

Abstract
Objective:
Previous research has demonstrated that the amygdala is enlarged in children with autism spectrum disorder (ASD). However, the precise onset of this enlargement during infancy, how it relates to later diagnostic behaviors, whether the timing of enlargement in infancy is specific to the amygdala, and whether it is specific to ASD (or present in other neurodevelopmental disorders, such as fragile X syndrome) are all unknown.
Methods:
Longitudinal MRIs were acquired at 6–24 months of age in 29 infants with fragile X syndrome, 58 infants at high likelihood for ASD who were later diagnosed with ASD, 212 high-likelihood infants not diagnosed with ASD, and 109 control infants (1,099 total scans).
Results:
Infants who developed ASD had typically sized amygdala volumes at 6 months, but exhibited significantly faster amygdala growth between 6 and 24 months, such that by 12 months the ASD group had significantly larger amygdala volume (Cohen’s d=0.56) compared with all other groups. Amygdala growth rate between 6 and 12 months was significantly associated with greater social deficits at 24 months when the infants were diagnosed with ASD. Infants with fragile X syndrome had a persistent and significantly enlarged caudate volume at all ages between 6 and 24 months (d=2.12), compared with all other groups, which was significantly associated with greater repetitive behaviors.
Conclusions:
This is the first MRI study comparing fragile X syndrome and ASD in infancy, demonstrating strikingly different patterns of brain and behavior development. Fragile X syndrome–related changes were present from 6 months of age, whereas ASD-related changes unfolded over the first 2 years of life, starting with no detectable group differences at 6 months. Increased amygdala growth rate between 6 and 12 months occurs prior to social deficits and well before diagnosis. This gradual onset of brain and behavior changes in ASD, but not fragile X syndrome, suggests an age- and disorder-specific pattern of cascading brain changes preceding autism diagnosis.
The defining behavioral features of autism spectrum disorder (ASD) emerge over the latter part of the first and second years of life, and generally do not consolidate into the full syndrome, enabling clinical diagnosis, until 24–36 months of age (1–4). Many behaviors characteristic of ASD are indistinguishable at 6 months between infants later diagnosed with ASD and those with typical development (2, 4). Group differences in behaviors begin to emerge by 12 months (5–8). These emerging changes in early behavioral development parallel dynamic changes in brain development in ASD during infancy. For example, we previously reported hyperexpansion of cortical surface area from 6 to 12 months of age in infants who were later diagnosed with ASD (9), consistent with cross-sectional reports of increased cortical surface area in later childhood (10). Similarly, we have described age-specific differences in the development of white matter microstructure (i.e., fractional anisotropy), observed to be increased in infants later diagnosed with ASD compared with control infants at 6 months, but showing no difference at 12 months, and then decreased at 24 months (11). We have also noted developmental changes in the corpus callosum, with increased volume of the anterior corpus callosum at 6 months that normalized in size by 24 months (12), consistent with reports of decreased corpus callosum size in cross-sectional studies at older ages (13). The trajectory of extra-axial CSF volume represents another developmental pattern of being persistently increased from 6 months of age in children diagnosed with ASD (14, 15) through 4 years of age (16).
Taken together, these studies illustrate a range of age-specific changes in early postnatal brain development in ASD, along with dynamic changes in behavior. This suggests that early, presymptomatic brain changes in infancy may represent a cascade of linked brain and behavior changes that lead to the emergence of the full syndrome of autism, consolidating into a clinically diagnosable condition in the 2nd and 3rd years of life (17). Further delineation of the nature and sequence of brain changes would provide important clues to elucidating the pathogenesis underlying this condition and could inform the development of targeted interventions aimed at these developmental trajectories.
Although subcortical structures, and in particular the amygdala, have long been implicated in ASD through structural and functional neuroimaging and postmortem studies, no studies have examined the nature and timing of subcortical brain development during infancy in ASD. Neuroimaging studies have demonstrated amygdala enlargement in 2- to 4-year-old children with ASD (18–22), and postmortem studies indicate an excess number of amygdala neurons (23) and increased dendritic spine density (24) as possible cellular processes contributing to early amygdala overgrowth. However, the vast majority of neuroimaging studies are cross-sectional and are conducted on children after diagnosis (i.e., 2 years of age and older), leaving unknown the developmental time course of amygdala enlargement, how it relates to the timing of emerging diagnostic features and eventual diagnosis, and whether enlargement is specific to the amygdala or also occurs during infancy in other subcortical structures, such as the basal ganglia. In addition, neuroimaging studies in infants who develop ASD have not examined the specificity of brain findings to ASD in relationship to other neurodevelopmental disorders.
In this study, we examined longitudinal structural MRIs of selected subcortical structures (amygdala, caudate, putamen, globus pallidus, thalamus) to contrast the early postnatal brain development of four groups: infants with fragile X syndrome (FXS); infants at high likelihood for ASD (due to having an older sibling with autism) who later developed ASD; high-likelihood infants who did not develop ASD; and control infants at lower likelihood who had typical development. The study design examined the issue of disorder specificity by contrasting the brain and behavioral development of idiopathic ASD, a behaviorally defined developmental disorder, with a genetically defined disorder, FXS, which has overlapping cognitive and behavioral features (25). In addition, we note the distinction that this study contrasts infants with familial autism (a subgroup of ASD whose etiology is most often attributed to common polygenic inheritance [26]), with FXS—a genetic developmental disorder and the most common inherited cause of intellectual disability, occurring in approximately 1 in every 2,000–5,000 live births (27). FXS is caused by a mutation on the X chromosome, resulting in the disrupted expression of the fragile X mental retardation gene 1 (FMR1) (27). The FMR1 mutation causes diminished production of the protein FMRP, which plays a vital role in neuronal development and synaptic plasticity and is highly expressed in the caudate (28–31).
Although idiopathic ASD and FXS share some overlapping behavioral features, this is the first study to compare their brain growth trajectories in infancy. Previous research has demonstrated that the caudate (but not the amygdala) is enlarged in 2- to 4-year-old children with FXS (18, 27, 32) and, conversely, that the amygdala (but not the caudate) is enlarged in children with idiopathic ASD at the same age (18–22). Importantly, the neural phenotype of FXS was consistent regardless of whether FXS children met behavioral criteria for ASD (18, 32). The timing and onset of these brain changes in infancy, and their relationship to behavior, are unknown. Contrasting developmental trajectories of key subcortical brain structures, along with their relationship to relevant behaviors during early postnatal development, will elucidate our understanding of the pathogenesis of both conditions and may provide important clues to rational approaches to targeted early intervention.
This study was guided by the following hypotheses: 1) infants later diagnosed with ASD would exhibit amygdala enlargement; 2) infants with FXS would exhibit caudate enlargement; and 3) amygdala enlargement in ASD would be associated with social deficits, and caudate enlargement in FXS would be associated with greater repetitive behaviors, based on the existing literature.
METHODS
Participants
Infants with FXS were enrolled, and the presence of full-mutation FXS was verified by medical records or genetic testing (polymerase chain reaction and Southern blot). High-likelihood (HL) infants were defined by having an older sibling with a diagnosis of ASD made by their clinical provider, corroborated by the Autism Diagnostic Interview–Revised (ADI-R) (33). Infants at lower likelihood (LL) had a typically developing older sibling and no siblings with ASD at the time of enrollment (1). Inclusion and exclusion criteria have been detailed elsewhere (34, 35) (see also the online supplement). Parents provided informed consent, and the institutional review board at each site approved the research protocol.
Behavioral Assessment
Infants underwent MRI scanning between 6 and 24 months of age. At 24 months, infants were evaluated with an assessment of cognitive ability (the Mullen Scales of Early Learning [36]) and the Repetitive Behavior Scale–Revised (RBS-R) (37), a questionnaire of repetitive behavior with demonstrated clinical utility in this sample (5, 38, 39). HL infants were classified at 24 months as either having ASD or not, based on expert clinical judgment using DSM-IV-TR criteria (40), the Autism Diagnostic Observation Schedule–Generic (ADOS) (41), and the ADI-R.
This yielded four outcome groups: 1) infants with FXS (N=29; 23 of them male); 2) HL infants who were diagnosed with ASD (HL-ASD: N=58; 46 of them male); 3) HL infants who were negative for ASD (HL-negative: N=212; 119 of them male); and 4) LL infants who were negative for ASD (LL-negative: N=109; 66 of them male). (For participant characteristics by group, see Table S1 in the online supplement.) This study of infants 6–24 months of age did not classify the FXS group into subsets according to whether they met diagnostic criteria for ASD. Because of low cognitive and social ability, the ADOS is not consistently reliable or valid in 24-month-olds with FXS (42); in the present sample, the ADOS at 24 months was available for only eight participants with FXS for whom brain data were available. Therefore, ASD diagnosis could not be confidently obtained in the FXS group. Importantly, in our previous neuroimaging studies of older children with FXS (in whom reliable ASD classification could be made), the distinct brain features of the FXS group were driven by the FXS genetic mutation itself and not by whether the individuals with FXS met criteria for ASD (i.e., FXS children with and without ASD were indistinguishable from each other) (18, 32).
