

ABSTRACT
Snake toxins are widely studied owing to their importance in snakebite accidents, a serious public health issue in tropical countries, and their broad therapeutic potential. Isolated fractions from venom produced by snakes of the genus Crotalus sp. present a wide variety of pharmacological uses such as antifungal, antiviral, antibacterial, and antitumor properties, among other therapeutic potentialities. Given the direct effect of this venom on tumor cells, isolation of its compounds is important for the characterization of its anticarcinogenic actions. Crotalus durissus terrificus venom and its toxins have been widely evaluated as potential candidates for the development of new antineoplastic therapies that are efficient against different tumor lines and cellular targets. This review highlights the venom toxins of this species, with a focus on their antineoplastic properties.
Keywords: Snake Venom, Cancer, Antitumor, Crotalid Venoms, Crotalus
INTRODUCTION
Currently, approximately 11,341 reptile species are recognized worldwide 1 , with 1,116 species found in Australia, 974 in Mexico, and 830 in Brazil. Crotalus comprises of the venomous Viperidae snakes 2 - 7 from the subfamily Crotalinae, also known as rattlesnakes. These are distributed across South America, mainly from Colombia to Argentina 5 , 7 - 9 , with the following six subspecies found in Brazil: Crotalus durissus cascavella, C. d. collilineatus, C. d. terrificus, C. d. marajoensis, C. d. ruruima, and C. d. durissus 4 , 6 , 9 .
These snakes are primarily nocturnal 5 and solenoglyphic dentition 5 , 10 presents loreal pits, a thermoreceptor organ of viperid species, visible as openings between the eye and the nostril of the animal head, which are of great importance for the detection of temperature variations, particularly of prey and predators 5 . The most striking characteristic of Crotalus snakes is the presence of a rattle at the end of their tails (Figure 1) 5 , 6 .
FIGURE 1: (A) Crotalus durissus terrificus; (B) Rattle detail; (C) Loreal Pit (yellow circle).

Crotalus snakes cause frequent and severe accidents, represent a serious public health problem in tropical countries, and the snakebites are considered a neglected disease by the World Health Organization 6 , 11 - 13 . However, venom is an important biotechnological tool because of the specialization and efficiency of its components, which affect a large number of targets with high selectivity and affinity 14 - 16 .
CROTALUS DURISSUS TERRIFICUS VENOM: COMPOSITION, GENERAL PHARMACOLOGICAL ACTIONS AND ANTINEOPLASTIC APPLICATIONS
Snake venom is one of the richest sources of bioactive substances in nature and is therefore of great interest for the development of new drugs 4 , 14 - 28 . Snake venoms are composed of a mixture of proteins, organic compounds, inorganic ions, carbohydrates, lipid fractions, and other substances 4 , 14 , 16 , 17 , 20 , 21 , 27 , 29 , 30 .
Proteins account for approximately 90% of the dry weight of snake venom 4 , 21 , 29 , 31 , 32 . C. d. terrificus (Cdt) venom is mainly composed of phospholipase A2 (PLA2), serinoproteases, hyaluronidases, L-amino acid oxidases, peptides, low molecular weight organic compounds, inorganic ions, and enzyme inhibitors 4 , 33 . The main toxins found in Cdt venom are Crotoxin, Convulxin, Gyroxin, and Crotamine 4 , 6 - 8 , 34 - 38 . This venom also contains more than 60 different protein families 23 . Envenomation generates local manifestations of pain, edema, erythema, paresthesia, and systemic manifestations such as eyelid ptosis, facial muscle paralysis, and myoglobinuria, among other clinical signs 4 , 6 , 25 , 31 , 35 , 39 , because of its neurotoxic, coagulant, and myotoxic actions 4 , 25 , 31 , 33 , 35 .
There is a wide variety of pharmacological uses of the different fractions of Crotalus sp. venom, including antifungal, antiviral, antibacterial, antitumor, and antiprotozoal activities 4 , 15 , 26 - 28 , 37 , 40 - 43 .
CROTOXIN
Crotoxin represents approximately 40%-60% of the dry weight of the Cdt venom 4 , 8 , 19 , 33 , 36 , 42 - 44 and is a potent neurotoxin formed by PLA2 and crotapotin, forming a complex of high toxicity 4 , 8 , 32 , 38 , 42 - 54 , and exhibits myotoxic, nephrotoxic, and cardiotoxic effects 4 , 37 , 38 , 43 , 44 , 46 , 48 , 50 .
This neurotoxic action is mainly attributed to the inhibitory mechanism of acetylcholine release in presynaptic neurons 48 , 52 , 54 . Desensitization of postsynaptic nicotinic receptors is another mechanism that reduces the response to acetylcholine 48 , 52 . Thus, crotoxin acts by blocking potassium channels and prolonging the action potential at neuromuscular junctions, thereby increasing calcium influx into the channels, mainly due to the presence and high activity of PLA2 in its composition 8 , 48 , 52 .
Crotoxin has been widely studied for its immunomodulatory, anti-inflammatory, antitumor, antimicrobial, and analgesic actions 4 , 40 , 43 , 44 , 46 , 48 , 50 - 54 . In vivo studies have demonstrated its ability to inhibit the production of pro-inflammatory and anti-inflammatory cytokines from the injection of the toxin in rats, including IL-10, IL-4, IL-6, and tumor necrosis factor 36 , 43 . This immunomodulatory activity may be associated with the production of anti-inflammatory mediators via the lipoxygenase pathway, such as lipoxin A4 (LXA4), and the activation of formyl peptide receptors, in addition to its regulatory role in macrophages 36 , 43 , 44 , 51 .
In vitro and in vivo studies have described activating mechanisms of cell apoptosis in different cancer cell lines 19 , 47 - 51 , 53 , 55 induced by cellular autophagy mechanisms 47 , 53 . Both cell death pathways activated by crotoxin (apoptosis and autophagy) can occur simultaneously or sequentially through mechanisms such as changes in mitochondrial membrane potential and release of intracellular cytochrome C. Another important factor related to the mechanism of action of crotoxin is its apparent selectivity for cells with high expression of epidermal growth factor receptors (EGFR) 19 , 21 , 47 , 50 , 56 .
The cytotoxic action on glioblastoma and benign pituitary adenoma cells was partially attributed to crotoxin, which is also cytotoxic to human mammary duct carcinoma and human lung adenocarcinoma cell lines 4 , 19 , 47 - 51 , 55 , 57 . The application of portions of the toxin to murine erythroleukemia cells demonstrated the potential to reduce the viability of the strain 38 . To observe cytotoxicity, the B subunit of crotoxin was separated from PLA2 and used alone 38 .
The isolated crotoxin is cytotoxic to different cell lines, with different cell response 53 . The mechanisms evaluated involved changes in the mitochondrial membrane potential, release of cytochrome C, and activation of caspase-3, a protease essential for the process of cell apoptosis 47 - 50 , 52 , 53 , 55 . Furthermore, it was possible to conclude that the toxin did not interfere with the viability of keratinocytes, which are highly affected by current antineoplastic therapies 53 .
