

Abstract
Ligand targeted therapy (LTT) is a precision medicine strategy that can selectively target diseased cells while minimizing off‐target effects on healthy cells. Integrin‐targeted LTT has been developed recently for angiogenesis‐related diseases. However, the clinical success is based on the optimal design of the nanoparticles for inducing receptor clustering within the cell membrane. The current study focused on determining the surface density of Ser‐Asp‐Val containing anti‐integrin heptapeptide on poly (ethylene glycol)‐b‐poly(propylene sulfide) micelles (MC) required for anti‐angiogenic effects on HUVECs. Varying peptide density on PEG‐b‐PPS/Pep‐PA MCs (Pep‐PA‐Peptide‐palmitoleic acid) was used in comparison to a random peptide (SGV) and cRGD (cyclic‐Arginine‐Glycine‐Aspartic acid) construct at 5%‐density on MCs. Immunocytochemistry using CD51/CD31 antibody was performed to study the integrin blocking by MCs. In addition, the expression of VWF and PECAM‐1, cell migration and tube formation was evaluated in the presence of PEG‐b‐PPS/Pep‐PA MCs. The results show PEG‐b‐PPS/SDV‐PA MCs with 5%‐peptide density to achieve significantly higher αvβ3 blocking compared to random peptide as well as cRGD. In addition, αvβ3 blocking via MCs further reduced the expression of vWF and PECAM‐1 angiogenesis protein expression in HUVECs. Although a significant level of integrin blocking was observed for 1%‐peptide density on MCs, the cell migration and tube formation were not significantly affected. In conclusion, the results of this study demonstrate that the peptide surface density on PEG‐b‐PPS/Pep‐PA MCs has a significant impact in integrin blocking as well as inhibiting angiogenesis during LTT. The outcomes of this study provides insight into the design of ligand targeted nanocarriers for various disease conditions.
Keywords: anti‐angiogenic peptide, LTT, micelles, PEG‐b‐PPS, αvβ3
1. INTRODUCTION
Ligand targeted therapy (LTT) is a promising strategy to maximize the efficacy and safety with little affinity toward normal cells. 1 , 2 Several LTT approaches have been developed using antibodies, antibody fragments, and peptides that can target different receptors based on the application. A considerable number of investigations have focused on integrin‐targeted therapeutics, demonstrating applications in anti‐angiogenic treatments for cancer, Crohn's disease and retinal disease. 3 , 4 , 5 , 6 , 7 , 8 , 9 , 10 Integrins are a class of transmembrane receptors and integrin‐mediated interactions are key contributors to angiogenesis. αvβ3 integrins are one of the most prominent classes of integrins involved in this process. 11 , 12 , 13 , 14 Previous investigations demonstrated that angiogenesis could be inhibited using anti‐integrin therapeutic peptides or integrin agonists. 15 , 16 , 17 A PDZ binding motif on an anti‐angiogenic peptide can be used to bind to αvβ3 integrins and thereby inhibit angiogenesis. 18 Since these peptides require large concentrations to elicit the desired response, a nanoscale delivery system that presents a high valency of these peptides on its surface can be used to bring about integrin clustering, and in turn elicit stronger intracellular signaling at low concentrations. 19 , 20 Poly(ethylene glycol)‐b‐poly(propylene sulfide) (PEG‐b‐PPS) micelles (MCs) have been used for a wide range of LTT strategies, including targeted intracellular delivery of latrunculin A for treating glaucoma as well as vitamin D in the treatment of vascular inflammation. 21 , 22 In studies conducted by Kumar et al, antagonists EMD4787 23 and SCH 221153 24 were found to inhibit angiogenesis successfully by binding to αvβ3 integrins using the chick chorioallantoic membrane (CAM) assay and rat models, respectively.
LTT provided promising opportunities to improve the selective treatment strategy and hence over 100 LTTs are currently under clinical trial. 25 αvβ3 based LTT demonstrated an advantage over non‐targeted systems in the tumor. 26 , 27 In addition, novel high‐affinity targeting moieties have been identified through phage‐display techniques which is crucial in LTT design. Generally, any receptor with threefold higher expression in diseased cells can be ideal for LTT. 28 In the case of αvβ3, the overexpression was significantly higher in abnormal angiogenesis. 29 However, a major design constraint in LTT is the stable and free presentation of ligands on nanomaterials, especially in the case of small peptide ligands. Ligand–receptor interactions at the bio/nanointerface between nanomaterials and cells depend on several factors, and the density of ligand molecules on the surface of the nanomaterials has been found to be particularly critical. 1 , 22 , 30 The peptide density should be specified to achieve optimal integrin clustering to block focal adhesion formation and subsequent cell adhesion and migration. Previous studies show that the optimal lateral spacing of the integrin ligands is approximately 60–70 nm. 31 It is necessary to know the optimum density of the peptides for triggering cellular responses. 1 , 12 , 13 , 14 , 32 The nature of the surface, like the hydrophobicity, size, charge, and surface chemistry responsible for exerting steric and electro‐steric effects together determine which biomolecules form the protein corona in blood and mediate subsequent cellular interactions. 33 , 34 , 35 As a result, receptor‐mediated endocytosis occurs. For ligand‐mediated uptake to occur, the membrane receptors must diffuse to the adhesion site to form a critical number of interactions leading to clathrin‐coated pit formation. 36
In a previous investigation, we used a Ser‐Asp‐Val (SDV) containing heptapeptide as an antagonist to αvβ3 integrins, which showed efficient anti‐angiogenesis when employing PEG‐b‐PPS MCs as a vehicle for peptide delivery compared to heptapeptide alone. 37 In addition, it was observed that the amount of peptide required to inhibit angiogenesis was significantly reduced when displayed on MCs compared to peptide alone, suggesting that integrin clustering plays a role in LTT. 37 In this study, we aim to address the role of varying SDV peptide density on PEG‐b‐PPS MCs on their anti‐angiogenic activity. We varied the ratio of peptide to polymer during MC assembly without affecting the MC structural integrity to develop an optimum formulation for an enhanced anti‐angiogenic effect.