MRI Acquisition and Processing
A total of 1,099 scans were collected between ages 6 and 24 months across the four groups (see Table S2 in the online supplement for the breakdown by group and time point). T1- and T2-weighted scans (1-mm3 voxels) underwent standard preprocessing (43–45). A graph-based multi-atlas segmentation method developed by our laboratory (46) was employed in the data set of infant MRIs by generating study- and age-specific atlases at ages 6, 12, and 24 months to segment the subcortical structures (46). (See reference 46 and the online supplement for complete details, and Figure S1 in the online supplement for an illustration of the segmentations.)
Statistical Analysis
A longitudinal mixed-effects model for repeated measures with unstructured covariance matrices was employed to analyze trajectories of subcortical structures from ages 6 to 24 months. Independent variables of interest included main effect of group, linear effect of age and sex, and group interactions with each of these variables. Total cerebral volume was included as a covariate given its relationship to subcortical volumes, and to control for brain size. Scan site was included as another control variable. The random effects were each individual’s age at each time point. Following significant omnibus results of the primary model described above, pairwise comparisons tested for cross-sectional group differences at each time point (6, 12, and 24 months) and were corrected for multiple comparisons (Tukey-Kramer). Percent differences in model-adjusted volumes at each time point and effect sizes (Cohen’s d) are reported relative to the LL-negative group. In the ASD group, linear regression was used to test whether amygdala volume growth rate between 6 and 12 months was associated with social deficits at 24 months (ADOS social affect calibrated severity score) or, alternatively, with restricted and repetitive behaviors at 24 months (ADOS restricted, repetitive behaviors calibrated severity score and RBS-R overall score) (47–49). (See the online supplement for the formula for amygdala volume growth rate.) Amygdala growth rate in the ASD group between 6 and 12 months was unrelated to amygdala volume at 6 months. In the FXS group, linear regression was used to test whether caudate volume at 12 months (the time point with the highest number of FXS scans) was associated with repetitive behaviors at 24 months (RBS-R overall score). Brain-behavior analyses were restricted to those subjects for whom the required imaging data and behavioral data were available at each time point of interest.
RESULTS
Descriptive Statistics and Cognitive Ability From 6 to 24 Months
The participant characteristics by group are summarized in Table S1 in the online supplement. There were no significant group differences in age at each MRI time point. The trajectories of overall cognitive ability (using the Mullen Early Learning composite score) were examined as an exemplar of a behavioral characteristic that is important to track in both ASD and FXS and because the same metric can be reliably measured in both disorders across different ages in infancy. Cognitive ability was selected for analysis over other behaviors (e.g., social deficits) that undergo age-related changes in how they are exhibited and measured across the 6- to 24-month age interval and would thus complicate longitudinal comparisons between groups during this time period. There was a significant group-by-age interaction in cognitive ability, indicating that the groups had different trajectories of cognitive ability from 6 to 24 months (p<0.0001), and a significant main effect of group, indicating overall group differences (p<0.01) (Figure 1). Compared with all other groups, infants with FXS had significantly lower cognitive ability, starting at 6 months of age, and remaining significantly lower at 12 and 24 months (p<0.0001). In contrast, the HL-ASD group showed no differences in cognitive ability at 6 months relative to HL- and LL-negative control subjects, but showed significantly lower cognitive ability by 12 months (p<0.0001). The different timing and pattern of cognitive trajectories served as an illustration of behavioral differences in development between FXS and ASD, leading to the expectation that differential brain trajectories and timing of brain differences would be observed between the FXS and ASD groups.
FIGURE 1.
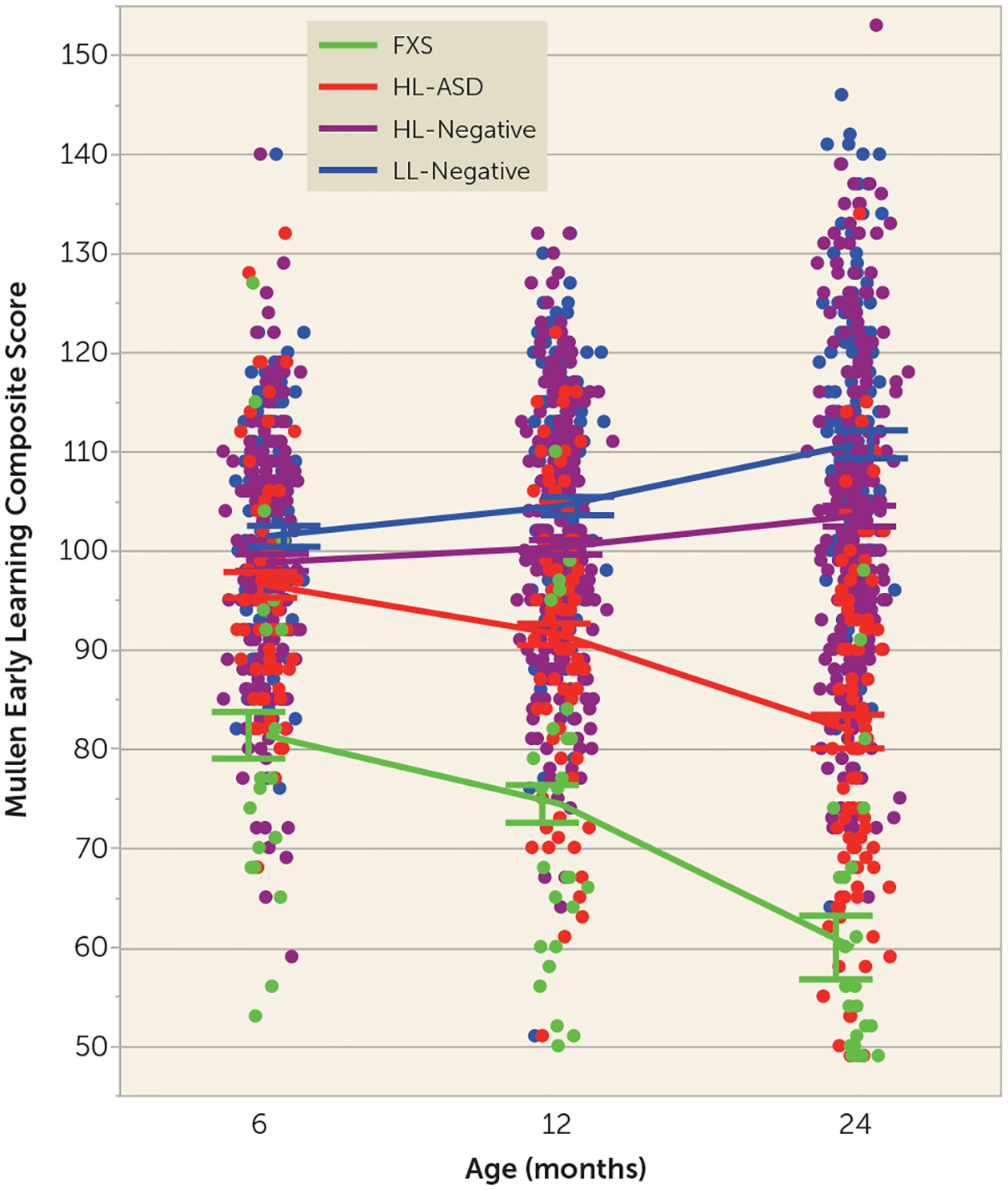
Cognitive ability from 6 to 24 months in infants at high or low likelihood for ASD who did or did not develop ASD and infants with fragile X syndromea
a ASD=autism spectrum disorder; FXS=fragile X syndrome; HL-ASD=infants at high likelihood for ASD who were later diagnosed with ASD; HL-negative=high-likelihood infants who were negative for ASD; LL-negative=low-likelihood infants who were negative for ASD. Compared with all other groups, the infants with fragile X syndrome had significantly lower cognitive ability (Mullen Early Learning composite score) starting at 6 months of age and remaining significantly lower at 12 and 24 months. The HL-ASD group showed no differences in cognitive ability at6 months relative to HL- and LL-negative control infants but showed significantly lower cognitive ability by 12 months of age. Error bars indicate standard error of the mean.
Subcortical Brain Growth Trajectories
Faster amygdala growth between 6 and 24 months and larger amygdala by 12 months in ASD.
