

ABSTRACT
The current study aimed to determine the role of a microRNA (miRNA), miR-140-3p, in the control of RhoA expression in bronchial smooth muscle cells (BSMCs). In cultured human BSMCs, incubation with interleukin-13 (IL-13) caused an up-regulation of RhoA protein concurrently with a down-regulation of miR-140-3p. Transfection of the cells with a miR-140-3p inhibitor caused an increase in basal RhoA protein level. Although a mimic of miR-140-3p had little effect on the basal RhoA level, its treatment inhibited the IL-13-induced up-regulation of RhoA. These findings suggest that RhoA expression is negatively regulated by miR-140-3p, and that the negative regulation is inhibited by IL-13 to cause an up-regulation of RhoA protein in BSMCs.
KEYWORDS: Microrna (miRNA), miR-140-3p, RhoA, asthma, bronchial smooth muscle hyperresponsiveness
Increased airway responsiveness to a wide variety of stimuli, called airway hyperresponsiveness (AHR), is a characteristic feature of allergic bronchial asthma. Airway smooth muscles (ASMs) of individual asthmatics exhibited an increased agonist-induced contraction [1–3], indicating that an abnormality of the property of ASM per se is involved in the exaggerated airway narrowing. Rapid relief from airway limitation by bronchorelaxants also suggests an involvement of augmented ASM contraction in the airway obstruction. It is thus important for the development of asthma therapy to understand changes in the contractile signalling of ASM cells associated with the disease.
A small GTPase RhoA is a key protein involved in the Ca2+ sensitization of smooth muscle contraction, including ASMs e.g. [4]. Increasing evidence suggests that the augmented RhoA-mediated Ca2+ sensitization of ASM contraction is associated with the AHR in experimental asthma [5–10]. An up-regulation of RhoA in the ASMs has also been demonstrated in animal models of allergic asthma [5–10]. An involvement of interleukin-13 (IL-13), one of the asthma-related cytokines, in the RhoA up-regulation has been suggested [11–13], although the mechanism is not fully understood.
MicroRNAs (miRNAs) are small single-stranded non-coding RNAs that negatively modulate gene expression [14–16]. There is increasing evidence that dysregulation of miRNAs is found in multiple human diseases, including chronic obstructive pulmonary disease (COPD) and asthma [17–20]. Although the miRNA roles in function and pathology of ASM remain largely unknown, increasing evidence suggests miR-140-3p as a candidate miRNA that contributes to dysfunction of ASM in asthma [21–24]. More recently, RhoA has been suggested as one of the targets of miR-140-3p in ASM cells [25]. Here, we show that IL-13 down-regulates miR-140-3p to cause up-regulation of RhoA in bronchial smooth muscle (BSM) cells.
Methods
Human bronchial smooth muscle cell (hBSMC) culture and sample collection
Normal human BSM cells (hBSMCs; Cambrex Bio Science Walkersville, Inc., Walkersville, MD) were maintained in SmBM medium (Cambrex) supplemented with 5% foetal bovine serum, 0.5 ng/mL human epidermal growth factor (hEGF), 5 µg/mL insulin, 2 ng/mL human fibroblast growth factor-basic (hFGF-b), 50 µg/mL gentamicin and 50 ng/mL amphotericin B. Cells were maintained at 37°C in a humidified atmosphere (5% CO2), fed every 48–72 hours, and passaged when cells reached 90–95% confluence. Then the hBSMCs (passages five through seven) were seeded in 6-well plates (Becton Dickinson Labware, Franklin Lakes, NJ) at a density of 3,500 cells/cm2 and, when 80–85% confluence was observed, cells were cultured without serum for 24 hours before addition of recombinant human IL-13 (100 ng/mL; PeproTech EC, Ltd., London, UK). At the indicated time after the IL-13 treatment, cells were washed with phosphate-buffered saline, immediately collected and disrupted with 1x SDS sample buffer (150 µL/well), and used for Western blot analyses. Total RNAs containing miRNAs were extracted using VantageTM total RNA purification kit (Origene Technologies, Inc., Rockville, MD) according to the manufacturer’s instructions.
