

Abstract
Background
Autism spectrum disorder (autism) is a complex neurodevelopmental condition with pronounced behavioral, cognitive, and neural heterogeneities across individuals. Here, our goal was to characterize heterogeneity in autism by identifying patterns of neural diversity as reflected in BOLD fMRI in the way individuals with autism engage with a varied array of cognitive tasks.
Methods
All analyses were based on the EU-AIMS/AIMS-2-TRIALS multisite Longitudinal European Autism Project (LEAP) with participants with autism (n = 282) and typically developing (TD) controls (n = 221) between 6 and 30 years of age. We employed a novel task potency approach which combines the unique aspects of both resting state fMRI and task-fMRI to quantify task-induced variations in the functional connectome. Normative modelling was used to map atypicality of features on an individual basis with respect to their distribution in neurotypical control participants. We applied robust out-of-sample canonical correlation analysis (CCA) to relate connectome data to behavioral data.
Results
Deviation from the normative ranges of global functional connectivity was greater for individuals with autism compared to TD in each fMRI task paradigm (all tasks p < 0.001). The similarity across individuals of the deviation pattern was significantly increased in autistic relative to TD individuals (p < 0.002). The CCA identified significant and robust brain-behavior covariation between functional connectivity atypicality and autism-related behavioral features.
Conclusions
Individuals with autism engage with tasks in a globally atypical way, but the particular spatial pattern of this atypicality is nevertheless similar across tasks. Atypicalities in the tasks originate mostly from prefrontal cortex and default mode network regions, but also speech and auditory networks. We show how sophisticated modeling methods such as task potency and normative modeling can be used toward unravelling complex heterogeneous conditions like autism.
Supplementary Information
The online version contains supplementary material available at 10.1186/s13229-022-00529-y.
Keywords: Autism, fMRI, Functional connectivity, Normative modeling, Heterogeneity, Canonical correlation analysis
Introduction
Autism spectrum disorder (henceforth ‘autism’) is a complex neurodevelopmental condition marked by difficulties with social communication, repetitive, restricted behaviors and interests and sensory processing atypicalities [1]. Cross-participant heterogeneity in autism has made understanding underlying mechanisms and the complex interrelation between neurobiology and cognitive profiles in autism challenging. Imaging studies in autism report both over-and under-connectivity of functional brain networks [2–4] on the basis of resting state fMRI data. Different task-fMRI studies, probing a variety of neural processes, report between-group differences with small effect size at best [5–7]. Crucially, little effort has been made to integrate the diverse findings both across different cognitive domains and between task-fMRI and resting state connectivity at the level of an individual participant. In order to better characterize heterogeneity both across cognitive domains and across individuals, we combine novel methodological approaches.
First, we propose an integrated analytical approach to characterize the task-specific cognitive demands in autistic individuals. We utilize a task potency approach which combines the unique aspects of both resting state fMRI (rs-fMRI) and task-fMRI [8]. Rs-fMRI provides insight into the large-scale ‘architecture’ of brain connectivity in an individual. Task-fMRI might however more directly probe the neural correlates of specific cognitive domains affected by the condition such as social/emotional processing and attention. We leverage the advantages offered by both views in task potency, which disentangles the relative contribution of task-induced functional connectivity from that of the baseline architecture at the individual level [9]. This allows for greater sensitivity to individual task-based functional connectivity (FC) effects as well as a more precise interpretation of findings as being related specifically to the cognitive load and not to differences in baseline.
Second, even though many cognitive/behavioral studies have been able to successfully show differences between individuals with autism and typically developing individuals across a range of cognitive domains such as social cognition, reward and emotion processing, and executive functioning [10], there appears to exist pronounced behavioral, cognitive, and neural heterogeneity across individuals with autism [11–13]. In order to parse the heterogeneous nature of autism neurobiology, we therefore apply normative modeling which will allow us to map atypicality of brain-derived features on an individual basis with respect to the distribution of those features in a group of similar typically developing controls. [14]. This approach has previously been applied in autism and yielded promising results [15–17]. This way, the analysis becomes more sensitive to idiosyncratic brain atypicalities.
In order to be able to characterize diversity in presentation across cognitive domains and individuals, we leverage the large-scale resource that has specifically been designed to capture a large, heterogenous and thus naturalistic autism sample—the EU-AIMS/AIMS-2-TRIALS Longitudinal European Autism Project (LEAP) [18–20]. It is designed to provide deep-phenotyping by including both various neuroimaging measures (such as rs-fMRI and task-fMRI), an extensive cognitive battery capturing social cognition, reward and emotion processing, and executive functioning and in-depth behavioral phenotyping. Due to the presence of multiple task paradigms in the dataset, we are able to contrast the spatial patterns of atypicality across these different tasks. We are especially interested in finding out whether posited patterns of task-specific functional connectome atypicality in autism are similar across cognitive domains—and conversely, how that similarity is expressed in typically developing controls.
We assess whether the patterns of brain atypicality we find in individuals relate to metrics of autism at a behavioral level—thereby assessing for each task whether task-specific functional connectome atypicality carries information relevant to finding brain-behavior relationships in autism. In order to relate high-dimensional brain data to behavioral data in a way without making prior assumptions on the most relevant features in a multivariate context, we will apply canonical correlation analysis (CCA) [21, 22].
By integrating complementary functional modalities and combining the aforementioned novel tools such as task potency and normative modelling, taking a unique look at identifying (a)typicality in the way individuals with autism engage with cognitive demands across tasks at the individual neural level is made possible. The multiple fMRI tasks present in the dataset enable a crucial cross-task perspective toward gauging to what extent such atypicalities exist across different cognitive loads, and whether this pattern is different in autism from typically developing controls.
