

Abstract
Heterozygous loss of function mutations in TWIST1 cause Saethre-Chotzen syndrome, which is characterized by craniosynostosis, facial asymmetry, ptosis, strabismus, and distinctive ear appearance. Individuals with syndromic craniosynostosis have high rates of strabismus and ptosis, but the underlying pathology is unknown. Some individuals with syndromic craniosynostosis have been noted to have absence of individual extraocular muscles or abnormal insertions of the extraocular muscles on the globe. Using conditional knock-out alleles for Twist1 in cranial mesenchyme, we test the hypothesis that Twist1 is required for extraocular muscle organization and position, attachment to the globe, and/or innervation by the cranial nerves. We examined the extraocular muscles in conditional Twist1 knock-out animals using Twist2-cre and Pdgfrb-cre drivers. Both are expressed in cranial mesoderm and neural crest. Conditional inactivation of Twist1 using these drivers leads to disorganized extraocular muscles that cannot be reliably identified as specific muscles. Tendons do not form normally at the insertion and origin of these dysplastic muscles. Knock-out of Twist1 expression in tendon precursors, using scleraxis-cre, however, does not alter EOM organization. Furthermore, developing motor neurons, which do not express Twist1, display abnormal axonal trajectories in the orbit in the presence of dysplastic extraocular muscles. Strabismus in individuals with TWIST1 mutations may therefore be caused by abnormalities in extraocular muscle development and secondary abnormalities in innervation and tendon formation.
Keywords: Twist1, extraocular muscles, strabismus, tendons, cranial nerves, craniosynostosis
Graphical Abstract
Introduction
TWIST1 is a basic helix-loop-helix transcription factor required for the specification of cranial mesoderm (Qin et al., 2012). Heterozygous loss of function of TWIST1 in humans causes Saethre-Chotzen syndrome, which is characterized by craniosynostosis, facial asymmetry, ptosis, strabismus, and distinctive ear appearance (Gallagher et al., 1993). Patients with craniosynostosis, especially syndromic craniosynostosis, have a very high incidence of V-pattern strabismus. Several potential mechanisms have been postulated, including malrotation/mispositioning of the orbits secondary to the craniosynostosis, absence of certain extraocular muscles (EOMs), or mispositioning of the EOMs on the globe (Rosenberg et al., 2013). Additionally, some individuals with TWIST1 variants have isolated ptosis, without clinically apparent craniosynostosis (Dollfus et al., 2002; Stoler et al., 2009). We therefore sought to evaluate the role of Twist1 in EOM differentiation and development.
The genetic pathways regulating extraocular muscle differentiation are distinct from those of somatic muscle and of adjacent craniofacial muscle (Buckingham and Rigby, 2014). In primordial EOM cells, PITX2 activates a myogenic regulatory factor (MRF) cascade involving MYF5 and MRF4, leading to the activation of MYOD, a myogenic determination factor. EOM precursors do not express PAX3 or PAX7, which activate MYOD expression in somitic muscle precursors (Sambasivan et al., 2009; Sambasivan et al., 2011). Thus, complete loss of either Pitx2, or combined loss of Myf5 and Mrf4, results in an absence of EOMs (Diehl et al., 2006; Gage et al., 1999; Kitamura et al., 1999; Sambasivan et al., 2009). EOMs differentiate from cranial mesodermal precursors in periocular mesenchyme (Bothe and Dietrich, 2006; Fan et al., 2016; Kuroda et al., 2021; Ziermann et al., 2018). The muscles initially form an anlage in the orbit. Between E11.75 and E12.5, they separate into distinct EOMs which originate at the annulus of Zinn and insert on the sclera (Comai et al., 2020; Michalak et al., 2017). The connective tissues at the insertion of the muscles on the sclera derive from neural crest and mesodermal precursors while the connective tissues at the origin of the muscles at the annulus of Zinn derive from mesodermal precursors (Comai et al., 2020; Grimaldi and Tajbakhsh, 2021; Kuroda et al., 2021). The cell-intrinsic factors and extrinsic cues which regulate the growth and organization of the EOMs are not well characterized.
Twist1 null mice die at mid gestation (~E10.5) and have complete failure of cranial neural fold fusion; chimeric mice with Twist1 null cells in the cranial mesenchyme also have exencephaly (Chen and Behringer, 1995). Heterozygous mice have limb abnormalities and premature fusion of the cranial sutures (Bourgeois et al., 1998; Carver et al., 2002). Due to the severe consequences of full loss of Twist1, several studies have used conditional alleles with a variety of cell-type specific cre recombinase lines to examine the role of Twist1 in various aspects of development (Bildsoe et al., 2009; Bildsoe et al., 2013). Loss of Twist1 in the cranial neural crest, with Wnt1-cre, leads to severe malformations of the facial and skull bones, as well as abnormalities of eye development (Bildsoe et al., 2009). Loss of Twist1 in cranial mesodermal progenitor cells via Mesp1-cre abolishes Pitx2 expression in the periocular area and leads to complete loss of the EOMs, including the initial muscle anlage. However, these animals, like the complete null, have neural tube closure defects (Bildsoe et al., 2013). Mesp1-cre is expressed in mesoderm starting at gastrulation (Saga et al., 1999); conditional knock-out of Twist1 under other mesenchymal cre drivers with later onset of expression allow neural tube closure and more complete head patterning (Tischfield et al., 2017).
To determine the role of Twist1 in extraocular muscle development without the confounding effects of exencephaly, we have examined the extraocular muscles in mice with loss of Twist1 in different subpopulations of mesenchyme and show that Twist1 expression is required for proper EOM patterning and tendon formation. Furthermore, developing motor neurons, which do not express Twist1, display abnormal branching in the presence of dysplastic EOMs.
Methods
Animals
Mouse lines used were: Twist1cKO (B6;129S7-Twist1tm2Bhr/Mmnc; MMRRC 016842-UNC) (Chen et al., 2007); Twist2-cre (B6.129X1-Twist2tm1.1(cre)Dor/J; Jackson Labs 008712) (Sosic et al., 2003); Scx-cre (Gift of Ronen Schweitzer; (Blitz et al., 2009)); Scx-GFP (Gift of Ronen Schweitzer; (Pryce et al., 2007)); IslMN-GFP (Tg(Isl1-EGFP*)1Slp; Jackson Labs 017952) (Lewcock et al., 2007); Pdgfrb-cre (Cuttler et al., 2011). All animal work was approved and completed in compliance with Boston Children’s Hospital Institutional Animal Care and Use Committee protocols (20-01-4061R).
