

Abstract
Per- and polyfluoroalkyl substances (PFAS) comprise a diverse class of chemicals used in industrial processes, consumer products, and fire-fighting foams which have become environmental pollutants of concern due to their persistence, ubiquity, and associations with adverse human health outcomes, including in pregnant persons and their offspring. Multiple PFAS are associated with adverse liver outcomes in adult humans and toxicological models, but effects on the developing liver are not fully described. Here we performed transcriptomic analyses in the mouse to investigate the molecular mechanisms of hepatic toxicity in the dam and its fetus after exposure to two different PFAS, perfluorooctanoic acid (PFOA) and its replacement, hexafluoropropylene oxide-dimer acid (HFPO-DA, known as GenX). Pregnant CD-1 mice were exposed via oral gavage from embryonic day (E) 1.5–17.5 to PFOA (0, 1, or 5 mg/kg-d) or GenX (0, 2, or 10 mg/kg-d). Maternal and fetal liver RNA was isolated (N = 5 per dose/group) and the transcriptome analyzed by Affymetrix Array. Differentially expressed genes (DEG) and differentially enriched pathways (DEP) were obtained. DEG patterns were similar in maternal liver for 5 mg/kg PFOA, 2 mg/kg GenX, and 10 mg/kg GenX (R2: 0.46–0.66). DEG patterns were similar across all 4 dose groups in fetal liver (R2: 0.59–0.81). There were more DEGs in fetal liver compared to maternal liver at the low doses for both PFOA (fetal = 69, maternal = 8) and GenX (fetal = 154, maternal = 93). Upregulated DEPs identified across all groups included Fatty Acid Metabolism, Peroxisome, Oxidative Phosphorylation, Adipogenesis, and Bile Acid Metabolism. Transcriptome-phenotype correlation analyses demonstrated > 1000 maternal liver DEGs were significantly correlated with maternal relative liver weight (R2 >0.92). These findings show shared biological pathways of liver toxicity for PFOA and GenX in maternal and fetal livers in CD-1 mice. The limited overlap in specific DEGs between the dam and fetus suggests the developing liver responds differently than the adult liver to these chemical stressors. This work helps define mechanisms of hepatic toxicity of two structurally unique PFAS and may help predict latent consequences of developmental exposure.
Keywords: PFAS, Developmental exposure, Liver disease, Emerging contaminants, Animal models
1. Introduction
Per- and polyfluoroalkyl substances (PFAS) are chemicals recently defined by the Organisation for Economic Co-operation and Development (OECD) as “any chemical with at least a perfluorinated methyl group (–CF3) or a perfluorinated methylene group (–CF2−)”, making this a family of over 12,000 unique chemical structures and counting (OECD, 2021; Wang et al., 2021) (see: https://comptox.epa.gov/dashboard/). Some well-studied chemicals in the broad universe of PFAS have been termed “forever chemicals” due to their persistence and bioaccumulation in humans, wildlife, and the environment (Sunderland et al., 2019). Exposure to several PFAS (primarily the legacy PFAS, PFOA and PFOS) is associated with adverse health outcomes across a variety of tissue systems and life stages in humans, including developmental deficits, disease of the liver, thyroid, and kidney, metabolic changes, obesity, immune suppression, and cancer, among others (Fenton et al., 2021). Exposure to these PFAS is positively and consistently associated with serum dyslipidemia, including increased total cholesterol and low-density lipoprotein cholesterol, in both occupational and non-occupational cohorts (ATSDR, 2020; Fenton et al., 2021), and in children (Blomberg et al., 2021). Another consistent adult health effect associated with exposures to these PFAS is adverse liver outcomes, such as fatty liver biomarkers, clinical biomarkers of liver damage, non-hepatitis liver disease, enlarged liver, cirrhosis, and steatosis (Fenton et al., 2021; Qi et al., 2020; Sen et al., 2022; Costello et al., 2022).
Some PFAS have been evaluated for their ability to cross the placenta during pregnancy as well as their distribution and accumulation in human fetal tissues. Mamsen and colleagues reported increasing accumulation of the PFAS they tested, in general, with increasing gestational age and accumulation of perfluorononanoic acid (PFNA), perfluorooctanoic acid (PFOA), and perfluorooctanesulfonic acid (PFOS) in the placenta and liver of second and third trimester fetuses (Mamsen et al., 2017, 2019). In recent studies, a number of PFAS measured in mothers’ serum during pregnancy were shown to be associated with elevated liver enzyme levels and increased liver injury risk in their 6–9 year old children (Stratakis et al., 2020) and higher PFAS serum levels was associated with more severe non-alcoholic fatty liver disease (NAFLD) in children aged 7–19 years (Jin et al., 2020).
Although evidence for PFAS-associated liver disease is limited in children, adult evidence is mounting. The strongest evidence for PFAS causing “Toxicant-Associated Fatty Liver Disease” (Qi et al., 2020) exists in rodent models (Costello et al., 2022). In rodents, NAFLD is associated with the inhibition of fatty acid β-oxidation, increased fatty acid uptake, and induction of oxidative stress. Numerous studies have shown that developmental exposure to relatively low doses (0.01–1 mg/kg-day) of perfluorooctanoic acid (PFOA) induces adverse metabolic health outcomes in multiple strains of mice. Evidence includes increased weight gain, increased serum leptin and insulin, increased fat mass, hepatocellular hypertrophy, and in some cases liver lesions, compared to vehicle controls (Filgo et al., 2015; Hines et al., 2009; Quist et al., 2015; Roth et al., 2021; van Esterik et al., 2016). Due to an abundance of evidence linking PFOA to detrimental developmental, reproductive, and systemic effects, PFOA was phased out of production in the United States beginning in 2006 (U.S. EPA, 2016), and alternative chemistry PFAS began being used in place of it.
Hexafluoropropylene oxide dimer acid (HFPO-DA) is a replacement PFAS for PFOA and one of the constituents of a specific processing aid technology called GenX, though HFPO-DA itself is now commonly referred to as “GenX”. GenX is structurally distinct from PFOA; PFOA is an 8-carbon perfluoroalkyl carboxylic acid whereas GenX is a 5-carbon perfluoropolyether carboxylic acid. GenX has not been as well studied as PFOA, but limited data have shown that GenX exposure can cause altered lipid metabolism, increased neonatal mortality rate, decreased birth weight, and increased maternal liver weight in rodents (Blake et al., 2020; Conley et al., 2019; Conley et al., 2021; Dupont Chem, 2010a; b). More recently, a constellation of liver lesions and metabolic disease was observed in mouse offspring exposed to GenX in utero (Cope et al., 2021). In the recent U.S. EPA human health toxicity value assessment for GenX chemicals, a similar constellation of liver lesions was observed across multiple animal studies and was the selected endpoint supporting the final toxicity values (U.S. EPA, 2021). Despite the structural and half-life differences between GenX (t ½ = 16–20 h in mice) and PFOA (t ½ = 18–22 d in mice) (Fenton et al., 2021), and the reported lack of liver bioaccumulation of GenX-exposed mice (Blake et a, 2020), both chemicals induced similar effects on the liver and the metabolic system.
NAFLD is an inflammatory disease defined by an accumulation of excessive fat in the liver that is closely associated with the development of metabolic disease and insulin resistance (Mitra et al., 2020). It has recently been suggested to rename NAFLD to metabolic dysfunction-associated fatty liver disease (MALFD); this is to more accurately represent the complex pathophysiology of this multisystem disorder (Eslam et al., 2020). With a global prevalence as high as one billion (25 %) and affecting between 80 and 100 million individuals in the United States, NAFLD is a growing global public health issue (Perumpail et al., 2017). In addition to being closely associated with Type 2 Diabetes, ~ 15 % of NAFLD patients progress to more severe liver complications including hepatocellular carcinoma (Mantovani et al., 2020). Further, non-alcoholic steatohepatitis (NASH), an additional complication caused by NAFLD, is the second leading cause of liver transplantation in the U.S. (Mantovani et al., 2020). Of growing concern, the NAFLD rate in children and adolescents is increasing rapidly (Welsh et al., 2013). Significant weight gain during the pubescent period carries a greater risk for developing NAFLD compared to adulthood, supporting the concept of unique life stage sensitivities to NAFLD and metabolic disease in early life. Though genetic risk contributes to the development of NAFLD, mounting evidence has suggested a link between prenatal exposure to environmental chemicals, possibly including PFAS, and the onset of NAFLD (Foulds et al., 2017). In fact, a recent systematic review evaluating PFAS exposure and evidence of liver injury found consistent evidence for PFAS hepatotoxicity in rodent studies and positive associations of PFAS and markers of liver function in observational human studies (Costello et al., 2022).
Considering the importance of the maternal liver in energy partitioning during pregnancy (Bauman and Currie, 1980), we hypothesized that PFAS-induced alterations in hepatic signaling within the maternal liver would be similar to altered pathways in the fetal liver. To better understand the mechanistic pathways involved in the hepatotoxic effects induced by PFOA and GenX, we used transcriptome-wide analysis to compare differentially regulated genes and pathways in matched maternal and fetal livers collected in the Blake et al. (2020) study. Because liver phenotypes were comparable between adult F0 and F1 mice gestationally exposed to PFOA or GenX in our previous studies (Blake et al., 2020; Cope et al., 2021), we hypothesized that PFOA and GenX disrupt similar biological pathways that ultimately lead to liver disease.
2. Methods
2.1. Animals
All tissues were obtained during experiments described in Blake et al. (2020), which were approved by the NIEHS Animal Care and Use Committee (ASP #2017–0022). Briefly, mice were bred in-house during single overnight cohabitation and copulatory plug-positive mice were identified as gestational day (GD) 0.5. Pregnant CD-1 mice from the NIEHS colony were singly housed in humidity (45–60 %) and temperature (25 °C)-controlled 12-h light cycle rooms in polypropylene cages with ad libitum access to NIH-31 diet and reverse osmosis deionized (RODI) water. Pregnant dams (2 blocks of equal size and total N = 10–13 per group) were exposed to 1 or 5 mg/kg perfluorooctanoic acid ammonium salt (PFOA, CAS #3825-26-1, Millipore Sigma), 2 or 10 mg/kg GenX (ammonium 2,3,3,3-tetrafluoro-2- (heptafluoropropoxy) propanoate, CAS# 62,037–80–3, SynQuest Laboratories), or vehicle (RODI water) via daily oral gavage from GD1.5 to GD17.5. CD-1 mice were selected for this work as they are an outbred strain and thus more representative of the genetic diversity of the human population, they produce large litters which requires the use of fewer animals to achieve the appropriate level of statistical power and thus aligns with the “reduction” aspect of the three “R’s” of animal research (replacement, reduction, refinement). CD-1 mice are more sensitive to PFAS than other strains and thus more health-protective in the context of human health risk assessment (Macon et al., 2011), and they have been previously used in toxicological studies of PFOA and GenX in support of human health risk assessment (U.S. EPA, 2016, 2021).