Transfection of hBSMCs with miR-140-3p inhibitor and mimic
The hBSMCs were plated at a density of 2 × 105 cells/well in a 6-well plate with SmGM medium. The next day, cells were transfected with 100 pmol (final 40 nM) of either an inhibitor of miR-140-3p (Peptide Nucleic Acids (PNAsTM) miRNA inhibitor; Cat. No.: PI-1171; Panagene Inc., Daejeon, Korea), a mimic of miR-140-3p (miCENTURY OX miNatural; Cat. No.: HN0000140A1-2; Cosmo Bio Co., Ltd., Tokyo, Japan), or a control RNA (Cosmo Bio Co., Ltd.) using the LipofectamineTM 2000 transfection reagent (Thermo Fisher Scientific, Waltham, MA) according to the manufacturer’s instructions. After a 4-hour transfection in Opti-MEMTM I reduced serum medium (Thermo Fisher Scientific), cells were cultured in SmGM medium for 20 hours and then in serum-free SmGM medium. Forty-eight hours after the transfection, IL-13 (100 ng/mL) or its vehicle (PBS) was administered to the cells.
Quantitative RT-PCR analyses
To synthesize cDNAs, 200 ng of total RNAs were poly-adenylated and cDNAs were synthesized using miRCURY LNATM Universal cDNA Synthesis Kit (#203,300, Exiqon A/S, Vedbaek, Denmark) according to the manufacturer’s instructions. Then the RT reaction mixture (1 µL) was subjected to real-time PCR analyses using StepOneTM real-time PCR system (Applied Biosystems, Foster City, CA) with Fast SYBR Green Master Mix (Applied Biosystems) according to the manufacturer’s instructions. The reactions were incubated in a 96-well optical plate at 95°C for 20 seconds, following by 43 cycles of 95°C for 3 seconds and 60°C for 30 seconds. The primer sets used were: LNATM PCR primer sets for human miR-140-3p (#204,304, Exiqon A/S) and for human U6 snRNA (#203,907, Exiqon A/S).
Western blot analyses
Protein samples were subjected to 15% sodium dodecyl sulphate-polyacrylamide gel electrophoresis and the proteins were then electrophoretically transferred to a PVDF membrane (WSE-4051CP; Atto, Co., Tokyo, Japan). After blocking with EzBlock ChemiTM (AE-1475CP; Atto, Co.), the PVDF membrane was incubated with polyclonal rabbit anti-RhoA (sc-418; 1:2,500 dilution; Santa Cruz Biotechnology, Inc., Santa Cruz, CA) antibody. Then the membrane was incubated with horseradish peroxidase-conjugated donkey anti-rabbit IgG (sc-2313; 1:2,500 dilution; Santa Cruz Biotechnology, Inc.), detected by EzWestBlueTM (AE-1490CP; Atto, Co.) and analysed by a densitometry system. Detection of house-keeping gene was also performed on the same membrane by using monoclonal mouse anti-GAPDH (sc-32233; 1:10,000 dilution; Santa Cruz Biotechnology, Inc.) to confirm the same amount of proteins loaded.
Data and statistical analyses
In the real-time PCR analyses, the comparative threshold cycle (CT) method was used for relative quantification of the target genes. Differences in the CT values (ΔCT) between miR-140-3p and U6 snRNA were calculated to determine the relative expression levels, using the following formula: ΔΔCT = (ΔCT of the treated sample) – (ΔCT of the control sample). The relative expression level between the samples was calculated according to equation 2−ΔΔCT.
All the data are expressed as the mean ± S.E. Statistical significance of difference was determined by unpaired Student’s t-test or one-way analysis of variance (ANOVA) with post hoc Bonferroni/Dunn (PrismTM 5 for Mac OS X; GraphPad Software, Inc., La Jolla, CA). A value of p < 0.05 was considered significant.