Frozen Section Immunohistochemistry
Embryos were fixed in 4% paraformaldehyde for 2–24 hours depending on the age, washed briefly with PBS, and transferred to a solution of 30% sucrose in PBS until they sank. Embryos were then embedded in Neg-50 (Richard Allan Scientific, San Diego, CA), frozen on dry ice, and stored at −80°C until they were sectioned. Immunohistochemistry was done using 20-um frozen sections cut on a Leica CM3050 S cryostat (Buffalo Grove, IL, USA). Prior to staining, sections were equilibrated to room temperature for at least 30 minutes. Antigen retrieval was performed using a citrate-based buffer (Antigen Unmasking Solution H-3300; Vector Laboratories, Burlingame, CA, USA) and steam for 20 minutes. Sections were then blocked in 2% normal goat serum in PBS with 0.1% Triton X-100 (PBS-TritonX), incubated in primary antibody overnight at 4°C, washed 3 × 3 minutes with PBS-TritonX, incubated in secondary antibody for 1 hour at room temperature, washed 2 × 3 minutes with PBS-TritonX, stained with DAPI (1:10,000) for 3’ in PBS, washed one last time for 3 minutes with PBS, and mounted in Fluoromount-G mounting media (Thermo Fisher Scientific, Waltham, MA, USA). Images were captured on a Zeiss LSM 700 series laser scanning confocal microscope (Carl Zeiss Meditec, Oberkochen, Germany).
Fluorescent Whole-Mount Embryo Immunohistochemistry
iDISCO
Whole-mount E13.5, 14.5, and E15.5 embryos were prepared using a modified version of the previously described iDISCO+ protocol (Renier et al., 2014; Whitman et al., 2019). Embryos were first collected and fixed overnight in 4% paraformaldehyde at 4°C, washed with PBS, dehydrated through a graded methanol series, bleached overnight in 5% H2O2 in methanol, rehydrated through a graded methanol series, and washed in PBS with 2% Triton-X. Embryos were then incubated at 37°C in permeabilization solution for 2 days, blocking solution for 2 days, and incubated in primary antibody diluted in PBS with 2% Tween-20 and heparin (PtWH), 5% DMSO, and 3% donkey serum for 7 days, washed with PtWH for one day at room temperature, and incubated with secondary antibody diluted in PtWH with 3% donkey serum at 37°C for 7 days. After staining, embryos were washed in PtWH, dehydrated through a graded methanol series, and incubated in a methanol and dichloromethane solution. Embryos were then cleared using dibenzyl ether (DBE). Embryos were mounted on coverslips in DBE and imaged using a Zeiss LSM 700 series laser scanning confocal microscope. Images were acquired with Zen Software (Carl Zeiss Micro-Imaging GmbH, Göttingen, Germany) and manipulated in three dimensions using Arivis software (Zeiss, Rostock, Germany). To facilitate 3D visualization of orbital structures at E15.5, arteries were selected using the find large objects feature and selectively cropped out of displayed images.
Quantification
Orbital Images were reconstructed in Arivis software (Zeiss, Rostock, Germany). EOMs were selected using the find large objects feature, and total volume of EOMs calculated. Graphs and statistical analysis (student’s t test) were done using GraphPad software.
Cleared Orbital Sections
E12.5-E15.5 mouse embryos were collected and heads were fixed overnight in 4% paraformaldehyde at 4°C with orbital shaking. After a wash with phosphate buffered saline (PBS), embryo heads were fixed overnight in Dent’s fix (80% Methanol/20%DMSO). Samples then incubated with primary antibody diluted in PBST (0.5% TritonX-100, 10% NGS) for 2–5 days (age dependent) at 4°C followed by several washes in PBST. Final washes done with PBS. anti-SMA used for wholemount EOM preps is Cy-3 conjugated, mitigating the need for a secondary antibody incubation. Samples were dehydrated through a series of 1 hr methanol incubations of increasing concentration (25%, 50%, 75%, 100%) and then cleared for several days in glass vials filled with 1:2 benzyl alcohol:benzyl benzoate (BABB) and stored at RT until imaging. Samples oriented in a 3D cut imaging chamber sandwiched between two glass coverslips and submerged in BABB. Images of EOMs were collected with a Zeiss LSM700 laser scanning confocal microscope and a 10X, 0.45 NA ApoChromat objective. Visualization of the orbit and annulus defined limits of the z-stack. Post-image processing was performed with Zen (Zeiss).
Antibodies
Mouse Anti-Actin, a-Smooth Muscle-Cy3 (1:750, RRID AB 476856; MilliporeSigma, Burlington, MA, USA), rabbit anti-GFP (1:500, RRID AB221569; Thermo Fisher Scientific, Boston, MA, USA), Mouse anti-MYOD (1:500, RRID AB 1098295; Thermo Fisher Scientific, Waltham, MA, USA), Mouse Anti-PITX2 (1:500, RRID AB 509266; Abnova, Taipei City, Taiwan), Alexa-Fluor 546 goat anti-mouse(1:1000, RRID AB 141370; Molecular Probes, Eugene, OR, USA), Alexa-Fluor 647 goat anti-rabbit (1:1000, RRID AB 141663; Invitrogen, Carlsbad, California, USA,), Alexa-Fluor 647 donkey anti-mouse (1:1000, RRID AB 162542; Molecular Probes, Eugene, OR, USA).