Two doses of PFOA were selected; a lower dose of 1 mg/kg/day and a higher dose of 5 mg/kg/day which has previously been shown to reduce birth weight in CD-1 mice (Lau et al., 2006; White et al., 2007). Based on the two-fold difference between the U.S. EPA 70 ppt PFOA drinking water health advisory level (U.S. EPA, 2016) and the provisional North Carolina state health goal for GenX of 140 ppt (NCDEQ/NCDHHS, 2018), we used 2 mg/kg/day and 10 mg/kg/day GenX as “PFOA-equivalent” doses. Due to the different rates at which PFOA and GenX are excreted by CD-1 mice, our goal was to select oral doses that would result in comparable serum concentrations so that measured endpoints could be compared based on the bioavailable dose. As reported in our prior work (Blake et al., 2020), the orally administered doses of PFOA and GenX resulted in comparable levels of each compound in maternal serum at gestational day 17.5 (1 mg/kg/day PFOA: 18.7 ± 3.2 μg PFOA/mL serum; 2 mg/kg/day GenX: 22.9 ± 17.1 μg GenX/mL serum; 5 mg/kg/day PFOA: 95.1 ± 14.1 μg PFOA/mL serum; 10 mg/kg/day GenX: 58.5 ± 34.6 μg GenX/mL serum).
2.2. Necropsy
A comprehensive overview of the necropsy procedures can be found in Blake et al. (2020). Briefly, on gestational day 17.5 pregnant dams were weighed and humanely euthanized by swift decapitation. Trunk blood was collected, and serum was used for clinical chemistry. Maternal livers were weighed, and a portion of the left lateral lobe was snap frozen in liquid nitrogen for subsequent RNA extraction. The uterus was removed, and viable embryos were collected in succession within a uterine horn and the livers were removed and immediately snap frozen in liquid nitrogen (n = 2–5 embryos/litter). A portion of fetal tissue corresponding to each snap frozen liver was reserved and genotyped by PCR amplification of the sex determining region-Y (SRY) gene (F: 5′ GCTTCAGTAATCTCAGCACCTAGAA 3′, R: 3′ CACATTGGCATGATAGCTCCAAATT 5′) to determine whether a fetus was genotypically male or female (TransnetYX, Inc). Only female fetal livers were used in this study to control for the potential impact of sex on the chemical exposure effects between maternal and fetal tissue due to the expansive sexual dimorphism in the organ (Lefebvre and Staels, 2021).
2.3. RNA isolation and quantification
Matching maternal and female fetal liver samples were used for this study. Five dams from the same block were randomly chosen for this evaluation. Fetal livers were selected at random when there was more than one female per litter collected. A total of N = 4–6 fetal liver per treatment group were used in the current study. Snap frozen tissues were trimmed on ice and weighed (< 30 mg) and then homogenized using a FastPrep-24™ 5 G (MP Biomedicals, Valiant Co., Ltd, China) in Buffer RLT (Qiagen, Hilden, Germany) within 2 mL, 2.4 mm metal bead homogenization tubes (Thermo Fisher Scientific, Waltham, MA). Homogenized liver samples were then snap frozen and stored at − 80 °C before use. RNA was extracted from maternal and fetal liver samples using RNeasy kits following manufacturer’s protocols (Qiagen, Hilden, Germany). Resulting purified RNA was subjected to quality control assessments using a NanoDrop™ 2000/2000c Spectrophotometer (Thermo Fisher Scientific, Waltham, MA) and Bioanalyzer High Sensitivity RNA Analysis (Agilent, Santa Clara, CA). All RNA samples used for transcriptomics analysis had a 260/280 ratio > 2.0 and RIN > 5.0. RNA isolates were diluted to a standard concentration of 100 ng RNA/ 1 μL H2O. RNA expression analysis was conducted using Affymetrix Mouse Clariom D arrays (Affymetrix, Santa Clara, CA). Total RNA (25 ng/sample) was first amplified and labeled using the Affymetrix WT Plus Reagent Kit (WT Plus Kit) per manufacturer’s instructions. Amplified biotin-cDNAs (5 μg/sample) were fragmented and hybridized to each array for 16 h at 45 °C in a rotating hybridization oven. Array slides were stained with streptavidin/phycoerythrin utilizing a double-antibody staining procedure and then washed for antibody amplification per the GeneChip Hybridization, Wash and Stain Kit user manual. Arrays were scanned in an Affymetrix Scanner 3000 and data was obtained using Transcriptome Analysis Console Software (Affymetrix, Santa Clara, CA).
2.4. Gene expression data analyses
A summary of the transcriptomic analysis workflow can be found in Fig. S1 and raw data are accessible on NCBI GEO (accession number GSE199233). The probe level raw intensity signal from the Affymetrix Mouse Clariom D Array (previously known as the Mouse Transcriptome Assay 1.0) was extracted. For each transcript these raw intensities were background adjusted, summarized using robust multi-array average (RMA) technique, and log2 transformed. The normalized expression was averaged across transcripts from the same gene to produce Entrez gene-level signal. Potential outliers were identified using a combination of approaches including principal component analysis, hierarchical cluster plots, and correlation plots, and none were detected in the current study (Fig. S2). We utilized Student’s t-test statistics to measure gene-level differential activity. A customized implementation of conventional Gene Set Enrichment Analysis (GSEA) (Subramanian et al., 2005) was employed to simultaneously identify differentially expressed genes (DEGs) and corresponding differentially enriched pathways (DEPs) for each chemical at each dose relative to the corresponding maternal or fetal vehicle control. Student’s t-test statistics were used to measure gene-level differential activity and perform GSEA tests on all Hallmark pathways from the Molecular Signature Database (MSigDB, version 6.2) for which five or more genes were present in the microarray (Liberzon et al., 2015; Subramanian et al., 2005). A total of 50 pathways were used for the gene set enrichment analysis. The significant p-values, false detection rate, and familywise error rate (FWER) for gene/pathway level activity were computed using 10,000 random permutations of sample labels. False detection rate and FWER calculations were performed separately for all pathways. For a gene to be considered a significant DEG, it was required to have an absolute fold change ≥ 2 and a distribution p-value ≤ 0.005. Similarly, a significant DEP required an absolute Normalized Enrichment Score (NES) ≥ 1.5 and a NES p-value ≤ 0.05. Matthews correlation coefficient was computed pairwise between each treatment group to assess overlap of each DEG list (Gorodkin, 2004; Matthews, 1975).
To further characterize the pathway level differences between PFOA and GenX and maternal and fetal tissue, gene-level results (fold changes and p-values) were imported into Ingenuity Pathway Analysis (IPA) software (Qiagen, Hilden, Germany) and analyzed using the Ingenuity Analysis Knowledgebase. The most significantly enriched gene networks and canonical pathways were identified using transcripts with p-value ≤ 0.005 and an absolute expression fold-change ≥ 2. Custom gene interaction networks were created to examine links between specific genes of interest and related pathways.
We then searched the IPA knowledgebase to identify genes associated with specific Hallmark Pathways that were previously determined from the MsigDB with significant levels of enrichment across multiple dose groups for both maternal and fetal liver, including those relating to peroxisomes (e.g. oxidation of fatty acids, peroxisome organization, formation, and proliferation), cholesterol and bile acid metabolism (e.g. synthesis, metabolism, and transport), and oxidative phosphorylation (e.g. lipolysis, lipid transport, fatty acid oxidation). The gene lists were then filtered to include only those identified as significant DEGs within our dataset. Diseases and biological functions associated with the gene lists and relevant to liver function were overlaid to construct custom gene networks, and the molecule activity predictor (MAP) tool was used to predict the activation state for the selected diseases and bio functions. To determine whether potential common transcriptional regulators were similarly activated or inhibited across different doses and tissues, IPA Upstream Regulator Analysis was used to identify the top 20 predicted upstream regulators (absolute z-score ≥2 and a p-value <0.05 were considered significant; Kräme et al., 2014).
2.5. Phenotypic correlation analysis
Gene expression data were correlated with phenotypic endpoints for maternal liver obtained during the same set of experiments and previously published in Blake et al. (2020). These endpoints included absolute and relative maternal liver weight, relative maternal weight gain from gestational day 0.5–11.5, relative maternal weight gain from gestational day 0.5–17.5, and maternal serum levels of albumin (ALB), alkaline phosphatase (ALP), alanine aminotransferase (ALT), aspartate aminotransferase (AST), triglycerides, high density lipoprotein (HDL), low density lipoprotein (LDL), cholesterol, total protein (TP), creatinine, glucose, and total bile acid (TBA) (see Blake et al., 2020 for a detailed description).
Phenotypic data from Blake et al. (2020) were subset to include only maternal (dam) liver samples that were in the transcriptomics dataset. First, phenotypic endpoints (listed above) were assessed within this subset to compare the 5 groups (vehicle control, GenX low dose (2 mg/kg-d GenX), GenX high dose (10 mg/kg-d GenX), PFOA low dose (1 mg/kg-d PFOA), and PFOA high dose (5 mg/kg-d PFOA). To directly compare each treatment group to the control group, the many-to-one (treatment groups to control group) Dunn Test was performed using a standard normal distribution. Endpoints with a treatment vs control p-value ≤ 0.05 were considered significant.
To assess the relationship between differential gene expression and phenotype, a correlation analysis was performed. Pearson’s product moment correlation test was performed for each gene/phenotype combination to measure the degree of linear dependence between expression and phenotype. Unadjusted and adjusted (to control the false discovery rate) p-values were calculated (Benjamini and Hochberg, 1995). Genes/phenotypes with correlations ≥ 0.8 and adjusted p-values ≤ 0.05 were considered significant.
2.6. Quantitative PCR validation of transcriptomics
Matching maternal and female fetal liver RNA samples were used to validate the transcriptomics analysis. RNA (1 μg) was transcribed into cDNA using High-Capacity cDNA synthesis kit (Applied Biosystems, Waltham, MA). Complimentary DNA (12.5 ng) was amplified for genes of interest in a QuantStudio 7 Flex Real Time PCR System (Applied Biosystems, Waltham, MA) using PowerUP SYBR Green 2x mastermix (Applied Biosystems, Waltham, MA). Primer sets for genes of interest were designed assays from IDT DNA (Coralville, IA) and are presented in the supplemental materials section (Table S1). Relative mRNA levels were calculated by using ΔΔCt method and presented as fold change over respective vehicle control. Data is presented as fold change over vehicle control. One-way ANOVA, with Dunnett’s multiple comparisons post hoc test (GraphPad Prism v.7), was performed to determine statistical significance between control and dose groups using ΔCt values (p-value <0.05) and 2-fold change or greater changes are presented as mean fold change for n = 4–5, biological replicates, and visualized as a heatmap and p-value table.