There was a significant group-by-age interaction (F=13.62, df=3, 562, p<0.0001), with the HL-ASD group having a faster growth rate of the amygdala between 6 and 24 months, such that by 12 months the HL-ASD group had significantly larger amygdala volumes compared with all other groups (p<0.005, corrected), controlling forage, sex, and total cerebral volume. The ASD group continued to have larger amygdala volumes at 24 months compared with all other groups (p<0.05, corrected). There was a main effect of sex on the amygdala (p=0.02) but no sex-by-group interaction (p=0.11). Direct model comparison was also employed to compare the model fit of the nested models, with and without the group-by-age interaction term. Using the fit statistic −2 log likelihood, the difference between the two models (or the “deviance”) was 39.62, and the resulting chi-square test yielded significant results (χ2=39.62, df=3, p<0.0001) that were consistent with the results of the F statistic reported above (p<0.0001), thereby providing convergent results indicating that the age trajectories were highly significantly different between groups. Figure 2 illustrates the group trajectories of amygdala volumes from 6 to 24 months, with percent differences and effect sizes between model-adjusted group means (relative to the LL-negative group) at each time point, controlling for age, sex, and total cerebral volume. The HL-ASD group had 5% larger amygdala volume at 12 months (d=0.56), and 4% larger at 24 months (d=0.40), relative to the LL-negative group. The FXS group did not differ significantly from the HL- or LL-negative groups.
FIGURE 2.
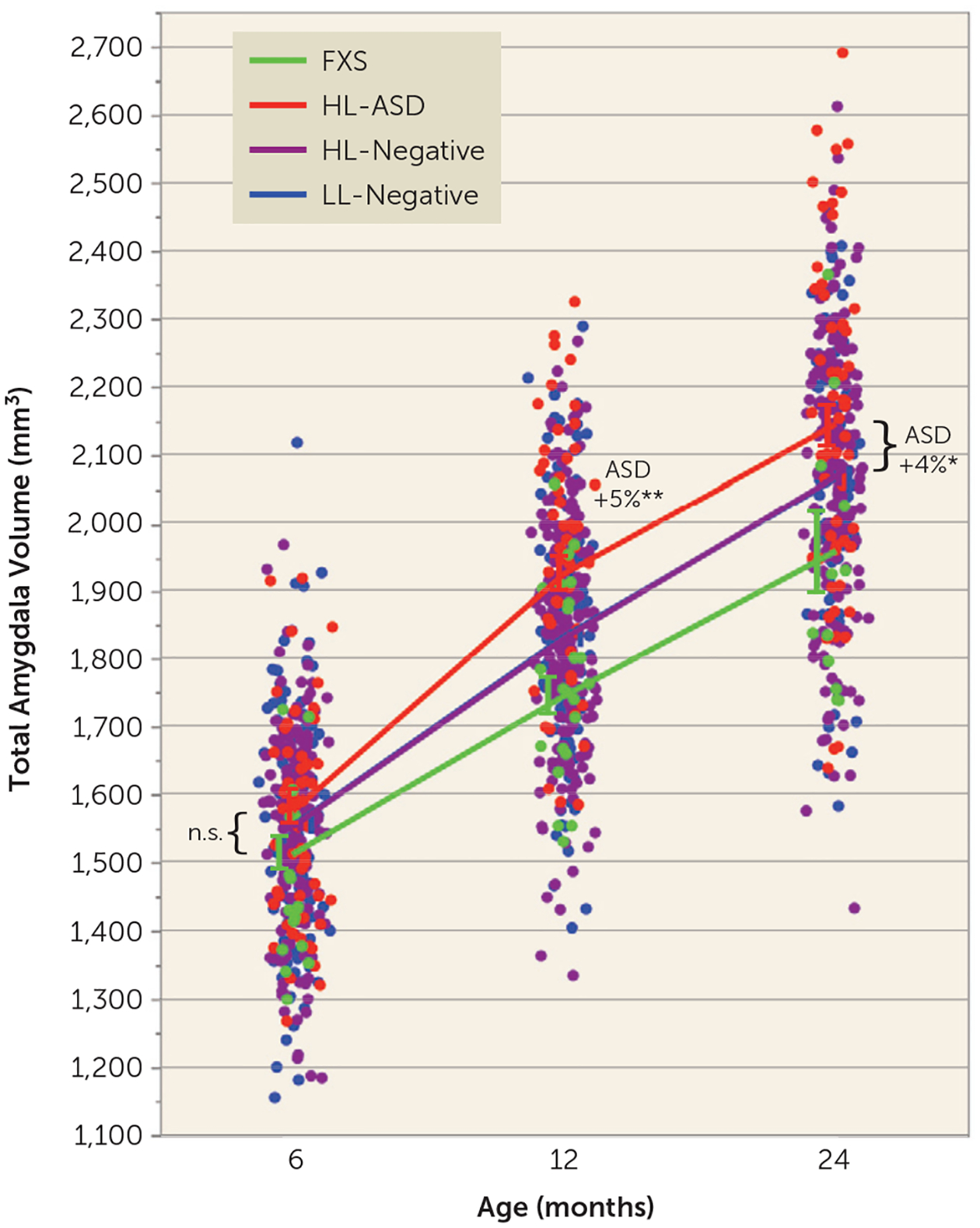
Amygdala growth between 6 and 24 months in infants at high or low likelihood for ASD who did or did not develop ASD and infants with fragile X syndromea
a ASD=autism spectrum disorder; FXS=fragile X syndrome; HL-ASD=infants at high likelihood for ASD who were later diagnosed with ASD; HL-negative=high-likelihood infants who were negative for ASD; LL-negative=low-likelihood infants who were negative for ASD. Infants who developed ASD had faster amygdala growth between 6 and 24 months; there were no group differences at 6 months, followed by significantly larger amygdala volume in the ASD group at 12 and 24 months, compared with all other groups; there was a significant group-by-age interaction (p<0.0001). Plots of the model-adjusted least-squares means are overlaid onto the raw data points of all participants. Percent differences in least-squares means are in relation to the LL-negative group. Effect sizes for the HL-ASD group relative to the LL-negative group: d=0.15 at 6 months; d=0.56 at 12 months; d=0.40 at 24 months. (Note that the LL-negative (blue) and HL-negative (purple) lines are overlapping.) Error bars indicate standard error of the mean. Asterisks indicate p values (corrected) compared with all other groups; n.s.=not significant.
*p<0.05. **p<0.005.
Caudate enlargement by 6 months in FXS.
There was a significant main effect of group on caudate volume (F=5.34, df=3, 559, p=0.001), controlling for age, sex, and total cerebral volume. Pairwise comparisons revealed that the FXS group had larger caudate volume at all ages from 6 to 24 months compared with all other groups (p<0.0001, corrected). There was no main effect of sex on caudate volume (p=0.19). Figure 3 illustrates the group trajectories of caudate volumes from 6 to 24 months, with percent differences between model-adjusted group means (relative to the LL-negative group) at each time point. The FXS group had 22% larger caudate volume at 6 months (d=2.12), 20% larger volume at 12 months (d=2.04), and 15% larger volume at 24 months (d=1.49).
FIGURE 3.