Results
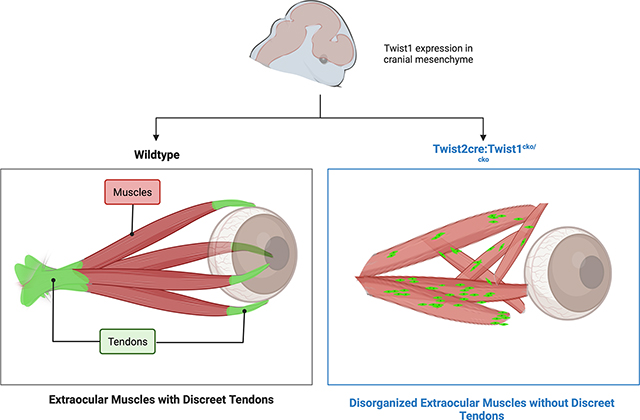
Loss of Twist1 in subpopulations of cranial mesenchyme results in disorganized EOMs.
To determine the role of Twist1 expression on EOM precursor differentiation and development, we produced conditional knock-outs using mesenchymal cre drivers with later, more limited expression than Mesp1-cre. In these animals, the neural tube closes and head formation is grossly normal (Tischfield et al., 2017).
We first examined conditional loss of Twist1 with Twist2-cre. TWIST2 is a bHLH transcription factor with high homology to Twist1. Spatially, Twist1 and Twist2 have similar expression patterns in mesenchymal tissues, but temporally Twist2 is expressed after Twist1 (Franco et al., 2011). Twist2 is expressed in medial cranial mesenchyme as early as E8.5 and E9.5 and is involved in maintenance of mesenchymal state (Bildsoe et al., 2016). Twist2-cre expression has been shown as early as E9.5 in mesenchymal tissues (He et al., 2017), Supplemental Figure 1). In these animals at E13.5, we found that the EOMs are present, but are severely disorganized compared to littermate controls that do not express cre and/or are wildtype for the Twist1 allele (Figure 1A–B). The annulus can be identified but the extraocular muscle fibers do not all appear to originate from it. Moreover, individual muscles cannot be identified beyond a putative retractor bulbi muscle (RB), and a muscle that may be the superior oblique, as it appears to go through the trochlea. Muscle fibers are present at a variety of orientations within the orbit, and the insertions to the globe cannot be identified. The total muscle volume of the EOMs was quantified and the mutant animals showed an increase in total volume (17.16+/−2.89 mm3 in mutants, vs 12.37 +/− 1.48 in controls, p=0.0002). One day earlier, at E12.5, the individual muscles cannot be identified in the mutants (not shown).
Figure 1.

Loss of Twist1 in different subsets of mesenchyme leads to varying degrees of EOM disorganization. Whole mounts from E13.5 embryos stained with anti-smooth muscle actin and imaged coronally show (A) the normal arrangement of the EOMs around the eye (dashed circle indicates left eye with red EOMs labeled) and the arteries of the anterior head. To the right of each image is a 3D reconstruction of the EOMs. (B) Loss of Twist1 in Twist2-cre expressing cells leads to profound disorganization of the EOMs. There is a muscle superiorly that appears to make a turn as if going through the trochlea, which may be the superior oblique (labeled SO?). This muscle, however, appears different from the wild-type superior oblique, in which muscle tissue extends only to the trochlea, not through it (compare to SO in panel A; in wild-type mice, a tendon (not labeled) extends from the globe to the trochlea). The RB and annulus of Zinn can be identified (arrow), but the other EOMs do not separate into distinct muscles. The total volume of EOMs is greater in the mutant animals (graph, inset). (C) Loss of Twist1 in Pdgfrb-cre expressing cells leads to mild disorganization of the EOMs. The SR, SO, and RB can be identified, but the other EOMs do not fully separate into distinct muscles, and there are muscle fibers oriented obliquely across the orbit, not originating at the apex (arrow). Consistent with the less severe phenotype, the total volume of the EOMs is not different from littermate controls (graph, inset). RB: retractor bulbi, IR: inferior rectus, MR: medial rectus, SR: superior rectus, LR: lateral rectus, SO: superior oblique, IO inferior oblique. ACA: anterior cerebral artery. Scale bar equals 200 um. n = 3–6 for each genotype. Control images are from littermate controls who do not express cre recombinase and/or have wildtype Twist1 alleles.
To confirm our findings, we examined knock-out of Twist1 with Pdgfrb-cre (Cuttler et al., 2011), a similar mesenchymal marker, which diffusely labels cells in the head, including neural crest and mesoderm-derived mesenchyme (Tischfield et al., 2017). These mice have a similar, although less severe, phenotype with disorganization of the EOMs (Figure 1C). The RB is positioned normally, and the superior rectus (SR) and superior oblique (SO) muscles also appear properly positioned. The medial rectus (MR), lateral rectus (LR) and inferior oblique (IO) muscles, however, do not separate properly into distinct muscles, and there are ectopic muscle fibers across the orbit (white arrows in Figure 1C). Consistent with the milder phenotype, the total volume of the EOMs was not different in mutant animals, compared to littermate controls (14.61 +/− 2.59 vs 14.03 +/− 2.32, p=0.67). Together, these findings indicate that expression of Twist1 in mesenchyme is necessary for EOM organization.
To further elucidate the extent to which Twist1 is required in mesenchymal tissues for EOM organization we focused the rest of our investigations on the animals with the more severe EOM organization phenotype, using the Twist2-cre driver.
The abnormal muscle has normal PITX2 and MYOD expression
The transcription factor PITX2 plays an important role in EOM differentiation and maintenance (Diehl et al., 2006; Zhou et al., 2009; Zhou et al., 2011), and loss of Twist1 under Mesp1-cre leads to loss of Pitx2 expression (Bildsoe et al., 2013). To determine if the disorganization of the EOMs seen with loss of Twist1 under Twist2-cre is mediated by PITX2, we examined PITX2 expression in Twist2:cre:Twist1cKO/cKO embryos. MYOD is a transcription factor downstream of PITX2 that is required for myogenic differentiation. PITX2 and MYOD expression are similar in EOMs of the mutant and control mice at E13.5 (Figure 2), a developmental time at which disorganization of the EOMs is already observed, indicating that there are additional factors downstream of TWIST1 that influence EOM organization.
Figure 2.

