

Abstract
Plants have evolved diverse reproductive allocation strategies and seed traits to aid in dispersal, persistence in the seed bank, and establishment. In particular, seed size, dormancy, and early seedling vigor are thought to be key functional traits with important recruitment and fitness consequences across abiotic stress gradients. Selection for favored seed‐trait combinations, or against maladaptive combinations, is likely an important driver shaping recruitment strategies. Here, we test for seed‐trait plasticity and patterns of recruitment using two genotypes representative of contrasting upland and lowland ecotypes of Panicum hallii with field experiments in native versus foreign habitats. Furthermore, we test whether seed traits have been under directional selection in P. hallii using the v‐test based on trait variance in a genetic cross. Finally, we evaluate the genetic architecture of ecotypic divergence for these traits with quantitative trait locus (QTL) mapping. Field experiments reveal little plasticity but support a hypothesis of adaptation divergence among ecotypes based on recruitment. Patterns of segregation within recombinant hybrids provides strong support for directional selection driving ecotypic divergence in seed traits. Genetic mapping revealed a polygenic architecture with evidence of genetic correlation between seed mass, dormancy, and seedling vigor. Our results suggest that the evolution of these traits may involve constraints that affect the direction of adaptive divergence. For example, seed size and germination percentage shared two colocalized QTL with antagonistic additive effects. This supports the hypothesis of a functional genetic relationship between these traits, resulting in either large seed/strong dormancy or small seed/weak dormancy trait combinations. Overall, our study provides insights into the factors facilitating and potentially constraining ecotypic differentiation in seed traits.
Keywords: Dormancy, ecotypic divergence, Panicum hallii, QTL, quantitative genetics, reciprocal transplantation, seed size
Impact Summary
Seed size and dormancy are key functional traits with important recruitment and fitness consequences. Theory suggests trade‐offs and plasticity in offspring quantity and quality are important in seed evolution. The genetics of seed size and dormancy traits have been studied extensively, but these studies are mostly limited to model systems or domesticated crops. We also know very little about the genetic architecture and evolution of seed‐based life‐history traits especially considering adaptation to xeric and mesic habitats. Here, we explored the genetic basis of trade‐offs between seed size and dormancy in a C4 perennial grass, Panicum hallii, endemic to North America. We planted seeds of recombinant inbred lines from a cross between a xeric and mesic ecotype of P. hallii and mapped quantitative trait loci (QTL) for seed size, dormancy, and seedling vigor traits. We detected a genetic basis of trade‐offs between seed size and dormancy, suggesting that natural selection strongly favored specific trait combinations in ecotype formation. We further explored the role of seed size variation on seedling and adult recruitment in contrasting habitats. Our data showed that seed size was under strong selection through recruitment. Overall, our results demonstrate that adaptive differentiation for seed size and early life stages are important factors in adaptation to contrasting habitats.
Local adaptation is an important component of responses to changing environments, as it reveals how environmental variation can drive phenotypic and genetic differentiation as a response to selection (Davis and Shaw 2001; Conover et al. 2009). In this process, plant populations may be selected for different resource allocation strategies and trait combinations based on habitat structure or climatic features across a species’ range. One hypothesis is that local adaptation results from genetic trade‐offs that result in maladaptation to alternative environments (Roff 2002; Kawecki and Ebert 2004; Lascoux et al. 2016). For example, plants have evolved diverse seed and seedling traits in response to heterogeneity in the amount of rainfall (Smith and Fretwell 1974; Venable 1992; Daws et al. 2007). These seed/seedling traits trade‐offs may maintain genetic variation among populations and drive phenotypic partitions within species (Futuyma and Moreno 1988; Jasmin and Kassen 2007). Thus, exploring the genetic basis of trade‐offs and how they shape ecological strategy is of major importance for evolutionary biology (Agrawal et al. 2010; Postma and Ågren 2016).
In plants, soil water availability plays a vital role maintaining global species distribution and determining genetic and phenotypic differences within a species’ range (Stebbins Jr 1952; Whittaker 1970; Woodward et al. 2004). Thus, plant adaptation to habitats differing in soil water availability can drive divergence and may promote ecological speciation by selecting distinct morphological and/or physiological characteristics that provide an environment‐specific fitness advantage (Baker 1972; Rundle and Nosil 2005; Roux et al. 2006; Juenger 2013). However, studies that have looked at the genetic basis of ecotypic differentiation are often based on seedling transplantation (Lowry and Willis 2010; Fournier‐Level et al. 2013) and so may exclude important aspects of divergence in earlier life stages (Donohue et al. 2010; Donohue 2014). This is surprising as early life stages are often crucial because this vulnerable period of development can suffer from high mortality rates (Kitajima and Fenner 2000; Donohue et al. 2010), thus selection may be severe during this part of the life cycle. Additionally, early life experiences often have nonlinear effects on later life‐history traits (Beckerman et al. 2002). Thus, it may be difficult or impossible to fully understand adaptation if juvenile stages are ignored.
Seed size plays an important role in early growth and establishment. It connects the ecology of reproductive strategies with seed dormancy, seedling establishment, and vegetative growth (Shipley et al. 1989; Leishman and Westoby 1992; Rees 1996; Grime et al. 2014). Thus, selection might favor different seed size strategies across heterogeneous habitats. For example, plants adapted to xeric habitats tend to have larger seeds compared to those occurring in mesic habitats (Baker 1972). A strategy producing fewer large seeds might ensure successful establishment by generating vigorous seedling growth, perhaps timed with pulses of seasonal precipitation. In contrast, a strategy producing many small seeds might arise in environments with highly predictable patterns of rainfall or where establishment and success are limited by other resources. Several studies have shown a positive correlation with seed mass and seedling root and shoot growth in different species (Wulff 1986; Jurado and Westoby 1992; Lloret et al. 1999). However, it is not clear whether seed and seedling traits jointly evolve by common pleiotropic effects or strong linkage disequilibrium or if these traits evolve independently.
Additionally, seed dormancy is another important trait tightly linked to plant life history (Venable 2007; Childs et al. 2010). A central idea is that dormancy is a bet‐hedging strategy for reproduction in an unpredictable and heterogeneous environment (Venable 2007; Childs et al. 2010). In systems with strong dormancy, specific environmental conditions must occur to break dormancy but also to ensure germination after dormancy is broken (Baskin and Baskin 1998). Empirical studies showed inconsistent relationships between seed mass and germination percentage traits. For example, studies conducted with Medicago truncatula and Brassica oleracea showed no genetic correlation between these two traits (Bettey et al. 2000; Dias et al. 2011). Dechaine et al. (2014) showed a positive genetic correlation between seed mass and germination percentage trait in Brassica rapa. In tomato, no genetic correlation was found between seed size and germination percentage when tested in greenhouse condition (Khan et al. 2012), but a negative genetic correlation was observed when tested under different nutritional conditions (Geshnizjani et al. 2020). Thus, it is not clear whether seed dormancy can evolve independently, or if dormancy is tightly linked with seed mass variation in natural systems.
Although seed and early life stages are likely critical in plant performance, our knowledge about the genetic basis of these traits, their plasticity, or their role in local adaptation is limited to only a few systems (Clauss and Aarssen 1994; Donohue et al. 2010; Donohue 2014). Seed mass and dormancy exhibit considerable phenotypic variation in nature (Baker 1972; Lacerda et al. 2004; Bradford and Nonogaki 2008; Bentsink et al. 2010; Harel et al. 2011). This variation might result from phenotypic plasticity or genetic differentiation. Based on habitat and seed size correlations, patterns in nature suggest seed size evolution is adaptive and is generally thought to be under stabilizing selection due to a seed size and number trade‐off (Smith and Fretwell 1974; Westoby and Rice 1982; Silvertown 1989; Venable 1992; Mojonnier 1998). Thus, it is not surprising that species reproductive allocation and establishment strategies are associated by the variation in seed mass. However, these conclusions are mostly based on evidence from experiments with crop plants grown in controlled environments (Silvertown 1989). Studies with wild plants often show marked phenotypic plasticity and low heritability of seed size (Primack and Antonovics 1981; Stanton 1984). If the variation is not plastic and instead caused by genetic differentiation, then exploring genetic architecture underlying this natural variation may help understand the impact of different selective forces driving seed size‐related phenotypic variation. It also helps to understand how adaptive evolution shaped genetic variability for these traits.
