

Abstract
Implicit learning about new words by picking up on associative information in the contexts they appear in is an important aspect of vocabulary growth. The current study investigated the neural correlates that underlie how school-aged children and adolescents identify the meaning of novel words embedded within sentence contexts. Importantly, we examine how differences in the brain response to novel words and their context differ as a function of 1) explicit learning success, i.e., whether novel word meanings can be correctly estimated in isolation after a learning opportunity, and 2) individual differences in offline language aptitude as well as age across our cohort (N = 82; 8–16 years). Using a regression-based analysis, we identified the unique influence of these individuals difference metrics by using both measures within the same series of models. The most notable finding from our analysis was a frequency-specific dissociation between the way age and language abilities held relationships with task-relevant oscillatory activity during the novel word meaning task: language abilities associated with task-relevant changes in beta band activity during sentence processing, while age associated with task-relevant changes in theta band activity during pseudoword processing. These effects reflect the how the neural correlates of mapping semantic meaning from sentence contexts—an important skill for word learning—is uniquely influenced by the maturity of language abilities as well as age
Keywords: EEG, Time frequency analysis, Word learning, Language Processing, Semantic processing
1. Introduction
Semantic learning represents a critical foundational step in a multi-stage process where a new word is being mapped to a novel, unique, and robust meaning representation (Nagy et al., 1985, Nagy et al., 1987, Fukkink, 2005, Horst, 2013, Schneider et al., 2021). As children progress beyond the first stages of language development, the meaning of a new word is often rooted in semantic representations that the child is already familiar with (e.g., flounder is a type of fish).
Throughout childhood, continuous exposure to words over time and across different contexts actively refines and reorganizes their meaning. This complex process not only relies upon the child’s existing ability to explicitly reason about what words mean and how they relate to one another but involves a sensitivity to linguistic and contextual information that influences language processing and operates on a rapid timescale, often without explicit awareness. This might be a particularly relevant skill for development, as some traditional conceptions of age- related changes in language learning processes posit that children learn language through more implicit means, while language learning in adulthood involves explicit learning strategies (see Lichtman, 2012). Additionally, both within and outside the domain of language learning, research suggests that the ability to learn from context through implicit means may change during development, with particular shifts during middle childhood (Janacsek et al., 2012, Nemeth et al., 2013).
Little is known about the neural underpinnings that support the development of implicit word learning abilities. EEG work in adults has demonstrated that novel words can adopt some level of meaning attachment after only a single presentation within a familiar sentence context (Borovsky et al., 2010, Borovsky et al., 2012). Using similar neurophysiological indices of online language processing, work has also shown how the semantic information that novel words become associated with becomes more elaborated and specified over repeated exposures in different contexts (Mestres-Missé et al., 2007, Abel et al., 2018). Importantly, extant research has often relied on neural signatures of semantic processing to make inferences about implicit meaning attachment to novel words, while ignoring how these measures might associate with learning success, i.e., whether or not the learned information can be demonstrated explicitly through verbal report. Additionally, the majority of previous studies have not attempted to understand how differences across individuals may relate to their propensity to learn semantic information about new words in this way. The current study addresses these issues by considering how continuous learning performance can be mapped onto neurophysiological measures of language processing, while also considering how this brain-behavior relationship is modulated by various developmental factors, such as language abilities and age.
1.1. Oscillatory dynamics supporting language processing
The investigation of neural oscillatory dynamics using electroencephalography (EEG) allows researchers to build inferences about how changes in synchronized activity of large-scale networks in the brain underlie cognitive and linguistic processes. Resting-state EEG activity points to a developmental shift in the maturation of domain-general neural networks, characterized by decreasing synchrony in low-frequency bands (e.g., theta) alongside increasing synchrony in higher frequency bands (i.e., alpha, beta, gamma; Marshall et al., 2004; Maguire and Schneider, 2019). These changes represent an important index of the maturity and efficiency of cortical networks, as they gauge the coordination of distributed neural activity that are functionally related to anatomical changes during development (Uhlhaas et al., 2010; Buzsáki and Draguhn, 2004). More specific to language-related neural networks, the neural oscillations supporting online semantic retrieval appear to become more similar to adults by age 8 (Schneider and Maguire, 2019) whereas those supporting more complex aspects of online sentence processing, such as integrating syntactic information, may continue to develop through middle childhood (Schneider et al., 2018).
Past studies of sentence processing indicate that frequency dynamics in the theta (4–7 Hz) and beta (14–20 Hz) frequency ranges are centrally involved in sentence comprehension and lexical processing (Maguire et al., 2022, Lam et al., 2016; Bastiaansen et al., 2005; 2006; Schneider et al., 2016, Schneider et al., 2018; Lewis and Bastiaansen, 2015). Specifically, theta is hypothesized to reflect semantic retrieval processes during single-word and sentence comprehension and is often positively associated with retrieval demands from long-term memory. For example, open-class compared to closed-class words, as well as words that violate online semantic expectations, elicit a relative enhancement of theta power (Bastiaansen et al., 2005). A functional role for beta band activity during language comprehension has been posited under the predictive coding framework (Lewis and Bastiaansen, 2015). This theory is supported by reported relationships between online beta desynchronization in response to lexical prediction errors, while increases in beta power are thought to reflect the top-down maintenance of predicted information in memory during meaning construction (e.g., Keilar et al., 2014). In this scheme, beta band enhancement occurs when information in the sentence aligns with what is predicted to occur next while beta suppression occurs when input does not align with prior expectations about upcoming words (Lewis and Bastiaansen, 2015, Lewis et al., 2016).
1.2. The current study
Although research into the development of oscillatory neural dynamics underpinning semantic learning is relatively scarce, recent work with school-aged children has provided evidence that theta and beta activity are sensitive to differences in the word learning process between weak and strong learners from low SES homes (Schneider et al., 2021). Thus, oscillatory activity in theta and beta can provide a useful physiological marker for cognitive processes that support semantic learning (see also Bakker et al., 2015). More generally, the current scarcity of empirical investigations into the oscillatory dynamics that support language learning during development serves as motivation for the current study’s focus on these measures, as opposed to event-related potentials. Specifically, it remains unknown how language aptitude uniquely relates to the neurocognitive mechanisms that support word-meaning inference from context. Indeed, both age-related changes in the brain could contribute to neural differences that could mask how language abilities influence the brain dynamics supporting contextual word learning. Considering this motivation, the current study is designed to examine 1) how oscillatory activity is differentially engaged during processing of words, and the sentence context preceding them, when meaning is and is not mapped to a semantic representation and 2) how these neural signatures relate uniquely to language abilities and age. These aims were accomplished using a meaning identification task that assessed children’s ability to quickly build meaning representations for previously unknown words presented in meaningful contexts.
