

Abstract
Simple Summary
Over the years, alternative and complementary medicine have garnered much attention all across the globe. To this end, phytochemicals have intriguing potential against myriads of disease conditions; nevertheless, as of yet, the molecular intricacies for their therapeutic potential is incompletely understood. It is widely acknowledged that proteomics technology has been ex-plored as a reliable approach to understand the molecular intricacies related to phytochemi-cal-based therapeutic interventions. Reckoning with this, the present review provides an overview of the proteomics studies performed to unravel the underlying molecular intricacies of various phytochemicals such as Curcumin, Genistein, and Tanshinone IIA.
Abstract
Phytochemicals possess various intriguing pharmacological properties against diverse pathological conditions. Extensive studies are on-going to understand the structural/functional properties of phytochemicals as well as the molecular mechanisms of their therapeutic function against various disease conditions. Phytochemicals such as curcumin (Cur), genistein (Gen), and tanshinone-IIA (Tan IIA) have multifaceted therapeutic potentials and various efforts are in progress to understand the molecular dynamics of their function with different tools and technologies. Cur is an active lipophilic polyphenol with pleiotropic function, and it has been shown to possess various intriguing properties including antioxidant, anti-inflammatory, anti-microbial, anticancer, and anti-genotoxic properties besides others beneficial properties. Similarly, Gen (an isoflavone) exhibits a wide range of vital functions including antioxidant, anti-inflammatory, pro-apoptotic, anti-proliferative, anti-angiogenic activities etc. In addition, Tan IIA, a lipophilic compound, possesses antioxidant, anti-angiogenic, anti-inflammatory, anticancer activities, and so on. Over the last few decades, the field of proteomics has garnered great momentum mainly attributed to the recent advancement in mass spectrometry (MS) techniques. It is envisaged that the proteomics technology has considerably contributed to the biomedical research endeavors lately. Interestingly, they have also been explored as a reliable approach to understand the molecular intricacies related to phytochemical-based therapeutic interventions. The present review provides an overview of the proteomics studies performed to unravel the underlying molecular intricacies of various phytochemicals such as Cur, Gen, and Tan IIA. This in-depth study will help the researchers in better understanding of the pharmacological potential of the phytochemicals at the proteomics level. Certainly, this review will be highly instrumental in catalyzing the translational shift from phytochemical-based biomedical research to clinical practice in the near future.
Keywords: phytochemicals, curcumin, genistein, tanshinone, proteomics, diseases, therapeutic intervention
1. Introduction
In recent years, alternative and/or complementary medicine have garnered much attention across the globe [1,2,3,4,5,6]. Phytochemicals are biologically active substances that embody various intriguing pharmacological properties against diverse pathological conditions including microbial infections, metabolic disorders, cancers, degenerative diseases, etc. The therapeutic potential of phytochemicals has been extensively investigated over the last few decades, and various reports have highlighted their interesting biological and therapeutic potentials [7,8,9,10,11]. Nevertheless, the molecular intricacies for their therapeutic potential are still an area of active research. Among various phytochemicals, curcumin (Cur), genistein (Gen), tanshinone IIA (Tan IIA), allicin, eugenol, apigenin, lycopene, anthocyanin, capsaicin, and shogaols share a history of high repute. Table 1 delineates the pharmacological properties, molecular functions, and therapeutic applications of various phytochemicals against diverse pathophysiological conditions. Of note, these phytochemicals embody diverse chemical space for drug discovery; to this end, various comprehensive online databases of phytochemicals have been developed, which enables computational approaches towards natural product-based drug discovery. These include TCM@Taiwan [12], KNAPSACK [13], TCMID [14], CVDHD [15], Nutrichem [16], TCM-Mesh [17], IMPPAT [18], etc. Interestingly, lately various newer therapeutic chemical moieties have been deciphered in pharmacology; interestingly, out of these identified therapeutic chemicals, approximately 50% are phytochemicals in nature, which depicts their importance in therapeutic interventions [19]. It is envisaged that dietary phytochemicals are extensively explored for therapeutic interventions due to a wide variety of reasons, including ease of availability, lower toxicity issues, characteristic biological effects, cost-effectiveness, and diversity of chemical components in plants. Several clinical studies have demonstrated a strong correlation between dietary intake of phytochemicals and reduced risk of cancer development and relapse [20,21,22,23,24]. A representative figure highlighting some of the therapeutic potentials of Cur, Gen, and Tan IIA are depicted in Figure 1.
Table 1.
Representative table highlighting various phytochemicals along with their pharmacological properties and therapeutic importance.
| S. No. | PhytochemicalName | Appearance | Chemical Structure | Molecular Weight, Chemical Formula, Pubchem CID, and IUPAC Name | Prospective Pharmacological Properties |
|---|---|---|---|---|---|
| 1. | Curcumin | Bright yellow-orange |
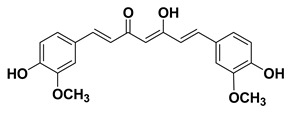
|
Mol wt: 368.38 g/mol Chemical formula: C21H20O6 Pubchem CID:969516 IUPAC name: 1,7-Bis(4-hydroxy-3-methoxyphenyl)hepta-1,6-diene-3,5-dione |
It acts as an antioxidant [25], anti-inflammatory [26] anti-bacterial [27], anti-fungal [28] antiviral [29], and anti-neoplastic agent [30]. It exhibits phototoxic and photodynamic activities [31,32,33], acts as a cyclo-oxygenase inhibitor [34], lipoxygenase inhibitor [35], iron chelator [36], immunomodulator [37], and neuroprotective agents [38,39,40,41]. |
| 2. | Genistein | Yellow |
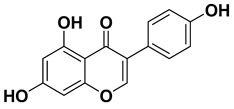
|
Mol wt: 270.24 g/mol Chemical formula: C15H10O5 Pubchem CID: 5280961 IUPAC name: 5,7-dihydroxy-3-(4-hydroxyphenyl)chromen-4-one |
It has antioxidant [42], anti-inflammatory and immunosuppressive activities [43]; it acts as anti-microbial agent [44], it embodies anti-carcinogenic and anti-metastatic properties [45]. It also acts as a phytoestrogen and a protein tyrosine kinase inhibitor [46], and neuroprotective agent [47,48,49,50]. |
| 3. | Tanshinone IIA | Red |
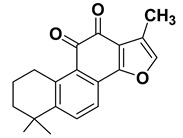
|
Mol wt: 294.3 g/mol Chemical formula: C19H18O3 Pubchem CID: 164676 IUPAC name: 1,6,6-trimethyl-8,9-dihydro-7H-naphtho [1,2-g] [1]benzofuran-10,11-dione |
It embodies antioxidant [51], anti -inflammatory [52], anti-microbial [53], anti-cancer [54], anti-angiogenic [55], and anti-adipogenic properties [56], and embodies neuroprotective properties [57,58,59,60]. |
| 4. | Allicin | Slightly yellow |
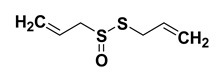
|
Mol wt: 162.3 g/mol Chemical formula: C6H10OS2, Pubchem CID: 65036 IUPAC name: 3-prop-2-enyl sulfinyl sulfanyl prop-1-ene |
It embodies free radical scavenging properties, viz., anti-oxidant [61], anti-bacterial [62,63,64], anti-fungal [63], and anti-viral properties [65]. It exhibits antihypertensive [66] and neuroprotective properties [67] and acts as hypo-lipidemic and hypo-glycemic [68,69] and anti-cancer agent [70,71,72]. |
| 5. | Eugenol | Pale yellow |
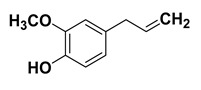
|
Mol wt: 164.2 g/mol Chemical formula: C10H12O2 Pubchem CID: 3314 IUPAC name: 2-methoxy-4-prop-2-enyl phenol |
It embodies antioxidant [73], anti-inflammatory [74], anti-microbial [75], anti-tumor [76,77,78,79], anti-mutagenic [76,77,78,79], anti-allergic [80], antipyretic [81] and analgesic characteristics [80]. It is a 5-lipoxygenase inhibitor [76], anti-hypercholesterolemic, and anti-atherogenic potential [82], antidiabetic [83], antiparasitic [84], and anti-leishmanial agent [85]. |
| 6. | Apigenin | Yellow |
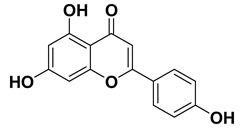
|
Mol wt: 270.24 g/mol Chemical formula: C15H10O5 Pubchem CID: 5280443 IUPAC name: 5,7-dihydroxy-2-(4-hydroxyphenyl) chromen-4-one |
It is an antioxidant [86], anti-inflammatory [87,88,89], anti-bacterial [90], anti-viral [91], anti-cancer [92], chemo-preventive agent, anti-invasive [92,93], and antidiabetic agent [94,95,96] and embodies neuroprotective [97] and vasodilatory action [98]. |
| 7. | Lycopene | Bright Red |
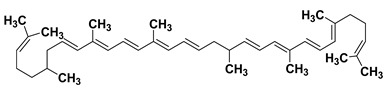
|
Mol wt: 536.873 g/mol Chemical formula: C40H56, Pubchem CID: 446925 IUPAC name: 2,6,10,14,19,23,27,31-octamethyldotriaconta-2,6,8,10,12,14,16,18,20,22,24,26,30-tridecaene |
It is an antioxidant [99], anti-inflammatory [100], anti-microbial [101], anticancer [102], radiation-protective agent [103], and embodies cardioprotective [104] and neuroprotective properties [105]. |
| 8. | Anthocyanin | Red, purple, and blue |
|
Mol wt: 207.24724 g/mol, Chemical formula: C15H11O+, Pubchem CID: 145858 IUPAC name: 2-phenylchromenylium |
It embodies antioxidant [106], anti-inflammatory [107], anti-microbial [108], antiviral [109], and anticancer properties [110]; it embodies hypouricemic and nephroprotective effects [111]. It acts as a cyto-protective [112] and neuroprotective agent [113]. |
| 9. | Capsaicin | Crystalline white |
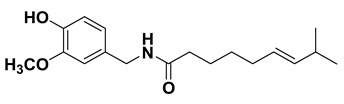
|
Mol wt: 305.4 g/mol, Chemical formula: C18H27NO3 Pubchem CID:1548943 IUPAC name: (E)-N-[(4-hydroxy-3-methoxyphenyl) methyl]-8-methylnon-6-enamide |
It act as an antioxidant [114], anti-inflammatory [115], anti-bacterial [116], anti-fungal [117], anti-viral [118], and anticancer agent [119]. It act as an analgesic [120], gastroprotective [121], anti-obesity [122], and antipruritic agent [123]. It embodies anti-proliferative and pro-apoptotic properties against cancer [124]. |
| 10. | Shogaols | Bright yellow |
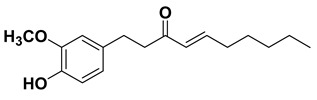
|
Mol wt: 276.376 g·mol−1, Chemical formula: C17H24O3, Pubchem CID: 5281794 IUPAC name: (E)-1-(4-hydroxy-3-methoxyphenyl)dec-4-en-3-one |
It has been found as an antioxidant [125,126,127], anti-inflammatory [128,129,130,131], anti-bacterial [132], anti-fungal [133], anti-viral [134], anticancer, and chemo-preventive agent [135] with anti-emetic and anti-thrombotic properties [136]. |
Figure 1.

Representative figure highlighting some of the therapeutic potentials of curcumin, genistein, and tanshinone IIA. These phytochemicals have been shown to possess intriguing anti-microbial, anti-inflammatory, anti-oxidant, and anti-cancer potentials. The figures are prepared with the BioRender Software (biorender.com).
1.1. Brief Overview on the General Characteristics of Various Phytochemicals
1.1.1. Curcumin
Cur is an active lipophilic polyphenol compound with various beneficial biological activities. The therapeutic properties of Cur have long been reported for centuries [137,138]. Basically, Cur is a bioactive component of Curcuma longa, which belongs to the ginger family. Cur has high potential to scavenge reactive oxygen species (ROS) which makes it an important antioxidant and therapeutic molecule [139,140]. The properties of Cur extend to numerous beneficial functions such as anti-inflammatory, anti-microbial, anti-genotoxic, anti-neoplastic, anti-mutagenic, and anti-tumor activities [141,142,143,144,145]. Moreover, it has been shown to possess phototoxic and photodynamic potential as well [31,32,33]. Although it has low systemic bioavailability issues; nevertheless, interesting data exist to support its intriguing clinical evaluation [146].
1.1.2. Genistein
Gen is an isoflavone mainly present in soy and soy-based food products that are consistently consumed by the Asian population [147,148]. Numerous epidemiological studies have indicated lower incidence of breast and prostate cancers in the Asian countries as compared to other Western countries. These observations have reinforced interest in focusing on the possible contribution of high dietary consumption of isoflavones and lower incidence of cancer [149,150]. It embodies striking structural similarity with the estrogen hormone as a result it is also known as phytoestrogen. Gen embodies a broad range of vital properties, such as antioxidant, anti-inflammatory, and anti-microbial; in addition, it is pro-apoptotic, anti-proliferative, and anti-angiogenic, which validates its chemo-preventive and chemo-therapeutic potential [45,151,152,153,154,155,156]. Interestingly, various clinical studies are on-going to ascertain its pharmacological potential, and there is great optimism that Gen formulation with better bioavailability could seemingly revolutionize Gen-based pharmacological interventions [42].
1.1.3. Tanshinone IIA (Tan IIA)
Tan IIA is a major lipophilic component extracted from Salvia miltiorrhiza Bunge. Accumulating evidence has shown that Tan IIA exhibits multiple biological functions, such as anti-oxidative, anti-inflammatory, and anti-angiogenic effects, as well as anticancer activity against various types of cancers [157,158]. Studies have shown that Tan IIA significantly inhibits the proliferation of several types of tumors, blocks the cell cycle, and induces apoptosis and autophagic death in addition to inhibiting cell migration and invasion. However, poor bioavailability has been a major challenge for pharmaceutical development of Tan IIA, since Tan IIA is challenging to absorb directly in the intestine. Therefore, various analogs and/or formulations have been developed to overcome its bioavailability issues [11].
1.1.4. Allicin
Allicin (diallylthiosulfonate) is obtained from Allium sativum (garlic). This sulfur compound provides a specific taste and smell to the freshly cut/crushed garlic. Allicin has displayed various intriguing pharmacological properties including potential anti-microbial agent. Accumulating evidence has shown that it embodies anti-bacterial activity against various Gram-positive and Gram-negative strains, and methicillin-resistant Staphylococcus aureus [64]. It also exhibits anti-fungal activity when used in in vitro and in vivo systems [159]. Studies have shown that allicin possess intriguing antioxidant potential; and the antioxidant potential of allicin could be plausibly ascribed to its ability to attenuate superoxide, nitric oxide (NO), and hydroxyl radicals [160]. Interestingly, it has been shown that the consumption of garlic in the diet was corelated with reduction of total cholesterol, low density lipoproteins, and triglycerides [161,162]. Moreover, it has been demonstrated to inhibit cholesterol biosynthesis seemingly owing to the inhibition of squalene-monooxygenase 85 and acetyl-CoA synthetase [68,69,163,164]. It has displayed intriguing anticancer potential against different types of cancers [70,71,72]. Moreover, it has been shown to exhibit an antihypertensive effect and neuroprotective potentials as well [66].
1.1.5. Eugenol
It is a phenolic compound (phenylpropanoid) obtained from the leaves and buds of Eugenia caryophyllata (clove). It is chemically 4-allyl-2-methoxyphenol and it imparts the spicy aroma to cloves [165,166]. Several studies have demonstrated different bioactivities of eugenol including anti-bacterial, anti-fungal, anti-viral, antioxidant, and anti-inflammatory properties [167,168,169,170]. Moreover, it has exhibited intriguing potential to combat various types of cancers including gastric, colon, prostate, skin, breast cancer and other cancers [77,78,80,171] through various intricate mechanisms. In addition, it has been shown to embody antidiabetic [83], antiparasitic [84], antileishmanial [85], antipyretic [81], analgesic [172,173] anti-hypercholesterolemic, and antiatherogenic potentials as well [82].
1.1.6. Apigenin
It is trihydroxyflavone found in chamomile, artichokes, celery, sorghum, parsley, oregano etc [174,175]. It embodies antioxidant properties [86] and it has been highlighted that the antioxidant potential of apigenin is plausibly mediated through modulation of antioxidant enzymes (catalase, superoxide dismutase, glutathione peroxidase, and phase II detoxification enzymes) and inhibition of the NF-κB signal transduction pathways [176,177]. Further, it is an intriguing anti-inflammatory agent [178], and the anti-inflammatory effects are seemingly mediated through inhibition of several cytokines including Th2 cytokines, IL-4, IL-10, NLRP3, interleukin 1β genes, etc besides other intricate mechanisms [89,179]. In addition, it also acts antibacterial [90], antiviral [91] and anticancer agent [180]. Similarly, the mechanism of anticancer potential is plausibly mediated through upregulation of STAT1 gene (tumor suppressor), and downregulation of IL-6, TNF-α, and CD40 (tumor causing genes) besides other intricate mechanisms [181]. Moreover, it exhibits antidiabetic activity [94] which has been seemingly attributed to various underlying mechanism including stimulation of insulin secretion, inhibition of gluconeogenesis, and increment in glycogen synthesis [94,95,96].