Here, we report on work exploring seed trait ecology and genetics in Panicum hallii, a self‐fertilizing C4 perennial bunch grass species. Panicum hallii is distributed across the southwestern part of the United States and northern Mexico (Smeins et al. 1976; Waller 1976; Hatch et al. 1999; Gould et al. 2018; Lovell et al. 2018b), with population genetic structure and putatively adaptive differentiation across its range (Lowry et al. 2013; Palacio‐Mejía et al. 2021a). However, the greatest differentiation within P. hallii occurs between ecotypes adapted to lowland/coastal and upland/southwestern regions (Lowry et al. 2015). These two ecotypes are P. hallii var. hallii and P. hallii var. filipes. hallii is the more widespread ecotype and occurs primarily in xeric upland calcareous soils, whereas filipes is located primarily in mesic and seasonal wet areas (Lovell et al. 2018a). In our study, we used field experiments to ask whether seed traits exhibit phenotypic plasticity in response to xeric and mesic habitat variation using two representative genotypes. Second, we used field seed addition experiments to test for trade‐offs based on seedling recruit and survivorship patterns. Finally, we used detailed growth chamber studies with a recombinant inbred population to explore the genetic architecture underlying ecotypic divergence in seed and seedling traits using quantitative trait loci (QTL) mapping. Together, our results provide insight in possible avenues of local adaptation mediated by seed traits.
Methods
PLANT MATERIALS AND RECIPROCAL TRANSPLANTATION
For plant material, we picked two representative inbred genotypes of the upland and lowland ecotypes (hallii; HAL2 genotype and filipes; FIL2 genotype, respectively) of Panicum hallii. The upland ecotype was collected from the Lady Bird Johnson Wildflower Center (Austin, TX, USA; 30.19°N, 97.87°W) and the lowland ecotype was collected from a natural area associated with the Corpus Christi Botanical Garden in South Texas (27.65°N, 97.40°W). Previous studies with natural populations collected across the P. hallii species range showed significant genetic and phenotypic divergence between these ecotypes, and the representative genotypes (HAL2 and FIL2) in this study were also used for genome assembly and ecotype characterization in several other studies (Lowry et al. 2013; Lowry et al. 2015; Gould et al. 2018; Lovell et al. 2018b; Khasanova et al. 2019; Palacio‐Mejía et al. 2021b). In a recent paper, Palacio‐Mejia et. al. (2021b) described the phenotypic variation within and divergence between ecotypes in P. hallii using natural accessions collected across its geographic ranges. This study revealed continuous divergence between lowland and upland ecotypes and the representative lines (HAL2 and FIL2) differed significantly from one another in multivariate space (Palacio‐Mejía et al. 2021b).
We reciprocally transplanted seedlings of hallii (HAL2) and filipes (FIL2) to their lowland (Nueces Delta Preserve [NDP], Odem, Texas) and upland natural habitats (Brackenridge Field Lab [BFL], Austin, Texas) to evaluate seed trait plasticity. The Austin site (30.28°N, 97.78°W) represents a typical upland habitat with dry and shallow calcareous soil, whereas the Odem site (27.92°N, 97.61°W) represents the lowland coastal prairie habitat. Both of these locations harbor natural populations of P. hallii, but we note here that these field sites are not the original location of origin for the HAL2 and FIL2 genotypes. We germinated seedlings in 1‐inch pot in a greenhouse located at the University of Texas, Austin and grew them to 20 days of age. We transferred 300 seedlings to the field sites in Fall 2018 (75 uplands + 75 lowlands = 150 × 2 sites = 300). After transplanting, we did not water or fertilize them but visited both sites in 30‐day intervals until November 2018, when we collected seeds from both sites. We later randomly picked seeds from 10 plants from each genotype to measure seed mass and dormancy. In total, we measured 40 seed lots (2 genotypes × 2 locations × 10 replicates = 40 seed lots). Seed mass was calculated by weighing 100 seeds per line using an analytical balance (Mettler Toledo, Ohio). We established a germination assay in a lab growth chamber with 15 seeds per petri dishes (unit of replication, total 40 petri dishes) and calculated the final percentage of germinated seeds for these lines.
To study P. hallii recruitment, we conducted another reciprocal transplant experiment at the same field sites. We established 14 plots (0.5 × 0.5 m2) at each site haphazardly arrayed over native habitat. In February 2019, we added 100 seeds per plot from either HAL2 or FIL2 at each of the two field locations (2 genotypes × 7 replicates × 2 field sites = 28 plots). We visited each site every 15 days from the date of seed addition and collected two seasons (spring and fall 2019) of seedling and adult (reproductive) recruitment data. During each visit, plots were inspected for new seedlings and tagged them using a mini rubber band. We also found other grass species emerging with P. hallii recruitment. To minimize confusion and misidentification, we also planted 10 HAL2 and 10 FIL2 in one‐gallon pots prepared at both field sites. We used these pots as a reference while identifying seedlings in the field. Once we identified and marked seedlings, we followed them from early establishment to flowering. We used flowering as a metric to count adult plants.
RIL POPULATION, LINKAGE MAP, AND EXPERIMENTAL SETUP
We used a recombinant inbred line (RIL) population generated from upland (hallii; HAL2 genotype) and lowland (filipes; FIL2 genotype) parental genotypes. A detailed description of the development of the RIL population and the genetic map construction are discussed in Khasanova et al. (2019). In brief, the RIL population was generated following a single seed descent method from F2 progeny of a single F1 hybrid to the F7 generation. A total of 335 RILs were re‐sequenced up to 30× coverage and used to build a high‐density linkage map after mapping against the FIL2 reference genome. The final map consists of 722 markers covering nine chromosomes. For this study, 295 RILs and both parental lines were collected from well‐watered plants grown as a seed increase at the Brackenridge Field Laboratory, Austin, Texas in September 2016. Seeds were collected from three maternal parents from each RIL. We carefully separated good seed from chaff, detritus, or undeveloped seeds and kept them at room temperature for 6 months before starting the experiment trials.
We used a large walk‐in growth chamber experiment to control environmental conditions throughout the experimental trials. Growth chamber temperature was set at 28°C during light and 22°C during dark conditions. The photoperiod for the chamber was set at 12 hours light/12 hours dark. Seeds were germinated in petri dishes (25 × 100 mm) with sterilized sand. We used 60 g of sands for each petri dish and added 12 mL tap water to each dish. Water was sprayed on the upper lid of the petri dishes to keep the inner environment moist, and each dish was sealed with parafilm. The experiment was replicated across three temporal blocks in which each block contained a single petri dish (our unit of replication) of each genetic line. In each petri dish, 15 seeds per lines were used. The complete experimental design was as follows: 295 RILs + 6 petri dishes of HAL2 + 6 petri dishes of FIL2 × 3 replication = 921 petri dishes.
Because we consistently observe strong seed dormancy in the hallii ecotype, we wanted to make sure an inference of dormancy in hallii (HAL2) is not simply the result of inviable/sterile seeds. Here, we applied scarification and found it to be effective for breaking dormancy in the upland hallii (HAL2) ecotype. We examined all nongerminated seeds from this germination assay to evaluate viability and dormancy by forcing germination after removing seed coats. We removed the seed coat by gently using sandpaper (P100) and then germinated them in petri dishes.
PHENOTYPIC TRAITS MEASUREMENT AT THE SEED, GERMINATION, AND EARLIER GROWTH STAGE
We measured six seed size and earlier seedling growth traits from our growth chamber experiment. Seed mass (SM) was measured by taking 100 seed weight per RIL line using an analytical balance. We recorded the day of first germination (GT) and germination percentage (GP) for each RIL as assayed in petri dish trials. Viability experiments with some random RIL lines and parental lines confirm that nongerminated seeds were dormant, and as such we view GP as a proxy for the degree of seed dormancy. Shoot length was measured twice on a representative seedling in a dish, at 5 days (SL5D) and at 10 days (SL10D) relative to their GT. Root length was also measured on a representative seedling per petri dish at 10 days (RL10D) relative to GT. A detailed description of our phenotypic data measurement is in File S1.