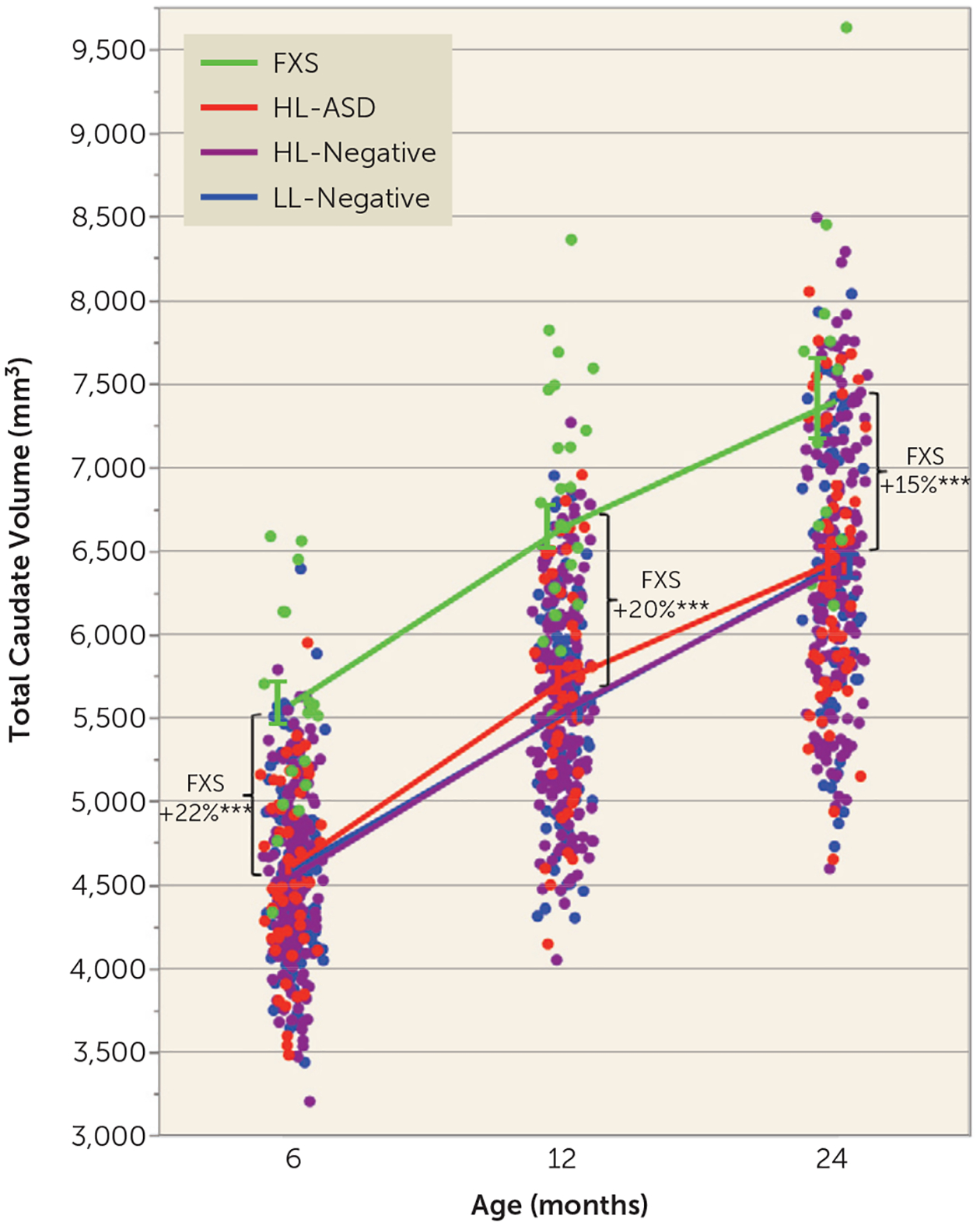
Caudate growth between 6 and 24 months in infants at high or low likelihood for ASD who did or did not develop ASD and infants with fragile X syndromea
a ASD=autism spectrum disorder; FXS=fragile X syndrome; HL-ASD=infants at high likelihood for ASD who were later diagnosed with ASD; HL-negative=high-likelihood infants who were negative for ASD; LL-negative=low-likelihood infants who were negative for ASD. Infants with fragile X syndrome had larger caudate volume at all ages between 6 and 24 months; there was a significant main effect of group (p=0.001). Plots of the model-adjusted least-squares means are overlaid onto the raw data points of all participants. Percent differences in least-squares means are in relation to the LL-negative group. Effect sizes for the FXS group relative to the LL-negative group: d=2.12 at 6 months; d=2.04 at 12 months; d=1.49 at 24 months. (Note that the lines for the LL-negative (blue) and HL-negative (purple) lines are overlapping.) Error bars indicate standard error of the mean. Asterisks indicate p values (corrected) compared with all other groups.
***p<0.0001.
Evaluating the volume of other basal ganglia structures.
Consistent with findings in the caudate, other basal ganglia structures (globus pallidus and putamen) were also significantly enlarged in the FXS group. The globus pallidus showed a significant main effect of group (p<0.0001), with the FXS group having significantly larger volume at 6 months (p<0.0001, corrected; 19% larger compared with the LL-negative group), 12 months (p<0.0001, corrected; 17% larger), and 24 months (p<0.005, corrected; 10% larger). Figure S2 in the online supplement illustrates the group trajectories of globus pallidus volumes from 6 to 24 months, with percent differences between model-adjusted group means (relative to the LL-negative group) at each time point. There was a main effect of sex on the globus pallidus (p=0.02) but no sex-by-group interaction (p=0.33). The putamen also showed a significant main effect of group (p<0.0001), with the FXS group having significantly larger volume at 6 months (p=0.005, corrected; 8% larger compared with the LL-negative group) and 12 months (p<0.005, corrected; 7% larger). There was no main effect of sex on the putamen (p=0.17). Figure S3 in the online supplement illustrates the group trajectories of putamen volumes from 6 to 24 months.
No differences in thalamus volume between the ASD or FXS groups.
We included an analysis of the thalamus as an additional subcortical structure to serve as a control. There were no significant differences between groups in thalamus volume (main effect of group, p=0.30), controlling for age, sex, and total cerebral volume. There was a main effect of sex on the thalamus (p=0.0005) but no sex-by-group interaction (p=0.52). Figure S4 in the online supplement illustrates the group trajectories of thalamus volumes from 6 to 24 months.
Brain-Behavior Associations
Given the extensive literature supporting the role of the amygdala in the development of social behavior (50), we tested the hypothesis that amygdala growth rate in the ASD group was associated with later social deficits (measured by the ADOS social affect calibrated severity score) at the time of diagnosis. Regression analyses indicated that faster amygdala growth rate in the HL-ASD group between 6 and 12 months was associated with greater social deficits at 24 months (F=11.57, df=1, 31, R2=0.272, r=0.52, p=0.002) (Figure 4A), but not with restricted and repetitive behaviors on either the ADOS (F=1.56, df=1, 31, R2=0.048, r=0.22, p=0.22) (Figure 4B) or the RBS-R (F=0.02, df=1, 24, R2=0.001, r=0.095, p=0.87). The association between 6- and 12-month amygdala growth and 24-month social deficits remained highly significant (p=0.002) even when the model controlled for growth rate of total cerebral volume between 6 and 12 months. Collectively, these results suggest that amygdala growth, above and beyond overall brain growth, was specifically associated with social deficits. The behavioral associations with the amygdala growth rate were not significant in the FXS group (all p values >0.57) (although this analysis was limited by eight FXS individuals who had imaging data at both time points and the requisite behavioral data at 24 months).
FIGURE 4.
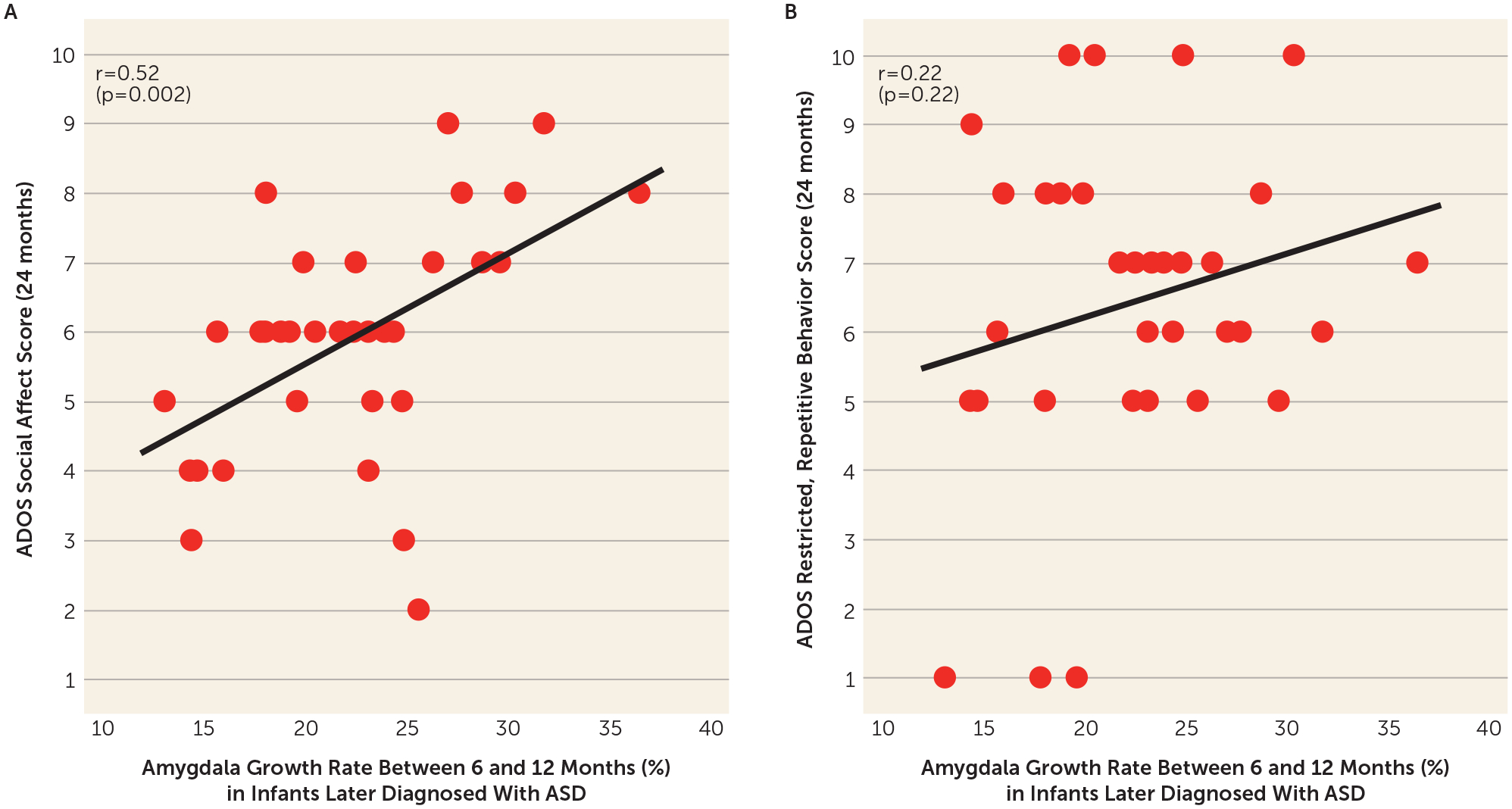
Relationship between amygdala growth between 6 and 12 months and social deficits and restricted and repetitive behaviors at 24 months in infants who developed ASDa
a ASD=autism spectrum disorder. Faster amygdala growth rate in the ASD group between 6 and 12 months was associated with greater social deficits at 24 months (based on the Autism Diagnostic Observation Schedule [ADOS] social affect calibrated severity score; panel A), but not restricted and repetitive behaviors at 24 months (based on the ADOS restricted, repetitive behavior calibrated severity score; panel B).
