

Abstract
Marine microalgae, diatoms, are considered a source of a wide range of high-value compounds, and numerous studies indicate their biotechnological potential in the food and feed industry, cosmetic industry, nanotechnology, pharmaceutical industry, biodiesel production, fertilizers, and wastewater treatment. The aim of this study was to compare the growth, chemical profiles, and antioxidant activity of the diatom Skeletonema grevillei cultivated in a bioreactor and an incubation-shaking cabinet at different growth phases (after 192 and 312 h). Growth was monitored by evaluating cell density with the Sedgewick Rafter chamber, and the collected biomass was extracted with 70% ethanol assisted by ultrasound. Extracts were evaporated to dryness and compounds were identified in derivatized form by gas chromatography and mass spectrometry (GC-MS) analysis, while antioxidant capacity was evaluated by DPPH and ORAC. Significantly faster growth was observed in the bioreactor than in the incubation-shaking cabinet. Oleamide, palmitelaidic acid, glycerol monostearate, myristic acid, cholesterol, eicosapentaenoic acid, 1-monopalmitin, and 24-methylene cholesterol were identified as the major compounds in both systems. Among them, oleamide was the dominant compound in both systems. It is also shown that prolonging the cultivation period had a direct effect on increasing the extract yield. The highest DPPH inhibition (11.4 ± 1%) and ORAC values (93.3 ± 8.4 mM TE) were obtained for the S. grevillei extract recovered from the bioreactor after 312 h. The obtained results contribute to the possibility of using S. grevillei for various biotechnological applications in the future.
Keywords: diatom, bioreactor, incubation-shaking cabinet, derivatized compounds, antioxidant activity
1. Introduction
Market demand is changing rapidly, directing research toward finding new ways to use marine resources as a renewable and sustainable source of compounds that can be used in a variety of industries. Among the great biodiversity found in the oceans and seas, marine microalgae are recognized as microorganisms with exceptional biotechnological potential, especially in industries directed to production of pharmaceuticals, functional food ingredients, and food products [1]. Although the application of microalgae is mainly related to the food industry, many species have potential for wastewater treatment, biodiesel production, and nutraceuticals [2]. Despite their great potential, the wider application of a larger number of species is still not economically viable due to numerous challenges, mainly related to technological barriers.
Diatoms are photosynthesizing microalgae with cell walls of transparent opaline silica containing compounds such as pigments, sterols, and fatty acids [3]. Currently, the most commercially important diatom species is Dunaliella salina. As the biotechnological potential of diatoms is constantly being discovered, the number of studies on different species is increasing. Diatoms have shown promising features that make them ideal candidates for this form of production, as their intracellular content can be used for the production of biodiesel and valuable food components, and the silicate shell can be used as a material for nanotechnology [4]. In terms of commercial use, the genus Skeletonema is an unconventional group of microalgae, but since it is non-toxic, it is suitable for use in the food industry [5]. In fact, the Food and Drug Administration considers Skeletonema sp. as generally recognized as safe (GRAS) [6]. Some species such as S. marinoii have been described in the literature [7,8]; however, there are no reports on S. grevillei. Nevertheless, the cultivation of microalgae for wider use slowed down due to technological obstacles, mainly related to the optimization of growth, harvesting, and extraction efficiency of targeted compounds [9]. In addition to the removing of technological barriers, cultivation should be directed toward the utilization of all production components (bio-based refinery), which would increase the economic profitability and environmental sustainability of the commercial use of diatoms.
Diatoms are among the most flexible and productive eukaryotic microalgae characterized by exceptional adaptability to changing environmental conditions (e.g., temperature and light). Mechanisms such as extracellular polysaccharide layers, cell wall thickness, and morphologically distinct resting stages minimize the potential damage caused by stress conditions [10,11,12]. On the other hand, their abundance in cultivation under laboratory conditions is much lower than in the natural environment. Therefore, optimization of cultivation and selection of suitable initiators for higher production of metabolites is of utmost importance. Increased production of primary and secondary metabolites is associated with stressful cultivation conditions (e.g., nutrient deficiency, non-optimal temperature, different light sources), while chemical characterization of the intracellular content (primary and secondary metabolites) is quite complex. Promoting metabolite production through nutrient deprivation and introducing other stress growth conditions often result in lower biomass yield, which is a common problem in microalgae production. The main challenges are obtaining a sufficient amount of biomass for biotechnological tests, problems related to the effects of nutrients and environmental changes on biosynthesis, and impurities [4]. Conventional cultivation in incubators typically produces little biomass, and the cost of the production effort exceeds the biomass yield and profitability. In 1997, Fukami et al. [13] developed the first bioreactor to promote biomass production of the diatom Nitzschia sp. In general, cultivation in bioreactors achieves greater homogeneity so that all cells have the same availability of key factors for successful growth, such as nutrients and light. While the production of higher amounts of primary metabolites is associated with optimal growth conditions, the accumulation of secondary metabolites is usually associated with stressful conditions [14]. Microalgae are known to produce high-value bioactive compounds such as polyunsaturated fatty acids (PUFA), carotenoids, phenolic compounds, sterols, and sulfated polysaccharides with many health and therapeutic benefits [6,15]. Nevertheless, there is little information on the cultivation potential and chemical profile of the genus Skeletonema. Some species such as S. marinoii, S. dohrnii and S. costatum have been previously studied [7,16,17,18]; however, there are no reports on S. gravileii. In a nutrient-deprived environment, Skeletonema species are able to increase lipid production [10], and even produce higher amounts of omega-3 eicosapentaenoic acid (EPA) [19]. Under light and temperature stress, these microorganisms produce components that exhibit antioxidant activity, while the selection of a suitable cultivation system can significantly increase biomass yields, i.e., extract yields [20,21]. In addition, metabolic processes in diatoms are extremely fast, and the production of various molecules may also depend on the growth phases [8].
Biotechnological improvements are possible by selecting suitable microalgal strains capable of producing large amounts of various bioactive compounds under optimal conditions. Such a form of microalgae cultivation would make commercialization faster and more profitable. Therefore, the aim of this study was to determine and compare the growth of S. grevillei in a standard F/2 medium using two cultivation systems (bioreactor and incubation-shaking cabinet). In addition, ethanolic extracts were prepared for each system at two growth stages and their chemical profiles (dominant compounds) and antioxidant potential were determined.
2. Results and Discussion
2.1. Comparison of Growth Curves in BRC and EIS
Considering that extending the duration of the cultivation period beyond the stationary phase would result in the decay of diatom cells, the end of cultivation in this study was defined as the time at which one of the systems reached maximum growth. This was considered the beginning of the stationary phase and in the bioreactor system (BRC) it was reached after 312 h.
The results of growth in a BRC and the incubation-shaking cabinet (EIS) system during the cultivation period are shown in Figure 1. First, the growth rate in each system over time was examined. After 144 h of cultivation, a statistically significant (p < 0.05) change (onset of exponential growth) was observed in the EIS system, while in the BRC system this was observed one day earlier. During the first 144 h, there was no statistically significant difference (p > 0.05) in cell number between the EIS and BRC systems. A statistically significant (p < 0.05) difference between the systems was first observed after 168 h and persisted until the end of cultivation. The total mass of dry extracts collected in the exponential and at the beginning of the stationary growth phase is shown in Figure 2. The weight of the dry extract was higher in the BRC system at both collection times. The results indicate that the higher homogeneity in the BRC system provided by the impeller and airflow was crucial for the uniform availability of light and nutrients, which consequently led to faster growth of diatoms in this system.
Figure 1.

Growth curves for Skeletonema grevillei in a bioreactor and incubation-shaking cabinet during cultivation period of 312 h.
Figure 2.

The average mass of dry Skeletonema grevillei extracts from bioreactor and incubation-shaking cabinet obtained from total diatom biomass after cultivation periods of 192 and 312 h.
Since the early days of diatom cultivation in bioreactors, it has been known that cell homogeneity allows maximum utilization of limiting nutrients [13]. Although the lack of nutrients often stimulates the metabolism to produce secondary metabolites, these conditions cause disturbances in the growth of diatom cultures. Deficiencies in nutrients such as N and Si negatively affect growth, motility, and often cell morphology, but they are found to be important in stimulating diatom metabolism to produce more valuable metabolites [22,23,24,25]. Ramirez et al. [26] compared the growth of two diatoms, N. epithemioides and Nitzschia sp. at different temperatures, light intensities and silicate levels. The addition of silicate was found to be more suitable to stimulate growth than modification of light and temperature. Raniello et al. [20] recorded greater biomass production of the diatom Cocconeis neothumensis under different light and nutrient conditions in a bioreactor than in a batch culture in Petri dishes. Significantly higher extract yield (24 ± 5 mg of dry extract) was obtained in the bioreactor compared to batch cultures (17 ± 3 mg of dry extract), which is comparable to the results of extract yield between BRC and EIS systems in this study [20].