DATA PROCESSING, STATISTICS, AND QTL MAPPING
We took two approaches to analyze our reciprocal field experiment data. To analyze seed mass and germination percentage in the first reciprocal transplant experiment, we fit linear models with genotype (G), site (E), and their interaction (G×E) as the effect variables. To analyze counts of seedling and adult recruitments in the second reciprocal transplant experiment, we fit generalized linear models with a Poisson distribution and log link. These models took the same form as the linear model above. We determined that the Poisson GLM adequately fit the data using the Pearson goodness of fit statistics. All models were fit in JMP version 15.1.0 (SAS Institute, Cary, NC).
For quantitative genetic studies, we fit linear models for the measured phenotypic traits treating RIL genotype as a fixed effect. We also included temporal block in the model as a fixed effect covariate when the block had a significant effect on phenotypic traits. Most of our collected phenotypes except SM and SL10D had a significant block effect. We used RIL means as estimates of breeding values to calculate the bivariate genetic correlation between traits using “multivariate” function in JMP version 15.1.0. Broad‐sense heritability was estimated using h2boot software fitting one‐way ANOVA among inbred lines with 1000 bootstrap runs (Phillips and Arnold 1999). Parental trait divergence was estimated by mean differences and significance tested with a t‐test in SAS. Resulting P‐values were adjusted for multiple testing via the Benjamini and Hochberg's (1995) method of false discovery rate estimation.
Fraser (2020) noted that the pattern of trait variance in recombinant populations can provide insight into whether the parental trait divergence is consistent with neutral divergence or more likely the result of natural selection. The approach is a generalization of the QTL sign test (Orr 1998) and rests on the realization that the phenotypic distribution in a segregating/recombinant population can be treated as a null model for the phenotypes expected under neutral evolution. In this framework, the pattern of underlying QTL effects under strong directional selection is expected to be complementary (in a consistent direction) if the trait has experienced consistent strong directional selection leading to parental divergence, whereas a mixed of positive and negative effects is expected if the trait evolved by neutral processes. By extension, if a trait has experienced strong directional selection driving parental divergence the variance among parental lines is expected to be larger than the segregating variance in the recombinant population, The v‐test was performed as described in equation (2) of Fraser (2020) for an RIL. In brief, to calculate v, we first estimated the trait variances within and between parents of the cross. Then, we estimated the variance among RIL means and used a c value of 1 (Fraser 2020).
All measured morphological traits in the RIL population were normally distributed except for SL10D and RL10D. These traits were log transformed before QTL mapping. We performed QTL mapping on RIL breeding values by stepwise fitting of multiple QTL models in R/qtl package (Broman et al. 2003). The input file for QTL mapping including the linkage map and phenotypic traits is in File S2. We used two functions to determine QTL positions and for the estimation of additive and pairwise epistatic effects. We first calculated penalties for main effects and their interactions using Haley‐Knott regression by 1000 permutations of the scan‐two function for each phenotypic trait. We then performed a forward/backward stepwise search for models with a maximum of six QTL that optimized the penalized logarithm of the odds (LOD) score criterion using the stepwise procedure. Significance thresholds were determined at an experiment wide Type 1 error rate of α = 0.05. Confidence intervals for significant QTLs were calculated as the 1.5 LOD drop interval by using the qtlStats function from qtlTools (github.com/jtlovell/qtlTools).
Results
SEED MASS AND EARLIER GROWTH TRAITS VARIED SIGNIFICANTLY BETWEEN PARENTAL LINES
Parental lines exhibited considerable phenotypic divergence in seed size, dormancy, and earlier growth traits (Fig. 1). In growth chamber experiments, parental lines significantly differed for all six measured traits (Table 1). The upland ecotype had higher values in all traits compared to the lowland ecotype except for the GP. For example, HAL2 had 56% greater SM than FIL2 and the GP in FIL2 was 63% greater than HAL2. The mean difference in GT between ecotypes was 60 hours (±2.7), with FIL2 germinating earlier than HAL2. Early shoot and root lengths in HAL2 were greater than FIL2. Shoot length after 5 days was 10% greater in HAL2 and the differences increased to 24% at the 10‐day measure. Ten days old root growth was 73% greater in HAL2 (Table 1; File S3).
Figure 1.
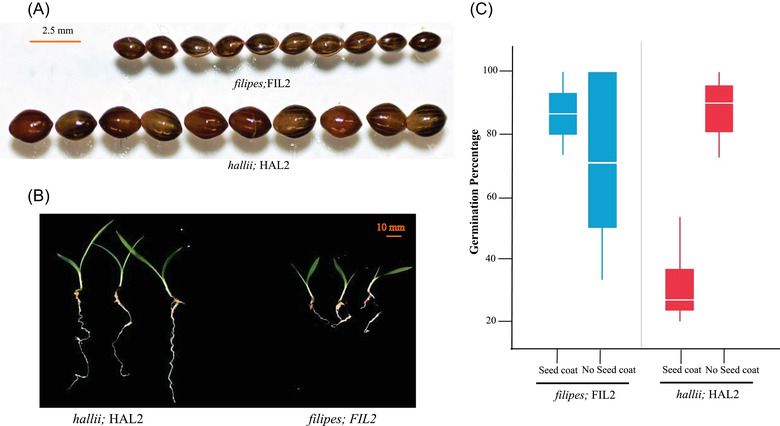
Seed and seedling size differences and seed viability tests between Panicum hallii upland (HAL2) and lowland (FIL2) representative genotypes. Panel A shows the representative differences in seed size between parental lines based on 10 random seeds for both parental lines. Panel B shows seedling growth differences for these parental lines at 10 days from the day of first germination. The upland ecotype grew larger than the lowland ecotype in both belowground and above ground biomass. Panel C shows the result of seed viability test between hallii (HAL2, red) and filipes (FIL2, blue). Ungerminated seeds were tested for germination by removing seed coats. The Y‐axis corresponds to the percent of germinated seeds and the X‐axis provides the genotype information. HAL2 has a higher germination rate compared to FIL2 if seed coats are removed.
Table 1.
Descriptive statistics for phenotypic traits for hallii; HAL2 and filipes; FIL2 parental lines and the recombinant inbred population (SE = standard error). The P‐value corresponds to a t‐test comparing the parental lines (HAL2 vs. FIL2)
| Trait | N | HAL2 (Mean ± SE) | N | FIL2 (Mean ± SE) | P | N | RIL Mean | RIL Range | H 2 ± SE |
|---|---|---|---|---|---|---|---|---|---|
| Seed mass (SM) (mg) | 13 | 1.29 ± 0.01 | 15 | 0.57 ± 0.01 | <0.0001 | 293 | 0.91 | 0.38–1.56 | 0.89 ± 0.01 |
| Germination percentage (GP) | 13 | 31.2 ± 2.39 | 15 | 86.21 ± 2.23 | <0.0001 | 278 | 55.38 | 6.61–98.37 | 0.24 ± 0.03 |
| Shoot length at 5 days (SL5D) (mm) | 13 | 12.57 ± 0.43 | 15 | 11.27 ± 0.40 | 0.0373 | 278 | 14.40 | 4.61–21.94 | 0.30 ± 0.04 |
| Shoot length at 10 days (SL10D) (mm) | 13 | 23.8 ± 0.82 | 15 | 18.04 ± 0.70 | <0.0001 | 278 | 22.04 | 12.66–31.33 | 0.39 ± 0.16 |
| Root length at 10 days (RL10D) (mm) | 13 | 41.75 ± 1.47 | 15 | 11.14 ± 1.37 | <0.0001 | 278 | 16.62 | 6.13–59.80 | 0.63 ± 0.05 |
| Germination time (GT) (hours) | 13 | 92.3 ± 2.8 | 15 | 30.4 ± 2.61 | <0.0001 | 278 | 74.22 | 36–240 | 0.76 ± 0.04 |
Additionally, we tested germination percentage for the nongerminated seeds by removing seed coats to check if the observed dormancy was due to seed inviability. Our data showed that this scarification technique effectively increased seed germination for the dormant HAL2 seeds. Upland HAL2 seeds germination was increased to 88.2% after removing seed coats, a 64% increase compared to their regular germination in the absence of seed coat removal (Fig. 1; File S4). This result confirmed that the apparent seed dormancy in the upland ecotype was not due to seed viability issues and may be related to inhibition associated with the seed coat.