Materials and methods
Sample
The dataset from the EU-AIMS/AIMS2TRIALS LEAP project was used for the current analyses—a large multi-center European project with an aim to identify biomarkers in [18–20]. Local ethical committees approved the study at their respective sites. Participant was extensively clinically phenotyped and underwent multiple MRI assessments. Data from participants with intellectual disability (IQ < 75) were not included in the current project. Furthermore, we removed participants from the analysis on the basis of data quality using the following criteria: Participants were required to have acceptable overlap (> 94%) with the MNI152 standard brain after image registration. We then excluded 57 participants due to poor overlap (< 50%) with one or more particular regions from the ICP brain parcellation atlas. In general, we followed the same rationale for brain region inclusion as used in [23]. Participants were furthermore excluded on the basis of incidental findings and incomplete scans (N = 18), and those in the top 5% in terms of head motion quantified through root mean square framewise displacement (N = 27) [24]. The above criteria resulted in the inclusion of data for analyses from the following participants: 282 participants with autism (age range 7.5–30.3 years; mean = 17.1; sd = 5.4; 72.3% male), and 221 typically developing controls (age range 6.9–29.8 years; mean = 17.0; sd = 5.5; 63.8% male). Not every participant was included in each analysis, for the reason that not all participants completed all fMRI scan sessions. See Additional file 1: Tables S1 through S5 for more details on the subsample characterization in each task.
Behavioral data
Total scores on seven behavioral variables were included in the multivariate canonical correlation analyses with the aim to broadly include information about the affected autism domains, co-occurring attention deficit hyperactivity disorder (ADHD), and adaptive functioning, as well as IQ. Three variables cover the primary affected domains in autism (social and communicative difficulties, repetitive/restricted behaviors and interests, and sensory atypicalities): Social Responsiveness Scale-2 (SRS) [25]—a quantitative scale of autism symptomatology over the past 6 months, the Repetitive Behaviors Scale-revised (RBS) [26]—a scale which assesses more specifically restricted and repetitive behaviors, and the Short Sensory Profile (SSP) [27]—a scale which assesses sensory processing atypicalities. Two further variables cover the ADHD-related behaviors: ADHD hyperactivity/impulsivity, and ADHD inattentiveness. These are the two components from the DSM-5 ADHD rating scale for behavior in the past 6 months. Furthermore, we included the Vineland-II Adaptive Behavior Composite (Vineland) [28] which assesses the level of real-life everyday adaptive functioning. Finally, we included Full scale intelligence quotient (IQ) as measured by the Wechsler Abbreviated scale intelligence-2 [29]. Missing data were imputed with random forest regression, as done in [17].
fMRI data
Participants performed a resting state fMRI (rs-fMRI) lasting approximately 6 min, and one or more of the following task-fMRI scans: Hariri emotion processing (Hariri) [30], Flanker and Go-NoGo (Flanker) [31], social reward anticipation (Reward_s), nonsocial reward anticipation (Reward_ns) [32], and animated shapes theory of mind (ToM) [33, 34]. See Additional file 1 for brief descriptions of these tasks. Participants were instructed to relax and fixate on a cross presented on the screen for the duration of the rs-fMRI scan. Additionally, each participant completed an anatomical scan for the purpose of registration. MRI data were acquired on 3T scanners at multiple sites in Europe—King’s College London (KCL), Radboud University Nijmegen Medical Centre (RUNMC), University Medical Centre Utrecht (UMCU), Autism Research Centre (ARC), University of Cambridge (UCAM), Central Institute of Mental Health, Mannheim (CIMH), and Karolinska Institutet (KI). Data from KI were not used due to a low number of participants. fMRI parameters are described in the supplementary information.
fMRI preprocessing
Preprocessing of both the resting state- and task-fMRI data was performed with tools from FSL [35]. The first five volumes for each acquisition were removed to allow for equilibration of the magnetization. To correct for head movement, we performed volume realignment to the middle volume using MCFLIRT. Next, global 4D mean intensity normalization and smoothing with a 6 mm FWHM kernel were applied. ICA-AROMA was used to identify and remove secondary motion-related artifacts [36, 37]. Next, signal from white matter and cerebrospinal fluid was regressed out and we applied a 0.01 Hz high-pass filter. For each participant, we registered acquisitions to their respective high-resolution T1 anatomical images by means of the Boundary-Based Registration tool from FSL-FLIRT [24]. The high-resolution T1 image belonging to each participant was registered to MNI152 space with FLIRT 12-degrees of freedom linear registration, and further refined using FNIRT nonlinear registration [38]. We used the inverse of these transformations to take a brain atlas to the native space of each participant, where all further analyses were performed. ComBat was used to clean the data for linear site effects [39]. This resulted in small changes to the functional connectivity matrices, see Additional file 1: Fig. S4A-E.
Task potency
We used the instantaneous connectivity parcellation (ICP) brain atlas with 168 brain regions (hierarchically situated in 11 larger-scale ’networks’) [40] to define brain regions in the native space of each subject. The ICP parcellation was chosen for its suitability to FC modeling and due to its hierarchical nature. This was done for each of the five task-fMRI acquisitions—as well as for resting state. For each participant, we calculated the regularized covariance [41] between the average BOLD time series extracted from each brain region pair. We then estimated partial correlations from the covariance matrix and consecutively applied the Fisher-Z transformation. This provided for each participant a connectivity matrix of size 168 × 168—one resting state matrix and one task-fMRI matrix per task. Mixture modeling offers an attractive way to analyze and model the noise in FC matrices without affecting reliable signal [42–44]. The main Gaussian from a mixture Gaussian-gamma model that was fitted on the distribution of edge values for each individual matrix supplied us with parameterized information about said distribution. We used these parameters to normalize the elements in each matrix by subtracting the main Gaussian mean and dividing by its standard deviation. In order to produce individual matrices of connectivity modulations induced by the task, i.e., task potency, we subtracted each participant’s resting state connectivity matrix from that participant’s task connectivity matrix. The resulting matrices are interpreted as containing the connectivity modulations away from the resting state baseline that the respective task induces in the brain—i.e., task potency [8, 45].
