. (2013) Swedish spring wheat varieties with the rare high grain protein alleles of NAM-B1 differ in leaf senescence and grain mineral content. PLoS One 8: e59704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bai M., Yuan J., Kuang H., Gong P., Li S., Zhang Z., et al. (2020) Generation of a multiplex mutagenesis population via pooled CRISPR-Cas9 in soya bean. Plant Biotechnol. J. 18: 721–731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bailey-Serres J., Parker J.E., Ainsworth E.A., Oldroyd G.E.D. and Schroeder J.I. (2019) Genetic strategies for improving crop yields. Nature 575: 109–118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baur E. (1914) Die Bedeutung der primitiven Kulturrassen und der wilden Verwandten unserer Kulturpflanzen für die Pflanzenzüchtung. pp. 104–110. Jahrbuch der DLG 29. [Google Scholar]
- Bazile D., Jacobsen S.E. and Verniau A. (2016) The global expansion of quinoa: Trends and limits. Front. Plant Sci 7: 622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benton T.G. and Bailey R. (2019) The paradox of productivity: agricultural productivity promotes food system inefficiency. Global Sustainability 2: 1–8. [Google Scholar]
- Benton T.G., Bieg C., Harwatt H., Pudasaini R. and Wellesley L. (2021) Food system impacts on biodiversity loss. Three levers for food system transformation in support of nature. Chatham House, the Royal Institute of International Affairs, London. https://www.ciwf.com/media/7443948/food-system-impacts-on-biodiversity-loss-feb-2021.pdf (January 29, 2022, date last accessed). [Google Scholar]
- Breseghello F. and Guedes Coelho A. (2013) Traditional and modern plant breeding methods with examples in rice (Oryza sativa L.). J. Agric. Food Chem. 61: 8277–8268. [DOI] [PubMed] [Google Scholar]
- Brink R.A. (1921) The genetic basis for improvement in self-fertilized crops. Sci. Agric. 2: 83–87. [Google Scholar]
- Cadima X., van Zonneveld M., Scheldeman X., Castañeda N., Patiño F., Beltran M., et al. (2014) Endemic wild potato (Solanum spp.) biodiversity status in Bolivia: reasons for conservation concerns. J. Nat. Conserv. 22: 113–131. [Google Scholar]
- Canadian Food Inspection Agency . (2004) Decision document DD2004-46: determination of the safety of the BASF Canada imidazolinone-tolerant lentil line RH44. [Online]. http://www.inspection.gc.ca/english/plaveg/bio/dd/dd0446e.shtml (March 6, 2022, date last accessed).
- Cassman K.G. and Grassini P. (2020) A global perspective on sustainable intensification research. Nat. Sustainability 3: 262–268. [Google Scholar]
- Charlton J.F.L, Douglas G.B., Wills B.J. and Prebble J.E. (2003) Farmer experience with tree fodder. InUsing Trees on Farms. Vol. 10, Edited by Charlton, J.F.L. pp. 7–16. Grassl. res. pract. ser. [Google Scholar]
- Che P., Chang S., Simon M.K., Zhang Z., Shaharyar A., Ourada J., et al. (2021) Developing a rapid and highly efficient cowpea regeneration, transformation and genome editing system using embryonic axis explants. Plant J. 106: 817–830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen K., Wang Y., Zhang R., Zhang H. and Gao C. (2019) CRISPR/Cas genome editing and precision plant breeding in agriculture. Annu. Rev. Plant Biol. 70: 667–697. [DOI] [PubMed] [Google Scholar]
- Chen S., Yang Y., Shi W., Ji Q., He F., Zhang Z., et al. (2008) Badh2, Encoding betaine aldehyde dehydrogenaase, inhbitis the biosynthesis of 2-acetyl-1-pyroline, a major component in rice fragrance. Plant Cell 20: 1850–1861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choudhury A. and Rajam M.V. (2021) Genetic transformation of legumes: an update. Plant Cell Rep. 40: 1813–1830. [DOI] [PubMed] [Google Scholar]
- Cohen M.N. (1977) The food crises in prehistory: overpopulation and the origins of agriculture. Yale University Press, New Haven. [Google Scholar]
- Comai L., Young K., Till B.J., Reynolds S.H., Greene E.A., Codomo C.A., et al. (2004) Efficient discovery of DNA polymorphisms in natural populations by Ecotilling. Plant J. 37: 778–786. [DOI] [PubMed] [Google Scholar]
- Cooper M., van Eeuwijk F.A., Hammer G.L., Podlich D.W. and Messina C. (2009) Modeling QTL for complex traits: detection and context for plant breeding. Curr. Opin. Plant Biol. 12: 231–240. [DOI] [PubMed] [Google Scholar]
- Cormier F., Throude M., Ravel C., Le Gouis J., Leveugle M., Lafarge S., et al. (2015) Detection of NAM-A1 natural variants in bread wheat reveals differences in haplotype distribution between a worldwide core collection and European elite germplasm. Agronomy 5: 143–151. [Google Scholar]