2. Methods
2.1. Participants and language assessments
104 right-handed school-aged children participated in both the behavioral testing and EEG portions of the study, described below (Mean age = 11 yrs 5mos.; σ = 2.5 years, Range: 8 years 0 mos. – 16 yrs 8 mos; 48 females). All children had normal or corrected-to-normal vision and no history of hearing disorder, language or learning disorder, traumatic brain injury or other significant neurological disorders (CVA, seizure disorder, tumor). Children were also either monolingual English speakers or had English as their primary language. To assess general language skill, all children were administered the Core Language subtests of the CELF-5 (Clinical Evaluation of Language Fundamentals, 5th Edition; Wiig et al., 2013). Due to the focus of this study on children with typical language development, a Core Language standard score at or above 85 (≥ −1 standard deviation) was used as an inclusion criterion. Data from 12 children were removed from the analysis due to scores below this cutoff. Three additional participant datasets were removed from the study due to excessive artifacts during EEG recording or because of technical difficulties during EEG stimulus presentation, as described below. A final cohort of 82 children were included in the final dataset (44 females; Mean age = 12 yrs 2 mos.; σ = 2.6 years, Range: 8 years 0 mos – 16 years 11mos; Mean CELF score = 111.3; σ = 11.3; range = 85–132). Parental consent and child assent was obtained in accordance with the Institutional Review Board of San Diego State University. This study was conducted according to the Good Clinical Practice Guidelines, the Declaration of Helsinki, and the U.S. Code of Federal Regulations.
2.2. Measure of language ability
Different subtests comprise the Core Language score for 8 year-olds, 9–15 year-olds, and 16 year-olds; however, all children completed the Formulating Sentences and Recalling Sentences subtests. Scores on these subtests were therefore combined to formulate our measure of language ability. These subtests contain developmentally appropriate scoring targets for children across a wide range of ages and could thus be used to compare relative abilities across our developmentally diverse sample. Raw scores derived from these subtests were z-scored to generate a continuous measure of language ability used in our statistical analysis.
2.3. Stimulus materials
Target words for the experiment were taken from a database of nouns commonly acquired before 3 years of age, and thus served as appropriate targets for even our youngest participants (MacArthur-Bates Communicative Developmental Inventory; Fenson et al., 2006). Sentence stimuli were written with these target nouns in the sentence-terminal position. All sentences were between 6 and 9 words in length and the target nouns were preceded by either a determiner (a/the) or possessive (my, your, his, her). They were all simple sentences containing early-acquired vocabulary (Fenson et al., 2006, Hall et al., 1980). Crucially, for the meaning identification task, the target nouns were replaced with pseudowords, as children were attempting to infer the meaning of the pseudoword using the surrounding information in each sentence context. Pseudowords that replaced the target nouns were generated using a database of CVC sequences (Storkel, 2013). Stimuli did not include any phonetic constraints for vowels or word-final consonants, and no pseudoword began with either /s/ or /sh/ in order to improve the time-locking accuracy to word onset. Pseudowords were included in the original sentence recordings, which was performed by a female native English speaker of General American English dialect at a natural speaking pace.
Three-hundred total sentences were normed offline by 248 adults using a sentence completion task to derive cloze probabilities for the target sentence-final nouns within their respective sentence contexts. Cloze probability for the target nouns was used to construct sentence triplets to be presented together within a single trial. The meaning identification task included 100 sentence triplets (300 total presentations of the pseudoword). Half of the 100 triplets involved a conditional manipulation that is not relevant to the current study, and thus only 50 total triplet presentations (150 presentations of the pseudoword) per participant were included in the current dataset (see description of conditions below). Sentence triplets were organized so that the first sentence provided relatively little contextual constraint for the particular target word, and successive sentences provided incrementally more constraint for the target word (Mean cloze probabilities for words in first sentence = 4%; second sentence = 40.6%; third sentence = 88.1%). Fig. 1 contains a depiction of a sample trial with an example sentence triplet.
Fig. 1.

Depiction of a single trial with an example sentence triplet used in the meaning identification task. Note that the target real word “bed” fits as a plausible ending for each sentence in the trial.
2.4. Procedure
To evaluate semantic learning, all children completed a meaning identification task during which their EEG was collected. The design of the meaning identification task was motivated by a paradigm used in previous word learning studies (Mestres-Missé, Rodriguez- Fornells & Münte, 2007; Abel et al., 2018; Maguire et al., 2018; Ralph et al., 2020; Schneider et al., 2021). While the majority of previous work has focused on the written modality for word and stimulus presentation, the current study used naturally-paced speech in order avoid potential constraints driven by differences in reading ability across children (see Momsen and Abel, 2022 for an identical study design in adults).
Participants listened to a series of 3 sentence, referred to here as a trial. Each trial ended with the same target pseudoword. This pseudoword served as the focus for the main experimental task—to verbally identify at the end of each trial whether a single real word could plausibly replace the pseudoword so that it fit all three sentence contexts.
The semantic relationship between the target pseudoword and sentence contexts was manipulated across 2 conditions: in the Meaning (+) condition, the target pseudoword stood to represent the same real word across all three sentences within the trial. In the Meaning (-) condition, the target pseudoword represented different real words within the trial, and thus the sentence contexts did not support a single meaning for the target. Only trials appearing in the Meaning (+) condition were used for the current study, therefore analyses related to the Meaning (-) condition will henceforth not be discussed.
Sentences were presented through two speakers flanking the computer monitor. As depicted in Fig. 1, each trial began with the presentation of fixation cross displayed centrally on the monitor for 600 ms. The fixation cross remained on the screen during each sentence to help minimize eye-movements during the recording. Following the offset of each sentence, a horizontal array of fixation crosses appeared, which served as a prompt for children to initiate the presentation of the next sentence (for sentences 1 and 2) or the test question (for sentence 3) at their pace with a single button push on a Logitech F310 game controller. This button push triggered a new single fixation cross for 600 ms before the onset of the subsequent sentence. This cycle repeated until the end of the presentation of the third sentence. Child-led pacing between sentences ensured that participants were able to comfortably perform the experimental task, thereby minimizing unwanted differences in task demands that might arise naturally across our diverse sample of children. After the completion of each trial, children were asked by a research assistant sitting in the room with them whether they believed the target pseudoword presented in that trial was a real word and, if so, what word they thought the pseudoword meant. Child responses were given verbally and recorded online. Trials were deemed correct if the provided word was identical to the target word that the stimulus was designed to represent or any alternative that reasonably fit in the sentence triplet (e.g., “truck” for the target word “car”). An incorrect response was either a response that there was no meaning for the pseudoword or a meaning that did not fit in the sentence triplet (e.g., “carrot” for the target word “car).
Participants completed two practice trials, one from each condition, providing feedback and repetition as necessary. No feedback or repetition was provided during the test trials.