Normative modelling
A normative model of potency edge values was built against age and sex from typically developing participants using the nispat implementation in python [14, 46]. This means that downstream analyses are also inherently corrected for age and sex. The model furthermore inherently models uncertainty that is present due to availability of datapoints in the age and sex distributions. The model was used to predict the range of (a)typical edge values in out-of-sample typically developing participants as well as the autistic participants. Task potency values were thereby transformed to Z-scores quantifying the atypicality per edge of each individual’s task potency matrix elements given the normative reference. This was done separately in each task. Participant mean Z-scores were not significantly related to participant scanner motion (FD), more details available in Additional file 1 (Fig. S3A–E). Percentage of connectome deemed atypical was identified by thresholding the Z-scores at z =|± 2.571|, nominally describing the 1% most extreme positive and negative values. The number of edges passing this threshold for each individual was then expressed as a percentage of the total amount of edges. Independent t tests were performed on the percentages to assess case–control differences in the tasks. All p values were FDR corrected for multiple comparisons across tasks. Cohen’s D was calculated for an effect size estimate in each task.
Cross-task similarity
We investigate similarity in the patterns of atypicality across the tasks in autism as well as typically developing controls. This is done for both diagnostic groups separately by constructing a Pearson correlation matrix from the mean edgewise atypicality scores across tasks. Differences in covariance values are compared between the groups with the Wilcoxon signed rank test [47].
Canonical correlation analysis
Canonical correlation analysis (CCA) is a technique for finding latent linear multivariate relationships between two sets of data [48–50]. CCA analysis was done only in the individuals with autism in order to assess whether the variation in the task atypicality patterns relates also to phenotypic description in individuals with autism. The following processing steps were done separately for each task. For the behavioral side of the CCA analysis, the seven variables previously described were used (SRS, RBS, SSP, ADHD hyperactivity/impulsivity, ADHD inattentiveness, Vineland, IQ). For the brain side of the CCA, principal components-based dimensionality reduction was applied to the brain subjects by edges data matrix in order to make the CCA well-posed. Aiming for a balance between model accuracy and complexity, the top 10 variance components were kept for input in the CCA. Stability and generalizability of CCA parameters was assessed through 1000 different splits of tenfold out-of-sample cross-validation—building a ‘test’ distribution of the first canonical correlation as well as the paired CCA weights. Significance of the relationship found was assessed through building an out-of-sample null distribution of CCA correlations, i.e., permutation testing. Subjects were randomly permuted (within scan site) to break up the original correlation structure and performing again 1000 different splits of tenfold cross-validation. Non-spuriousness of the original relationship found was assessed by assessing whether the mean of the test distribution is more extreme than the 95th percentile of the null distribution.
Results
General connectome atypicality levels
Figure 1 shows for each task distributions of subjects for the percentage of total edges that pass the atypicality threshold at 1% two tailed (z = 2.571) in the autism and TD groups. We interpret this metric as a global subject-level atypicality score in each task. We identify significantly greater levels of atypical modulation in individuals with autism in each of the tasks. Flanker—Cohen’s d: 0.61, p < 0.01. Hariri—d: 0.34, p < 0.01. Monetary reward—d: 0.19, p < 0.01. Social reward—d: 0.19, p < 0.01. Theory of mind—d: 0.36, p < 0.01. For full information see Additional file 1: Table T1. These findings provide a global view where the distribution of edges in autism is shifted toward greater atypicality in achieving the same cognitive states as represented by task potency.
Fig. 1.
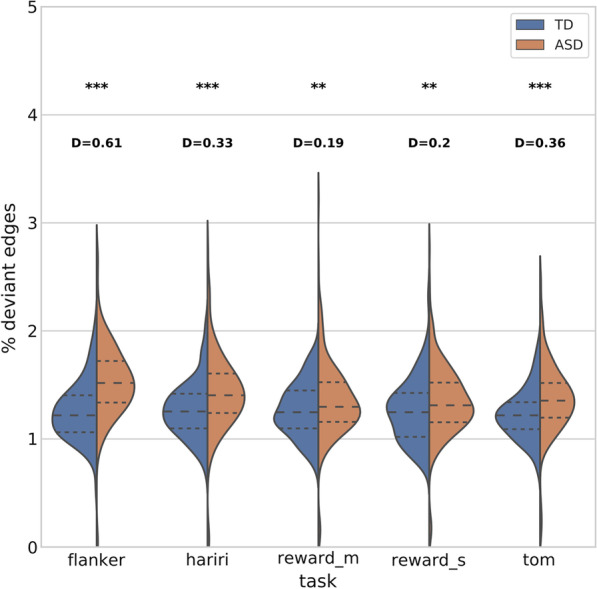
Violin plot of subject distributions for atypicality subject scores in each task. Independent t tests are performed between the subject distributions for each task. Derived p values (p), and Cohen’s d effect size (D) are displayed above the respective tasks. (**p < 0.01, ***p < 0.001)
Brain region atypicality levels
For the next analysis, we looked more closely at the spatial profile of connectivity modulation atypicality of autistic individuals with respect to typically developing controls. Figure 2 shows for each task the top 10% brain regions with the greatest atypicality scores in the autism group. To further identify the implicated cognitive terms associated with these regions, we used the Neurosynth online brain image decoder (October, 2021) to identify which networks and cognitive terms were represented in the atypicality data spatial pattern [51]. Because atypicality scores were initially estimated at the edge level, we computed the mean absolute atypicality value for all edges in a region, in order to present an atypicality score per brain region. For the Hariri task, the top matches were medial prefrontal cortex (r = 0.141), auditory (r = 0.132), and speech networks (r = 0.13), and superior temporal cortex (r = 0.119). In the Flanker task, the top matches between the pattern of atypicality and canonical brain networks were in order: medial prefrontal cortex (r = 0.144), default mode network (r = 0.12), and posterior cingulate (r = 0.11). For the monetary reward task, the top matches were again medial prefrontal cortex (r = 0.146), then somatosensory cortex (r = 0.116), default mode network (r = 0.115), and speech networks (r = 0.11). For the social reward task, the matches consisted of medial prefrontal cortex (r = 0.129), speech network (r = 0.11), and default mode network (r = 0.108). Finally, in the theory of mind task, the spatial pattern of atypical modulation was most closely matched to the speech network (r = 0.141), superior temporal cortex (r = 0.14), and auditory cortex (r = 0.139). The purely cognitive terms associated most with the atypicality scores were speech and listening/auditory regions across all tasks.
Fig. 2.