Second, we tested the a priori hypothesis that caudate enlargement in infants with FXS was associated with later repetitive behaviors, based on previous findings that preschool-age children with FXS had caudate enlargement associated with repetitive behaviors (51). In the FXS group, caudate volume at 12 months (the time point with the highest number of FXS scans) was significantly associated with greater repetitive behaviors at 24 months on the RBS-R (F=5.66, df=1, 10, R2=0.361, r=0.60, p=0.04) (Figure 5). This association was not significant in the HL-ASD group (F=0.22, df=1, 29, R2=0.007, r=0.09, p=0.64).
FIGURE 5.
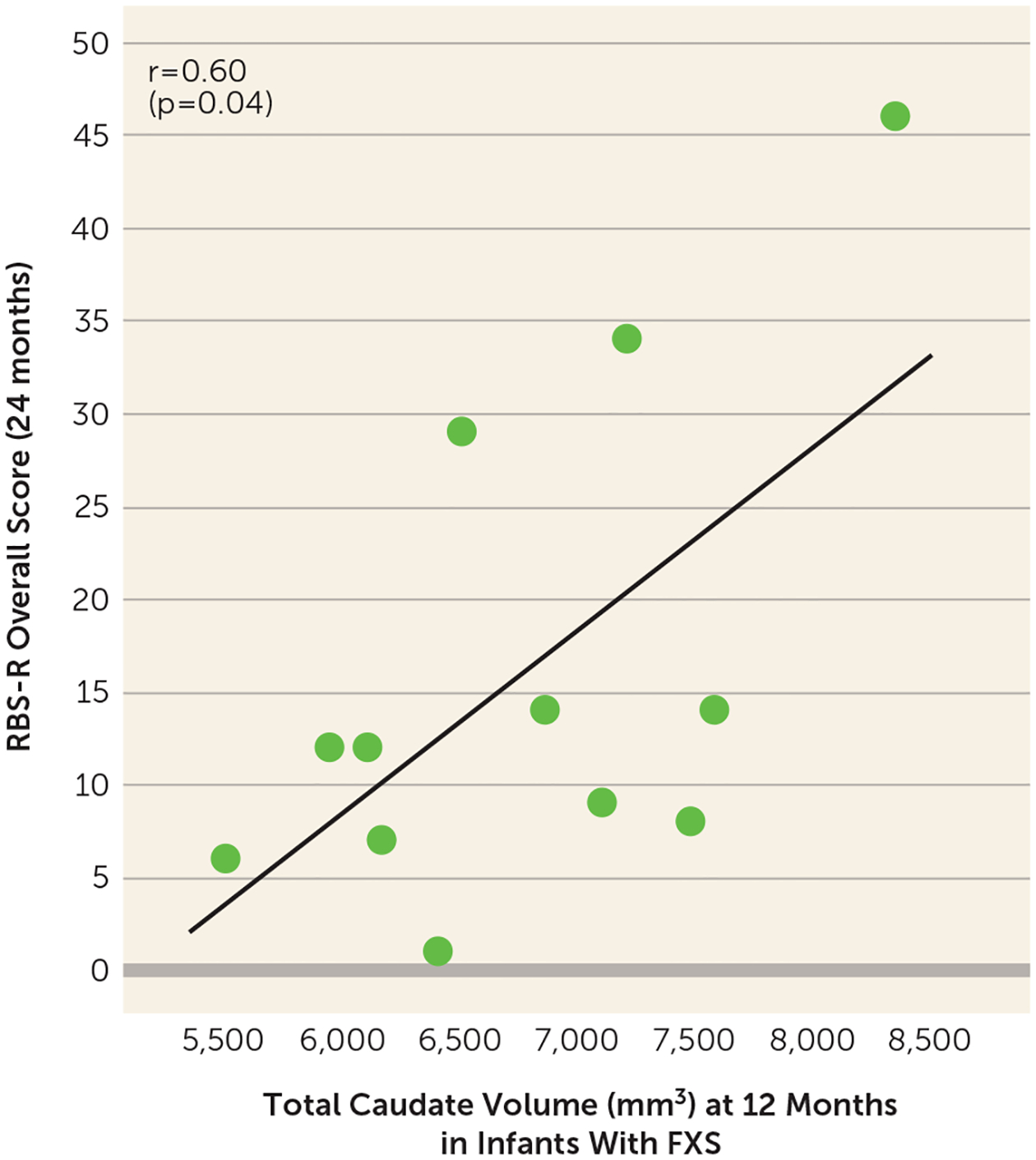
Relationship between caudate volume at 12 months and repetitive behaviors at 24 months in infants with fragile X syndromea
a In infants with fragile X syndrome (FXS), caudate enlargement at 12 months was associated with greater repetitive behaviors at 24 months (based on the overall score on the Repetitive Behavior Scale–Revised, RBS-R).
DISCUSSION
The results of this study demonstrate strikingly different timing and patterns of brain and behavior development in two neurodevelopmental disorders that share overlapping behavioral phenotypes. Notably, brain and behavior changes in FXS are present from the age of 6 months and persist until 24 months. In contrast, ASD characteristics change over the first 2 years of life, starting with a period of relatively typical brain and behavior development at 6 months (which coincides with data in this study as well as previous studies [1–4]), demonstrating relatively intact cognition and behavior at this young age in infants who later develop autism. The increased growth rate of the amygdala between 6 and 12 months occurs prior to the emergence of the social deficits that are diagnostic for autism (2, 4), and well before the typical age of consolidation of the symptoms of ASD into a diagnosable syndrome. This gradual onset of brain and behavior changes in ASD, but not FXS, suggests an age- and disorder-specific pattern of cascading brain changes leading to autism. More specifically, in ASD, we observed increased early postnatal growth rate of the amygdala, a brain structure often implicated in the social aspects of ASD, occurring prior to the consolidation of the defining social deficits in ASD, whereas, in contrast, in FXS we noted an earlier onset of enlargement of the caudate (consistent with findings in other basal ganglia structures), which remained stable over infancy—paralleling the temporal pattern of cognitive deficits in FXS that are evident by 6 months of age and not showing the same degree of decline over time observed in infants who later meet criteria for ASD. The present findings add to the growing evidence that the onset of ASD occurs in the early postnatal period, following a presymptomatic period when the defining features are not yet consolidated into the full, clinically defined syndrome that is typically observed later.
Potential Cellular and Developmental Processes Underlying the Specific Timing of Early Amygdala Overgrowth and Its Relation to Emerging Social Deficits in ASD
Our findings identify the onset and define the subsequent early development of the long-standing observation of enlarged amygdala volume in children with ASD. The specific timing of increased amygdala volume growth rate from 6 to 12 months of age raises the question of what cellular and developmental processes could be occurring that underlie this volume overgrowth. There is postmortem histological evidence in ASD demonstrating an excess of amygdala neurons in early childhood (23) (an age when amygdala enlargement has already been established in many children with autism), suggesting that early postnatal cellular growth may be dysregulated in individuals with ASD. Excess neuronal production could elicit a cascade of changes, such as increased dendritic arborization, leading to amygdala enlargement, dysfunctional communication between amygdala neurons and other brain regions, and clinical impairment (50). In the first year of life, dramatic dendritic growth and synapse formation takes place as dendrites establish synaptic connections with neurons. Therefore, more neurons in the amygdala during the first year of life in ASD might lead to an even greater volume and density of dendrites. Indeed, postmortem studies in ASD have reported excessive amygdala neurons (23) and increased dendritic spine density in the amygdala (24).