2. MATERIALS AND METHODS
2.1. Synthesis of peptide constructs
Custom‐made crude F‐moc protected peptides with embedded resin were purchased from Peptide 2.0. They are GHSDVHK, GHSGVHK (random peptide, by changing one amino acid in the active site), and cRGD (which has known anti‐angiogenic activity). Standard solid‐phase peptide synthesis techniques were used for the modification of the peptides. A PEG spacer Fmoc‐N‐amido‐dPEG was used to conjugate the peptide constructs to palmitoleic acid (Pep‐PA). Ninhydrin tests were carried out at each step to confirm the addition and deprotection of new residues. A solution of 95% Trifluoroacetic acid (TFA), 2.5% Thiosalicylic acid, and 2.5% ddH2O with a few drops of HCl was prepared and 10 ml of this was used to remove the resin from the SDV peptides. This was allowed to shake for 4 h, and the removal of resin was confirmed by LC–MS and by the solution changing color to yellow. To completely remove the resin, the modified peptide was washed 5 times with NMP (1‐methyl‐2‐pyrrolidinone). A rotary evaporator was used to evaporate the solvent, and ice‐cold diethyl ether was used to precipitate the modified peptide. After precipitation, mass spectrometry (MS) was carried out to confirm the stability and resin cleavage of the modified peptide. The cleaved peptide was lyophilized and stored at 4°C. RP‐HPLC (Waters 2545, Milford, MA, USA) was carried out to purify the cleaved peptide by adding 10 ml of 0.1% TFA to the modified peptide under isocratic mode with 2% TFA and 0.1% acetonitrile (Sigma, USA Cat No. 75‐05‐8) as mobile phase. At retention times of 42 and 45 min, the samples were obtained. This was followed by performing MS, which confirmed the presence of peptides of the expected molecular weight. Lyophilization was carried out on all the peptide samples, which were then stored at −20°C. To check the purity and the yield of the peptides, LC–MS was carried out.
2.2. Synthesis of PEG‐b‐PPS MCs
PEG‐b‐PPS MCs were synthesized according to a previously published protocol. 37 Briefly, PEG thioacetate was deprotected by sodium methoxide and used to initiate anionic ring‐opening polymerization of PPS. The co‐solvent evaporation method was used for the self‐assembly of PEG‐b‐PPS MCs. 20 mg of the polymer was dissolved in 1 ml of dichloromethane. This solution was added to 1 ml of PBS, which was stirred vigorously. Once the solvent completely evaporated, the resulting solution was transferred to sterile 2 ml Eppendorf tubes and stored at 4°C for further analysis.
2.3. Formulation of PEG‐b‐PPS‐anti‐αvβ3 peptides
Different densities of peptides on PEG‐b‐PPS MCs were prepared by varying the molar ratio of peptides to the polymer. To prepare SDV‐1%, SDV‐3%, and SDV‐5% PEG‐b‐PPS/ Pep‐PA MCs, the following formula was used:
The required amount of peptide construct was dissolved in 10 μl of DMSO, which was added to a phosphate‐buffered saline (PBS) solution of PEG‐b‐PPS MCs and mixed overnight in an end‐end shaker at room temperature. SGV‐5% and cRGD‐5% peptide‐bound MCs were prepared as controls. The PEG‐b‐PPS/Pep‐PA MCs were purified by Sephadex gravity chromatography where PBS was used as the mobile phase. The resulting fractions were concentrated by centrifugation in Millipore Sigma centrifugal filter units for 20 min at 4000 rpm at 20°C.
2.4. Characterization of PEG‐b‐PPS/Pep‐PA MCs
The average particle size of the MCs was determined using Dynamic Light Scattering (Malvern Instruments, Nano ZS90). The samples were diluted at ratios of 1:1000 with PBS prior to analysis. Cryogenic‐Transmission Electron Microscopy (Cryo‐TEM) was used to visualize the morphology of the MCs by using JEOL TEM‐1230 Electron Microscope, Peabody, MA, USA. To understand the difference in peptide density of each MC preparation, zeta potential was evaluated. The PEG‐b‐PPS/Pep‐PA MC and PEG‐b‐PPS MCs (termed blank MCs) were diluted in the ratio of 1:1000 with PBS. This solution was run in the (Malvern Instruments), Nano ZS90.
2.5. Cell culture
HUVEC—Human Umbilical Vein Endothelial cell line (Sigma, USA) was used to assess the anti‐angiogenic effect of the peptide display. The cell line was cultured in Endothelial Cell Growth Medium (Cell solutions, USA Cat No. 211‐500) with 1% Pen‐Strep Antibiotic (Gibco, USA Cat No. 15140122) and sub‐cultured at 80%–90% confluency by first washing the cells with Hank's Balanced Basic Salt Solution (HBSS, Sigma, USA Cat No. H6648) followed by trypsinization by Trypsin–EDTA (Sigma, USA Cat No. T3924) and neutralization with Trypsin inhibitor (Sigma, USA Cat No. T6414). The cell line was maintained at 37°C with 5% CO2 with three passages every week. The cells used in all the experiments were between passage No. 3–6 and the cell culture plates used for all the experiments were coated with denatured collagen coating solution (Sigma, USA Cat No. 125‐50).
2.6. Integrin αvβ3 expression
About 50,000 HUVECs/wells were seeded onto coverslips coated with the collagen coating solution. Once the coverslips were about 85% confluent, the cells were incubated with the MCs at 100 μg/ml for 4 h at 37°C and 5% CO2. After incubation, the cells were then fixed with 1% paraformaldehyde (PFA) in PBS for 5 min and then permeabilized with 0.1% Triton‐X 100 in PBS for 5 min. The coverslips were then washed with ice‐cold PBS for 2× and blocked with 1% bovine serum albumin (BSA) in PBS for 1.5 h. Anti‐Human CD51/CD61 (Integrin αvβ3) was diluted 1:1000 dilution in 1% BSA and incubated for 30 min in the dark. After incubation, the cells were washed 2× with PBS followed by mounting of the coverslip on the slide using fluoromount and imaged using confocal microscopy.