- Crain J., DeHaan L. and Poland J. (2021) Genomic prediction enables rapid selection of high-performing genets in an intermediate wheatgrass breeding program. Plant Genome 14: e20080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crossa J., Fritsche-Neto R., Montesinos-Lopez O.A., Costa-Neto G., Dreisigacker S., Montesinos-Lopez A., et al. (2021) The modern plant breeding triangle: Optimizing the use of genomics, phenomics, and enviromics data. Front. Plant. Sci 12: 651480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darwin C. (1868) The Variation of Animals and Plants under Domestication. John Murray, London. [Google Scholar]
- Davis J.M. (2008) Genetic improvement of poplar (Populus spp.) as a bioenergy crop. InGenetic Improvement of Bioenergy Crops. Edited by Vermerris, W. pp. 397–419. Springer, New York, NY.doi: 10.1007/978-0-387-70805-8_14. [DOI] [Google Scholar]
- Debernardi J.M., Tricoli D.M., Ercoli M.F., Hayta S., Ronald P., Palatnik J.F., et al. (2020) A GRF-GIF chimeric protein improves the regeneration efficiency of transgenic plants. Nat. Biotechnol. 38: 1274–1279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeHaan L., Larson S., López-Marqués R.L., Wenkel S., Gao C. and Palmgren M. (2020) Roadmap for accelerated domestication of an emerging perennial grain crop. Trends Plant Sci. 25: 525–537. [DOI] [PubMed] [Google Scholar]
- Del Mar Martínez-Prada M., Curtin S.J. and Gutiérrez-González J.J. (2021) Potato improvement through genetic engineering. GM Crops Food 12: 479–496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Do P.T., Lee H., Nelson-Vasilchik K., Kausch A. and Zhang Z.J. (2018) Rapid and efficient genetic transformation of sorghum via agrobacterium-mediated method. Curr. Protoc. Plant Biol. 3: e20077. [DOI] [PubMed] [Google Scholar]
- Dubcovsky J. and Dvorak J. (2007) Genome plasticity a key factor in the success of polyploid wheat under domestication. Science 316: 1862–1866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duguma L.A., Minang P.A. and van Noordwijk M. (2014) Climate change mitigation and adaptation in the land use sector: from complementarity to synergy. Environ. Manage. 54: 420–432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dyer G.A., López-Feldman A., Yúnez-Naude A. and Taylor J.E. (2014) Genetic erosion in maize’s center of origin. Proc. Natl. Acad. Sci. U.S.A. 111: 14094–14099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- East E.M. (1910) A Mendelian interpretation of variation that is apparently continuous. Am. Nat. 44: 65–82. [Google Scholar]
- Fans J.D., Simons K.J., Zhang Z. and Gill B.S. (2005) The wheat super domestication gene. Q. Front. Wheat Biosci. Memorial Issue, Wheat Inf. Serv. 100: 129–148. [Google Scholar]
- FAO Annual Report . (2004) Agrobiodiversity? Food and Agriculture Organization of the United Nations (FAO). What is happening to agrobiodiversity?
- FAO, IFAD, UNICEF, WFP & WHO . (2018) The State of Food Security and Nutrition in the World 2018. Building climate resilience for food security and nutrition. Rome, FAO. www.fao.org/3/I9553EN/i9553en.pdf (January 29, 2022, date last accessed). [Google Scholar]
- Feldman M. and Kislev M.E. (2007) Domestication of emmer wheat and evolution of free-threshing tetraploid wheat. Israel J. Plant Sci. 55: 207–221. [Google Scholar]
- Fereres E., Orgaz F., Gonzalez-Dugo V., Testi L. and Villalobos F.J. (2014) Balancing crop yield and water productivity tradeoffs in herbaceous and woody crops. Funct. Plant Biol. 41: 1009–1018. [DOI] [PubMed] [Google Scholar]
- Fernández R.J. and Reynolds J.F. (2000) Potential growth and drought tolerance of eight desert grasses: lack of a trade-off? Oecologia 123: 90–98. [DOI] [PubMed] [Google Scholar]
- Fisher R.A. (2015) Definitions and determination of crop yield, yield gaps, and of rates of change. Field Crops Res. 182: 9–18. [Google Scholar]
- Foley J.A., Ramankutty N., Brauman K.A., Cassidy E.S., Gerber J.S., Johnston M., et al. (2011) Solutions for a cultivated planet. Nature 478: 337–342. [DOI] [PubMed] [Google Scholar]
- Forster P., Torroni A., Renfrew C. and Röhl A. (2001) Phylogenetic star contraction applied to Asian and Papuan mtDNA evolution. Mol. Biol. Evol. 18: 1864–1881. [DOI] [PubMed] [Google Scholar]
- Friedman M., Roitman J.N. and Kozukue N. (2003) Glycoalkaloid and calystegine contents of eight potato cultivars. J. Agricult. Food Chem. 51: 2964–2973. [DOI] [PubMed] [Google Scholar]
- Gabriel D., Sait S.M., Kunin W.E. and Benton T.G. (2013) Food production vs. biodiversity: comparing organic and conventional agriculture. J. Applied Ecol. 50: 355–364. [Google Scholar]
- Gaillochet C., Develtere W. and Jacobs T.B. (2020) CRISPR screens in plants: approaches, guidelines, and future prospects. Plant Cell 33: 794–813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gambino P. and Vilela A. (2011) Morphological traits and allocation patterns related to stress-tolerance and seed-yield in wild and domesticated evening primrose (Oenothera L. Onagraceae). Ind. Crops Prod. 34: 1269–1276. [Google Scholar]