Previous work has shown that neurons in the amygdala mature in an experience-dependent fashion (52), raising the possibility that the excessive production of amygdala neurons (23) and increased dendritic spine density (24) in ASD may be related to altered activity-dependent maturation of amygdala neurons. During the first year of normative brain development, synapses compete for neural growth factors to survive: neural connections that are underactive are pruned in an activity-dependent manner, resulting in a functional network of efficient synaptic connections between neurons (53). However, emerging evidence suggests that sensory (particularly visual) processing may differ in infants who develop autism (1, 17, 54), during the early postnatal period when synaptic pruning is driven by sensory input. If there are excess neurons (23), and dendrites grow in a dysregulated manner by not undergoing efficient activity-dependent synaptic pruning (24), this could result in amygdala volume enlargement and aberrant signal transmission. This altered growth pattern would then be expected to lead to altered amygdala function and altered connectivity with brain regions that support sensory function, and thus contribute to social deficits. The amygdala is crucial for interpreting salient cues from the environment and coordinates multiple brain regions to detect threat and prepare an appropriate response: the visual system to detect a stimulus, the fusiform gyrus for face processing, and the orbitofrontal cortex for initiating goal-directed action. There are direct anatomical and functional connections between the amygdala, visual system, fusiform gyrus, and orbitofrontal cortex, all of which have been shown to be disrupted in young children with ASD (55).
During this period of increased amygdala growth from 6 to 12 months of age, we have observed contemporaneous hyperexpansion of surface area in the occipital cortex (9) and aberrant visual orienting (54) in this same sample of infants who later developed ASD. Aberrant growth of the amygdala and visual cortex, which is also temporally related to abnormal visual orienting, may represent an aberrant feedback loop between visual and attention regions and the amygdala. It is possible that altered sensory experience leads to hypertrophy of the amygdala and altered amygdala circuitry affecting sensory and social processing systems. The stress model of the amygdala, proposed by McEwen (56), posits that hypertrophy of the amygdala could be caused by stress. The amygdala stimulates the hypothalamic-pituitary-adrenal (HPA) system and stress response: sensory information enters the basolateral amygdala and is relayed to neurons in the central nucleus, and when neurons in the central nucleus are activated, the stress response ensues. The amygdala regulates the HPA axis by evoking the fight-or-flight response by increasing vigilance or alertness (56). The amygdala is involved in encoding memories of emotional and painful events, and thus fearful, distressing experiences can form quickly and be long-lasting. Neurons in the amygdala learn to respond to stimuli associated with fear and distress, which aids in recognizing similar fearful stimuli in the future and then evoking a fear response (57). For example, excessive activity of the amygdala is associated with anxiety disorders and autism (58–60). In autism, there is evidence that infants who later develop autism have more reactive temperament (61), which has been shown to be a precursor to later anxiety (62). Abnormal visual processing and atypical sensory experience (54) could result in distress and anxiety in infants who develop autism as their sensory experiences are altered. This early stress could drive hyperresponsiveness of the amygdala, increased HPA activity, and hypertrophy of the amygdala.
The amygdala works in concert with the parietal and occipital association cortices to regulate visual attention, such that disrupted development of this attentional network (54) and associated visual regions (e.g., middle occipital cortex [9]) could affect the normal development of important social behaviors (e.g., social eye gaze, interpreting another person’s intentions and movements, and perceiving the spatial relations between oneself and the surrounding environment [63]). This could contribute to the dysfunction of pivotal skills that are characteristic of social deficits in autism, including eye contact, response to name, and joint attention. For instance, joint attention comprises an important set of skills that support the development of language and social communication (64): the child needs to make eye contact, follow the eye gaze of another person, and direct and coordinate attention to share a visual experience (65). Joint attention impairment could relate to deficits in lower-level perceptual processes such as face processing, visual orienting and attention, and interpreting others’ actions (21). Indeed, Shen et al. (55) reported that preschool-age children with ASD have altered functional connectivity between the amygdala and areas important for social communication, including the medial prefrontal cortex (mPFC); this altered amygdala-mPFC connectivity was correlated with worse social deficits in ASD (55). The mPFC regulates emotional responses triggered by the amygdala by providing contextual and experiential input to the amygdala, which in turn uses this information to interpret social stimuli and prepare behavioral and emotional responses (55). Abnormal function of the amygdala and mPFC in children with ASD has been related to an exaggerated response of the amygdala to faces (66), alterations in social reward and social motivation (67), and increased ASD severity (55). The present study demonstrated that increased amygdala growth in the first year of life in ASD was associated with later social deficits. While this is consistent with the above-mentioned literature on the amygdala’s role in social behavior, this specific brain-behavior finding warrants replication in an independent sample of HL infants, which is currently under way.
Increased amygdala growth in ASD could also be related to neuroinflammation in the first years of life (68–70). Microglia play an important role in responding to neuroinflammation and neuronal injury. Resting microglia are characterized by small cell bodies and long thin processes; but when microglia are activated in response to immune challenges, they readily increase in number, their processes thicken, and the cell body swells to as much as two to four times its normal volume, while quickly moving to the site of infection, where microglia interact with neurons to fight infection (50, 68). Postmortem studies in autism have shown excessive microglia activation, both in number and size of microglia, in the amygdala (71).It is possible that increased amygdala size in the first year of life reflects 1) microglia becoming activated (and increasing in number and size) in an inflammatory response to overproliferation of amygdala neurons (23), both of which would contribute to early amygdala enlargement; or 2) increased number and size of microglial cells in a secondary response to some other postnatal neuroinflammatory insult (68–70).
What Could Underlie Caudate Enlargement in FXS and Its Relationship to Repetitive Behaviors?
The FMR1 mutation responsible for FXS causes diminished production of the protein FMRP expressed in neurons. FMRP binds to various mRNAs and plays a vital role in neural development and synaptic plasticity (27–30). Stereological analyses have shown that FMRP is highly expressed in the caudate and other basal ganglia structures and involves multiple neuronal processes: the nucleus, cytoplasm, dendrites, and dendritic spines (31). Disruption of FMRP could result in any number of prenatal and early postnatal events leading to increased caudate volume: increased neuron number, increased neuron size, decreased cell packing density, increased neuropil, reduced synaptic pruning, and greater volume and density of dendritic spines.
Previous research has identified a circuit-specific mechanism for repetitive behavior (72) whereby the D1 receptor regulates a specific loop between the caudate, globus pallidus, thalamus, and cortex that can increase repetitive behaviors. Excitatory/inhibitory activity imbalance in these regions (i.e., increased activity of excitatory neurons, decreased activity of inhibitory neurons) can lead to sensory dysfunction. For example, imbalance in activity between the direct and indirect pathways of the basal ganglia system—either reduced D2 activity (leading to reduced inhibition) or increased D1 activity (leading to increased excitation)—leads to increased excitability and greater repetitive behaviors (73). Cortico-striatal-thalamo-cortical circuits underlie behavioral features of many neurodevelopmental disorders, including motor stereotypies, compulsive and ritualistic behaviors, atypical reward processing, and sensory dysfunction (74, 75). The work of Ting and Feng (76) demonstrates that specific circuits and behavior domains have different critical periods, and therefore specific windows for interventions during periods of plasticity. For example, the striatal circuit described above has greater plasticity and a longer and later therapeutic window for treating repetitive behaviors, compared to social behaviors. This raises the potential that identifying the pathophysiological mechanisms underlying caudate enlargement could lead to targeted treatments for repetitive behaviors in FXS.
CONCLUSIONS
Amygdala growth between 6 and 12 months of age occurs during a cascading series of brain and behavior changes that precede and lead to ASD. Early aberrant amygdala growth occurs at the same time as sensorimotor and attention problems in infants who go on to develop ASD, and at the same time as hyperexpansion of cortical surface area in the visual cortex. It raises the possibility of whether visual attention deficits might lead to altered experience, with subsequent deficits in activity-dependent synaptic pruning, and amygdala overgrowth. Furthermore, the stark contrast in onset of cognitive deficits between ASD and FXS underscores that ASD is a disorder of early and gradual decline of cognitive function in infancy. This highlights the importance of studying the period before that decline and the subsequent brain changes, and suggests the importance of presymptomatic detection and intervention. Finally, the study findings suggest potential targets for early intervention (e.g., experience-dependent development or neuroinflammation) that may alter this developmental cascade in infancy.
