

Abstract
Aspergillus terreus is well-known for lovastatin and itaconic acid production with biomedical and commercial importance. The mechanisms of metabolite formation have been extensively studied to improve their yield through genetic engineering. However, the combined repertoire of carbohydrate-active enzymes (CAZymes), cytochrome P450s (CYP) enzymes, and secondary metabolites (SMs) in the different A. terreus strains has not been well studied yet, especially with respect to the presence of biosynthetic gene clusters (BGCs). Here we present a 30 Mb whole genome sequence of A. terreus ATCC 20541 in which we predicted 10,410 protein-coding genes. We compared the CAZymes, CYPs enzyme, and SMs across eleven A. terreus strains, and the results indicate that all strains have rich pectin degradation enzyme and CYP52 families. The lovastatin BGC of lovI was linked with lovF in A. terreus ATCC 20541, and the phenomenon was not found in the other strains. A. terreus ATCC 20541 lacked a non-ribosomal peptide synthetase (AnaPS) participating in acetylaszonalenin production, which was a conserved protein in the ten other strains. Our results present a comprehensive analysis of CAZymes, CYPs enzyme, and SM diversities in A. terreus strains and will facilitate further research in the function of BGCs associated with valuable SMs.
Subject terms: Microbial genetics, Microbiology
Introduction
Aspergillus spp. have the ability to adapt to various environments, likely, in part, because they produce a large array of secondary metabolites (SMs)1. SMs can be used in survival strategies against competitors via acidification, antifungal, and antiparasitic activity in nature and for invading hosts2,3. Since fungi represent a vast resource for the discovery of new valuable carbohydrate-active enzymes (CAZymes) and SMs2,4, there is a massive interest in sequencing new species and species isolates with the aim of finding new genes and products5. To this end, we note that the identification of genes for SM production is less complicated in fungi as all genes relevant for the production of a given SM are typically organized in biosynthetic gene clusters (BGCs), rather than being scattered around in the genome like in plants6,7. Typically, fungal BGCs contain a gene coding for a synthase that supplies the scaffold, which could be a non-ribosomal peptide, a terpene, or a polyketide; and genes coding for tailoring enzymes, which decorate the scaffold to produce the mature SM8.
Aspergillus terreus is commonly found in many diverse habitats including agricultural, mangrove, soil rhizospheres in tropical and subtropical regions, organs of living creatures, and even in the human lungs and sputum9–11. Importantly, A. terreus is a harmful pathogen to crops, which has been reported in rice (Oryza sativa L.), wheat (Triticum aestivum), potato (Solanum tuberosum L.), maize (Zea mays), and soybean (Glycine max L.)12,13, and it may even infect humans to cause detrimental invasive aspergillosis (IA)14,15. Though A. terreus causes severe agricultural damage and acts as a human pathogen, it is also a well-known industrial workhorse. For example, it is a well-known producer of itaconic acid16,17 and lovastatin18,19, the former of which is heavily used in the chemical industries, and the latter of which is an important cholesterol-lowering drug. A. terreus also produce many carbohydrate-active enzymes (CAZymes) including cellulases, xylanases, lipases, and phytases that are employed in plant biomass degradation20–24.
Given the large interest in A. terreus, ten whole genome sequences of A. terreus strains are available in the National Center for Biotechnology Information Search database (NCBI). This includes the whole genome of A. terreus NIH2624, which was the first reference genome for A. terreus. The genome of A. terreus strains M6925, ATCC 20542, ML-44, ASM-1, TN-484, IFO 6365, w25, 45A, and T3 Kankrej were subsequently published from 2016 to 202125–30. Many fungal genomes have been included in comparative studies aiming at improving the understanding of SM production, and discovering the evolutionary patterns in SM gene clusters diversity in fungal species, e.g., Aspergillus species (A. fumigatus, A. nidulans, and A. flavus), Fusarium species (F. oxysporum and F. fujikuroi) and Botrytis cinerea31–37. However, comprehensive comparisons of SM gene cluster polymorphisms of selected A. terreus strains have not be characterized.
In this study, we provide a thorough overview of the natural product repertoire of A. terreus. One future goal is to improve the yield of lovastatin and genomic differences may contribute useful insights. Towards this goal, we sequenced the genome of A. terreus isolate, ATCC 20541. We then compared all eleven A. terreus whole genomes, including genes encoding putative CAZymes, cytochrome P450s (CYPs), and BGC for SM production, which help to understand genomic diversity between A. terreus ATCC 20541 and other A. terreus strains. In particular, the variations in SM BGCs within A. terreus strains were analyzed. Overall, this study provides a high-quality whole genome assembly and annotation of A. terreus ATCC 20541 and a comprehensive analysis of CAZymes, CYPs, and SMs diversity in A. terreus strains.
Results
Assembly statistics and general features of A. terreus ATCC 20541 genome
We first fully genome sequenced A. terreus ATCC 20541 using Novaseq 6000 platforms (Illumina, USA) with 2 × 150 paired-end reads and obtained a total number of 69.7 million reads (~ 349 × coverage). The assembled genome represents 97.6% completeness (BUSCO) with a contig N50 length of 1.6 Mb and the longest contig of 3.8 Mb. The resulting genome assembly was based on 156 contigs with a 52.3% GC content, 10,410 protein-coding genes, 33 rRNA genes, and 180 tRNA genes. 1016 signal peptides and 2483 transmembrane helices were detected. Pfam domains were assigned to 8235 proteins based on InterProscan program. Subsequently, a total of 6689 proteins were assigned to the eukaryotic orthologous groups (KOG) databases (Table 1; Supplementary Fig. S1). The abundance of metabolism (III) was about 60%, which was the highest in these four categories. 2053 genes (19.7%) were annotated using KAAS and KEGG map, and KEGG annotations contained seven major pathways including BRITE hierarchies (36%), metabolism (34%), genetic information processing (13%), cellular processes (8%), environmental information processing (6%), human pathogenicity (2%), and organismal systems (2%). 18 genes were classified into the biosynthesis of “other” secondary metabolites (Supplementary Fig. S2). Gene ontology (GO) terms were divided into three major function categories: biological processes (39%), molecular functions (35%), and cellular components (26%). 8,671 genes (83%) were annotated and assigned to the three categories. 185 genes were classified into the secondary metabolite biosynthetic process (Supplementary Fig. S3; Supplementary Table S1). 1434 genes were unknown function in KOG, GO, and KEGG.