Crotoxin provokes possible damage to the cellular DNA of PANC-1 cells, associated with pancreatic tumors, by upregulating protein expression 53 . DNA damage has also been observed in glioma cell lines, leading to an increase in the percentage of cells undergoing apoptosis. Some in vitro studies have also reported a higher percentage of apoptosis among SK-MES-1 cells, a lung cancer cell line, in addition to damage such as nuclear condensation and fragmentation 50 , 57 .
When associated with tyrosine kinase inhibitors, crotoxin potentiates the antitumor effect of the drug against lung tumor cell lines 50 , 53 , 57 . In a dose-dependent manner, the toxin prevents DNA synthesis and interrupts the cell cycle in the S phase, suppressing the proliferation of SK-MES-1 cells both in vitro and in vivo 52 , 57 . One of the mechanisms identified was the increased expression and cleavage of caspase-3, which is responsible for inducing cell apoptosis 50 , 57 . Another mechanism observed was the induction of cytochrome C release, which increased the occurrence of cellular autophagy, a mechanism also observed in MCF-7 breast cancer lines 47 , 49 , 53 .
Crotoxin also induces the release of LXA4, pro-inflammatory eicosanoid lipoxin, and its analogs through the induction of its synthesis in macrophages 36 , 44 , 46 , 48 , 51 . In vivo studies of Walker 256 carcinoma cells concluded that this mechanism is responsible for the antineoplastic action of crotoxin on the lineage, and the concentration of lipoxin was 74% higher in the plasma of animals treated with crotoxin than in those treated with saline solution 51 . Lipoxins have been shown to be antineoplastic owing to their ability to inhibit tumor growth by inhibiting endothelial cell proliferation and reducing the production of angiogenic growth factors 46 , 51 .
Macrophages cultured in vitro in the presence of crotoxin secreted 47% less angiogenesis mediators than macrophages from a control group 46 , confirming the role of the toxin in reducing tumor blood vessel neoformation.
The efficacy of crotoxin in dose-dependent inhibition of human esophageal carcinoma tumor growth (Eca-109 cells) was demonstrated in vivo 55 , 57 . The toxin causes cellular damage to the lineage, such as formation of pyknotic cell nuclei, cell lysis, and DNA damage 55 . Exposure of tumor cells to crotoxin also resulted in an increase in the number of stagnant cells in the G1 phase of cell division 53 , 55 , 57 . Increased expression of caspase 3, p17, and p15 proteins and reduced production of Bcl-25 protein can be envolved 55 .
In vivo studies on the HL-60 leukemia cell line showed lower tumor growth inhibitory activity, suggesting that it acts preferentially on solid tumors 21 , 47 , 48 , 50 . The treatment of patients with solid tumors refractory to conventional antineoplastic therapies with the administration of different doses of crotoxin has demonstrated efficacy in reducing different types of carcinomas 21 . Mechanisms of mitochondrial collapse, cytochrome C release, and caspase 3 activation induced cell death in the human leukemia-associated K562 cell line, with the induction of apoptosis and autophagy observed 50 , 57 .
Crotoxin has been shown to be more cytotoxic than standard chemotherapeutic agents for the treatment of glioma, pancreatic cancer, esophageal cancer, and cervical cancer. Therefore, novel antineoplastic therapies are of great interest, particularly against leukemia, lung cancer, colon cancer, renal cancer, ovarian cancer, esophageal carcinoma, breast carcinoma, melanoma, and brain tumors, whose proliferation is already known to be preventable by the toxin 19 , 53 , 57 . New drugs derived from the toxin, such as VRCTC-310 and CB24, have already been studied in murine and human cell lines 16 , 17 , 21 , 41 , 48 .
PHOSPHOLIPASES A2
PLA2 are type 1 and type 2 enzymes associated with the induction of inflammatory processes, lipid membrane metabolism, and release of substances such as prostaglandins, prostacyclins, thromboxanes, and leukotrienes 16 , 21 , 58 - 60 .
These enzymes represent the largest family of proteins contained in the venom 23 , 58 , accounting for up to 80% of total proteins 24 . PLA2 induces processes such as edema, blockage of neuromuscular junctions, platelet aggregation, and muscle necrosis 21 , 59 . It has a substantial pharmacological interest owing to a wide range of biological actions 60 . Some enzymes have anticoagulant activity through mechanisms of hydrolysis of procoagulant phospholipids, antagonistic effects with coagulant proteins, and interaction with factor X 25 . Cotrim et al. (2011) suggested that PLA2 activity is attributable to its actions at different pharmacological sites, which are responsible for platelet aggregation, myotoxicity, and antibacterial activity, as well as anti-inflammatory and neurotoxic effects 58 .
PLA2 has shown anticancer properties by acting on epithelial growth factor receptors (EGFR), reducing the production of tumor necrosis factor, and inhibiting neoplastic growth in lung carcinoma, human breast carcinoma, and leukemia.
The Cdt crotoxin and Naja naja atra cardiotoxin association has been conducted to develop “VRCTC-310-Onco,” which aims to interfere with the signaling of EGRFs, reduce the production of tumor necrosis factor, and exert cytotoxic action on tumor cells 16 , 48 . The development of EGFR receptor inhibitor drugs represents a new type of therapy against epithelial neoplasms 61 , 62 given that the receptors act in the signaling responsible for the formation of epithelial cell tumors 61 .
Reduction in tumor necrosis factor production is also an important mechanism of anticancer action, since the presence of necrosis stimulates tumor phosphorescence mediators, favoring angiogenesis and tumor metastasis.
GYROXIN
Gyroxin, a member of the serinoprotease family, is a neurotoxic enzyme with coagulant action 4 , 6 , 25 , 45 , 63 , including thrombin-like action 4 , 37 , 45 , 63 , 64 , and represents the second most commonly found family of venoms 37 . Montoni et al. (2020) demonstrated that the toxin also has the ability to cross the blood-brain barrier 35 .
In vitro studies have revealed that the enzyme generates clotting in human plasma samples with citrate, with the speed of clot formation being directly proportional to the amount of gyroxin 25 , causing the breakdown of fibrinogen into fibrinopeptide A 25 . Gyroxin is the enzyme responsible for the coagulant activity of Cdt venom as it rapidly consumes circulating fibrinogen, making the blood incoagulable.
Brazilian researchers have used this activity to develop a biopolymer (Heterologous Fibrin Sealant, HFS), which consists of a fibrinogen-rich cryoprecipitate extracted from buffalo blood and a thrombin-like enzyme (gyroxin) purified from Crotalus durissus terrificus snake venom 27 , 63 - 65 . They successfully evaluated the safety and immunogenicity of HFS for the first time, estimated the optimum dose, and assessed its preliminary efficacy in the treatment of chronic venous ulcers (CVU) in a phase 2 clinical trial 27 , 63 - 65 .
As gyroxin can cross the blood-brain barrier, it can be an important tool for studies of tumors of the brain and central nervous system.
CONVULXIN
Convulxin is a high-molecular-mass glycoprotein of the C-type lectin family, which has potent platelet activating and aggregating action 4 , 6 , 66 , 67 , with high affinity for platelets 66 . However, its effect on human peripheral blood mononuclear cells (PBMCs) and the immune system remains unclear.