SEED PLASTICITY, SEEDLING RECRUITMENT, AND ADULT ESTABLISHMENT IN PARENTAL LINES
To explore the impact of seed development in native habitats on seed traits, we collected seeds from field plantings at reciprocal sites. We tested for genotype (G), site (E), and genotype‐by‐site (G×E) interaction effects on seed mass and germination traits. In this framework, we interpret significant G as evidence for ecotype divergence in seed traits, significant E as habitat driven seed plasticity, and G×E as ecotype variation in plastic responses. Seed mass revealed significant genotype main effects (P < 0.0001) but did not show significant effects of reciprocal sites (P = 0.86) or G×E interaction (P = 0.27) (Fig. 2A). We used field‐collected seeds in lab germination experiments to evaluate impacts on seed dormancy. As with seed mass, we observed strong genotype difference in germination percentage (P < 0.001), but no significant differences between reciprocal sites (P = 0.38) or for G×E interaction (P = 0.72) (Fig. 2B; File S5). These results indicate minimal phenotypic plasticity but strong genetic divergence in seed size and germination characteristics.
Figure 2.
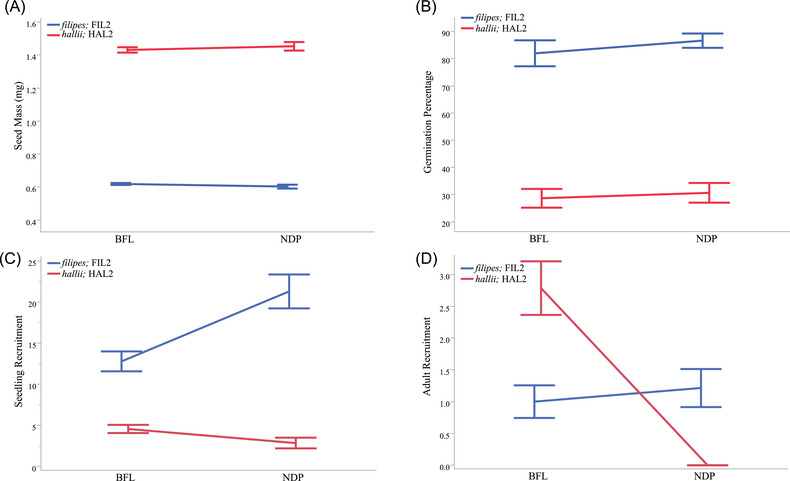
The effect of reciprocal transplantation on seed size, germination percentage, seedling recruitment, and survivorship between the upland, hallii (HAL2, red), and the lowland, filipes (FIL2, blue), ecotypes. Panels A and B show the effect of reciprocal transplantation on seed size and germination percentage between parental lines. The differences between ecotypes were significant but the location does not affect seed mass or germination. Panel C shows seedling recruitment and panel D shows the number of adult plants per plot in reciprocal habitats. The Y‐axis corresponds to the average number of seedling and adult plants per plot and X‐axis provides the location of the experiment (Brackenridge Field Lab [BFL], upland habitat; Nueces Delta Preserve [NDP], lowland habitat). Genotypes are indicated by color with hallii (HAL2, red) and filipes (FIL2, blue).
We also used seed addition plots to test for ecotypic differences in seed germination and seedling establishment. Overall, we observed many more seedlings (fivefold increase) from plots sown with FIL2 seed across both locations. Most recruitment occurred in the spring following the establishment of the experiment, with relatively few new seedlings in the following fall (Fig. 2C). Recruitment to the adult stage was low, especially for parental seed sown in foreign sites. We observed no HAL2 adult recruitment at the NDP (lowland) site and little FIL2 adult recruitment at either site (Fig. 2D). The majority of recruitment occurred for HAL2 seeds planted at the BFL (upland) field site. In factorial models, we detected significant G×E interaction for both seedling (P < 0.0001) and adult recruitment stages (P < 0.0001) (File S6). The form of the G×E suggests trade‐offs in early seedling recruitment favoring FIL2 across habitats, but later adult establishment favoring HAL2 at the upland habitat. Together, these trade‐offs are consistent with a model of adaptive divergence driven in part by early life‐history stages. Our field experiments provide compelling evidence for strong genetic differentiation in seed traits and differential recruitment and survival in home versus foreign habitats.
HERITABILITY AND GENETIC CORRELATIONS AMONG RILS
We estimated broad‐sense heritability (H 2) among RILs in the population as the proportion of total phenotypic variation due to genetic variation among RIL lines. All measured traits were heritable with heritability ranging from 24% to 89% (bootstrap‐based significance, P < 0.001). GP was the least (0.24) and the SM was the highest heritable traits (0.89). The heritability of GT and RL10D was also notably high (0.76 and 0.63, respectively). The estimated heritability for SL5D and SL10D was 0.30 and 0.39, respectively (Table 1).
We observed significant genetic correlation between most of the phenotypes measured in the RIL population. SM was significantly positively correlated to SL5D (r g = 0.15, P = 0.020), SL10D (r g = 0.23, P = 0.001), and RL10D (r g = 0.29, P = 0.001) but negatively correlated with GP (r g = −0.44, P < 0.001). GT was negatively covaried with GP (r g = −0.40, P < 0.001), SL5D (r g = −0.36, P < 0.001), and SL10D (r g = −0.21, P < 0.001). GP was positively correlated with SL5D (r g = 0.34, P < 0.001) and SL10D (r g = 0.17, P = 0.009) but negatively correlated with RL10D (r g = −0.13, P = 0.035). Both shoot length traits (SL5D and SL10D) were also positively correlated (r g = 0.67, P < 0.001) among RILs. In general, RILs with larger seeds initiated robust shoot and root growth but fewer of their seeds germinated over the course of petri dish trials (Fig. 3).
Figure 3.
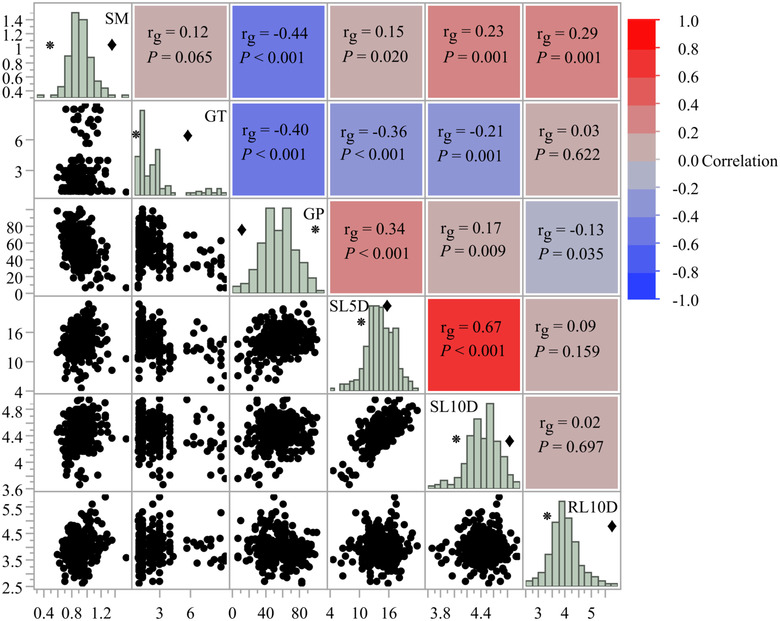
Genetic correlation coefficients (r g) and their pairwise significance (P‐value) for seed mass, germination rate, shoot length, and root length traits with RIL means. Seed mass showed an antagonistic relationship with germination percentage and showed a positive relationship with both shoot length and root length traits. Mean values for the upland genotype, hallii (HAL2), are shown as black diamonds (♦) and the lowland genotype, filipes (FIL2), are shown as the asterisk (٭) in the histograms.