2.7. Expression of angiogenic markers
To determine the angiogenic properties of the HUVECs immunostaining was performed to detect two angiogenic markers vWF (von Willebrand Factor) and Platelet Endothelial Cell Adhesion Molecule‐1 (PECAM‐1). Briefly, 50,000 cells/well were seeded onto denatured collagen‐coated coverslips and treated with 20 ng/ml of bFGF, and allowed to become confluent. Then the cells were incubated with the MCs at 100 μg/ml, 24 h at 37°C, and 5% CO2. The cells were then fixed with 1% paraformaldehyde (PFA) in PBS for 5 minutes and then permeabilized with 0.1% Triton‐X 100 in PBS for 5 min. The coverslips were then washed with ice‐cold PBS for 2X and blocked with 1% BSA in PBS for 1.5 h. Primary antibodies rabbit anti‐human vWF (Sigma USA Cat No. F3520) and mouse anti‐human CD31/PECAM‐1 (Novus Biologicals, USA Cat No. AF3628) were diluted 1:1000 dilution in 1% BSA and incubated for 1 h. Following incubation with primary antibody, the wells were washed 2× with 0.05% Tween‐20 in PBS. Secondary antibodies rabbit anti‐mouse AlexaFluor 594 (Invitrogen, USA Cat No. A27027) and goat anti‐rabbit AlexaFluor 488 (Invitrogen, USA Cat No. A‐21222) of 1:5000 dilution in 1% BSA was added and incubated for 1 h in the dark. After incubation, the cells were washed 2× with 0.05% Tween‐20 in PBS followed by mounting of the coverslip on the slide using fluoromount and imaged using confocal microscopy.
2.8. Migration by wound‐healing assay
Briefly, 50,000 cells/well were seeded in a 24‐well plate coated with the denatured collagen coating solution. The plate was then incubated at 37°C, 5% CO2, until confluent. Once the cells reached 100% confluency, a scratch was made using a 200 μl pipette and then treated with MCs. The plate was allowed to incubate for 24 and 48 h. Images of the scratch were taken at 0 and 24 h. They were analyzed and quantified with ImageJ and the Angiogenesis Analyzer plugin.
2.9. In vitro endothelial cell tube formation assay
In this experiment, a 24‐well plate was coated with denatured collagen and incubated at 37°C and 5% CO2 for 1 h in order to form a gel. About 12,000 cells/well of HUVEC's mixed with MCs were seeded onto the coated plates and incubated for 4 h at 37°C. Blank PEG‐b‐PPS MCs and HUVECs without any treatment were used as control. The tubes formed were observed under the inverted microscope. Angiogenesis analyzer plugin ImageJ was used to analyze and quantify the length of the tubes formed in each well. The tubes were quantified using tree structure detection method. Then the binary skeleton was done by an automatic analysis of branches and segments. The values less than 10 pixels were removed. Total length of branches and segments were quantified.
2.10. Statistics
Data obtained was performed in triplicates in three independent experiments. Student's t‐test and one‐way ANOVA with Tukey's multiple comparisons (GraphPad Prism) were used to compare the data for statistical significance among the experimental groups, and p < .05 was considered statistically significant. All the values are presented as mean ± standard deviation (mean ± SD).
3. RESULTS
3.1. PEG‐b‐PPS/Pep‐PA MCs
LC–MS was performed to determine the purity of the peptide constructs. This can significantly improve the analytical assessment of biological molecules. It is highly selective and can differentiate compounds depending on their mass to charge ratio (m/z). Based on the molecular weight of the required product, the peaks obtained on the extracted ion chromatograms (EICs) in the presence of the product can be determined. The purity of the sample is determined by comparing the sum of the area under the curve (AUC) of known compound peaks in total ion chromatogram (TICs) to the sum of all other peaks. It was calculated as shown below.
The LC–MS results show 1350, 1291, and 1174 g/mol respectively for SDV, SGV, and cRGD peptide constructs (Figure 1A) with 99% purity. The peaks obtained in the chromatograms correlate with the theoretical molecular weight of the SDV, cRGD, and the SGV peptide constructs. The modification of peptides with palmitoleic acid facilitates a hydrophobic moiety and thus inserts itself into the PEG‐b‐PPS MCs formed in the aqueous solution and interacts with the thiol group of PEG, leading to conjugation (Figure 1B). 38
FIGURE 1.

Peg‐b‐PPS/ Pep‐PA MCs synthesis and characterization. (A) The LCMS spectrum of SDV heptapeptide construct, SGV heptapeptide construct, and cRGD peptide construct indicates the 99% purity of the modified peptide constructs. (B) Schematic representation of synthesis of PEG‐b‐PPS/Pep‐PA MCs with varying peptide surface density. (C) DLS spectrum for (i) blank‐MC (ii) PEG‐b‐PPS/SDV‐PA MCs, (iii) PEG‐b‐PPS/SGV‐PA MCs, (iv) PEG‐b‐PPS/cRGD‐PA MCs. (D) Morphology of the (i) blank‐MC and (ii) PEG‐b‐PPS/ Pep‐PA MCs MCs by CryoTEM analysis. (E) Graph showing the zeta potential values of the PEG‐b‐PPS/Pep‐PA MCs with different densities on the MCs (SDV‐1 SDV‐3, SDV‐5 SGV‐5, and cRGD‐5 represents a different percentage [1%, 3% and 5%] of respective peptides on the MCs)
Dynamic light scattering was performed to determine the average particle size of the PEG‐b‐PPS/Pep‐PA MCs and the blank MCs (Figure 1C). In the case of blank MCs, about 77% were in the range of 20–50 nm with an average particle size of 33.6 ± 5 nm (PDI = 0.25). For SDV‐MC about 89.4% were in the range of 100 nm with an average size of 41.24 ± 7.6 nm (PDI = 0.215). For SGV MCs 89% of them were in the size range of 50–100 nm with an average size of 61.7 ± 5.8 nm (PDI = 0.27). In the case of cRGD MCs, 98% of the MCs were in the size range of 30–70 nm with an average size of 42.25 ± 6.9 nm (PDI = 0.189). Cryo‐TEM is used to visualize the morphology of the MCs at cryogenic temperatures of −160 to −180°C. In this technique, the samples can be visualized in their native form without any fixation, and it gives a high resolution of up to 1.8 Å. Figure 1D confirmed the structure of PEG‐b‐PPS/Pep‐PA MCs, as well as that of the blank MCs, to be spherical with a particle size between 20 and 80 nm in correlation to the DLS spectrum obtained.