The top 10% brain regions with the greatest atypicality score in autism as summed over edges. From top to bottom: Hariri, Flanker, social reward, nonsocial reward, and theory of mind
Atypicality similarity across tasks
Figure 3 shows the correlation matrix of the atypicality edge pattern across the different tasks in the typically developing controls and autism groups. Correlations are consistently across tasks higher for autism (mean correlation: 0.43) than for TD (mean correlation 0.07). Wilcoxon signed rank test of the difference shows a significant difference at p < 0.002.
Fig. 3.

Heatmap of cross-task Pearson correlation of the edgewise group-level mean atypicality pattern in the brain for TD and ASD. Correlations are higher across the board for autism (mean correlation: 0.43), than for TD (mean correlation 0.07). Wilcoxon signed rank test shows a significant difference at p < 0.002
Canonical correlation analysis
We then investigated whether these spatial patterns of atypicality in the different tasks are also meaningfully related to behavioral measures of autism, and how this may vary across tasks. This grounds the model into clinical relevance. Canonical correlation analyses found significant brain-behavior modes of covariation in each of the tasks. Loadings displayed very high stability under out-of-sample tenfold cross-validation (correlations > 0.99). Figure 4 displays CCA loadings in the behavioral domain for the respective tasks. Instead of revealing differential patterns for the different cognitive domains probed by the tasks, the patterns found are similar across tasks and load on to the behavioral variables in their respective direction of greater impairment. Because we include behavioral variables that form dimensional cornerstones of ASD diagnosis (SRS: social interaction, RBS: repetitive behavior, SSP: sensory atypicality), this suggest that our brain–behavior relationship follows a main positive–negative axis of autism impairment. The loadings are of comparable magnitude across the tasks with the exception of IQ, which does load in the modes revealed from the Hariri and Flanker tasks, but shows minimal involvement in (non)social reward and theory of mind. Figure 5 shows the top loading brain regions in the CCA. Figure 5 is effectively a rotation of the data from Fig. 2 under the added influence of autism behavioral scores. In this situation, for the Hariri task, the top Neurosynth matches were anterior cingulate cortex (r = 0.215), speech cortex (r = 0.143), and anterior insula (r = 0.134). For the Flanker task, these were temporal cortex (r = 0.152), language regions (r = 0.148), and superior temporal cortex (r = 0.148). In the monetary reward task, they were visual cortex (r = 0.185), occipital cortex (r = 0.177), and motor cortex (r = 0.162). In social reward, the best matches were prefrontal cortex (r = 0.135), linguistic regions (r = 0.132), and motor cortex (r = 0.132). Finally, for the theory of mind task, they consisted of dorsolateral cortex (r = 0.162), auditory cortex (r = 0.145), and anterior cingulate (r = 0.14). Additional file 1: Figure S1 shows the difference between the data from Figs. 2 and 5 visualized.
Fig. 4.

Behavioral loadings for each task in the CCA analysis
Fig. 5.

The top 10% brain regions with the greatest CCA loadings in autism as summed over edges. From top to bottom: Hariri, Flanker, social reward, nonsocial reward, and theory of mind
Discussion
Our aim in this study was to identify and map individual-level (a)typicality in neural patterns associated with processing across different cognitive domains. We employ a task potency approach (functional connectivity modulation away from resting state baseline) for individuals with autism as they engage in tasks with different cognitive demands.
We show that autism is paired with greater individual-level global atypicality of brain connectivity modulation in each of the tasks under review. This could indicate broadly atypical deployment of neural resources under cognitive loads in autism and reiterates the necessity to move beyond simple group comparisons through the normative modelling approach. We further show that the atypicalities we model in the brain relate to behavioral measures of autism and found a robust and stable primary relationship in each of the tasks. The behavioral loadings of this relationship can furthermore be interpreted as describing a main axis of impairment in autism. The relationship implies that the behavioral and fMRI data carry similar information. This means that our feature, consisting of atypical task modulation and which was developed independently of behavioral information, is nonetheless a descriptor of autism impairment as measured by behavioral testing.
Interestingly, a large amount of the brain regions with the greatest atypicality scores were involved with language comprehension and language production, possibly pointing toward language networks being central to autism brain presentation. Furthermore, the spatial patterns of atypicality in the autism group display high levels of cross-task correlation, which was mostly absent in the typically developing controls. This suggests a high level of similar atypicality in autism when dealing with the various cognitive demands. Interestingly, these findings in combination would suggest that while individuals with autism have a globally more atypical pattern of task potency relative to controls, the specific spatial pattern of this atypicality does show clear similarity across cognitive domains. As far as these authors are aware, this is a novel finding and could support cognitive theories of autism involving a common deficit across mental domains. In the context of heterogeneous samples, it furthermore lends validity to the concept of an autism as a grouping from a functional neurobiological perspective. Research simultaneously investigating multiple tasks has previously been demonstrated in an ADHD cohort [45], yet has not been applied to autistic individuals. Given our findings this could be a promising target for future research in autism. A deeper investigation of why and how it is the case that cross-task similarity in atypicalities are on average much more similar in autism than in controls, and how this might relate to literature of the lack of functional differentiation in autism [3, 52] is necessary. One explanation could be that the brain connectivity pattern of individuals with autism is less free to fluctuate and reorganize under different cognitive loads. This explains both the atypicality in relationship to controls as well as the similarity across tasks within autism as found in this paper.
The findings in this paper need to be contextualized with regards to some limitations. Recent work in functional connectivity has highlighted the presence of test–retest variability in functional connectivity [53, 54]. This variability can have different sources such as participant traits, time of day, level of caffeination, medication, and task-magnitude to name a few. This can set participants up toward different FC fingerprints in different such situations. In our research, we use every individual participant as their own baseline through task potency. In task potency, the participant task-scan is calibrated by using the participant resting state scan which was acquired during the same scan procedure. This offers an inherent method of controlling for some of these sources of variability. The brain plots displayed in this paper should not be regarded as directly equivalent to activation maps. While areas highlighted in the figures do imply involvement of said area, this involvement is through the up- or downregulation of its coupling with other areas—not necessarily its own activation in isolation. fMRI tasks used in the LEAP sample were chosen based on their relevance for autism research, however still other forms of cognitive engagement may be relevant for a complete cross-task perspective. Furthermore, though we view autism through individualized atypicality metrics, the normative range estimation necessitates that the typically developing participants are treated as a single group. This potentially masks subgroups in autism that are embedded in the typically developing range.