The Fraser v‐test statistics are highly significant (P < 0.01) for all traits except for SL5D (P = 0.57). Overall, the v‐test framework identified phenotypic patterns for all six traits that are suggestive of directional selection underlying seed and seedling trait divergence among P. hallii ecotypes. In all cases, variances of parental trait values were greater than the variances among RILs (Table 2). The most significant traits for directional selection were SM and RL10D (P < 0.0001), followed by GP (P = 0.0003), GT (P = 0.0018), and SL10D (P = 0.016). These results are consistent with strong directional selection pressures underlying the differences among these representative parents.
Table 2.
Selection test statistics on measured six traits. We used the v‐test from Fraser (2020) to test whether selection contributed to the divergence of the six measured traits in a recombinant population
| Traits | Variance Among Parents | Variance Among RILs | V‐Test Statistic | P |
|---|---|---|---|---|
| Seed mass (SM) (mg) | 3.5856422 | 0.021136 | 150.5 | <0.0001 |
| Germination percentage (GP) | 21020.79 | 369.6606 | 13.3 | 0.0003 |
| Shoot length at 5 days (SL5D) (mm) | 12.115722 | 7.636476 | 0.3 | 0.5712 |
| Shoot length at 10 days (SL10D) (mm) | 228.58812 | 11.05885 | 5.9 | 0.0163 |
| Root length at 10 days (RL10D) (mm) | 6517.6475 | 46.32988 | 85.9 | <0.0001 |
| Germination time (GT) (hours) | 46.338645 | 3.454001 | 9.9 | 0.0018 |
GENETIC ARCHITECTURE UNDERLYING SEED SIZE, GERMINATION, AND SEEDLING GROWTH TRAITS
We identified 20 QTL, including one epistatic QTL pair, from all measured traits using stepwise QTL models. SM, GP, and SL5D comprised over half (75%, 15 QTL) of the identified QTL. Most of the QTL (13 out of 20 QTL) had overlapping confidence intervals with a QTL for at least one other trait, suggesting that in many cases phenotypic correlations are reflective of underlying genetic correlations resulting from physical linkage or pleiotropy. About 84% of the identified QTL affected phenotypes in the expected direction based on observed evolutionary divergences. For example, in all cases the HAL2 allele increased trait values relative to FIL2 alleles for SM, SL10D, and RL10D. On the other hand, the FIL2 allele increased trait values for GP. We observed mixed QTL effects only for SL5D, where three of the QTL effects were increased by the HAL2 allele and two were increased by the FIL2 allele. Here, the direction of effects was more complex than expected based on ecotype divergence (Fig. 4). Details about individual QTL, including their location and confidence intervals, effects, and percent of variance explained by each QTL, can be found in File S7.
Figure 4.
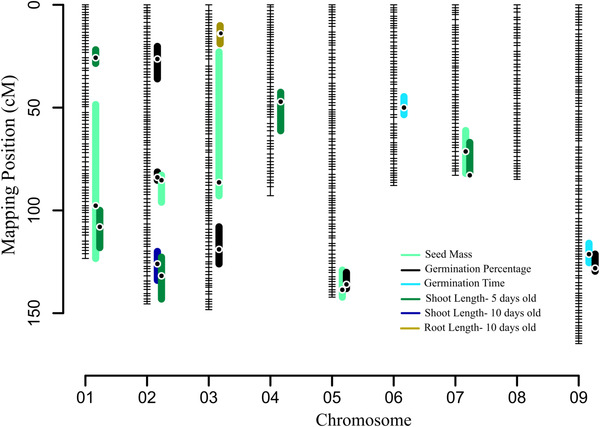
The RIL genetic linkage map of Panicum hallii generated from HAL2 × FIL2 mapping population. All identified QTL are presented as bars to the right of each linkage group. Each colored bar represents a QTL with length representing the 1.5‐LOD drop confidence intervals. Black dots in the colored bars indicate the position of the QTL based on the maximum LOD score.
Only single QTL were identified for SL10D and RL10D. The SL10D QTL overlapped with SL5D at Chr2@126 and the RL10D QTL had a unique position at Chr3@11.7. Despite these traits having relatively high heritability (Table 1), we only detected single QTL with a relatively low percent of variance explained for these traits (<10%), suggesting there may be many undetected loci controlling these traits or that our heritability estimates are inflated by maternal environmental or epigenetic effects.
QTL COLOCALIZATION, ALLELIC EFFECTS, GENETIC TRADE‐OFFS, AND EPISTASIS
Overall, we mapped 13 overlapping QTL regions. Colocalization of QTL could be indicative of loci with pleiotropic effects or genes in tight linkage that affect unique traits. We identified interesting and potentially functionally related combinations of QTL colocalization. For example, GP and GT trait QTL colocalized at Chr9@128 with antagonistic effects in the same direction as parental divergence. Here, the FIL2 allele increased trait values for GP but decreased values for GT. In another case, SM and GP shared two colocalized regions, one was at Chr2@85.3 and the other one was at Chr5@130. In both cases, the HAL2 allele increased trait values for SM and decreased GP trait values as expected from the observed evolutionary divergence between parental lines. We also observed QTL colocalization with mixed allelic recombination for same trait combination. SM and SL5D colocalized at two chromosomal regions. In the first colocalized regions at Chr1@108, HAL2 allele increased trait values for SM and decreased trait values for SL5D. But in the second region at Chr7@70, HAL2 allele increased traits values for both traits. We also found some expected traits colocalization. For example, shoot length measured at two time points, SL10D and SL5D, colocalized at Chr2@126 having additive effects in the same direction of parental divergence where the HAL2 allele increased trait values for both traits (File S7).
Only 25% of identified QTL (five out of 20) had unique position in the linkage map. From the trait measured, SL5D had two unique QTL (1@25.7 and 4@47.1). GP and GT had a single unique QTL occurring at 2@26.3 and 6@50, respectively (Fig. 4; File S7).
Finally, we detected one pairwise epistatic interaction for GT occurring between QTL on Chr6 and Chr9 (Fig. 5). Here, individuals that are homozygous for the HAL2 allele at both loci have delayed germination, but this pattern is masked in any of the alternative hybrid combinations.
Figure 5.
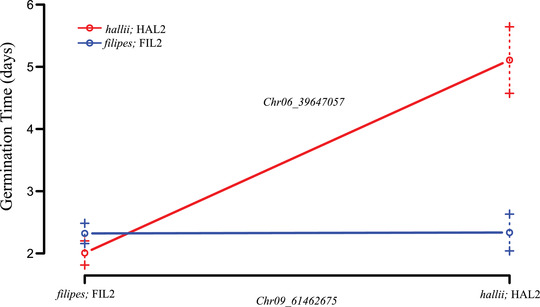
Pairwise epistatic QTL for the day of first germination (GT) between Chromosome 6 and 9 in the Panicum hallii RIL population. Plotted points indicate two‐locus genotype means ± 1 SE for the two loci containing GT between Chr6 and Chr9.
Discussion
Populations adapting to heterogeneous environments will develop genetic differences over time in response to selection. This adaptation may ultimately lead to divergence and ecotype formation with distinct morphological and/or physiological characteristics that provide an environment‐specific fitness advantage. In this study, we examined ecotypic divergence between xeric and mesic ecotypes at the seed and seedling stages in P. hallii to evaluate early life stages that may contribute to local adaptation. Field experiments reveal little plasticity but support a hypothesis of ecotypic adaptation driven by trade‐offs in seedling recruitment and adult establishment. We mapped QTL for six traits with several large effect QTL colocalizing to a relatively small number of genomic regions. Overall, the direction of QTL effects is predominantly in the direction of putative adaptive divergence. This pattern is consistent with a hypothesis of strong directional selection on seed‐related traits (Orr 1998; Rieseberg et al. 2002) as indicated by significant v‐tests (Fraser 2020). We also identified clear genetic trade‐offs between seed mass and germination percentage at two QTL based on QTL localization. These data suggest that seed size, dormancy, and seedling vigor traits may have jointly evolved in the process of ecotype divergence.