Table 1.
Genomic features of the A. terreus ATCC 20541.
| Features | Value |
|---|---|
| Total number of reads-Illumina (bp) | 69,681,640 |
| Genome coverage-Illumina | 349 X |
| Contigs/scaffolds | 156 |
| Largest contigs/scaffolds (bp) | 3,819,325 |
| N50 (bp) | 1,581,806 |
| Complete BUSCOs | 97.6% |
| Number of protein coding genes | 10,410 |
| rRNA genes | 33 |
| tRNA genes | 180 |
| Proteins with signal peptide | 1016 |
| Proteins with transmembrane helices | 2483 |
| Proteins with predicted Pfam domain | 8235 |
| Proteins assigned to KOG | 6689 |
| Predicted CAZymes proteins | 479 |
Phylogenetic tree and comparative genomics of A. terreus strains
In addition to the novel sequence of A. terreus ATCC 20541, we obtained the genomes of ten additional A. terreus strains accessed from the NCBI database. Previous research reported that A. terreus can be isolated in many different environments, including humans, soil, plant root, land, and marine creatures (Supplementary Fig. S4). Based on this, we found that the average genome size of the A. terreus strains was about 29.8 Mb and the average GC content was 52.5%. We observed three main clades in the phylogenetic tree of A. terreus. The first clade was constituted by A. terreus NIH2624, 45A, ATCC 20541, M6925, w25, ASM-1, ML-44, and T3 Kankrej strains. The second was A. terreus ATCC 20542, and the third was by A. terreus IFO 6365 and TN-484 (Supplementary Fig. S4). A. terreus T3 Kankrej showed the highest number of contigs/scaffolds and the lowest N50 value. We evaluated the completeness of A. terreus strains using BUSCO. BUSCO analysis indicated that the genome assembly contained > 95% complete single-copy BUSCOs and < 0.03% missing BUSCOs in all A. terreus strains, except for A. terreus T3 Kankrej with < 40% complete single-copy BUSCOs and > 40% missing BUSCOs (Supplementary Fig. S5). Therefore, we excluded A. terreus T3 Kankrej from subsequent analyses as the quality of genome assembly was unlikely sufficient for later analyses27.
The CAZymes and CYPs analysis among A. terreus strains
To evaluate the plant biomass-degrading ability across all A. terreus strains, the genes from the CAZyme family were predicted using dbCAN2 meta server with HAMMER, DIAMOND, and eCAMI38. CAZymes were divided into six main groups: glycoside hydrolases (GH), glycosyltransferases (GT), polysaccharide lyases (PL), carbohydrate esterases (CE), auxiliary activities (AA), and carbohydrate-binding modules (CBMs). According to De Vries et al.39, seven CAZymes families (cellulose, xylan, galactomannan, xyloglucan, pectin, starch, and inulin) were found in the plant biomass-degrading ability of Aspergillus spp. (Supplementary Table S2). All strains showed the highest and lowest percentage of value of genes associated with pectin degradation and inulin degradation, respectively. A. terreus M6925 showed 135 plant biomass-degrading related genes, and it was different from the other strains. The number of CAZymes genes in A. terreus ATCC 20541 was similar to those in A. terreus ATCC 20542 (Fig. 1). Fungal CYPs are associated with diverse biosynthesis including the production of primary and secondary metabolites and denitrification of xenobiotics40. In the CYP results, A. terreus ATCC 20541 had 150 CYPs, which could be classified into 22 families. The CYP52 family contained the highest number of genes (40–44 genes), and the second largest group including CYP504 and CYP58 family contained 15–22 genes in A. terreus strains (Table 2).
Figure 1.
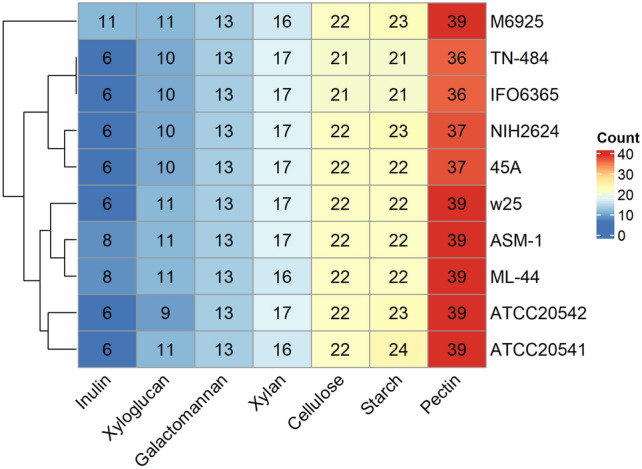
Number of genes related to degradation of different plant-based polysaccharides detected in A. terreus genomes.
Table 2.
Comparison of the cytochrome P450 genes in the A. terreus strains.
| CYP | ATCC 20541 | ATCC 20542 | 45A | ASM-1 | ML-44 | NIH2624 | w25 | M6925 | IFO 6365 | TN-484 |
|---|---|---|---|---|---|---|---|---|---|---|
| CYP52 | 41 | 40 | 40 | 40 | 40 | 41 | 41 | 42 | 44 | 44 |
| CYP504 | 18 | 18 | 18 | 18 | 18 | 18 | 18 | 22 | 18 | 18 |
| CYP58 | 15 | 19 | 14 | 17 | 17 | 19 | 19 | 21 | 18 | 18 |
| CYP3 | 11 | 12 | 12 | 13 | 12 | 11 | 11 | 12 | 11 | 11 |
| CYP51 | 11 | 11 | 13 | 10 | 10 | 13 | 10 | 10 | 7 | 7 |
| CYP65 | 10 | 10 | 12 | 11 | 11 | 10 | 10 | 14 | 13 | 13 |
| CYP61 | 9 | 9 | 6 | 9 | 9 | 9 | 6 | 9 | 6 | 6 |
| CYP68 | 9 | 6 | 4 | 7 | 7 | 6 | 6 | 6 | 6 | 6 |
| CYP5293 | 4 | 5 | 5 | 7 | 7 | 4 | 7 | 7 | 3 | 3 |
| CYP620 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| CYP5080 | 3 | 2 | 3 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| CYP505 | 3 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| CYP102 | 3 | 2 | 2 | 3 | 3 | 3 | 4 | 3 | 3 | 3 |
| CYP82 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 3 | 2 |
| CYP83 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| CYP541 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| CYP53 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| CYP2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 4 | 2 | 2 |
| CYP78 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| CYP71 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| CYP98 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| CYP4 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
Secondary metabolism and amino acid variants
A. terreus ATCC 20541 genome encoded 73 SM biosynthetic backbone genes according to the fungal antiSMASH v 6.0.1 database41. SM biosynthetic backbone genes types in A. terreus strains (Fig. 2) included type I polyketide synthase (T1PKS), non-ribosomal peptide synthetase (NRPS), PKS-NRPS hybrid, NRPS-like, T1PKS/NRPS-like, terpene (TC), indole (DMAT), betalactone, and siderophore. T1PKS were richly represented among A. terreus strains; and the itaconic acid producers, such as IFO 6365 and TN-484, showed the highest number of putative T1PKS. Additionally, A. terreus IFO 6365 and TN-484 lacked the SM cluster type of siderophore (Fig. 2).