1.1.7. Lycopene
It is a lipophilic carotenoid hydrocarbon compound found in orange, red, and pink coloured vegetables and fruits including tomatoes, melons, apricots, peaches, grapes, cranberries and papayas [182,183]. Lycopene exhibits various attractive potentials including antioxidant [184], anti-inflammatory [100], anticancer [185,186], cardioprotective [104], and neuroprotective potentials [105]. Studies have shown that lycopene displays intriguing antioxidant activity seemingly through enhancing the level of enzymatic antioxidants (catalase, peroxidase, and superoxide dismutase) as well as non-enzymatic antioxidants (vitamin C and E) [99,184]. Similarly, it displays anti-inflammatory activities plausibly through inhibition of several cytokines and chemokines including NF-κB, IL-6, IL-8, IL-1, TNF-α, nitric oxide (NO) etc. [187,188,189]. Lycopene acts as a cardioprotective agent seemingly through attenuation of oxidation of low density lipoproteins, increment of high density lipoproteinslevels besides other intricate mechanisms [190]. Likewise, the anticancer effect is attributed to induction of apoptosis, cell cycle arrest, and the amelioration of insulin-like growth factor 1 receptor (IGF-1R) signal transduction pathways and so-on [185,186,191].
1.1.8. Anthocyanins
These are group of natural water-soluble phenolic compounds that are broadly distributed in several plant families including Rosaceae, Vitaceae, Cruciferae, Caprifoliaceae, Ericaceae, Saxifragaceae, and Fabaceae [192,193]. Basically, they are subgroup of flavonoids and are found in various plant parts, particularly fruits and flowers [192,194,195]. These have demonstrated several important bioactivities including antioxidant, anti-inflammatory, anticancer, anti-viral, attenuation of neurodegenerative diseases and prevention/treatment of cardiovascular diseases besides other intriguing pharmacological properties [196]. Interestingly, anthocyanins have been reported more potent antioxidants when compared to vitamins C and E [197]. Mechanistically, its antioxidant potential has been associated with its ability to modulate the antioxidant defense system, stimulate glutathione synthesis, and activate antioxidant enzymes (catalase, SOD, glutathione peroxidase). In addition, it has been shown to chelate several metal ions such as iron and copper, and henceforth, reduce free radical production through Fenton and other reactions [198]. Further, the anti-inflammatory effect of anthocyanins is mediated through multiple intricate mechanisms including inhibition of NF-kB, COX-2, TNF-α, IL-1β, and IL-6 [199,200]. Likewise, the anticancer potential of anthocyanins has been attributed to modulation of NF-kB, PI3K/Akt pathway resulting in reduced proliferation of tumor cells and induction of apoptotic responses [110,201].
1.1.9. Capsaicin
It is an alkaloid found in the Capsicum genus. Capsaicin is lipophilic, colorless, odorless, crystalline capsaicinoid having the molecular formula C18H27NO3. Capsaicin exhibited various beneficial pharmacological properties. It has been shown to embody potent antioxidant activity; the antioxidant effect is seemingly attributed to modulation of antioxidant enzymes [202]. In addition, it has been shown to possess intriguing inflammatory potential and the anti-inflammatory potential of capsaicin has been attributed through inhibition of NF-kB, and cytokines levels [203]. Further, it has been highlighted to embody antidiabetic potential, which has been plausibly associated with its ability to induce improvement of glucose metabolism and glucose tolerance [204]. Moreover, capsaicin exhibits intriguing anticancer effect against different types of cancers [205,206]. The mechanism behind the anticancer activity is seemingly through induction of apoptotic responses, inhibition of angiogenesis and so-on [207]. It shows several other interesting pharmacological activities including gastroprotective [121], anti-obesity properties [122], and interestingly it is useful for chronic pain such as that occurring in diabetic neuropathy, rheumatoid arthritis, osteoarthritis, and musculoskeletal pain [208,209] with equal potency.
1.1.10. Shogaols
Shogaols are phenylakylketones found in Zingiber officinale. It has been found as intriguing antioxidant [125] and evidence has shown that shogaols ameliorates oxidative stress seemingly through upregulation of phase II antioxidant enzymes such as heme oxygenase I, glutathione, and thioredoxin I, antioxidant response element promotor functions via the Nrf2 signaling pathway, and so on [126,127,210]. Moreover, it embodies anti-inflammatory, anti-emetic, and anti-thrombotic potential [136] and studies have highlighted that it attenuates inflammatory responses plausibly through multiple intricate pathways involving nuclear factor-kappa B, mitogen-activated protein kinase cascades, activator protein-1, and peroxisome proliferator-activated receptor gamma, etc. [129,130,131]. Furthermore, it exhibits anticancer potentials against various forms of cancers including breast, prostate, bowel, ovary, pancreatic cancer etc mainly through induction of apoptosis and cell cycle arrest [211].
2. Proteomics-Based Interventions in Phytochemical Studies
Proteomic analysis is an unbiased perspective, which is a convenient approach to get a global overview about the effectiveness of bioactive molecules [212,213,214]. Interestingly, proteomic studies could be the intriguing intervention in search of precise phyto-therapeutic agents to combat various diseases. Reckoning with these, in this review, we have majorly concentrated on the proteomic-based studies of Cur, Gen, and Tan IIA bioactive molecules. This review updates the most relevant informations by incorporating an extensive amount of proteomic-based research studies related to these bioactive molecules. A comprehensive table delineating the proteomic studies related to Cur, Gen, and Tan IIA is highlighted in Table 2.
Table 2.
Representative table providing an overview of various proteomics studies to understand the potential role of Curcumin, Genistein, and Tanshinone IIA against various disease pathologies.
| SI. No. | Target Tissue or Cells | Proteomic Strategies |
Objective of the Study | Prospective Proteins Upregulated (↑) or Downregulated (↓) |
Disease and/or Condition Studied |
References |
|---|---|---|---|---|---|---|
| Proteomics studies to understand the molecular intricacies of Cur | ||||||
| 1 | Bacillus subtilis AH75 strain |
2D-DIGE, iTRAQ |
To investigate the proteome alterations in Bacillus subtilis following Cur treatment and identification of its molecular/cellular targets to understand the mechanism of action | UDP-N-acetyl glucosamine 1-carboxy vinyl transferase 2—↓ Putative septation protein (SpoVG)—↑ ATP-dependent zinc metalloprotease (FtsH)—↑ |
Antibacterial action |
[142] |
| 2 | Escherichia coli (ATCC 25922) |
LFQ, LC-MS/MS |
To investigate the mechanistic aspects of the antibacterial effects of Cur in the dark and upon illumination | Chaperone SeqB—↑ Gro-P like protein E (GrpE)—↑ Elongation factor (Tu1)—↑ Universal stress protein F (UspF)—↑ Probable quinol monooxygenase (YgiN)—↑ Uncharacterized oxidoreductase (YajO)—↑ |
Antibacterial action |
[220] |
| 3 | Imipenem- resistant Acinetobacter baumannii |
LFQ, LC-MS/MS |
To investigate the alteration in protein profile following exposure to blue light combined with Cur treatment | Carbonylated Omp38—↑ Carbonylated elongation factor Tu and P—↑ Carbonylated ribosome releasing factor—↑ |
Antimicrobial resistance |
[221] |
| 4 | Fathead minnow epithelial cells (FHM) |
LFQ, LC-MS/MS |
To evaluate the effect of Cur pretreatment in fathead minnow cells infected with viral hemorrhagic septicemia virus (VHSV) | Fibronectin (FN) 1—↓ Heat shock cognate 71 (HSC71)—↓ F-actin—↑ |
Viral Hemorrhagic Septicemia |
[29] |
| 5 | Human liver carcinoma cells (HepG2 cells) |
2D-DIGE, MALDI-TOF/ TOF/MS |
To understand the anticancer mechanism of natural borneol (NB) and Cur in combination |
Heterogeneous nuclear ribonucleoprotein (hnRNPC1/C2)—↓ Nucleophosmin (NPM)—↓ Proteasome 20S Subunit Alpha 5 (PSMA5)—↓ |
Liver cancer |
[222] |
| 6 | Human glioblastoma cells (U87 cells) |
2D-DIGE, iTRAQ, LFQ LC-MS/MS |
To understand the underlying intricacies of LLL12, a Cur derivative against glioblastoma multiforme |
Triose phosphate isomerase (TPI)—↓ Phosphoglycerate mutase 1 (PGAM1)—↓ Adaptor molecule (CRK2)—↓ protein DJ-1 (PARK7) —↓ Basic transcription factor 3 (BTF3)—↓ |
Glioblastoma multiforme |
[223] |
| 7 | Human colorectal carcinoma cells (HCT116 cells) | iTRAQ (TM) | To understand the molecular mechanism of action of Cur against colon cancer and try to identify its exact molecular targets |
Microtubule-associated proteins 1A/1B light chain 3B (LC3B)—↑ Lysosomal-associated membrane protein (Lamp1)—↑ Heat shock protein 70 (HSP70)—↑ |
Colorectal cancer |
[224] |
| 8 | Melanoma cells (LB24 Dagi cells) |
LFQ LC-MS/MS |
To investigate the changes in the protein profile of melanoma cells following treatment with D6 (Cur analog) |
PolyUbiquitin-C—↑ Heat shock 70 kDa protein 1A/1B—↑ DnaJ homolog subfamily B member 1—↑ Heterogeneous nuclear ribonucleoprotein Q—↓ Histone-H2A type 1-C—↓ |
Melanoma cancer |
[225] |
| 9 | Human liver carcinoma cells (HepG2 cells) |
MALDI- TOF/TOF/ MS |
To understand the efficacy of Cur/β-cyclodextrin polymer (CUR/CDP) inclusion complex against HepG2 and its possible molecular mechanisms of action |
Nucleophosmin (NPM1)—↓ Peroxiredoxin-6 (PRDX6)—↓ |
Liver cancer |
[226] |
| 10 | Human acute lymphocytic leukemia cells (MOLT-4 cells) |
2-DE, MALDI-TOF Pro |
To understand the role of Siah- interacting protein (SIP) in Cur-based therapeutic intervention |
Siah-interacting protein (SIP)—↓ | Leukemia | [227] |
| 11 | Human lung adenocarcinoma cells (A549 cells) |
2-DE, MALDI-TOF/ TOF MS |
To understand the precise molecular mechanism of Cur against human lung cancer |
Heat shock protein 90 (HSP-90)—↓ 14-3-3 protein—↓ |
Lung cancer |
[228] |
| 12 | Human colon adenocarcinoma cells (LOVO cells) |
MALDI- TOF/TOF MS |
To investigate the action of irinotecan and Cur against colorectal cancer (LOVO) cells |
Peroxiredoxin-4—↑ Glutathione S-transferase Mu 5—↓ Translocon associated protein subunit delta—↓ Calpain small subunit 1—↓ Protein disulfide-isomerase—↑ (Cur + irinotecan treatment) |
Colorectal cancer |
[229] |
| 13 | Human colorectal cancer cells (SW480 and SW620 cells) |
LFQ LC-MS/MS |
To understand the anti-metastatic properties of the conventional chemotherapeutic drugs and the phytochemicals through comparative proteomic approach |
Fatty acid synthase (FASN)—↓ Histone H4—↓ |
Colorectal cancer |
[230] |
| 14 | Human colon carcinoma cells (HCT-8/VCR cells) |
2-DE, MALDI- TOF/ MS |
To explore the differential proteomic profile of vincristine-resistant HCT-8/VCR cells with and without Cur treatment |
Glutathione S-transferase pi1 gene (GSTP1)—↓ | Colorectal cancer |
[231] |
| 15 | Human colorectal cancer cells (SW480 and SW620 cells) |
2-DE, LC-MS/MS |
To understand anticancer activity of Cur against colorectal cancer |
Mitogen-activated protein kinase (MEK1/2)—↑ Extracellular signal-regulated kinases (ERK1)—↑ Histone deacetylase C1 (HDAC1)—↓ Tumor protein 53 (P53)—↓ AMP-activated protein kinase (AMPKβ1)—↓ |
Colorectal cancer |
[232] |
| 16 | Human gastric cancer cells (BGC-823, MKN-45 and SCG-7901 cells) |
2-DE, MALDI-TOF/ TOF MS |
To investigate the potential of Cur as natural anticancer agent against gastric cancer |
Annexin A1 (ANXA1)—↑ Apoptosis Inducing Factor Mitochondria Associated 1 protein (AIFM1)—↑ Proliferation associated protein (A2G4)—↑ Protein phosphatase PP1-alpha catalytic subunit (PP1A)—↑ Glucose-regulated protein 75 (GRP75)—↓ T-complex protein 1 subunit alpha isoform a (TCPA)—↓ Eukaryotic initiation factor 4A-III (IF4A3)—↓ Thioredoxin domain-containing protein 5 (TXND5)—↓ |
Gastric cancer |
[233,234] |
| 17 | Human breast cancer cells (MCF-7, ZR-75-1) and TGF-β1 pretreated fibroblasts |
LFQ, LC-MS/MS |
To investigate the effects of Cur against breast cancer |
Heme Oxygenase-1 (HMOX1)—↑ Ras Related GTP Binding A (RRAGA)—↑ Ring Finger And CCCH-Type Domains 1 (RC3H1)—↓ (in MCF-7/CLC co-culture) Retrotransposon-derived protein (PEG10)—↓ (in ZR-75-1/CLC co-culture) |
Breast cancer |
[235] |
| 18 | Triple negative breast cancer (TNBC) cells (MDA-MB-231) |
LFQ, LC-MS/MS |
To gain insights into the molecular intricacies of the anticancer effects of combinatorial treatment of Cur and electrical pulses (Cur+EP) compared to solitary treatments |
Aldolase, Fructose-Bisphosphate A (ALDOA)—↓ Enolase 2 (ENO2)—↓ Lactate dehydrogenase A (LDHA)—↓ Lactate dehydrogenase B (LDHB)—↓ Phosphofructokinase platelet (PFKP)—↓ Phosphoglucomutase 1 (PGM1)—↓ (PGAM1)—↓ Phosphoglycerate kinase 1 (PGK1)—↓ |
Triple negative breast cancer |
[236] |
| 19 | Human oral adenosquamous carcinoma cells (CAL 27 cells) |
SILAC, LC-MS/MS |
To investigate the underlying molecular intricacies of tyrosine signaling in response to Cur |
Tyrosine-protein phosphatase non- receptor type 6 (PTPN6)—↑ Abelson tyrosine-protein kinase 2 (ABL2)—↑ Fyn-related Src family tyrosine kinase (FRK)—↓ Pseudopodium enriched atypical kinase (PEAK1)—↓ |
Head and Neck cancer |
[237] |
| 20 | Human Chronic myelogenous leukemia (CML) cells (K562 and LAMA84 cells) |
SWATH MS |
To investigate that how exosome proteins from Cur-treated K562 cells can mediate the anti-angiogenic effect on HUVECs |
Myristoylated Alanine Rich C- Kinase Substrate (MARCKS)—↓ Ras Homolog Family Member B (RhoB)—↓ Vascular cell adhesion protein 1 (VCAM1)—↓ |
Chronic myelogenous leukemia |
[238] |
| 21 | Human chronic myelogenous leukemia (CML) cells (K562 and LAMA84 cells) |
LFQ, LC-MS/MS |
To understand the pharmacological potential of Cur as a safe anti-tumor agent that can function as a chemosensitizer and a multi-targeted inhibitor |
Aldolase, Fructose-Bisphosphate A (ALDOA)—↓ pyruvate kinase muscle isozyme (PKM)—↓ Lactate dehydrogenase A (LDHA)—↓ Phosphoglycerate kinase 1 (PGK1)—↓ Importin-7 (IPO7)—↓ |
Chronic myelogenous leukemia |
[239] |
| 22 | Mouse macrophage cells (RAW264.7 cells) |
2-DE, MALDI-TOF/MS |
To study the anti-atherosclerosis mechanism of action of Cur |
ATP synthesis H+ transporting—↑ MHC class II—↑ Non-muscle myosin alkali light chain—↑ Cytochrome b5—↑ Phosphodiesterase 4D—↓ Eukaryotic initiation factor 3 (elF-3)—↓ Hnrpf protein—↓ Vimentin (VIME)—↓ Nucleophosmin—↓ Ran binding protein (Ranbp 1)—↓ |