The mechanism of action of convulxin involves the activation of phospholipase C and its rapid phosphorylation, which is similar to the mechanism induced by collagens in mediating platelet aggregation 66 .
In in vitro studies utilizing citrated human plasma samples, the protein generated clot formation without interfering with factors of the coagulation cascade 25 .
CROTAMINE
Crotamine is a non-enzymatic polypeptide with myotoxic and neurotoxic actions, responsible for causing cell death in skeletal muscles due to alterations in their sodium channels 4 , 7 , 10 , 18 , 28 , 37 , 68 - 71 .
A great curiosity is that this myotoxin is not present in all individuals of the species, being thus classified as crotamine-positive or crotamine-negative venom-producing animals 7 , 18 , 23 , 28 , 33 , 34 , 45 , 71 . In crotamine-positive venom-producing animals, the toxin comprises approximately 10%-15% of the venom 31 , 33 , 71 , 72 .
This toxin induces skeletal muscle contraction through its action on sodium channels, interfering with ion permeability in the sarcolemma and reducing the resting potential of the membranes 18 , 28 , 69 . The changes in permeability cause a greater influx of sodium and calcium ions, which are responsible for depolarization, muscle contraction, vacuolization of sarcoplasmic reticulum, rupture of actin and myosin muscle filaments, and muscle necrosis 18 , 28 , 69 , 71 .
Crotamine displays analgesic, antibacterial, antifungal, antiparasitic, and antitumor actions 4 , 7 , 18 , 26 , 28 , 71 , 73 - 76 . It can be classified as a cell-penetrating peptide, a protein transduction domain, a Trojan peptide, or a membrane translocation sequence 18 , 26 , 28 , 68 , 72 .
Translocation across cell membranes occurs by binding between crotamine and cell surface heparan sulfate proteoglycans, endocytosis, and accumulation of the toxin in intracellular vesicles 18 , 28 , 69 - 72 , 75 , 76 . To reach the cytoplasm, crotamine induces changes in the permeability of vesicles, causing it to be released and dispersed in the cell 18 , 69 , 72 , 76 . In the cytoplasm, it can bind to centrosomes in the G1 phase of cell proliferation and enables the diagnosis of cell division phases by functioning as a molecular marker 18 , 68 - 71 .
The antitumor and antimicrobial properties of crotamine are due to its ability to bind to surfaces and acidic cellular compartments such as lysosomes 28 , 74 - 76 . In tumor cells, the prevalence of negatively charged surface molecules, such as phospholipids and mucins, allows preferential binding with the toxin compared to that in healthy cells with electrically neutral surfaces 70 .
To develop new drugs, synthetic analogs of crotamine were produced, composed of peptides with smaller chains, and were used to study their functions in comparison to natural crotamine, revealing the possibility of producing crotamine derivatives with important antimicrobial and antineoplastic functions 18 , 74 .
Crotamine possesses preferential selectivity for proliferating cells and for certain phases of the cell cycle 18 , 69 , 71 , 72 , 74 - 77 . Both in vitro and in vivo studies have demonstrated specific and aggressive cytotoxicity against different tumor types 69 .
The role of crotamine against murine melanoma cells, human melanoma cells, and primary human pancreatic adenocarcinoma cells has been extensively studied 26 , 68 - 71 . Although it is cytotoxic to normal cells when administered at high doses, it is non-toxic at low doses 18 .
Crotamine administered via the intraperitoneal route at a concentration of one microgram per animal per day for 21 days demonstrated efficacy in reducing tumors in rats with subcutaneous melanoma 68 - 72 .
Crotamine’s action mechanisms to induce cell apoptosis include the activation of caspases, the reduction of mitochondrial membrane potential, and consequent alteration of organelle membrane permeability, inducing the release of intracellular calcium ions and the influx of extracellular calcium 28 , 68 , 70 , 71 , 73 , 76 . The activation of caspases is one of the mechanisms responsible for cell apoptosis signaling 78 . Its activation can occur by alterations in mitochondrial membrane permeability, which generates cytochrome C release that amplifies apoptosis signals 69 in HL-60 cells from human leukemia and urinary bladder tumors 69 .
Owing to the ability of the toxin to penetrate cells, it is a potential delivery mechanism for other drugs and antitumor agents 18 , 28 , 68 , 69 , 71 - 74 . In addition to representing a possible antineoplastic or adjuvant therapy, crotamine can be used as a diagnostic marker for cancer 70 , 73 , 76 . Crotamine can be used as a diagnostic marker in human epithelial carcinoma (HeLa), human pancreatic adenocarcinoma (BxPC-3), human breast carcinoma (BT-474), and human colorectal carcinoma (Caco2) cells.
L-AMINO OXIDASES (LAAOS)
LAAOs are flavoenzymes responsible for catalyzing amino acids, which generate alpha-keto acids, ammonia, and hydrogen peroxide 14 , 16 , 17 , 21 , 32 , 79 , 80 . Members of this enzyme class are highly toxic and have great pharmacological importance 16 , 79 , as they can cause platelet aggregation, hemorrhage, edema, cytotoxicity, and induction of apoptosis 14 , 16 , 17 , 37 , 79 , 80 .
These enzymes induce apoptosis in human leukemia cells. Their toxicity is mainly attributed to the formation of hydrogen peroxide during oxidative reactions, among other mechanisms 16 , 21 , 30 , 79 , 80 . Although cytotoxic to tumor cells, LAAOs do not affect healthy cells 21 , 80 .
The species-specific cytotoxicity of LAAOcdt was evaluated using nine human cancer cell lines, including pancreatic, esophageal, cervical, and glioblastoma tumors 80 .
Purified LAAOs can act on different targets of cellular mechanisms such as DNA fragmentation, chromatin condensation, and nuclear fragmentation. Another mechanism is the induction of P53 protein expression, which is synthesized from a tumor suppressor gene that is functionally deficient in more than half of the human tumors 78 , 79 , 81 . Moreover, the induction of protein expression would be relevant to stimulate the monitoring of genome integrity, allowing the identification of damage and repair, resulting in the reduced proliferation of cells with genetic mutations.
LECTINS
Lectins belong to a family of proteins and glycoproteins that generate platelet aggregation 10 , 16 , 20 , 67 . C-type lectins are non-enzymatic calcium-dependent proteins that affect cell adhesion, endocytosis, and neutralization of pathogens 67 . These proteins may also interfere with tumor proliferation, which has been observed using lectins from venom of other species, offering potential for antineoplastic therapy 16 .
METALLOPROTEASES
Metalloproteases present hemorrhagic action and induce coagulation alterations in the prey 16 , 21 , 82 . These proteins are copious in crotalid venom 82 but are present in low quantities in Cdt venom, thus conferring low proteolytic and hemorrhagic activity 33 .
This class of enzymes is composed of endopeptidases that degrade extracellular matrix proteins, blood components, and endothelial cells 21 . In addition, metalloproteases play a fibrinolytic role and act as prothrombin activators, blood coagulation factor X activators, apoptosis inducers, platelet aggregation inhibitors, pro-inflammatory agents, and inactivators of serinoprotease inhibitors 82 . Different groups of metalloproteases found in viperid and crotalid venoms are involved in tumor proliferation and angiogenesis processes 16 . However, specific studies on Cdt venom metalloproteinases have not yet been conducted.