3. Results
3.1. Maternal liver gene expression
Transcriptomics were conducted on RNA isolated from maternal liver after daily oral exposure from gestational day 1.5–17.5 to two doses of PFOA (1 mg/kg or 5 mg/kg) or GenX (2 mg/kg or 10 mg/kg), relative to vehicle control, to compare the effects of the two compounds relative to vehicle control. In general, exposure to PFOA or GenX resulted in a greater number of significantly upregulated differentially expressed genes (DEGs) than downregulated DEGs. Across the four exposure conditions, 5 mg/kg/day PFOA induced the greatest number of DEGs in maternal liver (N = 341; Table 1). Compared to 5 mg/kg/day PFOA, the 10 mg/kg/day GenX exposure resulted in fewer DEGs (N = 253), while 2 mg/kg/day GenX exposure resulted in a considerably greater number of DEGs (N = 93) relative to 1 mg/kg/day PFOA (N = 8; Table 1). A total of 407 unique genes were significantly differentially regulated in at least one experimental condition in maternal liver (Fig. S3).
Table 1.
Number of significant differentially expressed genes (DEGs) in maternal and fetal liver following exposure to PFOA or GenX.
| 1 mg/kg | 5 mg/kg | 2 mg/kg | 10 mg/kg | ||
|---|---|---|---|---|---|
| PFOA | PFOA | GenX | GenX | ||
| Maternal Liver | Up-regulated | 7 | 233 | 77 | 200 |
| Down-regulated | 1 | 108 | 16 | 53 | |
| Total | 8 | 341 | 93 | 253 | |
| Fetal Liver | Up-regulated | 65 | 107 | 117 | 129 |
| Down-regulated | 4 | 22 | 37 | 41 | |
| Total | 69 | 129 | 154 | 170 |
Note: DEGs were considered significant if absolute fold-change values were ≥ 2 and p ≤ 0.005.
Significant DEGs were most correlated between the 10 mg/kg GenX, 2 mg/kg GenX, and 5 mg/kg PFOA dose groups in the maternal liver for significant DEGs (R2: 0.46–0.66; Fig. 1). Across those three dose groups, 73 genes were upregulated and 7 genes were downregulated (Fig. 2A, Fig. 2B). The magnitude and direction of expression changes across these 80 genes tended to be most similar between the high doses (5 mg/kg PFOA and 10 mg/kg GenX) as well as similar between the low doses (1 mg/kg PFOA and 2 mg/kg GenX; Fig. 2C).
Fig. 1.
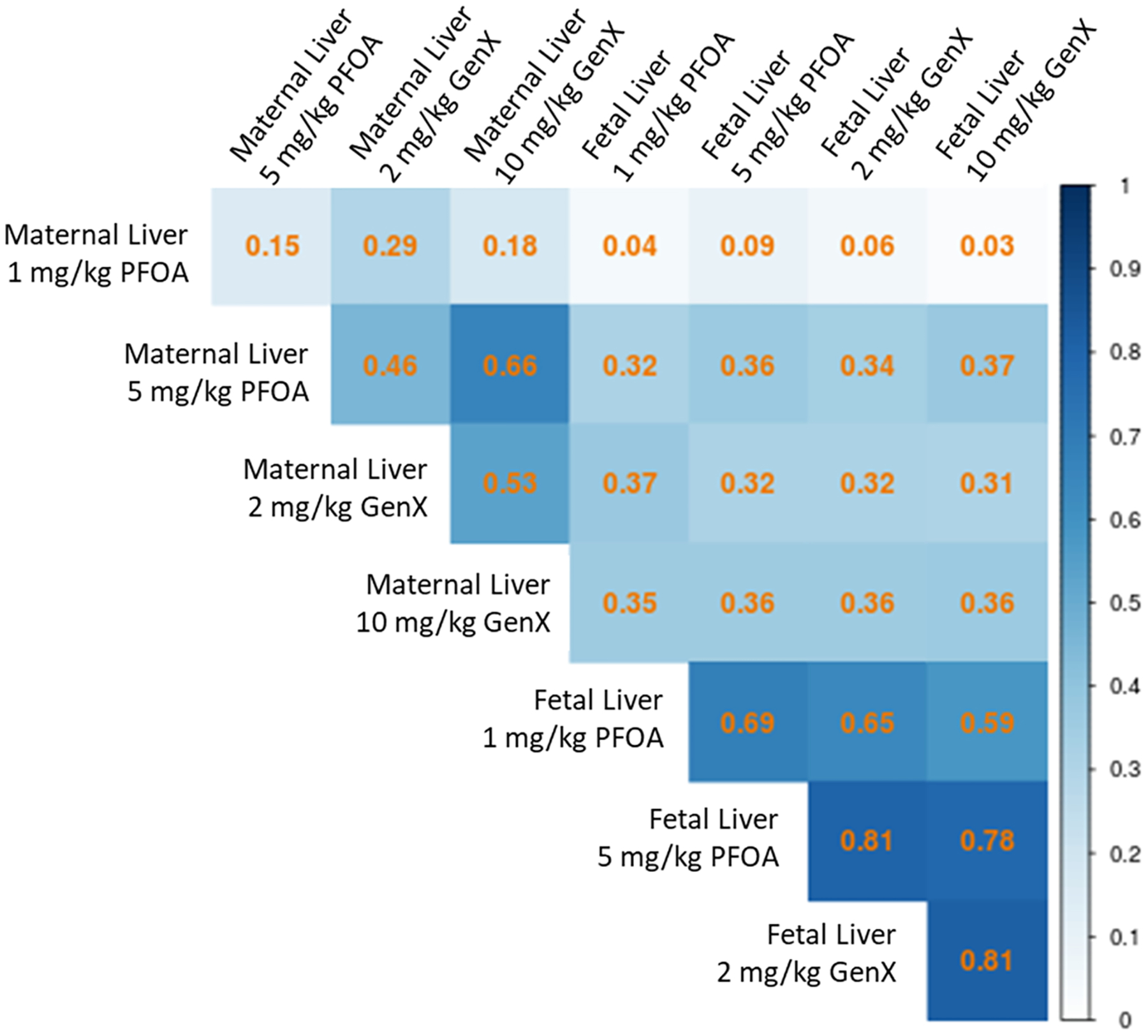
Matthews correlation coefficient plot demonstrating similarity between significant DEGs (shown in Table 1) across the four dose conditions for maternal and fetal liver. Gene expression changes tended to be more similar across dose groups in fetal liver (0.59–0.81) than across dose groups in maternal liver (0.15–0.66). Correlation coefficients between maternal and fetal liver across the dose conditions were least similar when comparing fetal dose groups to maternal 1 mg/kg PFOA (0.03–0.09) whereas all other correlation permutations were comparable (0.31–0.37).
Fig. 2.

Differentially expressed genes (DEGs) in maternal liver after gestational exposure to PFOA (1 and 5 mg/kg) or GenX (2 and 10 mg/kg). (A) Overlapping upregulated DEGs and (B) overlapping downregulated DEGs are shown in the Venn diagrams. (C) Heatmap showing row z-score of the normalized expression values for 80 genes significantly altered in the 5 mg/kg PFOA, 2 mg/kg GenX, and 10 mg/kg GenX exposure groups where each column represents an individual animal to highlight biologic variability within a given treatment group. Three non-coding genes were identified and include Gm11844, Gm15441, and Mir292b. DEGs were considered significant if absolute fold-change values were ≥ 2 and p ≤ 0.005.
3.2. Fetal liver gene expression
Transcriptomics were also conducted on female fetal liver samples after indirect (in utero) exposure to two doses of PFOA or GenX relative to control, to compare the effects of the two compounds on the developing liver. Significant DEGs were highly correlated when comparing across fetal liver dose groups (R2: 0.59–0.81; Fig. 1). In comparison, DEG correlations across maternal liver dose groups, were much less robust (R2: 0.15–0.66; Fig. 1), with the highest correlation between the 5 mg/kg PFOA and 10 mg/kg GenX groups. DEG correlations were the weakest when comparing maternal liver dose groups to fetal liver dose groups (R2: 0.03–0.37; Fig. 1), suggesting limited overlap between maternal liver DEGs and fetal liver DEGs. Compared to the maternal liver, there were fewer genes DEGs in fetal liver following exposure to the high doses of PFOA and GenX (Table 1). However, for both the low doses of PFOA and GenX, there was a greater number of DEGs compared to the maternal liver (Table 1). In the fetal liver, the high dose of GenX induced the greatest number of DEGs (total of 170), followed by the low dose of GenX (total of 154), the high dose of PFOA (total of 129), and the low dose of PFOA (total of 69; Table 1). Across all four experimental groups, a total of 202 shared genes were differentially expressed in at least one group in fetal liver (Fig. S4).
Examination of the shared up- and downregulated genes across all four fetal exposure groups revealed 60 upregulated and 1 down-regulated DEG (Fig. 3A, Fig. 3B). The relative fold changes across these 61 genes were highly similar across the PFAS exposures, as visualized by heatmapping (Fig. 3C). This similarity between the high and low dose exposures in the fetus were especially striking when compared to the common DEGs identified in the maternal liver, which exhibited more dose-specific responses (Fig. 2C).
Fig. 3.