Results
Effects of IL-13 on expression levels of RhoA protein and miR-140-3p in cultured human bronchial smooth muscle cells (hBSMCs)
Since IL-13 is capable of inducing an up-regulation of RhoA protein in hBSMCs [11,12], the effect of IL-13 on the expression of miR-140-3p was determined in the hBSMCs. Consistent with our previous studies [11,12], treatment of the cells with IL-13 (100 ng/mL) caused an up-regulation of RhoA protein (Figure 1(a)). As shown in Figure 1(b), the real-time RT-qPCR analyses revealed that the expression level of miR-140-3p was significantly decreased in the hBSMCs treated with IL-13 (p < 0.01 by unpaired Student’s t-test). The treatment also reduced the 5ʹ-strand of miR-140 duplex, miR-140-5p, significantly (1.06 ± 0.10 in Vehicle versus 0.68 ± 0.15 in IL-13, p < 0.05).
Figure 1.
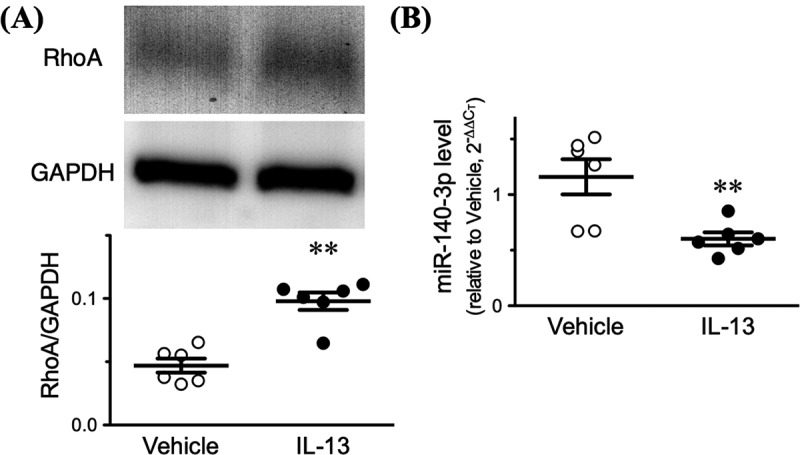
Interleukin-13 (IL-13) causes an up-regulation of RhoA protein concurrently with a down-regulation of miR-140-3p in cultured human bronchial smooth muscle cells. Cells were treated with IL-13 (100 ng/mL) or its vehicle for 24 hours, and proteins and total RNAs including miRNAs were extracted. (a) (Upper panel) Representative Western blots. The relative expression of RhoA to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) proteins in each sample was calculated and the data are summarized in lower panel. (b) miR-140-3p expression level determined by quantitative real-time reverse transcriptase-polymerase chain reaction. The relative gene expressions of miR-140-3p to U6 snRNA was calculated by the 2−ΔΔCT methods. Results are presented as mean ± S.E. from 6 independent experiments. **p < 0.01 versus Vehicle group by unpaired Student’s t-test
Effect of inhibition of miR-140-3p on RhoA protein expression in cultured human bronchial smooth muscle cells (hBSMCs)
To determine the role of miR-140-3p in RhoA expression of the ASM cells, cultured hBSMCs were transfected with its synthetic inhibitor, miR-140-3p inhibitor. A non-targeting 20–25 nt RNA was used as a control RNA. Immunoblot analyses of these cells revealed that RhoA protein expression was significantly increased when the cells were transfected with miR-140-3p inhibitor (Figure 2, ‘Control RNA’ versus ‘140–3p inhibitor’ groups), indicating that miR-140-3p is an endogenous modulator of RhoA protein expression in hBSMCs. On the other hand, transfection of the cells with miR-140-3p mimic had no effect on the expression level of RhoA protein (Figure 2, ‘Control RNA’ versus ‘140–3p mimic’ groups).
Figure 2.