2.2. Identification of Compounds by GC-MS
The compounds identified in a BRC, and EIS are listed in Table 1 and Table 2 along with their retention index, percentage of similarity, proportion, and molecular weight. The retention times and MS m/z of the compounds is given in the Supplementary Material (Table S1). The chromatograms obtained are shown in Figure 3. Since the method is not quantitative, the proportions of the compounds refer only to the injected sample. The amount of each compound in the injected sample volume is expressed as a percentage calculated based on the areas obtained. In Table 1 and Table 2, for the compounds identified in derivatized or non-derivatized form, only those with ≥85% similarity spectra and matching RI were considered. In addition, all compounds with similarity less than 90% were confirmed in non-derivatized form. All listed compounds were confirmed according to previous studies on the chemical profiling of disatoms [7,27,28].
Table 1.
List of tentatively identified compounds from Skeletonema grevillei extracts by GC-MS in bioreactor after 192 and 312 h.
| No. | Retention Index | Similarty (%) | Proportion (%) | Identified Compound | Molar Weight | |||
|---|---|---|---|---|---|---|---|---|
| BRC192 | BRC312 | BRC192 | BRC312 | BRC192 | BRC312 | |||
| 1 | 1242 | n.d. | 96 | n.d. | 0.05 | n.d. | Benzoic acid, TMS derivative | 194 |
| 2 | 1253 | n.d. | 86 | n.d. | 0.02 | n.d. | Octanoic acid, TMS derivative | 216 |
| 3 | 1269 | 1265 | 94 | 92 | 0.13 | 0.21 | Glycerol, 3TMS derivative | 308 |
| 4 | 1348 | 1345 | 95 | 92 | 0.08 | 0.05 | Nonanoic acid, TMS derivative | 230 |
| 5 | 1451 | 1450 | 95 | 91 | 0.02 | 0.02 | Decanoic acid, TMS derivative | 244 |
| 6 | n.d. | 1499 | n.d. | 94 | n.d. | 0.13 | Malic acid, 3TMS derivative | 350 |
| 7 | 1534 | n.d. | 91 | n.d. | 0.01 | n.d. | 1-Dodecanethiol | 202 |
| 8 | 1653 | 1499 | 97 | 93 | 0.05 | 0.03 | Dodecanoic acid, TMS derivative | 272 |
| 9 | 1691 | n.d. | 85 | n.d. | 0.03 | n.d. | Tetradecanenitrile | 326 |
| 10 | 1753 | 1752 | 94 | 94 | 0.10 | 0.14 | Tridecanoic acid, TMS derivative | 286 |
| 11 | 1767 | n.d. | 91 | n.d. | 0.01 | n.d. | 1-Tetradecanol, TMS derivative | 286 |
| 12 | 1788 | 1787 | 93 | 93 | 0.06 | 0.17 | Loliolide, TMS | 268 |
| 13 | 1853 | 1852 | 95 | 96 | 2.55 | 6.50 | Myristic acid, TMS derivative | 300 |
| 14 | n.d. | 1877 | n.d. | 90 | n.d. | 0.28 | (Z)-3-Hexenyl β -glucopyranoside, 4TMS derivative | 550 |
| 15 | 1879 | n.d. | 94 | n.d. | 0.11 | n.d. | Palmitoleonitrile | 235 |
| 16 | 1930 | n.d. | 86 | n.d. | 0.05 | n.d. | (E)-13-Methyltetradec-9-enoic acid, TMS derivative | 312 |
| 17 | 1952 | 1951 | 94 | 85 | 0.17 | 0.49 | Pentadecanoic acid, TMS derivative | 314 |
| 18 | 2009 | 2008 | 89 | 88 | 0.97 | 0.63 | Eicosapentaenoic acid, TMS derivative | 374 |
| 19 | 2031 | 2030 | 96 | 97 | 7.37 | 12.15 | Palmitelaidic acid, TMS | 326 |
| 20 | 2053 | 2052 | 96 | 95 | 2.31 | 2.73 | Palmitic acid, TMS derivative | 328 |
| 21 | 2076 | n.d. | 89 | n.d. | 0.43 | n.d. | 9-Octadecynenitrile | 261 |
| 22 | 2083 | 2082 | 97 | 97 | 2.96 | 0.38 | (Z)-9-Octadecenenitrile | 263 |
| 23 | 2151 | 2094 | 92 | 83 | 0.04 | 0.03 | Heptadecanoic acid, TMS derivative | 342 |
| 24 | n.d. | 2157 | n.d. | 93 | n.d. | 0.04 | Palmitoleamide | 253 |
| 25 | n.d. | 2163 | n.d. | 87 | n.d. | 0.01 | 1-Octadecanol, TMS derivative | 342 |
| 26 | n.d. | 2178 | n.d. | 85 | n.d. | 0.03 | Octadecanamide | 319 |
| 27 | 2184 | 2184 | 92 | 97 | 1.48 | 2.09 | Phytol, TMS derivative | 368 |
| 28 | 2216 | 2215 | 88 | 92 | 0.47 | 0.44 | Linoleic acid, TMS derivate | 352 |
| 29 | 2230 | 2229 | 94 | 94 | 2.36 | 0.90 | (Z)-Oleic acid, TMS derivative | 354 |
| 30 | 2251 | 2250 | 97 | 95 | 0.88 | 0.65 | Stearic acid, TMS derivative | 356 |
| 31 | n.d. | 2256 | n.d. | 86 | n.d. | 0.05 | 11-Methyloctadec-12-enoic acid, TMS derivative | 368 |
| 32 | n.d. | 2331 | n.d. | 90 | n.d. | 0.16 | (all-Z)-5,8,11,14,17-Eicosapentaenoic acid, methyl ester | 330 |
| 33 | 2337 | n.d. | 92 | n.d. | 0.10 | n.d. | (Z)-10-Nonadecenoic acid, TMS | 368 |
| 34 | 2365 | 2362 | 95 | 96 | 2.30 | 2.50 | 9-Octadecenamide | 281 |
| 35 | n.d. | 2373 | n.d. | 90 | n.d. | 0.68 | (Z)-5,8,11-Eicosatrienoic acid, TMS derivative | 378 |
| 36 | 2381 | 2380 | 94 | 96 | 3.29 | 6.00 | Eicosapentaenoic acid, TMS derivative | 374 |
| 37 | 2430 | 2422 | 83 | 82 | 49.84 | 33.65 | Oleamide, TMS derivative | 353 |
| 38 | 2450 | 2448 | 95 | 95 | 1.24 | 0.70 | Steramide, TMS derivative | 355 |
| 39 | 2564 | 2564 | 97 | 89 | 0.54 | 0.19 | Doconexent, TMS derivative | 400 |
| 40 | 2578 | 2578 | 93 | 87 | 1.39 | 1.09 | 2-Palmitoylglycerol, 2TMS derivative | 474 |
| 41 | 2610 | 2610 | 96 | 96 | 2.91 | 2.89 | 1-Monopalmitin, 2TMS derivative | 474 |
| 42 | 2771 | 2770 | 90 | 90 | 1.46 | 1.59 | 2-Monostearin, 2TMS derivative | 502 |
| 43 | 2787 | 2787 | 93 | 90 | 0.34 | 0.10 | 1-Monooleoylglycerol, 2TMS derivative | 500 |
| 44 | 2805 | 2804 | 97 | 96 | 5.92 | 9.30 | Glycerol monostearate, 2TMS derivative | 502 |
| 45 | 2826 | 2825 | 88 | 81 | 0.57 | 0.10 | (Z)-Docos-13-enamide, TMS | 409 |
| 46 | n.d. | 2997 | n.d. | 89 | n.d. | 0.15 | Eicosanoic acid, 2,3-bis-(OTMS) propyl ester | 530 |
| 47 | 2998 | n.d. | 93 | n.d. | 0.09 | n.d. | 2,3-Dihydroxypropyl icosanoate, 2TMS derivative | 530 |
| 48 | 3066 | 3066 | 89 | 88 | 0.09 | 0.10 | 2-Arachidonoylglycerol, 2TMS derivative | 522 |
| 49 | 3163 | 3162 | 96 | 96 | 3.97 | 6.21 | Cholesterol, TMS derivative | 458 |
| 50 | n.d. | 3201 | n.d. | 90 | n.d. | 0.40 | Desmosterol, TMS derivative | 456 |
| 51 | 3261 | 3260 | 90 * | 88 * | 3.03 | 5.09 | 24-Methylene cholesterol * | 398 * |
| 52 | 3380 | 3380 | 88 | 81 | 0.15 | 0.20 | Isofucosterol, TMS | 484 |
| 53 | 3672 | 3672 | 90 | 92 | 0.06 | 0.80 | Oleanolic acid 2TMS | 600 |
* A compound identified in non-derivatized form and its molecular weight (other data for this compound, RI, and proportion, were obtained for the derivatized form) as well as with literature data comparison for its derivatized form.; n.d.—not detected. BRC192—after 192 h in a bioreactor; BRC312—after 312 h in a bioreactor.