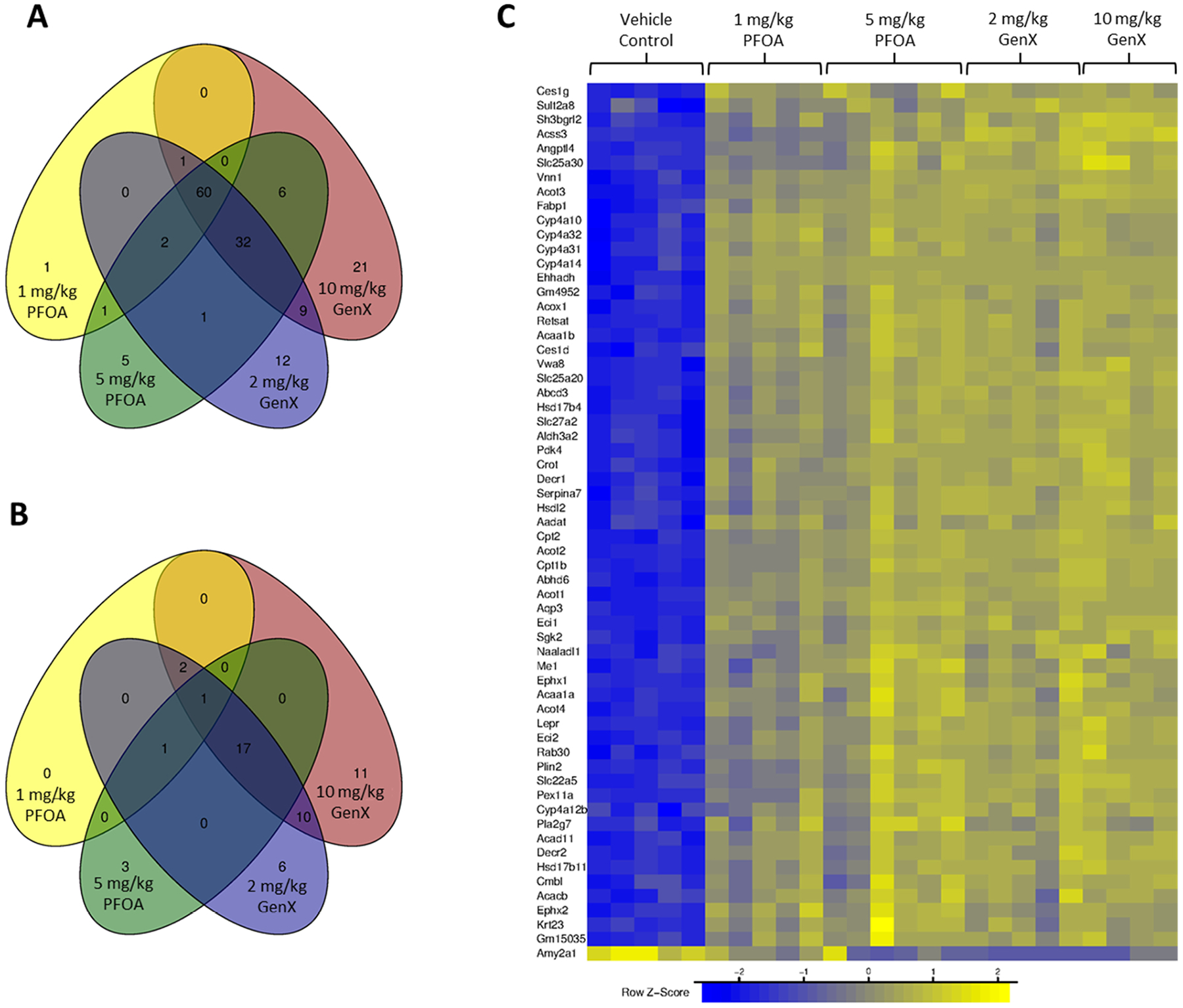
Differentially expressed genes (DEGs) in fetal liver after in utero exposure to PFOA (1 and 5 mg/kg) or GenX (2 and 10 mg/kg). (A) Overlapping upregulated DEGs and (B) overlapping downregulated DEGs are shown in the Venn diagrams. (C) Heatmap showing row z-score of the normalized expression values for 61 genes significantly altered in altered in all four exposure groups where each column represents an individual animal to highlight biologic variability within a given treatment group. One non-coding gene, Gm15035, was identified as a DEG across all treatment groups and one additional non-coding gene, 1300002E11Rik, was identified as a DEG across all treatment groups except 1 mg/kg PFOA. DEGs were considered significant if absolute fold-change values were ≥ 2 and p ≤ 0.005.
3.3. Shared coding and non-coding DEGs
To further examine the shared mechanisms of liver toxicity in the maternal and developing liver, common coding and non-coding DEGs were identified and compared. There were 30 significant DEGs across all conditions in maternal and fetal liver (except maternal 1 mg/kg PFOA which had very few DEGs; Table 2). The functions of these genes primarily correspond to the metabolism, peroxidation, synthesis, transport, and regulation of fatty acids and lipids. Other common DEGs include those that contribute to glucose metabolism through their action in the mitochondrial matrix (pyruvate dehydrogenase kinase 4, Pdk4) and regulation of peroxisome maintenance and proliferation (peroxisomal biogenesis factor 11 alpha, Pex11a).
Table 2.
Fold-change in 30 genes with significant differential expression in all maternal and fetal liver dose groups except maternal liver 1 mg/kg PFOA.
| Gene Symbol | Maternal Liver | Maternal Liver | Maternal Liver | Maternal Liver | Fetal Liver | Fetal Liver | Fetal Liver | Fetal Liver |
|---|---|---|---|---|---|---|---|---|
| 1 mg/kg PFOA | 5 mg/kg PFOA | 2 mg/kg GenX | 10 mg/kg GenX | 1 mg/kg PFOA | 5 mg/kg PFOA | 2 mg/kg GenX | 10 mg/kg GenX | |
| Cyp4a12b | ns | 136.1 | 28.6 | 97.5 | 3.7 | 5.7 | 5.6 | 6.5 |
| Cyp4a14 | ns | 18.6 | 15.5 | 17.4 | 35.3 | 38.7 | 38.7 | 38.5 |
| Pdk4 | ns | 26.9 | 14.8 | 27.7 | 4.3 | 5.3 | 5.7 | 6.4 |
| Acot1 | ns | 13.5 | 7.8 | 11.8 | 7.0 | 9.9 | 9.3 | 9.4 |
| Retsat | ns | 9.4 | 7.0 | 9.3 | 3.2 | 3.5 | 3.4 | 3.3 |
| Acot2 | ns | 12.2 | 6.2 | 10.3 | 7.2 | 12.4 | 13.2 | 13.8 |
| Serpina7 | ns | 8.6 | 5.8 | 8.6 | 4.7 | 6.6 | 6.9 | 7.2 |
| Acot3 | ns | 6.4 | 4.8 | 6.0 | 5.9 | 8.4 | 9.8 | 10.9 |
| Ehhadh | 2.6 | 5.8 | 4.7 | 5.5 | 9.9 | 11.5 | 11.4 | 11.7 |
| Vnn1 | ns | 6.8 | 4.7 | 6.0 | 8.1 | 11.0 | 13.3 | 13.9 |
| Cyp4a32 | ns | 6.6 | 4.6 | 5.8 | 12.6 | 12.9 | 11.5 | 11.0 |
| Acaa1b | ns | 5.8 | 4.2 | 5.2 | 4.9 | 6.1 | 6.1 | 6.1 |
| Hsd17b11 | ns | 5.6 | 4.0 | 6.0 | 2.5 | 2.9 | 3.2 | 3.7 |
| Pex11a | ns | 7.3 | 3.8 | 6.2 | 2.9 | 4.6 | 4.4 | 4.8 |
| Cyp4a10 | ns | 4.1 | 3.6 | 4.0 | 16.7 | 16.3 | 15.1 | 14.4 |
| Cyp4a31 | ns | 4.6 | 3.5 | 4.1 | 11.3 | 13.3 | 12.8 | 11.4 |
| Acot4 | ns | 5.7 | 3.2 | 4.9 | 3.0 | 4.3 | 3.7 | 3.8 |
| Aldh3a2 | ns | 4.0 | 2.9 | 3.7 | 2.2 | 2.6 | 2.8 | 2.8 |
| Slc22a5 | ns | 5.4 | 2.6 | 4.2 | 2.1 | 3.1 | 3.3 | 3.5 |
| Eci1 | ns | 3.3 | 2.5 | 3.3 | 2.8 | 3.1 | 3.4 | 3.6 |
| Vwa8 | ns | 3.0 | 2.4 | 3.0 | 2.7 | 3.1 | 3.2 | 3.5 |
| Eci2 | ns | 3.1 | 2.3 | 2.9 | 2.3 | 2.8 | 2.9 | 3.1 |
| Acox1 | ns | 3.2 | 2.3 | 3.0 | 3.2 | 3.6 | 3.4 | 3.7 |
| Ephx2 | ns | 3.0 | 2.3 | 2.8 | 3.1 | 3.3 | 2.6 | 2.9 |
| Slc25a20 | ns | 3.1 | 2.3 | 2.9 | 2.9 | 3.5 | 3.8 | 4.1 |
| Decr2 | ns | 2.3 | 2.1 | 2.6 | 2.1 | 2.1 | 2.2 | 2.4 |
| Decr1 | ns | 2.5 | 2.1 | 2.5 | 2.1 | 2.3 | 2.6 | 2.7 |
| Hsdl2 | ns | 2.6 | 2.1 | 2.5 | 2.1 | 2.6 | 2.8 | 3.0 |
| Acaa1a | ns | 2.9 | 2.0 | 2.6 | 2.1 | 2.6 | 2.5 | 2.4 |
| Cpt2 | ns | 2.7 | 2.0 | 2.6 | 2.5 | 3.3 | 3.4 | 3.5 |
Note: DEGs were considered significant if absolute fold-change values were ≥ 2 and p ≤ 0.005. Abbr: ns = not significant.
When comparing non-coding genes, there was less overlap between maternal and fetal liver and the four treatment conditions. To examine non-coding gene expression changes, non-coding genes that were significantly differentially expressed in at least one of the four treatment groups in maternal and fetal liver were included, resulting in 24 unique genes (Table S2).
3.4. Validation of gene expression changes
Subsequent to the transcriptomics analysis, a set of genes were chosen for validation by qPCR. Genes selected for validation met one of the following criteria: strong increase (e.g., cytochrome P450 A12B, Cyp4a12b) or decrease (e.g., carboxylesterase 1 C, Ces1c) in expression, or expression patterns that differed in intensity of gene expression changes when comparing maternal and fetal liver (e.g., cytochrome P450 aA14, Cyp4a14; flavin containing monooxygenase 3, Fmo3). Two additional genes were selected that were either not expressed in the female liver (testis expressed gene 101, Tex101) or were not significantly changed in the study (zinc finger protein 472, Zfp472). A 97 % concordance between the qPCR expression data and the transcriptomics data was confirmed for data points within defined thresholds (qPCR: 2-fold change, p-value<0.05; transcriptomics: 2-fold change, p-value<0.005). One hundred percent concordance for directionality and 93 % concordance for statistically changed genes exhibiting similar expression patterns were achieved (Fig. S5).
3.5. Pathway analysis in maternal and fetal liver
Pathway enrichment analyses revealed numerous MSigDB Hallmark Pathways with significant normalized enrichment scores (NES ≥ 1.5, p ≤ 0.05; Fig. 4, Fig. S6). For maternal liver, 22 hallmark pathways were significantly enriched by at least one exposure condition and 5 hallmark pathways were significantly enriched by all four exposure conditions: Bile Acid Metabolism, Peroxisome, Fatty Acid Metabolism, Adipo-genesis, and Oxidative Phosphorylation (Table S3). For fetal liver, 19 pathways were significantly enriched by at least one exposure condition and 11 pathways were significantly enriched by all four exposure conditions (all shown in Table S4). All 5 significantly enriched hallmark pathways identified in maternal liver across all four treatment groups were also significantly enriched in fetal liver across all four treatment groups (Fig. 4). IPA Upstream Regulator Analysis predicted upstream regulators that were common across most maternal and fetal dose groups and included regulators involved in regulation of liver gluco-neogenesis (Kruppel-like factor 15, KLF15; nuclear receptor interacting protein 1, NRIP1), energy metabolism and mitochondrial biogenesis (estrogen related receptor alpha, ESRRA), lipid metabolism and beta-oxidation (mediator complex subunit 1, MED1; B cell lymphoma 6, BCL6) and hepatocellular carcinoma (signal transducer and activator of transcription 6, STAT6; STAT5B; Table S5).
Fig. 4.

Normalized enrichment scores (NES) for significantly enriched MSigDB Hallmark Pathways in maternal (top panels) and fetal liver (bottom panels). Size of the dot indicates the log10(p value) with larger dots corresponding to more significant p values. Dot color gradient corresponds to the number of groups (1–8, with lighter colors corresponding to higher values) for which the pathway was significantly enriched and blank regions on the plot indicate no statistical significance of the pathway NES for that specific group. P ≤ 0.05 for all pathways shown.