The Zeta potential confirmed the surface density of the peptides on the micellar surface. The net charge of the peptide at pH 7.4 is 0.18 with 0.54 average hydrophilicity. The surface charge of blank MCs was −6.7 ± 0.00003 mv. For 1% SDV containing PEG‐b‐PPS/Pep‐PA MCS (PEG‐b‐PPS/SDV‐1‐PA), 3% SDV containing PEG‐b‐PPS/Pep‐PA MCs (PEG‐b‐PPS/SDV‐3‐PA), and 5% SDV containing PEG‐b‐PPS/Pep‐PA MCs (PEG‐b‐PPS/SDV‐5‐PA) the zeta values were −1.9 ± 0.96 mV, +1.5 ± 0.00 mV, and +7.4 ± 0.25 mV. There is a density‐dependent increase in the zeta potential observed in PEG‐b‐PPS/SDV‐PA. Similarly for PEG‐b‐PPS/cRGD‐5‐PA and PEG‐b‐PPS/SGV‐5‐PA, MCs the zeta potential values were +5.8 ± 0.0001 mV and +6.3 ± 0.0001 mV respectively (Figure 1E).
3.2. αvβ3 integrin blocking by PEG‐b‐PPS/SDV‐PA peptide MCs
The expression levels of integrin αvβ3 in HUVECs after overnight incubation with the Blank MCs and the PEG‐b‐PPS/Pep‐PA MCs were evaluated by confocal microscopy by staining with mouse anti‐human CD51/CD61 antibody. The results (Figure 2A,B) showed a significant difference in the anti‐αvβ3 antibody binding to the cells treated with PEG‐b‐PPS/SDV‐1‐PA MCs (59.512 ± 4.152 pixels, p < .05), PEG‐b‐PPS/SDV‐3‐PA MCs (49.360 ± 4.405 pixels), and PEG‐b‐PPS/SDV‐5‐PA MCs (36.807 ± 2.368 pixels) when compared to untreated control (84.439 ± 4.870 px). PEG‐b‐PPS/cRGD‐5‐PA MCs also blocked (53.94 ± 2.192 pixels, p < .01) the αvβ3 integrin (p < .01). No significant reduction in intensity was observed in blank and PEG‐b‐PPS/SGV‐5‐PA MCs (65.238 ± 5.077 pixels). Results show that PEG‐b‐PPS/SDV‐5‐PA MCs achieved the maximum αvβ3 integrin blocking when compared to control, Blank‐MCs PEG‐b‐PPS/SDV‐1‐PA MCs, and PEG‐b‐PPS/SDV‐5‐PA MCs and are comparable to PEG‐b‐PPS/cRGD‐5‐PA MCs.
FIGURE 2.
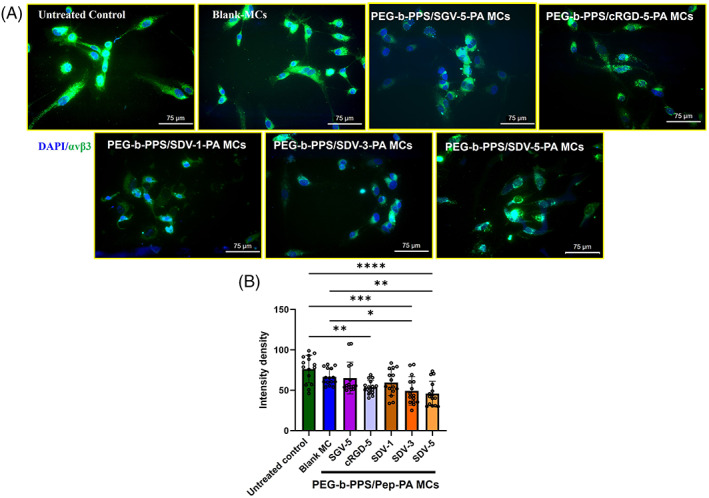
Inhibition of CD51/CD61 antibodies to αvβ3 receptor after short‐term incubation with PEG‐b‐PPS/Pep‐PA MCs. (A) confocal images of CD51/CD61 antibody staining and (B) quantitative expression of Integrin αvβ3 by ImageJ, where the color intensity recorded from individual cell and values were expressed as mean ± SD, n = 15 (SDV‐1 SDV‐3, SDV‐5 SGV‐5, and cRGD‐5 represent a different percentage [1%, 3% and 5%] of respective peptides on the MCs. The term “Pep” in figure B and C represents corresponding peptide sequence). p < .05 was considered significant. Statistical significance compared to control, and between the samples was determined by one‐way ANOVA, Tukey's model (*0.05, **0.01, ****0.0001). The scale bar is 75 μm
3.3. Suppression of vWF and PECAM‐1 expression by PEG‐b‐PPS /SDV‐PA MCs
HUVECs were stained with angiogenic markers vWF and PECAM‐1. The expression level of vWF and PECAM‐1 in HUVECs in the presence of Blank MCs and the PEG‐b‐PPS/Pep‐PA MCs are shown in Figure 3A. The intensity of vWF (Figure 3B) significantly reduced (p < .0001) in PEG‐b‐PPS/SDV‐1‐PA MCs (56.540 ± 2.443 pixels), PEG‐b‐PPS/SDV‐3‐PA MCs (49.837 ± 3.709 pixels), and PEG‐b‐PPS/SDV‐5‐PA MCs (37.270 ± 2.388 pixels), when compared to control (83.278 ± 5.56 pixels). PEG‐b‐PPS/cRGD‐5‐PA MCs (61.492 ± 2.99 pixels, p < .0001) also showed a significant decrease in vWF expression compared to control. Blank and PEG‐b‐PPS/SGV‐5‐PA MCs (54.600 ± 1.629 pixels) did not show a significant reduction. PEG‐b‐PPS/SDV‐5‐PA MCs treated cells showed significantly lower vWF expression compared to Blank‐MCs as well. Similarly, for PECAM‐1 (Figure 3C) PEG‐b‐PPS/SDV‐1‐PA MCs (72.536 ± 3.913 pixels, p < .001), PEG‐b‐PPS/SDV‐3‐PA MCs (52.193 ± 2.707 pixels, p < .0001), PEG‐b‐PPS/SDV‐5‐PA MCs (40.394 ± 3.335 pixels p < .0001) showed significantly lower expression compared to untreated control (101.732 ± 5.988 pixels); however, PEG‐b‐PPS/SGV‐5‐PA MCs and Blank MCs did not show a significant decrease in vWF and PECAM‐1 expression.
FIGURE 3.