Figure 2.

Core biosynthetic genes of A. terreus strains predicted by the antiSMASH database based on AUGUSTUS annotations. T1PKS polyketide synthase, NRPS nonribosomal peptides synthase, Hybrid PKS-NRPS hybrid, TC terpene cyclase, DMAT dimethylallyl tryptophan synthase.
We observed 65 conserved core SM proteins among six A. terreus strains, and 20 SM proteins among five A. terreus strains. A. terreus ATCC 20541 was predicted to encode a unique SM protein, which was similar to M6925 (Fig. 3; Supplementary Fig. S6). A. terreus TN-484 and ASM-1 were predicted to encode eight and three unique SM proteins, respectively. We noted that A. terreus TN-484 has the highest number of unique SM proteins among all strains, and A. terreus IFO 6365 and TN-484 were the most phylogenetically distant from other A. terreus strains (Fig. 3; Supplementary Fig. S6). Interestingly, our analyses indicated that the genome of A. terreus ATCC 20541, ATCC 20542, and w25 did not encode non-ribosomal peptide synthetase (AnaPS), terretonin (Trt4), and acetylaranotin (AtaP), respectively (Fig. 4; Supplementary Fig. S7).
Figure 3.

Comparison of the predicted secondary metabolism in A. terreus strains. Upset plot showing SM proteins from six representative genomes in each clade.
Figure 4.
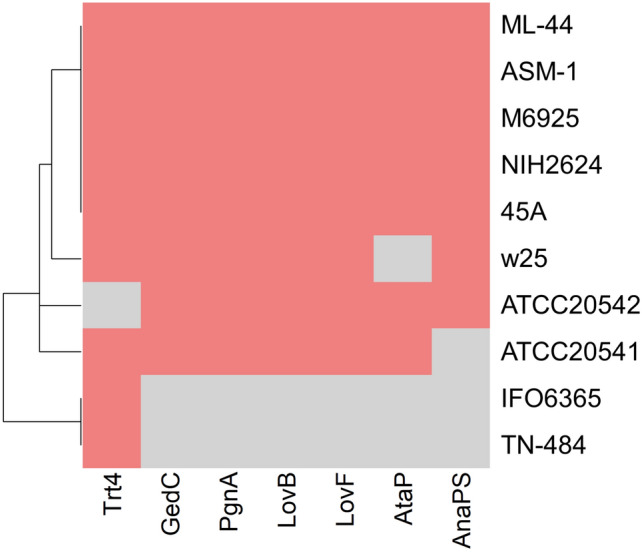
Heatmap represents the presence (pink) or absence (gray) matrix of orthologous A. terreus NIH2624 core biosynthetic proteins associated with SMs in A. terreus strains. The dendrogram was generated based on hierarchical clustering analysis. X-axis: orthologous A. terreus NIH2624 core SM proteins (Romsdahl and Wang, 2019); Y-axis: strain clustering. The completed results were given in Supplementary Fig. S7.
Lovastatin BGC comparison with A. terreus, M. pilosus and P. citrinum
Several filamentous fungi produce lovastatin, such as Monascus species (M. ruber, M. purpureus, M. sanguineus, and M. pilosus), Penicillium citrinum (P. citrinum), Trichoderma viride etc42–45. The lovastatin BGC comparison showed that lovI was absent in A. terreus w25 and M6925 strains, and lovI was linked with lovF in A. terreus ATCC 20541. In A. terreus ATCC 20542 lovI acts as a transport-related gene, but in A. terreus NIH2624, ASM-1, ML-44 and 45A lovI act as the “other genes” (note: the term “other genes” was shown in antiSMASH v. 6.0.1 database). In addition, the arrangement and composition of lovastatin BGC in A. terreus ATCC 20541, ASM-1, ML-44, ATCC 20542, NIH2624, 45A, M6925, and w25 were similar to the BGC in M. pilosus and P. citrinum (Fig. 5). LovB and lovF, the “core biosynthetic genes”, play crucial roles to form the lovastatin core structure46,47. 30 and 59 amino acid changes (referred to as a non-synonymous single nucleotide polymorphism (SNP) or mutation) were observed in lovB and lovF genomic regions in A. terreus ATCC 20541 (Supplementary Figs. S8, S9). There were 115 transpositions, including 84 additional amino acids in positions 395–566 and 31 missing amino acids in positions 1111–1141, and 15 indels in positions 1142–1156, were observed in lovF of A. terreus ATCC 20541, ML-44, and ASM-1 (Fig. 6; Supplementary Fig. S9).
Figure 5.

Genes involved in the biosynthesis of lovastatin (A. terreus strains), monacolin K (M. pilosus), and compactin (P. citrinum).
Figure 6.

Sequence alignment showing the amino acid sequence coded by lovF. A deletion and an indel region were observed. A. terreus M6925, ATCC 20541, ML-44, and ASM-1 lacked 31 amino acids (red line) and had 15 indels (green line) in lovF (position: 1111–1156).
Discussion
In this study, we present a high-quality whole genome assembly of A. terreus ATCC 20541 containing 10,410 protein coding genes with a total genome size of ~ 30 Mb. The largest scaffold size was 3.82 Mb and the N50 value of scaffolds was 1.58 Mb. In addition, all publicly available whole genome sequences of A. terreus strains were retrieved and further compared. The total genome size (28.0–31.8 Mb) and GC content (52.1–53.1%) of eleven A. terreus strains are similar to all Aspergillus genomes (genome sizes are 29–41 Mb and GC contents vary from 43 to 53%)36,39. Phylogenetic analysis indicates that A. terreus ATCC 20541 is positioned in a major clade including A. terreus NIH2624, 45A, M6925, w25, ASM-1, ML-44, and T3 Kankrej, so the genomic relationship in A. terreus ATCC 20541 is closely related to most A. terreus strains, especially A. terreus NIH2624 and 45A. Most A. terreus strains are divergent from A. terreus ATCC 20542, IFO 6365, and TN-484 (Supplementary Fig. S4).