PARENTAL DIVERGENCE AND TRAIT PLASTICITY
Plant populations are generally limited by either the abundance and quality of seed for seedling recruitment or by the availability of environmental microsites for seedling establishment. One hypothesis is that aspects of habitat quality and the local competitive environments may therefore play key selective roles in the evolution of seed and seedling phenotypes. Considering this theory, it is interesting to speculate on the patterns of ecotypic divergence in P. hallii. Panicum hallii var. hallii is a widespread ecotype and occurs primarily in xeric habitats in rocky, dry, and calcareous soils (Smeins et al. 1976; Waller 1976; Hatch et al. 1999) and with relatively low ground cover or competition. In contrast var. filipes is located primarily in mesic and seasonal wet areas (Gould 1975; Waller 1976) in dense coastal prairie habitats. Using representative inbred lines, our data show that hallii (HAL2) produces larger seeds with delayed germination and strong dormancy compared to filipes (FIL2). In addition, HAL2 roots grew 73% longer than FIL2 roots after 10 days of seedling growth (Fig. 1; Table 1). Typically, increased root growth from large seeds would result in an establishment benefit in resource poor sites. Thus, having larger seed mass, high dormancy, delayed germination time, and faster root growth may be selected for HAL2 to survive hazards of establishment in harsh southwestern xeric habitats. Reciprocal plantings of ecotypes revealed that local conditions have little impact on seed mass or germination percentage. Our seed addition experiment indicates that the lowland ecotype (filipes; FIL2) is most likely limited by seed or microsite availability in general, whereas the upland ecotype (hallii; HAL2) exhibited more striking habitat dependent establishment (Fig. 2; Files S5 and S6).
Our results support the model developed for maintenance of seed size diversity by Muller‐Landau (2010), where species coexist in heterogenous habitats by a tolerance‐fecundity trade‐off. Under this mechanism, species with larger seeds win in a stressful environment (e.g., too dry, too shady, etc.) due to their higher tolerance to stress, whereas small‐seeded species win in competitive habitats due to their higher fecundity. We find filipes ecotype commonly in competitive coastal prairie environments with a high density of other grasses (e.g., Bouteloua rigidiseta and Nassella leucotricha). The upland strategy centers on producing fewer larger seeds with high dormancy, and the lowland strategy centers on producing many small seeds with low dormancy. Larger seed mass in the upland ecotype is probably under strong selection to improve establishment in dry and calcareous soil, and seed number is possibly under strong selection in the lowland ecotype as a mechanism to provide opportunities for establishment. Here, selection may have favored the production of a large number of small seeds with attributes allowing establishment in rare disturbed patches of habitat. Future field studies exploring the impact of competitive vegetation, disturbance, or seasonality on seedling establishment will help better reveal possible mechanisms of selection on seed traits in P. hallii.
THE GENETICS OF ECOTYPIC DIVERGENCE
Ecotypic divergence caused by adaptation to xeric and mesic habitats provides ample evidence of ecological functions that contribute to the process of speciation (Clausen 1951; Kruckeberg 1986; Rajakaruna 2004; Lowry 2012; Lowry et al. 2015). Understanding the underlying genetic basis of ecotypic divergence helps to better understand the degree to which adaptive evolution was constrained or facilitated by the structure of genetic variation. In this study, we found colocalization of QTL for a common set of traits involved in xeric and mesic ecotypic divergence, suggesting that these traits may jointly evolve. We observed that seed mass QTL colocalized with all other measured traits, suggesting that in many cases seed mass, seed investment, and earlier growth‐related traits may function as an integrated suite of functional traits.
We also found evidence for genetic trade‐off between seed‐based life‐history traits. Most of the discovered QTL clusters in this study had antagonistic effects across traits. The antagonistic additive effects for QTL clusters implies that seed‐based related traits are under strong selection between P. hallii lowland and upland ecotypes. For example, seed mass and germination percentage colocalized at two chromosomal regions (Chromosomes 2 and 5) where the HAL2 allele increased seed mass but decreased germination percentage. Our result suggests that P. hallii ecotypes may have experienced strong selection pressures to evolve either large seed/strong dormancy or small seed/weak dormancy trait combinations. Additionally, GP and GT colocalized at Chr9 where the HAL2 allele decreased GP but increased GT. The observed genetic covariation in the RIL population suggests that the major axis of standing genetic variation would support the rapid evolution of the observed ecotypic divergence in seed traits, while potentially constraining evolution of the opposite trait combinations. In addition, large seeds with vigorous root growth might be favored in dryer and resource‐limited habitats (Fig. 3). The Fraser v‐test also supports this inference; the highly significant P‐values are consistent with directional selection for these trait combinations while adapted to lowland and upland habitats. Thus, our mapping data support the idea that large seeds/dormancy and longer root growth trait combination may have been an important adaptation for hallii (HAL2) as it diverged from a coastal ancestor and invaded dry, calcareous habitats across the southwest.
We have consistently found that upland and lowland ecotype differentiating traits colocalized in P. hallii to a relatively small number of chromosomal regions. Lowry et al. (2015) mapped five QTL to a locus on chromosome 5 with traits that are involved in ecotypic divergence using an earlier F2 population. These traits are mainly associated with flowering time, tiller characters, and leaf width. In our study, two traits (seed mass and germination percentage) colocalized to this region. Interestingly, all five traits that Lowry et al. (2015) mapped had additive effects in the same direction where the FIL2 allele increased the trait values. Khasanova et al. (2019) also mapped five shoot and root traits to the same region. In that study, all traits also had additive effects in the same direction with the FIL2 allele increasing trait values. In our study, this QTL had contrasting effects on seed size and germination, in that the HAL2 allele increased seed mass trait values and decreased germination percentage. Our study therefore provides further support for a major pleiotropic region that contributes to ecotype differentiation in P. hallii.
Conclusions
Seed biology plays an important role in plant ecology. It tells us how plant species invest resources into reproduction, and how natural selection has given rise to an incredible diversity of reproductive strategies. In this study, we explored several facets of ecotype differentiating traits from a seed biology perspective. Our study reinforces a pattern that has been observed in multiple species, wherein genetic studies find that ecotype differentiating traits colocalize to common genomic regions (Hall et al. 2006; Latta and Gardner 2009; Lowry and Willis 2010; Lovell et al. 2013; Lowry et al. 2015; Khasanova et al. 2019). Once fine mapping or transformation becomes feasible in this species, further work can identify candidate genes to gain better insight into the molecular details underlying ecotypic divergence in P. hallii. Overall, our data support the functional integration of seed size and seed dormancy traits, and an exciting next step will be to evaluate the fitness and population impact of the seed variation in different habitats.
AUTHOR CONTRIBUTIONS
SR and TEJ designed the experiment. SR conducted the experiment, analyzed the data, and wrote the manuscript. TEJ provided feedback on analyses and edited the manuscript.
CONFLICT OF INTEREST
The authors declare no conflict of interest.
Supporting information
Supplementary file S1
Supplementary file S2
Supplementary file S3
Supplementary file S4
Supplementary file S5
Supplementary file S6
Supplementary file S7
ACKNOWLEDGMENTS
The authors thank B. Campitelli, A. Hutt, and S. N. Bhathena for helping during seed collection. J. D. Palacio Mejía helped with the initial setup of the experiment. R. Heckman gave important comments and valuable insights during the work, analyses, and manuscript writing. A. MacQueen provided feedback on the manuscript. The authors thank J. Herring, Director of Land Conservation, from Coastal Bend Bays and Estuaries Program for allowing to conduct field plantings at the Nueces Delta Preserve, Odem Texas. R. Plowes provided space at the Brackenridge Field Lab site at UT Austin. S. Merrell provided technical support while setting up the experiment in the greenhouse facility. This research was supported by a National Science Foundation Plant Genome Research Program Grant (IOS‐0922457) to TEJ. The authors also acknowledge Texas Ecolab support to SR for conducting field work.