Table 2.
List of tentatively identified compounds by GC-MS in incubation-shaking cabinet after 192 and 312 h.
| No. | Retention Index | Similarity (%) | Proportion(%) | Identified Compound | Molar Weight | |||
|---|---|---|---|---|---|---|---|---|
| EIS192 | EIS312 | EIS192 | EIS312 | EIS192 | EIS312 | |||
| 1 | 1335 | n.d. | 95 | n.d. | 0.10 | n.d. | Nonanoic acid, TMS derivative | 230 |
| 2 | 1444 | 1379 | 94 | 92 | 0.03 | 0.03 | Decanoic acid, TMS derivative | 244 |
| 3 | n.d. | 1448 | n.d. | 75 | n.d. | 0.01 | Butanedioic acid, TMS derivate | 350 |
| 4 | 1650 | 1630 | 93 | 95 | 0.04 | 0.05 | Dodecanoic acid, TMS derivative | 272 |
| 5 | 1691 | n.d. | 93 | n.d. | 0.02 | n.d. | Tetradecanenitrile | 209 |
| 6 | 1751 | 1739 | 88 | 93 | 0.03 | 0.13 | Tridecanoic acid, TMS derivative | 286 |
| 7 | 1787 | 1773 | 86 | 92 | 0.01 | 0.22 | Loliolide, TMS | 268 |
| 8 | 1805 | 1796 | 88 | 80 | 0.05 | 0.02 | Azelaic acid, 2TMS derivative | 332 |
| 9 | n.d. | 1831 | n.d. | 86 | n.d. | 0.06 | Myristoleic acid, trimethylsilyl ester | 298 |
| 10 | 1852 | 1845 | 95 | 96 | 1.59 | 5.12 | Myristic acid, TMS derivative | 300 |
| 11 | 1877 | n.d. | 94 | n.d. | 0.08 | n.d. | Palmitoleonitrile | 235 |
| 12 | 1898 | n.d. | 96 | n.d. | 0.19 | n.d. | Heptadecanenitrile | 251 |
| 13 | n.d. | 1956 | n.d. | 82 | n.d. | 0.03 | Hexadecanenitrile | 251 |
| 14 | 1951 | 1947 | 93 | 94 | 0.10 | 0.23 | Pentadecanoic acid, TMS derivative | 314 |
| 15 | n.d. | 1960 | n.d. | 93 | n.d. | 0.05 | 1-Hexadecanol, TMS derivative | 314 |
| 16 | 1968 | 1964 | 96 | 95 | 0.66 | 0.25 | Tetradecanamide | 227 |
| 17 | 2031 | 2026 | 88 | 96 | 1.63 | 7.89 | Palmitelaidic acid, TMS | 326 |
| 18 | 2052 | 2049 | 96 | 96 | 1.65 | 2.07 | Palmitic acid, TMS derivative | 328 |
| 19 | 2076 | n.d. | 89 | n.d. | 0.43 | n.d. | 9-Octadecynenitrile | 261 |
| 20 | 2082 | 2078 | 97 | 97 | 6.10 | 0.37 | (Z)-9-Octadecenenitrile | 263 |
| 21 | 2151 | 2149 | 91 | 80 | 0.04 | 0.02 | Heptadecanoic acid, trimethylsilyl ester | 342 |
| 22 | 2158 | 2155 | 97 | 96 | 0.97 | 0.36 | Palmitoleamide | 253 |
| 23 | n.d. | 2161 | n.d. | 86 | n.d. | 0.05 | 1-Octadecanol, TMS derivative | 342 |
| 24 | 2178 | 2176 | 98 | 97 | 1.93 | 0.52 | Hexadecanamide | 255 |
| 25 | 2184 | 2183 | 93 | 97 | 0.20 | 2.06 | Phytol, TMS derivative | 368 |
| 26 | n.d. | 2213 | n.d. | 88 | n.d. | 0.24 | Linoleic, TMS derivative | 352 |
| 27 | 2251 | 2250 | 95 | 95 | 0.75 | 0.53 | Stearic acid, TMS derivative | 356 |
| 28 | n.d. | 2364 | n.d. | 96 | n.d. | 13,97 | 9-Octadecenamide | 281 |
| 29 | n.d. | 2380 | n.d. | 96 | n.d. | 3.78 | Eicosapentaenoic acid, TMS derivative | 374 |
| 30 | 2427 | 2422 | 84 | 82 | 73.31 | 35,53 | Oleamide, TMS derivative | 353 |
| 31 | 2449 | 2419 | 95 | 95 | 1.37 | 0.75 | Octadecanamide, N-TMS derivate | 355 |
| 32 | 2571 | n.d. | 93 | n.d. | 0.81 | n.d. | 13-Docosenamide, (Z)- | 337 |
| 33 | 2578 | 2577 | 93 | 94 | 0.32 | 1.88 | 2-Palmitoylglycerol, 2TMS derivative | 474 |
| 34 | 2610 | 2610 | 94 | 96 | 0.47 | 6.92 | 1-Monopalmitin, 2TMS derivative | 474 |
| 35 | 2771 | 2771 | 89 | 91 | 1.28 | 2.20 | 2-Monostearin, 2TMS derivative | 502 |
| 36 | 2804 | 2804 | 95 | 96 | 3.52 | 10,64 | Glycerol monostearate, 2TMS derivative | 502 |
| 37 | 2998 | 2997 | 87 | 80 | 0.04 | 0.10 | 2,3-Dihydroxypropyl icosanoate, 2TMS derivative | 530 |
| 38 | 3162 | 3160 | 96 | 96 | 1.21 | 1.97 | Cholesterol, TMS derivative | 458 |
| 39 | 3260 | 3529 | 88 * | 89 * | 1.05 | 1.59 | 24-Methylene cholesterol * | 398 * |
| 40 | n.d. | 3379 | n.d. | 88 | n.d. | 0.13 | Isofucosterol, O-TMS | 484 |
| 41 | n.d. | 3672 | n.d. | 90 | n.d. | 0.22 | Oleanolic acid 2TMS | 600 |
* A compound identified in non-derivatized form and its molecular weight (other data for this compound, RI, and proportion, were obtained for the derivatized form) as well as with literature data comparison for its derivatized form.; n.d.—not detected. EIS192—after 192 h in an incubation-shaking cabinet; EIS312—after 312 h in an incubation-shaking cabinet.
Figure 3.

The chromatograms of derivatized dominant compounds in Skeletonema grevillei extracts tentatively identified by GC-MS in: (A,C) bioreactor, (B,D) incubation-shaking cabinet (1—myristic acid, 2—palmitelaidic acid, 3—eicosapentaeonic acid, 4—oleamide or 9- octadecenamide, 5—1-monopalmitin, 6—glycerol monostearate, 7—cholesterol, 8—24-methylene cholesterol).
The difference in the number of compounds identified in the BRC system after 192 and 312 h of cultivation was 44 and 42 compounds, respectively. Compared to BRC, a lower number of compounds were identified in both phases of the EIS system, 33 after 192 h and 34 after 312 h of cultivation. Oleamide (also known as 9-octadecenamide or (Z)-9-octadecenamide) was dominant in both cultivation systems and at both phases. After 192 h, the percentage of oleamide in BRC and EIS was 49.84% and 73.31%, respectively. In both systems, there was a significant decrease with the extension of the cultivation period, by 16.19% in BRC and 37.78% in EIS.