Conclusions
To conclude, in this paper, we have applied innovative techniques to aid understanding of autism brain connectivity heterogeneity in a multi-task setting. These techniques reveal that individuals with autism engage with tasks in a globally atypical way, but that the particular pattern of this atypicality is nevertheless similar across tasks. Atypicalities across tasks originate mostly from prefrontal cortex and default mode network regions, but also speech and auditory networks. We furthermore validated the behavioral relevance of these techniques through showing significant relationships between brain and behavioral data. The similarities between atypicalities across the affected cognitive domains in autism may hold the key to furthering our understanding of the autistic brain. Further, we demonstrated the added value of innovative tools, i.e., task potency and normative modelling, with the goal to improve the interpretability of task-based fMRI functional connectivity and parse heterogeneity at the individual level in autism. We show that individuals with autism exhibit an atypical task-active functional connectome and we show that taking a cross-task perspective might help reveal a common pattern of atypicality in autism more broadly. Further research may focus on applying advanced discriminative and/or clustering procedures on the novel brain features that we have shown to be relevant for autism in order to predict and/or subtype autism on the basis of FC measurements. Additionally, the current research may be expanded to tasks beyond the five we take under consideration, as well as to further neurodevelopmental conditions.
Supplementary Information
Additional file 1: Scanning parameters, task description, subsample characterisations and additional figures.
Acknowledgements
We thank all participants and their families for participating in this study. We gratefully acknowledge the contributions of all members of the EU-AIMS LEAP group: Jumana Ahmad, Sara Ambrosino, Bonnie Auyeung, Tobias Banaschewski, Simon Baron-Cohen, Sarah Baumeister, Christian F. Beckmann, Sven Bölte, Thomas Bourgeron, Carsten Bours, Michael Brammer, Daniel Brandeis, Claudia Brogna, Yvette de Bruijn, Jan K. Buitelaar, Bhismadev Chakrabarti, Tony Charman, Ineke Cornelissen, Daisy Crawley, Flavio Dell’Acqua, Guillaume Dumas, Sarah Durston, Christine Ecker, Jessica Faulkner, Vincent Frouin, Pilar Garcés, David Goyard, Lindsay Ham, Hannah Hayward, Joerg Hipp, Rosemary Holt, Mark H. Johnson, Emily J.H. Jones, Prantik Kundu, Meng-Chuan Lai, Xavier Liogier D’ardhuy, Michael V. Lombardo, Eva Loth, David J. Lythgoe, René Mandl, Andre Marquand, Luke Mason, Maarten Mennes, Andreas Meyer-Lindenberg, Carolin Moessnang, Nico Mueller, Declan G.M. Murphy, Bethany Oakley, Laurence O’Dwyer, Marianne Oldehinkel, Bob Oranje, Gahan Pandina, Antonio M. Persico, Annika Rausch, Barbara Ruggeri, Amber Ruigrok, Jessica Sabet, Roberto Sacco, Antonia San José Cáceres, Emily Simonoff, Will Spooren, Julian Tillmann, Roberto Toro, Heike Tost, Jack Waldman, Steve C.R. Williams, Caroline Wooldridge, Iva Ilioska, Ting Mei and Marcel P. Zwiers.
Author contributions
TL, DF, AL, JB, and CB conceived and designed the analysis. TL, RC, AL, AM, and CB, contributed analysis tools. TL performed the analysis. TL wrote the paper. DF substantively revised the paper. AL and CB provided support in analyses. DM, TB, and TC have contributed to the coordination, data acquisition, funding, and coordination of the EU-AIMS LEAP project. All authors reviewed the manuscript.
Funding
The results leading to this publication have received funding from the Innovative Medicines Initiative 2 Joint Undertaking under Grant Agreement Nos. 115300 (for EU-AIMS) and 777394 (for AIMS-2-TRIALS). This Joint Undertaking receives support from the European Union's Horizon 2020 research and innovation programme and EFPIA and AUTISM SPEAKS, Autistica, SFARI. DLF is supported by funding from the European Union’s Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie Grant Agreement No. 101025785. This work has been further supported by the European Union Seventh Framework Programme Grant Nos. 602805 (AGGRESSOTYPE) (to JKB), 603016 (MATRICS) (to JKB), and 278948 (TACTICS) (to JKB); European Community’s Horizon 2020 Programme (H2020/2014-2020) Grant Nos. 643051 (MiND) (to JKB), 642996 (BRAINVIEW) (to JKB) and 847818 (CANDY) (to JKB and CFB); the Netherlands Organization for Scientific Research VICI Grant No. 17854 (to CFB); Wellcome Trust Collaborative Award Grant No. 215573/Z/19/Z (to CFB); the Autism Research Trust (to SBC), and the NWO Gravitation Programme Language in Interaction (Grant 024.001.006). The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results. Any views expressed are those of the author(s) and not necessarily those of the funders.
Availability of data and materials
At the moment of submission, data are not publicly available. The EU-AIMS consortium will make the data publicly available in the near future and the exact protocol to access this data is under discussion.
Declarations
Ethics approval and consent to participate
The ethics committee (London Queen Square Health Research Authority Research Ethics Committee) of KCL and University of Cambridge gave ethical approval for this work (13/LO/1156). The ethics committee (Radboud Universitair Medisch Centrum Instituut Waarborging Kwaliteit en Veiligheid Commissie Mensgebonden Onderzoek and Regio Arnhem-Nijmegen (Radboud University Medical Centre Institute Ensuring Quality and Safety Committee on Research Involving Human Subjects Arnhem-Nijmegen) of Radboud UMC gave ethical approval of this work (2013/455). The ethics committee of Central Institute of Mannheim (UMM Universitaetsmedizin Mannheim, Medizinische Ethik Commission II (UMM University Medical Mannheim, Medical Ethics Commission II) gave ethical approval for this work (2014-540N-MA). The ethics committee of University of Rome (Universita Campus Bio-Medica De Roma Comitato Etico (University Campus Bio-Medical Ethics Committee De Roma) gave ethical approval of this work (8/14 PAR ComET CBM).