Contributor Information
Samsad Razzaque, Email: Samsad.razzaque@utexas.edu.
Thomas E. Juenger, Email: tjuenger@austin.utexas.edu.
LITERATURE CITED
- Agrawal, A. A. , Conner J. K., and Rasmann S.. 2010. Tradeoffs and negative correlations in evolutionary ecology. Pp. 243–268 in Bell M. A., Eanes W. F., Futuyma D. J., and Levinton J. S., eds. Evolution since Darwin: the first 150 years. Sinauer Associates, Sunderland, MA. [Google Scholar]
- Baker, H. G. 1972. Seed weight in relation to environmental conditions in California. Ecology. 53:997–1010. [Google Scholar]
- Baskin, C. C. , and Baskin J. M.. 1998. Seeds: ecology, biogeography, and evolution of dormancy and germination. Elsevier, Amsterdam, The Netherlands. [Google Scholar]
- Beckerman, A. , Benton T. G., Ranta E., Kaitala V., and Lundberg P.. 2002. Population dynamic consequences of delayed life‐history effects. Trends Ecol. Evol. 17:263–269. [Google Scholar]
- Benjamini, Y. , and Hochberg Y.. 1995. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. B Stat. Methodol. 57:289–300. [Google Scholar]
- Bentsink L., Hanson J., Hanhart C. J., Blankestijn‐de Vries H., Coltrane C., Keizer P., et al. 2010. Natural variation for seed dormancy in Arabidopsis is regulated by additive genetic and molecular pathways. Proceedings of the National Academy of Sciences. 107:4264–4269 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bettey M., Finch‐Savage W., King G. J., and Lynn J.. 2000. Quantitative genetic analysis of seed vigour and pre‐emergence seedling growth traits in Brassica oleracea. New Phytologist. 148:277–286 [Google Scholar]
- Bradford K., and Nonogaki H.. 2008. Annual plant reviews, seed development, dormancy and germination, vol 27. John Wiley & Sons, [Google Scholar]
- Broman K. W., Wu H., Sen Ś., and Churchill G. A.. 2003. R/qtl: QTL mapping in experimental crosses. Bioinformatics. 19:889–890 [DOI] [PubMed] [Google Scholar]
- Childs D. Z., Metcalf C. J. E., and Rees M.. 2010. Evolutionary bet‐hedging in the real world: empirical evidence and challenges revealed by plants. Proceedings of the Royal Society B: Biological Sciences. 277: 3055–3064 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clausen, J. 1951. Stages in the evolution of plant species. Cornell Univ. Press, Ithaca, NY. [Google Scholar]
- Clauss M., and Aarssen L.. 1994. Phenotypic plasticity of size–fecundity relationships in Arabidopsis thaliana . Journal of Ecology. 82:447–455 [Google Scholar]
- Conover D. O., Duffy T. A., and Hice L. A.. 2009. The covariance between genetic and environmental influences across ecological gradients. Annals of the New York Academy of Sciences. 1168:100–129 [DOI] [PubMed] [Google Scholar]
- Davis M. B., and Shaw R. G.. 2001. Range shifts and adaptive responses to quaternary climate change. Science (New York, N.Y.). 292:673–679 [DOI] [PubMed] [Google Scholar]
- Daws M. I., Ballard C., Mullins C. E., Garwood N. C., Murray B., Pearson T. R., et al. 2007. Allometric relationships between seed mass and seedling characteristics reveal trade‐offs for neotropical gap‐dependent species. Oecologia. 154:445–454 [DOI] [PubMed] [Google Scholar]
- Dechaine J. M., Brock M. T., and Weinig C.. 2014. QTL architecture of reproductive fitness characters in Brassica rapa . BMC Plant Biology. 14:66 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dias P. M. B., Brunel‐Muguet S., Dürr C., Huguet T., Demilly D., Wagner M.‐H., et al. 2011) QTL analysis of seed germination and pre‐emergence growth at extreme temperatures in Medicago truncatula . Theoretical and Applied Genetics. 122:429–444 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donohue K. 2014. Why ontogeny matters during adaptation: developmental niche construction and pleiotorpy across the life cycle in Arabidopsis thaliana . Evolution; International Journal of Organic Evolution. 68:32–47 [DOI] [PubMed] [Google Scholar]
- Donohue K., Rubio de Casas R., Burghardt L., Kovach K., and Willis C. G.. 2010. Germination, postgermination adaptation, and species ecological ranges. Annual Review of Ecology, Evolution, and Systematics. 41:293–319 [Google Scholar]
- Fournier‐Level A., Wilczek A. M., Cooper M. D., Roe J. L., Anderson J., Eaton D., et al. 2013. Paths to selection on life history loci in different natural environments across the native range of Arabidopsis thaliana . Molecular Ecology. 22:3552–3566 [DOI] [PubMed] [Google Scholar]
- Fraser H. B. 2020. Detecting selection with a genetic cross. Proceedings of the National Academy of Sciences. 117:22323–22330 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Futuyma D. J., and Moreno G.. 1988. The evolution of ecological specialization. Annual review of ecology and systematics. 19:207–233 [Google Scholar]
- Geshnizjani N., Snoek B. L., Willems L. A., Rienstra J. A., Nijveen H., Hilhorst H. W., et al. 2020. Detection of QTLs for genotype × environment interactions in tomato seeds and seedlings. Plant, Cell & Environment. 43:1973–1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gould B. A., Palacio‐Mejia J. D., Jenkins J., Mamidi S., Barry K., Schmutz J., et al. 2018. Population genomics and climate adaptation of a C4 perennial grass, Panicum hallii (Poaceae). BMC Genomics [Electronic Resource]. 19:792 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gould F. 1975. The grasses of Texas. Texas A&M Univ. Press, College Station, TX. [Google Scholar]
- Grime J. P., Hodgson J. G., and Hunt R.. 2014. Comparative plant ecology: a functional approach to common british species. Springer, Berlin. [Google Scholar]
- Hall M. C., Basten C. J., and Willis J. H.. 2006. Pleiotropic quantitative trait loci contribute to population divergence in traits associated with life‐history variation in Mimulus guttatus. Genetics. 172:1829–1844 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harel D., Holzapfel C., and Sternberg M.. 2011. Seed mass and dormancy of annual plant populations and communities decreases with aridity and rainfall predictability. Basic and Applied Ecology. 12:674–684 [Google Scholar]
- Hatch S. L., Schuster J. L., and Drawe D. L.. 1999. Grasses of the Texas Gulf prairies and marshes. vol 24. Texas A&M Univ. Press, College Station, TX. [Google Scholar]
- Jasmin J. N., and Kassen R.. 2007. On the experimental evolution of specialization and diversity in heterogeneous environments. Ecology Letters. 10:272–281 [DOI] [PubMed] [Google Scholar]
- Juenger T. E. 2013. Natural variation and genetic constraints on drought tolerance. Current opinion in Plant Biology. 16:274–281 [DOI] [PubMed] [Google Scholar]
- Jurado E., and Westoby M.. 1992. Seedling growth in relation to seed size among species of arid Australia. Journal of Ecology. 80:407–416 [Google Scholar]
- Kawecki T. J., and Ebert D.. 2004. Conceptual issues in local adaptation. Ecology Letters. 7:1225–1241 [Google Scholar]
- Khan N., Kazmi R. H., Willems L. A., Van Heusden A. W., Ligterink W., and Hilhorst H. W.. 2012. Exploring the natural variation for seedling traits and their link with seed dimensions in tomato. Plos One. 7:e43991 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khasanova A., Lovell J. T., Bonnette J., Weng X., Jenkins J., Yoshinaga Y., et al. 2019. The genetic architecture of shoot and root trait divergence between mesic and xeric ecotypes of a perennial grass. Frontiers in Plant Science. 10:366 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kitajima K., and Fenner M.. 2000. Ecology of seedling regeneration. Seeds: the Ecology of Regeneration in Plant Communities. 2:331–359 [Google Scholar]
- Kruckeberg A. R. 1986. An essay: the stimulus of unusual geologies for plant speciation. Systematic Botany. 455–463 [Google Scholar]
- Lacerda D. R., Lemos Filho J. P., Goulart M. F., Ribeiro R. A., and Lovato M. B.. 2004. Seed‐dormancy variation in natural populations of two tropical leguminous tree species: Senna multijuga (Caesalpinoideae) and Plathymenia reticulata (Mimosoideae). Seed Science Research. 14:127–135 [Google Scholar]