Atherosclerosis | [240] |
| 23 | Mouse fibroblast cells (3T3-L1) and Primary white adipocytes |
2-DE, MALDI-TOF/MS |
To understand the proteomic changes in cultured white adipocytes in response to Cur treatment and to identify the target proteins responsible for the fat-browning effects of Cur |
Hormone-sensitive lipase (HSL)—↑ |
Weight management |
[241] |
| 24 | C57BL/6 mice | LFQ, LC-MS/MS |
To understand the therapeutic efficacy of Cur against pulmonary fibrosis | Tumor protein (p53)—↓ Urokinase-type Plasminogen Activator (Upa)—↑ Plasminogen activator inhibitor-1 (PAI-I protein)—↓ |
Pulmonary fibrosis |
[242] |
| 25 | Human neuroblastoma (NB) cells (SH-SY5Y cells) |
LFQ, LC-MS/MS |
To explore the anticancer activity of Cur against human neuroblastoma |
Heat shock protein 70 (Hsp70)—↑ Peroxiredoxin 1 (PRDX1)—↓ Peroxiredoxin 6 (PRDX6)—↓ |
Neuroblastoma | [243] |
| 26 | Human lens epithelial B3 cells (HLE- B3 cells) |
SELDI-TOF/MS | To understand the effect of Cur on HLE-B3 cell proliferation |
Chemotactic factor A17—↓ Chemotactic factor A22—↓ IL-8—↓ Neutrophil active peptide-2—↓ |
Posterior capsular opacification post cataract complications |
[244] |
| 27 | Human colorectal adenoma | LC-MS/MS | To evaluate the effect of Cur on intestinal Uridine diphosphate glucuronosy ltransferase (UGT) expression |
Uridine diphosphate glucuronosyltransferase (UGTs)—not affected by oral Cur | Colorectal adenoma |
[245] |
| 28 | Hepatic liver tissue and Murine hepatocyte cells (AML12 cells) |
2-DE, MALDI–TOF/MS |
To investigate the underlying intricacies of the effect of Cur against non-alcoholic fatty liver disease (NAFLD) |
Superoxide dismutase 1 (SOD1)—↑ Sirtuin 1 (SIRT1)—↑ |
Non-alcoholic fatty liver disease |
[246] |
| 29 | Male hamsters | LFQ, LC-MS/MS |
To elucidate the potential use of Cur and to identify its novel molecular targets |
S100A6—↓ Lumican—↓ Plastin-2—↓ 14-3-3 zeta/delta—↓ Vimentin (VIME)—↓ |
Cholangiocarcinoma | [247] |
| 30 | Sprague Dawley rats |
2-DE, LC-MS/MS |
To investigate whether Cur regulates γ- enolase expression in focal cerebral ischemic injury in rats |
γ-enolase—attenuation of its decreased expression induced by ischemic injury |
Cerebral Ischemia |
[248] |
| 31 | Adult male rats |
2-DE; LC-MS/MS |
To identify various proteins that are differentially expressed by Cur treatment in focal cerebral ischemia |
Ubiquitin carboxy-terminal hydrolase L1 (UCH-L1)—↑ Isocitrate dehydrogenase (ICDH)—↑ Adenosyl homocysteinase (AHC)— ↑ Eukaryotic initiation factor 4A (eIF4A)— ↑ Pyridoxal phosphate phosphatase (PPP)—↓ |
Cerebral Ischemia |
[249] |
| 32 | Mouse models of human inflammtory bowel disease (IBD) |
2-DE, LC-MS/MS |
To understand the role of nutrient–gene interactions in human inflammatory bowel disease (IBD) |
Proteins involved in digestion, excretion, and metabolism—↓ Cellular stress and immune response proteins—↑ |
Inflammatory bowel disease |
[250] |
| Proteomics studies to understand the molecular intricacies of Gen | ||||||
| 33 | Triple negative breast cancer (TNBC) cells (MDA- MB-231 cells) |
TMT | To elucidate anticancer effects of Gen against TNBC cells |
Cyclin-dependent kinase inhibitor (p21WAF1)—↑ Bcl-2-associated X protein (Bax)—↑ B-cell lymphoma-2 (Bcl-2)—↓ Tumor p53 protein (p53)—↓ Cyclin-dependent kinase 1 (CDK1)—↓ |
Breast cancer |
[251] |
| 34 | Human breast cancer cells (T47D cells) |
SILAC | To understand the effect of a varying intracellular ERα/ERβ ratio on Gen-induced genes and protein expression profile |
Myosin (MYH10, MYH14, MYL12B, MYH9, and MYL6)—↑ S100 family Ca2+ binding proteins (S100A8, S100A9)—↓ Prolactin Induced Protein (PIP)—↓ |
Breast cancer |
[252] |
| 35 | Human hepatocellular carcinoma cells (SNU-449 cells) |
2-DE, LC-MS/MS |
To understand the anticancer effects of Gen against SNU-449 cells |
B-cell lymphoma-2 (Bcl-2)—↓ Thioredoxin 1 (Trx1)—↓ |
Hepatocellular carcinoma |
[253] |
| 36 | Human gastric cancer cells (SGC-7901 cells) |
SDS-PAGE, LC- MS/MS |
To understand Gen-induced protein alterations in gastric cancer cells and to investigate the molecular mechanism responsible for the anticancer actions |
Kinesin family proteins (KIFs)—↓ KIF11, KIF20A, KIF22, KIF23—↓ Centromere Protein F (CENPF)—↓ |
Gastric cancer |
[254] |
| 37 | Human promyelocytic leukemia cells (HL-60 cells) |
In-gel digestion, MALDI-TOF/TOF/MS | To elucidate the changes in protein profile following treatment with Gen in human leukemia cells |
Hsp70 protein 8—↑ Heterogeneous nuclear ribonucleoprotein (hnRNP) H1—↑ Ras-related protein (Rab14)—↓ Heterogeneous nuclear ribonucleo protein C (hnRNP C)—↓ Stathmin-1—↓ |
Leukemia | [17] |
| 38 | Human promyelocytic leukemia cells (HL-60 cells) and Human AML cells (MV4-11 cells) |
iTRAQ | To investigate the prospect of using Gen as an effective alternative therapy for AML |
Eukaryotic translation initiation factor 4E-binding protein 1 (4EBP-1)—↓ |
Acute myeloid leukemia |
[255] |
| 39 | Mouse Cardiomyocyte cells (HL-1 cells) |
2-DE, LC-MS/MS |
To study the concentration-dependent effects of Gen treatments on cardiomyocytes |
Heat shock protein 27 (HSP27)—↑ Cathepsin D—↑ Heat shock protein (HSP70)—↑ Glucose-regulated protein (GRP78)—↑ Voltage-dependent anion-selective channel protein 1 (VDAC-2)—↑ Preprotein translocase of the inner membrane of mitochondria (TIM50)—↑ Bcl-2-associated athanogene 2 (BAG2)—↓ N-Myc Downstream Regulated 1 (NDRG1)—↓ |
Cardiac Ischemic stress |
[256] |
| 40 | Human endothelial cells (EA. hy 926 cells) |
2-DE, LC-MS/MS |
Gen, on changes in protein expression levels induced by the endothelial stressor homocysteine (Hcy) in EA.hy 926 endothelial cells | End binding 1 (EB1)—↓ Cathepsin B—↓ |
Atherosclerosis | [257] |
| 41 | Rat Hepatic Stellate cells (HSC-T6 cells) |
2-DE, LC-MS/MS |
To elucidate the antifibrotic mechanism of combinatorial treatment of Gen, Taurine, and epigallocatechin gallate (EGCG) | Hexokinase 2 (HK2)—↓ Lysosomal-associated membrane protein (LAMP1)—↑ Cathepsin D—↑ |
Liver fibrosis |
[258] |
| 42 | Rats | 2-DE, LC-MS/MS and/or MALDI-TOF/TOF |
To understand the role of Gen in breast cancer prevention |
Endoplasmic reticulum resident protein 29 (ERp29)—↑ Guanine deaminase—↑ Fetuin-B—↑ Annexin A1, A2—↓ |
Breast cancer |
[259] |
| 43 | Rats | 2-DE, LC-MS/MS |
To investigate Gen mechanisms of action against chemically induced mammary cancer |
GTP cyclohydrolase 1 (GTP-CH1)—↑ Tyrosine hydroxylase (TH)—↑ Vascular endothelial growth factor receptor 2 (VEGFR2)—↓ |
Breast cancer |
[260] |
| 44 | Prepubertal girls blood and urine |
TMT | To identify protein biomarkers of the effect and susceptibility for cancer from the blood of girls exposed to select environmental chemicals |
Endothelin-converting enzyme (ECE-1)—↓ Eukaryotic translation initiation factor 3 subunit J (EIF-3)—↓ Nucleolar 7—↑ PR domain zinc finger 5 (PRDM5)—↑ |
Mammary cancer |
[261] |
| 45 | Sprague Dawley rats |
TMT | To identify protein biomarkers of susceptibility from blood sera of rats exposed prepubertally to Bisphenol A (BPA) or Gen |
Neurosecretory protein VGF 8a (VGF)—↓ Rho-associated coiled-coil containing protein kinase 2 (ROCK2)—↓ Matrix metalloproteinase 3 (MMP3)—↓ Protein tyrosine phosphatase receptor type K (PTPRK)—↑ SET domain containing 2 (SETD2)—↑ Ubiquitin carboxyl-terminal hydrolase—↑ |
Cancer | [262] |
| 46 | C57BL/6J female mice (INTACT) and castrated females (CAST) |
2-DE, LC-MS/MS |
To assess the impact of Gen on the cardiac proteome in ovariectomized female mice | Myosin 6—↑ Myosin regulatory light chain 4 (MLC-4)—↑ Moesin—↑ |
Cardiovascular disease |
[263] |
| 47 | Mice | 2-DE, LC-MS/MS |
To characterize an animal model for alternative hormone replacement with Gen as a natural estrogenic compound |
LIM domain-binding protein—↑ Desmin—↑ |
Animal model characterization |
[264] |
| 48 | Sprague Dawley rat model of liver fibrosis |
iTRAQ | To understand the anti-fibrotic mechanisms of combination therapy of Gen, taurine, and epigallocatechin gallate (EGCG) |
Annexin A2 (Anxa2)—↑ Thioredoxin domain-containing protein (Txn1)—↑ Proteoglycan 4 (Prg4)—↑ polymeric immunoglobulin receptor (Pigr)—↑ fibulin-1 (Fbln1)—↑ Triosephosphate isomerase (Tpi1)—↑ |
Liver fibrosis | [265] |
| 49 | Rat model of liver fibrosis |
iTRAQ | To understand the possible therapeutic mechanism of combination therapy against liver fibrosis |
Thioredoxin domain-containing protein (Txn1)—↑ DEAD box protein family (Ddx39a)—↑ 17-beta-hydroxysteroid dehydrogenase type 6 (Hsd17b6)—↑ Cysteine Conjugate-beta Lyase 2 (Ccbl2)—↑ Magnesium cation transporter protein (Magt1)—↑ Cytochrome P450 4A14 (Cyp4a14)—↑ Glutathione S-transferase A1 (Gsta1)—↑ |
Liver fibrosis |
[266] |
| 50 | Adults male Wistar rats |
SDS-PAGE, LC- MS/MS |
Morphological response of reactive astrocytes positive for glial fibrillary acidic protein (GFAP) in rats |
Glial fibrillary acidic protein (GFAP)—↓ |
Astrogliosis | [267] |
| Proteomics studies to understand the molecular intricacies of Tan IIA | ||||||
| 51 | Human cervical cancer cells (HeLa cells) |
In-gel digestion, MALDI-TOF MS |
To investigate the prospective potential of Tan IIA as a potential anti-tumor agent |
Proliferating cell nuclear antigen (PCNA)—↓ Heat shock protein 27 (HSP27)—↓ Vimentin (VIME)—↓ β-tubulin—↑ Superoxide dismutase (MgSOD)—↑ Glucose-regulated protein 75 (GRP75)—↑ Prohibitin—↑ |
Cervical cancer |
[268] |
| 52 | Human hepatocellular carcinoma cells (MHCC97-H cells) and Chang liver cells |
LFQ, LC-MS/MS |
To elucidate the Tan IIA-induced protein profile alteration in MHCC97-H cells |
Keratin, type II cytoskeletal 8—↓ Keratin, type I cytoskeletal 18, 19, and 20—↓ Cathespin D—↓ Profilin 1—↓ Nucleoside diphosphate kinase A—↑ Annexin A1, A2—↑ |
Liver cancer |
[269] |
| 53 | Human gastric cancer cells (AGS cells) |
iTRAQ | To understand the mechanism of action of TanIIA against gastric cancer |
Tumor p53 protein (P53)—↑ Serine/threonine-protein kinase (AKT)—↓ |
Gastric cancer |
[270] |
| 54 | Immortalized rat myofibroblast cells (HSC-T6 cells) and Human Hepatocellular carcinoma cells (HepG2 cells) |
In-gel digestion, MALDI-TOF/ TOF/ MS |
To explore the mechanism of apoptosis induced by Tan IIA on activated rat Hepatic Stellate Cells (HSCs) |
Prohibitin—↑ Translational Controlled Tumor Protein–(TCTP)—↓ GDP-dissociation inhibitor 1 (GDIR1)—↓ 14-3-3ε—↓ |
Liver fibrosis |
[271] |
| 55 | Osteosarcoma cancer cells (MG-63 cells) |
2-DE, LC-MS/MS |
To understand the molecular mechanisms of anticancer effects of Ginsenoside Rg1, Cinnamic acid, and Tan IIA and to know their targets | Prohibitin—↓ | Osteosarcoma | [272] |
| 56 | Human gastric cancer cells (AGS cells) |
Phosphoproteomics LTQ LC-MS/MS |
To understand the molecular signal transduction pathway associated with the anticancer potential of Tan IIA |
Phosphorylation of heat shock protein 27 Heat shock factor 1 (HSF1)—↑ |
Gastric cancer |
[273] |
| 57 | Human papillomavirus type 16 (HPV-16)-positive cells (CaSki cells) and Human cervical cancer cells (HeLa and SiHa cells) |
In-gel digestion, MALDI-TOF/ TOF MS |
To evaluate the growth inhibitory effect of Tan IIA on CaSki cells |
Protein disulfide-isomerase A1 (PDIA1)—↓ Glucose-regulated protein 78 (Grp78)—↓ Whey proteins (TERA)—↓ Glucose-regulated protein (Grp94)—↓ Vimentin (VIME)—↓ Glucosidase II Alpha Subunit (GANAB)—↑ Cytoskeletal protein (VINC)—↑ The Putative Coupling Protein (TCPA)—↑ The brucella effector protein B (TCPB)—↑ Keratin 2C7 (K2C7)—↑ |
Cervical cancer |
[274] |
| 58 | Bone marrow- derived mesenchymal cells (BM-MSC) and Wharton’s Jelly- derived mesenchymal cells (WJ- MSC) |
LFQ, LC-MS/MS |
To understand the ability of BM- and WJ-MSC to differentiate towards the osteogenic lineage |
Proteins of BMP signaling—↑ | Orthopedic disease |
[275] |
| 59 | Mice myocardial cells |
LFQ, MALDI- TOF/TOF/MS |
To investigate the effect of Tan IIA on transverse aortic constriction (TAC)-induced heart failure |
NADPH Oxidase 4 (Nox4)—↓ P38—↓ Nuclear factor erythroid 2- Related factor 2 (Nrf2)—↑ |
Myocardial apoptosis |
[276] |
| 60 | Lung cancer Radioresistant cells (H358-IR and H157-IR cells) |
SILAC MS |
To evaluate the potential of Tan I as a potential radiation sensitizer in lung cancer |
Phosphoribosyl Pyrophosphate Amidotransferase (PPAT)—↓ B-cell lymphoma-2 (BCL2)—↓ Caspase 8—↑ |
Lung cancer |
[277] |
| 61 | Adult male KM mice |
2-DE, LC-MS/MS |
To understand the potentials of Tan II A sodium sulfonate (TSNIIA-SS) against Doxorubicin (DXR)-induced nephropathy |
Myo-inositol oxygenase—↑ Glutathione peroxidases (GSH-Pxs)—↓ Proteasome alpha 5—↓ |
Nephropathy | [278] |
Abbreviations: LFQ: Label free quantification; MALDI-TOF/TOF/MS: Matrix Assisted Laser Desorption/Ionization-Time of Flight/Time of Flight/Mass Spectrometry; 2-DE: Two-dimensional Electrophoresis; iTRAQ: Isobaric tag for relative and absolute quantitation; LC-MS/MS: Liquid Chromatography-Mass Spectrometry/Mass Spectrometry; 2D-DIGE: Two-dimensional-Difference In Gel Electrophoresis; SILAC: Stable Isotope Labeling by/with Amino acids in Cell culture; SELDI-TOF-MS: Time of Flight Mass Spectrometry; SDS-PAGE: Sodium dodecyl-sulfate polyacrylamide gel electrophoresis; TMT: Tandem Mass Tag.
Over the last two decades, various mass spectrometry (MS)-based high throughput proteomic approaches have been widely utilized for a plethora of applications [215,216]. As a matter of fact, the proteomic analysis deals with the profiling, identification, and quantification of proteins as well as peptides in different biological samples [217]. Conventionally, it comprises of various gel-based techniques such as two-dimensional gel electrophoresis (2-DE), two-dimensional differential gel electrophoresis (2D-DIGE), and gel free techniques. For example, these include stable isotope labeling by amino acids in cell culture (SILAC), isobaric tag for relative and absolute quantitation (iTRAQ), tandem mass tags (TMT), label free quantitation (LFQ) analysis, and multiple reaction monitoring (MRM). For a comprehensive review on proteomics-based technology, please refer to articles by Beck et al., [218] and Domon et al., [219]. Here, we have described the proteomics-based studies of Cur, Gen, and Tan IIA in detail. An overview of the proteomics strategies exploited to study the underlying molecular intricacies for phytochemical-based therapeutic interventions has been depicted in Figure 2.
Figure 2.