PITX2 and MYOD expression are normal in the disorganized EOMs. Sagittal frozen sections stained with anti-PITX2 antibody (green) and anti-MYOD antibody (red) show equal PITX2 and MYOD expression in the Twist2-cre:Twist1cKO/cKO embryos (B) compared to littermate controls (A). A and B are low-magnification views of the eye and orbit, demonstrating the disorganized muscles in B. A’ and B’ show high magnification images of individual EOMs, with individual channels displayed to the right. Scale bar in B equals 200 um for A and B; scale bar in B’ equals 50um for all other panels. n = 6 for each genotype.
Twist1 expression is required for the proper formation of EOM tendons
The insertion of EOMs onto their appropriate scleral target requires coordinated development of EOM tendons. Thus, to examine the tendons, origins, and scleral insertions of the disorganized muscles, we used Scx-GFP reporter mice, which express GFP in tendons throughout the body (Pryce et al., 2007). In the periocular region of E15.5 control mice, GFP signal is discretely concentrated at the origin of the EOMs (annulus of Zinn) and at the points of insertion on the sclera (Figure 3A,C,E). By contrast, the disorganized muscles of E15.5 Twist2cre:Twist1cKO/cKO embryos do not have identifiable tendons inserting on sclera (Figure 3B, D). There are areas of GFP expression along the belly of some EOMs (white arrowheads in 3B, 3F). In some cases, there appears to be a tendon condensation at the end of a disorganized muscle, but it is not inserting on the sclera (white arrow in 3B, see 3F).
Figure 3.

The disorganized EOMs have poorly organized tendons. Heads from E15.5 embryos expressing GFP from the scx promoter were whole mounted, stained with anti-GFP (green) and anti-smooth muscle actin (red) antibodies, cleared, and imaged coronally on a confocal microscope. Max intensity projections are shown in A and B, orthogonal views in C-F. In control embryos (A,C,E), tendons are at the apex, scleral insertion of each muscle, origin of the SO and IO muscles (blue arrows) and at the trochlea. In the disorganized EOMs caused by loss of Twist1 in Twist2 expressing cells (B,D,F), there are no discrete identifiable tendons inserting on the eyeball. There is a condensation of scx-expressing cells at the end of a muscle (white arrow) in the middle of the orbit, and there is diffuse expression of scx (white arrowheads) along the length of some muscles. The orthogonal views in C-F are at the level indicated in the upper right inset of each panel. The lower left shows a single Z-plane, with orthogonal views along the red and green lines to the right and above, respectively. C and D are at the same level, through the insertion of the LR and MR in the control. It can be seen in D that there are no discreet muscle insertions with tendons. F is of the area of the lower white arrowhead in B, and E is an equivalent plane in the control. Scale bar equals 200um. For A and B, images were reconstructed in Arivis software and the arteries were cropped out to facilitate visualization. n=4 for each genotype.
Twist1 expression is not required in tendon precursors for normal EOM or tendon development
Since the tendons do not form properly in the disorganized EOMs, we next asked whether Twist1 expression in the tendon precursor cells is necessary for proper EOM and tendon development. Scleraxis-cre (Scx-cre)(Blitz et al., 2009) is expressed in tendon precursor cells of the EOMs by E11.5 (Comai et al., 2020). In limb muscles, Scx-cre also marks a small subset of muscle cells, concentrated near the tendon (Esteves de Lima et al., 2021). We have noted sparse labeling of both mesenchymal and neuroectodermal tissue with Scx-cre as early as E9.5 (Supplemental Figure 2). When Twist1 is knocked out under the control of Scx-cre, EOMs develop normally, with normal final patterning (Figure 4). Quantification of the EOM volume showed no significant differences between Scx-cre:Twist1cko/cko embryos and littermate controls who either do not express cre or have a wild-type Twist1 allele (E14.5: 22.0 +/− 3.3 vs 25.1 +/− 3.8, p=0.17). The tendons also form normally, as evidenced by Scx-GFP expression at the origin and insertion of each EOM (Figure 4C–D). Therefore, the abnormal EOM and tendon organization seen with mesenchymal loss of Twist1 is not secondary to loss of Twist1 in the tendon precursors. Additionally, tendon formation is not dependent on Twist1 expression in tendon precursor cells and the tendon abnormalities seen in Twist2-cre:Twist1cko/cko animals are secondary to abnormal EOM development.
Figure 4.

Loss of Twist1 in Scx expressing cells does not disrupt EOM organization or tendon formation. Thick sections from E14.5 embryos stained with anti-smooth muscle actin (red) and imaged coronally show the normal arrangement of the EOMs in both control embryos (A) and Scx-cre:Twist1cKO/cKO embryos (B). Orbits from whole mounts of E13.5 embryos expressing GFP from the scleraxis promoter show normal tendon formation at the apex (arrow) and end of each muscle in both control embryos (C) and Scx-cre:Twist1cKO/cKO embryos (D). Images in C and D were reconstructed in Arivis and are oriented looking into the orbit towards the apex. Abbreviations as per Figure 1. n= 5 Scx-cre+, 3 littermate controls. Scale bar in B equals 100um.
Abnormal configuration of the EOMs leads to abnormal branching of the cranial motor neurons
The EOMs are innervated by the axons of oculomotor (CN3), trochlear (CN4), and abducens (CN6) cranial motor neurons that develop as the muscles are developing. At E10.5, axons have exited the motor nuclei and begun growing towards the orbit. A few pioneer axons contact PITX2+ cells ventrolateral to the eye as early as E10.0 (Bjorke et al., 2021). The majority of axons reach the orbit around E11.5, as the muscle anlage is differentiating (Bjorke et al., 2021; Michalak et al., 2017). Normally, CN3 travels ventrally from the oculomotor nucleus in the midbrain, through the region that will become the cavernous sinus, and enters the orbit. The superior division branches off from the main oculomotor trunk to innervate the superior rectus and levator palpabrae superioris muscles. The main trunk becomes the inferior branch and forms a decision region, which sends short branches to the medial rectus and inferior rectus muscles and a long branch to the inferior oblique muscle. CN4 exits the midbrain dorsally, crosses the midline, then courses ventrally to the opposite orbit, where it innervates the superior oblique muscle. CN6 travels from its origin in the hindbrain, along the retractor bulbi, to innervate the lateral rectus muscle (Figure 5A). We have previously shown that in the absence of the EOMs, the cranial nerves target the orbit correctly, but fail to make terminal branches (Michalak et al., 2017). In the presence of disorganized EOMs in the Twist2-cre:Twist1cko/cko, the cranial nerves also target the orbit correctly, but then branch abnormally (Figure 5B–D). CN4 is defasciculated and sends multiple branches to the superior aspect of the orbit, some of which do not appear to be contacting muscle tissue. CN3 forms a large decision region near the origin of the EOMs from which multiple aberrant branches extend. In some embryos, a branch resembling the wildtype branch to the inferior oblique forms, but exits the inferior decision region at an abnormal angle and appears to make two right-angle turns to course aberrantly towards the anterior orbit (Figure 5B–C). CN6 can be seen coursing along the retractor bulbi muscle (which appears the most normal and identifiable of the EOMs), but does not contact any muscles in the orbit. These observations suggest that appropriate EOM organization is required for the terminal branching and insertion of cranial motor neurons.