Based on the results of the MSigDB Hallmark Pathway analysis, related pathways for Peroxisome and Oxidative Phosphorylation were used to construct custom gene networks in IPA. Significant DEGs were populated into the resulting gene networks to visualize effects in the maternal and fetal liver. Consistent with our other results, fetal liver low dose group effects on DEGs and pathway enrichment tended to be stronger than maternal liver low dose group effects (oxidative phosphorylation, Fig. 5; peroxisomes, Fig. S7; cholesterol homeostasis, Fig. S8). In general, DEGs and pathway enrichment was more evident in the maternal liver high exposure groups, particularly for oxidative phosphorylation and peroxisomes (Figs. S9–S11).
Fig. 5.

Gene networks for the Oxidative Phosphorylation Hallmark Pathway and related Hallmark Pathways constructed in Ingenuity Pathway Analysis using statistically significant DEGs in (A) maternal liver after gestational exposure to 1 mg/kg PFOA, (B) fetal liver after in utero exposure to 1 mg/kg PFOA, (C) maternal liver after gestational exposure to 2 mg/kg GenX, and (D) fetal liver after in utero exposure to 2 mg/kg GenX. DEGs were considered significant if absolute fold-change values were ≥ 2 and p ≤ 0.005.
3.6. Correlation between maternal phenotype and gene expression
Phenotypic data for maternal liver weight, body weight, and clinical chemistry previously reported in Blake et al. (2020) were correlated with transcriptome-wide gene expression (for the individual animals used in these studies) to determine whether the expression of specific genes correlated with phenotypic anchors (Table 3). The maternal phenotype with the greatest number of correlated genes was relative liver weight, with 1085 correlated genes (868 positively correlated, 217 negatively correlated; Table 3 and Table S6). Other endpoints with gene correlations included low density lipoprotein (LDL; 18 positively correlated, 9 negatively correlated), aspartate aminotransferase (AST; 35 positively correlated, 3 negatively correlated), serum triglycerides (10 positively correlated, 3 negatively correlated), alkaline phosphatase (ALP; 3 positively correlated, 1 negatively correlated), albumin (ALB; 1 positively correlated, 1 negatively correlated), sorbitol dehydrogenase (SDH; 1 positively correlated, 1 negatively correlated), total protein (TP; 2 positively correlated), and maternal weight gain at gestational day 11.5 (1 positively correlated, 1 negatively correlated; Table 3 and Table S6).
Table 3.
Transcriptome-wide transcript-phenotype correlation analysis of maternal liver gene expression and relevant phenotypic endpoints.
| Phenotype | Genes with expression levels significantly * correlated to phenotype | Significantly correlated genes identified as significant DEGs in at least one treatment group (%) | Significantly correlated genes not identified as significant DEGs in at least one treatment group (%) | Overlap between genes with significant correlation to phenotype and DEGs shown in Fig. 2C |
|---|---|---|---|---|
| Relative liver weight (% body weight) | 1085 | 220 (20.3 %) | 865 (79.7 %) | 63.8 % (51/80 genes) |
| Absolute liver weight (g) | 434 | 120 (27.6 %) | 314 (72.3 %) | 18.8 % (15/80 genes) |
| Triglycerides (mg/dL) | 13 | 0 | 13 (100 %) | 0 |
| Weight gain from GD 0.5–11.5 (% change) | 2 | 0 | 2 (100 %) | 0 |
| LDL (mg/dL) | 27 | 0 | 27 (100 %) | 0 |
| ALB (mg/dL) | 2 | 0 | 2 (100 %) | 0 |
| ALP (U/L) | 4 | 2 (50 %) | 2 (50 %) | 0 |
| TP (g/dL) | 2 | 0 | 2 (100 %) | 0 |
| SDH (U/L) | 2 | 0 | 2 (100 %) | 0 |
| AST (U/L) | 38 | 0 | 38 (100 %) | 0 |
Abbr: GD = gestational day, LDL = lactate dehydrogenase, ALB = albumin, ALP = alkaline phosphatase, TP = total protein, SDH = sorbitol dehydrogenase, AST = aspartate aminotransferase.
Pearson’s product moment correlation tests of transcriptome-phenotype correlations ≥ 0.8 and adjusted p-values ≤ 0.05 were considered significant.
The top 10 genes with the most significant positive correlation between gene expression and relative maternal liver weight included: Aconitase 2 (Aco2), Predicted gene 5421 (Gm5421), Solute carrier family 22 member 21 (Slc22a21), Mitochondrial ribosomal protein L15 (Mrpl15), Platelet activating factor acetylhydrolase 2 (Pafah2), Methylthioadenosine phosphorylase (Mtap), Proteasome 20 S subunit alpha 7 (Psma7), Keratin 23 (Krt23), Imprinted and ancient gene protein homolog (Impact), and 5-Oxoprolinase, ATP-hydrolyzing (Oplah; Fig. 6). Of the genes significantly negatively correlated with relative maternal liver weight, the top 10 included: Coagulation factor XI (F11), Amylase alpha 1 A (Amy1), Syntaxin binding protein 6 (Stxbp6), Flavin containing dimethylaniline monooxygenase 3 (Fmo3), RIKEN cDNA 4930594M22 gene (4930594M22Rik), Complement C4B (C4b), Metallophosphoesterase 1 (Mppe1), Inter-alpha-trypsin inhibitor heavy chain 1 (Itih1), Phospholipase A1 member A (Pla1a), and Complement C4A (C4a; Fig. 6). Of the 80 significant DEGs across the high dose groups for PFOA and GenX and the GenX low dose group (shown in Fig. 2), 51 of these DEGs were significantly correlated with relative maternal liver weight (48 positively correlated, 3 negatively correlated; Table S6). There were 220 genes that were significantly differentially regulated in at least one treatment group and significantly correlated with maternal liver weight (166 positively correlated and 54 negatively correlated; Table S6). There were 865 genes whose expression was significantly correlated with relative maternal weight, but not identified as significant DEGs in any treatment group (702 positively correlated, 163 negatively correlated; Table S6).
Fig. 6.

Correlation between individual dam gene expression and phenotype for two example genes, Aco2 and F11, and relative liver weight. Box and whiskers plot showing dam relative liver weights (A) and normalized expression of (B) Aco2 and (C) F11 in the liver, with treatment groups significantly different from the Vehicle Control in solid blue. Transcriptome-phenotype correlation plots, r-squared values, and adjusted p-values for dam relative liver weight and (D) Aco2 and (E) F11 expression. N = 5 dams per treatment group.
4. Discussion
A challenge faced by PFAS researchers is the sheer number of unique chemical structures, with over 12,000 currently identified in the U.S. EPA CompTox Chemicals Dashboard (https://comptox.epa.gov/dashboard/chemical-lists/PFASMASTER). It is not feasible to conduct in vivo studies to evaluate the toxicological effects of this many chemicals, therefore in vitro, in silico, and -omics strategies will be integral to keep up with the growing universe of PFAS. In the present work, we have provided a rich database of transcriptomic and transcriptome-phenotype correlation analyses for two PFAS of high public interest; PFOA, a legacy chemical that has been phased out of use due to its unfavorable toxicological profile, and GenX, a replacement chemical of concern due to its high levels of contamination in multiple communities (Gebbink and van Leeuwen, 2020; Hopkins et al., 2018; Sun et al., 2016). These data can be leveraged in subsequent predictive toxicology efforts; for example, the group of 30 DEGs common across nearly all dose groups in the maternal and fetal liver and the top 10 genes positively or negatively correlated with maternal relative liver weight could serve as a useful resource for designing high-throughput toxicity screens, particularly those with endpoints relevant to fatty acid, lipid, and bile acid synthesis, metabolism, and oxidative phosphorylation in the liver.
Legacy PFAS, namely PFOA and PFOS, are confirmed developmental toxicants in rodents and humans (Blake and Fenton, 2020; Fenton et al., 2021; Lam et al., 2014; U.S. EPA, 2016; NCDEQ/NCDHHS, 2018). However, following the phase out of these chemicals, the potential health effects of replacement PFAS, such as GenX, are not as well characterized as their predecessors. The liver is a primary target of several PFAS (Costello et al., 2022), with reported perturbation of PPARα-dependent and PPARα-independent mechanisms that can lead to liver disease, metabolic disruption, and dyslipidemia [see (Armstrong and Guo, 2019; Fenton et al., 2021) for recent reviews]. There is also growing evidence that exposure to some PFAS during development may program an increased risk for metabolic syndrome and NAFLD (Braun et al., 2016; Halldorsson et al., 2012; Høyer et al., 2015; Janis et al., 2021; Manzano-Salgado et al., 2017; Cope et al., 2021).
Our prior work illustrated the phenotypic similarities induced by exposure to PFOA or GenX in the livers of pregnant CD-1 mice at the organ level (increased liver weight), and the cellular level (adverse constellation of lesions characterized by increased vacuolation, increased numbers of cytoplasmic organelles consistent with mitochondria and peroxisomes, reduced glycogen stores and reduction of rough endoplasmic reticulum ribosomes) (Blake et al., 2020). Some of these maternal findings have been confirmed in rats and Balb/c mice (Conley et al., 2019; Xu et al., 2022). We have also reported that prenatal exposure to PFOA or GenX at levels lower than those in the present study induced adverse metabolic and hepatic phenotypes in adult offspring (Cope et al., 2021), building upon earlier work demonstrating the effects of in utero exposure to PFOA or other legacy PFAS on mouse fetal liver gene expression (Rosen et al., 2007, 2009, 2017; Abbott et al., 2012). Collectively, these studies suggest the potential for PFAS-impaired molecular signaling in the developing fetal liver. The current work bridges a critical knowledge gap by providing evidence for similarities in PFOA and GenX-induced liver changes at the molecular level through analyzing transcriptome-wide gene expression changes and identifying underlying genomic pathways in both the maternal and developing fetal liver, which primarily included those involved in the synthesis, metabolism, and oxidative phosphorylation of fatty acids, lipids, and bile acids.
The maternal liver undergoes numerous physiological changes to support pregnancy, several of which can mirror conditions observed in nonpregnant individuals with liver disease, such as hyper-dynamic circulation resulting from increased heart rate, increased cardiac output, increased plasma volume, and reduced peripheral vascular resistance (Westbrook et al., 2016), suggesting the maternal liver may be particularly vulnerable to hepatotoxicants. Previous studies have implicated a role for maternal exposure to several PFAS in multisystem pregnancy disorders that involve liver dysfunction, such as preeclampsia (Bommarito et al., 2021; Huang et al., 2019; Wikström et al., 2019), but a direct connection between human exposure to PFAS during pregnancy and disrupted maternal liver function has yet to be established. Although these data are sorely needed, a recent study reported that serum levels of some PFAS were associated with disruptions in key hepatic metabolic pathways, including lipid and bile acid metabolism in a nonpregnant cohort (Sen et al., 2022). Here we provide insight into the underlying biological pathways of PFOA- and GenX-induced maternal hepatotoxicity in pregnant mice, which, at the gene expression level, primarily involved disruptions in lipid and bile acid metabolism and oxidative phosphorylation. Our findings are consistent with maternal liver gene expression in Sprague-Dawley rats after gestational exposure to GenX, which resulted in up-regulation of genes involved in fatty acid metabolism, adipogenesis, and PPAR signaling (Conley et al., 2019). These biological processes are compromised in MAFLD (Eslam et al., 2020; Shao et al., 2020). Furthermore, MAFLD during pregnancy is associated with increased risk for preeclampsia, pregnancy-induced hypertension, and gestational diabetes mellitus (Lee et al., 2021).