An overview of the proteomics strategies exploited to study the underlying intricacies for phytochemical-based therapeutic interventions. The figures are prepared with the BioRender Software (biorender.com). Abbreviations: SWATH: Sequential Windowed Acquisition of All Theoretical Fragment; DDA: Data-Independent Acquisition; DIA: Data-Dependent Acquisition; MRM: Multiple-Reaction Monitoring; SRM: Selective Reaction Monitoring; iTRAQ: Isobaric tag for Relative and Absolute Quantitation; SILAC: Stable Isotope Labeling by/with Amino acids in Cell culture; TMT: Tandem Mass Tag; 2-DE: Two-dimensional Electrophoresis; 2D-DIGE: Two-dimensional-Difference in Gel Electrophoresis; MS: Mass spectrometry.
2.1. Proteomic Approaches to Understand the Function of Curcumin as Therapeutic Intervention
2.1.1. Proteomic Studies to Explore Its Potential against Microbial Diseases
Accumulating evidence has highlighted that Cur has a broad spectrum of anti-microbial effects including antibacterial, antifungal, and antiviral action [29,142,220,221]; it has been envisaged to exhibit synergistic effects in combinatorial therapeutic regimes. In this regard, to explore the antibacterial activities of Cur against Bacillus subtilis, Reddy et al. treated B. subtilis AH75 strain with Cur (20 μM) at different time intervals, and performed a comprehensive proteomic analysis using 2D-DIGE and iTRAQ to analyze the differential expression profile. Interestingly, differential proteomics profiling revealed alterations in various proteins including putative septation protein SpoVG, UDP-N-acetylglucosamine 1-carboxy vinyl transferase 1, and the ATP-dependent Clp protease proteolytic subunit. Moreover, alteration in the universal chaperone system (GroEL), required for the tubulin homologue protein filamenting temperature-sensitive mutant Z (FtsZ) folding, as well as the major protease (Clp family) system that targets FtsZ for degradation was also observed. Further, bioinformatics analysis revealed that Cur treatment considerably altered various cellular processes including central metabolism, fatty acid metabolism, and cell wall synthesis pathways, all of which have an important role for bacterial viability. Collectively, the study provided a plausible understanding of the mechanism of action and the putative targets of Cur, suggesting that treatment of Cur majorly affects cell division, cell wall synthesis, chaperones, and central metabolism in B. subtilis AH75 strain [142].
Reckoning with its antiviral action, it has been envisaged that although Cur has been widely studied in the background of the antiviral mechanism, the available literature does not clearly explain the effect of Cur in the early stages of viral infection. However, the underlying intricacies employed for interactions among viruses, cells, and antiviral compounds are incredibly diversified. Thus, it is important to comprehensively analyze the diverse protein–protein interactions in host cells during the viral entry phase. To this end, Jeong and colleagues investigated the underlying intricacies; for this, they pre-treated the head minnow cells with Cur (15–240 μM) followed by a viral hemorrhagic septicemia virus (VHSV) infection [29]. Thereafter, they performed a comparative proteomic study on the animal models with VHSV-infected, and Cur-treated VHSV-infected animals. Proteomics analysis revealed alterations in protein expression of several proteins including heat shock cognate 71 (HSC71), elongation factor 1 (EEF1), alpha cardiac muscle (ACTC1) protein and actin protein. Further pathway analysis through ingenuity pathways analysis (IPA) provided clues that HSC71 could be the primary candidate interacting with actin proteins (ACTB, ACTG, and F-actin), fibronectin (FN)-1, and gelsolin (GSN) in both VHSV-infected and Cur-treated VHSV-infected organisms. All these data provided evidence that Cur downregulates the expression of HSC71, which consequently increased virally infected cell viability, and inhibited the VHSV replication. Furthermore, Cur induced an alteration in the ratio of F-actin/G-actin, this represents another interesting connecting link that indicates the plausible mechanism to inhibit viral entry [29].
Collectively, all these studies provided a plausible understanding of the mechanism of action and the putative targets of Cur for their antimicrobial potential.
2.1.2. Proteomic Studies to Explore Its Potential against Cancer
Several promising studies have shown the protective potential of Cur against many disease conditions; nevertheless, their anticancer potential is the most researched topic. Cur has been shown to inhibit cancer cell growth, invasion, and the metastasis properties of various types of cancer [233,279]. Further exploration of the underlying intricacies highlighted that the target proteins of Cur were found to be involved in many different processes including cell proliferation, apoptotic responses, nucleic acid processing, protein folding, protein translational machinery, proteolysis process, cytoskeleton organization, and signal transduction pathways [233].
Cur is well known for its therapeutic activities; nevertheless, it has poor systemic bioavailability. Therefore, various chemical analogs and/or Cur formulations have been developed with the aim to improve their bioavailability issues and enhance its efficacy thereof [280]. To this end, in order to increase the absorption of Cur, Natural borneol (NB), the bicyclic organic compound, has been formulated and tested against liver cancer. Interestingly, the treatment of Cur/NB in HepG2 cells resulted in differentially expressed proteins (17 proteins upregulated and 12 downregulated) that were functionally associated with the cell cycle and apoptosis, as well as the p53 pathway (hnRNPC1/C2, NPM, and PSMA5). To this end, decreased levels of hnRNPC1/C2 and NPM eventually leads to phosphorylation of the p53 protein; the activated p53, along with differentially expressed PSMA5, consequently increased the level of p21. Moreover, NB/Cur also enhances ROS synthesis, which is involved in the G2/M cell arrest mechanism. Collectively, it is reasonable that this proteomic study provided a strong evidence and better understanding regarding the anticancer property of Cur [222].
Further, Cur derivatized inhibitory compound, LLL12, demonstrated intriguing potential against glioblastoma multiforme (GBM) [223]. LLL12 is a known inhibitor of signal transducer and activator of transcription 3 (STAT3), which is constitutively active in various types of cancers. Basically, the global effects of targeting STAT3 using LLL12 were identified using 2D-DIGE and iTRAQ, suggesting intriguing anti-tumorigenic activity of LLL12. Interestingly, LLL12 treatment exhibited downregulation of phosphoglycerate mutase 1 (PGAM1), triosephosphate isomerase (TPI), adaptor molecule cysteine-rich receptor-like protein kinase 2 (CRK2), basic transcription factor 3 (BTF3), and protein DJ-1 (PARK7), which suggested that these targets may serve as prognostic or predictive markers in GBM [223]. Concomitantly, this study revealed Cur involvement in various cellular responses such as apoptosis induction, cellular metabolism, and anti-angiogenic activities.
Further, to better understand the molecular target of Cur, an intriguing investigation was performed by Wang and group in 2015. In their study, they utilized a cell-permeable Cur probe (Cur-P) coupled with an alkyne moiety that can be tagged with biotin for further enrichment. The researchers performed a quantitative proteomics approach to identify specific binding targets. This study revealed 197 proteins that were seemingly identified as Cur-binding targets. Further investigation divulged the target distribution and enrichment in different organelles such as mitochondria, nucleus, and plasma membrane. IPA divulged the anticancer effects of Cur, which suggest the involvement of Cur in a myriad of biological functions including mTOR signaling, mitochondrial dysfunction pathways, as well as regulation of eIF4/p70S6K and EIF2 proteins. Later, functional validation established that Cur induces autophagy, suppresses cellular protein synthesis, and increases ROS production and lysosomal activation, which leads to cell death of cancerous cells and confirms its anticancer potential [224].
Another investigation highlighting the antitumor activity of Cur D6 (hydroxylated biphenyl compound) on primary melanoma LB24Dagi cells, employed proteomics and mass spectrometry analysis. The altered proteins exhibited strong activation of a cellular stress response, with upregulation of several HSPs and triggered ubiquitin-proteasome pathways. The researchers concluded that Cur seemingly altered the majority of cellular functions and finally drives the cells to apoptotic pathways, without affecting normal healthy cells [225].
Another interesting study utilizing a gel-based proteomic approach showed decreased spot intensity (up to 70–90%) of SIP (Siah-interacting protein) in the Cur-treated (Cur sensitive) compared to the Cur-resistant human acute lymphocytic leukemia (MOLT-4) cells that caught the attention of the researchers. The study indicated that seemingly SIP is an important player in Cur-induced apoptosis in Cur sensitive cells and plays a critical role in Cur resistance [227].
Further, treatment of hypotriploid human epithelial lung carcinoma cells, viz., A549 cells with the Cur analog T63 (4-arylidene), revealed ~66 proteins with altered expression patterns. It seems that T63 contains a diverse range of molecular targets including HSP90 and 14-3-3 proteins as revealed by 2-DE and Ultraflex II MALDI-TOF/TOF MS analysis. Overall, the study proposed that T63-triggered cell cycle arrest and apoptotic responses involving mitochondrial dysfunction and ROS generation; and inhibition of the proteasomal machinery [228].
Further, besides being employed in solitary treatment regimes, Cur has also been employed in combinatorial treatment regimes. To this end, a report investigated the combined action of Cur and irinotecan on colorectal cancer (CRC) cells (LOVO cell) using in-gel protein digestion and MALDI-TOF/TOF MS [229]. Interestingly, it was found that out of a total of 54 protein spots differentially expressed, four exhibited protein–protein interactions. The cocktail could seemingly enhance the expression of protein disulphide isomerase (PDI) and peroxiredoxin-4 (PRDX4) which disarranged the formation and reduction of disulphides, which consequently leads to enhanced apoptotic responses in LOVO cells. The authors speculated that Cur may lead to the suppression of glutathione S-transferase Mu 5 (GSTM5) expression that helps in enhancing the lethal effect of irinotecan. Another study concluded that Cur enhanced the effect of irinotecan against CRC cells through ROS generation and activation of the Endoplasmic Reticulum (ER) stress pathway. Proteomic analysis through MALDI-TOF/TOF MS revealed 11 repeated protein nodes, which are involved in intracellular calcium pathways, intracellular redox reaction pathways, and intracellular endoplasmic reticulum (ER) stress [281]. Furthermore, anti-metastasis activity of Cur, ginsenoside 20 (S)-Rg3, and oxaliplatin were comparatively evaluated using proteomic analysis in isogenic primary (SW480) and metastatic colon (SW620) cell lines. This combinational therapy demonstrated the suppressive effect of all three bioactive substances on fatty acid synthase and histone H4 expression. There was a significant reduction in migratory activity of SW620 cells, which suggests that theyteffectively retards cell migration in colon cancer [230].
Cumulatively, it is reasonable to envisage that all these proteomic studies provides a better understanding of the underlying intricacies for the anticancer potential of Cur.
2.1.3. Proteomic Studies to Explore Its Potential against Various Other Disease Pathologies
Accumulating evidence has highlighted the intriguing role of Cur against atherosclerosis [240,282]. In order to understand the molecular intricacies, the proteomic analysis ofmonocyte/macrophage-like cells (RAW264.7) cultured in the presence of Cur revealed considerable alteration in the proteome profile. This included increased expression of cytochrome b5 (cb5), ATP synthase, non-muscle myosin alkali light chain, and MHC class II protein moieties in RAW264.7 cells. On the other hand, decreased expression for various key players such as ran binding protein (RanBP)-1, phosphodiesterase 4D, eukaryotic initiation factor 3 (elF-3), nucleophosmin, vimentin, and heterogeneous nuclear ribonucleoprotein F (Hnrpf) protein were found as well. These data indicated the involvement of Cur in a myriad of functions including modulation of cell inflammation, reduction in the accumulation of intracellular cholesterol, antioxidant activity, and inhibition of cholesterol transport in RAW264.7 cells. Collectively, this study conclusively supports the anti-atherosclerosis mechanism of Cur seemingly through regulation of the accumulation of intracellular cholesterol levels and its transport [240].
Furthermore, Cur effectiveness and beneficial properties have also been investigated in weight management employing proteomic approaches. As a matter of fact, browning of white adipose tissue is an intriguing approach to combat obesity by enhancing energy expenditure. To demonstrate the protein involved in the fat-browning effect, proteomic modifications were analyzed in cultured white adipocytes under Cur treatment. Analysis through 2-DE combined with MALDI-TOF-MS revealed differential expression of ~58 protein spots among the control and Cur-treated adipocytes; out of which, hormone-sensitive lipase (HSL), an interacting partner of another two browning markers, uncoupling protein 1 (UCP1) and Peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α) were found to be prominently associated with the browning phenotype. Overall, this study suggests that Cur induces the HSL level in white adipocytes, which in turn induces fat browning [241].
Further, as a matter of fact, pulmonary fibrosis is an impaired fibrinolytic system that is associated with inflammation of the alveoli, which thereby leads to deposition of extracellular matrix (ECM) components and myofibroblasts. In order to investigate the therapeutic potential of Cur against pulmonary fibrosis disease progression, Gouda et al. employed high throughput Q-Orbitrap MS technology. In their study, basically C57BL/6 mice were injected with Bleomycin (BLM), followed by Cur treatment for 24 and 48 h time intervals [242]. The results of proteomic analysis revealed fascinating outcomes; it was found that BLM-exposed mice showed gradual weight loss and altered lung morphology. On the other hand, these symptoms were considerably reversed following Cur treatment. The proteomic analysis suggested strong interaction of Cur with p53, PAI-I, and uPA proteins as an expression of IL-17A-mediated inflammation in the impairment of the p53-fibrinolytic system and alveolar epithelial cell (AEC) apoptosis, which is a critical pathophysiological hallmark of pulmonary fibrosis. The results suggested that Cur could act as a potential therapeutic candidate to target the fibrinolytic system during pulmonary fibrosis, alongside its protective role against the progression of pulmonary fibrosis [242].
Further, accumulating evidence has highlighted the intriguing role of Cur as an effective therapeutic agent against various neurodegenerative diseases [243,283,284,285]. To this end, Urbani and group highlighted an intriguing molecular investigation of the main proteome rearrangements involved in the cellular response to Cur in human neuroblastoma cells sensitive to cisplatin and its resistant counterpart through shotgun proteomics analysis. Interestingly, the comparative proteomics analysis revealed that 66 proteins were differentially expressed following Cur treatment in sensitive cells. On the other hand, 32 proteins were differentially expressed in resistant treated cells. Further, gene ontology studies revealed that proteins involved in cellular assembly and organization, biosynthesis, and glycolysis were downregulated following Cur treatment. Moreover, proteome changes were also associated with cell cycle arrest in the G2/M phase and accumulation of polyubiquitinated proteins. As a matter of fact, the polyubiquitination of proteins influences a wide range of cellular processes; thus, the inhibition of the ubiquitin–proteasome system might be the major way through which Cur performs its multifactorial effects [243].
Further, studies have shown that Cur protects against Alzheimer’s disease plausibly via binding to sensile plaques and thereby inhibiting plaque pathology, Aβ plaque aggregation, and reduction in amyloid levels [38,39,41]. Moreover, Cur has been demonstrated to attenuate parkinsonism as well, seemingly through modulation of human α7-nicotinic acetylcholine receptor (α7-nAChRs) [40,286].
Further, Cur has been found to be an intriguing agent to be used following cataract surgery. Basically, a cataract is an opacification (cloudy appearance or opaqueness) of the eye lens that causes a decrease in vision. After cataract operation (i.e., lens replacement), some patients develop the symptom of faded vision. The condition occurs due to posterior capsular opacification (PCO) after cataracts removal. Unfortunately, it is a prevalent side effect following lens replacement. Therefore, attenuation of proliferation of lens epithelial cells (LECs) could plausibly prevent and/or repress PCO. To this end, Hu et al., used MS to investigate the inhibitory action of Cur against the proliferation of human lens epithelial B3 cells (HLE-B3). The proliferation of HLE-B3 cells was induced through administration of recombinant human basic fibroblast growth factor (rhbFGF), followed by treatment with Cur (20 mg/L). Interestingly, the results of this study showed that Cur acts as an effective inhibitor of the HLE-B3 cell proliferation induced by rhbFGF [244]; this plausibly endorses its effectiveness in management of post-cataract complications.
As already mentioned, alternative and/or complementary medicine have shown intriguing potential against various disease pathologies including cancer. Albeit often perceived as innocuous, these phytochemicals can seemingly interact with various metabolic enzymes including cytochrome P450s (CYPs), UDP glucuronosyl-transferases (UGTs), and drug transporters (e.g., P-gp, MRP, OATP). These intricacies have highlighted the need to understand potential phytochemical–molecular interactions To this end, a critical study evaluated the effect of oral Cur on intestinal uridine diphosphate glucuronosyltransferase (UGTs) expression in healthy volunteers aged between 40–80 years through LC-MS/MS [245]. In this study, all volunteers consumed daily curcuminoid extract (4 g) for 30 days. Interestingly but not surprisingly, proteomic data analysis did not reveal any significant differences in rectal mucosal UGT concentrations before and after Cur administration. Concomitantly, this study indicates that daily Cur use is unlikely to alter colonic UGT expression, especially in colon cancer [245]. Nevertheless, whether this is true with other forms of cancer as well seemingly requires further investigations to truly understand the Cur–metabolic enzyme interactions.