Compounds with a proportion higher than 5% were palmitelaidic acid, glycerol monostearate, cholesterol, 24-methylenecholesterol, myristic acid, and EPA acid in BRC and glycerol monostearate, palmitelaidic acid, myristic acid, and 1-monopalmitin in EIS. The second most abundant compound in BRC was palmitelaidic acid, while in EIS it was glycerol monostearate. Between the two samplings, the proportion of palmitelaidic acid increased from 7.37% to 12.15% in BRC and from 1.63% to 7.89% in EIS. On the other hand, the proportion of glycerol monostearate increased 3-fold in EIS (from 3.52% to 10.64%) while the increase was lower in BRC at the end of the cultivation period (from 5.92% to 9.30%). To our knowledge, this is the first time it has been identified in marine diatoms. In both systems, myristic acid increased by 3.95% in EIS and 3.53% in BRC, with an additional culture period of 120 h. After 312 h of cultivation, EPA was identified in both systems but the proportion in BRC was higher than in EIS, 6.40% and 3.78%, respectively. In addition, after 192 h, EPA was not detected in the EIS, while in BRC, it was recorded in proportion of 3.29%. The proportion of 1-monopalmitin in BRC remained almost unchanged (from 2.91% to 2.89%), and the same compound showed a sharp increase in EIS (from 0.47% to 6.92%) with the extension of cultivation.
In addition, an increase in tridecanoic acid, loliolide, pentadecanoic acid, palmitic acid, phytol, 2-monostearin, and loliolide was observed in both systems over cultivation time, while the proportion of decanoic acid remained unchanged. At the end of the cultivation period, some newly identified compounds in a proportion of less than 1%, were malic acid, (Z)-3-hexenyl-β-glucopyranoside, palmitoleamide, 1-octadecanol, desmosterol in BRC and butanedioic acid, hexadecanenitrile, 1-hexadecanol in EIS.
There were eight compounds that were predominant in both systems. These are listed in Table 3, with chemical formula and structure and biological activity reported in recent studies.
Table 3.
The most dominant compounds identified from bioreactor and incubation-shaking cabinet in Skeletonema grevillei extracts.
| No. | Compounds | Molecular formula | Structure | Properties |
|---|---|---|---|---|
| 1. | Oleamide | C18H35NO |
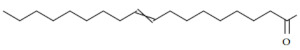
|
Anti-inflammatory, antialgal, antimicrobial and antifungal [30,31,34,35,36] |
| 2. | Palmitelaidic acid | C16H30O2 |
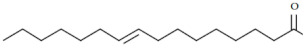
|
Antibiofilm activity [37] |
| 3. | Glycerol monostearate | C21H42O4 |
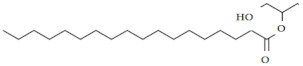
|
Anthelmintic [41] |
| 4. | Myristic acid | C14H28O2 |
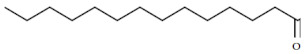
|
Antifungal, antiviral, anticancer and antiparasitic [40] |
| 5. | Cholesterol | C27H46O |
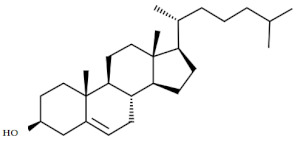
|
Anticancer, anticardiac, anti-inflammatory, antimicrobial, anti-psychotic, antioxidative [42] |
| 6. | Eicosapentaenoic acid | C20H30O2 |
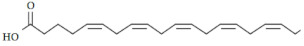
|
Cardioprotective, neuroprotective, anti-inflammatory, anticancer, antimicrobial and antioxidative [43,44,45,46] |
| 7. | 1-monopalmitin | C19H38O4 |
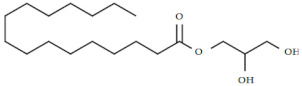
|
Antiviral [47] |
| 8. | 24-methylene cholesterol | C28H46O |
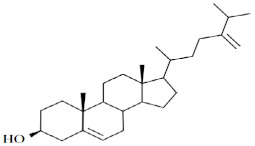
|
Anticancer [48] |
Oleamide is an amide derived from oleic acid. It is found in green algae such as Chromochloris zofingiensis and in few terrestrial plants. There are numerous biological activities related to this compound [29,30,31]. In general, oleamide from green algae is used in the treatment/prevention of arthritis, atherosclerosis, thrombosis, and cancer [32]. It was reported that oleamide from terrestrial plants (Ipomoea and Dillenia species) excretes anti-inflammatory properties [29]. Furthermore, oleamide from the fungus Colletotrichum gloeosporioides has confirmed antimicrobial activity against Staphylococcus aureus (zone of inhibition = 25 mm) [33]. The extract of Diaporthe schini, in which oleamide was a dominant metabolite, also showed antimicrobial activity against S. aureus (MIC = 125 uL/mL) [34]. Furthermore, antifungal, and antimicrobial activity of oleamide from cinnamon bark was recorded against Aspergillus flavus and Klebsiella pneumonia [35]. It is also recognized as a potential algicide in the control of cyanobacterial blooms, especially against Microcystis aeruginosa NIES -843, a toxin-producing cyanobacterium [30]. Oleamide was found to be the main metabolite among the various bioactive compounds found in the green microalgae Tetraselmis tetrathela when cultivated at a salinity of 30 ppm [36].
Fatty acids are known for their antimicrobial and antibiofilm activity. Monounsaturated palmitelaidic acid showed antibiofilm activity against Escherichia coli, inhibiting 25% of biofilm formation at 256 µg/mL, while inhibition of S. aureus was 21% at a concentration of 16 µg/mL [37]. In general, diatoms are known producers of long-chain fatty acids such as omega-3 EPA [38]. EPA is known for its health-beneficial effects, so it is used in dietary supplements and nutraceuticals. During the stationary growth phase of diatoms, the limited amount of nutrients from the culturing media is promoting the production of EPA in the cells [39]. In our study, we reported a higher accumulation of EPA in a BRC during the stationary growth phase, which was likely caused by nutrient deficiency. Moreover, myristic acid was identified as the most abundant fatty acid in S. grevillei for both cultivation systems. Marzec et al. [27] also found the myristic acid to be the most abundant fatty acid in the diatom Halamphora cf. salinicola (strain SZCZM1454A). Myristic acid has been described for a variety of biological activities, including antifungal, antiviral, anticancer, and antiparasitic activities [40].
An important cellular metabolite in triacylglycerol (TAG) biosynthesis is 1-monopalmitin. When TAGs are synthesized more intensively, a decrease in the content of 1-monopalmitin can be observed [49]. In S. grevillei we observed the decrease of 1-monopalmitin, probably due to the synthesis of fatty acids such as eicosaonic, EPA, and 5,8,11-eicosatrienoic acids.
Glycerol monostearate is known as the glycerol ester of stearic acid. There is no available literature on this compound from diatoms. So far it has been extracted from plant Dichapetalum filicaule and reported for antihelminthic activity against hookworms [41]. In general, glycerol monostearate from terrestrial plants is widely used to extend shelf-life and as an emulsifier in the food industry.
One of the most interesting pieces of information about the metabolic system of diatoms is their ability to synthesize animal and plant sterols [15]. In this study, the results showed a three-fold higher production of sterols in the BRC (12.70%) system compared to the EIS (3.69%) at the end of the cultivation period. The same ratios in sterol production between BRC and EIS can also be seen after 192 h of cultivation. The most abundant animal sterol in diatoms is cholesterol, which has a wide range of biological activities [42,50]. We identified three sterol compounds in derivatized extracts of S. grevillea, cholesterol, desmosterol, and isofucosterol. In addition to cholesterol, 24-methylene cholesterol was found among the dominant compounds. The proportion of 24-methylene cholesterol in BRC was 3.44% higher than in EIS at the end of the cultivation period. The mass spectra of the fourth sterol compound, 24-methylenecholesterol in derivatized form (Figure 4) was identified at a retention index of 3259. Since it was not available from the commercial libraries, the compound was confirmed in its non-derivatized form and in alignment with the literature data. Yang et al. [28] have provided the GC-MS spectrum for BSTFA derivatized 24-methylene cholesterol, which is in accordance with our results, while Brooks et al. [51] provided RI for the BSTFA derivatized 24-methylene cholesterol, which is also in alignment with our data.
Figure 4.

Mass spectra of derivatized sterol compound 24-methylene cholesterol in Skeletonema grevillei extracts.
A wide range of biological activities such as anticancer, anticardiac, anti-inflammatory, antimicrobial, anti-psychotic, and antioxidative have been reported for cholesterol [42]. In addition, Cutignano et al. [48] reported the anticancer activity of 24-methylene cholesterol against breast and lung cancer.