Supplementary Material
Acknowledgments
The Infant Brain Imaging Study (IBIS) Network is an NIH-funded Autism Center of Excellence project and consists of a consortium of 10 universities in the United States and Canada. Members and components of the IBIS Network include J. Piven (IBIS Network principal investigator). Clinical sites: Children’s Hospital of Philadelphia: R.T. Schultz, J. Pandey, J. Parish-Morris, B. Tunç, W. Guthrie; University of Minnesota (UMN): J.T. Elison, J.J. Wolff; University of North Carolina (UNC): J. Piven, H.C. Hazlett, M.D. Shen, J.B. Girault, R. Grzadzinski; University of Washington (UW): S.R. Dager, A.M. Estes, T. St. John, D. Shaw; Washington University School of Medicine in St. Louis (WU): K.N. Botteron, R.C. McKinstry, J.N. Constantino, N. Marrus. Admin Core: WU: Alicia Rocca; UNC: J.C. Chappell. Behavior Core: UW: A.M. Estes, T. St. John; University of Alberta: L. Zwaigenbaum; UMN: J.T. Elison, J.J. Wolff, C. Burrows; University of Texas at Dallas: M.R. Swanson. MRI Core: UNC: M.A. Styner, M.D. Shen; New York University: G. Gerig; WU: J.R. Pruett, Jr., R.C. McKinstry; UMN: J.T. Elison; UW: S.R. Dager. Data Coordinating Center: Montreal Neurological Institute: A.C. Evans, L.C. MacIntyre, S. Torres-Gomez, S. Das. Statistical Analysis Core: UNC: K. Truong. Environmental Risk Core: Johns Hopkins University (JHU): H. Volk. Genetics Core: JHU: M.D. Fallin; UNC: M.D. Shen.
Supported by NIH grants R01-HD055741, R01-HD059854, R01-MH118362-01, R01-MH118362-02S1, T32-HD040127, U54-HD079124, P50-HD103573 (project ID 8084), R01-EB021391, and U54-HD086984; by Autism Speaks; and by the Simons Foundation (grant 140209). Dr. Shen was supported by an NIH career development award (K12-HD001441).
The authors are grateful to all the families and children who have participated in the IBIS study.
Dr. McKinstry has served on a medical advisory board for Nous Imaging and has equity in the company, and he has received travel support from Siemens Healthineers and Philips Healthcare. The other authors report no financial relationships with commercial interests.
REFERENCES
- 1.Estes A, Zwaigenbaum L, Gu H, et al. : Behavioral, cognitive, and adaptive development in infants with autism spectrum disorder in the first 2 years of life. J Neurodev Disord 2015; 7:24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ozonoff S, Iosif A-M, Baguio F, et al. : A prospective study of the emergence of early behavioral signs of autism. J Am Acad Child Adolesc Psychiatry 2010; 49:256–266.e1–2 [PMC free article] [PubMed] [Google Scholar]
- 3.Landa RJ, Gross AL, Stuart EA, et al. : Developmental trajectories in children with and without autism spectrum disorders: the first 3 years. Child Dev 2013; 84:429–442 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Zwaigenbaum L, Bryson S, Rogers T, et al. : Behavioral manifestations of autism in the first year of life. Int J Dev Neurosci 2005; 23:143–152 [DOI] [PubMed] [Google Scholar]
- 5.Wolff JJ, Botteron KN, Dager SR, et al. : Longitudinal patterns of repetitive behavior in toddlers with autism. J Child Psychol Psychiatry 2014; 55:945–953 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Filliter JH, Longard J, Lawrence MA, et al. : Positive affect in infant siblings of children diagnosed with autism spectrum disorder. J Abnorm Child Psychol 2014; 43:567–575 [DOI] [PubMed] [Google Scholar]
- 7.Miller M, Iosif A-M, Hill M, et al. : Response to name in infants developing autism spectrum disorder: a prospective study. J Pediatr 2017; 183:141–146.e1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Stallworthy I, Lasch C, Berry D, et al. : Variability in responding to joint attention cues in the first year is associated with autism outcome. J Am Acad Child Adolesc Psychiatry 2022; 61:413–422 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hazlett HC, Gu H, Munsell H, et al. : Early brain development in infants at high risk for autism spectrum disorder. Nature 2017; 542: 348–351 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ohta H, Nordahl CW, Iosif A-M, et al. : Increased surface area, but not cortical thickness, in a subset of young boys with autism spectrum disorder. Autism Res 2015; 9:232–248 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Wolff JJ, Gu H, Gerig G, et al. : Differences in white matter fiber tract development present from 6 to 24 months in infants with autism. Am J Psychiatry 2012; 169:589–600 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wolff JJ, Gerig G, Lewis JD, et al. : Altered corpus callosum morphology associated with autism over the first 2 years of life. Brain 2015; 138:2046–2058 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Frazier TW, Keshavan MS, Minshew NJ, et al. : A two-year longitudinal MRI study of the corpus callosum in autism. J Autism Dev Disord 2012; 42:2312–2322 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Shen MD, Nordahl CW, Young GS, et al. : Early brain enlargement and elevated extra-axial fluid in infants who develop autism spectrum disorder. Brain 2013; 136:2825–2835 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Shen MD, Kim SH, McKinstry RC, et al. : Increased extra-axial cerebrospinal fluid in high-risk infants who later develop autism. Biol Psychiatry 2017; 82:186–193 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Shen MD, Nordahl CW, Li DD, et al. : Extra-axial cerebrospinal fluid in high-risk and normal-risk children with autism aged 2–4 years: a case-control study. Lancet Psychiatry 2018; 5:895–904 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Piven J, Elison JT, Zylka MJ: Toward a conceptual framework for early brain and behavior development in autism. Mol Psychiatry 2017; 22:1385–1394 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hazlett HC, Poe MD, Lightbody AA, et al. : Teasing apart the heterogeneity of autism: same behavior, different brains in toddlers with fragile X syndrome and autism. J Neurodev Disord 2009; 1:81–90 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Sparks BF, Friedman SD, Shaw DW, et al. : Brain structural abnormalities in young children with autism spectrum disorder. Neurology 2002; 59:184–192 [DOI] [PubMed] [Google Scholar]
- 20.Schumann CM, Barnes CC, Lord C, et al. : Amygdala enlargement in toddlers with autism related to severity of social and communication impairments. Biol Psychiatry 2009; 66:942–949 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Mosconi MW, Cody-Hazlett H, Poe MD, et al. : Longitudinal study of amygdala volume and joint attention in 2- to 4-year-old children with autism. Arch Gen Psychiatry 2009; 66:509–516 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Nordahl CW, Scholz R, Yang X, et al. : Increased rate of amygdala growth in children aged 2–4 years with autism spectrum disorders: a longitudinal study. Arch Gen Psychiatry 2012; 69:53–61 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Avino TA, Barger N, Vargas MV, et al. : Neuron numbers increase in the human amygdala from birth to adulthood, but not in autism. Proc Natl Acad Sci USA 2018; 115:3710–3715 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Weir RK, Bauman MD, Jacobs B, et al. : Protracted dendritic growth in the typically developing human amygdala and increased spine density in young ASD brains. J Comp Neurol 2017; 526:262–274 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Niu M, Han Y, Dy ABC, et al. : Autism symptoms in fragile X syndrome. J Child Neurol 2017; 32:903–909 [DOI] [PubMed] [Google Scholar]
- 26.Gaugler T, Klei L, Sanders SJ, et al. : Most genetic risk for autism resides with common variation. Nat Genet 2014; 46:881–885 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Abrams MT, Reiss AL: The neurobiology of fragile X syndrome. Ment Retard Dev Disabil Res Rev 1995; 1:269–275 [Google Scholar]
- 28.Jones R: FMRP and its many partners. Nat Rev Neurosci 2003; 4: 245 [Google Scholar]
- 29.Miyashiro KY, Beckel-Mitchener A, Purk TP, et al. : RNA cargoes associating with FMRP reveal deficits in cellular functioning in Fmr1 null mice. Neuron 2003; 37:417–431 [DOI] [PubMed] [Google Scholar]
- 30.Zalfa F, Giorgi M, Primerano B, et al. : The fragile X syndrome protein FMRP associates with BC1 RNA and regulates the translation of specific mRNAs at synapses. Cell 2003; 112:317–327 [DOI] [PubMed] [Google Scholar]
- 31.Zorio DAR, Jackson CM, Liu Y, et al. : Cellular distribution of the fragile X mental retardation protein in the mouse brain. J Comp Neurol 2017; 525:818–849 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Hazlett HC, Poe MD, Lightbody AA, et al. : Trajectories of early brain volume development in fragile X syndrome and autism. J Am Acad Child Adolesc Psychiatry 2012; 51:921–933 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lord C, Rutter M, Le Couteur A: Autism Diagnostic Interview–Revised: a revised version of a diagnostic interview for caregivers of individuals with possible pervasive developmental disorders. J Autism Dev Disord 1994; 24:659–685 [DOI] [PubMed] [Google Scholar]