- Gao C. (2021) Genome engineering for crop improvement and future agriculture. Cell 184: 1621–1635. [DOI] [PubMed] [Google Scholar]
- Gao L. (2003) The conservation of Chinese rice biodiversity: genetic erosion, ethnobotany and prospects. Genetic Res. Crop Evol. 50: 17–32. [Google Scholar]
- Gepts, P., Famula, T.R., Bettinger, R.L., Brush, S.B., Damania, A.B., McGuire, P.E., et al. (eds.) (2012) Biodiversity in Agriculture: Domestication, Evolution, and Sustainability. Cambridge Univ. Press, Cambridge. [Google Scholar]
- Girija A., Jifar H., Jones C., Yadav R., Doonan J. and Mur L.A.J. (2021) Tef: a tiny grain with enormous potential. Trends Plant Sci. 27: 220–223. [DOI] [PubMed] [Google Scholar]
- Gleadow R.M. and Møller B.L. (2014) Cyanogenic glycosides: synthesis, physiology, and phenotypic plasticity. Annu. Rev. Plant Biol. 65: 155–185. [DOI] [PubMed] [Google Scholar]
- Glover J.D., Reganold J.P., Bell L.W., Borevitz J., Brummer E.C., Buckler E.S., et al. (2010) Agriculture. Increased food and ecosystem security via perennial grains. Science 328: 1638–1639. [DOI] [PubMed] [Google Scholar]
- Godfray H.C., Beddington J.R., Crute I.R., Haddad L., Lawrence D., Muir J.F., et al. (2010) Food security: the challenge of feeding 9 billion people. Science 327: 812–818. [DOI] [PubMed] [Google Scholar]
- Goodwin S.B., Cohen B.A. and Fry W.E. (1994) Panglobal distribution of a single clonal lineage of the Irish potato famine fungus. Proc. Natl. Acad. Sci. 91: 11591–11595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graham N., Patil G.B., Bubeck D.M., Dobert R.C., Glenn K.C., Gutsche A.T., et al. (2020) Plant genome editing and the relevance of off-target changes. Plant Physiol. 183: 1453–1471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gross B.L. and Olsen K.M. (2010) Genetic perspectives on crop domestication. Trends Plant Sci. 15: 529–537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haas M., Schreiber M. and Mascher M. (2019) Domestication and crop evolution of wheat and barley: genes, genomics, and future directions. J. Integr. Plant Biol. 61: 204–225. [DOI] [PubMed] [Google Scholar]
- Hagenblad J., Asplund L., Balfourier F., Ravel C. and Leino M.W. (2012) Strong presence of the high grain protein content allele of NAM-B1 in Fennoscandian wheat. Theor. Appl. Genet. 125: 1677–1686. [DOI] [PubMed] [Google Scholar]
- Halford N.G. (2019) Legislation governing genetically modified and genome-edited crops in Europe: the need for change. J. Sci. Food Agric. 99: 8–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammer K., Knüpffer H., Xhuveli L. and Perrino P. (1996) Estimating genetic erosion in landraces—two case studies. Genet. Resour. Crop Evol. 43: 329–336. [Google Scholar]
- Harlan J.R. (1975) Crops and Man. American Society of Agronomy and Crop Science Society of America, Madison, WI. [Google Scholar]
- Harwood W.A. and Smedley M.A. (2009) Barley transformation using biolistic techniques. Methods Mol. Biol. 478: 125–136. [DOI] [PubMed] [Google Scholar]
- Hashimoto T. and Yamada Y. (2003) New genes in alkaloid metabolism and transport. Curr. Opin. Biotechnol. 14: 163–168. [DOI] [PubMed] [Google Scholar]
- Haudry A., Cenci A., Ravel R., Batallion T., Brunel D., Poncet C., et al. (2007) Grinding up wheat: a massive loss of nucleotide diversity since domestication. Mol. Biol. Evol. 24: 1506–1517. [DOI] [PubMed] [Google Scholar]
- Hedden P. (2003) The genes of the green revolution. Trends Genet. 19: 5–9. [DOI] [PubMed] [Google Scholar]
- Henikoff S., Till B.J. and Comai L. (2004) TILLING. Traditional mutagenesis meets functional genomics. Plant Physiol. 135: 630–636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holme I.B., Gregersen P.L. and Brinch-Pedersen H. (2019) Induced genetic variation in crop plants by random or targeted mutagenesis: convergence and differences. Front. Plant Sci. 10: 68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hufford M.B., Xu X., van Heerwaarden J., Pyhäjärvi T., Chia J.M., Cartwright R.A., et al. (2012) Comparative population genomics of maize domestication and improvement. Nat. Genet. 44: 808–811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Humphry M., Reinstådler A., Ivanov S., Bisseling T. and Panstruga R. (2011) Durable broad-spectrum powdery mildew resistance in pea er1 plants is conferred by natural loss-of-function mutations in PsML01. Mol. Plant Pathol. 12: 866–878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hunt H.V., Denyer K., Packman L.C., Jones M.K. and Howe C.J. (2010) Molecular basis of the waxy endosperm starch phenotype in broomcorn miller (Panicum miliaceum L.). Mol. Biol. Evol. 27: 1478–1494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hyams G., Abadi S., Lahav S., Avni A., Halperin E., Shani E., et al. (2018) CRISPys: optimal sgRNA design for editing multiple members of a gene family using the CRISPR system. J. Mol. Biol. 430: 2184–2195. [DOI] [PubMed] [Google Scholar]