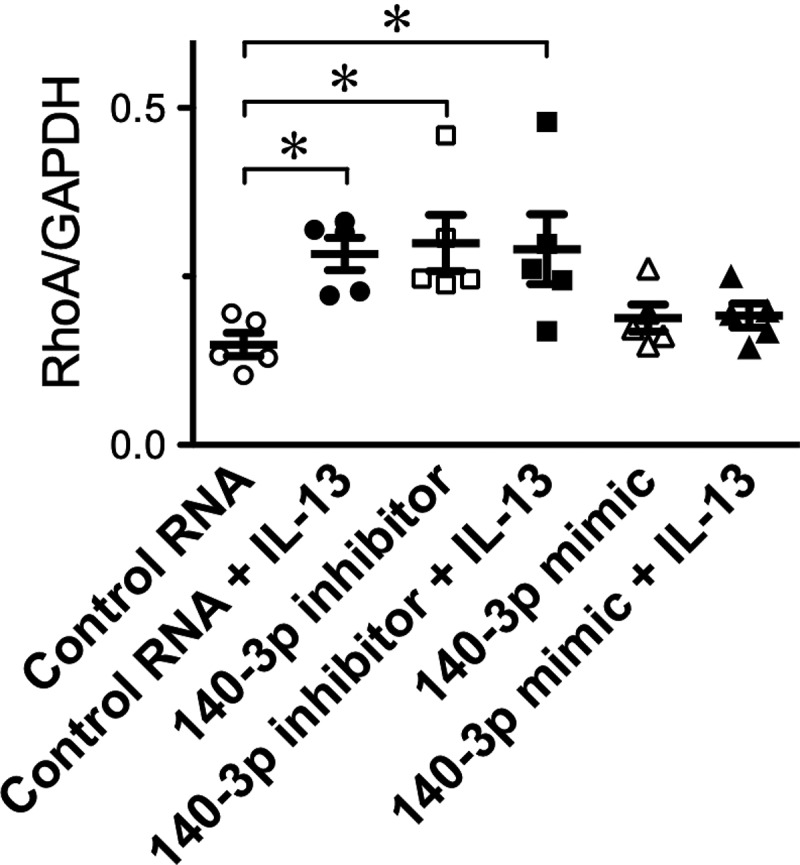
Effects of miR-140-3p inhibitor and mimic on the interleukin-13 (IL-13)-induced up-regulation of RhoA protein in cultured human bronchial smooth muscle cells. Cells were transfected with a non-targeting 20–25 nt RNA (Control RNA), a miR-140-3p inhibitor (140–3p inhibitor), or a miR-140-3p mimic (140–3p mimic), and then treated with IL-13 (100 ng/mL) or its vehicle 48 hours after the transfection. Twenty-four hours after the IL-13 treatment, total protein samples were prepared and immunoblot analyses were performed. The relative expression of RhoA to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) in each sample was calculated and the data are summarized. Results are presented as mean ± S.E. from 5 independent experiments. *p < 0.05 versus Control RNA only group by Bonferroni/Dunn test. Note that the RhoA up-regulation induced by IL-13 (Control RNA + IL-13 group) was inhibited by the transfection of miR-140-3p mimic (140–3p mimic + IL-13 group)
Effects of miR-140-3p inhibitor and mimic on the IL-13-induced up-regulation of RhoA protein in cultured human bronchial smooth muscle cells (hBSMCs)
To determine the role of miR-140-3p on the IL-13-induced up-regulation of RhoA protein, the hBSMCs transfected with miR-140-3p inhibitor or mimic were stimulated by IL-13 (100 ng/mL). As a result, the IL-13-induced up-regulation of RhoA protein was slightly but not significantly augmented when the cells were transfected with miR-140-3p inhibitor (Figure 2: ‘Control RNA + IL-13’ versus ‘140–3p inhibitor + IL-13’). On the other hand, transfection of the cells with miR-140-3p mimic inhibited the RhoA up-regulation induced by IL-13, almost completely (Figure 2): the significant increase in RhoA induced by IL-13 (‘Control RNA’ versus ‘Control RNA + IL-13’) was not observed in the miR-140-3p-transfected cells (‘Control RNA’ versus ‘140–3p mimic + IL-13’).