The ability to connect gene expression to phenotype is critical for establishing causal relationships between exposures and outcomes and for building confidence in toxicological mechanisms of action. Herein, the relationship between gene expression and relevant phenotypic endpoints were measured within the same mouse. In doing so, we were able to identify specific genes for which expression levels were tightly correlated to these phenotypic anchors, with relative maternal liver weight exhibiting the greatest number of significant transcriptome-phenotype correlations. A strength of the phenotype-transcript correlation analysis reported in this work was the inclusion of all genes captured in the transcriptome-wide expression analysis, rather than limiting the correlation analysis to only genes meeting the criteria for significant differential expression. Approximately 80 % of the genes that were significantly correlated with maternal relative liver weight were not identified as significant DEGs, meaning these genes were not captured in our pathway analyses (criteria for pathway analysis were genes with fold-change ≥ 2 and P ≤ 0.005). Interestingly, Proteasome 20 S subunit alpha 7 (Psma7), Metallophosphoesterase 1 (Mppe1), and Inter-alpha-trypsin inhibitor heavy chain 1 (Itih1) were some of the top-ranking genes in the correlation analysis yet did not have P-values < 0.005 and thus were not captured in DEG analyses. Aberrant expression of these genes is associated with hepatocellular carcinoma (Tu et al., 2019; Chen et al., 2020; Chang et al., 2021; Qian et al., 2020), a disease not identified in our pathway analyses. Additionally, upstream regulator analyses predicted down-regulation of STAT6 and STAT5B in both maternal and fetal liver samples; decreased expression of these genes has been implicated in hepatocellular carcinoma in mice and in humans (Dong et al., 2019; Cui et al., 2007). Had the transcriptome-phenotype correlation analysis been limited to genes meeting the DEG statistical cutoff, Krt23 would have been the only top 10 gene identified in the correlation analysis that is associated with hepatocellular carcinoma in mice (Kim et al., 2019). Gene expression changes not meeting our strict statistical cutoffs may have important biological implications.
Overall, there were substantially fewer phenotype-transcriptome correlations for other liver-related endpoints (e.g., ALP, SDH) when compared with liver weight, which may be the result of obtaining these measurements from serum rather than from the liver, or could result from the relatively brief exposure duration (~16 days). However, our phenotype-transcriptome correlation analyses showed dose-responsive shifts in both gene expression and phenotype for PFOA and GenX and identified genes correlated with phenotypic endpoints that were not altered in a statistically significant manner (e.g., AST). A recent multi-omics study of the hepatotoxic mechanism of perfluorohexanoic acid (PFHxA) in adult male mice identified fatty acid biosynthesis and degradation as key biological processes leading to oxidative stress and subsequent disrupted metabolism and liver injury, evidenced by increased liver weight and liver pathology (Jiang et al., 2021). Taken together with the current study, these data corroborate the link between PFAS-induced disruptions in liver transcriptome and phenotype for certain compounds (i.e., PFOA, GenX, and PFHxA) and build confidence in the role of fatty acid metabolism as a key mechanistic component of hepatotoxicity induced by these PFAS and possibly other PFAS.
The maternal liver does not function in isolation during pregnancy. Maternal adipose tissue, the digestive system (e.g., the intestines), and the placenta all function in collaboration with the liver to sustain a healthy pregnancy and modulate appropriate fetal growth and development. Our prior work demonstrated that the weight and histopathology of the placenta and maternal liver were altered by gestational exposure to PFOA (1 or 5 mg/kg-d) or GenX (2 or 10 mg/kg-d) in CD-1 mice (Blake et al., 2020). In a recent study, gestational exposure to 1 mg/kg-d PFOA or 2 mg/kg-d GenX in Balb/c mice also increased maternal liver weight and induced similar histopathologic alterations, as well as disrupted the intestinal mucosal barrier and permeability (Xu et al., 2022). This suggests the gut-liver axis may be vulnerable to PFAS exposure. Future work should investigate the molecular mechanisms through which gestational exposure to PFAS might perturb biological systems directly interconnected with the maternal liver, such as the maternal gut-liver axis and the placenta.
The health of a developing fetus depends on the ability of the placenta and maternal liver to metabolize and transport critical nutrients and hormones. Thus, disruptions in maternal liver function have a negative impact on the developing fetus and are thought to developmentally program MAFLD in the offspring (Brumbaugh and Friedman, 2014). Here we provide evidence that prenatal exposure to PFOA or GenX disrupts biological pathways in the developing liver that mirror disrupted pathways in the maternal liver, but with different subsets of genes driving these changes and a greater sensitivity of these gene changes to the lower doses of PFOA and GenX. This was evidenced by greater numbers of DEGs in the fetal liver low dose groups relative to the maternal liver low dose groups, including a more consistent response in altered gene expression across the four fetal liver groups compared with the four maternal liver groups. Additionally, hallmark pathway and IPA pathway analyses indicated stronger pathway enrichment in fetal liver low dose groups compared with maternal liver high dose groups. We believe this indicates two important findings: 1) gene expression and pathway changes in fetal livers are consistent with adverse liver outcomes and could predict liver disease and/or program metabolic disease in the offspring, and 2) the fetal liver is uniquely sensitive to the hepatotoxic effects of PFAS, particularly at lower doses.
These findings align with our prior work showing adverse metabolic and liver phenotypes in adult offspring after in utero exposure to doses of PFOA or GenX less than or equal to the low doses in the present study (≤2 mg/kg GenX and ≤1 mg/kg PFOA; Cope et al., 2021). Our findings are also consistent with increased relative liver weights, reduced liver glycogen, and disrupted serum glucose, liver enzymes, cholesterol, and triglycerides in rat offspring developmentally exposed to GenX (Conley et al., 2021). In a human birth cohort, increasing exposure to several PFAS during pregnancy was associated with an increased risk for liver injury in children, including increased serum AST and increased gamma-glutamyltransferase (GGT), a marker indicating damage to the liver’s bile ducts (Stratakis et al., 2020). Our data indicate fetal livers exhibited enrichment of biological and hallmark pathways in a similar manner to maternal livers, with stronger responses in the fetal liver low dose groups relative to the maternal liver low dose groups. Taken together, this growing evidence base suggests developmental exposure to a range of PFAS may damage the developing liver and program an adverse metabolic phenotype in the offspring.
In animal studies, the externally administered dose of a chemical may not reflect tissue-specific bioavailable concentrations of the test chemical due to compound-specific differences in its absorption, distribution, metabolism, and excretion. For PFOA and GenX, elimination rates differ in mice (PFOA half-life = 17–19 days, GenX half-life = 20 h; Gannon et al., 2016; Lau et al., 2012) and it is possible that there are chemical-specific differences in serum protein binding. As reported in Blake et al. (2020), the orally administered doses of PFOA and GenX differed by a factor of two, but the mean maternal serum concentrations of these chemicals were similar at the high (95.1 μg/mL for 5 mg/kg PFOA and 58.5 μg/mL for 10 mg/GenX) and low (18.7 μg/mL for 1 mg/kg PFOA and 22.9 μg/mL 2 mg/kg GenX) doses. However, mean PFOA accumulation in the maternal liver was 40 times greater than GenX at the low doses (4.56 μg/g for 2 mg/kg GenX and 181.1 μg/g for 1 mg/kg PFOA) and 11 times greater at the high doses (14.2 μg/g for 10 mg/kg GenX and 159.2 μg/g for 5 mg/kg PFOA). For fetuses, whole-body dosimetry showed that GenX accumulated about two times less than PFOA at the high and low doses on average (3.2 and 7.7 μg/g for 2 and 10 mg/kg GenX; 5.8 and 16.4 μg/g for 1 and 5 mg/kg PFOA). In comparing maternal serum dosimetry to whole-fetus dosimetry, PFOA levels were roughly four times lower in the fetus compared to dam serum while GenX levels were roughly seven times lower in the fetus relative to dam serum. However, it is difficult to interpret the significance of these values in the absence of tissue specific dosimetry for fetal liver or serum. The disparate accumulation of PFOA and GenX in the maternal liver considered in the context of the current work underscores the extent to which GenX is a far more potent inducer of hepatic gene expression changes than PFOA in CD-1 mice. Future studies should conduct tissue-specific dosimetry to determine the extent to which PFOA and GenX (or other PFAS) accumulate in the developing fetal liver and serum.
There are several limitations to consider in the present study. First, the sample sizes are relatively small and represent a subset of animals evaluated in the Blake et al. (2020) study (N = 4–6 samples per tissue type and treatment group in the present study, N = 10–13 dams per treatment group in the Blake et al., 2020 study). Although a larger sample size would impart greater confidence in the reported findings, the associations reported herein align with our prior work and that of others. However, an important strength of the current work is that the transcriptomic and pathway analyses are bolstered by the phenotypic data which was obtained from the same animals. Additionally, maternal and female fetal livers were collected from the same maternal-fetal units, creating a unique continuity across the maternal phenotype, maternal gene expression, and fetal gene expression data. Although we did not have internal dosimetry data for the serum and livers of the exact mice included in the present study (due to prioritized use of serum for clinical chemistry), our prior work presented these data for mice that underwent the same exposures (Blake et al., 2020), providing useful context for interpreting the results of the present work. It is possible that fetal liver gene expression changes may be an indirect consequence of the exposure (e.g., due to alterations in maternal lipid metabolism and transport to the fetus) rather than a direct consequence of the fetal liver responding to the chemical stressor, or some combination of the two. Lastly, this study only compares transcriptomic and pathway alterations in maternal and female fetal liver, but not male fetal liver. This decision was made to eliminate sex-specific differences in order to compare effects between the maternal and fetal livers but came at the cost of being unable to compare effects between the sexes after developmental exposure. Particularly with regard to the developing liver, sex differences in PFAS-induced hepatotoxicity are important, as were identified in our recent work on some of these chemicals (Cope et al., 2021) and should be considered in future work.
In this work we demonstrated that alterations in hepatic signaling within the maternal liver by two PFAS with important differences in chemical structure, PFOA and GenX, mirrored pathways altered in the fetal liver. We simultaneously investigated livers obtained from the same maternal-fetal units and identified synthesis and metabolism of fatty acids, lipids, and bile acids to be key molecular pathways in the hepatotoxic effects of two different PFAS in pregnant mice and their developing offspring. Furthermore, we identified several non-coding genes that were significantly altered in both maternal and fetal livers and may serve as useful reference points to compare across different species and PFAS. Importantly, gene expression and pathway enrichment in the developing liver was more sensitive to lower dose effects of both PFOA and GenX relative to the maternal liver. We demonstrated that these PFAS-induced alterations in hepatic signaling within the maternal liver may be predictive of altered pathways in the fetal liver, which may allow for the identification of key molecular events in the developmental toxicity of these and other PFAS. Future work should validate the DEG sets identified here to determine their ability to predict PFAS-induced hepatotoxicity. Our findings are consistent with disruptions in the developing liver that would program increased risk for metabolic dysfunction-associated fatty liver disease and suggest that prenatal exposure to the currently studied PFAS (and likely others) may predispose offspring to liver disease.