- Jacobs T.B., Zhang N., Patel D. and Martin G.B. (2017) Generation of a collection of mutant tomato lines using pooled CRISPR libraries. Plant Physiol. 174: 2023–2037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jamnadass R., Mumm R.H., Hale I., Hendre P., Muchugi A., Dawson I.K., et al. (2020) Enhancing African orphan crops with genomics. Nat. Genet. 52: 356–360. [DOI] [PubMed] [Google Scholar]
- Jankowicz-Cieslak J., Mba C., Till B.J. (2017) Mutagenesis for crop breeding and functional genomics. InBiotechnologies for Plant Mutation Breeding. Edited by Jankowicz-Cieslak, J., et al. pp. 3–18. International Atomic Energy Agency, Springer, Cham. [Google Scholar]
- Jankowicz-Cieslak J. and Till B.J. (2016) Chemical mutagenesis of seed and vegetatively propagated plants using EMS. Curr. Protoc. Plant Biol. 1: 617–635. [DOI] [PubMed] [Google Scholar]
- Jarvis D.E., Ho Y.S., Lightfoot D.J., Schmöckel S.M., Li B., Borm T.J.A., et al. (2017) The genome of Chenopodium quinoa. Nature 542: 307–312. [DOI] [PubMed] [Google Scholar]
- Jarvis D.I., Brown A.H., Cuong P.H., Collado-Panduro L., Latournerie-Moreno L., Gyawali S., et al. (2008) A global perspective of the richness and evenness of traditional crop-variety diversity maintained by farming communities. Proc. Natl. Acad. Sci. U.S.A. 105: 5326–5331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones G., Kluyver T., Preece C., Swarbrick J., Forster E., Wallace M., et al. (2021) The origins of agriculture: Intentions and consequences. J. Archaeol. Sci. 125: 105290. [Google Scholar]
- Jørgensen I.H. (1992) Discovery, characterization and exploitation of Mlo powdery mildew resistance in barley. Euphytica 63: 141–152. [Google Scholar]
- Jørgensen M.E., Crocoll C., Halkier B.A. and Nour-Eldin H.H. (2017) Uptake assays in Xenopus laevis oocytes using liquid chromatography-mass spectrometry to detect transport activity. Bio. Protoc. 7: e2581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jørgensen M.E., Nour-Eldin H.H. and Halkier B.A. (2015) Transport of defense compounds from source to sink: lessons learned from glucosinolates. Trends Plant Sci. 20: 508–514. [DOI] [PubMed] [Google Scholar]
- Kaiser N., Douches D., Dhingra A., Glenn K.C., Herzig P.R., Stowe E.C., et al. (2020) The role of conventional plant breeding in ensuring safe levels of naturally occurring toxins in food crops. Trends Food Sci. Technol. 100: 51–66. [Google Scholar]
- Keneni G., Bekele E., Imtiaz M. and Dagne K. (2012) Genetic vulnerability of modern crop cultivars: causes, mechanism and remedies. Int. J. Plant Res. 2: 69–79. [Google Scholar]
- Khanday I., Skinner D., Yang B., Mercier R. and Sundaresan V. (2019) A male-expressed rice embryogenic trigger redirected for asexual propagation through seeds. Nature 565: 91–95. [DOI] [PubMed] [Google Scholar]
- Khlestkina E.K., Huang X.Q., Quenum F.J., Chebotar S., Röder M.S. and Börner A. (2004) Genetic diversity in cultivated plants-loss or stability? Theor. Appl. Genet. 108: 1466–1472. [DOI] [PubMed] [Google Scholar]
- Khoury C.K., Brush S., Costich D.E., Curry H.A., de Haan S., Engels J.M.M., et al. (2022) Crop genetic erosion: understanding and responding to loss of crop diversity. New Phytol. 233: 84–118. [DOI] [PubMed] [Google Scholar]
- Kingsbury N. (2009) Hybrid: The History and Science of Plant Breeding. University of Chicago Press, Chicago, IL. [Google Scholar]
- Knudsen S., Lok F., Braumann I. (2020) Developing barley crops for improved brewing quality. InAchieving Sustainable Cultivation of Barley. Edited by Fox, G. and Li, C.. Burleigh Dodds Science, Cambridge. ISBN 978-1-78676-308-2. [Google Scholar]
- Knudsen S., Wendt T., Dockter C., Thomsen H.C., Rasmussen M., Jørgensen M.E., et al. (2021) FIND-IT: Ultrafast mining of genome diversity. BioRxiv.doi: 10.1101/2021.05.20.444969. [DOI] [Google Scholar]
- Kong J., Martin-Ortigosa S., Finer J., Orchard N., Gunadi A., Batts L.A., et al. (2020) Overexpression of the transcription factor GROWTH-REGULATING FACTOR5 improves transformation of dicot and monocot species. Front. Plant Sci. 11: 572319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koziol L., Rieseberg L.H., Kane N. and Bever J.D. (2012) Reduced drought tolerance during domestication and the evolution of weediness results from tolerance-growth trade-offs. Evolution 66: 3803–3814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kozukue N., Kozukue E. and Mizuno S. (1987) Glycoalkaloids in potato plants and tubers. Hortic. Sci. 22: 294–296. [Google Scholar]