Several Aspergillus spp. can degrade plant biomass polysaccharides which is a major carbon source for many fungal species. The fungal CAZymes family has been utilized in industrial applications by the hydrolysis of plant biomass for the subsequent production of biofuels and high-value biochemicals48. Our results indicate that A. terreus strains secrete high levels of pectinases, so they are capable of degrading pectin which is a complex polysaccharide in plant cell walls49. Abundant genes in GH31 (starch), GH32 (inulin), GH28 (pectin), GH78 (pectin), PL1 (pectin), and AA9 (cellulose) of the CAZymes family were observed, which ranged from 10 to 11, 6 to 11, 7 to 8, 5 to 7, 7, and 10 genes, respectively (Supplementary Table S2).
Fungi have diversified CYP450 families, which contribute to survival strategies, to adapt to varying environments, and to facilitate detoxification50. The CYP52 family is the largest (40 to 44 genes) in the A. terreus strains (Table 2). This CYP takes part in degrading n-hexadecane (HXD) when it is used as the sole carbon source in Aspergillus sp. RFC-151. Previous studies reported that the CYP504 family including 239 proteins can catalyze phenylacetate catabolism and may be related to the diversified functions of xenobiotic compounds detoxification52,53. In addition, the CYP58 family including 274 proteins in A. terreus strains was related to aflatoxin biosynthesis, and the CYP58 family also can be found in A. flavus and A. parasiticusv54. CYP51 family (CYP51A, CYP51B, and CYP51C) including 720 proteins in A. flavus is an antifungal drug target for controlling pest and fungal plant diseases50. Interestingly, the CYP51 family, the target of azole drugs, was found in different A. terreus strains in this study. Several studies have already detailed the role of CYP51 variation in azole resistance in A. terreus55 and have identified three CYP51 paralogs56.
Apart from A. terreus IFO 6365 and TN-484, other A. terreus strains contained the BGC of siderophore production. Siderophore is one of the metabolites secreted by A. terreus57 and A. fumigatus58. These metabolites sequester iron from the host microbes and cycle this metal nutrient to themself59. In addition, A. terreus IFO 6365 and TN-484 are itaconic acid producers, and LovF, LovB, GedC, PgnA, AtaP, and AnaPS are not found in their core of SM proteins. A. terreus ATCC 20541 lacks an SM AnaPS, which participates in acetylaszonalenin production60. A. terreus strains produce several interesting bioactive compounds such as butyrolactones (BtyA), asterriquinone (AtqA), terreic acid (AtX), citreoviridin (CtvA), terrein (TerA), Trt4, and geodin (GedC) (Supplementary Fig. S7), which could be potential sources of biosynthetic enzymes, antibiotics, or antitumor in the future61–67.
We also compared the lovastatin BGC in A. terreus strains. The lovastatin BGC including lovA, lovB, lovC, lovD, lovE, lovF, lovG, lovI, and IvrA in A. terreus strains are similar to the monacolin K BGC in M. pilosus BCRC 38072 and to the compactin BGC in P. citrinum (Fig. 5). The hypolipidemic agents monacolin K and compactin (mevastatin) serve the same function as lovastatin, which act as competitive inhibitors of HMG-CoA reductases68,69. Additionally, we found that there were significantly different types of lovI gene variants among the A. terreus strains. The lovI gene is a hypothetical protein HFD88_005927 composed of 517 amino acid residues in A. terreus ATCC 20542. lovI gene had approximately 400 amino acid deletions in A. terreus NIH2624, ASM-1, ML-44 and 45A against the corresponding gene of A. terreus ATCC 20542. Notably, lovD and lvrA are fused into a single gene in A. terreus ATCC 20542, which is different from other A. terreus stains. A similar phenomenon also showed in Bacillus subtilis strains. For example, ppsB 3′ and ppsB 5′ are fused into a single ppsB in plipastatin BGC of B. subtilis, which could inhibit the Fusarium species (F. oxysporum and F. graminearum)70. This mechanism in A. terreus still requires further investigations.
In conclusion, a high-quality whole genome assembly of A. terreus ATCC 20541 was presented in this study. Compared with genomic sequences among A. terreus strains, diversified conserved BGCs were identified. Amino acid deletions and indels observed in the lovB and lovF genes played an important role in the lovastatin formation. Our comparative analyses may motivate further investigation to study the function of BGCs associated with valuable SMs, and understand the genomic diversity in A. terreus.
Methods
Strains and genomic DNA isolation
A. terreus ATCC 20541 used in this study was collected from American Type Culture Collection (Manassas, VA, USA). The strain was cultured on potato dextrose agar (PDA) at 28 °C. The A. terreus ATCC 20541 genomic DNA was extracted as described in Aamir71 with the following modifications. First, the harvested fungal tissue was homogenized in lysis buffer (50 mM EDTA, 3% SDS, 100 mM Tris–HCl, and pH 8). The homogenate was centrifuged at 13,000 rpm for 10 min, and the supernatant was then mixed with an equal volume of phenol: chloroform: Isoamyl alcohol (25:24:1). After centrifugation, the aqueous layer was collected in a new eppendorf tube, followed by ethanol precipitation and centrifugation. The DNA pellet was dissolved in Tris–HCl buffer, pH 8.5. The genomic DNA quality was assessed on an Agilent 2100 Bioanalyzer (Agilent Technologies, USA), and DNA integrity was checked using 1% agarose gel electrophoresis then for sequencing by Illumina sequencing platform.
Genome sequencing, assembly and quality assessment
The DNA library was constructed using the Illumina TruSeq Nano DNA High Throughput Library Prep Kit (Illumina, USA) according to the manufacturer’s instructions. Whole-genome shotgun sequencing was performed using Novaseq 6000 platforms (Illumina, USA) with 2 × 150 paired-end reads. The raw reads were trimmed by removing adaptor sequences and low-quality sequences with Q < 20 using Trimmomatic v.0.39 with parameters of “SLIDINGWINDOW:4:20”72, and MultiQC v.1.273 summarized the sequence of quality control. The de novo genome assembly was carried out with SPAdes v.3.10.174, and the assembly was polished with Pilon v1.2375. The assembly was evaluated using QUAST v.4.576, and the gVolante was used to assess the completeness of the genome assembly, according to the Eurotiomycetes database77.