These results are consistent with previous studies that characterized the genus Skeletonema by a high content of sterols. Sterols with chemotypes of animals (cholesterol and desmosterol), plants (24-methylene cholesterol) and alga (fucosterol) were previously identified in the diatom species S. marinoi [7]. Cultivation conditions, especially a decrease in the amount of available nutrients, cause stress to microalgal cells that stimulates the metabolic production of sterols [52]. The modification of the cultivation conditions significantly affects the sterol profile [15]. A similar impact was observed in this study, at the beginning of the stationary phase in BRC when the amount of nutrients was limited higher sterol content was observed. Based on the sterol profile, it was originally hypothesized that diatoms could synthesize sterols by cyclization of lanosterol and cycloartneoids [7,50,53]. However, Gallo et al. [7] confirmed that the S. marinoi strain does not possess genes for lanosterol synthetase, but that desmosterol and cholesterol are produced via cycloartenoid pathways [15]. Since S. marinoi belongs to the same order as S. grevillei, it can be hypothesized that the biosynthesis of sterols occurs in a similar manner.
2.3. Antioxidation Activity Assays
The results of antioxidant activity (DPPH and ORAC) of diatom extracts are shown in Figure 5. At the tested concentration (10 mg dry extract/mL), the DPPH radical inhibition of the extracts from both systems was low. The highest DPPH inhibition (11.4 ± 1%) was observed for the extract from BRC after 312 h. Extracts from the stationary phase showed higher inhibition results for both systems.
Figure 5.

DPPH inhibition (left) and ORAC (right) results of Skeletonema grevillei extracts from bioreactor and incubation-shaking cabinet after 192 and 312 h. Different letters denote statistical difference (p < 0.05).
In addition to the DPPH method, the ORAC method was chosen because it best simulates in vivo conditions and provides results for antioxidant levels equivalent to those in biological systems [54]. For the ORAC assay, the diatom extracts were diluted 10-fold. The highest ORAC value of 93.3 ± 8.4 mM TE was obtained for the extract from BRC after 312 h. Similar to DPPH inhibition results, ORAC values increased over cultivation period.
Fatty acids and sterols are recognized as compounds with strong antioxidant activity [42,55]. The increase in EPA and cholesterol over the cultivation period might be the reason why the antioxidant activity of S. grevillei extracts also increased. A statistically significant difference (p < 0.05) in antioxidant activity was detected between cultivation systems at the same growth stage for ORAC, but not for DPPH.
Antioxidant activity of aqueous and ethanolic extracts of marine microalgae Tetraselmis sp. IMP3, Tetraselmis sp. CTP4, and Skeletonema sp. was evaluated using the methods ferric ion reducing antioxidant power (FRAP), DPPH, and 2,2′-azino-bis (3-ethylbenzothiazoline-6-sulphonic acid) (ABTS) [56]. The highest antioxidant activity was found with the ABTS method, reaching a reduction of more than 80% in aqueous extracts of Skeletonema sp. while the results of ethanolic extracts obtained with the DPPH method were comparable to those obtained in the present study.
3. Materials and Methods
3.1. Chemicals
LiCrosolv Ethanol (gradient grade), Sigma Aldrich (St. Louis, MO, USA) was used for extraction. In GC/MS analysis NO-Bis(trimethylsilyl)trifluoroacetamide (BSTFA) from Sigma-Aldrich (St. Louis, USA) was used as a derivatization reagent while dichlormethane (Gram-mol, Croatia) was used as a solvent for the non-derivatized sample.
3.2. Experimental Design
The strain of S. grevillei (CIM876), isolated from the northern Adriatic Sea, was donated from the culture collection of the Center for Marine Research of the Ruđer Bošković Institute (Rovinj, Croatia). The diatom was cultured in F/2 medium in two parallel systems: (i) in a bioreactor (BRC) (BiostatB Twin, Sartorius, Goettingen, Germany), (ii) in a 5-L Erlenmeyer flask set in an incubation-shaking cabinet (EIS) (Certomar T plus, Sartorius, Goettingen, Germany). Two replicas were used for each system, each containing 2 L of F/2 medium inoculated with 50 mL of S. gravillei (9 × 104 cells/mL). The medium was prepared according to the previously described recipe [57].
In both systems, diatoms were cultured at 18 °C with a light–dark photoperiod set to 12 h. The light intensity was 7500 lux in both cultivation systems. In BRC, a LED light (Led GNC Minu Deep AM140, Sicce, Pozzoleone, Italy) was used (a combination of cool white and blue), and fluorescent lamps (Fluora T8, Osram, Garching, Germany) were used in EIS. In BRC, the diatoms were stirred with an impeller at 70 rpm and the air flow rate set at 1 L/min, while in EIS they were shaken at a low speed (50 rpm) to avoid spillage.
3.3. The Growth Curve Determination
To compare the growth rate in BRC and EIS systems, 5 mL of the cultures were collected every 24 h of the cultivation period under sterile conditions. The growth curve was determined by counting the cells in a Sedgewick Rafter chamber using a light microscope (Sundew MCXI600, Micros, St. Veit/Glan, Austria) at 200xX magnification [58].
3.4. Collection and Extraction of Diatom Biomass
The biomass was collected at the exponential and at the beginning of stationary growth phases. These were determined at 192 and 312 h of culturing. Using the filtration glass microfiber filters (Grade GF/F Whatman) at 3.12 psi pressure the biomass was collected and transferred to falcon tubes and re-suspended in 10 mL of 70% ethanol. Extraction was performed in duplicate by ultrasound-assisted extraction (UAE) using an ultrasound probe (Sonoplus HD 3200, Bandalin, Berlin, Germany) set at 20 kHz ± 500 Hz. The extraction was performed for 5 min, at an amplitude of 30% and treatments of 20 s. The break between treatments was 40 s. The samples were centrifuged (Lynx 4000 centrifuge, Thermo Scientific, Waltham, MA, USA) at 29,097× g, at 4 °C for 30 min to separate the silicate frustule. The collected supernatant was filtered through a sterile CA filter with a pore diameter of 0.2 µm (LGG, Meckenheim, Germany) and evaporated in a centrifugal evaporator (RC10-22, Jouan, Herblain, France) until completely dry. The mass of the dry extract was recorded.
3.5. Identification of Compounds by GC/MS
The identification of compounds was done according to Torras-Claveria et al. with minor modifications [59]. Briefly, derivatizing agent (BSTFA) (50 μL) was added to the dry extracts. The identification of the compounds was determined using gas chromatography (GC, Nexis GC-2030, Shimadzu, Kyoto, Japan), coupled with a mass spectrometry (MS) detector (Shimadzu QP2020 NX), equipped with a split/splitless injection port. The analyses were performed using a fused silica capillary column (length 30 m × inside diameter 0.25 mm i.d., film thickness 0.25 m) (SH-5MS, Shimazu, Japan). Ultra-pure helium was used as carrier gas with a 1 mL/min flow rate. Analyses were performed with MS full scan (35–750 m/z). A perfluorotributylamine was used for calibration of the mass spectrometer, at an electron impact ionization energy of 70 eV. The column temperature program was oven equilibration time of 3 min; initial temperature 120 °C for 3 min, increased to 292 °C at a rate of 5 °C /min, then increased to 320 °C at a rate of 30 °C/min and held isothermal for 17 min. Identification of the compounds in derivatized extracts was performed by comparing their trimethylsilyl (TMS) derivative mass spectra and GC retention times with Wiley 12 & NIST 2020 databases [60,61]. All samples were injected and analyzed in duplicate.
Since commercial libraries did not offer adequate mass spectra for the identification of compounds in derivatized forms, the same samples were analyzed in non-derivatized form as well. The analyses were performed according to the same protocol, but 50 μL dichloromethane was used instead of the derivatizing agent. The derivatized and non-derivatized compounds were identified by their retention index and mass spectra using the Wiley 12 & NIST 2020 database.
3.6. Antioxidant Activity of Diatom Extracts
Prior to analyses, dried extracts were dissolved in 70% ethanol at a concentration of 10 mg/mL. Two assays, 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging and oxygen radical absorbance capacity (ORAC) were used to determine the antioxidant potential of the extracts.
DPPH radical scavenging ability of diatom extracts was measured in 96-well microplates, following the method previously reported by Čagalj et al. [62]. Briefly, DPPH radical solution (290 µL) and prepared sample (10 µL) were pipetted in microplate wells. The absorbance was measured at 517 nm and the initial absorbance of the DPPH solution was set at 1.2. After 1 h, the decrease in the absorbance was measured using the plate reader (Synergy HTX Multi-Mode Reader, BioTek Instruments, Inc., Winooski, VT, USA). The antioxidant activity of extracts was expressed as DPPH radical inhibition percentages (% inhibition). ORAC method was performed using previously published protocols [63,64]. The samples were diluted in a 1:10 ratio and 25 µL of the prepared sample and 150 µL of fluorescein were added to each well of the microtiter plate. For 30 min, the plates were thermostated at 37 °C followed by the addition of 25 µL of AAPH. The measurements were recorded every minute over 80 min, at excitation and emission wavelengths of 485 and 520 nm. Results are expressed as µM Trolox equivalents (mM TE). Both assays were done in triplicate.