- Krasileva K.V., Vasquez-Gross H.A., Howell T., Bailey P., Paraiso F., Clissold L., et al. (2017) Uncovering hidden variation in polyploid wheat. Proc. Natl. Acad. Sci. U.S.A. 114: E913–E921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuppu S., Ron M., Marimuthu M.P.A., Li G., Huddleson A., Siddeek M.H., et al. (2020) A variety of changes, including CRISPR/Cas9-mediated deletions, in CENH3 lead to haploid induction on outcrossing. Plant Biotechnol. J. 18: 2068–2080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kushwah A., Gupta S., Bindra S., Johal N., Singh I., Bharadwaj C. et al. (2020) Gene pyramiding and multiple character breeding. InChickpea: Crop Wild Relatives for Enhancing Genetic Gains. Edited by Singh, M. pp. 131–165. Academic Press, Cambridge, MA. [Google Scholar]
- Lambein F., Travella S., Kuo Y.H., Van Montagu M. and Heijde M. (2019) Grass pea (Lathyrus sativus L.): orphan crop, nutraceutical or just plain food? Planta 250: 821–838. [DOI] [PubMed] [Google Scholar]
- Larson G., Piperno D.R., Allaby R.G., Purugganan M.D., Andersson L., Arroyo-Kalin M., et al. (2014) Current perspectives and the future of domestication studies. Proc. Natl. Acad. Sci. 111: 6139–6146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lev-Yadun S., Gopher A. and Abbo S. (2000) The cradle of agriculture. Science 288: 1602–1603. [DOI] [PubMed] [Google Scholar]
- Li T., Yang X., Yu Y., Si X., Zhai X., Zhang H., et al. (2018) Domestication of wild tomato is accelerated by genome editing. Nat. Biotechnol. 36: 1160–1163. [DOI] [PubMed] [Google Scholar]
- Liebman M. (2018) Polyculture cropping systems. InAgroecology. The science of sustainable agriculture. Edited by Altieri, M.A. pp. 205–218. CRC Press, Boca Raton, FL. [Google Scholar]
- Liu H.J., Jian L., Xu J., Zhang Q., Zhang M., Jin M., et al. (2020) High-throughput CRISPR/Cas9 mutagenesis streamlines trait gene identification in maize. Plant Cell 32: 1397–1413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- López-Marqués R.L., Nørrevang A.F., Ache P., Moog M., Visintainer D., Wendt T., et al. (2020) Prospects for the accelerated improvement of the resilient crop quinoa. J. Exp. Bot. 71: 5333–5347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lowe K., La Rota M., Hoerster G., Hastings C., Wang N., Chamberlin M., et al. (2018) Rapid genotype “independent” Zea mays L. (maize) transformation via direct somatic embryogenesis. Vitro Cell. Dev. Biol. Plant. 54: 240–252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lowe K., Wu E., Wang N., Hoerster G., Hastings C., Cho M.J., et al. (2016) Morphogenic regulators Baby boom and Wuschel improve monocot transformation. Plant Cell 28: 1998–2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu Y., Ye X., Guo R., Huang J., Wang W., Tang J., et al. (2017) Genome-wide targeted mutagenesis in rice using the CRISPR/Cas9 system. Mol. Plant 10: 1242–1245. [DOI] [PubMed] [Google Scholar]
- Luo G. and Palmgren M. (2021) GRF-GIF chimeras boost plant regeneration. Trends Plant Sci. 26: 201–204. [DOI] [PubMed] [Google Scholar]
- Luo M.-C., Yang Z.-L., You F.M., Kawahara R., Waines J.G. and Dvorak J. (2007) The structure of wild and domesticated emmer wheat populations, gene flow between them, and the site of emmer domestication. Theor. Appl. Genet. 114: 947–959. [DOI] [PubMed] [Google Scholar]
- Ma L., Kong F., Sun K., Wang T. and Guo T. (2021) From classical radiation to modern radiation: past, present, and future of radiation mutation breeding. Front. Public Health 9: 768071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackay T.F.C., Stone E.A. and Ayroles J.F. (2009) The genetics of quantitative traits: challenges and prospects. Nat. Rev. Genet. 10: 565–577. [DOI] [PubMed] [Google Scholar]
- MacKey J. (1954) Neutron and X-ray experiments in wheat and a revision of the speltoid problem. Hereditas 40: 65–180. [Google Scholar]
- Maher M.F., Nasti R.A., Vollbrecht M., Starker C.G., Clark M.D. and Voytas D.F. (2020) Plant gene editing through de novo induction of meristems. Nat. Biotechnol. 38: 84–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marthe C., Kumlehn J. and Hensel G. (2015) Barley (Hordeum vulgare L.) transformation using immature embryos. Methods Mol. Biol. 1223: 71–83. [DOI] [PubMed] [Google Scholar]
- Martin-Merino M. (2021) The Neolithic revolution: agriculture, sedentary lifestyle and its consequences. Cambridge Open Engage. 10.33774/coe-2021-2589h. [Google Scholar]