2.5. EEG recording
EEG data was recorded from a 64 electrode Neuroscan Quickcap arranged in the 10–20 International Standard configuration. Participants were seated in a sound-attenuated recording chamber approximately 1 m from a color monitor. Data was sampled at 1 kHz during the recording session, and a centroparietal site near the scalp vertex was used as an online reference electrode. Recorded datasets were processed in MATLAB using the EEGLAB toolbox (Delorme and Makeig, 2004). Recorded data was resampled at 512 Hz after recording. Offline re-referencing was performed using the average of two near-mastoid electrodes (P9, P10), and these sites were removed from each dataset before statistical analysis. Continuous data was filtered using a 50 Hz low-pass filter and 0.01 Hz high-pass filter and was also trimmed to remove large sections of task-irrelevant data before further processing. Independent Component Analysis was used on each dataset to identify and remove sources of non-cortical activity representing blink or saccade related artifacts (Infomax ICA; Bell and Sejnowski, 1995). Independent Components were manually identified and removed from continuous datasets, while blocked or excessively noisy electrodes were corrected using spherical interpolation within the EEGLAB toolbox (Mean number of components removed per subject = 2.2).
2.6. Event‐Related Spectral Perturbation (ERSP) Analysis
Data epochs were created for each of the three sentences within a trial during the meaning identification task. EEG data from − 1000–1000 ms relative to the onset of the target pseudoword in each sentence rendered 150 total data epochs for each per participant (3 sentences per trial; 50 total trials). Data epochs containing remaining artifacts were identified via automated artifact detection (Moving window peak-to-peak, step-like threshold = 12 µV) and removed before statistical analysis. Considering the age range included in the current analysis (8–16 years), inter-individual differences in brain development and physical maturation during puberty may introduce amplitude variation into the EEG signals across our dataset (Arain et al., 2013, Hagemann et al., 2008, Myslobodsky et al., 1989). To account for this, automatic data flagging was accomplished after standardizing the EEG signals within each participant and channel to create conformity across datasets during artifact detection. This was achieved by z-scoring amplitude values at each time point within a single channel for each participant before artifact flagging. Data was adjusted back into non-standardized amplitudes after artifact flagging before further processing (Mean percent trials rejected per subject = 6.4%; σ = 6.7%).
Time-frequency representations (TFRs) were calculated using code adopted from Fieldtrip (Oostenveld, Fries, Maris, and Schoffelen, 2011). The transform used a single taper Hanning window (Rommers et al., 2017) with a length of 500 ms across 32 ms steps of the 2 s epochs. This resulted in TFRs across a frequency range of 3–30 Hz in 2 Hz steps which were then interpolated to render estimates across all integers between 3 and 30 Hz (1 Hz steps). The resulting transformation resulted segments of time-frequency data spanning from ∼742 ms prior to and following pseudoword onset (∼1485 ms total). Rather than using a pre-pseudoword baseline, EEG data recorded prior to the onset of the respective sentence was used to examine the neural activity during both sentence and pseudoword processing. The decision to use pre-sentence EEG activity as a baseline was motivated by the fact that this can dissociate how task-related changes in EEG activity, including potential interactions with child-age or language abilities, manifest in association with sentence processing vs novel word processing directly. Pre-sentence baseline windows for all subjects were separately extracted from each sentence and each trial, and the same time-frequency transformation pipeline was used to render data spanning from − 1000–1000 ms relative to sentence onset. Decibel correction for each sentence was performed by log transforming a portion of this baseline activity (400–100 ms pre-sentence onset) against the entire segment of sentence-related activity (∼1485 ms) across all trials and subjects individually (10 *log10(activity/baseline)). Throughout the remainder of the results and discussion, reported time information is relative to pseudoword onset at the 0 ms timestamp within all epochs. The baseline correction procedure was performed on a single-trial basis, resulting in 4D data structures containing Trial x Channel x Frequency x Time information for each subject. The same dB correction procedure was performed for all presented figures, albeit collapsing within sentence condition rather than keeping single-trial data information.
2.7. Statistical analysis
Non-parametric approaches to ERSP analysis, while popular for their strength in ameliorating problems associated with Type 1 error control (see Maris and Oostenveld, 2007) are less flexible in incorporating covariates and continuous variables that may interact with effects of interest. Motivated by our interest in separately modeling the influence of individual differences related to age and language abilities across children, we used single-trial multilevel modeling to model the influence of these variables on measures of neural activity during the online meaning identification task. The implemented analysis is an extension of the regression-based ERP approach to the time-frequency domain (Aurnhammer et al., 2021, Smith and Kutas, 2015). The rERP estimation technique is based on a series of regression models fit across each channel as well as each time sample across participants to assess the influence of experimental variables on the raw EEG waveforms. We apply a variation of this technique by using a series of generalized linear mixed effects models (GLMER) at each scalp location and time point (see Urbach et al., 2020). Random intercepts for each experimental item and subject were included to reduce estimation error unrelated to variables of interest (Baayen et al., 2008). Additionally, we reversed the typical directionality of these models in our approach by using a binomial representation of behavioral performance on the meaning identification task (correct vs incorrect word identification) as the outcome variable that the oscillatory signal and individual difference measures (age and language ability) were used to predict. The resulting set of models allowed us to identify the neural signature of online sentence processing that was behaviorally relevant for word identification. Importantly, any interactions between the oscillatory signal and individual difference measures would indicate how task relevant neural activity either during sentence or pseudoword processing changes uniquely as a function of age or language ability, which would provide evidence for relationships between language-specific vs age-related factors and online speech processing abilities in service of inferential word learning.
Following a priori predictions, two separate windows of ERSP activity were used in each analysis and corresponded to the frequency bands of interest: theta (4–7 Hz) and low beta (14–20 Hz), which are commonly modulated in similar language tasks both in adults (e.g., Bakker et al., 2015; Momsen and Abel, 2022) and school-aged children (e.g., Schneider et al., 2016). Single- trial measurements of ERSP data corresponding to these two frequency bands served as independent variables that were permitted to interact uniquely with z-scored vectors associated with child age as well as z-transformed raw scores on the CELF-5 subtests, as described in Section 1.1. Location information from 60 scalp electrodes was grouped into clusters based on general scalp location (see Fig. 2 for cluster configuration details). To reduce the computational load associated with our regression analysis, time-frequency data was averaged at each time sample across electrodes located within each of the 9 cluster locations.
Fig. 2.
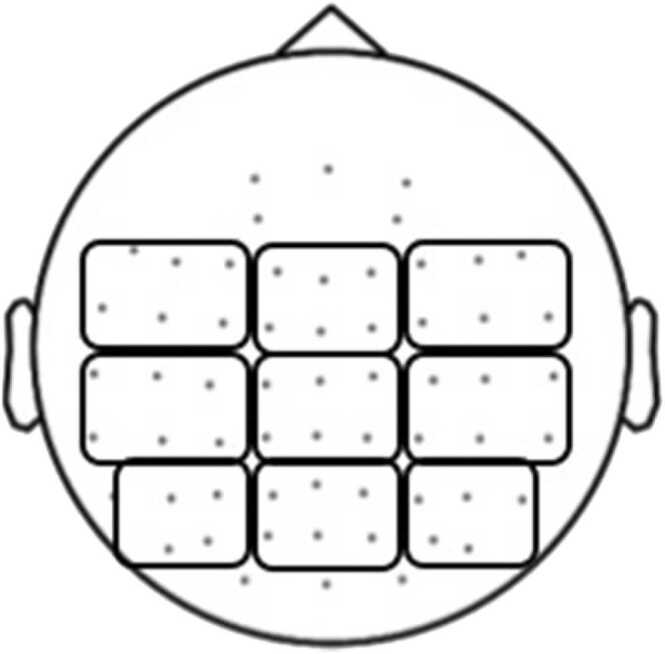
EEG data collected from 52 scalp electrodes were aggregated within 9 cluster regions for the regression-based ERSP analysis.