3.7. Statistical Analyses
Cell density data, collected and recorded using the Sedgewick Rafter method, were used for the analysis. The analyses of variance (one-way ANOVA followed by Fisher’s least significant difference test) was used to determine the significant changes (p < 0.05) in the growth rate in each system over time and the difference between the EIS and BRC systems, as well as antioxidant activities [65]. Analyses were performed using Statgraphics Centurion-Ver.16.1.11 (StatPoint Technologies, Inc., Warrenton, VA, USA).
4. Conclusions
This study confirms that bioreactor cultivation is a successful system for the cultivation of diatoms. Optimal growth conditions in bioreactors allowed faster entry into the exponential and stationary growth phases. It could be hypothesized that prolongation of cultivation would result in slightly higher biomass in the bioreactor; however, the prolongation would also result in the death of diatom cells. The results of the chemical profiles revealed that oleamide was most abundant compound in the BRC and EIS systems, and that palmitelaidic acid, glycerol monostearate, myristic acid, cholesterol, eicosapentaenoic acid, 1-monopalmitin and 24-methylene cholesterol were also abundant. All of the mentioned compounds have been previously characterized for their biological activities, which is why some of them are already used in industry, while the other compounds have the potential for biotechnological applications. Moreover, in this study, tentative identification of derivatized 24-methylene cholesterol was obtained, which was not previously found in its derivatized form in the commercial libraries but is known from the literature. For precise identification techniques such HR-MS or NMR should be used. In addition, ethanolic extracts of S. grevillei showed low antioxidant activity at the tested concentration, so further studies should focus on the biological activity at higher concentrations and quantitative determination of the biologically active compounds. It should also be emphasized that, in addition to the extracts, silicate diatom frustules are also used in nanotechnology, which will certainly affect the profitability of the production of these microalgae in the future. Future research should focus on finding conditions under which it is possible to maximize the production of metabolites that have the potential for biotechnological applications.
Acknowledgments
The authors would like to thank Daniela Marić Pfannkuchen from the Laboratory for Evolutionary Ecology of the Center for Marine Research of the Ruđer Bošković Institute (Rovinj, Croatia) for the donation of the Skeletonema grevillei strain from the institute culture collection, and BIOCentre–Incubation Centre for Biosciences for technical support.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/md20110697/s1. Additional information of identified compound such as, retention time and m/z, available in Supplementary Table S1 List of identified compounds from Skeletonema grevillei extracts by GC-MS in incubation-shaking cabinet after 312 hours.
Author Contributions
Conceptualization, R.F. and V.Š.; methodology, R.F., M.P. and V.Š.; validation, M.P., V.Š. and T.S.; formal analysis, R.F. and M.P.; investigation, R.F. and V.Š.; data curation, R.F., V.Š. and M.P.; writing—original draft preparation, R.F. and V.Š.; writing—review and editing, V.Š., M.P. and T.S.; visualization, R.F. and V.Š.; supervision, V.Š. and T.S. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data available on request.
Conflicts of Interest
The authors declare no conflict of interest.
Funding Statement
This research was partially supported under the project STIM–REI, Contract Number: KK.01.1.1.01.0003, a project funded by the EU through the ERDF–the Operational Programme Competitiveness and Cohesion 2014-2020 (KK.01.1.1.01).
Footnotes
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Sathasivam R., Radhakrishnan R., Hashem A., Abd_Allah E.F. Microalgae Metabolites: A Rich Source for Food and Medicine. Saudi J. Biol. Sci. 2019;26:709–722. doi: 10.1016/j.sjbs.2017.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ummalyma S.B., Sirohi R., Udayan A., Yadav P., Raj A., Sim S.J., Pandey A. Sustainable Microalgal Biomass Production in Food Industry Wastewater for Low-Cost Biorefinery Products: A Review. Phytochem. Rev. 2022;3:1–23. doi: 10.1007/s11101-022-09814-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Spaulding S.A., Potapova M.G., Bishop I.W., Lee S.S., Gasperak T.S., Jovanoska E., Furey P.C., Edlund M.B. Diatoms.Org: Supporting Taxonomists, Connecting Communities. Diatom Res. 2021;36:291–304. doi: 10.1080/0269249X.2021.2006790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Stonik V.S., Stonik I. Low-Molecular-Weight Metabolites from Diatoms: Structures, Biological Roles and Biosynthesis. Mar. Drugs. 2015;13:3672–3709. doi: 10.3390/md13063672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Benavente-Valdés J.R., Méndez-Zavala A., Hernández-López I., Carreón-González B.A., Velázquez-Arellano M.E., Morales-Oyervides L., Montañez-Saénz J.C. Unconventional Microalgae Species and Potential for Their Use in the Food Industry. Cultured Microalgae for the Food Industry. Current and Potential Applications; Academic Press; Cambridge, MA, USA: 2021. pp. 49–71. [DOI] [Google Scholar]
- 6.Silva M., Kamberovic F., Uota S.T., Kovan I.M., Viegas C.S.B., Simes D.C., Gangadhar K.N., Varela J., Barreira L. Microalgae as Potential Sources of Bioactive Compounds for Functional Foods and Pharmaceuticals. Appl. Sci. 2022;12:5877. doi: 10.3390/app12125877. [DOI] [Google Scholar]
- 7.Gallo C., Landi S., D’Ippolito G., Nuzzo G., Manzo E., Sardo A., Fontana A. Diatoms Synthesize Sterols by Inclusion of Animal and Fungal Genes in the Plant Pathway. Sci. Rep. 2020;10:4204. doi: 10.1038/s41598-020-60993-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Vidoudez C., Pohnert G. Comparative Metabolomics of the Diatom Skeletonema Marinoi in Different Growth Phases. Metabolomics. 2012;8:654–669. doi: 10.1007/s11306-011-0356-6. [DOI] [Google Scholar]
- 9.Gonzalez-Fernandez C., Muñoz R. Microalgae-Based Biofuels and Bioproducts: From Feedstock Cultivation to End-Products. Woodhead Publishing; Sawston, UK: 2017. (Woodhead Publishing Series in Energy). [Google Scholar]
- 10.Gao G., Wu M., Fu Q., Li X., Xu J. A Two-Stage Model with Nitrogen and Silicon Limitation Enhances Lipid Productivity and Biodiesel Features of the Marine Bloom-Forming Diatom Skeletonema Costatum. Bioresour. Technol. 2019;289:121717. doi: 10.1016/j.biortech.2019.121717. [DOI] [PubMed] [Google Scholar]
- 11.Yuan W., Gao G., Shi Q., Xu Z., Wu H. Combined Effects of Ocean Acidification and Warming on Physiological Response of the Diatom Thalassiosira Pseudonana to Light Challenges. Mar. Environ. Res. 2018;135:63–69. doi: 10.1016/j.marenvres.2018.01.016. [DOI] [PubMed] [Google Scholar]
- 12.Souffreau C., Vanormelingen P., Verleyen E., Sabbe K., Vyverman W. Tolerance of Benthic Diatoms from Temperate Aquatic and Terrestrial Habitats to Experimental Desiccation and Temperature Stress. Phycologia. 2010;49:309–324. doi: 10.2216/09-30.1. [DOI] [Google Scholar]