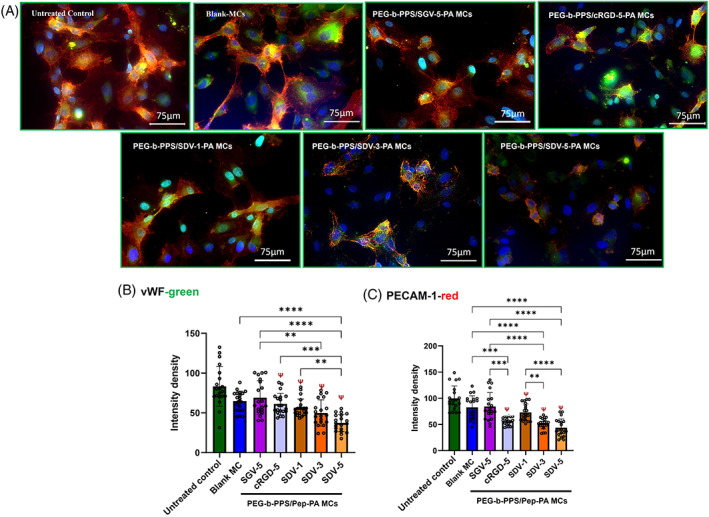
Expression of Platelet Endothelial Cell Adhesion molecule (PECAM‐1) and von Willebrand factor (vWF‐red) bu HUVEC cells. (A) Confocal microscopic image of HUVEC cells expressing PECAM‐1 (green) and vWF (red) after PEG‐b‐PPS/Pep‐PA MCs and Blank MCs. Quantitative expression of vWF and PECAM‐1 by ImageJ, where the color intensity recorded from individual cell and values were expressed as mean ± SD, n = 20 (B) PECAM‐1 and (C) vWF. (SDV‐1 SDV‐3, SDV‐5 SGV‐5, and cRGD‐5 represent different percentages [1%, 3% and 5%] of respective peptides on the MCs. The term “Pep” in figure B and C represents corresponding peptide sequence). p < .005 was considered significant. Statistical significance compared to control (ψ), and between the samples (*) was determined by one‐way ANOVA, Tukey's model. The scale bar is 75 μm
3.4. Peptide density‐dependent inhibition of migration
Migration plays a very crucial role in angiogenesis. The images (Figure 4A) show the inhibition in HUVEC migration by PEG‐b‐PPS/SDV‐1‐PA MCs, PEG‐b‐PPS/SDV‐3‐PA MCs, and PEG‐b‐PPS/SDV‐5‐PA MCs with a maximum inhibition at 5% peptide surface density on MCs. In control, 91.46% of wound healing occurred while in PEG‐b‐PPS/SDV‐1‐PA MCs, PEG‐b‐PPS/SDV‐3‐PA MCs, and PEG‐b‐PPS/SDV‐5‐PA MCsabout 93.93%, 23.42%, and 10.04% of wound healing (Figure 4B) respectively, was observed. PEG‐b‐PPS/SGV‐5‐PA MCs show 52.75% of wound healing and PEG‐b‐PPS/cRGD‐5‐PA MCs show 37.84% when compared to control.
FIGURE 4.
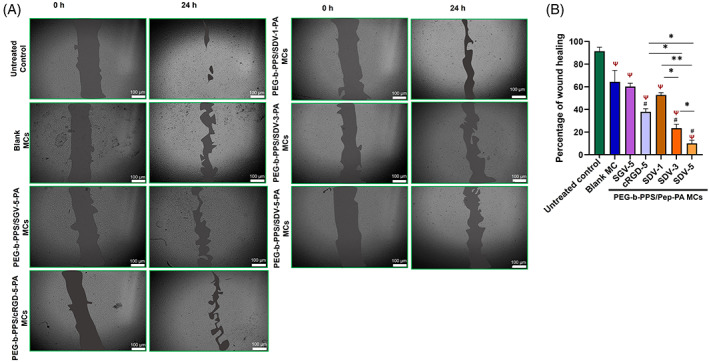
Inhibition of HUVEC cell migration after PEG‐b‐PPS/Pep‐PA MCs treatment. (A) Microscopic image of HUVECs treated with Blank MCs and PEG‐b‐PPS/Pep‐PA MCs at 0 and 24 h. (B) The percentage of wound healing was analyzed using ImageJ software, where the area of the scratched space was recorded and values were expressed as mean ± SD, n = 3). (SDV‐1 SDV‐3, SDV‐5 SGV‐5, and cRGD‐5 represent different percentages [1%, 3% and 5%] of respective peptides on the MCs. The term “Pep” in figure B and C represents corresponding peptide sequence). The scale bar is 100 μm. Statistical significance compared to untreated control (ψ), between the samples (*) and compared to blank‐MCs (#), was determined by one‐way ANOVA, Tukey's model. p < .05 is considered significant
3.5. Disruption of endothelial tube formation by an increase in peptide density on MCs
Inhibition of tube formation in a dose‐dependent manner was observed at all the peptide densities tested. Figure 5A,B shows that at 100 μg/ml concentration, PEG‐b‐PPS/SDV‐1‐PA MCs (52.88 ± 0.003 pixels, p < .01), PEG‐b‐PPS/SDV‐3‐PA MCs (0.122 ± 0.27 pixels, p < .0001), PEG‐b‐PPS/SDV‐5‐PA MCs (0.04 ± 0.15 pixels, p < .0001), and PEG‐b‐PPS/cRGD‐5‐PA MCs (0.08 ± 0.27 pixels, p < .0001) significantly inhibited tube formation compared to untreated control. In addition, blank MCs and PEG‐b‐PPS/SGV‐5‐PA MCs MCs treatment also reduced the tube formation in endothelial cells though it was not significant compared to other anti‐angiogenic PEG‐b‐PPS/Pep‐PA MCs.
FIGURE 5.