2.2. Proteomic Approaches to Understand the Function of Genistein as Therapeutic Intervention
Gen embodies broad range of vital properties, including antioxidant, anti-inflammatory, anti-microbial, anti-cancer, and so on [45,151,152,153]. As a matter of fact, a type of breast cancer classified as triple-negative breast cancer (TNBC) is estrogen receptor-negative, progesterone receptor-negative, and Her2-negative. Overall the survival, whether in early-stage or advanced disease stage, is poor in TNBC patients. Unfortunately, there are shortage of targeted therapies for TNBC. Of note, Gen is known for its estrogenic potential and accumulating data has highlighted its anticancer potential in breast cancer. To explore more about the anticancer potential of Gen against TNBC; Fang et al. performed phosphoproteomics studies. Interestingly, they identified approximately 5445 phosphorylation sites on 2008 phosphoproteins following Gen treatment. Further, bioinformatics analysis indicated the presence of 332 Gen-regulated phosphorylation sites on 226 proteins. Thus, proteomic data revealed that Gen may be involved in the critical cell cycle processes, including DNA replication, cohesin complex cleavage, and kinetochore formation. Additionally, Gen potentiates the activation of DNA damage responses, such as activation of ataxia telangiectasia serine/threonine-protein kinase (ATR) and breast cancer susceptibility gene 1 (BRCA1) complex. Conclusively, this phosphoproteomics study revealed the complex role of Gen in the regulation of the cell cycle and DNA damage response [251].
Another study explored concentration-dependent anticancer activity of Gen against SNU-449 cells [253]. The study reported its apoptosis-associated signature characteristics including involvement in caspase-3 activation as well as DNA fragmentation. Proteomics analysis revealed the involvement of antioxidant protein, thioredoxin-1, in Gen-induced apoptosis. Of note, thioredoxin-1 levels were found to be downregulated following Gen treatment, resulting in increased accumulation of ROS intracellularly. Moreover, Gen potentiated activation of different signalling mediator proteins including c-Jun N-terminal kinases (JNK), apoptosis signal-regulating kinase 1, and p38. Interestingly, prior treatment of JNK and p38 inhibitors can considerably abolish Gen-induced apoptotic responses. Concomitantly, the study concluded that Gen induces apoptotic responses in SNU-449 cells were plausibly through reduction in the thioredoxin-1 concentration and activation of JNK, apoptosis signal-regulating kinase 1, and p38 kinase thereof [253].
Another study explored the proteomics alterations in rat mammary glands following Gen treatment. Basically, the female rats were exposed to Gen by different routes through lactating dams and thereafter, the mammary glands were collected at day 21 and 50 post treatment and subjected to proteomic studies. The study revealed alteration in expression of ~23 proteins. Wherein, proteins such as Annexin A2, Gelsolin, Phosphoglycerate kinase-1 (P1), protein disulfide isomerase A3 (PDIA3), vascular endothelial growth factor receptor 2 (VEGF-R2), and epidermal growth factor receptor (EGF-R) were further validated through immunoblot assay. Of note, differential expressions of these proteins at different time points were found; for instance, expression of annexin A2 were found to be increased at the 21st day and reduced at the 50th day. On the other hand, PGK1 levels remained unchanged at the 21st day but decreased around the 50th day. Similarly, fetuin B expression was unaltered until the 21st day but increased on the 50th day, whereas the expression of VEGF-R2 and EGF-R were decreased at the 50th day in the mammary gland [259].
One more study by Wang et al. focused on the protein biomarkers for both effectiveness and susceptibility to breast cancer in blood and urine of prepubertal girls exposed to selected environmental chemicals with high urine concentrations of Gen, BPA, mono-ethyl hexyl phthalate (MEHP), and mono-benzyl phthalate (MBzP). Proteomics data suggested that the differentially regulated cancer-related proteins in girls with high concentrations of BPA and Gen corelated well with previously reported functions of BPA in carcinogenesis and of Gen in mammary cancer prevention, respectively [261].
Further, another proteomics study was performed to identify the prospective protein biomarkers and their association with carcinogenesis upon exposure to bisphenol A (BPA, a cancer-causing agent) and/or Gen to prepubertal rats. The results of the study suggested that Gen pre-exposed rats showed decreased expression of matrix metalloproteinase-3 (MMP3), rho associated coiled-coil containing protein kinase 2 (ROCK2), VGF nerve growth factor inducible, and Alpha-1 antitrypsin (SERPINA1), whose overexpression has previously been associated with carcinogenesis in various types of human cancers. Similarly, three tumor suppressor proteins (UCH1, SETD2, and PTPRK) were found to be upregulated in Gen-exposed rats. Thus, decreased expression of carcinogenesis inducing protein and increased expression of a tumor suppressor protein seems to be responsible for the chemoprotective action of Gen in animal model [262].
Gen treatment has also been employed against various other human disease pathologies such as cardiovascular diseases, liver fibrosis, and so on. Due to its potential role in the cardiovascular system, Gen has been reported to be associated with a lower blood pressure condition either directly or indirectly. It has been shown that Gen considerably attenuates vascular contraction; thereby regulating vascular tone and blood pressure plausibly via regulation of myosin light chain (MLC) phosphorylation, mediated through myosin light chain kinase (MLCK) or the RhoA signaling cascade. Further, it has been highlighted that oral consumption of Gen altered the level of the various cytoskeletal and contractile proteins in ovariectomized female mice, and also increased the phosphorylation of MLC. This study, contrary to others, suggests that Gen does not inhibit the MLCK or RhoA pathway [263]. Thus, it necessitates further investigation to fill in these gaps and plausibly resolve these discrepancies.
It is widely acknowledged that after menopause, estrogen elevates the risk of cardiovascular disease in women. Therefore, a novel strategy is to replace estrogens with alternative hormones like phytochemical Gen that acts as a natural estrogenic compound and reduces the effect. To this end, a 2-DE/ESI-LC-MS approach was used to investigate the effects of a dietary supplement with the phytoestrogen Gen on the cardiac proteome pattern for young, adult, and castrated male and female mice. Basically, the protein species diversification and their alteration were studied following Gen intake. The authors noticed substantial effect on the relative abundance of estrogen receptors, even through oral consumption. This investigation revealed expression of several fatty acid metabolism associated enzymes, and interestingly, their transcriptional regulators varied in male and female mice at both the transcriptional as well as at the protein level. Moreover, they also noticed that Gen increased the protein levels in male mice, which was found to be closely associated with oxidative phosphorylation and generation of ROS. On the other hand, in female mice, Gen elevates the level of two isoforms of LIM (LIN-11, Isl-1, and MEC-3) domain-binding protein and one isoform of desmin, which is associated with cardiac hypertrophy [264]. Taken together, this research endeavor revealed a complex influence of Gen on the proteome of the murine heart and warrants further investigations for a better understanding of the influence of Gen in myocardial pathology. In analogy, another study further investigated the effects of fixed concentrations of Gen on HL-1 cardiomyocyte cells. They noticed that various proteins were differentially expressed upon treatment with 1 μM and 50 μM Gen, and that both concentrations of Gen impacted the regulation of ATPase activity and glucose catabolic processes. Nevertheless, at lower concentrations, Gen significantly influences the heat shock proteins and anti-apoptotic responses. Furthermore, a higher concentration reduces glycolytic proteins and antioxidant enzymes, which consequently leads to energy depletion and apoptotic responses, making the cardiomyocytes potentially more susceptible [256].
It is widely envisaged that liver fibrotic conditions ensue when healthy tissue of the liver becomes scarred, which can turn into chronic liver diseases at later stages. Researchers have evaluated the anti-fibrotic activity of Gen on gastric cancer cell line AGS by proteomic analysis using combination therapy including Gen and other phytochemicals such as taurine, epigallocatechin, and gallate. A proteomics study revealed the involvement of these phytochemicals in the improvement of liver function. A total of 89 protein alterations were reported, out of which four differentially expressed proteins (Tpi1, Txn1, Fgb, and F7) were involved in the glycolysis pathway, coagulation cascade pathway, and antioxidant defense system. Further investigation revealed reduced expression of aspartate transaminase (AST), alanine aminotransferase (ALT), transforming growth factor-β1 (TGF-β1), and collagen I, and increased expression of superoxide dismutase (SOD), total antioxidative capacity (T-AOC), and glutathione peroxidase (GSH-Px). This study suggested the use of combination therapy as an alternative treatment against liver fibrosis [265]. Furthermore, another study provided combinational therapy to rats followed by proteomic analysis of collected liver tissue. Intriguingly, proteomic alteration of 115 proteins was detected, in which 31 proteins were found to be downregulated, whereas 84 were differentially up-regulated. Out of these, three proteins including Txn1, Ctsd, and Cdk4 were selected for further investigation through real-time PCR and Western blotting. Conclusively, this study suggested the significant correlation of these proteins with liver fibrosis and also clarified the role of combination therapy as a potential intervention for the treatment of liver fibrosis [266].
Besides these, Gen has been found to be a promising candidate against neurodegenerative diseases as well. Interestingly, it has displayed antioxidant potential via annihilation of free radicals [287] and amelioration of antioxidant enzyme activity [288], and thus plausibly leads to prevention and/or treatment of Alzheimer’s disease pathology [47]. It has been shown to antagonize the toxicity of amyloid β-protein (Aβ), and thereby could be useful as an intriguing neuroprotective agent. It has been demonstrated that Gen considerably decreases Aβ production seemingly through inhibition of Beta-site (βsite) amyloid precursor protein (APP) cleaving enzyme 1 (BACE1) [48,49]. It also counteracts the progression of Parkinson’s disease through several intricate mechanisms [50].
Collectively, as more and more data are gleaned, proteomics studies will be highly instrumental in broadening our understanding regarding the molecular intricacies of Gen-based therapeutic interventions.
2.3. Proteomic Approaches to Understand the Function of Tan IIA as Therapeutic Intervention
Tan IIA has been widely known for its therapeutic potential against myriads of disease pathologies [6,11]. Interestingly, but not surprisingly, analysis of its therapeutic potential from a proteomic perspective would be highly instrumental for a comprehensive understanding of their mechanisms of action. Accordingly, several reports have highlighted the molecular intricacies underlying the therapeutic potential of Tan IIA against various disease pathologies employing proteomic studies. To this end, Pan et al. focused on the changes in the proteome of HeLa cancer cells treated with Tan IIA using MALDI-TOF analysis. Their proteomic data revealed the alteration of 12 differentially regulated proteins in the HeLa cancer cells following treatment with Tan IIA. The expression levels of proteins such as heat shock protein 27 (HSP27), vimentin, tubulin, and vinculin that play an important role in signal transduction pathways, energy metabolism, motility and microtubule assembly were found to be considerably modified. They proposed that these protein molecules could be related to HeLa cell growth inhibition [268]. Interestingly, the authors envisaged that the plausible contributions of these proteins to the cytotoxicity of Tan IIA seemingly provides intriguing opportunities for the development of Tan IIA-based cancer therapeutics.
In another study, Long et al. performed label free nano-LC-MS/MS-based proteomic analysis to identify the proteome changes in hepatocellular cancer cells (MHCC97-H) in response to Tan IIA treatment. They identified ~41 significantly altered proteins; most of them were associated with various cellular functions such as microtubule movement, stress resistance, cytoskeletal organization, and translational and transcriptional regulation. Furthermore, they proposed that these proteins could play an important role in hepatocellular cancer tumorigenesis [269]. Further, in a similar kind of study, Lin et al. performed iTRAQ-based proteomics studies along with RNA-seq transcriptomics studies and showed that the suppression of AGS gastric cancer cell growth was seemingly due to alteration of glucose metabolism. They further identified ~102 altered signature proteins. Gene enrichment analysis led to the discovery of the role of various dysregulated proteins in alteration of several key cellular functions such as apoptosis, cell cycle, DNA damage, carbohydrate metabolism, and cytoskeleton reorganization functions. In particular, they identified down-regulation of L-lactate dehydrogenase B chains and glucose-6-phosphate isomerase proteins. These results suggested that Tan IIA plays a key role in the blocking of glucose metabolism, thereby inhibiting cell proliferation [270]. Similarly, 2-DE proteomic analysis of MG-63 osteosarcoma cancer cells with and without treatment with Tan IIA along with other two anticancer agents, i.e., Ginsenoside Rg1 and Cinnamic acid, identified prohibitin as a dramatically down-regulated protein in the nuclear matrix in the treated cells. Authors validated this protein using Western blot and immunogold electro-microscopy analysis. They also stated that prohibitin acts as a molecular chaperone and regulates several oncogenes as well as tumor suppressor genes, thereby playing an important role in cancer treatment [272]. Further, Yin et al. performed phosphoproteomics analysis for the Tan IIA treated AGS gastric cell line through a label free proteomic approach using linear ion trap (LTQ)-Orbitrap. In this study, they identified HSP27 phosphorylation at serine 82 in response to Tan IIA treatment. Moreover, they reported that phosphorylation of HSP27 leads to the production of ROS in the gastric cancer cells, emphasizing its importance in cancer cell apoptosis [273].
Further, other proteomic studies have highlighted the molecular intricacies of Tan IIA against various other human diseases such as liver fibrosis, cardiac failure, and nephropathy. To this end, Pan et al. used a 2-DE-based proteomic approach followed by MALDI-TOF analysis to identify the global proteome modifications of HSC-T6 cell line treated with Tan IIA. In this study, they found 13 proteins with altered expression patterns and among these proteins, prohibitin showed an upregulated pattern. They further validated this study using Western blotting analysis, while knockdown studies revealed its role in attenuation of apoptosis in liver fibrosis [271]. In another study, Yan et al. studied the proteomic alterations in transverse aortic constriction (TAC)-mediated cardiac failure and the cardio protective function of Tan IIA using label free Liquid chromatography-matrix-assisted laser desorption/ionization mass spectrometry (LC-MALDI-MS) proteomic approach. They identified 44 differentially regulated proteins common in control vs. saline-TAC and Tan IIA-TAC vs. saline-TAC. Most of these proteins were associated with mitochondrial function of myocardial cells. Proteins such as carnitine palmitoyl transferase I (CPT-1) and glucose transporter type 4 (GLUT-4) were downregulated when the myocardial cells were treated with TAC, whereas their expression levels recovered to normal after treatment with Tan IIA. These proteins are involved in the metabolic activity of mitochondria by regulating free fatty acids and glucose transfers [276]. In a similar study, Liu et al. performed the 2-DE-based proteomic approach followed by MALDI-TOF MS/MS analysis to identify the protein alterations in doxorubicin nephropathy vs. Tan IIA mediated kidney protection. They identified 17 altered proteins in the control male Kunming (KM)-mice as compared to doxorubicin-treated mice. Twelve of these proteins exhibited downregulation while five of them showed upregulation. Further analysis revealed that out of these twelve downregulated proteins, eight were significantly reversed when doxorubicin-treated male KM-mice were further treated with Tan IIA. Similarly, out of five upregulated proteins, expression of three proteins were reversed following treatment with Tan IIA. These proteins were found to be mainly involved in various cellular activities such as oxidative stress, protein synthesis, cytoskeleton synthesis, etc. [278].
Furthermore, Tan IIA has shown intriguing neuroprotective potential against Alzheimer’s, Parkinson’s, and multiple sclerosis [289,290]. Tan IIA has displayed inhibition of acetylcholinesterase and butyrylcholinestrase, which cause degradation of acetylcholine and thus disrupts cholinergic neurotransmission [57,58]. Interestingly, it has shown promising activity as a learning and memory booster and neuroprotectant against the Aβ plaque- and APP-induced AD symptoms in rodents. Moreover, the neuroprotective effect of Tan IIA is exerted by its anti-inflammatory effect in the brain as indicated by attenuation of astrocytic and microglial activation, proinflammatory cytokines (TNF-α, IL-1β, and IL-6) production, and NF-κB signaling in the cortex and hippocampus in the brains of mice [59]. Likewise, Tan IIA has been demonstrated to ameliorate 6-hydroxydopamine (6-OHDA)-induced dopaminergic neuronal loss seemingly through activation of the NF-E2-related factor 2 (Nrf2)–antioxidant response element (ARE) signal transduction pathways [60].
In summation, it is envisaged that proteomics studies have been highly instrumental in broadening our understanding regarding the molecular intricacies of Tan IIA-based therapeutic interventions.
3. Conclusions
Accumulating evidence has highlighted the therapeutic potential of various phytochemicals for the treatment and management of various disease conditions. It is reasonable to argue that the pharmaceutical industry is seeking a gradual shift from chemically derived drugs to phytochemically derived drugs. Numerous proteomics-based studies have highlighted the promising effects of Cur, Gen, and Tan IIA against various pathological conditions including microbial infections, metabolic disorders, cancer, neurodegenerative diseases, and soon; and provided a molecular rationale for their therapeutic potentials. It is envisaged that proteomics as a technology is evolving at a fast pace; henceforth, with the continued technical advancements, it would be highly instrumental in orchestrating much deeper insights in phytochemical-based therapeutic interventions.
Acknowledgments
We acknowledge the funding support by center-based grant from Zayed Center for Health Sciences, United Arab Emirates University (UAEU-ZCHS) (Grant number #31R235) and the UAEU postdoc grant (Grant number #31R241).