Genome annotation
Genome assemblies of ten A. terreus strains were obtained from NCBI (access date: November 2021). All the A. terreus strains were annotated using AUGUSTUS v. 3.4.078 with “Aspergillus terreus” as a training dataset, and the parameters were based on Takahashi et al.29. Ribosomal RNA (rRNA) and transfer RNA (tRNA) genes were predicted using barrnap v. 0.9 and tRNAscan-SE v. 2.0.979. Signal peptide and transmembrane helices were functionally annotated using SignalP v. 5.0b80 and Phobius server81 (access date: November 2021). The predicted proteins were assigned by the eukaryotic orthologous group (KOG) using the webMGA server82 with a cut-off e-value ≤ 1e − 5. KEGG Automated Annotation Server (KAAS) was used for pathway mapping of Eurotiomycetes species with a bi-directional best hit (BBH)83,84. Gene ontology (GO) was predicted using PANNZER web server85. Proteinortho program v. 6.0.3186 with “blastp” function was used to detect orthologous genes within the A. terreus strains.
CAZymes and CYP family in A. terreus strains
CAZymes annotation was performed using dbCAN2 meta server38 (access date: November 2021) with HAMMER, DIAMOND, and eCAMI. The cytochrome P450 gene family classifications in A. terreus strains were performed based on A. terreus NIH2624 using biocatnet CYPED v 6.087 with cut off e-value ≤ 1e − 5 and identity > 40%.
Construction of phylogenetic trees
Sequence similarity, the Average Nucleotide Identity (ANI) values of A. terreus strains were calculated using PYANI v.0.2.11 based on BLAST + program88. The phylogenetic tree of A. terreus strains was constructed using R language.
Secondary metabolite gene clusters and amino acid variants detection
The whole genome sequence of A. terreus strains was analyzed by using the antiSMASH v. 6.0.1 (fungal version)41 with default parameters to identify the potential secondary metabolite gene clusters. Amino acid variants were identified by pair-wise alignment of conservation gene using ClustalW in Bioedit v. 7.2.6.
Ethics approval and consent to participate
This study does not contain any experiments with human and animals performed by all the authors.
Supplementary Information
Acknowledgements
Whole-genome shotgun sequencing was performed at the Genomics BioSci & Tech Co., Ltd. (Taipei, Taiwan).
Author contributions
H.Y.W., U.H.M., and H.Y.T. analyzed the data and prepared the manuscript. H.Y.T., U.H.M., and F.R.C. conceived the investigation. All authors have read and agreed to the manuscript.
Funding
This study was supported by the Ministry of Science and Technology, Taiwan (MOST 108-2320-B-110-007-MY3).
Data availability
The Whole Genome Shotgun project has been deposited at GenBank in NCBI. The accession number is JANHGT000000000 and BioProject is PRJNA861866. The deposited data will be released upon acceptance. The submission confirmation letter sent by GenBank NCBI is attached in the related file.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-022-27311-7.
References
- 1.De Aguirre L, et al. Rapid differentiation of Aspergillus species from other medically important opportunistic molds and yeasts by PCR-enzyme immunoassay. J. Clin. Microbiol. 2004;42:3495–3504. doi: 10.1128/JCM.42.8.3495-3504.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Keller NP. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2019;17:167. doi: 10.1038/s41579-018-0121-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Magnuson, J. K. & Lasure, L. L. Organic acid production by filamentous fungi. Adv. Fungal Biotechnol. Ind. Agric. Med. 307–340 (2004).
- 4.Drula E, et al. The carbohydrate-active enzyme database: Functions and literature. Nucleic Acids Res. 2022;50:D571–D577. doi: 10.1093/nar/gkab1045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Nordberg H, et al. The genome portal of the Department of Energy Joint Genome Institute: 2014 updates. Nucleic Acids Res. 2014;42:D26–D31. doi: 10.1093/nar/gkt1069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Medema MH, Osbourn A. Computational genomic identification and functional reconstitution of plant natural product biosynthetic pathways. Nat. Prod. Rep. 2016;33:951–962. doi: 10.1039/C6NP00035E. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Nützmann H-W, Scazzocchio C, Osbourn A. Metabolic gene clusters in eukaryotes. Annu. Rev. Genet. 2018;52:159–183. doi: 10.1146/annurev-genet-120417-031237. [DOI] [PubMed] [Google Scholar]
- 8.Mózsik, L., Iacovelli, R., Bovenberg, R. A. L. & Driessen, A. J. M. Transcriptional activation of biosynthetic gene clusters in filamentous fungi. Front. Bioeng. Biotechnol.10, (2022). [DOI] [PMC free article] [PubMed]
- 9.Palonen EK, et al. Transcriptomic complexity of Aspergillus terreus velvet gene family under the influence of butyrolactone I. Microorganisms. 2017;5:12. doi: 10.3390/microorganisms5010012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kithsiri Wijeratne EM, et al. Cytotoxic constituents of Aspergillus terreus from the rhizosphere of opuntia versicolor of the Sonoran Desert. J. Nat. Prod. 2003;66:1567–1573. doi: 10.1021/np030266u. [DOI] [PubMed] [Google Scholar]
- 11.Abdel-Azeem, A. M. et al. Biodiversity of the genus Aspergillus in different habitats. New Futur. Dev. Microb. Biotechnol. Bioeng. Aspergillus Syst. Prop. Appl. 3–28 (2016).