- Lascoux M., Glémin S., and Savolainen O.. 2016. Local adaptation in plants. Pp. 1–7 in eLS. John Wiley, Chichester, U.K. [Google Scholar]
- Latta R. G., and Gardner K. M.. 2009. Natural selection on pleiotropic quantitative trait loci affecting a life‐history trade‐off in Avena barbata . Evolution: International Journal of Organic Evolution. 63:2153–2163 [DOI] [PubMed] [Google Scholar]
- Leishman M. R., and Westoby M.. 1992. Classifying plants into groups on the basis of associations of individual traits–evidence from Australian semi‐arid woodlands. Journal of Ecology. 80:417–424 [Google Scholar]
- Lloret F., Casanovas C., and Peñuelas J.. 1999. Seedling survival of Mediterranean shrubland species in relation to root: shoot ratio, seed size and water and nitrogen use. Functional Ecology. 13:210–216 [Google Scholar]
- Lovell J. T., Jenkins J., Lowry D. B., Mamidi S., Sreedasyam A., Weng X., et al. 2018a. The genomic landscape of molecular responses to natural drought stress in Panicum hallii . Nature Communications. 9:5213 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lovell J. T., Jenkins J., Lowry D. B., Mamidi S., Sreedasyam A., Weng X., et al. 2018b. The genomic landscape of molecular responses to natural drought stress in Panicum hallii. Nature Communications. 9:1–10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lovell J. T., Juenger T. E., Michaels S. D., Lasky J. R., Platt A., Richards J. H., et al. 2013. Pleiotropy of FRIGIDA enhances the potential for multivariate adaptation. Proceedings of the Royal Society B: Biological Sciences. 280: 20131043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lowry D. B. 2012. Ecotypes and the controversy over stages in the formation of new species. Biological Journal of the Linnean Society. 106:241–257 [Google Scholar]
- Lowry D. B., Hernandez K., Taylor S. H., Meyer E., Logan T. L., Barry K. W., et al. 2015. The genetics of divergence and reproductive isolation between ecotypes of Panicum hallii. New Phytologist. 205:402–414 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lowry D. B., Purmal C. T., and Juenger T. E.. 2013. A population genetic transect of Panicum hallii (Poaceae). American Journal of Botany. 100:592–601 [DOI] [PubMed] [Google Scholar]
- Lowry D. B., and Willis J. H.. 2010. A widespread chromosomal inversion polymorphism contributes to a major life‐history transition, local adaptation, and reproductive isolation. Plos Biology. 8:e1000500 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mojonnier L. 1998. Natural selection on two seed‐size traits in the common Morning Glory Ipomoea purpurea (Convolvulaceae): patterns and evolutionary consequences. The American Naturalist. 152:188–203 [DOI] [PubMed] [Google Scholar]
- Muller‐Landau H. C. 2010. The tolerance–fecundity trade‐off and the maintenance of diversity in seed size. Proceedings of the National Academy of Sciences. 107:4242–4247 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orr H. A. 1998. Testing natural selection vs. genetic drift in phenotypic evolution using quantitative trait locus data. Genetics. 149:2099–2104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palacio‐Mejía J. D., Grabowski P. P., Ortiz E. M., Silva‐Arias G. A., Haque T., Des Marais D. L., et al. 2021a. Geographic patterns of genomic diversity and structure in the C4 grass Panicum hallii across its natural distribution. AoB Plants. 13:plab002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palacio‐Mejía J. D., Grabowski P. P., Ortiz E. M., Silva‐Arias G. A., Haque T., Des Marais D. L., et al. 2021b. Geographic patterns of genomic diversity and structure in the C4 grass Panicum hallii across its natural distribution. AoB Plants. 13:plab002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phillips P. C., and Arnold S. J.. 1999. Hierarchical comparison of genetic variance‐covariance matrices. I. Using the Flury hierarchy. Evolution; international journal of organic evolution. 53:1506–1515 [DOI] [PubMed] [Google Scholar]
- Postma F. M., and Ågren J.. 2016. Early life stages contribute strongly to local adaptation in Arabidopsis thaliana . Proceedings of the National Academy of Sciences. 113:7590–7595 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Primack R. B., and Antonovics J.. 1981. Experimental ecological genetics in Plantago. V. Components of seed yield in the ribwort plantain Plantago lanceolata L. Evolution; international journal of organic evolution. 35:1069–1079 [DOI] [PubMed] [Google Scholar]
- Rajakaruna N. 2004. The edaphic factor in the origin of plant species. International Geology Review. 46:471–478 [Google Scholar]
- Rees M. 1996. Evolutionary ecology of seed dormancy and seed size. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences. 351:1299–1308 [Google Scholar]
- Rieseberg L. H., Widmer A., Arntz A. M., and Burke J. M.. 2002. Directional selection is the primary cause of phenotypic diversification. Proceedings of the National Academy of Sciences. 99:12242–12245 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roff D. A. 2002. Life history evolution. Sinauer Associates, Sunderland, MA: [Google Scholar]
- Roux F., Touzet P., Cuguen J., and Le Corre V.. 2006. How to be early flowering: an evolutionary perspective. Trends in Plant Science. 11:375–381 [DOI] [PubMed] [Google Scholar]
- Rundle H. D., and Nosil P.. 2005. Ecological speciation. Ecology Letters. 8:336–352 [Google Scholar]
- Shipley B., Keddy P., Moore D., and Lemky K.. 1989. Regeneration and establishment strategies of emergent macrophytes. The Journal of Ecology. 77:1093–1110 [Google Scholar]
- Silvertown J. 1989. The paradox of seed size and adaptation. Trends in Ecology & Evolution. 4:24–26 [DOI] [PubMed] [Google Scholar]
- Smeins F. E., Taylor T. W., and Merrill L. B.. 1976. Vegetation of a 25‐year exclosure on the Edwards Plateau, Texas. Rangeland Ecology & Management/Journal of Range Management Archives. 29:24–29 [Google Scholar]
- Smith C. C., and Fretwell S. D.. 1974. The optimal balance between size and number of offspring. The American Naturalist. 108:499–506 [Google Scholar]
- Stanton M. L. 1984. Developmental and genetic sources of seed weight variation in Raphanus raphanistrum L.(Brassicaceae). American Journal of Botany. 71:1090–1098 [Google Scholar]
- Stebbins G. L. Jr 1952. Aridity as a stimulus to plant evolution. The American Naturalist. 86:33–44 [Google Scholar]
- Venable D. L. 1992. Size‐number trade‐offs and the variation of seed size with plant resource status. The American Naturalist. 140:287–304 [Google Scholar]
- Venable D. L. 2007. Bet hedging in a guild of desert annuals. Ecology. 88:1086–1090 [DOI] [PubMed] [Google Scholar]
- Waller F. R. 1976. A biosystematic study of Panicum section Diffusa (Poaceae) in North America. Ph.D. thesis, Texas A&M University, College Station, TX. [Google Scholar]
- Westoby M., and Rice B.. 1982. Evolution of the seed plants and inclusive fitness of plant tissues. Evolution; International Journal of Organic Evolution. 36:713–724 [DOI] [PubMed] [Google Scholar]
- Whittaker R. H. 1970. Communities and ecosystems. Macmillan, New York. [Google Scholar]
- Woodward F. I., Lomas M. R., and Kelly C. K.. 2004. Global climate and the distribution of plant biomes. Philosophical Transactions of the Royal Society of London Series B: Biological Sciences. 359:1465–1476 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wulff R. D. 1986. Seed size variation in Desmodium paniculatum: II. Effects on seedling growth and physiological performance. The Journal of Ecology. 74:99–114 [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary file S1
Supplementary file S2
Supplementary file S3
Supplementary file S4
Supplementary file S5
Supplementary file S6
Supplementary file S7