- 34.Hazlett HC, Gu H, McKinstry RC, et al. : Brain volume findings in 6-month-old infants at high familial risk for autism. Am J Psychiatry 2012; 169:601–608 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Swanson MR, Wolff JJ, Shen MD, et al. : Development of white matter circuitry in infants with fragile X syndrome. JAMA Psychiatry 2018; 75:505–513 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Mullen EM: Mullen Scales of Early Learning. Circle Pines, Minn, American Guidance Service, 1995 [Google Scholar]
- 37.Bodfish JW, Symons FJ, Parker DE, et al. : Varieties of repetitive behavior in autism: comparisons to mental retardation. J Autism Dev Disord 2000; 30:237–243 [DOI] [PubMed] [Google Scholar]
- 38.Elison JT, Wolff JJ, Reznick JS, et al. : Repetitive behavior in 12-month-olds later classified with autism spectrum disorder. J Am Acad Child Adolesc Psychiatry 2014; 53:1216–1224 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Wolff JJ, Swanson MR, Elison JT, et al. : Neural circuitry at age 6 months associated with later repetitive behavior and sensory responsiveness in autism. Mol Autism 2017; 8:8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.American Psychiatric Association: Diagnostic and Statistical Manual of Mental Disorders (Text Revision), 4th ed. Washington, DC, American Psychiatric Association, 2000 [Google Scholar]
- 41.Lord C, Risi S, Lambrecht L, et al. : The Autism Diagnostic Observation Schedule–Generic: a standard measure of social and communication deficits associated with the spectrum of autism. J Autism Dev Disord 2000; 30:205–223 [PubMed] [Google Scholar]
- 42.Roberts JE, Tonnsen BL, McCary LM, et al. : Brief report: autism symptoms in infants with fragile X syndrome. J Autism Dev Disord 2016; 46:3830–3837 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Fonov VS, Janke A, Caramanos Z, et al. : Improved precision in the measurement of longitudinal global and regional volumetric changes via a novel MRI gradient distortion characterization and correction technique. Med Imag Augment Reality 2010; 6326: 324–333 [Google Scholar]
- 44.Sled JG, Zijdenbos AP, Evans AC: A nonparametric method for automatic correction of intensity nonuniformity in MRI data. IEEE Trans Med Imaging 1998; 17:87–97 [DOI] [PubMed] [Google Scholar]
- 45.Collins DL, Neelin P, Peters TM, et al. : Automatic 3D intersubject registration of MR volumetric data in standardized Talairach space. J Comp Assisted Tomography 1994; 18:192–205 [PubMed] [Google Scholar]
- 46.Wang J, Vachet C, Rumple A, et al. : Multi-atlas segmentation of subcortical brain structures via the AutoSeg software pipeline. Front Neuroinform 2014; 8:7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Gotham K, Pickles A, Lord C: Standardizing ADOS scores for a measure of severity in autism spectrum disorders. J Autism Dev Disord 2009; 39:693–705 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Gotham K, Risi S, Pickles A, et al. : The Autism Diagnostic Observation Schedule: revised algorithms for improved diagnostic validity. J Autism Dev Disord 2007; 37:613–627 [DOI] [PubMed] [Google Scholar]
- 49.Gotham K, Pickles A, Lord C: Trajectories of autism severity in children using standardized ADOS scores. Pediatrics 2012; 130: e1278–e1284 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Schumann CM, Nordahl CW: Bridging the gap between MRI and postmortem research in autism. Brain Res 2011; 1380: 175–186 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Wolff JJ, Hazlett HC, Lightbody AA, et al. : Repetitive and self-injurious behaviors: associations with caudate volume in autism and fragile X syndrome. J Neurodev Disord 2013; 5:12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Fudge JL, deCampo DM, Becoats KT: Revisiting the hippocampal-amygdala pathway in primates: association with immature-appearing neurons. Neuroscience 2012; 212:104–119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.McAllister AK: Dynamic aspects of CNS synapse formation. Annu Rev Neurosci 2007; 30:425–450 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Elison JT, Paterson SJ, Wolff JJ, et al. : White matter microstructure and atypical visual orienting in 7-month-olds at risk for autism. Am J Psychiatry 2013; 170:899–908 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Shen MD, Li DD, Keown CL, et al. : Functional connectivity of the amygdala is disrupted in preschool-aged children with autism spectrum disorder. J Am Acad Child Adolesc Psychiatry 2016; 55: 817–824 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.McEwen BS: Structural plasticity of the adult brain: how animal models help us understand brain changes in depression and systemic disorders related to depression. Dialogues Clin Neurosci 2004; 6:119–133 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Adolphs R: Cognitive neuroscience of human social behaviour. Nat Rev Neurosci 2003; 4:165–178 [DOI] [PubMed] [Google Scholar]
- 58.Tottenham N, Hertzig ME, Gillespie-Lynch K, et al. : Elevated amygdala response to faces and gaze aversion in autism spectrum disorder. Soc Cogn Affect Neurosci 2014; 9:106–117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Green SA, Hernandez L, Tottenham N, et al. : Neurobiology of sensory overresponsivity in youth with autism spectrum disorders. JAMA Psychiatry 2015; 72:778–786 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Amaral DG, Bauman MD, Mills Schumann C: The amygdala and autism: implications from non‐human primate studies. Genes Brain Behav 2003; 2:295–302 [DOI] [PubMed] [Google Scholar]
- 61.Paterson SJ, Wolff JJ, Elison JT, et al. : The importance of temperament for understanding early manifestations of autism spectrum disorder in high-risk infants. J Autism Dev Disord 2019; 49: 2849–2863 [DOI] [PubMed] [Google Scholar]
- 62.Shephard E, Bedford R, Milosavljevic B, et al. : Early developmental pathways to childhood symptoms of attention-deficit hyperactivity disorder, anxiety, and autism spectrum disorder. J Child Psychol Psychiatry 2019; 60:963–974 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Shultz S, Klin A, Jones W: Inhibition of eye blinking reveals subjective perceptions of stimulus salience. Proc Natl Acad Sci USA 2011; 108:21270–21275 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Tomasello M, Carpenter M, Call J, et al. : Understanding and sharing intentions: the origins of cultural cognition. Behav Brain Sci 2005; 28:675–691; discussion 691–735 [DOI] [PubMed] [Google Scholar]
- 65.Mundy P, Newell L: Attention, joint attention, and social cognition. Curr Dir Psychol Sci 2007; 16:269–274 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Swartz JR, Wiggins JL, Carrasco M, et al. : Amygdala habituation and prefrontal functional connectivity in youth with autism spectrum disorders. J Am Acad Child Adolesc Psychiatry 2013; 52:84–93 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Chevallier C, Kohls G, Troiani V, et al. : The social motivation theory of autism. Trends Cogn Sci 2012; 16:231–239 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Morgan JT, Chana G, Pardo CA, et al. : Microglial activation and increased microglial density observed in the dorsolateral prefrontal cortex in autism. Biol Psychiatry 2010; 68:368–376 [DOI] [PubMed] [Google Scholar]
- 69.Pardo CA, Vargas DL, Zimmerman AW: Immunity, neuroglia, and neuroinflammation in autism. Int Rev Psychiatry 2005; 17:485–495 [DOI] [PubMed] [Google Scholar]
- 70.Vargas DL, Nascimbene C, Krishnan C, et al. : Neuroglial activation and neuroinflammation in the brain of patients with autism. Ann Neurol 2004; 57:67–81 [DOI] [PubMed] [Google Scholar]
- 71.Morgan JT, Barger N, Amaral DG, et al. : Stereological study of amygdala glial populations in adolescents and adults with autism spectrum disorder. PLoS One 2014; 9:e110356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Burguière E, Monteiro P, Mallet L, et al. : Striatal circuits, habits, and implications for obsessive-compulsive disorder. Curr Opin Neurobiol 2015; 30:59–65 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Ting JT, Feng G: Glutamatergic synaptic dysfunction and obsessive-compulsive disorder. Curr Chem Genom 2008; 2:62–75 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Dichter GS, Felder JN, Green SR, et al. : Reward circuitry function in autism spectrum disorders. Soc Cogn Affect Neurosci 2012; 7:160–172 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Dichter GS, Damiano CA, Allen JA: Reward circuitry dysfunction in psychiatric and neurodevelopmental disorders and genetic syndromes: animal models and clinical findings. J Neurodev Disord 2012; 4:19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Ting JT, Feng G: Unfolding neurodevelopmental disorders: found in translation. Nat Med 2011; 17:1352–1353 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.