DISINTEGRINS
Disintegrins are also important for the inhibition of tumor cells, together with metalloproteases, by acting against angiogenesis and metastasis 16 . This group of non-enzymatic proteins of low molecular mass can interact with integrins expressed by different cells 16 , 17 , 20 , 83 , important cell surface receptors that are involved in interactions between cells and between cells and the extracellular matrix 16 , 17 , 20 , 21 , 83 .
Aggrastat® (Tirofiban, Merck & Co.) and Integrilin® (Eptifibatide, Cor Therapeutics, now part of Millennium Pharmaceuticals) were developed based on snake disintegrins such as echistatin from the saw-scaled viper Echis carinatus and barbourin from the southeastern pygmy rattlesnake Sistrusus miliarius barbouiri 14 , 20 , 84 .
Integrins, one of the most important targets of antineoplastic action, are cell surface adhesion molecules that function as receptors and transmitters of cellular signals for migration, invasion and cell proliferation 16 , 83 . Inhibition of integrins is important because it affects cell proliferation, angiogenesis, and metastasis and is a widely studied antineoplastic treatment option 16 , 83 .
Disintegrins isolated from Cdt venom inhibit the interaction between tumor cells, impairing their motility, and preventing the invasion of other tissues 21 . One of the mechanisms involved is the deposition of fibrin around the tumor, which limits its growth.
PHOSPHODIESTERASES
These enzymes are less abundant in the venom, representing only approximately 2% of its total 33 . Despite being present in the venom in low quantities, it is responsible for inducing important clinical signs of intoxication 33 , and its antitumor activity has not yet been evaluated.
Figure 2, Figure 3, and Figure 4 summarize the main mechanisms of antineoplastic action for each component of Crotalus durissus terrificus venom.
FIGURE 2: Main antineoplastic actions of Crotalus durissus terrificus venom associated with Crotoxin.
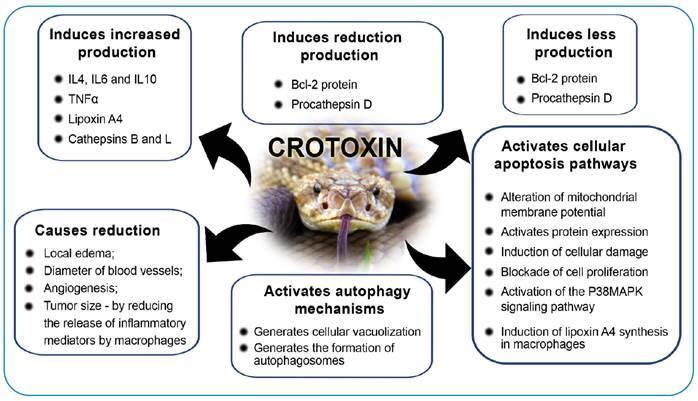
FIGURE 3: Main antineoplastic actions of Crotalus durissus terrificus venom associated with Crotamin.
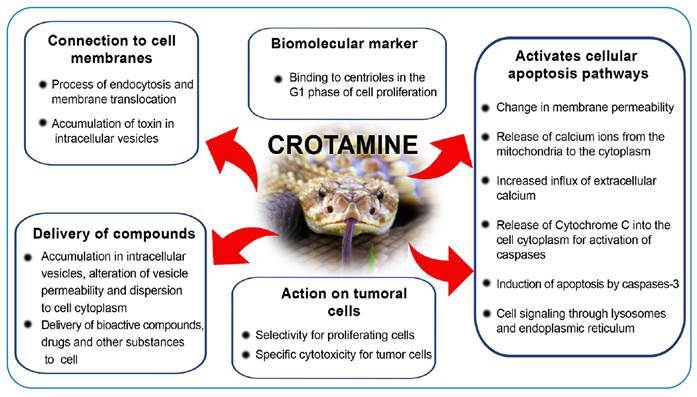
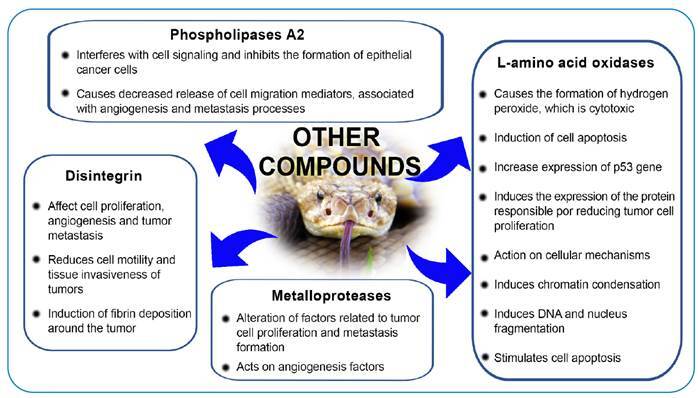
FIGURE 4: Main antineoplastic actions of Crotalus durissus terrificus venom associated with other venom compounds.
CLINICAL TRIALS
Crotoxin was administered to patients with solid tumors that were refractory to standard therapy in a phase 1 clinical trial that observed a partial response of more than 50% reduction in tumor mass and a complete response in three of the 23 evaluated patients 21 , 48 , 77 . The authors concluded that crotoxin is a new class of anticancer agents that acts through a novel mechanism of action and thought that neurotoxicity could be the principal toxic effect and appears to be manageable. They recommended 0.18 mg/m2 a therapeutic dose for Phase II studies 77 .
The same research team proposed an innovative design for a phase 1 trial with intra-patient dose escalation to study crotoxin 85 . As recorded on the clinical trial platform ClinicalTrial.gov, 18 patients were recruited for this study between 2015 and 2018. The researchers stated that the results would be published shortly 86 .
CONCLUSIONS
After elucidating the various mechanisms of action of the C. d. terrificus venom, it may be stated that this venom is a potential candidate for the development of new antineoplastic therapies that are efficient against different tumor lines and act on different cellular targets.
Considering the selective cytotoxicity of venom components for tumor cells to the detriment of healthy cells, the development of innovative therapies against cancer, based on the bioactive compounds of the rattlesnake, may present greater benefits compared to current therapeutic protocols, such as chemotherapy and radiotherapy, which are known to cause alterations in the normal cells of cancer patients.
The therapeutic use of compounds from Crotalus durissus terrificus snake venom also represents an alternative for the treatment of tumors resistant to drugs currently available on the market.
Therefore, one can conclude that the improvement of studies of the different fractions of ophidian venom is of great pharmacological interest, with potential for immense impact on the future of therapeutic medicine.