Consent for publication
Not applicable.
Competing interests
JKB has been a consultant to, advisory board member of, and a speaker for Janssen Cilag BV, Eli Lilly, Shire, Lundbeck, Roche, and Servier. He is not an employee of any of these companies, and not a stock shareholder of any of these companies. He has no other financial or material support, including expert testimony, patents or royalties. CFB is director and shareholder in SBGNeuro Ltd. The present work is unrelated to the above grants and relationships. TB served in an advisory or consultancy role for ADHS digital, Infectopharm, Lundbeck, Medice, Neurim Pharmaceuticals, Oberberg GmbH, Roche, and Takeda. He received conference support or speaker’s fee by Medice and Takeda. He received royalities from Hogrefe, Kohlhammer, CIP Medien, Oxford University Press. TC has received consultancy from Roche and Servier and received book royalties from Guildford Press and Sage. The other authors report no biomedical financial interests or potential conflicts of interest.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Tristan Looden, Email: tristan.looden@donders.ru.nl.
the AIMS-2-TRIALS group:
Jumana Ahmad, Sara Ambrosino, Bonnie Auyeung, Tobias Banaschewski, Simon Baron-Cohen, Sarah Baumeister, Christian F. Beckmann, Sven Bölte, Thomas Bourgeron, Carsten Bours, Michael Brammer, Daniel Brandeis, Claudia Brogna, Yvette de Bruijn, Jan K. Buitelaar, Bhismadev Chakrabarti, Tony Charman, Ineke Cornelissen, Daisy Crawley, Flavio Dell’ Acqua, Guillaume Dumas, Sarah Durston, Christine Ecker, Jessica Faulkner, Vincent Frouin, Pilar Garcés, David Goyard, Lindsay Ham, Hannah Hayward, Joerg Hipp, Rosemary Holt, Mark H. Johnson, Emily J. H. Jones, Prantik Kundu, Meng-Chuan Lai, Xavier Liogier D’ardhuy, Michael V. Lombardo, Eva Loth, David J. Lythgoe, René Mandl, Andre Marquand, Luke Mason, Maarten Mennes, Andreas Meyer-Lindenberg, Carolin Moessnang, Nico Mueller, Declan G. M. Murphy, Bethany Oakley, Laurence O’Dwyer, Marianne Oldehinkel, Bob Oranje, Gahan Pandina, Antonio M. Persico, Annika Rausch, Barbara Ruggeri, Amber Ruigrok, Jessica Sabet, Roberto Sacco, Antonia San José Cáceres, Emily Simonoff, Will Spooren, Julian Tillmann, Roberto Toro, Heike Tost, Jack Waldman, Steve C. R. Williams, Caroline Wooldridge, Iva Ilioska, Ting Mei, and Marcel P. Zwiers
References
- 1.American Psychiatric Association . Diagnostic and statistical manual of mental disorders (DSM-5®) American Psychiatric Pub; 2013. [Google Scholar]
- 2.Oldehinkel M, Mennes M, Marquand A, Charman T, Tillmann J, Ecker C, et al. Altered connectivity between cerebellum, visual, and sensory-motor networks in autism spectrum disorder: results from the EU-AIMS longitudinal european autism project. Biol Psychia Cogn Neurosci Neuroimag. 2019;4(3):260–270. doi: 10.1016/j.bpsc.2018.11.010. [DOI] [PubMed] [Google Scholar]
- 3.Picci G, Gotts SJ, Scherf KS. A theoretical rut: revisiting and critically evaluating the generalized under/over-connectivity hypothesis of autism. Dev Sci. 2016;19:524–549. doi: 10.1111/desc.12467. [DOI] [PubMed] [Google Scholar]
- 4.Uddin LQ, Supekar K, Menon V. Reconceptualizing functional brain connectivity in autism from a developmental perspective. Front Human Neurosci. 2013 doi: 10.3389/fnhum.2013.00458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Deshpande G, Libero LE, Sreenivasan KR, Deshpande HD, Kana RK. Identification of neural connectivity signatures of autism using machine learning. Front Hum Neurosci. 2013;7:1–15. doi: 10.3389/fnhum.2013.00670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Just MA, Cherkassky VL, Buchweitz A, Keller TA, Mitchell TM. Identifying autism from neural representations of social interactions: neurocognitive markers of autism. PLoS ONE. 2014;9:1–22. doi: 10.1371/journal.pone.0113879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Moessnang C, Baumeister S, Tillmann J, Goyard D, Charman T, Ambrosino S, et al. Social brain activation during mentalizing in a large autism cohort: the Longitudinal European Autism Project. Mol Autism. 2020;11:1–17. doi: 10.1186/s13229-020-0317-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Chauvin RJ, Mennes M, Llera A, Buitelaar JK, Beckmann CF. Disentangling common from specific processing across tasks using task potency. Neuroimage. 2019;184:632–645. doi: 10.1016/j.neuroimage.2018.09.059. [DOI] [PubMed] [Google Scholar]
- 9.Mennes M, Kelly C, Zuo XN, Di Martino A, Biswal BB, Castellanos FX, Milham MP. Inter-individual differences in resting-state functional connectivity predict task-induced BOLD activity. Neuroimage. 2010;50:1690–1701. doi: 10.1016/j.neuroimage.2010.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hull JV, Jacokes ZJ, Torgerson CM, Irimia A, Van Horn JD, Aylward E, et al. Resting-state functional connectivity in autism spectrum disorders: a review. Front Psychiatry. 2017 doi: 10.3389/fpsyt.2016.00205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Brunsdon VE, Happé F. Exploring the ‘fractionation’ of autism at the cognitive level. Autism. 2014;18:17–30. doi: 10.1177/1362361313499456. [DOI] [PubMed] [Google Scholar]