Random effects structures for all models were defined with random intercepts for subject, and task Item (the target pseudoword presented for each trial). GLMER model coefficients can be interpreted as the change in log-odds associated with the behavioral response for any given trial, i.e., a change in the probability that a child would have correctly identified the meaning of the pseudoword for a particular trial. Task Accuracy was coded as 0 for incorrect response and 1 for a correct response (see Section 1.3), so that a positive change in log-odds indicates an increase in the probability that a trial would have been associated with an acceptable verbal response for the meaning of the pseudoword based on the sentence contexts in the trial. All regression modeling was performed in R (RStudio Team, 2018) using the lme4 package (Bates et al., 2014). Table 1 and 2 contain descriptions of test statistics from best fit models from all analyses. To correct for multiple comparisons across cluster location and time samples, significance estimates for all parameters were corrected using the false discovery rate adjustment (Benjamini and Hochberg, 1995). To further control for false alarms and to improve the interpretability of the data, effects were only deemed significant if the models returned significant parameter values across more than 6 consecutive time points, as this would correspond to the length of one cycle within the highest frequency band measured (20 Hz; approximately 50 ms). Detailed model outputs for all main and interaction effects identified in sentences 1, 2 and 3 can be found in Supplementary Tables 1, 2, and 3, respectively.
Table 1.
Results from the GLMER analysis using ERSP activity to predict Task Accuracy at the single-trial level across each of the 3 Sentence conditions. Timing information for all effects are reported relative to pseudoword onset corresponding to 0 ms.
| GLMER analysis results | Sentence 1 | Sentence 2 | Sentence 3 |
|---|---|---|---|
| Main effect of Beta Band activity | Left Frontal: − 39–23 ms Left Central: − 625 to − 461 ms Mid Parietal: − 664 to − 555 ms |
n.s. | n.s. |
| Beta by Age interaction | n.s. | n.s. | n.s. |
| Beta by Language Ability interaction | Mid Frontal: − 547 to − 477 ms Mid Central: − 508 to − 422 ms |
Right Central: − 516 to − 461 ms | Left Frontal: − 680 to − 586 ms Right Frontal: − 453 to − 391 ms; − 39–70 ms Right Central: − 664 to − 578 ms Right Parietal: − 695 to − 586 ms |
| Main effect of Theta Band activity | Right Parietal: − 352 to − 242 ms | n.s. | n.s. |
| Theta by Age interaction | n.s. | Left Frontal: − 31–63 ms Left Central: − 39–180 ms Left Parietal: 164–234 ms |
n.s. |
| Theta by Language Ability interaction | n.s. | n.s. | n.s. |
In summary, these models were set up to achieve two goals: 1) identify neural activity related to pseudoword processing during our meaning identification task that was directly related to task performance (the effect of theta and beta band activity on Task Accuracy), and 2) test the hypothesis that individual differences related to age and language abilities uniquely influence the online activity relevant for implicit language learning (a modulatory influence of Age or Language abilities on the relationship between ERSP activity Task Accuracy).
3. Results
3.1. Performance on the Meaning Identification Task
As described above, Task Accuracy was assessed as the proportion of trials where participants verbally reported a correct response (50 trials total). Mean task accuracy across all children was generally high (Mean = 79.1%; σ = 8.9%; Range: 58–94%). A simple linear regression including Age and Language Ability as continuous independent variables predicting Task Accuracy revealed a relationship between both Age and Language Ability with Task Accuracy (Adjusted R2 = 0.46; p < 0.001). Both variables significantly contributed to variance in Task Accuracy. Age was positively associated with task performance (β = 3.5%; SE = 1.1%; p < 0.01). Our index of Language Ability, i.e., z-transformed raw scores from the CELF-5 subtests, was also positively related to task performance and accounted for unique variance in Task Accuracy (β = 4.1%; SE = 1.1%; p < 0.001). Fig. 3 provides a visualization of the relationship between Age and Task Accuracy (3 A), as well as Language ability and Task Accuracy (3B). Fig. 5.
Fig. 3.

A) Simple linear regression between Age and performance on the meaning identification task. B) Simple linear regression between Language Abilities, as indexed by raw scores on the Formulating Sentences and Recalling Sentences subtests of the CELF-5 battery, and performance on the meaning identification task. Reported test statistics used a model with z-transformed proportion of total correct trials on the meaning identification task, but raw proportions are represented in the scatterplots for clarity. Both Age and Language Abilities held a significant positive relationship with Task Accuracy.
Fig. 5.

A) Time series of regression model weights corresponding to the main effects of beta (blue) and theta (orange) band activity on task accuracy identified during Sentence 1. B) Time series of regression model weights corresponding to the interaction effects between theta band activity and age on task accuracy during Sentence 2. C) Time series of regression model weights corresponding to interaction effects between beta band activity and language abilities on task accuracy during Sentences 1 (left), 2 (middle), and 3 (right). Shaded areas surrounding the plotted lines represent the standard error for the corresponding model estimate at each time point. Significant model estimates are marked by the shaded boxes. Pseudoword onset occurred at 0 ms.
3.2. ERSP results
3.2.1. Main effects of theta and beta band activity on task accuracy
A binary representation of Task Accuracy was used as a dependent variable in two GLMER models used to assess the task relevance of beta and theta band activity across each of the three sentences comprising each trial. The reported effects from each sentence analysis were corrected for multiple comparisons (see Table 1). Results from our series of regression models revealed a main effect of beta band activity on Task Accuracy across three cluster locations in sentence one: Left frontal, Left central, and Middle parietal (see Supplementary Table 1 for model output). The main effects of beta band activity manifested prior to the onset of the pseudoword in each of the identified cluster regions (earliest onset = −664 ms; latest offset = 23 ms), i.e., during the spoken sentences before novel words were heard. These effects were associated with a negative change in log-odds related to task accuracy, indicating that greater beta band suppression prior to pseudoword onset at these scalp locations was associated with better task performance.
A main effect of theta band activity on Task Accuracy was identified only during neural activity recorded during the first sentence presentation in one cluster region: Right parietal (see Supplementary Table 1 for model output). This effect was also prior to pseudoword onset (−352 to −242 ms). The theta band effect was associated with a positive change in log-odds related to Task Accuracy, indicating that greater theta band power at right-parietal electrodes prior to pseudoword onset was associated with a higher probability of giving a correct response on the meaning identification task.