- 13.Fukami K., Nishimura S., Ogusa M., Asada M., Nishijima T. Continuous Culture with Deep Seawater of a Benthic Food Diatom Nitzschia Sp. Hydrobiologia. 1997;358:245–249. doi: 10.1023/A:1003141104693. [DOI] [Google Scholar]
- 14.Vuppaladadiyam A.K., Prinsen P., Raheem A., Luque R., Zhao M. Microalgae Cultivation and Metabolites Production: A Comprehensive Review. Biofuels Bioprod. Biorefining. 2018;12:304–324. doi: 10.1002/bbb.1864. [DOI] [Google Scholar]
- 15.Fagundes M.B., Vendruscolo R.G., Wagner R. Sterols from Microalgae. Elsevier Inc.; Amsterdam, The Netherlands: 2020. [DOI] [Google Scholar]
- 16.Iglesias S., Míguez C., Sánchez A., Cancela A., Álvarez X. Thalassiosira Pseudonana and Skeletonema Costatum Biomass Optimization: Cultivation, Harvesting, Extraction of Oils and Biodiesel and Pelletization of the Residue. J. Sea Res. 2022;187:102243. doi: 10.1016/j.seares.2022.102243. [DOI] [Google Scholar]
- 17.Sharmin T., Monirul Hasan C.M., Aftabuddin S., Rahman M.A., Khan M. Growth, Fatty Acid, and Lipid Composition of Marine Microalgae Skeletonema Costatum Available in Bangladesh Coast: Consideration as Biodiesel Feedstock. J. Mar. Biol. 2016;2016:6832847. doi: 10.1155/2016/6832847. [DOI] [Google Scholar]
- 18.Thangaraj S., Sun J. The Biotechnological Potential of the Marine Diatom Skeletonema Dohrnii to the Elevated Temperature and PCO2. Mar. Drugs. 2020;18:259. doi: 10.3390/md18050259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Jiang X., Han Q., Gao X., Gao G. Conditions Optimising on the Yield of Biomass, Total Lipid, and Valuable Fatty Acids in Two Strains of Skeletonema Menzelii. Food Chem. 2016;194:723–732. doi: 10.1016/j.foodchem.2015.08.073. [DOI] [PubMed] [Google Scholar]
- 20.Raniello R., Iannicelli M.M., Nappo M., Avila C., Zupo V. Production of Cocconeis Neothumensis (Bacillariophyceae) Biomass in Batch Cultures and Bioreactors for Biotechnological Applications: Light and Nutrient Requirements. J. Appl. Phycol. 2007;19:383–391. doi: 10.1007/s10811-006-9145-4. [DOI] [Google Scholar]
- 21.Ingebrigtsen R.A., Hansen E., Andersen J.H., Eilertsen H.C. Light and Temperature Effects on Bioactivity in Diatoms. J. Appl. Phycol. 2016;28:939–950. doi: 10.1007/s10811-015-0631-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Bondoc K.G.V., Heuschele J., Gillard J., Vyverman W., Pohnert G. Selective Silicate-Directed Motility in Diatoms. Nat. Commun. 2016;7:10540. doi: 10.1038/ncomms10540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Leynaert A., Fardel C., Beker B., Soler C., Delebecq G., Lemercier A., Pondaven P., Durand P.E., Heggarty K. Diatom Frustules Nanostructure in Pelagic and Benthic Environments. Silicon. 2018;10:2701–2709. doi: 10.1007/s12633-018-9809-0. [DOI] [Google Scholar]
- 24.Liu Y., Song X., Cao X., Yu Z. Responses of Photosynthetic Characters of Skeletonema Costatum to Different Nutrient Conditions. J. Plankton Res. 2013;35:165–176. doi: 10.1093/plankt/fbs080. [DOI] [Google Scholar]
- 25.Gao G., Shi Q., Xu Z., Xu J., Campbell D.A., Wu H. Global Warming Interacts with Ocean Acidification to Alter PSII Function and Protection in the Diatom Thalassiosira Weissflogii. Environ. Exp. Bot. 2018;147:95–103. doi: 10.1016/j.envexpbot.2017.11.014. [DOI] [Google Scholar]
- 26.Ramirez E.E., Gonzales M.A., Cifuentes A.S., Inostroza I., Urrutia R.E. Culture and Growth of Two Benthic Diatoms Species Isolated from the Salar Del Huasco (North of Chile, 20° S) at Different Conditions of Temperature, Light and Nutrient. Gayana Bot. 2015;72:165–176. doi: 10.4067/S0717-66432015000200001. [DOI] [Google Scholar]
- 27.Marzec M., Dąbek P., Witkowski A., Monedeiro F., Pomastowski P., Buszewski B., Nowak I. Lipid Constituents of Diatoms (Halamphora) as Components for Production of Lipid Nanoparticles. Pharmaceutics. 2022;14:1171. doi: 10.3390/pharmaceutics14061171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Yang J., Li C., Zhang Y. Engineering of Saccharomyces Cerevisiae for 24-Methylene-Cholesterol Production. Biomolecules. 2021;11:1710. doi: 10.3390/biom11111710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Ameamsri U., Chaveerach A., Sudmoon R., Tanee T., Peigneur S., Tytgat J. Oleamide in Ipomoea and Dillenia Species and Inflammatory Activity Investigated through Ion Channel Inhibition. Curr. Pharm. Biotechnol. 2020;22:254–261. doi: 10.2174/1389201021666200607185250. [DOI] [PubMed] [Google Scholar]
- 30.Shao J., He Y., Li F., Zhang H., Chen A., Luo S., Gu J.D. Growth Inhibition and Possible Mechanism of Oleamide against the Toxin-Producing Cyanobacterium Microcystis Aeruginosa NIES-843. Ecotoxicology. 2016;25:225–233. doi: 10.1007/s10646-015-1582-x. [DOI] [PubMed] [Google Scholar]
- 31.Leitner P.D., Jakschitz T., Gstir R., Stuppner S., Perkams S., Kruus M., Trockenbacher A., Griesbeck C., Bonn G.K., Huber L.A., et al. Anti-Inflammatory Extract from Soil Algae Chromochloris Zofingiensis Targeting TNFR/NF-ΚB Signaling at Different Levels. Cells. 2022;11:1407. doi: 10.3390/cells11091407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Moon S.M., Lee S.A., Hong J.H., Kim J.S., Kim D.K., Kim C.S. Oleamide Suppresses Inflammatory Responses in LPS-Induced RAW264.7 Murine Macrophages and Alleviates Paw Edema in a Carrageenan-Induced Inflammatory Rat Model. Int. Immunopharmacol. 2018;56:179–185. doi: 10.1016/j.intimp.2018.01.032. [DOI] [PubMed] [Google Scholar]
- 33.Premjanu N., Jaynthy C. Identification and Characterization of Antimicrobial Metabolite from an Endophytic Fungus, Colletotrichum Gloeosporioides Isolated from Lannea Corammendalica. Int. J. ChemTech Res. 2015;7:369–374. [Google Scholar]
- 34.Dos Reis C.M., da Rosa B.V., da Rosa G.P., do Carmo G., Morandini L.M.B., Ugalde G.A., Kuhn K.R., Morel A.F., Jahn S.L., Kuhn R.C. Antifungal and Antibacterial Activity of Extracts Produced from Diaporthe Schini. J. Biotechnol. 2019;294:30–37. doi: 10.1016/j.jbiotec.2019.01.022. [DOI] [PubMed] [Google Scholar]
- 35.Hameed I.H., Altameme H.J., Mohammed G.J. Evaluation of Antifungal and Antibacterial Activity and Analysis of Bioactive Phytochemical Compounds of Cinnamomum Zeylanicum (Cinnamon Bark) Using Gas Chromatography-Mass Spectrometry. Orient. J. Chem. 2016;32:1769–1788. doi: 10.13005/ojc/320406. [DOI] [Google Scholar]
- 36.El-Kassas H.Y., El-Sheekh M.M. Induction of the Synthesis of Bioactive Compounds of the Marine Alga Tetraselmis Tetrathele (West) Butcher Grown under Salinity Stress. Egypt. J. Aquat. Res. 2016;42:385–391. doi: 10.1016/j.ejar.2016.10.006. [DOI] [Google Scholar]
- 37.Yuyama K.T., Rohde M., Molinari G., Stadler M., Abraham W.R. Unsaturated Fatty Acids Control Biofilm Formation of Staphylococcus Aureus and Other Gram-Positive Bacteria. Antibiotics. 2020;9:788. doi: 10.3390/antibiotics9110788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Yang R., Wei D., Xie J. Diatoms as Cell Factories for High-Value Products: Chrysolaminarin, Eicosapentaenoic Acid, and Fucoxanthin. Crit. Rev. Biotechnol. 2020;40:993–1009. doi: 10.1080/07388551.2020.1805402. [DOI] [PubMed] [Google Scholar]