Discussion
Our previous study revealed a decrease in the level of miR-140-3p in bronchial smooth muscles (BSMs) of a murine asthma model [25] that possesses BSM hyperresponsiveness together with up-regulations of RhoA protein and IL-13 [11,26]. Analyses using a public database tool suggested that RhoA mRNA is one of the targets of miR-140-3p [25]. The vector-based analyses also demonstrated that miR-140-3p interacts with 3ʹ-UTR of RhoA mRNA to negatively regulate the RhoA protein expression [25]. In the present study, inhibition of endogenous miR-140-3p by its synthetic inhibitor caused an up-regulation of RhoA protein in cultured hBSMCs (Figure 2). Treatment of hBSMCs with IL-13 caused an up-regulations of RhoA protein together with a down-regulation of miR-140-3p (Figure 1). The IL-13-induced up-regulation of RhoA protein was inhibited when the cells were transfected with miR-140-3p mimic (Figure 2). These observations indicate that the expression level of RhoA is negatively regulated by endogenous miR-140-3p and that IL-13 could induce RhoA up-regulation via inhibition of miR-140-3p expression at least in part.
Increasing evidence suggests that dysregulation of miRNA is involved in multiple human diseases including asthma [27,28]. Among the miRNA families, miR-140-3p has been suggested as a candidate miRNA that contributes to the dysfunction of ASM in asthma [21–25]. Kannan and colleagues demonstrated that miR-140-3p negatively controls the expression of CD38 [22], a regulatory protein of the ASM contractility [29,30]. The current study revealed that miR-140-3p also regulates the RhoA expression in BSM. It is thus possible that miR-140-3p controls the expression of several genes, such as CD38 and RhoA, in the ASM to regulate its contractility.
The ASM is a crucial effector tissue regulating bronchomotor tone. It has been suggested that modulation of ASM by inflammatory mediators such as cytokines plays an important role in the development of AHR [31–34]. Our previous findings that both the increased BSM contractility and the up-regulation of RhoA protein observed in the antigen-induced murine asthma model were reproduced by IL-13 [11] suggest that IL-13 is a key mediator for the induction of AHR. The IL-13-induced up-regulation of RhoA protein was inhibited both by a synthetic inhibitor and a small interfering RNA (siRNA) for STAT6 [26]. However, their inhibitory effects were only partial, whereas complete inhibition of STAT6 activity was observed [26], suggesting that some STAT6-independent mechanisms might also contribute to the IL-13-induced up-regulation of RhoA. We show here that IL-13 induced a down-regulation of miR-140-3p in hBSMCs (Figure 1(b)). Based on the current result that a miR-140-3p inhibitor caused an up-regulation of RhoA (Figure 2), the IL-13-induced down-regulation of miR-140-3p could be a cause of the IL-13-induced up-regulation of RhoA protein. Our preliminary observations that neither inhibitor nor mimic of miR-140-3p had effect on the mRNA expression of RhoA (data not shown) support an idea that miR-140-3p controls translation of RhoA, although further detailed studies are required.
In conclusion, the current findings suggest that RhoA protein expression is negatively regulated by miR-140-3p in hBSMCs. IL-13 is capable of reducing the miR-140-3p expression in BSMs. The reduced expression of miR-140-3p might cause an up-regulation of RhoA in BSM cells, resulting in an augmentation of the contraction, which is one of the causes of AHR in asthmatics.
Funding Statement
This work was partly supported by Grant-in-Aid for Scientific Research (C) Grant Number 15583093 (Y.C.) and 18988016 (M.H.) from the Japan Society for the Promotion of Science (JSPS).
Abbreviations
ACh acetylcholine
AHR airway hyperresponsiveness
ANOVA analysis of variance
ASM airway smooth muscle
BSM bronchial smooth muscle
cDNA complementary DNA
COPD chronic obstructive pulmonary disease
CT threshold cycle
GAPDH glyceraldehyde 3-phosphate dehydrogenase
hBSMC human bronchial smooth muscle cell
hEGF human epidermal growth factor
hFGF-b human fibroblast growth factor-basic
IL-13 interleukin-13
LNA locked nucleic acid
miRNA microRNA
mRNA messenger RNA
nt nucleotide
PCR polymerase chain reaction
PNA peptide nucleic acid
PVDF polyvinylidene difluoride
RT-PCR reverse transcriptase-polymerase chain reaction
RT-qPCR quantitative RT-PCR
SDS sodium dodecyl sulphate
siRNA small interfering RNA
snRNA small nuclear RNA
UTR untranslated region
Author contributions
Conception and design: Y.C., H.S. and M.H. Analysis and interpretation: Y.C., Y.A., Y.K. and H.S. Drafting the manuscript for important intellectual content: Y.C., H.S. and M.H. All authors provided final approval of the final version of the manuscript.