REFERENCES
- 1.Uetz P, Hošek J. The Reptile Database. Peter Uetz; May 22, 2021. [2020 october 27]. Available from: http://www.reptile-database.org . [Google Scholar]
- 2.Fraga R, Lima AP, Prudente ALC, Magnusson WE. Guide to the snakes of the Manaus region - Central Amazonia. 1st ed. Manaus: Editora Inpa; 2013. 303 p [Google Scholar]
- 3.Mader DR. Reptile Medicine and Surgery. 2nd ed. Elsevier; 2005. 1264 p [Google Scholar]
- 4.Santos IGC, Fortes-Dias CL, Dos-Santos MC. Pharmacological applications of Brazilian snake venoms with emphasis in Crotalus durissus terrificus and Crotalus durissus ruruima. Sci Amazon. 2017;6(1):42–53. [Google Scholar]
- 5.Marques OAV, Medeiros CR. Our Incredible Serpents: Characterization, Biology, Accidents and Conservation. 1st ed. Cotia: Ponto A; 2018. 80 p [Google Scholar]
- 6.Frare BT, Resende YKS, Dornelas BC, Jorge MT, Ricarte VAS, Alves LM, et al. Clinical, Laboratory, and Therapeutic Aspects of Crotalus durissus (South American Rattlesnake) Victims: A Literature Review. Biomed Res Int. 2019 doi: 10.1155/2019/1345923. 1345923:1-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Tasima LJ, Serino-Silva C, Hatakeyama DM, Nishiduka ES, Tashima AK, Sant’Anna SS, et al. Crotamine in Crotalus durissus: distribution according to subspecies and geographic origin, in captivity or nature. J Venom Anim Toxins Incl Trop Dis. 2020;26:e20190053. doi: 10.1590/1678-9199-jvatitd-2019-0053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Rangel-Santos A, Dos-Santos EC, Lopes-Ferreira M, Lima C, Cardoso DF, Mota I. A comparative study of biological activities of crotoxin and CB fraction of venoms from Crotalus durissus terrificus, Crotalus durissus cascavella and Crotalus durissus collilineatus. Toxicon. 2004;43(7):801–810. doi: 10.1016/j.toxicon.2004.03.011. [DOI] [PubMed] [Google Scholar]
- 9.Wüster W, Ferguson JE, Quijada-Mascareñas JA, Pook CE, Salomão MDG, Thorpe RS. Tracing an invasion: Landbridges, refugia, and the phylogeography of the Neotropical rattlesnake (Serpentes: Viperidae: Crotalus durissus) Mol Ecol. 2005;14(4):1095–1080. doi: 10.1111/j.1365-294X.2005.02471.x. [DOI] [PubMed] [Google Scholar]
- 10.Fry BG, Sunagar K, Casewell NR, Kochva E, Roelants K, Scheib H, et al. The origin and evolution of the toxicofera reptile venom system. In: Fry BG. Venomous reptiles & their toxins: Evolution, pathophysiology & biodiscovery. New York: Oxford University Press; 2015. pp. 1–31. [Google Scholar]
- 11.WHO - World Health Organization . Report of the Tenth Meeting of the WHO Strategic and Technical Advisory Group for Neglected Tropical Diseases. Geneva: WHO; 2017. [2020 Oct 27]. Available from: http://www.who.int/neglected_diseases/NTD_STAG_report_2017.pdf?ua=1 . [Google Scholar]
- 12.Grego KF, Vieira SEM, Vidueiros JP, Serapicos EO, Barbarini CC, Silveira GPM, et al. Maintenance of venomous snakes in captivity for venom production at Butantan Institute from 1908 to the present: a scoping history. J Venom Anim Toxins Incl Trop Dis. 2021;27:e20200068. doi: 10.1590/1678-9199-jvatitd-2020-0068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chippaux JP. Snakebite envenomation turns again into a neglected tropical disease. J Venom Anim Toxins Incl Trop Dis. 2017;8(23):38–38. doi: 10.1186/s40409-017-0127-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.El-Aziz TMA, Soares AG, Stockand JD. Snake Venoms in Drug Discovery: Valuable Therapeutic Tools for Life Saving. Toxins. 2019;11(10):1–25. doi: 10.3390/toxins11100564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Almeida JR, Resende LM, Watanabe RK, Carregari VC, Huancahuire-Vega S, Caldeira CAS, et al. Snake Venom Peptides and Low Mass Proteins: Molecular Tools and Therapeutic Agents. Curr Med Chem. 2017;24(30):3254–3282. doi: 10.2174/0929867323666161028155611. [DOI] [PubMed] [Google Scholar]
- 16.Calderon LA, Sobrinho JC, Zaqueo KD, Moura AA, Grabner NA, Mazzi MV, et al. Antitumoral activity of snake venom proteins: New trends in cancer therapy. BioMed Res Int. 2014;2014:203639–203639. doi: 10.1155/2014/203639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Gomes A, Bhattacharjee P, Mishra R, Biswas AK, Dasgupta SC, Giri B. Anticancer potential of animal venoms and toxins. Indian J Exp Biol. 2010;48(2):93–103. [PubMed] [Google Scholar]
- 18.Kerkis I, Silva F, Pereira A, Kerkis A, Rádis-Baptista G. Biological versatility of crotamine--a cationic peptide from the venom of a South American rattlesnake. Expert Opin Investig Drugs. 2010;19(12):1515–1525. doi: 10.1517/13543784.2010.534457. [DOI] [PubMed] [Google Scholar]
- 19.Soares MA, Pujatti PB, Fortes-Dias CI, Antonelli L, Santos RG. Crotalus durissus terrificus venom as a source of antitumoral agents. J Venom Anim Toxins Incl Trop Dis. 2010;16(3):480–492. doi: 10.1590/S1678-91992010000300015. [DOI] [Google Scholar]
- 20.Vonk FJ, Jackson K, Doley R, Madaras F, Mirtschin PJ, Vidal N. Snake venom: From fieldwork to the clinic: Recent insights into snake biology, together with new technology allowing high-throughput screening of venom, bring new hope for drug discovery. Bioessays. 2011;33(4):269–279. doi: 10.1002/bies.201000117. [DOI] [PubMed] [Google Scholar]
- 21.Jain D, Kumar S. Snake venom: a potent anticancer agent. Asian Pac J Cancer Prev. 2012;13(10):4855–4860. doi: 10.7314/APJCP.2012.13.10.4855. [DOI] [PubMed] [Google Scholar]
- 22.Kollipara PS, Won DH, Hwang CJ, Jung YY, Yoon HS, Park MH, et al. Enhanced Anti-Cancer Effect of Snake Venom Activated NK Cells on Lung Cancer Cells by Inactivation of NF-κB. Biomol Ther. 2014;22(2):106–113. doi: 10.4062/biomolther.2013.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Melani RD, Araujo GDT, Carvalho PC, Goto L, Nogueira FCS, Junqueira M, et al. Seeing beyond the tip of the icebeg: A deep analysis of the venome of the Brazilian Rattlesnake, Crotalus durissus terrificus. EuPA Open Proteom. 2015;8:144–156. doi: 10.1016/j.euprot.2015.05.006. [DOI] [Google Scholar]