Figure 5.

Disorganization of the EOMs leads to abnormal branching of the innervating cranial motor neurons. E13.5 embryos expressing GFP from the Isl1MN promoter were whole mounted, stained with anti-GFP (green) and anti-smooth muscle actin (red, labels muscles and arteries) antibodies, cleared, and imaged sagittally on a confocal microscope. In a control embryo (A) CN4 innervates the SO muscle, CN3 innervates the MR, IR, IO and SR, and CN6 innervates the LR. In Twist2-cre:Twist1cKO/cKO embryos (B-D, examples of three different embryos), the EOMs are disorganized and the ocular motor nerves have abnormal branching patterns. CN4 is defasciculated and sends aberrant branches to the superior aspect of the orbit, as well as to muscle fibers that may be SO. CN3 enters the orbit in the correct location but then extends exuberant aberrant branches in addition to normal branches, and CN6 enters the orbit but cannot be visualized innervating any muscle. Scale bar in A equals 300um in A and B. Scale bar in C equals 300um in C and D. Abbreviations as in Figure 1. n=6 of each genotype.
Discussion
The EOMs are a unique muscle group based on their molecular composition and function (Porter, 2002). EOM and orbital development is also unique because tissues are derived from different embryological origins. Orbital connective tissue at the level of the EOM insertions is derived from neural crest cells (NCCs), while the connective tissue at the level of the EOM origin at the annulus of Zinn originates from the cranial mesoderm (Comai et al., 2020). Although much is known about the myogenic transcription factors required for initial EOM differentiation, little is known about how the distinct muscles form and project from the initial anlage of muscle precursors. Here, we show that mesenchymal expression of the transcription factor TWIST1 is necessary for the proper organization of EOMs into distinct muscles and formation of normal tendon insertions onto the sclera.
Knock-out of Twist1 under either Pdgfrb-cre or Twist2-cre leads to disorganization of the EOMs, of varying severity. Both Pdgfrb-cre and Twist2-cre are expressed in cranial mesenchyme of both mesodermal and cranial neural crest origin. Twist2 expression begins as early as E8.5 and is relatively enriched in the cranial mesoderm versus the cranial neural crest, although it is expressed in both (Bildsoe et al., 2016). Pdgfrb is expressed in mesenchyme throughout the body as early as E10.5 (Shinbrot et al., 1994). The conditional loss of Twist1 under these cre-recombinase-expressing lines allows for the interrogation of Twist1’s role in EOM development distinct from its role in neural tube closure, unlike Mesp1-cre. The disorganization of the EOMs we show here with these conditional knockouts of Twist1 reveals that Twist1 expression is necessary for proper differentiation of the EOMs into distinct muscles. The Twist2:cre mutants also displayed a larger total EOM volume. This could be the result of disruption of the normal control of muscle precursor division, leading to overproduction of muscle cells, or could indicate that the process of individual muscle separation/differentiation also influences muscle size.
We hypothesized that the effects of loss of Twist1 may be mediated by Pitx2 for several reasons: Pitx2 is required for EOM development (Gage et al., 1999); Pitx2 expression is regulated by retinoic acid (Bohnsack et al., 2012; Kumar and Duester, 2010); and blockade of retinoic acid signaling, either genetically or pharmacologically during early development of the initial EOM anlage, also leads to abnormal EOM patterning (Bohnsack et al., 2012; Comai et al., 2020). PITX2 expression was not altered in the mutant muscles, however, nor was its downstream target MYOD, indicating both that PITX2 and MYOD expression are not sufficient for normal EOM differentiation and that Twist1 is acting through an alternate pathway to control EOM patterning.
In addition to the disorganization of the EOMs, we also show that tendons do not form properly in Twist2-cre:Twist1cko/cko mice. Tendons form from Scx-expressing precursors (Murchison et al., 2007; Schweitzer et al., 2001). In the limbs, the tendons attach muscle to bone; bone superstructures provide anchoring points for tendons. These bony superstructures initially form independent of the muscle from precursors that express Scx and Sox9 (Blitz et al., 2009; Sugimoto et al., 2013); later maintenance and growth of these bony structures requires signals from both tendon and muscle (Huang, 2017). Unlike limb muscles, however, EOMs insert onto sclera of the eyeball rather than onto bone and there are no bony superstructures for the tendons to insert onto. Analogous to development of the bony superstructures, Scx and Sox9 are also expressed in the periocular mesenchyme, initially broadly and then restricted to the tendon insertion sites by E13.5 (Comai et al., 2020). In Twist2-cre:Twist1cko/cko mice, instead of distinct, Scx-expressing tendons at the origin and insertion of each EOM, mutant animals have Scx-expressing cells in clumps throughout the EOMs. Some clumps do appear to be at the end of a muscle, but not attached to sclera. Scx-GFP has been reported to also label muscle connective tissue (Comai et al., 2020) and some myogenic cells come from a Scx lineage (Esteves de Lima et al., 2021), so this ectopic GFP expression may represent abnormal muscle or connective tissue differentiation in the mutant animals, with resulting abnormal expression of scleraxis. To determine the role of Twist1 expression in the tendon precursor cells, we knocked out Twist1 under the control of Scx-cre, but this did not result in any abnormalities of EOM or tendon formation. These results suggest that the disorganized tendons are not caused by lack of Twist1 expression in the tendon precursor cells, but instead that normal EOM organization is required for proper tendon formation. Thus, the lack of tendon formation is a secondary effect, and tendon formation is dependent on proper EOM formation.