Disclosure of potential conflictof interest
The authors declare that they have no conflict of interest.
References
- [1].Seow CY, Schellenberg RR, Pare PD.. Structural and functional changes in the airway smooth muscle of asthmatic subjects. Am J Respir Crit Care Med. 1998;158(supplement_2):S179–86. [DOI] [PubMed] [Google Scholar]
- [2].Martin JG, Duguet A, Eidelman DH.. The contribution of airway smooth muscle to airway narrowing and airway hyperresponsiveness in disease. Eur Respir J. 2000;16(2):349–354. [DOI] [PubMed] [Google Scholar]
- [3].Sutcliffe A, Hollins F, Gomez E, et al. Increased nicotinamide adenine dinucleotide phosphate oxidase 4 expression mediates intrinsic airway smooth muscle hypercontractility in asthma. Am J Respir Crit Care Med. 2012;185(3):267–274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Somlyo AP, Somlyo AV. Signal transduction by G-proteins, rho-kinase and protein phosphatase to smooth muscle and non-muscle myosin II. J Physiol. 2012;522(2):177–185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Chiba Y, Takada Y, Miyamoto S, et al. Augmented acetylcholine-induced, Rho-mediated Ca 2+++ sensitization of bronchial smooth muscle contraction in antigen-induced airway hyperresponsive rats. Br J Pharmacol. 1999;127(3):597–600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Schaafsma D, Gosens R, Bos IS, et al. Allergic sensitization enhances the contribution of Rho-kinase to airway smooth muscle contraction. Br J Pharmacol. 2004;143(4):477–484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Chiba Y, Ueno A, Shinozaki K, et al. Involvement of RhoA-mediated Ca2+ sensitization in antigen-induced bronchial smooth muscle hyperresponsiveness in mice. Respir Res. 2005;6(1):4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Kudo M, Melton AC, Chen C, et al. IL-17A produced by αβT cells drives airway hyper-responsiveness in mice and enhances mouse and human airway smooth muscle contraction. Nat Med. 2015;18(4):547–554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Wei B, Shang YX, Li M, et al. Cytoskeleton changes of airway smooth muscle cells in juvenile rats with airway remodeling in asthma and the RhoA/ROCK signaling pathway mechanism. Genet Mol Res. 2014;13(1):559–569. [DOI] [PubMed] [Google Scholar]
- [10].Balenga NA, Klichinsky M, Xie Z, et al. A fungal protease allergen provokes airway hyper-responsiveness in asthma. Nat Commun. 2015;6(1):6763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Chiba Y, Nakazawa S, Todoroki M, et al. Interleukin-13 augments bronchial smooth muscle contractility with an up-regulation of RhoA protein. Am J Respir Cell Mol Biol. 2009;40(2):159–167. [DOI] [PubMed] [Google Scholar]
- [12].Chiba Y, Tanabe M, Goto K, et al. Down-regulation of miR-133a contributes to up-regulation of Rhoa in bronchial smooth muscle cells. Am J Respir Crit Care Med. 2009;180(8):713–719. [DOI] [PubMed] [Google Scholar]
- [13].Kudo M, Khalifeh Soltani SM, Sakuma SA, et al. Mfge8 suppresses airway hyperresponsiveness in asthma by regulating smooth muscle contraction. Proc Natl Acad Sci USA. 2013;110(2):660–665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Ambros V. The functions of animal microRNAs. Nature. 2004;431(7006):350–355. [DOI] [PubMed] [Google Scholar]
- [15].Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–297. [DOI] [PubMed] [Google Scholar]
- [16].Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136(2):215–233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Wu DD, Song J, Bartel S, et al. The potential for targeted rewriting of epigenetic marks in COPD as a new therapeutic approach. Pharmacol Ther. 2018;182:1–14. [DOI] [PubMed] [Google Scholar]