- 12.DeLucca AJ. Harmful fungi in both agriculture and medicine. Rev. Iberoam. Micol. 2007;24:3–13. [PubMed] [Google Scholar]
- 13.Kü Ck U, Bloemendal S, Teichert I. Putting fungi to work: Harvesting a cornucopia of drugs, toxins, and antibiotics. PLoS Pathog. 2014;10:e1003950. doi: 10.1371/journal.ppat.1003950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Iwen PC, Rupp ME, Langnas AN, Reed EC, Hinrichs SH. Invasive pulmonary aspergillosis due to Aspergillus terreus: 12-year experience and review of the literature. Clin. Infect. Dis. 1998;26:1092–1097. doi: 10.1086/520297. [DOI] [PubMed] [Google Scholar]
- 15.Iwen PC, et al. Nosocomial invasive aspergillosis in lymphoma patients treated with bone marrow or peripheral stem cell transplants. Infect. Control Hosp. Epidemiol. 1993;14:131–139. doi: 10.2307/30148476. [DOI] [PubMed] [Google Scholar]
- 16.Saha BC, Kennedy GJ, Bowman MJ, Qureshi N, Nichols NN. Itaconic acid production by Aspergillus terreus from glucose up to pilot scale and from corn stover and wheat straw hydrolysates using new manganese tolerant medium. Biocatal. Agric. Biotechnol. 2022;43:102418. doi: 10.1016/j.bcab.2022.102418. [DOI] [Google Scholar]
- 17.Nemestóthy N, et al. Carbohydrate to itaconic acid conversion by Aspergillus terreus and the evaluation of process monitoring based on the measurement of CO2. Waste Biomass Valoriz. 2020;11:1069–1075. doi: 10.1007/s12649-019-00729-3. [DOI] [Google Scholar]
- 18.Hasan H, et al. Increasing lovastatin production by re-routing the precursors flow of Aspergillus terreus via metabolic engineering. Mol. Biotechnol. 2022;64:90–99. doi: 10.1007/s12033-021-00393-w. [DOI] [PubMed] [Google Scholar]
- 19.Barrios-González J, Pérez-Sánchez A, Bibián ME. New knowledge about the biosynthesis of lovastatin and its production by fermentation of Aspergillus terreus. Appl. Microbiol. Biotechnol. 2020;104:8979–8998. doi: 10.1007/s00253-020-10871-x. [DOI] [PubMed] [Google Scholar]
- 20.Sharma R, Singh Kocher G, Singh Bhogal R, Oberoi HS. Cellulolytic and xylanolytic enzymes from thermophilic Aspergillus terreus RWY. J. Basic Microbiol. 2014;54:1367–1377. doi: 10.1002/jobm.201400187. [DOI] [PubMed] [Google Scholar]
- 21.Pierce BC, Agger JW, Zhang Z, Wichmann J, Meyer AS. A comparative study on the activity of fungal lytic polysaccharide monooxygenases for the depolymerization of cellulose in soybean spent flakes. Carbohydr. Res. 2017;449:85–94. doi: 10.1016/j.carres.2017.07.004. [DOI] [PubMed] [Google Scholar]
- 22.Narra M, Dixit G, Divecha J, Madamwar D, Shah AR. Production of cellulases by solid state fermentation with Aspergillus terreus and enzymatic hydrolysis of mild alkali-treated rice straw. Bioresour. Technol. 2012;121:355–361. doi: 10.1016/j.biortech.2012.05.140. [DOI] [PubMed] [Google Scholar]
- 23.Chantasingh D, Pootanakit K, Champreda V, Kanokratana P, Eurwilaichitr L. Cloning, expression, and characterization of a xylanase 10 from Aspergillus terreus (BCC129) in Pichia pastoris. Protein Expr. Purif. 2006;46:143–149. doi: 10.1016/j.pep.2005.09.013. [DOI] [PubMed] [Google Scholar]
- 24.Varga J, et al. Evolutionary relationships among Aspergillus terreus isolates and their relatives. Antonie Van Leeuwenhoek. 2005;88:141–150. doi: 10.1007/s10482-005-3870-6. [DOI] [PubMed] [Google Scholar]
- 25.Kanamasa S, et al. Draft genome sequence of Aspergillus terreus high-itaconic-acid-productivity mutant TN-484. Microbiol. Resour. Anno. 2019;8:e01170–e1219. doi: 10.1128/MRA.01170-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Palanivel M, et al. Whole-genome sequencing of Aspergillus terreus species complex. Mycopathologia. 2020;185:405–408. doi: 10.1007/s11046-020-00433-0. [DOI] [PubMed] [Google Scholar]
- 27.Ryngajłło M, Boruta T, Bizukojć M. Complete genome sequence of lovastatin producer Aspergillus terreus ATCC 20542 and evaluation of genomic diversity among A. terreus strains. Appl. Microbiol. Biotechnol. 2021;105:1615–1627. doi: 10.1007/s00253-021-11133-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Savitha J, Bhargavi SD, Praveen VK. Complete genome sequence of soil fungus Aspergillus terreus (KM017963), a potent lovastatin producer. Genome Announc. 2016;4:e00491–e516. doi: 10.1128/genomeA.00491-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Takahashi H, et al. Draft genome sequence of the Aspergillus terreus high-itaconic-acid-productivity strain IFO6365. Microbiol. Resour. Announc. 2020;9:e00080–e120. doi: 10.1128/MRA.00080-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wu, C. et al. Aspulvinones suppress postprandial hyperglycemia as potent α-glucosidase inhibitors from Aspergillus terreus ASM-1. Front. Chem.9, (2021). [DOI] [PMC free article] [PubMed]
- 31.Chang PK, Horn BW, Dorner JW. Sequence breakpoints in the aflatoxin biosynthesis gene cluster and flanking regions in nonaflatoxigenic Aspergillus flavus isolates. Fungal Genet. Biol. 2005;42:914–923. doi: 10.1016/j.fgb.2005.07.004. [DOI] [PubMed] [Google Scholar]
- 32.Chiara M, et al. Genome sequencing of multiple isolates highlights subtelomeric genomic diversity within Fusarium fujikuroi. Genome Biol. Evol. 2015;7:3062–3069. doi: 10.1093/gbe/evv198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Drott MT, et al. Diversity of secondary metabolism in Aspergillus nidulans clinical isolates. Msphere. 2020;5:e00156–e220. doi: 10.1128/mSphere.00156-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lind AL, et al. Drivers of genetic diversity in secondary metabolic gene clusters within a fungal species. PLoS Biol. 2017;15:1–26. doi: 10.1371/journal.pbio.2003583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Schumacher J, et al. A functional bikaverin biosynthesis gene cluster in rare strains of Botrytis cinerea is positively controlled by VELVET. PLoS ONE. 2013;8:e53729. doi: 10.1371/journal.pone.0053729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kjærbølling I, et al. A comparative genomics study of 23 Aspergillus species from section Flavi. Nat. Commun. 