Our results also provide further evidence that EOM identity influences axon guidance decisions of the innervating cranial motor neurons. In the presence of disorganized muscles, the axon decision regions also appear disorganized. Neither Twist1 nor Twist2-cre are expressed in motor neurons, so these neuronal changes must be secondary to the muscle changes. In the complete absence of EOMs, motor neurons target the orbit correctly, but then fail to make correct terminal branches and many later die (Bjorke et al., 2021; Michalak et al., 2017). The exact branching errors depend on the genetic strategy used to ablate the EOMs. With the Myf5 cre/cre allele, which disrupts both Myf5 and Mrf4, axons fail to make terminal branches and many die (Michalak et al., 2017). In Myf5-DTA mutants, in which Myf5 expressing cells are killed, some embryos have a similar lack of branches and some form ectopic branches (Bjorke et al., 2021). By contrast, Pitx2−/− embryos, which disrupt an earlier stage of muscle development, show more severe axon errors, with a failure to stop near the eye and long, misdirected branches (Bjorke et al., 2021). Here, we find that in the presence of disorganized muscles, instead of failing to make branches, the motor neurons make aberrant branches, and often these branches do not appear to be contacting any muscle. In this way, they are most similar to the Pitx2 knockouts. The EOMs in the Twist mutants are not just spatially disorganized, but are also missing key elements of EOM differentiation. Thus, these findings suggest that individual EOMs express or secrete cues that guide the appropriate incoming axons to them, and that such expression is disrupted by loss of Twist1 or the resulting disorganization and lack of individual muscle identity. The identity of the cues that lead incoming axons to the proper EOM are not currently known.
We have shown that mesenchymal expression of Twist1 is necessary for the proper separation of the EOM muscle anlage into distinct EOMs. Secondarily, properly formed EOMs are required for appropriate tendon formation and for axon guidance of the cranial motor nerves. Strabismus in patients with TWIST1 variants, therefore, may be caused by abnormal development of the EOMs and resulting abnormalities in tendon formation and innervation.
Supplementary Material
Supplemental Figure 1: Twist2-cre expression. Coronal images from E9.5 Twist2-cre:TdTomato fluorescent reporter embryos shows sparse labeling in the mesenchyme surrounding the optic vesicle (A, B, D) and more robust labeling in more posterior mesenchyme (B,C). There is no labeling in neuronal tissues. Scale bar in C equals 200um in A-C. Scale bar in D equals 50um.
Supplemental Figure 2: Scx-cre expression. Coronal images from E9.5 Scx-cre:TdTomato fluorescent reporter embryos shows sparse labeling in the mesenchyme (A-C). Note the labeling in some neuronal tissues. Scale bar in C equals 200um in A-C.
Highlights.
Loss of TWIST1 in cranial mesenchyme leads to disorganized extraocular muscles
Disorganized extraocular muscles have abnormal tendon formation
TWIST1 is not required in tendon precursors for normal extraocular muscle and tendon development
Cranial motor neurons make abnormal branches in the presence of disorganized muscles
Acknowledgements
We thank Ronen Schweitzer for the ScxGFP & ScxCre mice. Funding provided by the National Eye Institute (NEI) 5K08EY027850 (MCW), Children’s Hospital Ophthalmology Foundation (MCW), Boston Children’s Hospital Faculty Development Award (MCW), Boston Children’s Hospital Intellectual and Developmental Disabilities Research Center (NICHD 1U54HD090255). National Institute of General Medical Sciences (NMG) (T32GM007753). ECE is a Howard Hughes Medical Institute Investigator. The content is solely the responsibility of the authors and does not necessarily represent the official views of the national Institute of General medical Sciences or the National Institutes of Health. This article is subject to HHMI’s Open Access to Publications policy. HHMI lab heads have previously granted a nonexclusive CC BY 4.0 license to the public and a sublicensable license to HHMI in their research articles. Pursuant to those licenses, the author-accepted manuscript of this article can be made freely available under a CC BY 4.0 license immediately upon publication.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Bildsoe H, Fan X, Wilkie EE, Ashoti A, Jones VJ, Power M, Qin J, Wang J, Tam PPL, Loebel DAF, 2016. Transcriptional targets of TWIST1 in the cranial mesoderm regulate cell-matrix interactions and mesenchyme maintenance. Dev Biol 418, 189–203. [DOI] [PubMed] [Google Scholar]
- Bildsoe H, Loebel DA, Jones VJ, Chen YT, Behringer RR, Tam PP, 2009. Requirement for Twist1 in frontonasal and skull vault development in the mouse embryo. Dev Biol 331, 176–188. [DOI] [PubMed] [Google Scholar]
- Bildsoe H, Loebel DA, Jones VJ, Hor AC, Braithwaite AW, Chen YT, Behringer RR, Tam PP, 2013. The mesenchymal architecture of the cranial mesoderm of mouse embryos is disrupted by the loss of Twist1 function. Dev Biol 374, 295–307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bjorke B, Weller KG, Jones LE, Robinson GE, Vesser M, Chen L, Gage PJ, Gould TW, Mastick GS, 2021. Oculomotor nerve guidance and terminal branching requires interactions with differentiating extraocular muscles. Dev Biol 476, 272–281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blitz E, Viukov S, Sharir A, Shwartz Y, Galloway JL, Pryce BA, Johnson RL, Tabin CJ, Schweitzer R, Zelzer E, 2009. Bone ridge patterning during musculoskeletal assembly is mediated through SCX regulation of Bmp4 at the tendon-skeleton junction. Developmental cell 17, 861–873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bohnsack BL, Kasprick DS, Kish PE, Goldman D, Kahana A, 2012. A zebrafish model of axenfeld-rieger syndrome reveals that pitx2 regulation by retinoic acid is essential for ocular and craniofacial development. Investigative ophthalmology & visual science 53, 7–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bothe I, Dietrich S, 2006. The molecular setup of the avian head mesoderm and its implication for craniofacial myogenesis. Developmental dynamics : an official publication of the American Association of Anatomists 235, 2845–2860. [DOI] [PubMed] [Google Scholar]