- Matsumoto T., Wu J., Kanamori H., Katayose Y., Fujisawa M., Namiki N., et al. (2005) The map-based sequence of the rice genome. Nature 436: 793–800. [DOI] [PubMed] [Google Scholar]
- Mayer K.F.X., Waugh R., Langridge P., Close T.J., Wise R.P., Graner A., et al. (2012) A physical, genetic and functional sequence assembly of the barley genome. Nature 491: 711–716. [DOI] [PubMed] [Google Scholar]
- Mba C., Guimaraes E.P. and Ghosh K. (2012) Re-orienting crop improvement for the changing climatic conditions of the 21st century. Agric. Food Secur. 1: 7. [Google Scholar]
- McCallum C.M., Comai L., Greene E.A. and Henikoff S. (2000) Targeted screening for induced mutations. Nat. Biotechnol. 18: 455–457. [DOI] [PubMed] [Google Scholar]
- Meng X., Yu H., Zhang Y., Zhuang F., Song X., Gao S., et al. (2017) Construction of a genome-wide mutant library in rice using CRISPR/Cas9. Mol. Plant 10: 1238–1241. [DOI] [PubMed] [Google Scholar]
- Meuwissen T.H.E., Hayes B.J. and Goddard M.E. (2001) Prediction of total genetic value using genome-wide dense marker maps. Genetics 157: 1819–1829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer R.S., DuVal A.E. and Jensen H.R. (2012) Patterns and processes in crop domestication: an historical review and quantitative analysis of 203 global food crops. New Phytol. 196: 29–48. [DOI] [PubMed] [Google Scholar]
- Meyer R.S. and Purugganan M.D. (2013) Evolution of crop species: genetics of domestication and diversification. Nat. Rev. Genet. 14: 840–852. [DOI] [PubMed] [Google Scholar]
- Minkenberg B., Wheatley M. and Yang Y. (2017) CRISPR/Cas9-enabled multiplex genome editing and its application. Prog. Mol. Biol. Transl. Sci. 149: 111–132. [DOI] [PubMed] [Google Scholar]
- Molnar T.J., Kahn P.C., Ford T.M., Funk C.J. and Funk C.R. (2013) Tree crops, a permanent agriculture: concepts from the past for a sustainable future. Resources 2: 457–488. [Google Scholar]
- Monna L., Kitazawa N., Yoshino R., Suzuki J., Masuda H., Maehara Y., et al. (2002) Positional cloning of rice semidwarfing gene, sd-1: rice “Green Revolution Gene” encodes a mutant enzyme involved in gibberellin synthesis. DNA Res. 9: 11–17. [DOI] [PubMed] [Google Scholar]
- Moreira X., Abdala-Roberts L., Gols R. and Francisco M. (2018) Plant domestication decreases both constitutive and induced chemical defenses by direct selection against defensive traits. Sci. Rep. 8: 12678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muckey E. (2019) Kernza® in Southern Minnesota: Assessing local viability of intermediate wheatgrass. Co-publication of the University of Minnesota’s Center for Urban and Regional Affairs (CURA), Southwest Regional Sustainable Development Partnership (RSDP), and Rural Advantage. [Google Scholar]
- Mus F., Crook M.B., Garcia K., Garcia Costas A., Geddes B.A., Kouri E.D., et al. (2016) Symbiotic nitrogen fixation and the challenges to its extension to nonlegumes. Appl. Environ. Microbiol. 82: 3698–3710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neelakandan A.K. and Wang K. (2012) Recent progress in the understanding of tissue culture-induced genome level changes in plants and potential applications. Plant Cell Rep. 31: 597–620. [DOI] [PubMed] [Google Scholar]
- Nishimura A., Aichi I. and Matsuoka M. (2007) A protocol for Agrobacterium-mediated transformation in rice. Nat. Protoc. 1: 2796–2802. [DOI] [PubMed] [Google Scholar]
- Nour-Eldin H.H., Madsen S.R., Engelen S., Jørgensen M.E., Olsen C.E., Andersen J.S., et al. (2017) Reduction of antinutritional glucosinolates in Brassica oilseeds by mutation of genes encoding transporters. Nat. Biotechnol. 35: 377–382. [DOI] [PubMed] [Google Scholar]
- Oladosu Y., Rafii M.Y., Abdullah N., Hussin G., Ramli A., Rahim H.A., et al. (2015) Principle and application of plant mutagenesis in crop improvement: a review. Biotechnol. Biotechnol. Equip. 30: 1–16. [Google Scholar]
- Orman-Ligeza B., Harwood W., Hedley P.E., Hinchcliffe A., Macaulay M., Uauy C., et al. (2020) TRA1: A locus responsible for controlling Agrobacterium-mediated transformability in barley. Front. Plant Sci. 11: 355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Østerberg J.T., Xiang W., Olsen L.I., Edenbrandt A.K., Vedel S.E., Christiansen A., et al. (2017) Accelerating the domestication of new crops: feasibility and approaches. Trends Plant Sci. 22: 373–384. [DOI] [PubMed] [Google Scholar]
- Palmgren M.G., Edenbrandt A.K., Vedel S.E., Andersen M.M., Landes X., Østerberg J.T., et al. (2015) Are we ready for back-to-nature crop breeding? Trends Plant Sci. 20: 155–164. [DOI] [PubMed] [Google Scholar]