Abbreviations
| ACTC1 | Alpha cardiac muscle protein |
| AEC | Alveolar epithelial cells |
| ALT | Alanine aminotransferase |
| AST | Aspartate transaminase |
| ATR | Ataxia telangiectasia serine/threonine-protein kinase |
| BLM | Bleomycin |
| BPA | Bisphenol A |
| BRCA1 | Breast cancer susceptibility gene 1 |
| BTF3 | Basic transcription factor 3 |
| CPT-1 | Carnitine palmitoyl transferase I |
| CRC | Colorectal cancer |
| CRK2 | Cysteine-rich receptor-like protein kinase |
| Cur-P | Curcumin probe |
| CUR/CDP | Curcumin/β-cyclodextrin polymer |
| 2-DE | Two-dimensional gel electrophoresis |
| 2D-DIGE | Two-dimensional differential gel electrophoresis |
| 2-DE/ESI-LC-MS | Two-dimensional liquid chromatography/electrospray ionization mass spectrometry |
| DJ-1 (PARK7) | Protein deglycase DJ-1, also known as Parkinson disease protein 7 |
| DMSO | Dimethyl sulfoxide |
| ECM | Extracellular matrix |
| eEF1 | Eukaryotic elongation factor 1 |
| EGF-R | Epidermal growth factor receptor |
| eIF2 | Eukaryotic Initiation Factor 2 |
| elF-3 | Eukaryotic initiation factor 3 |
| ER | Endoplasmic reticulum |
| FN-1 | Fibronectin-1 |
| FtsZ | Filamenting temperature-sensitive mutant Z |
| GBM | Glioblastoma multiforme |
| GLUT-4 | Glucose transporter type 4 |
| GSH-Px | Glutathione peroxidase |
| GSN | Gelsolin |
| GSTM5 | Glutathione S-transferase Mu 5 |
| HNRPF | Heterogeneous nuclear ribonucleoprotein F |
| HSC71 | Heat shock cognate 71 |
| HSL | Hormone-sensitive lipase |
| HSP27 | Heat shock protein 27 |
| IPA | Ingenuity pathways analysis |
| iTRAQ | Isobaric tags for relative and absolute quantitation |
| JNK | c-Jun N-terminal kinase |
| KM-mice | Kunming mice |
| LC-MALDI-MS | Liquid chromatography-matrix-assisted laser desorption/ionization mass spectrometry |
| LC-MS/MS | Liquid Chromatography with tandem mass spectrometry |
| LECs | Lens epithelial cells |
| LFQ | Label free quantitation |
| LTQ-Orbitrap | Linear ion trap-Orbitrap |
| MALDI-TOF/TOF-MS | Matrix-assisted laser desorption ionization tandem time-of-flight mass spectrometry |
| MBzP | Mono-benzyl phthalate |
| MEHP | Mono-ethyl hexyl phthalate |
| MLC | Myosin light chain |
| MLCK | Myosin light chain kinase |
| MMP3 | Matrix metalloproteinase-3 |
| MRM | Multiple reaction monitoring |
| MS | Mass spectrometry |
| mTOR | Mammalian target of rapamycin |
| NPM1 | Nucleophosmin |
| PCO | Posterior capsular opacification |
| PDI | Protein disulphide isomerase |
| PDIA3 | Protein disulfide isomerase A3 |
| PGAM1 | Phosphoglycerate mutase 1 |
| PGC-1α | Peroxisome proliferator-activated receptor gamma coactivator 1-alpha |
| PGK1 | Phosphoglycerate kinase-1 |
| PRDX4 | Peroxiredoxin-4 |
| PRDX6 | Peroxiredoxin-6 |
| Q-Orbitrap MS | Quadrupole-Orbitrap Mass Spectrometer |
| RanBP-1 | Ran binding protein-1 |
| rhbFGF | Recombinant human basic fibroblast growth factor |
| ROCK2 | Rho associated coiled-coil containing protein kinase 2 |
| ROS | Reactive oxygen species |
| SILAC | Stable isotope labeling by amino acids in cell culture |
| SOD | Superoxide dismutase |
| STAT3 | Signal transducer and activator of transcription 3 |
| SWATH-MS | Sequential window acquisition of all theoretical mass spectra |
| SDS-PAGE | Sodium dodecyl-sulfate polyacrylamide gel electrophoresis |
| T-AOC | Total antioxidant capacity |
| TAC | Transverse aortic constriction |
| Tan IIA | Tanshinone IIA |
| TGF-β1 | Transforming growth factor-β1 |
| TMT | Tandem mass tags |
| TNBC | Triple-negative breast cancer |
| TPI | Triosephosphate isomerase |
| UCP1 | Uncoupling protein 1 |
| UGT | Uridine diphosphate glucuronosyltransferase |
| VEGF-R2 | Vascular endothelial growth factor receptor 2 |
| VHSV | Viral hemorrhagic septicemia virus |
Author Contributions
Conceptualization: F.B.K., P.S., S.A.A. and M.A.A.; Writing—original draft preparation: F.B.K., P.S., S.A.A., H.K., M.K.J. and M.A.A.; Writing—review and editing: F.B.K., P.S., Y.F.J., S.A.A., A., S.U., Q.Z., M.K.J., M.K., M.O., C.Y.H., V.C., C.A., L.C.M., W.A., H.K. and M.A.A. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Funding Statement
This research received no external funding.
Footnotes
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content.
References
- 1.Choudhary N., Tewari D., Nabavi S.F., Kashani H.R.K., Lorigooini Z., Filosa R., Khan F.B., Masoudian N. Plant based food bioactives: A boon or bane for neurological disorders. Crit. Rev. Food Sci. Nutr. 2022:1–47. doi: 10.1080/10408398.2022.2131729. [DOI] [PubMed] [Google Scholar]
- 2.Khan F.B., Ansari M.A., Uddin S., Palakott A.R., Anwar I., Almatroudi A., Alomary M.N., Alrumaihi F., Alkhayl F.F.A., Alghamdi S., et al. Prospective Role of Bioactive Molecules and Exosomes in the Therapeutic Potential of Camel Milk against Human Diseases: An Updated Perspective. Life. 2022;12:990. doi: 10.3390/life12070990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Patil K., Khan F.B., Akhtar S., Ahmad A., Uddin S. The plasticity of pancreatic cancer stem cells: Implications in therapeutic resistance. Cancer Metastasis Rev. 2021;40:691–720. doi: 10.1007/s10555-021-09979-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Chiang J.-T., Badrealam K.F., Shibu M.A., Kuo C.-H., Huang C.-Y., Chen B.-C., Lin Y.-M., Viswanadha V.P., Kuo W.-W. Eriobotrya japonica ameliorates cardiac hypertrophy in H9c2 cardiomyoblast and in spontaneously hypertensive rats. Environ. Toxicol. 2018;33:1113–1122. doi: 10.1002/tox.22589. [DOI] [PubMed] [Google Scholar]
- 5.Chiang J.-T., Badrealam K.F., Shibu M.A., Cheng S.-F., Shen C.-Y., Chang C.-F., Lin Y.-M., Viswanadha V.P., Liao S.-C., Huang C.-Y. Anti-Apoptosis and Anti-Fibrosis Effects of Eriobotrya Japonica in Spontaneously Hypertensive Rat Hearts. Int. J. Mol. Sci. 2018;19:1638. doi: 10.3390/ijms19061638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ansari M.A., Khan F.B., Safdari H.A., Almatroudi A., Alzohairy M.A., Safdari M., Amirizadeh M., Rehman S., Equbal M.J., Hoque M. Prospective therapeutic potential of Tanshinone IIA: An updated overview. Pharmacol. Res. 2020;164:105364. doi: 10.1016/j.phrs.2020.105364. [DOI] [PubMed] [Google Scholar]
- 7.Forni C., Facchiano F., Bartoli M., Pieretti S., Facchiano A., D’Arcangelo D., Norelli S., Valle G., Nisini R., Beninati S., et al. Beneficial Role of Phytochemicals on Oxidative Stress and Age-Related Diseases. BioMed Res. Int. 2019;2019:8748253. doi: 10.1155/2019/8748253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ansari M.A., Badrealam K.F., Alam A., Tufail S., Khalique G., Equbal M.J., Alzohairy M.A., Almatroudi A., Alomary M.N., Pottoo F.H. Recent Nano-based Therapeutic Intervention of Bioactive Sesquiterpenes: Prospects in Cancer Therapeutics. Curr. Pharm. Des. 2020;26:1138–1144. doi: 10.2174/1381612826666200116151522. [DOI] [PubMed] [Google Scholar]
- 9.Abdullah , Khan M.A., Ahmad W., Ahmad M., Adhikari A., Ibrar M., Rehman M.U., Asif M.J.D., Toxicology C. Antioxidant, antinociceptive, anti-inflammatory, and hepatoprotective activities of pentacyclic triterpenes isolated from Ziziphus oxyphylla Edgew. Drug Chem. Toxicol. 2021;45:1796–1807. doi: 10.1080/01480545.2021.1880427. [DOI] [PubMed] [Google Scholar]
- 10.Abdullah M.A.K., Ahmad W., Adhikari A., Ibrar M., ur Rehman M., Khan M., Kamil A. Exploration of Hepatoprotective Potential and Phytochemicals of Ziziphus oxyphylla Edgew. Pak. Vet. J. 2020;40:431–436. doi: 10.29261/pakvetj/2020.054. [DOI] [Google Scholar]
- 11.Alam S.S.M., Uddin F., Khan F.B., Kamal M.A., Hoque M. Therapeutic and pharmacological potential of Tanshinones against lung cancer: A systematic review. Phytomed. Plus. 2022;2:100202. doi: 10.1016/j.phyplu.2021.100202. [DOI] [Google Scholar]
- 12.Chen K.-J., Hui K.K., Lee M.S., Xu H. The Potential Benefit of Complementary/Alternative Medicine in Cardiovascular Diseases. Evid. -Based Complement. Altern. Med. 2012;2012:1. doi: 10.1155/2012/125029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Afendi F.M., Okada T., Yamazaki M., Hirai-Morita A., Nakamura Y., Nakamura K., Ikeda S., Takahashi H., Amin A.U., Darusman L.K., et al. KNApSAcK Family Databases: Integrated Metabolite–Plant Species Databases for Multifaceted Plant Research. Plant Cell Physiol. 2011;53:e1. doi: 10.1093/pcp/pcr165. [DOI] [PubMed] [Google Scholar]
- 14.Xue R., Fang Z., Zhang M., Yi Z., Wen C., Shi T. TCMID: Traditional Chinese medicine integrative database for herb molecular mechanism analysis. Nucleic Acids Res. 2012;41:D1089–D1095. doi: 10.1093/nar/gks1100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gu J., Gui Y., Chen L., Yuan G., Xu X. CVDHD: A cardiovascular disease herbal database for drug discovery and network pharmacology. J. Chemin. 2013;5:51. doi: 10.1186/1758-2946-5-51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Jensen K., Panagiotou G., Kouskoumvekaki I. Integrated text mining and chemoinformatics analysis associates diet to health benefit at molecular level. PLoS Comput. Biol. 2014;10:e1003432. doi: 10.1371/annotation/96a702bd-85a5-49d9-8fcc-3aad7aa4afa7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Zhang D., Tai Y.-C., Wong C.-H.S., Tai L.-K., Koay E.S.-C., Chen C.-S. Molecular response of leukemia HL-60 cells to genistein treatment, a proteomics study. Leuk. Res. 2007;31:75–82. doi: 10.1016/j.leukres.2006.02.026. [DOI] [PubMed] [Google Scholar]
- 18.Mohanraj K., Karthikeyan B.S., Vivek-Ananth R.P., Chand R.P.B., Aparna S.R., Mangalapandi P., Samal A. IMPPAT: A curated database of Indian Medicinal Plants, Phytochemistry And Therapeutics. Sci. Rep. 2018;8:4329. doi: 10.1038/s41598-018-22631-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Newman D.J., Cragg G.M. Natural products as sources of new drugs over the last 25 years. J. Nat. Prod. 2007;70:461–477. doi: 10.1021/np068054v. [DOI] [PubMed] [Google Scholar]
- 20.Cutler G.J., Nettleton J.A., Ross J.A., Harnack L.J., Jacobs D.R., Jr., Scrafford C.G., Barraj L.M., Mink P.J., Robien K. Dietary flavonoid intake and risk of cancer in postmenopausal women: The Iowa Women’s Health Study. Int. J. Cancer. 2008;123:664–671. doi: 10.1002/ijc.23564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Key T.J., Sharp G.B., Appleby P.N., Beral V., Goodman M.T., Soda M., Mabuchi K. Soya foods and breast cancer risk: A prospective study in Hiroshima and Nagasaki, Japan. Br. J. Cancer. 1999;81:1248–1256. doi: 10.1038/sj.bjc.6690837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hussain Y., Abdullah , Alsharif K.F., Aschner M., Theyab A., Khan F., Saso L., Khan H. Therapeutic Role of Carotenoids in Blood Cancer: Mechanistic Insights and Therapeutic Potential. Nutrients. 2022;14:1949. doi: 10.3390/nu14091949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Le Marchand L., Murphy S.P., Hankin J.H., Wilkens L.R., Kolonel L.N. Intake of Flavonoids and Lung Cancer. Gynecol. Oncol. 2000;92:154–160. doi: 10.1093/jnci/92.2.154. [DOI] [PubMed] [Google Scholar]
- 24.Su L.J., Arab L.J. Tea consumption and the reduced risk of colon cancer–results from a national prospective cohort study. Public Health Nutr. 2002;5:419–425. doi: 10.1079/PHN2001314. [DOI] [PubMed] [Google Scholar]
- 25.Jakubczyk K., Drużga A., Katarzyna J., Skonieczna-Żydecka K. Antioxidant Potential of Curcumin—A Meta-Analysis of Randomized Clinical Trials. Antioxidants. 2020;9:1092. doi: 10.3390/antiox9111092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Peng Y., Ao M., Dong B., Jiang Y., Yu L., Chen Z., Hu C., Xu R. Anti-Inflammatory Effects of Curcumin in the Inflammatory Diseases: Status, Limitations and Countermeasures. Drug Des. Dev. Ther. 2021;ume 15:4503–4525. doi: 10.2147/DDDT.S327378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Adamczak A., Ożarowski M., Karpiński T.M. Curcumin, a Natural Antimicrobial Agent with Strain-Specific Activity. Pharmaceuticals. 2020;13:153. doi: 10.3390/ph13070153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Moghadamtousi S.Z., Kadir H.A., Hassandarvish P., Tajik H., Abubakar S., Zandi K. A Review on Antibacterial, Antiviral, and Antifungal Activity of Curcumin. BioMed Res. Int. 2014;2014:186864. doi: 10.1155/2014/186864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Jeong E.-H., Vaidya B., Cho S.-Y., Park M.-A., Kaewintajuk K., Kim S.R., Oh M.-J., Choi J.-S., Kwon J., Kim D. Identification of regulators of the early stage of viral hemorrhagic septicemia virus infection during curcumin treatment. Fish Shellfish Immunol. 2015;45:184–193. doi: 10.1016/j.fsi.2015.03.042. [DOI] [PubMed] [Google Scholar]
- 30.Tomeh M.A., Hadianamrei R., Zhao X. A Review of Curcumin and Its Derivatives as Anticancer Agents. Int. J. Mol. Sci. 2019;20:1033. doi: 10.3390/ijms20051033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ribeiro A.P.D., Pavarina A.C., Dovigo L.N., Brunetti I.L., Bagnato V.S., Vergani C.E., Costa C.A.D.S. Phototoxic effect of curcumin on methicillin-resistant Staphylococcus aureus and L929 fibroblasts. Lasers Med. Sci. 2012;28:391–398. doi: 10.1007/s10103-012-1064-9. [DOI] [PubMed] [Google Scholar]
- 32.Hegge A.B., Nielsen T.T., Larsen K.L., Bruzell E., Tønnesen H.H. Impact of Curcumin Supersaturation in Antibacterial Photodynamic Therapy—Effect of Cyclodextrin Type and Amount: Studies on Curcumin and Curcuminoides XLV. J. Pharm. Sci. 2012;101:1524–1537. doi: 10.1002/jps.23046. [DOI] [PubMed] [Google Scholar]
- 33.Santezi C., Reina B.D., de Annunzio S.R., Calixto G., Chorilli M., Dovigo L.N. Photodynamic potential of curcumin in bioadhesive formulations: Optical characteristics and antimicrobial effect against biofilms. Photodiagnosis Photodyn. Ther. 2021;35:102416. doi: 10.1016/j.pdpdt.2021.102416. [DOI] [PubMed] [Google Scholar]
- 34.Goel A., Boland C., Chauhan D.P. Specific inhibition of cyclooxygenase-2 (COX-2) expression by dietary curcumin in HT-29 human colon cancer cells. Cancer Lett. 2001;172:111–118. doi: 10.1016/S0304-3835(01)00655-3. [DOI] [PubMed] [Google Scholar]
- 35.Bezáková L., Košťálová D., Obložinský M., Hoffman P., Pekárová M., Kollárová R., Holková I., Mošovská S., Sturdík E.S. Inhibition of 12/15 lipoxygenase by curcumin and an extract from Curcuma longa L. Ceska Slov. Farm. 2014;63:26–31. [PubMed] [Google Scholar]