2020;11:1106. doi: 10.1038/s41467-019-14051-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kjærbølling I, Vesth T, Andersen MR. Resistance gene-directed genome mining of 50 Aspergillus species. mSystems. 2019;4:e00085–e119. doi: 10.1128/mSystems.00085-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Zhang H, et al. DbCAN2: A meta server for automated carbohydrate-active enzyme annotation. Nucleic Acids Res. 2018;46:W95–W101. doi: 10.1093/nar/gky418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.DeVries RP, et al. Comparative genomics reveals high biological diversity and specific adaptations in the industrially and medically important fungal genus Aspergillus. Genome Biol. 2017;18:28. doi: 10.1186/s13059-017-1151-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Shin J, Kim J-E, Lee Y-W, Son H. Fungal cytochrome P450s and the P450 complement (CYPome) of Fusarium graminearum. Toxins (Basel). 2018;10:112. doi: 10.3390/toxins10030112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Blin K, et al. AntiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021;49:W29–W35. doi: 10.1093/nar/gkab335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Wen Q, et al. An overview of Monascus fermentation processes for monacolin K production. Open Chem. 2020;18:10–21. doi: 10.1515/chem-2020-0006. [DOI] [Google Scholar]
- 43.Chen Y-P, et al. Identification of the high-yield monacolin K strain from Monascus spp. and its submerged fermentation using different medicinal plants. Bot. Stud. 2022;63:20. doi: 10.1186/s40529-022-00351-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Mulder KCL, et al. Lovastatin production: From molecular basis to industrial process optimization. Biotechnol. Adv. 2015;33:648–665. doi: 10.1016/j.biotechadv.2015.04.001. [DOI] [PubMed] [Google Scholar]
- 45.Liu A, et al. Investigation of citrinin and monacolin K gene clusters variation among pigment producer Monascus species. Fungal Genet. Biol. 2022;160:103687. doi: 10.1016/j.fgb.2022.103687. [DOI] [PubMed] [Google Scholar]
- 46.Zhang Y, et al. An overview on the biosynthesis and metabolic regulation of monacolin K/lovastatin. Food Funct. 2020;11:5738–5748. doi: 10.1039/D0FO00691B. [DOI] [PubMed] [Google Scholar]
- 47.Guo CJ, Wang CCC. Recent advances in genome mining of secondary metabolites in Aspergillus terreus. Front. Microbiol. 2014;5:717. doi: 10.3389/fmicb.2014.00717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Alazi E, Ram AFJ. Modulating transcriptional regulation of plant biomass degrading enzyme networks for rational design of industrial fungal strains. Front. Bioeng. Biotechnol. 2018;6:133. doi: 10.3389/fbioe.2018.00133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Benoit I, et al. Degradation of different pectins by fungi: Correlations and contrasts between the pectinolytic enzyme sets identified in genomes and the growth on pectins of different origin. BMC Genom. 2012;13:1–11. doi: 10.1186/1471-2164-13-321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Pratiwi RA, Yahya NSW, Chi Y. Bio function of Cytochrome P450 on fungus: A review. IOP Conf. Ser. Earth Environ. Sci. 2022;959:012023. doi: 10.1088/1755-1315/959/1/012023. [DOI] [Google Scholar]
- 51.Al-Hawash AB, et al. Biodegradation of n-hexadecane by Aspergillus sp. RFC-1 and its mechanism. Ecotoxicol. Environ. Saf. 2018;164:398–408. doi: 10.1016/j.ecoenv.2018.08.049. [DOI] [PubMed] [Google Scholar]
- 52.Porter TD, Coon MJ. Cytochrome P-450: Multiplicity of isoforms, substrates, and catalytic and regulatory mechanisms. J. Biol. Chem. 1991;266:13469–13472. doi: 10.1016/S0021-9258(18)92717-1. [DOI] [PubMed] [Google Scholar]
- 53.Rodríguez-Sáiz M, et al. Reduced function of a phenylacetate-oxidizing cytochrome p450 caused strong genetic improvement in early phylogeny of penicillin-producing strains. J. Bacteriol. 2001;183:5465–5471. doi: 10.1128/JB.183.19.5465-5471.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Črešnar B, Petrič Š. Cytochrome P450 enzymes in the fungal kingdom. Biochim. Biophys. Acta Proteins Proteom. 2011;1814:29–35. doi: 10.1016/j.bbapap.2010.06.020. [DOI] [PubMed] [Google Scholar]
- 55.Zoran, T. et al. Azole-resistance in Aspergillus terreus and related species: An emerging problem or a rare phenomenon? Front. Microbiol.9, (2018). [DOI] [PMC free article] [PubMed]
- 56.Pérez-Cantero A, López-Fernández L, Guarro J, Capilla J. Azole resistance mechanisms in Aspergillus: Update and recent advances. Int. J. Antimicrob. Agents. 2020;55:105807. doi: 10.1016/j.ijantimicag.2019.09.011. [DOI] [PubMed] [Google Scholar]
- 57.Waqas M, et al. Endophytic infection alleviates biotic stress in sunflower through regulation of defence hormones, antioxidants and functional amino acids. Eur. J. Plant Pathol. 2015;141:803–824. doi: 10.1007/s10658-014-0581-8. [DOI] [Google Scholar]
- 58.Haas H. Fungal siderophore metabolism with a focus on Aspergillus fumigatus. Nat. Prod. Rep. 2014;31:1266–1276. doi: 10.1039/C4NP00071D. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Khasheii B, Mahmoodi P, Mohammadzadeh A. Siderophores: Importance in bacterial pathogenesis and applications in medicine and industry. Microbiol. Res. 2021;250:126790. doi: 10.1016/j.micres.2021.126790. [DOI] [PubMed] [Google Scholar]
- 60.Yin WB, Grundmann A, Cheng J, Li SM. Acetylaszonalenin biosynthesis in Neosartorya fischeri: Identification of the biosynthetic gene cluster by genomic mining and functional proof of the genes by biochemical investigation. J. Biol. Chem. 2009;284:100–109. doi: 10.1074/jbc.M807606200. [DOI] [PubMed] [Google Scholar]
- 61.Bräse S, Encinas A, Keck J, Nising CF. Chemistry and biology of mycotoxins and related fungal metabolites. Chem. Rev. 2009;109:3903–3990. doi: 10.1021/cr050001f. [DOI] [PubMed] [Google Scholar]