- Pourkheirandish M., Hensel G., Kilian B., Senthil N., Chen G., Sameri M., et al. (2015) Evolution of the grain dispersal system in barley. Cell 162: 527–539. [DOI] [PubMed] [Google Scholar]
- Price T.D. and Bar-Yosef O. (2011) The origins of agriculture: new data, new ideas: an introduction to supplement 4. Curr. Anthropol. 52: S163–S174. [Google Scholar]
- Rakszegi M., Kisgyörgy B.N., Tearall K., Shewry P.R., Láng L., Phillips A., et al. (2010) Diversity of agronomic and morphological traits in a mutant population of bread wheat studied in the Healthgrain program. Euphytica 174: 409–421. [Google Scholar]
- Raman R. (2017) The impact of Genetically Modified (GM) crops in modern agriculture: a review. GM Crops Food 8: 195–208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ray D.K., Ramankutty N., Mueller N.D., West P.C. and Foley J.A. (2012) Recent patterns of crop yield growth and stagnation. Nat. Commun. 3: 1293. [DOI] [PubMed] [Google Scholar]
- Razzaq A., Saleem F., Wani S.H., Abdelmohsen S.A.M., Alyousef H.A., Abdelbacki A.M.M., et al. (2021) De-novo domestication for improving salt tolerance in crops. Front Plant Sci. 12: 681367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Renard D. and Tilman D. (2019) National food production stabilized by crop diversity. Nature 571: 257–260. [DOI] [PubMed] [Google Scholar]
- Roberts H.F. (1919) The founders of the art of breeding. J. Heredity 10: 99–106. [Google Scholar]
- Rodríguez-Leal D., Lemmon Z.H., Man J., Bartlett M.E. and Lippman Z.B. (2017) Engineering quantitative trait variation for crop improvement by genome editing. Cell 2: 470–480.e8. [DOI] [PubMed] [Google Scholar]
- Roy S., Liu W., Nandety R.S., Crook A., Mysore K.S., Pislariu C.I., et al. (2020) Celebrating 20 years of genetic discoveries in legume nodulation and symbiotic nitrogen fixation. Plant Cell 32: 15–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rull V. (2010) Food security: green revolution drawbacks. Science 328: 169. [DOI] [PubMed] [Google Scholar]
- Sahoo K.K., Tripathi A.K., Pareek A., Sopory S.K. and Singla-Pareek S.L. (2011) An improved protocol for efficient transformation and regeneration of diverse indica rice cultivars. Plant Methods 7: 1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sailer C., Schmid B. and Grossniklaus U. (2016) Apomixis allows the transgenerational fixation of phenotypes in hybrid plants. Curr. Biol. 26: 331–337. [DOI] [PubMed] [Google Scholar]
- Sánchez-León S., Gil-Humanes J., Ozuna C.V., Giménez M.J., Sousa C., Voytas D.F., et al. (2018) Low-gluten, nontransgenic wheat engineered with CRISPR/Cas9. Plant Biotechnol. J. 16: 902–910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schlegel R.H.J. (2018) History of Plant Breeding. CRC Press, Boca Raton, FL. [Google Scholar]
- Sharma S., Schulthess A.W., Bassi F.M., Badeaeva E.D., Neumann K., Graner A., et al. (2021) Introducing beneficial alleles from plant genetic resources into the wheat germplasm. Biology 10: 982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaw L.M., Turner A.S., Henry L., Griffiths S. and Laurie D.A. (2013) Mutant alleles of Photoperiod-1 in wheat (Triticum aestivum L.) that confer a later flowering phenotype in long days. PLoS One 8: e79459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shu Q.Y., Forster B.P. and Nakagawa H. (2012) Plant Mutation Breeding and Biotechnology. p. 608. CABI, Wallingford, UK. [Google Scholar]
- Sleper D.A. and Poehlman J.M. (2006) Breeding Field Crops. p. 424. Blackwell Publishing, Oxford. [Google Scholar]
- Slinkard A.E., Vanderberg A. and Holm F.A. (2007) Lentil plants having increased resistance to imidazolinone herbicides. U.S. Patent, 2007. http://www.uspto.gov/patft/index.html (March 6, 2022 date last accessed). [Google Scholar]
- Spielmeyer W., Ellis M.H. and Chandler P.M. (2002) Semidwarf (sd-1), “green revolution” rice, contains a defective gibberellin 20-oxidase gene. Proc. Natl. Acad. Sci. U.S.A. 99: 9043–9048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Symko S. (1999) From a Single Seed. Tracing the Marquis Wheat Success Story in Canada to Its Roots in the Ukraine. Agriculture and Agri-Food Canada. Stategic Promotion, Research Branch, Agriculture and Agri-Food Canada, Ottawa, Canada. [Google Scholar]
- Talamè V., Bovina R., Sanguineti M.C., Tuberosa R., Lundqvist U. and Salvi S. (2008) TILLMore, a resource for the discovery of chemically induced mutants in barley. Plant Biotechnol. J. 6: 477–485. [DOI] [PubMed] [Google Scholar]
- Tester M. and Langridge P. (2010) Breeding technologies to increase crop production in a changing world. Science 327: 818–822. [DOI] [PubMed] [Google Scholar]