- 36.Rainey N.E., Moustapha A., Saric A., Nicolas G., Sureau F., Petit P.X. Iron chelation by curcumin suppresses both curcumin-induced autophagy and cell death together with iron overload neoplastic transformation. Cell Death Discov. 2019;5:150. doi: 10.1038/s41420-019-0234-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Catanzaro M., Corsini E., Rosini M., Racchi M., Lanni C. Immunomodulators Inspired by Nature: A Review on Curcumin and Echinacea. Molecules. 2018;23:2778. doi: 10.3390/molecules23112778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Lim G.P., Chu T., Yang F., Beech W., Frautschy S.A., Cole G.M. The Curry Spice Curcumin Reduces Oxidative Damage and Amyloid Pathology in an Alzheimer Transgenic Mouse. J. Neurosci. 2001;21:8370–8377. doi: 10.1523/JNEUROSCI.21-21-08370.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.McClure R., Ong H., Janve V., Barton S., Zhu M., Li B., Dawes M., Jerome W.G., Anderson A., Massion P., et al. Aerosol Delivery of Curcumin Reduced Amyloid-β Deposition and Improved Cognitive Performance in a Transgenic Model of Alzheimer’s Disease. J. Alzheimer’s Dis. 2016;55:797–811. doi: 10.3233/JAD-160289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Del Tredici K., Braak H. Sporadic Parkinson’s disease: Development and distribution of α-synuclein pathology. Neuropathol. Appl. Neurobiol. 2016;42:33–50. doi: 10.1111/nan.12298. [DOI] [PubMed] [Google Scholar]
- 41.El Nebrisi E., Javed H., Ojha S.K., Oz M., Shehab S. Neuroprotective Effect of Curcumin on the Nigrostriatal Pathway in a 6-Hydroxydopmine-Induced Rat Model of Parkinson’s Disease is Mediated by α7-Nicotinic Receptors. Int. J. Mol. Sci. 2020;21:7329. doi: 10.3390/ijms21197329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Sharifi-Rad J., Quispe C., Imran M., Rauf A., Nadeem M., Gondal T.A., Ahmad B., Atif M., Mubarak M.S., Sytar O., et al. Genistein: An Integrative Overview of Its Mode of Action, Pharmacological Properties, and Health Benefits. Oxidative Med. Cell. Longev. 2021;2021:150. doi: 10.1155/2021/3268136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Goh Y.X., Jalil J., Lam K.W., Husain K., Premakumar C.M. Genistein: A Review on its Anti-Inflammatory Properties. Front. Pharmacol. 2022;13:820969. doi: 10.3389/fphar.2022.820969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Hong H., Landauer M.R., Foriska M.A., Ledney G.D. Antibacterial activity of the soy isoflavone genistein. J. Basic Microbiol. 2006;46:329–335. doi: 10.1002/jobm.200510073. [DOI] [PubMed] [Google Scholar]
- 45.Tuli H.S., Tuorkey M.J., Thakral F., Sak K., Kumar M., Sharma A.K., Sharma U., Jain A., Aggarwal V., Bishayee A. Molecular Mechanisms of Action of Genistein in Cancer: Recent Advances. Front. Pharmacol. 2019;10:1336. doi: 10.3389/fphar.2019.01336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Tanjak P., Thiantanawat A., Watcharasit P., Satayavivad J. Genistein reduces the activation of AKT and EGFR, and the production of IL6 in cholangiocarcinoma cells involving estrogen and estrogen receptors. Int. J. Oncol. 2018;53:177–188. doi: 10.3892/ijo.2018.4375. [DOI] [PubMed] [Google Scholar]
- 47.Polkowski K., Mazurek A.P. Biological properties of genistein. A review of in vitro and in vivo data. Acta Pol. Pharm. -Drug Res. 2000;57:30–38. [PubMed] [Google Scholar]
- 48.Li R., He P., Cui J., Staufenbiel M., Harada N., Shen Y. Brain Endogenous Estrogen Levels Determine Responses to Estrogen Replacement Therapy via Regulation of BACE1 and NEP in Female Alzheimer’s Transgenic Mice. Mol. Neurobiol. 2012;47:857–867. doi: 10.1007/s12035-012-8377-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Youn K., Park J.-H., Lee S., Lee S., Lee J., Yun E.-Y., Jeong W.-S., Jun M. BACE1 Inhibition by Genistein: Biological Evaluation, Kinetic Analysis, and Molecular Docking Simulation. J. Med. Food. 2018;21:416–420. doi: 10.1089/jmf.2017.4068. [DOI] [PubMed] [Google Scholar]
- 50.Arbabi E., Hamidi G., Talaei S.A., Salami M. Estrogen agonist genistein differentially influences the cognitive and motor disorders in an ovariectomized animal model of Parkinsonism. Iran. J. Basic Med. Sci. 2016;19:1285–1290. doi: 10.22038/IJBMS.2016.7911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Rhiu S., Chae M.K., Lee E.J., Lee J.B., Yoon J.S., Huang W., Zhang X. Effect of Tanshinone IIA in an In Vitro Model of Graves’ Orbitopathy. Investig. Opthalmol. Vis. Sci. 2014;55:5900–5910. doi: 10.1167/iovs.14-14008. [DOI] [PubMed] [Google Scholar]
- 52.Chen Z., Xu H. Anti-Inflammatory and Immunomodulatory Mechanism of Tanshinone IIA for Atherosclerosis. Evid. -Based Complement. Altern. Med. 2014;2014:267976. doi: 10.1155/2014/267976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Wang D., Zhang W., Wang T., Li N., Mu H., Zhang J., Duan J. Unveiling the Mode of Action of Two Antibacterial Tanshinone Derivatives. Int. J. Mol. Sci. 2015;16:17668–17681. doi: 10.3390/ijms160817668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Fang Z., Zhang M., Liu J.-N., Zhao X., Zhang Y.-Q., Fang L. Tanshinone IIA: A Review of its Anticancer Effects. Front. Pharmacol. 2021;11:611087. doi: 10.3389/fphar.2020.611087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Xing Y., Tu J., Zheng L., Guo L., Xi T. Anti-angiogenic effect of tanshinone IIA involves inhibition of the VEGF/VEGFR2 pathway in vascular endothelial cells. Oncol. Rep. 2014;33:163–170. doi: 10.3892/or.2014.3592. [DOI] [PubMed] [Google Scholar]
- 56.Park Y.-K., Obiang-Obounou B.W., Lee J., Lee T.-Y., Bae M.-A., Hwang K.-S., Lee K.-B., Choi J.-S., Jang B.-C. Anti-Adipogenic Effects on 3T3-L1 Cells and Zebrafish by Tanshinone IIA. Int. J. Mol. Sci. 2017;18:2065. doi: 10.3390/ijms18102065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Ozhan Kocakaya Ş., Ertaş A., Yener I., Ercan B., Oral E., Akdeniz M., Kaplaner E., Topcu G., Kolak U. Selective in-vitro enzymes’ inhibitory activities of fingerprints compounds of Salvia species and molecular docking simulations. Iran. J. Pharm. Res. 2020;19:187–198. doi: 10.22037/ijpr.2020.112498.13801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ślusarczyk S., Senol Deniz F.S., Abel R., Pecio Ł., Pérez-Sánchez H., Cerón-Carrasco J.P., den-Haan H., Banerjee P., Preissner R., Krzyżak E., et al. Norditerpenoids with Selective Anti-Cholinesterase Activity from the Roots of Perovskia atriplicifolia Benth. Int. J. Mol. Sci. 2020;21:4475. doi: 10.3390/ijms21124475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Ding B., Lin C., Liu Q., He Y., Ruganzu J.B., Jin H., Peng X., Ji S., Ma Y., Yang W. Tanshinone IIA attenuates neuroinflammation via inhibiting RAGE/NF-κB signaling pathway in vivo and in vitro. J. Neuroinflamm. 2020;17:302. doi: 10.1186/s12974-020-01981-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Zhang X.S., Ha S., Wang X.L., Shi Y.L., Duan S.S., Li Z.A. Tanshinone IIA protects dopaminergic neurons against 6-hydroxydopamine-induced neurotoxicity through miR-153/NF-E2-related factor 2/antioxidant response element signaling pathway. Neuroscience. 2015;303:489–502. doi: 10.1016/j.neuroscience.2015.06.030. [DOI] [PubMed] [Google Scholar]
- 61.Chung L.Y. The antioxidant properties of garlic compounds: Allyl cysteine, alliin, allicin, and allyl disulfide. J. Med. Food. 2006;9:205–213. doi: 10.1089/jmf.2006.9.205. [DOI] [PubMed] [Google Scholar]
- 62.Ankri S., Mirelman D. Antimicrobial properties of allicin from garlic. Microbes Infect. 1999;1:125–129. doi: 10.1016/S1286-4579(99)80003-3. [DOI] [PubMed] [Google Scholar]
- 63.Kim Y.-S., Kim K.S., Han I., Kim M.-H., Jung M.H., Park H.-K. Quantitative and Qualitative Analysis of the Antifungal Activity of Allicin Alone and in Combination with Antifungal Drugs. PLoS ONE. 2012;7:e38242. doi: 10.1371/journal.pone.0038242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Cutler R.R., Wilson P. Antibacterial activity of a new, stable, aqueous extract of allicin against methicillin-resistant Staphylococcus aureus. Br. J. Biomed. Sci. 2004;61:71–74. doi: 10.1080/09674845.2004.11732646. [DOI] [PubMed] [Google Scholar]
- 65.Rouf R., Uddin S.J., Sarker D.K., Islam M.T., Ali E.S., Shilpi J.A., Nahar L., Tiralongo E., Sarker S.D. Antiviral potential of garlic (Allium sativum) and its organosulfur compounds: A systematic update of pre-clinical and clinical data. Trends Food Sci. Technol. 2020;104:219–234. doi: 10.1016/j.tifs.2020.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Benavides G.A., Squadrito G.L., Mills R.W., Patel H.D., Isbell T.S., Patel R.P., Darley-Usmar V.M., Doeller J.E., Kraus D.W. Hydrogen sulfide mediates the vasoactivity of garlic. Proc. Natl. Acad. Sci. USA. 2007;104:17977–17982. doi: 10.1073/pnas.0705710104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Li X.-H., Li C.-Y., Lu J.-M., Tian R.-B., Wei J. Allicin ameliorates cognitive deficits ageing-induced learning and memory deficits through enhancing of Nrf2 antioxidant signaling pathways. Neurosci. Lett. 2012;514:46–50. doi: 10.1016/j.neulet.2012.02.054. [DOI] [PubMed] [Google Scholar]
- 68.Eilat S., Oestraicher Y., Rabinkov A., Ohad D., Mirelman D., Battler A., Eldar M., Vered Z. Alteration of lipid profile in hyperlipidemic rabbits by allicin, an active constituent of garlic. Coron. Artery Dis. 1995;6:985–990. [PubMed] [Google Scholar]
- 69.Gebhardt R., Beck H., Wagner K.G., Metabolism L. Inhibition of cholesterol biosynthesis by allicin and ajoene in rat hepatocytes and HepG2 cells. Biochim. Et Biophys. Acta (BBA) Lipids Lipid Metab. 1994;1213:57–62. doi: 10.1016/0005-2760(94)90222-4. [DOI] [PubMed] [Google Scholar]
- 70.Oommen S., Anto R.J., Srinivas G., Karunagaran D. Allicin (from garlic) induces caspase-mediated apoptosis in cancer cells. Eur. J. Pharmacol. 2003;485:97–103. doi: 10.1016/j.ejphar.2003.11.059. [DOI] [PubMed] [Google Scholar]
- 71.Park S.-Y., Cho S.-J., Kwon H.-C., Lee K.-R., Rhee D.-K., Pyo S. Caspase-independent cell death by allicin in human epithelial carcinoma cells: Involvement of PKA. Cancer Lett. 2005;224:123–132. doi: 10.1016/j.canlet.2004.10.009. [DOI] [PubMed] [Google Scholar]
- 72.Bat-Chen W., Golan T., Peri I., Ludmer Z., Schwartz B. Allicin Purified From Fresh Garlic Cloves Induces Apoptosis in Colon Cancer Cells Via Nrf2. Nutr. Cancer. 2010;62:947–957. doi: 10.1080/01635581.2010.509837. [DOI] [PubMed] [Google Scholar]
- 73.Gülçin I. Antioxidant Activity of Eugenol: A Structure–Activity Relationship Study. J. Med. Food. 2011;14:975–985. doi: 10.1089/jmf.2010.0197. [DOI] [PubMed] [Google Scholar]
- 74.Barboza J.N., Da Silva Maia Bezerra Filho C., Silva R.O., Medeiros J.V.R., de Sousa D.P. An Overview on the Anti-inflammatory Potential and Antioxidant Profile of Eugenol. Oxidative Med. Cell. Longev. 2018;2018:3957262. doi: 10.1155/2018/3957262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Marchese A., Barbieri R., Coppo E., Orhan I.E., Daglia M., Nabavi S.M., Izadi M., Abdollahi M., Ajami M. Antimicrobial activity of eugenol and essential oils containing eugenol: A mechanistic viewpoint. Crit. Rev. Microbiol. 2017;43:668–689. doi: 10.1080/1040841X.2017.1295225. [DOI] [PubMed] [Google Scholar]
- 76.Zari A.T., Zari T.A., Hakeem K.R. Anticancer Properties of Eugenol: A Review. Molecules. 2021;26:7407. doi: 10.3390/molecules26237407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Hussain A., Brahmbhatt K., Priyani A., Ahmed M., Rizvi T.A., Sharma C. Eugenol Enhances the Chemotherapeutic Potential of Gemcitabine and Induces Anticarcinogenic and Anti-inflammatory Activity in Human Cervical Cancer Cells. Cancer Biother. Radiopharm. 2011;26:519–527. doi: 10.1089/cbr.2010.0925. [DOI] [PubMed] [Google Scholar]
- 78.Kaur G., Athar M., Alam M.S. Eugenol precludes cutaneous chemical carcinogenesis in mouse by preventing oxidative stress and inflammation and by inducing apoptosis. Mol. Carcinog. 2009;49:290–301. doi: 10.1002/mc.20601. [DOI] [PubMed] [Google Scholar]
- 79.Vidhya N., Devaraj S.N. Induction of apoptosis by eugenol in human breast cancer cells. J. Pharmacol. Exp. Ther. 2011;49:871–878. [PubMed] [Google Scholar]
- 80.Pramod K., Ansari S.H., Ali J. Eugenol: A Natural Compound with Versatile Pharmacological Actions. Nat. Prod. Commun. 2010;5:1999–2006. doi: 10.1177/1934578X1000501236. [DOI] [PubMed] [Google Scholar]
- 81.Brodin P., Røed A. Effects of eugenol on rat phrenic nerve and phrenic nerve-diaphragm preparations. Arch. Oral Biol. 1984;29:611–615. doi: 10.1016/0003-9969(84)90130-4. [DOI] [PubMed] [Google Scholar]
- 82.Venkadeswaran K., Muralidharan A.R., Annadurai T., Ruban V.V., Sundararajan M., Anandhi R., Thomas P.A., Geraldine P. Antihypercholesterolemic and Antioxidative Potential of an Extract of the Plant, Piper betle, and Its Active Constituent, Eugenol, in Triton WR-1339-Induced Hypercholesterolemia in Experimental Rats. Evid. -Based Complement. Altern. Med. 2014;2014:478973. doi: 10.1155/2014/478973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Khalil A.A., Rahman U.U., Khan M.R., Sahar A., Mehmood T., Khan M. Essential oil eugenol: Sources, extraction techniques and nutraceutical perspectives. RSC Adv. 2017;7:32669–32681. doi: 10.1039/C7RA04803C. [DOI] [Google Scholar]
- 84.El-Kady A.M., Ahmad A.A., Hassan T.M., El-Deek H.E.M., Fouad S.S., Al-Thaqfan S.S. Eugenol, a potential schistosomicidal agent with anti-inflammatory and antifibrotic effects against Schistosoma mansoni, induced liver pathology. Infect. Drug Resist. 2019;ume 12:709–719. doi: 10.2147/IDR.S196544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Ueda-Nakamura T., Mendonça-Filho R.R., Morgado-Díaz J.A., Maza P.K., Filho B.P.D., Cortez D.A.G., Alviano D.S., Rosa M.D.S.S., Lopes A.H.C., Alviano C.S., et al. Antileishmanial activity of Eugenol-rich essential oil from Ocimum gratissimum. Parasitol. Int. 2005;55:99–105. doi: 10.1016/j.parint.2005.10.006. [DOI] [PubMed] [Google Scholar]
- 86.Kim J.K., Park S.U.J. Recent insights into the biological functions of apigenin. Indian J. Exp. Biol. 2020;19:984–991. doi: 10.17179/excli2020-2579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Lee J.-H., Zhou H.Y., Cho S.Y., Kim Y.S., Lee Y.S., Jeong C.S. Anti-inflammatory mechanisms of apigenin: Inhibition of cyclooxygenase-2 expression, adhesion of monocytes to human umbilical vein endothelial cells, and expression of cellular adhesion molecules. Arch. Pharmacal Res. 2007;30:1318–1327. doi: 10.1007/BF02980273. [DOI] [PubMed] [Google Scholar]