- 12.Nunes AS, Peatfield N, Vakorin V, Doesburg SM. Idiosyncratic organization of cortical networks in autism spectrum disorder. Neuroimage. 2018 doi: 10.1016/j.neuroimage.2018.01.022. [DOI] [PubMed] [Google Scholar]
- 13.Wolfers T, Floris DL, Dinga R, van Rooij D, Isakoglou C, Kia SM, et al. From pattern classification to stratification: towards conceptualizing the heterogeneity of Autism Spectrum Disorder. Neurosci Biobehav Rev. 2019;104:240–254. doi: 10.1016/j.neubiorev.2019.07.010. [DOI] [PubMed] [Google Scholar]
- 14.Marquand AF, Kia SM, Zabihi M, Wolfers T, Buitelaar JK, Beckmann CF. Conceptualizing mental disorders as deviations from normative functioning. Mol Psychiatry. 2019;24:1415–1424. doi: 10.1038/s41380-019-0441-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Bethlehem RAI, Seidlitz J, Romero-Garcia R, Trakoshis S, Dumas G, Lombardo MV. A normative modelling approach reveals age-atypical cortical thickness in a subgroup of males with autism spectrum disorder. Commun Biol. 2020;3:486. doi: 10.1038/s42003-020-01212-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Floris DL, Wolfers T, Zabihi M, Holz NE, Zwiers MP, Charman T, et al. A typical brain asymmetry in autism—a candidate for clinically meaningful stratification. Biol Psychiatry Cogn Neurosci Neuroimaging. 2020;43:1–11. doi: 10.1016/j.bpsc.2020.08.008. [DOI] [PubMed] [Google Scholar]
- 17.Zabihi M, Oldehinkel M, Wolfers T, Frouin V, Goyard D, Loth E, et al. Dissecting the heterogeneous cortical anatomy of autism spectrum disorder using normative models. Biol Psychiatry Cogn Neurosci Neuroimaging. 2019;4:567–578. doi: 10.1016/j.bpsc.2018.11.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Charman T, Loth E, Tillmann J, Crawley D, Wooldridge C, Goyard D, et al. The EU-AIMS Longitudinal European Autism Project (LEAP): clinical characterisation. Mol Autism. 2017;8:1–21. doi: 10.1186/s13229-017-0145-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Charman T, Loth E, Tillmann J, Crawley D, Wooldridge C, Goyard D, et al. The EU-AIMS Longitudinal European Autism Project (LEAP): methods. Mol Autism. 2017;8:1–19. doi: 10.1186/s13229-017-0145-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Loth E, Charman T, Mason L, Tillmann J, Jones EJH, Wooldridge C, et al. The EU-AIMS Longitudinal European Autism Project (LEAP): design and methodologies to identify and validate stratification biomarkers for autism spectrum disorders. Mol Autism. 2017;8:24. doi: 10.1186/s13229-017-0146-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Mihalik A, Ferreira FS, Moutoussis M, Ziegler G, Adams RA, Rosa MJ, et al. Multiple holdouts with stability: improving the generalizability of machine learning analyses of brain–behavior relationships. Biol Psychiatry. 2020;87:368–376. doi: 10.1016/j.biopsych.2019.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Wang HT, Smallwood J, Mourao-Miranda J, Xia CH, Satterthwaite TD, Bassett DS, Bzdok D. Finding the needle in a high-dimensional haystack: canonical correlation analysis for neuroscientists. Neuroimage. 2020;216:116745. doi: 10.1016/j.neuroimage.2020.116745. [DOI] [PubMed] [Google Scholar]
- 23.Chauvin RJ, Mennes M, Buitelaar JK, Beckmann CF. Assessing age-dependent multi-task functional co-activation changes using measures of task-potency. Dev Cogn Neurosci. 2017;33:0–1. doi: 10.1016/j.dcn.2017.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Jenkinson M, Bannister P, Brady M, Smith S. Improved optimization for the robust and accurate linear registration and motion correction of brain images. Neuroimage. 2002;17:825–841. doi: 10.1006/nimg.2002.1132. [DOI] [PubMed] [Google Scholar]
- 25.Constantino JN. Social responsiveness scale. In: Volkmar FR, editor. Encyclopedia of Autism Spectrum Disorders. New York, NY: Springer; 2013. pp. 2919–2929. [Google Scholar]
- 26.Bodfish JW, Symons FJ, Parker DE, Lewis MH. Varieties of repetitive behavior in autism: comparisons to mental retardation. J Autism Dev Disord. 2000;30:237–243. doi: 10.1023/A:1005596502855. [DOI] [PubMed] [Google Scholar]
- 27.Tomchek SD, Dunn W. Sensory processing in children with and without autism: a comparative study using the short sensory profile. Am J Occup Ther. 2007;61:190–200. doi: 10.5014/ajot.61.2.190. [DOI] [PubMed] [Google Scholar]
- 28.Sparrow SS. Vineland adaptive behavior scales. In: Kreutzer JS, DeLuca J, Caplan B, editors. Encyclopedia of clinical neuropsychology. New York, NY: Springer; 2011. pp. 2618–2621. [Google Scholar]
- 29.Wechsler D, Zhou X, Psychological Corporation., Assessment Library Materials (University of Lethbridge. Faculty of Education. Curriculum Laboratory) (2011) WASI-II : Wechsler Abbreviated Scale of Intelligence.