- 24.Boldrini-França J, Cologna CT, Pucca MB, Bordon KCF, Amorim FG, Anjolette FAP, et al. Minor snake venom proteins: Structure, function and potential applications. Biochim Biophys Acta Gen Subj. 2017;1861(4):824–838. doi: 10.1016/j.bbagen.2016.12.022. [DOI] [PubMed] [Google Scholar]
- 25.Sousa IDL, Barbosa AR, Salvador GHM, Frihling BEF, Santa-Rita PH, Soares AM, et al. Secondary homeostasis studies of crude venom and isolated proteins from the snake Crotalus durissus terrificus. Int J Biol Macromol. 2019;131:127–133. doi: 10.1016/j.ijbiomac.2019.03.059. [DOI] [PubMed] [Google Scholar]
- 26.Pérez-Peinado C, Defaus S, Andreu D. Hitchhiking with Nature: Snake Venom Peptides to Fight Cancer and Superbugs. Toxins. 2020;12(4):1–23. doi: 10.3390/toxins12040255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Santos L, Oliveira C, Vasconcelos BM, Vilela D, Melo L, Ambrósio L, et al. Good management practices of venomous snakes in captivity to produce biological venom-based medicines: achieving replicability and contributing to pharmaceutical industry. J Toxicol Environ Health B Crit Rev. 2021;24(1):30–50. doi: 10.1080/10937404.2020.1855279. [DOI] [PubMed] [Google Scholar]
- 28.Cunha DB, Silvestrini AVP, Silva ACG, Estevam DMP, Pollettini FL, Navarro JO, et al. Mechanistic insights into functional characteristics of native crotamine. Toxicon. 2018;146:1–12. doi: 10.1016/j.toxicon.2018.03.007. [DOI] [PubMed] [Google Scholar]
- 29.Vyas VK, Brahmbhatt K, Bhatt H, Parmar U. Therapeutic potential of snake venom in cancer therapy: current perspectives. Asian Pac J Trop Biomed. 2013;3(2):156–162. doi: 10.1016/S2221-1691(13)60042-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Shanbhag VKL. Applications of snake venoms in treatment of cancer. Asian Pac J Trop Biomed. 2015;5(4):275–276. doi: 10.1016/S2221-1691(15)30344-0. [DOI] [Google Scholar]
- 31.Devi A. In: Venomous Animals and their Venoms. 1st ed. Bucherl W, Buckley EE, Deulofeu V, editors. New York: Academic Press; 1968. The Protein and Nonprotein Constituents of Snake Venoms; pp. 119–165. [DOI] [Google Scholar]
- 32.Sarkar NK, Devi A. Bucherl W, Buckley EE, Deulofeu V. Enzymes in Snake Venoms. New York: Academic Press; 1968. pp. 167–216. [DOI] [Google Scholar]
- 33.Fusco LS, Neto EB, Francisco AF, Alfonso J, Soares A, Pimenta DC, et al. Fast venomic analysis of Crotalus durissus terrificus from northeastern Argentina. Toxicon X. 2020;7:100047–100047. doi: 10.1016/j.toxcx.2020.100047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lourenço A, Jr, Creste CFZ, Barros LC, dos Santos LC, Pimenta DC, Barraviera B, et al. Individual venom profiling of Crotalus durissus terrificus specimens from a geographically limited region: Crotamine assessment and captivity evaluation on the biological activities. Toxicon. 2013;69:75–81. doi: 10.1016/j.toxicon.2013.01.006. [DOI] [PubMed] [Google Scholar]
- 35.Montoni F, Andreotti DZ, Eichler RAS, Santos WS, Kisaki CY, Arcos SSS, et al. The impact of rattlesnake venom on mice cerebellum proteomics points to synaptic inhibition and tissue damage. J Proteomics. 2020;221:103779–103779. doi: 10.1016/j.jprot.2020.103779. [DOI] [PubMed] [Google Scholar]
- 36.Lima TS, Neves CL, Zambelli VO, Lopes F, Sampaio SC, Cirillo MC. Crotoxin, a rattlesnake toxin, down-modulates functions of bone marrow neutrophils and impairs the Syk-GTPase pathway. Toxicon. 2017;136:44–55. doi: 10.1016/j.toxicon.2017.07.002. [DOI] [PubMed] [Google Scholar]
- 37.Tasima LJ, Hatakeyama DM, Serino-Silva C, Rodrigues C, de Lima E, Sant'Anna SS, et al. Comparative proteomic profiling and functional characterization of venom pooled from captive Crotalus durissus terrificus specimens and the Brazilian crotalic reference venom. Toxicon. 2020;185:26–35. doi: 10.1016/j.toxicon.2020.07.001. [DOI] [PubMed] [Google Scholar]
- 38.Corin RE, Viskatis LJ, Vidal JC, Etcheverry MA. Cytotoxicity of crotoxin on murine erythroleukemia cells in vitro. Invest New Drugs. 1993;11(1):11–15. doi: 10.1007/BF00873905. [DOI] [PubMed] [Google Scholar]
- 39.Bordon KCF, Perino MG, Giglio JR, Arantes EC. Isolation, enzymatic characterization and antiedematogenic activity of the first reported rattlesnake hyaluronidase from Crotalus durissus terrificus venom. Biochimie. 2012;94(12):2740–2748. doi: 10.1016/j.biochi.2012.08.014. [DOI] [PubMed] [Google Scholar]
- 40.Faure G, Bakouh N, Lourdel S, Odolczyk N, Premchandar A, Servel N, et al. Rattlesnake Phospholipase A2 increases CFTR-chloride channel current and corrects F508CFTR dysfunction: impact in Cystic Fibrosis. J Mol Biol. 2016;428(14):2898–2915. doi: 10.1016/j.jmb.2016.05.016. [DOI] [PubMed] [Google Scholar]
- 41.Marcussi S, Santos PRS, Menaldo DL, Silveira LB, Santos-Filho NA, Mazzi MV, et al. Evaluation of the genotoxicity of Crotalus durissus terrificus snake venom and its isolated toxins on human lymphocytes. Mutat Res. 2011;724(1-2):59–63. doi: 10.1016/j.mrgentox.2011.06.004. [DOI] [PubMed] [Google Scholar]
- 42.Muller VDM, Russo RR, Cintra ACO, Sartim MA, Alves-Paiva RM, Figueiredo LTM, et al. Crotoxin and phospholipases A₂ from Crotalus durissus terrificus showed antiviral activity against dengue and yellow fever viruses. Toxicon. 2012;59(4):507–515. doi: 10.1016/j.toxicon.2011.05.021. [DOI] [PubMed] [Google Scholar]