Suppression in endothelial tube formation in vitro by PEG‐b‐PPS/Pep‐PA MCs. (A) Representative image of the effect of density of peptide on PEG‐b‐PPS MCs on the tube formation after 4 h of incubation. Where SGV‐5, cRGD‐5, SDV‐1, SDV‐3, and SDV‐5 are representing corresponding PEG‐b‐PPS/Pep‐PA MCs. (SDV‐1 SDV‐3, SDV‐5 SGV‐5, and cRGD‐5 represent different percentages [1%, 3% and 5%] of respective peptides on the MCs. The term “Pep” in figure B and C represents corresponding peptide sequence)). Scale bar 100 μM. (b) The bar graph represents the quantification of the tubes formed using ImageJ an angiogenesis analyzer plugin where values were expressed as mean ± SD, n = 3, p < .05 was considered significant. Statistical significance between the samples (*) and compared to control (ψ) was determined by one‐way ANOVA, Tukey's model
4. DISCUSSION
The PEG‐b‐PPS block‐copolymer is a versatile delivery platform that can self‐assemble into thermodynamically stable structures like MCs, polymersomes, and filomicelles. 21 , 22 , 37 These systems have been extensively studied in terms of their sustained drug release kinetics, toxicity, and cell uptake both in vitro and in vivo. 22 , 30 , 39 In our previous study, we showed that an anti‐integrin (anti‐αvβ3) SDV heptapeptide bound to PEG‐b‐PPS MCs could efficiently inhibit angiogenesis with less peptide concentration in micellar form compared to the bare peptide. 37 This demonstrated the new approach where the targeting ligand can act as therapeutic moiety as well.
Another important design consideration is the surface density of ligands on nanomaterials. However, there is no specific number that can be followed since it depends primarily on the structure of the ligand. 40 A dense hydrophilic corona can support both stability and non‐specific protein binding in nanocarriers. Steric shielding of the surface‐bound peptide using polyethylene glycol functionalization to prevent off‐target binding was previously demonstrated by Fleischmann et al 41 Further receptor binding and endocytosis require thermodynamically favored membrane curvature for receptor clustering. In this research, there is an increase in the size of PEG‐b‐PPS/Pep‐PA MCs compared to Blank‐ MCs (Figure 1C). This increase in the size of the peptide conjugated to the MCs when compared to the blank MCs occurs due to the surface display of peptides on the MC surfaces as reported by Lin et al 42 and Nagaraj et al 37 We have generated MCs with the varying surface density (1%–5% peptide) to optimize the formulation design. We demonstrated that the zeta potential values of the PEG‐b‐PPS/Pep‐PA MCs and Blank MCs lie within −10 to 10 mV (Figure 1E), and hence high stability of the peptide constructs can be expected. The surface potential increase with the peptide surface density from 1% to 5% on MCs. cRGD (5%) and SGV (5%) peptide‐bound MCs showed higher zeta potential values when compared to blank MCs with no peptide.
In addition, the binding of the peptides on MCs does not affect the nanocarrier stability and allows delivery of the therapeutically required load to the target site with similar biodistributions in vivo as shown previously. 43 The integrin bocking by SDV peptide was consistent with Bang et al, 18 where the peptides showed a significant reduction in αvβ3 receptor expression. However, the concentration of the peptide required to induce the blocking was significantly reduced when presented on the micellar surface. 37 From this study, there is clear evidence of density‐dependent blocking of integrin receptors and that can be attributed to the enhanced receptor clustering, which subsequently improved the anti‐angiogenic efficiency of the peptide‐bound MCs by reducing the expression of angiogenic markers, vWF, and PECAM‐1. Studies demonstrate that the inhibition of vWF decreases angiogenesis by HUVECs. 44 In addition, there is a clear correlation between the membrane expression of PECAM‐1 with the expression of β3 integrin on the cell surface in endothelial cells. 45 Our results showed a positive correlation between αvβ3 blocking and PECAM‐1 and vWF expression in SDV‐3 and SDV‐5‐ MCs in a density‐dependent manner (Figure 3).
The previous investigation on PTX‐loaded liposome carrying RGD peptide and neuropilin‐a (NRP‐1, a VEGF receptor) demonstrated a bimodal liposome which enabled higher internalization than liposome carrying single ligand. 46 In addition, the cRGD‐based LTTs using Poly e‐caprolactone, poly (lactic acid), and Poly (lactic‐co‐glycolic acid) with and without Polyethylene glycol as a hydrophilic component of MCs were investigated extensively for drug delivery. 47 , 48 , 49 Where the binding with the receptor will allow the internalization of nanomaterials followed by drug diffusion through a polymer matrix. 50 , 51 However, cRGD can bind to multiple integrins and is not specific to abnormal angiogenesis. A more specific ligand against abnormal angiogenesis would be beneficial to avoid adverse effects on normal blood vessels. PEG‐b‐PPS/SDV‐PA MCs with 5% and 3% densities of peptides induce a 99% inhibition in tube formation in αvβ3 overexpressed endothelial cells. Further analysis of endothelial cell migration, one of the critical stages in the process of angiogenesis, also showed a peptide density‐dependent reduction in cell migration, demonstrating the anti‐angiogenic potential of the platform. PEG‐b‐PPS/SDV‐5‐PA MCs showed 90.04% inhibition in cell migration when compared to control (Figure 4). Studies by Bang et al and Naik et al showed that endothelial cell migration during angiogenesis is mediated by autophosphorylation of FAK. The anti‐angiogenic peptide after binding with integrin αvβ3 inhibits the phosphorylation of FAK and hence inhibits the migration of endothelial cells. 18 , 52 The results obtained for proliferation, migration, and tube formation suggest that PEG‐b‐PPS/Pep‐PA MCs show a density‐dependent inhibition of all the processes of angiogenesis (Figure 5). Even though the random peptide used in this study did not induce any toxicity in proliferation assay, 37 it has some minor effects on migration and tube formation assays. The reason for such an effect needs further investigation.
5. CONCLUSIONS
This study demonstrates the effect of varying the surface density of anti‐αvβ3 peptides on the efficiency of anti‐angiogenic LTT nano therapy. The increase in peptide density on MCs was observed through zeta potential analysis. From the optimization studies carried out, 5% density of peptides on MCs shows the highest integrin αvβ3 blocking when compared to 1% and 3%. These PEG‐b‐PPS/SDV‐5‐PA MCs showed inhibition in migration and tube formation, which are crucial events in angiogenesis. The integrin blocking directly influences the expression of vWF and PECAM‐1 in HUVEC cells. The developed PEG‐b‐PPS‐/SDV‐PA MCs could be an efficient platform for LTT and present novel treatment options for patients afflicted with angiogenesis‐related disease conditions.
CONFLICT OF INTEREST
The authors declare no conflict of interest.
ACKNOWLEDGMENTS
The authors acknowledge the support from Regenerative Medicine and Disability Lab at Biomedical Sciences. The authors also thank the support from NUANCE/SHyNE facilities at Northwestern University for CryoTEM analysis. The authors acknowledge the financial support by NIH/NIBIB R03EB032602, Blazer foundation, and Medical Biotechnology Program at Biomedical Sciences, UIC Rockford. The authors than thank Dr. Fanfan Du, Biomedical Engineering, Northwestern University for helping with polymer synthesis.