Supplementary Material
Acknowledgements
The authors thank Dr. Justin Conley (U.S. EPA), Dr. Michelle Angrish (U.S. EPA), and Dr. Charles Breeze (NCI) for their review of this manuscript. The authors thank Dr. Kevin Gerrish (NIEHS) for his support with the transcriptomics work.
Disclaimer
This work was supported by NIEHS funding Z0ES102785 (SEF). This project was also supported in part by an appointment to the Research Participation Program at the Center for Public Health and Environmental Assessment at the U.S. EPA administered by the Oak Ridge Institute for Science and Education through an interagency agreement between the U.S. Department of Energy and US EPA. The research described in this article was reviewed and approved for publication by NIEHS and the Center for Public Health and Environmental Assessment at the U.S. EPA. The views expressed in this article are those of the authors and do not necessarily represent the views or policies of the U.S. EPA, the Department of Health and Human Services, nor the U.S. government. The mention of trade names or commercial products does not constitute endorsement or recommendation for use.
Footnotes
CRediT authorship contribution statement
BEB: Conceptualization, Methodology, Investigation, Data curation, Writing – original draft, Visualization, Supervision, Project administration. CNM: Conceptualization, Writing – original draft, Funding acquisition. HN: Data curation, Writing – original draft. VAC: Writing – review & editing, Software, Formal analysis, Data curation. TPP: Writing – original draft. DPP: Software, Formal analysis, Resources, Data curation, Writing – review & editing. MRB-M: Software, Formal analysis, Resources, Data curation, Writing – review & editing. DM: Software, Formal analysis, Resources, Data curation. RRS: Software, Formal analysis, Resources, Data curation. SEF: Conceptualization, Methodology, Writing – review & editing, Visualization, Supervision, Project administration, Funding acquisition.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Appendix A. Supporting information
Supplementary data associated with this article can be found in the online version at doi:10.1016/j.ecoenv.2022.114314.
Data availability
All data will be made publicly available upon acceptance.
References
- Abbott BD, Wood CR, Watkins AM, Tatum-Gibbs K, Das KP, Lau C, 2012. Effects of perfluorooctanoic acid (PFOA) on expression of peroxisome proliferator-activated receptors (PPAR) and nuclear receptor-regulated genes in fetal and postnatal CD-1 mouse tissues. Reprod. Toxicol 33 (4), 491–505, 1. [DOI] [PubMed] [Google Scholar]
- Armstrong LE, Guo GL, 2019. Understanding environmental contaminants’ direct effects on non-alcoholic fatty liver disease progression. Curr. Environ. Health Rep 6, 95–104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ATSDR Toxicological Profile for Perfluoroalkyls Agency for Toxic Substances and Disease Registry 2020.https://www.atsdr.cdc.gov/toxprofiles/tp200.pdf. [PubMed]
- Bauman DE, Currie WB, 1980. Partitioning of nutrients during pregnancy and lactation: a review of mechanisms involving homeostasis and homeorhesis. J. Dairy Sci 63, 1514–1529. [DOI] [PubMed] [Google Scholar]
- Benjamini Y, Hochberg Y, 1995. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B (Methodol.) 57, 289–300. [Google Scholar]
- Blake BE, Cope HA, Hall SM, Keys RD, Mahler BW, McCord J, et al. , 2020. Evaluation of maternal, embryo, and placental effects in cd-1 mice following gestational exposure to perfluorooctanoic acid (pfoa) or hexafluoropropylene oxide dimer acid (hfpo-da or genx). Environ. Health Perspect 128, 027006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blake BE, Fenton SE, 2020. Early life exposure to per-and polyfluoroalkyl substances (pfas) and latent health outcomes: A review including the placenta as a target tissue and possible driver of peri-and postnatal effects. Toxicology 443, 152565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blomberg AJ, Shih Y-H, Messerlian C, Jørgensen LH, Weihe P, Grandjean P, 2021. Early-life associations between per-and polyfluoroalkyl substances and serum lipids in a longitudinal birth cohort. Environ. Res 200, 111400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bommarito PA, Ferguson KK, Meeker JD, McElrath TF, Cantonwine DE, 2021. Maternal levels of perfluoroalkyl substances (pfas) during early pregnancy in relation to preeclampsia subtypes and biomarkers of preeclampsia risk. Environ. Health Perspect 129, 107004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Braun JM, Chen A, Romano ME, Calafat AM, Webster GM, Yolton K, et al. , 2016. Prenatal perfluoroalkyl substance exposure and child adiposity at 8 years of age: the home study. Obesity 24, 231–237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brumbaugh DE, Friedman JE, 2014. Developmental origins of nonalcoholic fatty liver disease. Pediatr. Res 75, 140–147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang Q-H, Mao T, Tao Y, Dong T, Tang X-X, Ge G-H, et al. , 2021. Pan-cancer analysis identifies ITIH1 as a novel prognostic indicator for hepatocellular carcinoma. Aging. 10.18632/aging.202765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen X, Xu J, Wang P, Shang L, Guo J, Huang L, Jiang YA, Chen J, Chen H, Shang Y, Zhang Q, et al. , 2020. Metallophosphoesterase 1, a novel candidate gene in hepatocellular carcinoma malignancy and recurrence. Cancer Biol Ther. 10.1080/15384047.2020.1824480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conley JM, Lambright CS, Evans N, McCord J, Strynar MJ, Hill D, et al. , 2021. Hexafluoropropylene oxide-dimer acid (hfpo-da or genx) alters maternal and fetal glucose and lipid metabolism and produces neonatal mortality, low birthweight, and hepatomegaly in the sprague-dawley rat. Environ. Int 146, 106204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conley JM, Lambright CS, Evans N, Strynar MJ, McCord J, McIntyre BS, et al. , 2019. Adverse maternal, fetal, and postnatal effects of hexafluoropropylene oxide dimer acid (genx) from oral gestational exposure in sprague-dawley rats. Environ. Health Perspect 127, 037008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cope HA, Blake BE, Love C, McCord J, Elmore SA, Harvey JB, et al. , 2021. Latent, sex-specific metabolic health effects in cd-1 mouse offspring exposed to pfoa or hfpo-da (genx) during gestation. Emerg. Contam 7, 219–235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Costello E, Rock S, Stratakis N, Eckel SP, Walker DI, Valvi D, et al. , 2022. Exposure to per-and polyfluoroalkyl substances and markers of liver injury: a systematic review and meta-analysis. Environ. Health Perspect 130 (4), 046001 10.1289/EHP10092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cui Y, Hosui A, Sun R, Shen K, Gavrilova O, Chen W, Cam MC, Gao B, Robinson GW, Hennighausen L, 2007. Loss of signal transducer and activator of transcription 5 leads to hepatosteatosis and impaired liver regeneration. Hepatology 46, 504–513. [DOI] [PubMed] [Google Scholar]
- Dong Z, Chen Y, Yang C, Zhang M, Chen A, Yang J, Huang Y, 2019. STAT gene family mRNA expression and prognostic value in hepatocellular carcinoma. OncoTargets Ther. 12, 7175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dupont Chem C, 2010a. Dupont-18405-841: an Oral (Gavage) Prenatal Developmental Toxicity Study of h-28548 in Rats, Ashland, Ohio. [Google Scholar]
- Dupont Chem C, 2010b. Dupont-18405-1037: an Oral (Gavage) Reproduction/developmental Toxicity Screening Study of h-28548 in Mice, Ashland, Ohio. [Google Scholar]
- Eslam M, Newsome PN, Sarin SK, Anstee QM, Targher G, Romero-Gomez M, et al. , 2020. A new definition for metabolic dysfunction-associated fatty liver disease: an international expert consensus statement. J. Hepatol 73, 202–209. [DOI] [PubMed] [Google Scholar]
- Fenton SE, Ducatman A, Boobis A, DeWitt JC, Lau C, Ng C, et al. , 2021. Per-and polyfluoroalkyl substance toxicity and human health review: current state of knowledge and strategies for informing future research. Environ. Toxicol. Chem 40, 606–630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Filgo AJ, Quist EM, Hoenerhoff MJ, Brix AE, Kissling GE, Fenton SE, 2015. Perfluorooctanoic acid (pfoa)–induced liver lesions in two strains of mice following developmental exposures: Pparα is not required. Toxicol. Pathol 43, 558–568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Foulds CE, Treviño LS, York B, Walker CL, 2017. Endocrine-disrupting chemicals and fatty liver disease. Nat. Rev. Endocrinol 13, 445–457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gebbink WA, van Leeuwen SP, 2020. Environmental contamination and human exposure to pfass near a fluorochemical production plant: review of historic and current pfoa and genx contamination in the netherlands. Environ. Int 137, 105583. [DOI] [PubMed] [Google Scholar]
- Gorodkin J, 2004. Comparing two k-category assignments by a k-category correlation coefficient. Comput. Biol. Chem 28, 367–374. [DOI] [PubMed] [Google Scholar]
- Halldorsson TI, Rytter D, Haug LS, Bech BH, Danielsen I, Becher G, et al. , 2012. Prenatal exposure to perfluorooctanoate and risk of overweight at 20 years of age: a prospective cohort study. Environ. Health Perspect 120, 668–673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hines EP, White SS, Stanko JP, Gibbs-Flournoy EA, Lau C, Fenton SE, 2009. Phenotypic dichotomy following developmental exposure to perfluorooctanoic acid (pfoa) in female cd-1 mice: low doses induce elevated serum leptin and insulin, and overweight in mid-life. Mol. Cell. Endocrinol 304, 97–105. [DOI] [PubMed] [Google Scholar]
- Hopkins ZR, Sun M, DeWitt JC, Knappe DR, 2018. Recently detected drinking water contaminants: genx and other per-and polyfluoroalkyl ether acids. J. Water Works Assoc 110, 13–28. [Google Scholar]
- Høyer BB, Ramlau-Hansen CH, Vrijheid M, Valvi D, Pedersen HS, Zviezdai V, et al. , 2015. Anthropometry in 5-to 9-year-old greenlandic and ukrainian children in relation to prenatal exposure to perfluorinated alkyl substances. Environ. Health Perspect 123, 841–846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang R, Chen Q, Zhang L, Luo K, Chen L, Zhao S, et al. , 2019. Prenatal exposure to perfluoroalkyl and polyfluoroalkyl substances and the risk of hypertensive disorders of pregnancy. Environ. Health 18, 1–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janis JA, Rifas-Shiman SL, Seshasayee SM, Sagiv S, Calafat AM, Gold DR, et al. , 2021. Plasma concentrations of per-and polyfluoroalkyl substances and body composition from mid-childhood to early adolescence. J. Clin. Endocrinol. Metab 106, e3760–e3770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang L, Hong Y, Xie G, Zhang J, Zhang H, Cai Z, 2021. Comprehensive multi-omics approaches reveal the hepatotoxic mechanism of perfluorohexanoic acid (pfhxa) in mice. Sci. Total Environ 790, 148160. [DOI] [PubMed] [Google Scholar]