- 43.Sartim MA, Menaldo DL, Sampaio SV. Immunotherapeutic potential of crotoxin: anti-inflammatory and immunosuppressive properties. J Venom Anim Toxins Incl Trop Dis. 2018;24:1–13. doi: 10.1186/s40409-018-0178-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Pimenta LA, Almeida MES, Bretones ML, Cirillo MC, Curi R, Sampaio SC. Crotoxin promotes macrophage reprogramming towards an antiangiogenic phenotype. Sci Rep. 2019;9(1):1–15. doi: 10.1038/s41598-019-40903-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Silva-Júnior LN, Abreu LS, Rodrigues CFB, Galizio NC, Aguiar WS, Serino-Silva C, et al. Geographic variation of individual venom profile of Crotalus durissus snakes. J Venom Anim Toxins Incl Trop Dis. 2020;26:e20200016. doi: 10.1590/1678-9199-jvatitd-2020-0016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Sampaio SC, Brigatte P, Sousa-e-Silva MCC, dos-Santos EC, Rangel-Santos AC, Curi R, et al. Contribution of crotoxin for the inhibitory effect of Crotalus durissus terrificus snake venom on macrophage function. Toxicon. 2003;41(7):899–907. doi: 10.1016/S0041-0101(03)00069-2. [DOI] [PubMed] [Google Scholar]
- 47.Yang C, Yang Y, Qin Z, Gu Z, Reid P, Liang Z. Autophagy is involved in cytotoxic effects of crotoxin in human breast cancer cell line MCF-7 cells. Acta Pharmacol Sin. 2007;28(4):540–548. doi: 10.1111/j.1745-7254.2007.00530.x. [DOI] [PubMed] [Google Scholar]
- 48.Sampaio SC, Hyslop S, Fontes MRM, Prado-Francechi J, Zambelli VO, Magro AJ, et al. Crotoxin: Novel activities for a classic β-neurotoxin. Toxicon. 2010;55(6):1045–1060. doi: 10.1016/j.toxicon.2010.01.011. [DOI] [PubMed] [Google Scholar]
- 49.Ye B, Xie Y, Qin ZH, Wu JC, Han R, He JK. Anti-tumor activity of CrTX in human lung adenocarcinoma cell line A549. Acta Pharmacol Sinica. 2011;32(11):1397–1401. doi: 10.1038/aps.2011.116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Wang J, Xie Y, Wu J, Han R, Reid PF, Qin Z, et al. Crotoxin enhances the antitumor activity of gefinitib (Iressa) in SK-MES-1 human lung squamous carcinoma cells. Oncol Rep. 2012;27(5):1341–1347. doi: 10.3892/or.2012.1677. [DOI] [PubMed] [Google Scholar]
- 51.Brigatte P, Faiad OJ, Nocelli RCF, Landgraf RG, Palma MS, Cury Y, et al. Walker 256 Tumor Growth Suppression by Crotoxin Involves Formyl Peptide Receptors and Lipoxin A₄. Mediators Inflamm. 2016;2016:2457532–2457532. doi: 10.1155/2016/2457532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Cavalcante WLG, Matos JBN, Timoteo MA, Fontes MRM, Gallacci M, de Sa PC. Neuromuscular paralysis by the basic phospholipase A2 subunit crotoxin from Crotalus durissus terrificus snake venom needs its acid chaperone to con-currently inhibit acetylcholine release and produce muscle blockage. Toxicol Appl Pharmacol. 2017;334:8–17. doi: 10.1016/j.taap.2017.08.021. [DOI] [PubMed] [Google Scholar]
- 53.Muller SP, Silva V, Silvestrini A, Macedo LH, Caetano GF, Reis RM, et al. Crotoxin from Crotalus durissus terrificus venom: In vitro cytotoxic activity of a heterodimeric phospholipase A2 on human cancer-derived cell lines. Toxicon. 2018;156:13–22. doi: 10.1016/j.toxicon.2018.10.306. [DOI] [PubMed] [Google Scholar]
- 54.Andrade CM, Rey FM, Cintra A, Sampaio SV, Torqueti MR. Effects of crotoxin, a neurotoxin from Crotalus durissus terrificus snake venom, on human endothelial cells. Int J Biol Macromol. 2019;134:613–621. doi: 10.1016/j.ijbiomac.2019.05.019. [DOI] [PubMed] [Google Scholar]
- 55.He J, Wu X, Wang Y, Han R, Qin Z, Xie Y. Growth inhibitory effects and molecular mechanisms of crotoxin treatment in esophageal Eca-109 cells and transplanted tumors in nude mice. Acta pharmacol Sin. 2013;34(2):295–300. doi: 10.1038/aps.2012.156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Donato NJ, Martin CA, Perez M, Newman RA, Vidal JC, Etcheverry M. Regulation of epidermal growth factor receptor activity by crotoxin, a snake venom phospholipase A2 toxin. A novel growth inhibitory mechanism. Biochem pharmacol. 1996;51(11):1535–1543. doi: 10.1016/0006-2952(96)00097-4. [DOI] [PubMed] [Google Scholar]
- 57.Han R, Liang H, Qin ZH, Liu CY. Crotoxin induces apoptosis and autophagy in human lung carcinoma cells in vitro via activation of the p38MAPK signaling pathway. Acta Pharmacol Sin. 2014;35(10):1323–1332. doi: 10.1038/aps.2014.62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Cotrim CA, Oliveira SCB, Filho EBSD, Fonseca FV, Jr LB, Antunes E, et al. Quercetin as an inhibitor of snake venom secretory phospholipase A2. Chem Biol Interact. 2011;189(1-2):9–16. doi: 10.1016/j.cbi.2010.10.016. [DOI] [PubMed] [Google Scholar]
- 59.Sunagar K, Jackson TNW, Reeks T, Fry BG. Fry BG. Venomous reptiles & their toxins: Evolution, pathophysiology & biodiscovery. New York: Oxford University Press; 2015. Group I phospholipase A2 enzymes; pp. 327–334. [Google Scholar]
- 60.Sunagar K, Tsai IH, Lomonte B, Jackson TNW, Fry BG. Fry BG. Venomous reptiles & their toxins: Evolution, pathophysiology & biodiscovery. New York: Oxford University Press; 2015. Group II phospholipase A2 enzymes; pp. 335–340. [Google Scholar]
- 61.Herbst RS. Review of epidermal growth factor receptor biology. Int J Radiat Oncol Biol Phys. 2004;59(2):2–26. doi: 10.1016/j.ijrobp.2003.11.041. [DOI] [PubMed] [Google Scholar]
- 62.Sharma SV, Bell DW, Settleman J, Haber DA. Epidermal growth factor receptor mutations in lung cancer. Nat Rev Cancer. 2007;7:169–181. doi: 10.1038/nrc2088. [DOI] [PubMed] [Google Scholar]
- 63.Ferreira RS, de Barros LC, Abbade LPF, Barraviera SRCS, Silvares MRC, de Pontes LG, et al. Heterologous fibrin sealant derived from snake venom: from bench to bedside - an overview. J Venom Anim Toxins Incl Trop Dis. 2017;23:21–21. doi: 10.1186/s40409-017-0109-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Abbade LPF, Barraviera SRCS, Silvares MRC, Lima ABBCO, Haddad GR, Gatti MAN, et al. Treatment of Chronic Venous Ulcers with Heterologous Fibrin Sealant: A Phase I/II Clinical Trial. Front Immunol. 2021;12:627541–627541. doi: 10.3389/fimmu.2021.627541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Abbade LPF, Ferreira RS, Jr, dos Santos LD, Barraviera B. Chronic venous ulcers: a review on treatment with fibrin sealant and prognostic advances using proteomic strategies. J Venom Anim Toxins Incl Trop Dis. 2020;26:e20190101. doi: 10.1590/1678-9199-jvatitd-2019-0101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Francischetti IMB, Ghazaleh FA, Reis RAM, Carlini CR, Guimarães JA. Convulxin induces platelet activation by a tyrosine-kinase-dependent pathway and stimulates tyrosine phosphorylation of platelet proteins, including PLCγ2, independently of integrin αIIBβ3. Arch biochem biophys. 1998;353(2):239–250. doi: 10.1006/abbi.1998.0598. [DOI] [PubMed] [Google Scholar]