- 39.Zulu N.N., Zienkiewicz K., Vollheyde K., Feussner I. Current Trends to Comprehend Lipid Metabolism in Diatoms. Prog. Lipid Res. 2018;70:1–16. doi: 10.1016/j.plipres.2018.03.001. [DOI] [PubMed] [Google Scholar]
- 40.Javid S., Purohit M.N., Yogish Kumar H., Ramya K., Mithuna N.F.A., Salahuddin M.D., Prashantha Kumar B.R. Semisynthesis of Myristic Acid Derivatives and Their Biological Activities: A Critical Insight. J. Biol. Act. Prod. Nat. 2020;10:455–472. doi: 10.1080/22311866.2020.1865836. [DOI] [Google Scholar]
- 41.Chama M.A., Dziwornu G.A., Waibel R., Osei-Safo D., Addae-Mensah I., Otchere J., Wilson M. Isolation, Characterization, and Anthelminthic Activities of a Novel Dichapetalin and Other Constituents of Dichapetalum Filicaule. Pharm. Biol. 2016;54:1179–1188. doi: 10.3109/13880209.2015.1059861. [DOI] [PubMed] [Google Scholar]
- 42.Zhang K., Li T., Shan X., Lu R., Zhang S., Xu H. Cholesterol: Bioactivities, Structural Modification, Mechanisms of Action, and Structure-Activity Relationships. Mini-Rev. Med. Chem. 2021;21:1830–1848. doi: 10.2174/1389557521666210105123320. [DOI] [PubMed] [Google Scholar]
- 43.Ceccarelli V., Ronchetti S., Marchetti M.C., Calvitti M., Riccardi C., Grignani F., Vecchini A. Molecular Mechanisms Underlying Eicosapentaenoic Acid Inhibition of HDAC1 and DNMT Expression and Activity in Carcinoma Cells. Biochim. Biophys. Acta—Gene Regul. Mech. 2020;1863:194481. doi: 10.1016/j.bbagrm.2020.194481. [DOI] [PubMed] [Google Scholar]
- 44.Bie N., Han L., Meng M., Zhang Y., Guo M., Wang C. Anti-Tumor Mechanism of Eicosapentaenoic Acid (EPA) on Ovarian Tumor Model by Improving the Immunomodulatory Activity in F344 Rats. J. Funct. Foods. 2020;65:103739. doi: 10.1016/j.jff.2019.103739. [DOI] [Google Scholar]
- 45.Ferreira M., Teixeira C., Abreu H., Silva J., Costas B., Kiron V., Valente L.M.P. Nutritional Value, Antimicrobial and Antioxidant Activities of Micro- and Macroalgae, Single or Blended, Unravel Their Potential Use for Aquafeeds. J. Appl. Phycol. 2021;33:3507–3518. doi: 10.1007/s10811-021-02549-2. [DOI] [Google Scholar]
- 46.Ribeiro-Vidal H., Sánchez M.C., Alonso-Español A., Figuero E., Ciudad M.J., Collado L., Herrera D., Sanz M. Antimicrobial Activity of Epa and Dha against Oral Pathogenic Bacteria Using an in Vitro Multi-Species Subgingival Biofilm Model. Nutrients. 2020;12:2812. doi: 10.3390/nu12092812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Cheng J.-C., Liaw C.-C., Lin M.-K., Chen C.-J., Chao C.-L., Chao C.-H., Kuo Y.-H., Chiu Y.-P., Peng Y.-S., Hui-Chi H. Anti-Influenza Virus Activity and Chemical. Molecules. 2020;25:4427. doi: 10.3390/molecules25194427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Cutignano A., Conte M., Tirino V., del Vecchio V., de Angelis R., Nebbioso A., Altucci L., Romano G. Cytotoxic Potential of the Marine Diatom Thalassiosira Rotula: Insights into Bioactivity of 24-Methylene Cholesterol. Mar. Drugs. 2022;20:595. doi: 10.3390/md20100595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Shi Y., Chen Z., Li Y., Cao X., Yang L., Xu Y., Li Z., He N. Function of ORFC of the Polyketide Synthase Gene Cluster on Fatty Acid Accumulation in Schizochytrium Limacinum SR21. Biotechnol. Biofuels. 2021;14:163. doi: 10.1186/s13068-021-02014-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Rampen S.W., Abbas B.A., Schouten S., Damsté J.S.S. A Comprehensive Study of Sterols in Marine Diatoms (Bacillariophyta): Implications for Their Use as Tracers for Diatom Productivity. Limnol. Oceanogr. 2010;55:91–105. doi: 10.4319/lo.2010.55.1.0091. [DOI] [Google Scholar]
- 51.Brooks C.J.W., Cole W.J., McIntyre H.B., Smith A.G. Selective Reactions in the Analysis and Characterization of Steroids by Gas Chromatography-Mass Spectrometry. Lipids. 1980;15:745–755. doi: 10.1007/BF02534028. [DOI] [PubMed] [Google Scholar]
- 52.Kumari P., Kumar M., Reddy C.R.K., Jha B. Algal Lipids, Fatty Acids and Sterols. Woodhead Publishing Ltd.; Sawston, UK: 2013. [DOI] [Google Scholar]
- 53.Barrett S.M., Volkman J.K., Dunstan G.A., LeRoi J.-M. Sterols of 14 Species of Marine Diatoms (Bacillariophyta) J. Phycol. 1995;31:360–369. doi: 10.1111/j.0022-3646.1995.00360.x. [DOI] [Google Scholar]
- 54.Bank B.G., Schauss A. Antioxidant Testing: An ORAC Update. Nutraceuticals World. 2004;7:68–71. [Google Scholar]
- 55.Xiao B., Li Y., Lin Y., Lin J., Zhang L., Wu D., Zeng J., Li J., Liu J.w., Li G. Eicosapentaenoic Acid (EPA) Exhibits Antioxidant Activity via Mitochondrial Modulation. Food Chem. 2022;373:131389. doi: 10.1016/j.foodchem.2021.131389. [DOI] [PubMed] [Google Scholar]
- 56.Cardoso C., Pereira H., Franca J., Matos J., Monteiro I., Pousão-Ferreira P., Gomes A., Barreira L., Varela J., Neng N., et al. Lipid Composition and Some Bioactivities of 3 Newly Isolated Microalgae (Tetraselmis Sp. IMP3, Tetraselmis Sp. CTP4, and Skeletonema Sp.) Aquac. Int. 2020;28:711–727. doi: 10.1007/s10499-019-00489-w. [DOI] [Google Scholar]
- 57.Andersen R.A. Algal Culturing Techniques. Elsevier Inc.; Amsterdam, The Netherlands: 2010. [Google Scholar]
- 58.Karlson B., Caroline C., Bresnan E. Microscopic and Molecular Methods for Quantitative Phytoplankton Analysis. United Nations Educational, Scientific and Cultural Organization (UNESCO); Paris, France: 2010. [Google Scholar]
- 59.Torras-Claveria L., Berkov S., Jáuregui O., Caujapé J., Viladomat F., Codina C., Bastida J. Metabolic Profiling of Bioactive Pancratium Canariense Extracts by GC-MS. Phytochem. Anal. 2010;21:80–88. doi: 10.1002/pca.1158. [DOI] [PubMed] [Google Scholar]
- 60.NIST NIST Chemistry WebBook. [(accessed on 11 October 2022)]; Available online: https://webbook.nist.gov/chemistry/
- 61.Oberacher H. Wiley Registry of Tandem Mass Spectral Data, MS for ID. Wiley; Hoboken, NJ, USA: 2012. [Google Scholar]
- 62.Čagalj M., Skroza D., del Carmen Razola-Díaz M., Verardo V., Bassi D., Frleta R., Generalić Mekinić I., Tabanelli G., Šimat V. Variations in the Composition, Antioxidant and Antimicrobial Activities of Cystoseira Compressa during Seasonal Growth. Mar. Drugs. 2022;20:64. doi: 10.3390/md20010064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Burčul F., Generalić Mekinić I., Radan M., Rollin P., Blažević I. Isothiocyanates: Cholinesterase Inhibiting, Antioxidant, and Anti-Inflammatory Activity. J. Enzym. Inhib. Med. Chem. 2018;33:577–582. doi: 10.1080/14756366.2018.1442832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Prior R.L., Hoang H., Gu L., Wu X., Bacchiocca M., Howard L., Hampsch-Woodill M., Huang D., Ou B., Jacob R. Assays for Hydrophilic and Lipophilic Antioxidant Capacity (Oxygen Radical Absorbance Capacity (ORACFL)) of Plasma and Other Biological and Food Samples. J. Agric. Food Chem. 2003;51:3273–3279. doi: 10.1021/jf0262256. [DOI] [PubMed] [Google Scholar]
- 65.Šimat V., Vlahović J., Soldo B., Generalić Mekinić I., Čagalj M., Hamed I., Skroza D. Production and Characterization of Crude Oils from Seafood Processing By-Products. Food Biosci. 2020;33:100484. doi: 10.1016/j.fbio.2019.100484. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data available on request.