3.3. Unique influence of age on ERSP signatures supporting meaning identification
Interactions between both beta and theta band activity and age across each of the three sentences within a trial were tested as independent predictors of the binary representation of Task Accuracy. These interactions indicate that age held a unique influence on the task relevance of neural activity recorded the meaning identification task. Because age and language abilities were entered in the same regression analysis, significant interactions indicate how the brain response during our meaning identification task changed as a function of age while controlling for language ability, and vice versa.
Our analysis revealed that Age, over and above Language Abilities, influenced the way that theta band activity associated with Task Accuracy. Interactions between Age and theta band activity on Task Accuracy were identified in left hemisphere activity during sentence two, and these interactions manifested primarily during and after the onset of pseudowords during these sentences (earliest onset = −39 ms; latest offset = 234 ms; see Table 1 and Supplementary Table 2). These interactions indicate that relative theta band enhancement was negatively associated with task performance as a function of increasing age. This means that for a given trial, greater theta band power in left hemisphere channels was more likely to occur during incorrect trials, and this was especially the case for older, relative to younger children (Fig. 6). Notably, there was no influence of Age on the relationship between beta activity and Task Accuracy.
Fig. 6.

Main effects of theta and beta power on Task Accuracy. Graphs depict predicted power plotted across time as a function of Task Accuracy to visualize the relationship between ERSP activity and task performance. All main effects were identified during initial sentence in each trial (Sentence 1). A) Main effects of beta band activity on task performance were identified in middle parietal, left frontal, and left central pre-pseudoword activity during sentence 1. The GLMER analysis revealed that greater beta band suppression was associated with better task performance. B) A main effect of theta activity on Task Accuracy was identified over right parietal electrodes during sentence 1. The GLMER analysis revealed that greater theta band enhancement was associated with better task performance. Shaded regions denote the time windows that significant main effects of theta and beta activity on Task Accuracy were identified. Pseudoword onset occurred at 0 ms.
3.4. Unique influence of language abilities on ERSP signatures supporting meaning identification
Interactions between both beta and theta band activity and language abilities across each of the three sentences within a triplet were tested as independent predictors of the binary representation of Task Accuracy. These interactions indicate that language abilities held a unique influence on the task relevance of neural activity recorded the meaning identification task. As mentioned previously, because age and language abilities were entered in the same regression analysis, identified interactions indicate how the brain response during our meaning identification task changed as a function of language abilities while controlling for the influence of age.
Our analysis revealed that language abilities, over and above age, influenced the way that beta activity contributed to Task Accuracy. Interactions between language abilities and beta band activity on Task Accuracy were identified in middle frontal and middle central channels during Sentence 1, right frontal channels in Sentence 2, and in left frontal, right frontal, right central, and right parietal channels during Sentence 3 (see Fig. 4 and Supplementary Tables 1–3). In contrast to relationships between age and theta band activity, interactions between language abilities and beta activity manifested in pre-pseudoword activity. The interactions were in the same direction as the interactions identified between theta activity and Age–for a given trial, beta band power was negatively associated with task accuracy as an increasing function of language abilities. In other words, greater values of beta band activity were more likely to occur during incorrect trials particularly for children with higher language abilities, whereas higher levels of beta power were more likely to occur during correct trials for children who performed less-well on the language assessment (Fig. 7) Fig. 8.
Fig. 4.

Summary figure depicting the distribution of the task relevant theta and beta band activity identified by the GLMER analysis. Age modulated the effect of theta band activity on task performance while language abilities held a modulatory effect on the relationship between beta band activity and task performance.
Fig. 7.

A series of graphs showing predicted theta band power across correct and incorrect trials as a function of child age during the second sentence. Predicted fits were obtained in a series of LMER models that modeled theta band activity as a function of Task Accuracy. Interactions between Task Accuracy and age were included to visualize the theta band by Age interactions identified in the GLMER analysis using Task Accuracy as a binomial outcome variable. Shaded regions represent the significant time points identified in the GLMER analysis. Pseudoword onset occurred at 0 ms.
Fig. 8.

A series of graphs showing predicted beta band power across correct and incorrect trials as a function of language abilities during the first (left) and third (right) sentences. Predicted fits were obtained in a series of LMER models that modeled beta band activity as a function of Task Accuracy. Interactions between Task Accuracy and language abilities were included to visualize the beta band by Language Ability interactions identified in the GLMER analysis using Task Accuracy as a binomial outcome variable. Shaded regions represent the significant time points identified in the GLMER analysis. Pseudoword onset occurred at 0 ms.
4. Discussion
4.1. Neural correlates of meaning identification from prior context
The current study set out to investigate how dynamic oscillatory activity varies as a function of behavioral performance during a meaning identification task requiring contextual inference. Importantly, we sought to investigate how task-relevant activity in the EEG varies as a function of both age and language ability in school-age children. We found that theta band and low beta band activity were associated with performance on our meaning identification task, but that they held relatively unique relationships with age and language abilities, respectively. When analyzing our large sample (N = 82) as a whole, we found that relative beta suppression and theta band enhancement during processing of the first sentence in a trial was associated with a higher probability that the pseudoword for a particular trial would be correctly identified. While this was the case for sentence one, the way that neural activity was associated with meaning identification performance varied substantially across our cohort especially during sentences two and three in a trial: theta band activity, particularly over left hemisphere electrodes, held a relationship with task performance as a function of age during sentence 2, whereas beta band activity held a relationship with task performance as a function of language abilities during all sentences. These unique relationships between task-relevant brain activation patterns and individual differences in our child cohort also manifested during relatively different times over the course of spoken sentence processing, with age-mediated changes in theta band activity being approximated to pseudoword presentation, and language aptitude-mediated changes in beta band activity predominantly occurring during sentence processing itself, i.e., prior to pseudoword onset. We will discuss each of these relationships and their relevance for our understanding of language development in turn.
4.2. Age-mediated differences in memory activity to novel words in context
A general extrapolation that can be made from these results is that, within a group of children with typical language skills, age, over and above language ability, mediates the way that theta band engagement to novel words in context is predictive of task performance on a trial-by trial basis, i.e., whether meaning is successfully mapped to those unfamiliar words. Our data suggest that greater theta band power to pseudowords is less ideal for task performance as children get older, while it is more ideal for task performance for younger children.
In line with similar work investigating the developmental trajectory of neural oscillations during written sentence comprehension (Maguire et al., 2022) and word reading (Spironelli and Angrilli, 2010) tasks, we found age-related changes in task-relevant theta band activity near the onset of pseudowords, predominantly over left-hemisphere regions. Changes in theta band activity over left-hemisphere regions aligns with the majority of adult studies in the language processing literature (e.g., Hald et al., 2006). Typically, theta band enhancement is found in response to lexical items that are either unrelated to a preceding context, or for words that contain stronger semantic associations (open-class vs closed-class words (Bastiaansen and Hagoort, 2006, Bastiaansen et al., 2005). While our task did put working memory demands on children, the lateralization of the relationship between theta power and task accuracy suggests this activity is associated with age-related changes in semantic memory activation to pseudowords in context, rather than primarily reflecting domain-general processes associated with our task, such as changing demands on working memory processes (e.g. Fernández et al., 2021).