- [18].Stolzenburg LR, Harris A. The role of microRNAs in chronic respiratory disease: recent insights. Biol Chem. 2018;399:219–234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Tost J. A translational perspective on epigenetics in allergic diseases. J Allergy Clin Immunol. 2018;142(3):715–726. [DOI] [PubMed] [Google Scholar]
- [20].Svitich OA, Sobolev VV, Gankovskaya LV, et al. The role of regulatory RNAs (miRNAs) in asthma. Allergol Immunopathol (Madr). 2018;46(2):201–205. [DOI] [PubMed] [Google Scholar]
- [21].Kuhn AR, Schlauch K, Lao R, et al. MicroRNA expression in human airway smooth muscle cells: role of miR-25 in regulation of airway smooth muscle phenotype. Am J Respir Cell Mol Biol. 2010;42(4):506–513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Jude JA, Dileepan M, Subramanian S, et al. miR-140-3p regulation of TNF-α-induced CD38 expression in human airway smooth muscle cells. Am J Physiol Lung Cell Mol Physiol. 2012;303(5):L460–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Deshpande DA, Guedes AGP, Graeff R, et al. CD38/cADPR Signaling Pathway in Airway Disease: regulatory Mechanisms. Mediators Inflamm. 2018;2018:8942042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Specjalski K, Jassem E. MicroRNAs: potential biomarkers and targets of therapy in allergic diseases? Arch Immunol Ther Exp. 2019;67:213–223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Chiba Y, Ando Y, Fujii S, et al. Down-regulation of miR-140-3p is a cause of up-regulation of RhoA protein in bronchial smooth muscle of murine experimental asthma. Am J Respir Cell Mol Biol. 2021;64(1):138–140 [DOI] [PubMed] [Google Scholar]
- [26].Chiba Y, Todoroki M, Nishida Y, et al. A novel STAT6 inhibitor AS1517499 ameliorates antigen-induced bronchial hypercontractility in mice. Am J Respir Cell Mol Biol. 2009;41(5):516–524. [DOI] [PubMed] [Google Scholar]
- [27].Mousavi SR, Ahmadi A, Jamalkandi SA, et al. Involvement of microRNAs in physiological and pathological processes in asthma. J Cell Physiol. 2019;234:21547–21559. [DOI] [PubMed] [Google Scholar]
- [28].Taka S, Tzani-Tzanopoulou P, Wanstall H, et al. MicroRNAs in asthma and respiratory infections: identifying common pathways. Allergy Asthma Immunol Res. 2020;12:4–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Deshpande DA, Walseth TF, Panettieri RA, et al. CD38/cyclic ADP-ribose-mediated Ca2+ signaling contributes to airway smooth mus- cle hyper-responsiveness. Faseb J. 2003;17:452–454. [DOI] [PubMed] [Google Scholar]
- [30].Deshpande DA, White TA, Guedes AG, et al. Altered airway responsiveness in CD38-deficient mice. Am J Respir Cell Mol Biol. 2005;32:149–156. [DOI] [PubMed] [Google Scholar]
- [31].Amrani Y, Panettieri RA Jr.. Modulation of calcium homeostasis as a mechanism for altering smooth muscle responsiveness in asthma. Curr Opin Allergy Clin Immunol. 2002;2(1):39–45. [DOI] [PubMed] [Google Scholar]
- [32].Kardas G, Daszyńska-Kardas A, Marynowski M, et al. Role of platelet-derived growth factor (PDGF) in asthma as an immunoregulatory factor mediating airway remodeling and possible pharmacological target. Front Pharmacol. 2020;11:47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Fang L, Sun Q, Roth M. Immunologic and non-immunologic mechanisms leading to airway remodeling in asthma. Int J Mol Sci. 2020;21(3):E757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [34].Lam M, Lamanna E, Bourke JE. Regulation of airway smooth muscle contraction in health and disease. Adv Exp Med Biol. 2019;1124:381–422. [DOI] [PubMed] [Google Scholar]