- Bourgeois P, Bolcato-Bellemin AL, Danse JM, Bloch-Zupan A, Yoshiba K, Stoetzel C, Perrin-Schmitt F, 1998. The variable expressivity and incomplete penetrance of the twist-null heterozygous mouse phenotype resemble those of human Saethre-Chotzen syndrome. Human molecular genetics 7, 945–957. [DOI] [PubMed] [Google Scholar]
- Buckingham M, Rigby PW, 2014. Gene regulatory networks and transcriptional mechanisms that control myogenesis. Developmental cell 28, 225–238. [DOI] [PubMed] [Google Scholar]
- Carver EA, Oram KF, Gridley T, 2002. Craniosynostosis in Twist heterozygous mice: a model for Saethre-Chotzen syndrome. Anat Rec 268, 90–92. [DOI] [PubMed] [Google Scholar]
- Chen YT, Akinwunmi PO, Deng JM, Tam OH, Behringer RR, 2007. Generation of a Twist1 conditional null allele in the mouse. Genesis 45, 588–592. [DOI] [PubMed] [Google Scholar]
- Chen ZF, Behringer RR, 1995. twist is required in head mesenchyme for cranial neural tube morphogenesis. Genes & development 9, 686–699. [DOI] [PubMed] [Google Scholar]
- Comai GE, Tesarova M, Dupe V, Rhinn M, Vallecillo-Garcia P, da Silva F, Feret B, Exelby K, Dolle P, Carlsson L, Pryce B, Spitz F, Stricker S, Zikmund T, Kaiser J, Briscoe J, Schedl A, Ghyselinck NB, Schweitzer R, Tajbakhsh S, 2020. Local retinoic acid signaling directs emergence of the extraocular muscle functional unit. PLoS Biol 18, e3000902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cuttler AS, LeClair RJ, Stohn JP, Wang Q, Sorenson CM, Liaw L, Lindner V, 2011. Characterization of Pdgfrb-Cre transgenic mice reveals reduction of ROSA26 reporter activity in remodeling arteries. Genesis 49, 673–680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diehl AG, Zareparsi S, Qian M, Khanna R, Angeles R, Gage PJ, 2006. Extraocular muscle morphogenesis and gene expression are regulated by Pitx2 gene dose. Investigative ophthalmology & visual science 47, 1785–1793. [DOI] [PubMed] [Google Scholar]
- Dollfus H, Biswas P, Kumaramanickavel G, Stoetzel C, Quillet R, Biswas J, Lajeunie E, Renier D, Perrin-Schmitt F, 2002. Saethre-Chotzen syndrome: notable intrafamilial phenotypic variability in a large family with Q28X TWIST mutation. Am J Med Genet 109, 218–225. [DOI] [PubMed] [Google Scholar]
- Esteves de Lima J, Blavet C, Bonnin MA, Hirsinger E, Comai G, Yvernogeau L, Delfini MC, Bellenger L, Mella S, Nassari S, Robin C, Schweitzer R, Fournier-Thibault C, Jaffredo T, Tajbakhsh S, Relaix F, Duprez D, 2021. Unexpected contribution of fibroblasts to muscle lineage as a mechanism for limb muscle patterning. Nat Commun 12, 3851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fan X, Loebel DAF, Bildsoe H, Wilkie EE, Qin J, Wang J, Tam PPL, 2016. Tissue interactions, cell signaling and transcriptional control in the cranial mesoderm during craniofacial development. AIMS Genetics 3, 74–98. [Google Scholar]
- Franco HL, Casasnovas J, Rodriguez-Medina JR, Cadilla CL, 2011. Redundant or separate entities?--roles of Twist1 and Twist2 as molecular switches during gene transcription. Nucleic acids research 39, 1177–1186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gage PJ, Suh H, Camper SA, 1999. Dosage requirement of Pitx2 for development of multiple organs. Development 126, 4643–4651. [DOI] [PubMed] [Google Scholar]
- Gallagher ER, Ratisoontorn C, Cunningham ML, 1993. Saethre-Chotzen Syndrome, in: Adam MP, Mirzaa GM, Pagon RA, Wallace SE, Bean LJH, Gripp KW, Amemiya A (Eds.), GeneReviews((R)), Seattle (WA). [PubMed] [Google Scholar]
- Grimaldi A, Tajbakhsh S, 2021. Diversity in cranial muscles: Origins and developmental programs. Curr Opin Cell Biol 73, 110–116. [DOI] [PubMed] [Google Scholar]
- He Y, Sun X, Wang L, Mishina Y, Guan JL, Liu F, 2017. Male germline recombination of a conditional allele by the widely used Dermo1-cre (Twist2-cre) transgene. Genesis 55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang AH, 2017. Coordinated development of the limb musculoskeletal system: Tendon and muscle patterning and integration with the skeleton. Dev Biol 429, 420–428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kitamura K, Miura H, Miyagawa-Tomita S, Yanazawa M, Katoh-Fukui Y, Suzuki R, Ohuchi H, Suehiro A, Motegi Y, Nakahara Y, Kondo S, Yokoyama M, 1999. Mouse Pitx2 deficiency leads to anomalies of the ventral body wall, heart, extra- and periocular mesoderm and right pulmonary isomerism. Development 126, 5749–5758. [DOI] [PubMed] [Google Scholar]
- Kumar S, Duester G, 2010. Retinoic acid signaling in perioptic mesenchyme represses Wnt signaling via induction of Pitx2 and Dkk2. Dev Biol 340, 67–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuroda S, Adachi N, Kusakabe R, Kuratani S, 2021. Developmental fates of shark head cavities reveal mesodermal contributions to tendon progenitor cells in extraocular muscles. Zoological Lett 7, 3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewcock JW, Genoud N, Lettieri K, Pfaff SL, 2007. The ubiquitin ligase Phr1 regulates axon outgrowth through modulation of microtubule dynamics. Neuron 56, 604–620. [DOI] [PubMed] [Google Scholar]