Bhushan NP, Stack T, Scott EA, Shull KR, Mathew B, Bijukumar D. In vitro assessment of varying peptide surface density on the suppression of angiogenesis by micelles displaying αvβ3 blocking peptides. J Biomed Mater Res. 2023;111(2):343‐353. doi: 10.1002/jbm.b.35154
Funding information National Institute of Health, Grant/Award Number: R03EB032602
DATA AVAILABILITY STATEMENT
The data that support the findings of this study are available from the corresponding author upon reasonable request.
REFERENCES
- 1. Elias DR, Poloukhtine A, Popik V, Tsourkas A. Effect of ligand density, receptor density, and nanoparticle size on cell targeting. Nanomedicine. 2013;9(2):194‐201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Srinivasarao M, Low PS. Ligand‐targeted drug delivery. Chem Rev. 2017;117(19):12133‐12164. [DOI] [PubMed] [Google Scholar]
- 3. Bhise NS, Shmueli RB, Sunshine JC, Tzeng SY, Green JJ. Drug delivery strategies for therapeutic angiogenesis and antiangiogenesis. Expert Opin Drug Deliv. 2011;8(4):485‐504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Seah I, Zhao X, Lin Q, et al. Use of biomaterials for sustained delivery of anti‐VEGF to treat retinal diseases. Eye. 2020;34(8):1341‐1356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Karumanchi DK, Gaillard ER. Sustained release drug delivery for treating ocular angiogenesis. Invest Ophthalmol Vis Sci. 2015;56(7):4157. [Google Scholar]
- 6. Geng H, Song H, Qi J, Cui D. Sustained release of VEGF from PLGA nanoparticles embedded thermo‐sensitive hydrogel in full‐thickness porcine bladder acellular matrix. Nanoscale Res Lett. 2011;6(1):1‐8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Liu P, Gao C, Chen H, et al. Receptor‐mediated targeted drug delivery systems for treatment of inflammatory bowel disease: opportunities and emerging strategies. Acta Pharm Sin B. 2021;11(9):2798‐2818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Park SC, Jeen YT. Anti‐integrin therapy for inflammatory bowel disease. World J Gastroenterol. 2018;24(17):1868‐1880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Chung CH, Chang CH, Hsu CC, Lin KT, Peng HC, Huang TF. Aggretin venom polypeptide as a novel anti‐angiogenesis agent by targeting integrin alpha2beta1. Sci Rep. 2017;7(1):1‐11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Bhatwadekar AD, Kansara V, Luo Q, Ciulla T. Anti‐integrin therapy for retinovascular diseases. Expert Opin Investig Drugs. 2020;29(9):935‐945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Sepp NT, Li LJ, Lee KH, et al. Basic fibroblast growth factor increases expression of the αvβ3 integrin complex on human microvascular endothelial cells. J Investig Dermatol. 1994;103(3):295‐299. [DOI] [PubMed] [Google Scholar]
- 12. Kluza E, Van Der Schaft DW, Hautvast PA, et al. Synergistic targeting of αvβ3 integrin and galectin‐1 with heteromultivalent paramagnetic liposomes for combined MR imaging and treatment of angiogenesis. Nano Lett. 2010;10(1):52‐58. [DOI] [PubMed] [Google Scholar]
- 13. Bianchini F, De Santis A, Portioli E, et al. Integrin‐targeted AmpRGD sunitinib liposomes as integrated antiangiogenic tools. Nanomedicine. 2019;18:135‐145. [DOI] [PubMed] [Google Scholar]
- 14. Jiang Z, Guan J, Qian J, Zhan C. Peptide ligand‐mediated targeted drug delivery of nanomedicines. Biomater Sci. 2019;7(2):461‐471. [DOI] [PubMed] [Google Scholar]
- 15. Raab‐Westphal S, Marshall JF, Goodman SL. Integrins as therapeutic targets: successes and cancers. Cancer. 2017;9(9):110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. McLean LP, Cross RK. Integrin antagonists as potential therapeutic options for the treatment of Crohn's disease. Expert Opin Investig Drugs. 2016;25(3):263‐273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Millard M, Odde S, Neamati N. Integrin targeted therapeutics. Theranostics. 2011;1:154‐188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Bang JY, Kim EY, Kang DK, et al. Pharmacoproteomic analysis of a novel cell‐permeable peptide inhibitor of tumor‐induced angiogenesis. Mol Cell Proteomics. 2011;10(8):M110.005264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Sorriento D, Santulli G, Ciccarelli M, et al. The amino‐terminal domain of GRK5 inhibits cardiac hypertrophy through the regulation of calcium‐calmodulin dependent transcription factors. Int J Mol Sci. 2018;19(3):861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Cao Z, Suo X, Chu Y, et al. Peptides derived from the integrin β cytoplasmic tails inhibit angiogenesis. Cell Commun Signal. 2018;16(1):1‐15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Stack T, Vahabikashi A, Johnson M, Scott E. Modulation of Schlemm's canal endothelial cell stiffness via latrunculin loaded block copolymer micelles. J Biomed Mater Res A. 2018;106(7):1771‐1779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Yi S, Zhang X, Sangji MH, et al. Surface engineered polymersomes for enhanced modulation of dendritic cells during cardiovascular immunotherapy. Adv Funct Mater. 2019;29(42):1904399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Fu Y, Ponce ML, Thill M, Yuan P, Wang NS, Csaky KG. Angiogenesis inhibition and choroidal neovascularization suppression by sustained delivery of an integrin antagonist, EMD478761. Invest Ophthalmol Vis Sci. 2007;48(11):5184‐5190. [DOI] [PubMed] [Google Scholar]
- 24. Kumar CC, Malkowski M, Yin Z, et al. Inhibition of angiogenesis and tumor growth by SCH221153, a dual αvβ3 and αvβ5 integrin receptor antagonist. Cancer Res. 2001;61(5):2232‐2238. [PubMed] [Google Scholar]
- 25. Allen TM. Ligand‐targeted therapeutics in anticancer therapy. Nat Rev Cancer. 2002;2(10):750‐763. [DOI] [PubMed] [Google Scholar]