- Jin R, McConnell R, Catherine C, Xu S, Walker DI, Stratakis N, et al. , 2020. Perfluoroalkyl substances and severity of nonalcoholic fatty liver in children: an untargeted metabolomics approach. Environ. Int 134, 105220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krämer A, Green J, Pollard J Jr, Tugendreich S, 2014. Causal analysis approaches in ingenuity pathway analysis. Bioinformatics 4, 523–530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lam J, Koustas E, Sutton P, Johnson PI, Atchley DS, Sen S, et al. , 2014. The navigation guide—evidence-based medicine meets environmental health: integration of animal and human evidence for pfoa effects on fetal growth. Environ. Health Perspect 122, 1040–1051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lau C, Thibodeaux JR, Hanson RG, Narotsky MG, Rogers JM, Lindstrom AB, et al. , 2006. Effects of perfluorooctanoic acid exposure during pregnancy in the mouse. Toxicol. Sci 90, 510–518. [DOI] [PubMed] [Google Scholar]
- Lee SM, Jung YM, Choi ES, Kwak SH, Koo JN, Oh IH, et al. , 2021. Metabolic dysfunction-associated fatty liver disease and subsequent development of adverse pregnancy outcomes. Clin. Gastroenterol. Hepatol [DOI] [PubMed] [Google Scholar]
- Lefebvre P, Staels B, 2021. Hepatic sexual dimorphism—implications for non-alcoholic fatty liver disease. Nat. Rev. Endocrinol 17, 662–670. [DOI] [PubMed] [Google Scholar]
- Liberzon A, Birger C, Thorvaldsdóttir H, Ghandi M, Mesirov JP, Tamayo P, 2015. The molecular signatures database hallmark gene set collection. Cell Syst. 1, 417–425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macon MB, Villanueva LR, Tatum-Gibbs K, Zehr RD, Strynar MJ, Stanko JP, White SS, Helfant L, Fenton SE, 2011. Prenatal perfluorooctanoic acid exposure in CD-1 mice: low-dose developmental effects and internal dosimetry. Toxicol. Sci 1, 134–145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mamsen LS, Björvang RD, Mucs D, Vinnars MT, Papadogiannakis N, Lindh CH, et al. , 2019. Concentrations of perfluoroalkyl substances (pfass) in human embryonic and fetal organs from first, second, and third trimester pregnancies. Environ. Int 124, 482–492. [DOI] [PubMed] [Google Scholar]
- Mamsen LS, Jönsson BA, Lindh CH, Olesen RH, Larsen A, Ernst E, et al. , 2017. Concentration of perfluorinated compounds and cotinine in human foetal organs, placenta, and maternal plasma. Sci. Total Environ 596, 97–105. [DOI] [PubMed] [Google Scholar]
- Mantovani A, Scorletti E, Mosca A, Alisi A, Byrne CD, Targher G, 2020. Complications, morbidity and mortality of nonalcoholic fatty liver disease. Metabolism 111s, 154170. [DOI] [PubMed] [Google Scholar]
- Manzano-Salgado CB, Casas M, Lopez-Espinosa M-J, Ballester F, Iñiguez C, Martinez D, et al. , 2017. Prenatal exposure to perfluoroalkyl substances and cardiometabolic risk in children from the spanish inma birth cohort study. Environ. Health Perspect 125, 097018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matthews BW, 1975. Comparison of the predicted and observed secondary structure of t4 phage lysozyme. Biochim. Biophys. Acta (BBA)-Protein Struct 405, 442–451. [DOI] [PubMed] [Google Scholar]
- Mitra S, De A, Chowdhury A, 2020. Epidemiology of non-alcoholic and alcoholic fatty liver diseases. Transl. Gastroenterol. Hepatol 5, 16–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- NCDEQ/NCDHHS, 2018. Secretaries’ Science Advisory Board Review of the North Carolina Drinking Water Provisional Health Goal for Genx. North Carolina Department of Environmental Quality and North Carolina Department of Health and Human Services. [Google Scholar]
- OECD, 2021. Reconciling terminology of the universe of per- and polyfluoroalkyl substances: recommendations and practical guidance. Series on Risk Management no. 61. https://www.oecd.org/chemicalsafety/portal-perfluorinated-chemicals/terminology-per-and-polyfluoroalkyl-substances.pdf 2022]. [Google Scholar]
- Perumpail BJ, Khan MA, Yoo ER, Cholankeril G, Kim D, Ahmed A, 2017. Clinical epidemiology and disease burden of nonalcoholic fatty liver disease. World J. Gastroenterol 23, 8263–8276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qi W, Clark JM, Timme-Laragy AR, Park Y, 2020. Per-and polyfluoroalkyl substances and obesity, type 2 diabetes and non-alcoholic fatty liver disease: a review of epidemiologic findings. Toxicol. Environ. Chem 102, 1–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qian X, Bao Z-M, Yao D, Shi Y, et al. , 2022. Lysine demethylase 5C epigenetically reduces transcription of ITIH1 that results in augmented progression of liver hepatocellular carcinoma. The Kaohsiung Journal of Medical Sciences. 10.1002/kjm2.12501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quist EM, Filgo AJ, Cummings CA, Kissling GE, Hoenerhoff MJ, Fenton SE, 2015. Hepatic mitochondrial alteration in cd-1 mice associated with prenatal exposures to low doses of perfluorooctanoic acid (pfoa). Toxicol. Pathol 43, 546–557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosen MB, Das KP, Rooney J, Abbott B, Lau C, Corton JC, 2017. PPARα-independent transcriptional targets of perfluoroalkyl acids revealed by transcript profiling. Toxicology 387, 95–107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosen MB, Schmid JE, Das KP, Wood CR, Zehr RD, Lau C, 2009. Gene expression profiling in the liver and lung of perfluorooctane sulfonate-exposed mouse fetuses: comparison to changes induced by exposure to perfluorooctanoic acid. Reprod. Toxicol 27 (3–4), 278–288. [DOI] [PubMed] [Google Scholar]
- Rosen MB, Thibodeaux JR, Wood CR, Zehr RD, Schmid JE, Lau C, 2007. Gene expression profiling in the lung and liver of PFOA-exposed mouse fetuses. Toxicology 239 (1–2), 15–33. [DOI] [PubMed] [Google Scholar]
- Roth K, Yang Z, Agarwal M, Liu W, Peng Z, Long Z, et al. , 2021. Exposure to a mixture of legacy, alternative, and replacement per-and polyfluoroalkyl substances (pfas) results in sex-dependent modulation of cholesterol metabolism and liver injury. Environ. Int 157, 106843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sen P, Qadri S, Luukkonen PK, Ragnarsdottir O, McGlinchey A, Jäntti S, et al. , 2022. Exposure to environmental contaminants is associated with altered hepatic lipid metabolism in non-alcoholic fatty liver disease. J. Hepatol 76, 283–293. [DOI] [PubMed] [Google Scholar]
- Shao M, Ye Z, Qin Y, Wu T, 2020. Abnormal metabolic processes involved in the pathogenesis of non-alcoholic fatty liver disease. Exp. Ther. Med 20, 1–1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stratakis N, C. D,V, Jin R, Margetaki K, Valvi D, Siskos AP, et al. , 2020. Prenatal exposure to perfluoroalkyl substances associated with increased susceptibility to liver injury in children. Hepatology 72, 1758–1770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subramanian A, Tamayo P, Mootha VK, Mukherjee S, Ebert BL, Gillette MA, et al. , 2005. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 102, 15545–15550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun M, Arevalo E, Strynar M, Lindstrom A, Richardson M, Kearns B, et al. , 2016. Legacy and emerging perfluoroalkyl substances are important drinking water contaminants in the cape fear river watershed of north carolina. Environ. Sci. Technol. Lett 3, 415–419. [Google Scholar]
- Sunderland EM, Hu XC, Dassuncao C, Tokranov AK, Wagner CC, Allen JG, 2019. A review of the pathways of human exposure to poly-and perfluoroalkyl substances (pfass) and present understanding of health effects. J. Expo. Sci. Environ. Epidemiol 29, 131–147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- U.S. Environmental Protection Agency (U.S. EPA), 2016. Drinking water health advisory for perfluorooctanoic acid (pfoa). EPA Document 822-R-16-005.
- U.S. Envrionmental Protection Agency (U.S. EPA), 2021. Human health toxicity values for hexafluoropropylene oxide (HFPO) dimer acid and its ammonium salt (CASRN 13252-13-6 and CASRN 62037-80-3) also known as “GenX chemicals”. EPA Document 822R-21-010.
- Tu J, Chen J, He M, Tong H, Liu H, Zhou B, Liao Y, Wang Z, et al. , 2019. Bioinformatics analysis of molecular genetic targets and key pathways for hepatocellular carcinoma. Onco Targets Ther. 10.2147/OTT.S198802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Esterik J, Sales LB, Dollé M, Håkansson H, Herlin M, Legler J, et al. , 2016. Programming of metabolic effects in c57bl/6jxfvb mice by in utero and lactational exposure to perfluorooctanoic acid. Arch. Toxicol 90, 701–715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Z, Buser AM, Cousins IT, Demattio S, Drost W, Johansson O, et al. , 2021. A new oecd definition for per-and polyfluoroalkyl substances. Environ. Sci. Technol 55, 15575–15578. [DOI] [PubMed] [Google Scholar]
- Welsh JA, Karpen S, Vos MB, 2013. Increasing prevalence of nonalcoholic fatty liver disease among united states adolescents, 1988–1994 to 2007–2010. J. Pediatr 162, 496–500.e491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Westbrook RH, Dusheiko G, Williamson C, 2016. Pregnancy and liver disease. J. Hepatol 64, 933–945. [DOI] [PubMed] [Google Scholar]
- White SS, Calafat AM, Kuklenyik Z, Villanueva L, Zehr RD, Helfant L, et al. , 2007. Gestational pfoa exposure of mice is associated with altered mammary gland development in dams and female offspring. Toxicol. Sci 96, 133–144. [DOI] [PubMed] [Google Scholar]
- Wikström S, Lindh CH, Shu H, Bornehag C-G, 2019. Early pregnancy serum levels of perfluoroalkyl substances and risk of preeclampsia in swedish women. Sci. Rep 9, 1–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu L-L, Chen Y-K, Zhang Q-Y, Chen L-J, Zhang K-K, Li J-H, et al. , 2022. Gestational exposure to genx induces hepatic alterations by the gut-liver axis in maternal mice: a similar mechanism as pfoa. Sci. Total Environ, 153281 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
All data will be made publicly available upon acceptance.