- 67.Arlinghaus FT, Fry BG, Sunagar K, Jackson TNW, Eble JA, Reeks T, et al. Fry BG. Lectin proteins. New York: Oxford University Press; 2015. pp. 299–311. [Google Scholar]
- 68.Kerkis I, Hayashi MAF, da Silva ARBP, Pereira A, Júnior PLS, Zaharenko AJ, et al. State of the art in the studies on crotamine, a cell penetrating peptide from South American rattlesnake. Biomed Res Int. 2014;2014:675985–675985. doi: 10.1155/2014/675985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Rádis-Baptista G, Kerkis I. Crotamine, a small basic polypeptide myotoxin from rattlesnake venom with cell-penetrating properties. Curr Pharm Des. 2011;17(38):4351–4361. doi: 10.2174/138161211798999429. [DOI] [PubMed] [Google Scholar]
- 70.Falcao CB, Rádis-Baptista G. Crotamine and crotalicidin, membrane active peptides from Crotalus durissus terrificus rattlesnake venom, and their structurally-minimized fragments for applications in medicine and biotechnology. Peptides. 2020;126:170234–170234. doi: 10.1016/j.peptides.2019.170234. [DOI] [PubMed] [Google Scholar]
- 71.Silva ARBP, Fry BG, Sunagar K, Scheib H, Jackson TNW, Rádis-Baptista G, Zaharenko AJ, Jr PLS, Pereira A, Oguiura N, Hayashi MAF, Kerkis A, Yamane T, Kerkis I. Beta-defensins. In: Fry BG. Venomous reptiles & their toxins: Evolution, pathophysiology & biodiscovery. New York: Oxford University Press; 2015. pp. 228–238. [Google Scholar]
- 72.Campeiro JD, Marinovic MP, Carapeto FC, Mas CD, Monte GG, Porta LC, et al. Oral treatment with a rattlesnake native polypeptide crotamine efficiently inhibits the tumor growth with no potential toxicity for the host animal and with suggestive positive effects on animal metabolic profile. Amino Acids. 2018;50(2):267–278. doi: 10.1007/s00726-017-2513-3. [DOI] [PubMed] [Google Scholar]
- 73.Vu TTT, Jeong B, Yu J, Koo B, Jo S, Robinson RC, et al. Soluble prokaryotic expression and purification of crotamine using an N-terminal maltose-binding protein tag. Toxicon. 2014;92:157–165. doi: 10.1016/j.toxicon.2014.10.017. [DOI] [PubMed] [Google Scholar]
- 74.Dal Mas C, Pinheiro DA, Campeiro JD, Mattei B, Oliveira V, Oliveira EB, et al. Biophysical and biological properties of small linear peptides derived from crotamine, a cationic antimicrobial/antitumoral toxin with cell penetrating and cargo delivery abilities. Biochim Biophys Acta Biomembr. 2017;1859(12):2340–2349. doi: 10.1016/j.bbamem.2017.09.006. [DOI] [PubMed] [Google Scholar]
- 75.Yamane ES, Bizerra FC, Oliveira EB, Moreira JT, Rajabi M, Nunes GL, et al. Unraveling the antifungal activity of a South American rattlesnake toxin crotamine. Biochimie. 2013;95(2):231–240. doi: 10.1016/j.biochi.2012.09.019. [DOI] [PubMed] [Google Scholar]
- 76.Nascimento FD, Sancey L, Pereira A, Rome C, Oliveira V, Oliveira EB, et al. The natural cell-penetrating peptide crotamine targets tumor tissue in vivo and triggers a lethal calcium-dependent pathway in cultured cells. Mol Pharm. 2012;9(2):211–221. doi: 10.1021/mp2000605. [DOI] [PubMed] [Google Scholar]
- 77.Cura JE, Blanzaco DP, Brisson C, Cura MA, Cabrol R, Larrateguy L, et al. Phase I and pharmacokinetics study of crotoxin (cytotoxic PLA(2), NSC-624244) in patients with advanced cancer. Clin Cancer Res. 2002;8(4):1033–1041. [PubMed] [Google Scholar]
- 78.Anazetti MC, Melo PS. Apoptosis Cell Death: biochemistry and molecular aspects. Metrocamp Pesquisa. 2007;1(1):37–58. [Google Scholar]
- 79.Tan NH, Fry BG, Sunagar K, Jackson TNW, Reeks T, Fung SY. Fry BG. Venomous reptiles & their toxins: Evolution, pathophysiology & biodiscovery. New York: Oxford University Press; 2015. L-Amino acid oxidase enzymes; pp. 291–298. [Google Scholar]
- 80.Teixeira TL, Silva VAO, Cunha DB, Polettini FL, Thomaz CD, Pianca AA, et al. Isolation, characterization and screening of the in vitro cytotoxic activity of a novel L-amino acid oxidase (LAAOcdt) from Crotalus durissus terrificus venom on human cancer cell lines. Toxicon. 2016;119:203–217. doi: 10.1016/j.toxicon.2016.06.009. [DOI] [PubMed] [Google Scholar]
- 81.Fett-Conte AC, Salles ABCF. The importance of the p53 gene in human carcinogenesis. Rev Bras Hematol Hemoter. 2002;24(2):85–89. doi: 10.1590/S1516-84842002000200004. [DOI] [Google Scholar]
- 82.Markland JR, Francis S, Swenson S. Snake venom metalloproteinases. Toxicon. 2013;62:3–18. doi: 10.1016/j.toxicon.2012.09.004. [DOI] [PubMed] [Google Scholar]
- 83.Galán JA, Sánchez EE, Rodríguez-Acosta A, Soto JG, Bashir S, McLane MA, et al. Inhibition of lung tumor colonization and cell migration with the disintegrin crotatroxin 2 isolated from the venom of Crotalus atrox. Toxicon. 2008;51(7):1186–1196. doi: 10.1016/j.toxicon.2008.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Pennington MW, Czerwinski A, Norton RS. Peptide therapeutics from venom: Current status and potential. Bioorg Med Chem. 2018;26(10):2738–2758. doi: 10.1016/j.bmc.2017.09.029. [DOI] [PubMed] [Google Scholar]
- 85.Medioni J, Brizard M, Elaidi R, Reid PF, Benlhassan K, Bray D. Innovative design for a phase 1 trial with intra-patient dose escalation: The Crotoxin study. Contemp Clin Trials Commun. 2017;7:186–188. doi: 10.1016/j.conctc.2017.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Gil-Delgado MA. Identifier NCT01481532, Open Label Clinical Trial of Intravenous Crotoxin. National Library of Medicine (US); Nov 28, 2011. [2022 Sep 21]. Available from: https://clinicaltrials.gov/ct2/history/NCT01481532?V_5=View#StudyPageTop . [Google Scholar]