To further understand the relationship between child age and task-related changes in theta band activity, it is noteworthy to consider that sentences 2 and 3 provided more contextual constraint than sentence 1 in our paradigm, which would afford more precise predictions about the semantic content of upcoming speech. Because the task-related changes of theta band activity and its relationship with age manifest after sentence 1, our results point to differences across children in their sensitivity to how contextual constraints shape memory encoding or activation to novel words during language comprehension (Rommers et al., 2017). In line with this interpretation, Armeni et al. (2019) identified that oscillatory activity during spoken discourse comprehension in beta and theta bands were differentially associated with the amount of predictive affordance that the discourse context provided, and theta band increases were found in left temporoparietal and medial temporal regions when there was less certainty about which words would be heard next.
Other research investigating neural signatures of word learning from context in school-aged children found that age modulated how sentence constraint influenced the neural response to unfamiliar words in context, as well as how those same novel words could be used as semantic primes after an initial learning procedure (Vergilova et al., 2022). More specifically, children under 10 showed sensitivity to measures of constraint, as measured by N400 responses to pseudowords embedded within sentence contexts, but showed relatively weak semantic priming effects when those words were later used as primes for known, real words. Along with the current work, age, but not language abilities more generally, may act as a limiting factor not simply for the way that contextual constraint can prime the language system to receive upcoming words, but for how robustly novel words are integrated within the lexicon and interacts with language comprehension mechanisms during subsequent exposure to recently learned words. While these previous findings support an interpretation that age influences how novel words are processed as a function of contextual predictability, more research will be necessary to identify why these effects manifested during sentence 2, but not sentence 3 in our study.
An alternative, but possibly compatible, interpretation is that theta band fluctuations in response to pseudowords signify changes in semantic memory activation as a result of competition between simultaneously activated items in semantic memory that the sentence context could have primed for (Strauß et al., 2014). For older children, increased activation of an alternative word meaning that fits, for example, the context in sentence 2, may lead to inaccurate meaning estimation at the end of trials, which required a more global consideration about how the pseudoword could feasibly relate to all three sentence contexts. Although we did not run follow up tests within pre-defined groups of younger or older participants, visual inspection of the age by theta band interaction suggests that theta band activity may have been less associated with task performance in younger children, as the difference between successful and unsuccessful trials appears less pronounced in younger children according to the model-fitted data (see the left parietal cluster in Fig. 6). Further research is necessary to identify whether the relationship between left hemisphere theta band activity and task performance is non-linear—which would suggest they reflect mechanisms that emerge as a function of age.
4.3. Language skills relate to the efficacy of online processing abilities during sentence comprehension
The current data indicate that successful pseudoword identification was also associated with differences in beta power across successful versus unsuccessful trials, such that children with higher language scores showed lower beta power on successful relative to unsuccessful trials. Likewise, children with lower language scores were more likely to show greater beta power on unsuccessful trials compared to successful trials. Notably, and unlike the effects identified in the theta band, the interaction between task performance and language abilities were identified prior to pseudoword onset, i.e., during the spoken sentence context (see Fig. 7). In addition, whereas the age-related changes in theta band activity arose during sentence 2 during our meaning identification task, relationships between language ability and beta band activity were identified in all three sentences across trials (see Fig. 4).
In the context of language processing, beta band activity is hypothesized to reflect predictive coding dynamics relevant for semantic or syntactic information during sentence comprehension. In particular, increases in beta are thought to reflect the active maintenance of linguistic information in memory as the sentence-level meaning is constructed (Lewis and Bastiaansen, 2015). In this scheme, beta enhancement occurs when information in the sentence aligns with what is predicted to occur next, while beta suppression occurs when input does not align with prior expectations about upcoming words (Lewis and Bastiaansen, 2015, Lewis et al., 2016). In conjunction with the time course of these interactions (prior to pseudoword onset), these results suggest that the way language abilities shape behavioral performance on our task is less related to how sentence constraint influences language processing or how children process novel words per se, and instead may be more related to speech processing and sentence comprehension more generally.
Visual inspection of hemispheric differences in the beta band analysis during sentence 3 suggests that task-related differences in left hemispheric beta band activity were more pronounced in children with lower language scores, while those with higher language scores showed more pronounced task-related changes over right hemisphere sites, especially over frontal and parietal regions (see Fig. 7). This trend may indicate qualitative differences in task-relevant brain activity as a function of language abilities. Extant research has reported hemispheric asymmetries in beta band activity during language comprehension tasks in children and adults (e.g., Spironelli and Angrilli, 2010). Notably, beta activity during a semantic processing task was right lateralized over anterior electrodes in a typically developing control group compared with children with dyslexia (Spironelli et al., 2008). The controls in that study showed more lateralized beta band effects compared to children with language processing deficits, and this is notable considering the current data shows that stronger language abilities associated with more pronounced differences in right hemispheric beta band activity during sentence processing as a function of behavioral performance. Future work should explicitly investigate the hemispheric asymmetries in oscillatory activity underlying speech processing during development to elucidate their potential functional significance.
5. Limitations
To minimize the risk of overfitting in our GLMER analysis, we compared the fit our omnibus model with an intercept-only model as well as a model including main effects of theta and beta band activity without the addition of interactions with Age or Language Ability. Model comparison was based on AIC values for each model averaged across timesteps for each cluster location, which provide an estimate of model performance sensitive to the tradeoff between fit and model complexity (see Supplementary Tables 4–6). While this analysis suggested that omnibus models provided better or similar fits of the data compared to the simpler, nested models, without further cross-validation procedures we cannot quantify the generalizability of our models to new data.
6. Conclusions
These results are important as they begin to shed light on the way that different developmental variables uniquely shape the neural correlates underlying language processing and learning. Importantly, irrespective of particular interpretations about the directionality of the interaction effects, the current study identifies a novel distinction in the way that task-relevant brain activity scales uniquely with age and with language abilities in school-aged children, such that age held a relationship with theta band dynamics, and language abilities held a relationship with beta band dynamics. Our regression-based analysis allowed us to identify these relationships while controlling for any confounding influence of the alternative individual difference measure. As both older children and children with better language abilities were more likely to perform well on the meaning identification task, the mechanistic differences observed in the EEG data were relevant for task performance overall despite the fact that each was associated with different developmental factors across our cohort of children. Our data support the notion that while language abilities may confer certain benefits for language processing and learning, such as online predictive processing abilities during speech processing, but may be limited in terms of supporting controlled memory-retrieval processes that depend on how sentences constrain the space of possible meanings for upcoming words. Learning how different variables during development associate with online language processing and learning mechanisms is vital for understanding how language development unfolds in relation to both language-specific aptitudes and biological or maturational constraints.
Funding
This work was funded in part by National Institute for Deafness and Other Communication Disorders (NIH-NIDCD 1R21DC018865) to A. A.