- Michalak SM, Whitman MC, Park JG, Tischfield MA, Nguyen EH, Engle EC, 2017. Ocular Motor Nerve Development in the Presence and Absence of Extraocular Muscle. Investigative ophthalmology & visual science 58, 2388–2396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murchison ND, Price BA, Conner DA, Keene DR, Olson EN, Tabin CJ, Schweitzer R, 2007. Regulation of tendon differentiation by scleraxis distinguishes force-transmitting tendons from muscle-anchoring tendons. Development 134, 2697–2708. [DOI] [PubMed] [Google Scholar]
- Porter JD, 2002. Extraocular muscle: cellular adaptations for a diverse functional repertoire. Annals of the New York Academy of Sciences 956, 7–16. [DOI] [PubMed] [Google Scholar]
- Pryce BA, Brent AE, Murchison ND, Tabin CJ, Schweitzer R, 2007. Generation of transgenic tendon reporters, ScxGFP and ScxAP, using regulatory elements of the scleraxis gene. Developmental dynamics : an official publication of the American Association of Anatomists 236, 1677–1682. [DOI] [PubMed] [Google Scholar]
- Qin Q, Xu Y, He T, Qin C, Xu J, 2012. Normal and disease-related biological functions of Twist1 and underlying molecular mechanisms. Cell Res 22, 90–106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Renier N, Wu Z, Simon DJ, Yang J, Ariel P, Tessier-Lavigne M, 2014. iDISCO: a simple, rapid method to immunolabel large tissue samples for volume imaging. Cell 159, 896–910. [DOI] [PubMed] [Google Scholar]
- Rosenberg JB, Tepper OM, Medow NB, 2013. Strabismus in craniosynostosis. Journal of pediatric ophthalmology and strabismus 50, 140–148. [DOI] [PubMed] [Google Scholar]
- Saga Y, Miyagawa-Tomita S, Takagi A, Kitajima S, Miyazaki J, Inoue T, 1999. MesP1 is expressed in the heart precursor cells and required for the formation of a single heart tube. Development 126, 3437–3447. [DOI] [PubMed] [Google Scholar]
- Sambasivan R, Gayraud-Morel B, Dumas G, Cimper C, Paisant S, Kelly RG, Tajbakhsh S, 2009. Distinct regulatory cascades govern extraocular and pharyngeal arch muscle progenitor cell fates. Developmental cell 16, 810–821. [DOI] [PubMed] [Google Scholar]
- Sambasivan R, Kuratani S, Tajbakhsh S, 2011. An eye on the head: the development and evolution of craniofacial muscles. Development 138, 2401–2415. [DOI] [PubMed] [Google Scholar]
- Schweitzer R, Chyung JH, Murtaugh LC, Brent AE, Rosen V, Olson EN, Lassar A, Tabin CJ, 2001. Analysis of the tendon cell fate using Scleraxis, a specific marker for tendons and ligaments. Development 128, 3855–3866. [DOI] [PubMed] [Google Scholar]
- Shinbrot E, Peters KG, Williams LT, 1994. Expression of the platelet-derived growth factor beta receptor during organogenesis and tissue differentiation in the mouse embryo. Developmental dynamics : an official publication of the American Association of Anatomists 199, 169–175. [DOI] [PubMed] [Google Scholar]
- Sosic D, Richardson JA, Yu K, Ornitz DM, Olson EN, 2003. Twist regulates cytokine gene expression through a negative feedback loop that represses NF-kappaB activity. Cell 112, 169–180. [DOI] [PubMed] [Google Scholar]
- Stoler JM, Rogers GF, Mulliken JB, 2009. The frequency of palatal anomalies in Saethre-Chotzen syndrome. Cleft Palate Craniofac J 46, 280–284. [DOI] [PubMed] [Google Scholar]
- Sugimoto Y, Takimoto A, Akiyama H, Kist R, Scherer G, Nakamura T, Hiraki Y, Shukunami C, 2013. Scx+/Sox9+ progenitors contribute to the establishment of the junction between cartilage and tendon/ligament. Development 140, 2280–2288. [DOI] [PubMed] [Google Scholar]
- Tischfield MA, Robson CD, Gilette NM, Chim SM, Sofela FA, DeLisle MM, Gelber A, Barry BJ, MacKinnon S, Dagi LR, Nathans J, Engle EC, 2017. Cerebral Vein Malformations Result from Loss of Twist1 Expression and BMP Signaling from Skull Progenitor Cells and Dura. Developmental cell 42, 445–461 e445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitman MC, Miyake N, Nguyen EH, Bell JL, Matos Ruiz PM, Chan WM, Di Gioia SA, Mukherjee N, Barry BJ, Bosley TM, Khan AO, Engle EC, 2019. Decreased ACKR3 (CXCR7) function causes oculomotor synkinesis in mice and humans. Human molecular genetics 28, 3113–3125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou Y, Cheng G, Dieter L, Hjalt TA, Andrade FH, Stahl JS, Kaminski HJ, 2009. An altered phenotype in a conditional knockout of Pitx2 in extraocular muscle. Investigative ophthalmology & visual science 50, 4531–4541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou Y, Liu D, Kaminski HJ, 2011. Pitx2 regulates myosin heavy chain isoform expression and multi-innervation in extraocular muscle. J Physiol 589, 4601–4614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ziermann JM, Diogo R, Noden DM, 2018. Neural crest and the patterning of vertebrate craniofacial muscles. Genesis 56, e23097. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplemental Figure 1: Twist2-cre expression. Coronal images from E9.5 Twist2-cre:TdTomato fluorescent reporter embryos shows sparse labeling in the mesenchyme surrounding the optic vesicle (A, B, D) and more robust labeling in more posterior mesenchyme (B,C). There is no labeling in neuronal tissues. Scale bar in C equals 200um in A-C. Scale bar in D equals 50um.
Supplemental Figure 2: Scx-cre expression. Coronal images from E9.5 Scx-cre:TdTomato fluorescent reporter embryos shows sparse labeling in the mesenchyme (A-C). Note the labeling in some neuronal tissues. Scale bar in C equals 200um in A-C.