- 30.Hariri AR, Tessitore A, Mattay VS, Fera F, Weinberger DR. The amygdala response to emotional stimuli: a comparison of faces and scenes. Neuroimage. 2002;17:317–323. doi: 10.1006/nimg.2002.1179. [DOI] [PubMed] [Google Scholar]
- 31.Meyer-Lindenberg A, Weinberger DR. Intermediate phenotypes and genetic mechanisms of psychiatric disorders. Nat Rev Neurosci. 2006;7:818–827. doi: 10.1038/nrn1993. [DOI] [PubMed] [Google Scholar]
- 32.Delmonte S, Balsters JH, McGrath J, Fitzgerald J, Brennan S, Fagan AJ, Gallagher L. Social and monetary reward processing in autism spectrum disorders. Mol Autism. 2012;3:1–13. doi: 10.1186/2040-2392-3-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Castelli F, Frith C, Happé F, Frith U. Autism, Asperger syndrome and brain mechanisms for the attribution of mental states to animated shapes. Brain J Neurol. 2002;125:1839–1849. doi: 10.1093/brain/awf189. [DOI] [PubMed] [Google Scholar]
- 34.White SJ, Coniston D, Rogers R, Frith U. Developing the Frith-Happé animations: a quick and objective test of Theory of Mind for adults with autism. Autism Res. 2011;4:149–154. doi: 10.1002/aur.174. [DOI] [PubMed] [Google Scholar]
- 35.Jenkinson M, Beckmann CF, Behrens TEJ, Woolrich MW, Smith SM. FSL. Neuroimage. 2012;62:782–790. doi: 10.1016/j.neuroimage.2011.09.015. [DOI] [PubMed] [Google Scholar]
- 36.Pruim RHR, Mennes M, Buitelaar JK, Beckmann CF. Evaluation of ICA-AROMA and alternative strategies for motion artifact removal in resting state fMRI. Neuroimage. 2015;112:278–287. doi: 10.1016/j.neuroimage.2015.02.063. [DOI] [PubMed] [Google Scholar]
- 37.Pruim RHR, Mennes M, van Rooij D, Llera A, Buitelaar JK, Beckmann CF. ICA-AROMA: a robust ICA-based strategy for removing motion artifacts from fMRI data. Neuroimage. 2015;112:267–277. doi: 10.1016/j.neuroimage.2015.02.064. [DOI] [PubMed] [Google Scholar]
- 38.Anderson ML. The massive redeployment hypothesis and the functional topography of the brain. Philos Psychol. 2007;20:143–174. doi: 10.1080/09515080701197163. [DOI] [Google Scholar]
- 39.Johnson WE, Li C, Rabinovic A. Adjusting batch effects in microarray expression data using empirical Bayes methods. Biostatistics. 2007;8:118–127. doi: 10.1093/biostatistics/kxj037. [DOI] [PubMed] [Google Scholar]
- 40.van Oort ESB, Mennes M, Navarro Schröder T, Kumar VJ, Zaragoza Jimenez NI, Grodd W, et al. Functional parcellation using time courses of instantaneous connectivity. Neuroimage. 2017;170:1–10. doi: 10.1016/j.neuroimage.2017.07.027. [DOI] [PubMed] [Google Scholar]
- 41.Ledoit O, Wolf M. Nonlinear shrinkage estimation of large-dimensional covariance matrices. Ann Stat. 2012;40:1024–1060. doi: 10.1214/12-AOS989. [DOI] [Google Scholar]
- 42.Bielczyk NZ, Walocha F, Ebel PW, Haak KV, Llera A, Buitelaar JK, et al. Thresholding functional connectomes by means of mixture modeling. Neuroimage. 2018 doi: 10.1016/j.neuroimage.2018.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Llera A, Vidaurre D, Pruim RHR, Beckmann CF (2016) Variational mixture models with gamma or inverse-gamma components. Retrieved from http://arxiv.org/abs/1607.07573.
- 44.Feinberg DA, Moeller S, Smith SM, Auerbach E, Ramanna S, Glasser MF, et al. Multiplexed echo planar imaging for sub-second whole brain FMRI and fast diffusion imaging. PLoS ONE. 2010;5:e15710. doi: 10.1371/journal.pone.0015710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Chauvin RJ, Buitelaar JK, Sprooten E, Oldehinkel M, Franke B, Hartman C, et al. Task-generic and task-specific connectivity modulations in the ADHD brain: an integrated analysis across multiple tasks. Transl Psychiatry. 2021;11:159. doi: 10.1038/s41398-021-01284-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Marquand AF, Rezek I, Buitelaar J, Beckmann CF. Understanding heterogeneity in clinical cohorts using normative models: beyond case-control studies. Biol Psychiatry. 2016;80:552–561. doi: 10.1016/j.biopsych.2015.12.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Wilcoxon F. Individual comparisons by ranking methods. Biom Bull. 1945;1:80–83. doi: 10.2307/3001968. [DOI] [Google Scholar]
- 48.Hotelling H. Relations between two sets of variates. Biometrika. 1936;28:321. doi: 10.1093/biomet/28.3-4.321. [DOI] [Google Scholar]
- 49.Pedregosa F, Varoquaux G, Gramfort A, Michel V, Thirion B, Grisel O, et al. Scikit-learn: machine learning in Python. J Mach Learn Res. 2011;12:2825–2830. [Google Scholar]
- 50.Smith SM, Nichols TE, Vidaurre D, Winkler AM, Behrens TEJ, Glasser MF, et al. A positive-negative mode of population covariation links brain connectivity, demographics and behavior. Nat Neurosci. 2015;18:1565–1567. doi: 10.1038/nn.4125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Gorgolewski KJ, Varoquaux G, Rivera G, Schwarz Y, Ghosh SS, Maumet C, et al. NeuroVault.org: a web-based repository for collecting and sharing unthresholded statistical maps of the human brain. Front Neuroinform. 2015;0:8. doi: 10.3389/fninf.2015.00008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Nebel MB, Joel SE, Muschelli J, Barber AD, Caffo BS, Pekar JJ, Mostofsky SH. Disruption of functional organization within the primary motor cortex in children with autism. Hum Brain Mapp. 2012;35:567–580. doi: 10.1002/hbm.22188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Noble S, Scheinost D, Constable RT. A decade of test-retest reliability of functional connectivity: a systematic review and meta-analysis. Neuroimage. 2019;203:116157. doi: 10.1016/j.neuroimage.2019.116157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Noble S, Scheinost D, Constable RT. A guide to the measurement and interpretation of fMRI test–retest reliability. Curr Opin Behav Sci. 2021;40:27–32. doi: 10.1016/j.cobeha.2020.12.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1: Scanning parameters, task description, subsample characterisations and additional figures.
Data Availability Statement
At the moment of submission, data are not publicly available. The EU-AIMS consortium will make the data publicly available in the near future and the exact protocol to access this data is under discussion.