- 62.Chang H-Y, et al. Ectopic ATP synthase blockade suppresses lung adenocarcinoma growth by activating the unfolded protein response. Cancer Res. 2012;72:4696–4706. doi: 10.1158/0008-5472.CAN-12-0567. [DOI] [PubMed] [Google Scholar]
- 63.Kaji A, Saito R, Nomura M, Miyamoto K, Kiriyama N. Relationship between the structure and cytotoxic activity of asterriquinone, an antitumor metabolite of Aspergillus terreus, and its alkyl ether derivatives. Biol. Pharm. Bull. 1998;21:945–949. doi: 10.1248/bpb.21.945. [DOI] [PubMed] [Google Scholar]
- 64.Liao W-Y, et al. Asperjinone, a nor-neolignan, and terrein, a suppressor of ABCG2-expressing breast cancer cells, from thermophilic Aspergillus terreus. J. Nat. Prod. 2012;75:630–635. doi: 10.1021/np200866z. [DOI] [PubMed] [Google Scholar]
- 65.Wu CJ, et al. Terretonin D1, a new meroterpenoid from marine-derived Aspergillus terreus ML-44. Nat. Prod. Res. 2019;33:2262–2265. doi: 10.1080/14786419.2018.1493583. [DOI] [PubMed] [Google Scholar]
- 66.Yavlovich A, et al. Ectopic ATP synthase facilitates transfer of HIV-1 from antigen-presenting cells to CD4 target cells. J. Am. Soc. Hematol. 2012;12:1246–1253. doi: 10.1182/blood-2011-12-399063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Rao KV, et al. Butyrolactones from Aspergillus terreus. Chem. Pharm. Bull. 2000;48:559–562. doi: 10.1248/cpb.48.559. [DOI] [PubMed] [Google Scholar]
- 68.Endo A. Discovery and development of statins. Nat. Prod. Commun. 2017;12:1153–1156. [Google Scholar]
- 69.Endo A. Monacolin K, a new hypocholesterolemic agent that specifically inhibits 3-hydroxy-3-methylglutaryl coenzyme A reductase. J. Antibiot. (Tokyo) 1980;33:334–336. doi: 10.7164/antibiotics.33.334. [DOI] [PubMed] [Google Scholar]
- 70.Kiesewalter HT, et al. Genomic and chemical diversity of Bacillus subtilis secondary metabolites against plant pathogenic fungi. mSystems. 2021;6:1–15. doi: 10.1128/mSystems.00770-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Aamir S. A rapid and efficient method of fungal genomic DNA extraction, suitable for PCR based molecular methods. Plant Pathol. Quar. 2015;5:74–81. doi: 10.5943/ppq/5/2/6. [DOI] [Google Scholar]
- 72.Bolger AM, Lohse M, Usadel B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics. 2014;30:2114–2120. doi: 10.1093/bioinformatics/btu170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Ewels P, Ns Magnusson M, Lundin S, Aller MK. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics. 2016;32:3047–3048. doi: 10.1093/bioinformatics/btw354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Nurk, S. et al. Assembling genomes and mini-metagenomes from highly chimeric reads. Res. Comput. Mol. Biol. 158–170 (2013). [DOI] [PMC free article] [PubMed]
- 75.Walker BJ, et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE. 2014;9:e112963. doi: 10.1371/journal.pone.0112963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Gurevich A, Saveliev V, Vyahhi N, Tesler G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics. 2013;29:1072–1075. doi: 10.1093/bioinformatics/btt086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Nishimura, O., Hara, Y. & Kuraku, S. Evaluating genome assemblies and gene models using gVolante. Methods Mol. Biol. 247–256 (2019). [DOI] [PubMed]
- 78.Stanke M, et al. AUGUSTUS: Ab initio prediction of alternative transcripts. Nucleic Acids Res. 2006;34:W435–W439. doi: 10.1093/nar/gkl200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Chan PP, Lin BY, Mak AJ, Lowe TM. TRNAscan-SE 2.0: Improved detection and functional classification of transfer RNA genes. Nucleic Acids Res. 2021;49:9077–9096. doi: 10.1093/nar/gkab688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Almagro Armenteros JJ, et al. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019;37:420–423. doi: 10.1038/s41587-019-0036-z. [DOI] [PubMed] [Google Scholar]
- 81.Käll L, Krogh A, Sonnhammer ELL. Advantages of combined transmembrane topology and signal peptide prediction-the Phobius web server. Nucleic Acids Res. 2007;35:429–432. doi: 10.1093/nar/gkm256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Wu S, Zhu Z, Fu L, Niu B, Li W. WebMGA: A customizable web server for fast metagenomic sequence analysis. BMC Genom. 2011;12:444. doi: 10.1186/1471-2164-12-444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Moriya Y, Itoh M, Okuda S, Yoshizawa AC, Kanehisa M. KAAS: An automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 2007;35:W182–W185. doi: 10.1093/nar/gkm321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Kanehisa, M., Furumichi, M., Sato, Y., Kawashima, M. & Ishiguro-Watanabe, M. KEGG for taxonomy-based analysis of pathways and genomes. Nucleic Acids Res. gkac963 (2022). [DOI] [PMC free article] [PubMed]
- 85.Törönen P, Holm L. PANNZER—A practical tool for protein function prediction. Protein Sci. 2022;31:118–128. doi: 10.1002/pro.4193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Lechner M, et al. Proteinortho: Detection of (Co-)orthologs in large-scale analysis. BMC Bioinform. 2011;12:1–9. doi: 10.1186/1471-2105-12-124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Gricman Ł, Vogel C, Pleiss J. Identification of universal selectivity-determining positions in cytochrome P450 monooxygenases by systematic sequence-based literature mining. Proteins Struct. Funct. Bioinform. 2015;83:1593–1603. doi: 10.1002/prot.24840. [DOI] [PubMed] [Google Scholar]
- 88.Pritchard L, Glover RH, Humphris S, Elphinstone JG, Toth IK. Genomics and taxonomy in diagnostics for food security: Soft-rotting enterobacterial plant pathogens. Anal. Methods. 2016;8:12–24. doi: 10.1039/C5AY02550H. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The Whole Genome Shotgun project has been deposited at GenBank in NCBI. The accession number is JANHGT000000000 and BioProject is PRJNA861866. The deposited data will be released upon acceptance. The submission confirmation letter sent by GenBank NCBI is attached in the related file.