CRediT authorship contribution statement
Jacob Momsen: Conceptualization, Software, Formal analysis, Data curation, Writing – original draft, Visualization. Julie Schneider: Conceptualization, Methodology, Software, Formal analysis, Writing – review & editing. Alyson Abel: Conceptualization, Methodology, Investigation, Resources, Data Curation, Writing – review & editing, Supervision, Project administration, Funding acquisition.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Footnotes
Supplementary data associated with this article can be found in the online version at doi:10.1016/j.dcn.2022.101185.
Appendix A. Supplementary material
Supplementary material
.
Supplementary material
.
Supplementary material
.
Supplementary material
.
Data Availability
Data will be made available on request.
References
- Abel A.D., Schneider J., Maguire M.J. N400 response indexes word learning from linguistic context in children. Lang. Learn. Dev. 2018;14(1):61–71. [Google Scholar]
- Arain M., Haque M., Johal L., Mathur P., Nel W., Rais A., Sandhu R., Sharma S. Maturation of the adolescent brain. Neuropsychiatr. Dis. Treat. 2013;9:449–461. doi: 10.2147/NDT.S39776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Armeni K., Willems R.M., Van den Bosch A., Schoffelen J.M. Frequency-specific brain dynamics related to prediction during language comprehension. NeuroImage. 2019;198:283–295. doi: 10.1016/j.neuroimage.2019.04.083. [DOI] [PubMed] [Google Scholar]
- Aurnhammer C., Delogu F., Schulz M., Brouwer H., Crocker M.W. Retrieval (N400) and integration (P600) in expectation-based comprehension. PLOS ONE. 2021;16(9) doi: 10.1371/journal.pone.0257430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baayen R.H., Davidson D.J., Bates D.M. Mixed-effects modeling with crossed random effects for subjects and items. J. Mem. Lang. 2008;59(4):390–412. [Google Scholar]
- Bakker I., Takashima A., van Hell J.G., Janzen G., McQueen J.M. Changes in theta and beta oscillations as signatures of novel word consolidation. J. Cogn. Neurosci. 2015;27(7):1286–1297. doi: 10.1162/jocn_a_00801. [DOI] [PubMed] [Google Scholar]
- Bastiaansen M.C., Hagoort P. In: Oscillatory neuronal dynamics during language comprehension. Neuper C., Klimesch W., editors. Elsevier; 2006. pp. 179–196. (Event-related dynamics of brain oscillations) (Event-related dynamics of brain oscillations) [DOI] [PubMed] [Google Scholar]
- Bastiaansen M.C., Linden M.V., Keurs M.T., Dijkstra T., Hagoort P. Theta responses are involved in lexicalsemantic retrieval during language processing. J. Cogn. Neurosci. 2005;17(3):530–541. doi: 10.1162/0898929053279469. [DOI] [PubMed] [Google Scholar]
- Bates, D., Maechler, M., Bolker, B., & Walker, S. (2014). lme4: Linear mixed-effects models using Eigen and S4. R package version 1.1–7.
- Bell A.J., Sejnowski T.J. An information-maximization approach to blind separation and blind deconvolution. Neural Comput. 1995;7(6):1129–1159. doi: 10.1162/neco.1995.7.6.1129. [DOI] [PubMed] [Google Scholar]
- Benjamini Y., Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995;57(1):289–300. [Google Scholar]
- Borovsky A., Kutas M., Elman J. Learning to use words: Event-related potentials index single-shot contextual word learning. Cognition. 2010;116(2):289–296. doi: 10.1016/j.cognition.2010.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borovsky A., Elman J.L., Kutas M. Once is enough: N400 indexes semantic integration of novel word meanings from a single exposure in context. Lang. Learn. Dev. 2012;8(3):278–302. doi: 10.1080/15475441.2011.614893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buzsáki G., Draguhn A. Neuronal oscillations in cortical networks. Science. 2004;304(5679):1926–1929. doi: 10.1126/science.1099745. [DOI] [PubMed] [Google Scholar]
- Delorme A., Makeig S. EEGLAB: an open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods. 2004;134(1):9–21. doi: 10.1016/j.jneumeth.2003.10.009. [DOI] [PubMed] [Google Scholar]
- Fenson, L., Marchman, V.A., Thal, D.J., Dale, P.S., Reznick, J.S., & Bates, E. (2006). The MacArthur-Bates communicative development inventories user’s guide and technical manual (2nd ed.). Brookes.
- Fukkink R.G. Deriving word meaning from written context: a process analysis. Learn. Instr. 2005;15(1):23–43. [Google Scholar]
- Hagemann D., Hewig J., Walter C., Naumann E. Skull thickness and magnitude of EEG alpha activity. Clin. Neurophysiol. 2008;119(6):1271–1280. doi: 10.1016/j.clinph.2008.02.010. [DOI] [PubMed] [Google Scholar]
- Hald L.A., Bastiaansen M.C., Hagoort P. EEG theta and gamma responses to semantic violations in online sentence processing. Brain Lang. 2006;96(1):90–105. doi: 10.1016/j.bandl.2005.06.007. [DOI] [PubMed] [Google Scholar]
- Hall, W.S., Nagy, W.E., & Linn, R.L. (1980). Spoken words. Center for the Study of Reading Technical Report; no. 177.