- 26. Danhier F, Le Breton A, Préat V. RGD‐based strategies to target alpha (v) beta (3) integrin in cancer therapy and diagnosis. Mol Pharm. 2012;9(11):2961‐2973. [DOI] [PubMed] [Google Scholar]
- 27. Fu S, Xu X, Ma Y, Zhang S, Zhang S. RGD peptide‐based non‐viral gene delivery vectors targeting integrin αvβ3 for cancer therapy. J Drug Target. 2019;27(1):1‐11. [DOI] [PubMed] [Google Scholar]
- 28. Al‐Mansoori L, Elsinga P, Goda SK. Bio‐vehicles of cytotoxic drugs for delivery to tumor specific targets for cancer precision therapy. Biomed Pharmacother. 2021;144:112260. [DOI] [PubMed] [Google Scholar]
- 29. Mezu‐Ndubuisi OJ, Maheshwari A. The role of integrins in inflammation and angiogenesis. Pediatr Res. 2021;89(7):1619‐1626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Stack T, Vincent M, Vahabikashi A, et al. Targeted delivery of cell softening micelles to Schlemm's canal endothelial cells for treatment of glaucoma. Small. 2020;16(43):2004205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Mendes PM. Cellular nanotechnology: making biological interfaces smarter. Chem Soc Rev. 2013;42(24):9207‐9218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Zhao J, Santino F, Giacomini D, Gentilucci L. Integrin‐targeting peptides for the design of functional cell‐responsive biomaterials. Biomedicine. 2020;8(9):307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Lin J, Miao L, Zhong G, Lin CH, Dargazangy R, Alexander‐Katz A. Understanding the synergistic effect of physicochemical properties of nanoparticles and their cellular entry pathways. Commun Biol. 2020;3(1):1‐10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Vincent MP, Bobbala S, Karabin NB, et al. Surface chemistry‐mediated modulation of adsorbed albumin folding state specifies nanocarrier clearance by distinct macrophage subsets. Nat Commun. 2021;12(1):1‐18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Vincent MP, Karabin NB, Allen SD, et al. The combination of morphology and surface chemistry defines the immunological identity of nanocarriers in human blood. Adv Therap. 2021;4(8):2100062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Nel AE, Mädler L, Velegol D, et al. Understanding biophysicochemical interactions at the nano–bio interface. Nat Mater. 2009;8(7):543‐557. [DOI] [PubMed] [Google Scholar]
- 37. Nagaraj R, Stack T, Yi S, et al. High density display of an anti‐angiogenic peptide on micelle surfaces enhances their inhibition of αvβ3 integrin‐mediated neovascularization in vitro. Nanomaterials. 2020;10(3):581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Karabin NB, Allen S, Kwon HK, et al. Sustained micellar delivery via inducible transitions in nanostructure morphology. Nat Commun. 2018;9(1):1‐13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Burke JA, Zhang X, Bobbala S, et al. Subcutaneous nanotherapy repurposes the immunosuppressive mechanism of rapamycin to enhance allogeneic islet graft viability. Nat Nanotechnol. 2022;17(3):319‐330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Gao H, Shi W, Freund LB. Mechanics of receptor‐mediated endocytosis. Proc Natl Acad Sci U S A. 2005;102(27):9469‐9474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Fleischmann D, Maslanka Figueroa S, Goepferich A. Steric shielding of cRGD‐functionalized nanoparticles from premature exposition to off‐target endothelial cells under a physiological flow. ACS Appl Biol Mater. 2020;4(1):640‐650. [Google Scholar]
- 42. Lin WJ, Chien WH. Peptide‐conjugated micelles as a targeting nanocarrier for gene delivery. J Nanopart Res. 2015;17(9):1‐14. [Google Scholar]
- 43. Yin H, Gong C, Shi S, Liu X, Wei Y, Qian Z. Toxicity evaluation of biodegradable and thermosensitive PEG‐PCL‐PEG hydrogel as a potential in situ sustained ophthalmic drug delivery system. J Biomed Mater Res Part B. 2010;92(1):129‐137. [DOI] [PubMed] [Google Scholar]
- 44. Starke RD, Ferraro F, Paschalaki KE, et al. Endothelial von Willebrand factor regulates angiogenesis. Blood. 2011;117(3):1071‐1080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Imamaki R, Ogawa K, Kizuka Y, et al. Glycosylation controls cooperative PECAM‐VEGFR2‐β3 integrin functions at the endothelial surface for tumor angiogenesis. Oncogene. 2018;37(31):4287‐4299. [DOI] [PubMed] [Google Scholar]
- 46. Nasongkla N, Shuai X, Ai H, et al. cRGD‐functionalized polymer micelles for targeted doxorubicin delivery. Angew Chem. 2004;116(46):6483‐6487. [DOI] [PubMed] [Google Scholar]
- 47. Wang Y, Tong L, Wang J, et al. cRGD‐functionalized nanoparticles for combination therapy of anti‐endothelium dependent vessels and anti‐vasculogenic mimicry to inhibit the proliferation of ovarian cancer. Acta Biomater. 2019;94:495‐504. [DOI] [PubMed] [Google Scholar]
- 48. Martínez‐Jothar L, Barendrecht AD, de Graaff AM, et al. Endothelial cell targeting by cRGD‐functionalized polymeric nanoparticles under static and flow conditions. Nanomaterials. 2020;10(7):1353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Cai M, Ye M, Shang X, et al. cRGD‐functionalized redox‐sensitive micelles as potential doxorubicin delivery carriers for α v β 3 integrin over expressing tumors. RSC Adv. 2015;5(112):92292‐92302. [Google Scholar]
- 50. Danhier F, Vroman B, Lecouturier N, et al. Targeting of tumor endothelium by RGD‐grafted PLGA‐nanoparticles loaded with paclitaxel. J Control Release. 2009;140(2):166‐173. [DOI] [PubMed] [Google Scholar]
- 51. Danhier F, Ansorena E, Silva JM, Coco R, Le Breton A, Préat V. PLGA‐based nanoparticles: an overview of biomedical applications. J Control Release. 2012;161(2):505‐522. [DOI] [PubMed] [Google Scholar]
- 52. Naik MU, Mousa SA, Parkos CA, Naik UP. Signaling through JAM‐1 and αvβ3 is required for the angiogenic action of bFGF: dissociation of the JAM‐1 and αvβ3 complex. Blood. 2003;102(6):2108‐2114. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.