- Thanh P.T., Vladutu C.I., Kianian S.F., Thanh P.T., Ishii T., Nitta M., et al. (2014) Molecular genetic analysis of domestication traits in emmer wheat. I: map construction and QTL analysis using an F2 Population. Biotechnol. Biotechnol. Equip. 27: 3627–3637. [Google Scholar]
- The Government Office for Science. (2011) Foresight: the future of food and farming. Final project report. The Government Office for Science, London. [Google Scholar]
- Till B.J., Cooper J., Tai T.H., Colowit P., Greene E.A., Henikoff S., et al. (2007) Discovery of chemically induced mutations in rice by TILLING. BMC Plant Biol. 7: 19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tilman D., Balzer, C., Hill, J., and Befort, B.L. (2011) Global food demand and the sustainable intensification of agriculture. Proc Natl Acad Sci U.S.A. 108: 20260–20264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toensmeier E. (2016) The Carbon Farming Solution: A Global Toolkit of Perennial Crops and Regenerative Agriculture Practices for Climate Change Mitigation and Food Security. p. 512. Chelsea Green Publishing, White River Junction, VT. [Google Scholar]
- Tsegaye B. and Berg T. (2007) Genetic erosion of Ethiopian tetraploid wheat landraces in Eastern Shewa, Central Ethiopia. Genet. Res. Crop Evol. 54: 715–726. [Google Scholar]
- Uauy C., Paraiso F., Colasuonno P., Tran R.K., Tsai H., Berardi S., et al. (2009) A modified TILLING approach to detect induced mutations in tetraploid and hexaploid wheat. BMC Plant Biol. 9: 1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Udall J.A. (2003) Breeding for quantitative traits in plants. Crop Sci. 43: 1578–1580. [Google Scholar]
- van de Wouw M., Kik C., van Hintum T., van Treuren R. and Visser B. (2010) Genetic erosion in crops: concept, research results and challenges. Plant Genet. Res. 8: 1–15. [Google Scholar]
- Vogel K.E. (2009) Backcross-breeding. Methods Mol. Biol. 526: 161–169. [DOI] [PubMed] [Google Scholar]
- Voytas D.F. (2013) Plant genome engineering with sequence-specific nucleases. Annu. Rev. Plant Biol. 64: 327–350. [DOI] [PubMed] [Google Scholar]
- Wang C., Liu Q., Shen Y., Hua Y., Wang J., Lin J., et al. (2019) Clonal seeds from hybrid rice by simultaneous genome engineering of meiosis and fertilization genes. Nat. Biotechnol. 37: 283–286. [DOI] [PubMed] [Google Scholar]
- Wang K., Shi L., Liang X., Zhao P., Wang W., Liu J., et al. (2022) The gene TaWOX5 overcomes genotype dependency in wheat genetic transformation. Nat. Plants 8: 110–117.doi: 10.1038/s41477-021-01085-8. [DOI] [PubMed] [Google Scholar]
- Wang Y., Cheng X., Shan Q., Zhang Y., Liu J., Gao C., et al. (2014) Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat. Biotechnol. 32: 947–951. [DOI] [PubMed] [Google Scholar]
- Wani S.H., Dutta T., Neelapu N.R.R. and Surekha C. (2017) Transgenic approaches to enhance salt and drought tolerance in plants. Plant Gene 11: 219–231. [Google Scholar]
- Wendt T., Holme I., Dockter C., Preu A., Thomas W., Druka A., et al. (2016) HvDep1 Is a positive regulator of culm elongation and grain size in barley and impacts yield in an environment-dependent manner. PLoS One 11: e0168924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Westerbergh A., Lerceteau-Köhler E., Sameri M., Bedada G. and Lundquist P.-O. (2018) Towards the development of perennial barley for cold temperate climates—evaluation of wild barley relatives as genetic resources. Sustainability 10: 1969. [Google Scholar]
- Willmann M.R. (2019) Seeds without sex: clonal seed production in rice. CRISPR J. 2: 11–14. [DOI] [PubMed] [Google Scholar]
- Wingen L.U., West C., Leverington-Waite M., Collier S., Orford S., Goram R., et al. (2017) Wheat landrace genome diversity. Genetics 205: 1657–1676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolter F., Schindele P. and Puchta H. (2019) Plant breeding at the speed of light: the power of CRISPR/Cas to generate directed genetic diversity at multiple sites. BMC Plant Biol. 19: 176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu H., Lin T., Meng X., Du H., Zhang J., Liu G., et al. (2021) A route to de novo domestication of wild allotetraploid rice. Cell 184: 1156–1170.e14. [DOI] [PubMed] [Google Scholar]
- Zhang Y., Pribil M., Palmgren M. and Gao C. (2020) A CRISPR way for accelerating improvement of food crops. Nat. Food 1: 200–205. [Google Scholar]
- Zhong Y., Liu C., Qi X., Jiao Y., Wang D., Wang Y., et al. (2019) Mutation of ZmDMP enhances haploid induction in maize. Nat. Plants. 5: 575–580. [DOI] [PubMed] [Google Scholar]
- Zsögön A., Čermák T., Naves E.R., Notini M.M., Edel K.H., Weinl S., et al. (2018) De novo domestication of wild tomato using genome editing. Nat. Biotechnol. 36: 1211–1216. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
No new datasets were generated or analyzed in this study.