- Horst J.S. Context and repetition in word learning. Front. Psychol. 2013;4:149. doi: 10.3389/fpsyg.2013.00149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janacsek K., Fiser J., Nemeth D. The best time to acquire new skills: age‐related differences in implicit sequence learning across the human lifespan. Dev. Sci. 2012;15(4):496–505. doi: 10.1111/j.1467-7687.2012.01150.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lam N.H., Schoffelen J.M., Uddén J., Hultén A., Hagoort P. Neural activity during sentence processing as reflected in theta, alpha, beta, and gamma oscillations. Neuroimage. 2016;142:43–54. doi: 10.1016/j.neuroimage.2016.03.007. [DOI] [PubMed] [Google Scholar]
- Lewis A.G., Bastiaansen M. A predictive coding framework for rapid neural dynamics during sentence-level language comprehension. Cortex. 2015;68:155–168. doi: 10.1016/j.cortex.2015.02.014. [DOI] [PubMed] [Google Scholar]
- Lewis A.G., Schoffelen J.M., Schriefers H., Bastiaansen M. A predictive coding perspective on beta oscillations during sentence-level language comprehension. Front. Hum. Neurosci. 2016;10 doi: 10.3389/fnhum.2016.00085. Article 85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lichtman K.M. University of Illinois at Urbana-Champaign,; 2012. Child-adult Differences in Implicit and Explicit Second Language Learning. [Google Scholar]
- Maguire M.J., Schneider J.M. Socioeconomic status related differences in resting state EEG activity correspond to differences in vocabulary and working memory in grade school. Brain Cogn. 2019;103619 doi: 10.1016/j.bandc.2019.103619. [DOI] [PubMed] [Google Scholar]
- Maguire M.J., Schneider J.M., Middleton A.E., Ralph Y., Lopez M., Ackerman R.A., Abel A.D. Vocabulary knowledge mediates the link between socioeconomic status and word learning in grade school. J. Exp. Child Psychol. 2018;166:679–695. doi: 10.1016/j.jecp.2017.10.003. [DOI] [PubMed] [Google Scholar]
- Maguire M.J., Schneider J.M., Melamed T.C., Ralph Y.K., Poudel S., Raval V.M., Mikhail D., Abel A.D. Temporal and topographical changes in theta power between middle childhood and adolescence during sentence comprehension. Dev. Cogn. Neurosci. 2022;53 doi: 10.1016/j.dcn.2021.101056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maris E., Oostenveld R. Nonparametric statistical testing of EEG-and MEG- data. J. Neurosci. Methods. 2007;164(1):177–190. doi: 10.1016/j.jneumeth.2007.03.024. [DOI] [PubMed] [Google Scholar]
- Marshall P.J., Fox N.A., Group B.C. A comparison of the electroencephalogram between institutionalized and community children in Romania. J. Cogn. Neurosci. 2004;16(8):1327–1338. doi: 10.1162/0898929042304723. [DOI] [PubMed] [Google Scholar]
- Mestres-Missé A., Rodriguez-Fornells A., Münte T.F. Watching the brain during meaning acquisition. Cereb. Cortex. 2007;17(8):1858–1866. doi: 10.1093/cercor/bhl094. [DOI] [PubMed] [Google Scholar]
- Momsen J.P., Abel A.D. Neural oscillations reflect meaning identification for novel words in context. Neurobiol. Lang. 2022;3(1):132–148. doi: 10.1162/nol_a_00052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Myslobodsky M.S., Coppola R., Bar-Ziv J., Karson C., Daniel D., Praag H.V., Weinberger D.R. EEG asymmetries may be affected by cranial and brain parenchymal asymmetries. Brain Topogr. 1989;1(4):221–228. doi: 10.1007/BF01129599. [DOI] [PubMed] [Google Scholar]
- Nagy W.E., Herman P.A., Anderson R.C. Learning words from context. Read. Res. Q. 1985:233–253. [Google Scholar]
- Nagy W.E., Anderson R.C., Herman P.A. Learning word meanings from context during normal reading. Am. Educ. Res. J. 1987;24(2):237–270. [Google Scholar]
- Nemeth D., Janacsek K., Fiser J. Age-dependent and coordinated shift in performance between implicit and explicit skill learning. Front. Comput. Neurosci. 2013;7:147. doi: 10.3389/fncom.2013.00147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oostenveld R., Fries P., Maris E., Schoffelen J.M. FieldTrip: open source software for advanced analysis of MEG, EEG, and invasive electrophysiological data. Comput. Intell. Neurosci. 2011;2011 doi: 10.1155/2011/156869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ralph Y.K., Schneider J.M., Abel A.D., Maguire M.J. Using the N400 event- related potential to study word learning from context in children from low-and higher- socioeconomic status homes. J. Exp. Child Psychol. 2020;191 doi: 10.1016/j.jecp.2019.104758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rommers J., Dickson D.S., Norton J.J., Wlotko E.W., Federmeier K.D. Alpha and theta band dynamics related to sentential constraint and word expectancy. Lang. Cogn. Neurosci. 2017;32(5):576–589. doi: 10.1080/23273798.2016.1183799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider J.M., Maguire M.J. Developmental differences in the neural correlates supporting semantics and syntax during sentence processing. Dev. Sci. 2019;22(4) doi: 10.1111/desc.12782. [DOI] [PubMed] [Google Scholar]
- Schneider J.M., Abel A.D., Ogiela D.A., Middleton A.E., Maguire M.J. Developmental differences in beta and theta power during sentence processing. Dev. Cogn. Neurosci. 2016;19:19–30. doi: 10.1016/j.dcn.2016.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider J.M., Abel A.D., Ogiela D.A., McCord C., Maguire M.J. Developmental differences in the neural oscillations underlying auditory sentence processing in children and adults. Brain Lang. 2018;186:17–25. doi: 10.1016/j.bandl.2018.09.002. [DOI] [PubMed] [Google Scholar]
- Schneider J.M., Abel A.D., Momsen J., Melamed T.C., Maguire M.J. Neural oscillations reveal differences in the process of word learning among school-aged children from lower socioeconomic status backgrounds. Neurobiol. Lang. 2021;2(3):372–388. doi: 10.1162/nol_a_00040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith N.J., Kutas M. Regression‐ based estimation of ERP waveforms: I. The rERP framework. Psychophysiology. 2015;52(2):157–168. doi: 10.1111/psyp.12317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spironelli C., Angrilli A. Developmental aspects of language lateralization in delta, theta, alpha and beta EEG bands. Biol. Psychol. 2010;85(2):258–267. doi: 10.1016/j.biopsycho.2010.07.011. [DOI] [PubMed] [Google Scholar]
- Spironelli C., Penolazzi B., Angrilli A. Dysfunctional hemispheric asymmetry of theta and beta EEG activity during linguistic tasks in developmental dyslexia. Biol. Psychol. 2008;77(2):123–131. doi: 10.1016/j.biopsycho.2007.09.009. [DOI] [PubMed] [Google Scholar]
- Storkel H.L. A corpus of consonant–vowel–consonant real words and nonwords: Comparison of phonotactic probability, neighborhood density, and consonant age of acquisition. Behav. Res. Methods. 2013;45:1159–1167. doi: 10.3758/s13428-012-0309-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strauß A., Kotz S.A., Scharinger M., Obleser J. Alpha and theta brain oscillations index dissociable processes in spoken word recognition. NeuroImage. 2014;97:387–395. doi: 10.1016/j.neuroimage.2014.04.005. [DOI] [PubMed] [Google Scholar]
- Uhlhaas P.J., Roux F., Rodriguez E., Rotarska-Jagiela A., Singer W. Neural synchrony and the development of cortical networks. Trends Cogn. Sci. 2010;14(2):72–80. doi: 10.1016/j.tics.2009.12.002. [DOI] [PubMed] [Google Scholar]
- Urbach T.P., DeLong K.A., Chan W.H., Kutas M. An exploratory data analysis of word form prediction during word-by-word reading. Proc. Natl. Acad. Sci. 2020;117(34):20483–20494. doi: 10.1073/pnas.1922028117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vergilova Y., Jachmann T.K., Mani N., Kray J. Age‐related differences in expectation‐based novel word learning. Psychophysiology. 2022 doi: 10.1111/psyp.14030. [DOI] [PubMed] [Google Scholar]
- Wiig E.H., Semel E., Secord W.A. NCS Pearson; Bloomington, MN: 2013. Clinical Evaluation of Language Fundamentals– Fifth Edition (CELF-5) [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary material
Supplementary material
Supplementary material
Supplementary material
Data Availability Statement
Data will be made available on request.