- 88.Escande C., Nin V., Price N.L., Capellini V., Gomes A.P., Barbosa M.T., O’Neil L., White T.A., Sinclair D.A., Chini E.N. Flavonoid apigenin is an inhibitor of the NAD+ ase CD38: Implications for cellular NAD+ metabolism, protein acetylation, and treatment of metabolic syndrome. Diabetes. 2013;62:1084–1093. doi: 10.2337/db12-1139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Cardenas H., Arango D., Nicholas C., Duarte S., Nuovo G.J., He W., Voss O.H., Gonzalez-Mejia M.E., Guttridge D.C., Grotewold E., et al. Dietary apigenin exerts immune-regulatory activity in vivo by reducing NF-κB activity, halting leukocyte infiltration and restoring normal metabolic function. Int. J. Mol. Sci. 2016;17:323. doi: 10.3390/ijms17030323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Kim S., Woo E.-R., Lee D.G. Apigenin promotes antibacterial activity via regulation of nitric oxide and superoxide anion production. J. Basic Microbiol. 2020;60:862–872. doi: 10.1002/jobm.202000432. [DOI] [PubMed] [Google Scholar]
- 91.Shibata C., Ohno M., Otsuka M., Kishikawa T., Goto K., Muroyama R., Kato N., Yoshikawa T., Takata A., Koike K. The flavonoid apigenin inhibits hepatitis C virus replication by decreasing mature microRNA122 levels. Virology. 2014;462–463:42–48. doi: 10.1016/j.virol.2014.05.024. [DOI] [PubMed] [Google Scholar]
- 92.Yan X., Qi M., Li P., Zhan Y., Shao H. Apigenin in cancer therapy: Anti-cancer effects and mechanisms of action. Cell Biosci. 2017;7:50. doi: 10.1186/s13578-017-0179-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Ashrafizadeh M., Bakhoda M.R., Bahmanpour Z., Ilkhani K., Zarrabi A., Makvandi P., Khan H., Mazaheri S., Darvish M., Mirzaei H. Apigenin as Tumor Suppressor in Cancers: Biotherapeutic Activity, Nanodelivery, and Mechanisms With Emphasis on Pancreatic Cancer. Front. Chem. 2020;8:829. doi: 10.3389/fchem.2020.00829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Osigwe C.C., Akah P.A., Nworu C.S., Okoye F.B.C. Apigenin: A methanol fraction component of Newbouldia laevis leaf, as a potential antidiabetic agent. J. Phytopharm. 2017;6:38–44. doi: 10.31254/phyto.2017.6106. [DOI] [Google Scholar]
- 95.Kalra S., Mukherjee J.J., Venkataraman S., Bantwal G., Shaikh S., Saboo B., Das A.K., Ramachandran A. Hypoglycemia: The neglected complication. Indian J. Endocrinol. Metab. 2013;17:819–834. doi: 10.4103/2230-8210.117219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Cazarolli L.H., Folador P., Moresco H.H., Brighente I.M.C., Pizzolatti M.G., Silva F.R.M.B. Mechanism of action of the stimulatory effect of apigenin-6-C-(2″-O-α-l-rhamnopyranosyl)-β-l-fucopyranoside on 14C-glucose uptake. Chem. Interact. 2009;179:407–412. doi: 10.1016/j.cbi.2008.11.012. [DOI] [PubMed] [Google Scholar]
- 97.Balez R., Steiner N., Engel M., Sanz Muñoz S., Lum J.S., Wu Y., Wang D., Vallotton P., Sachdev P., O’Connor M., et al. Neuroprotective effects of apigenin against inflammation, neuronal excitability and apoptosis in an induced pluripotent stem cell model of Alzheimer’s disease. Sci. Rep. 2016;6:31450. doi: 10.1038/srep31450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Ali F., Rahul , Naz F., Jyoti S., Siddique Y.H. Health functionality of apigenin: A review. Int. J. Food Prop. 2017;20:1197–1238. doi: 10.1080/10942912.2016.1207188. [DOI] [Google Scholar]
- 99.Imran M., Ghorat F., Ul-Haq I., Ur-Rehman H., Aslam F., Heydari M., Shariati M.A., Okuskhanova E., Yessimbekov Z., Thiruvengadam M., et al. Lycopene as a Natural Antioxidant Used to Prevent Human Health Disorders. Antioxidants. 2020;9:706. doi: 10.3390/antiox9080706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Campos K.K.D., Araújo G.R., Martins T.L., Bandeira A.C.B., Costa G.D.P., Talvani A., Garcia C.C.M., Oliveira L.A.M., Costa D.C., Bezerra F.S. The antioxidant and anti-inflammatory properties of lycopene in mice lungs exposed to cigarette smoke. J. Nutr. Biochem. 2017;48:9–20. doi: 10.1016/j.jnutbio.2017.06.004. [DOI] [PubMed] [Google Scholar]
- 101.Alshahrani M.Y., Ibrahim E.H., Asiri M., Kilany M., Alkhathami A.G., Chandramoorthy H.C. Lycopene augments and enhances anti-oxidant/antibacterial efficiency of ethanolic leaf extract of Helianthus annuus over multidrug-resistant bacterial isolates. J. King Saud Univ. -Sci. 2022;34:102250. doi: 10.1016/j.jksus.2022.102250. [DOI] [Google Scholar]
- 102.Ono M., Takeshima M., Nakano S. Mechanism of the Anticancer Effect of Lycopene (Tetraterpenoids) Enzymes. 2015;37:139–166. doi: 10.1016/bs.enz.2015.06.002. [DOI] [PubMed] [Google Scholar]
- 103.Islamian J.P., Mehrali H. Lycopene as A Carotenoid Provides Radioprotectant and Antioxidant Effects by Quenching Radiation-Induced Free Radical Singlet Oxygen: An Overview. Cell J. 2015;16:386–391. doi: 10.22074/cellj.2015.485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Jacques P.F., Lyass A., Massaro J.M., Vasan R.S., D’Agostino Sr R.B. Relationship of lycopene intake and consumption of tomato products to incident CVD. Br. J. Nutr. 2013;110:545–551. doi: 10.1017/S0007114512005417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Chen D., Huang C., Chen Z. A review for the pharmacological effect of lycopene in central nervous system disorders. Biomed. Pharmacother. 2019;111:791–801. doi: 10.1016/j.biopha.2018.12.151. [DOI] [PubMed] [Google Scholar]
- 106.Kähkönen M.P., Heinonen M. Antioxidant Activity of Anthocyanins and Their Aglycons. J. Agric. Food Chem. 2003;51:628–633. doi: 10.1021/jf025551i. [DOI] [PubMed] [Google Scholar]
- 107.Ma Z., Du B., Li J., Yang Y., Zhu F. An Insight into Anti-Inflammatory Activities and Inflammation Related Diseases of Anthocyanins: A Review of Both In Vivo and In Vitro Investigations. Int. J. Mol. Sci. 2021;22:11076. doi: 10.3390/ijms222011076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Cisowska A., Wojnicz D., Hendrich A.B. Anthocyanins as Antimicrobial Agents of Natural Plant Origin. Nat. Prod. Commun. 2011;6:149–156. doi: 10.1177/1934578X1100600136. [DOI] [PubMed] [Google Scholar]
- 109.Palungwachira P., Tancharoen S., Phruksaniyom C., Klungsaeng S., Srichan R., Kikuchi K., Nararatwanchai T. Erratum to “Antioxidant and Anti-Inflammatory Properties of Anthocyanins Extracted from Oryza sativa L. in Primary Dermal Fibroblasts”. Oxidative Med. Cell. Longev. 2020;2020:1. doi: 10.1155/2020/6306104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Fragoso M.F., Romualdo G.R., Vanderveer L.A., Franco-Barraza J., Cukierman E., Clapper M.L., Carvalho R.F., Barbisan L.F. Lyophilized açaí pulp (Euterpe oleracea Mart) attenuates colitis-associated colon carcinogenesis while its main anthocyanin has the potential to affect the motility of colon cancer cells. Food Chem. Toxicol. 2018;121:237–245. doi: 10.1016/j.fct.2018.08.078. [DOI] [PubMed] [Google Scholar]
- 111.Qian X., Wang X., Luo J., Liu Y., Pang J., Zhang H., Xu Z., Xie J., Jiang X., Ling W. Hypouricemic and nephroprotective roles of anthocyanins in hyperuricemic mice. Food Funct. 2019;10:867–878. doi: 10.1039/C8FO02124D. [DOI] [PubMed] [Google Scholar]
- 112.Aboonabi A., Singh I., Meyer R.R. Cytoprotective effects of berry anthocyanins against induced oxidative stress and inflammation in primary human diabetic aortic endothelial cells. Chem. Interact. 2020;317:108940. doi: 10.1016/j.cbi.2020.108940. [DOI] [PubMed] [Google Scholar]
- 113.Mattioli R., Francioso A., Mosca L., Silva P. Anthocyanins: A Comprehensive Review of Their Chemical Properties and Health Effects on Cardiovascular and Neurodegenerative Diseases. Molecules. 2020;25:3809. doi: 10.3390/molecules25173809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Joshi R., Gangabhagirathi R. Antioxidant activity of capsaicin on radiation-induced oxidation of murine hepatic mitochondrial membrane preparation. Res. Rep. Biochem. 2015;5:163–171. doi: 10.2147/RRBC.S84270. [DOI] [Google Scholar]
- 115.Kim C.-S., Kawada T., Kim B.-S., Han I.-S., Choe S.-Y., Kurata T., Yu R. Capsaicin exhibits anti-inflammatory property by inhibiting IkB-a degradation in LPS-stimulated peritoneal macrophages. Cell. Signal. 2002;15:299–306. doi: 10.1016/S0898-6568(02)00086-4. [DOI] [PubMed] [Google Scholar]
- 116.Füchtbauer S., Mousavi S., Bereswill S., Heimesaat M.M. Antibacterial properties of capsaicin and its derivatives and their potential to fight antibiotic resistance—A literature survey. Eur. J. Microbiol. Immunol. 2021;11:10–17. doi: 10.1556/1886.2021.00003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Veloso J., Prego C., Varela M.M., Carballeira R., Bernal A., Merino F., Díaz J. Properties of capsaicinoids for the control of fungi and oomycetes pathogenic to pepper. Plant Biol. 2014;16:85–177. doi: 10.1111/j.1438-8677.2012.00717.x. [DOI] [PubMed] [Google Scholar]
- 118.Tang K., Zhang X., Guo Y. Identification of the dietary supplement capsaicin as an inhibitor of Lassa virus entry. Acta Pharm. Sin. B. 2020;10:789–798. doi: 10.1016/j.apsb.2020.02.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Clark R., Lee S.-H. Anticancer Properties of Capsaicin Against Human Cancer. Anticancer Res. 2016;36:837–843. [PubMed] [Google Scholar]
- 120.Fattori V., Hohmann M.S.N., Rossaneis A.C., Pinho-Ribeiro F.A., Verri W.A. Capsaicin: Current understanding of its mechanisms and therapy of pain and other pre-clinical and clinical uses. Molecules. 2016;21:844. doi: 10.3390/molecules21070844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Holzer P., Lippe I. Stimulation of afferent nerve endings by intragastric capsaicin protects against ethanol-induced damage of gastric mucosa. Neuroscience. 1988;27:981–987. doi: 10.1016/0306-4522(88)90201-1. [DOI] [PubMed] [Google Scholar]
- 122.Kawada T., Hagihara K.-I., Iwai K. Effects of Capsaicin on Lipid Metabolism in Rats Fed a High Fat Diet. J. Nutr. 1986;116:1272–1278. doi: 10.1093/jn/116.7.1272. [DOI] [PubMed] [Google Scholar]
- 123.Andersen H.H., Marker J.B., Hoeck E.A., Elberling J., Arendt-Nielsen L. Antipruritic effect of pretreatment with topical capsaicin 8% on histamine-and cowhage-evoked itch in healthy volunteers: A randomized, vehicle-controlled, proof-of-concept trial. Br. J. Dermatol. 2017;177:107–116. doi: 10.1111/bjd.15335. [DOI] [PubMed] [Google Scholar]
- 124.Caetano B.F.R., Tablas M.B., Pereira N.E.F., de Moura N.A., Carvalho R.F., Rodrigues M.A.M., Barbisan L.F. Capsaicin reduces genotoxicity, colonic cell proliferation and preneoplastic lesions induced by 1,2-dimethylhydrazine in rats. Toxicol. Appl. Pharmacol. 2018;338:93–102. doi: 10.1016/j.taap.2017.11.008. [DOI] [PubMed] [Google Scholar]
- 125.Chrubasik S., Pittler M., Roufogalis B. Zingiberis rhizoma: A comprehensive review on the ginger effect and efficacy profiles. Phytomedicine. 2005;12:684–701. doi: 10.1016/j.phymed.2004.07.009. [DOI] [PubMed] [Google Scholar]
- 126.Kim J.-K., Jang H.-D. 6-shogaol attenuates H2O2-induced oxidative stress via upregulation of Nrf2-mediated γ-glutamylcysteine synthetase and heme oxygenase expression in HepG2 cells. Food Sci. Biotechnol. 2016;25:319–327. doi: 10.1007/s10068-016-0045-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Bak M.-J., Ok S., Jun M., Jeong W.-S. 6-Shogaol-Rich Extract from Ginger Up-Regulates the Antioxidant Defense Systems in Cells and Mice. Molecules. 2012;17:8037–8055. doi: 10.3390/molecules17078037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Dugasani S., Pichika M.R., Nadarajah V.D., Balijepalli M.K., Tandra S., Korlakunta J.N. Comparative antioxidant and anti-inflammatory effects of [6]-gingerol, [8]-gingerol, [10]-gingerol and [6]-shogaol. J. Ethnopharmacol. 2010;127:515–520. doi: 10.1016/j.jep.2009.10.004. [DOI] [PubMed] [Google Scholar]
- 129.Han Q., Yuan Q., Meng X., Huo J., Bao Y., Xie G. 6-Shogaol attenuates LPS-induced inflammation in BV2 microglia cells by activating PPAR-γ. Oncotarget. 2017;8:42001. doi: 10.18632/oncotarget.16719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Chen F., Tang Y., Sun Y., Veeraraghavan V.P., Mohan S.K., Cui C. 6-shogaol, a active constiuents of ginger prevents UVB radiation mediated inflammation and oxidative stress through modulating NrF2 signaling in human epidermal keratinocytes (HaCaT cells) J. Photochem. Photobiol. B Biol. 2019;197:111518. doi: 10.1016/j.jphotobiol.2019.111518. [DOI] [PubMed] [Google Scholar]
- 131.Annamalai G., Suresh K. [6]-Shogaol attenuates inflammation, cell proliferation via modulate NF-κB and AP-1 oncogenic signaling in 7, 12-dimethylbenz [a] anthracene induced oral carcinogenesis. Biomed. Pharmacother. 2018;98:484–490. doi: 10.1016/j.biopha.2017.12.009. [DOI] [PubMed] [Google Scholar]
- 132.Rampogu S., Baek A., Gajula R.G., Zeb A., Bavi R.S., Kumar R., Kim Y., Kwon Y.J., Lee K.W. Ginger (Zingiber officinale) phytochemicals—Gingerenone-A and shogaol inhibit SaHPPK: Molecular docking, molecular dynamics simulations and in vitro approaches. Ann. Clin. Microbiol. Antimicrob. 2018;17:16. doi: 10.1186/s12941-018-0266-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Kim H., Eom Y. Antifungal and anti-biofilm effects of 6-shogaol against Candida auris. J. Appl. Microbiol. 2020;130:1142–1153. doi: 10.1111/jam.14870. [DOI] [PubMed] [Google Scholar]
- 134.Hayati R.F., Better C.D., Denis D., Komarudin A.G., Bowolaksono A., Yohan B., Sasmono R.T. [6]-Gingerol Inhibits Chikungunya Virus Infection by Suppressing Viral Replication. BioMed Res. Int. 2021;2021:6623400. doi: 10.1155/2021/6623400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Zadorozhna M., Mangieri D. Mechanisms of chemopreventive and therapeutic proprieties of ginger extracts in cancer. Int. J. Mol. Sci. 2021;22:6599. doi: 10.3390/ijms22126599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Khaki A., Fathiazad F., Nouri M., Afshin Khaki A., Ozanci C.C., Ghafari-Novin M., Hamadeh M. The effects of Ginger on spermatogenesis and sperm parameters of rat. Int. J. Reprod. BioMed. 2009;7:7. doi: 10.1016/S1472-6483(12)60133-3. [DOI] [Google Scholar]
- 137.Smith M.R., Gangireddy S.R., Narala V.R., Hogaboam C.M., Standiford T.J., Christensen P.J., Kondapi A.K., Reddy R.C. Curcumin inhibits fibrosis-related effects in IPF fibroblasts and in mice following bleomycin-induced lung injury. Am. J. Physiol. Cell. Mol. Physiol. 2010;298:L616–L625. doi: 10.1152/ajplung.00002.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Stanić Z. Curcumin, a Compound from Natural Sources, a True Scientific Challenge—A Review. Plant Foods Hum. Nutr. 2017;72:1–12. doi: 10.1007/s11130-016-0590-1. [DOI] [PubMed] [Google Scholar]
- 139.López-Malo D., Villarón-Casares C.A., Alarcón-Jiménez J., Miranda M., Díaz-Llopis M., Romero F.J., Villar V.M. Curcumin as a Therapeutic Option in Retinal Diseases. Antioxidants. 2020;9:48. doi: 10.3390/antiox9010048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Rainey N.E., Moustapha A., Petit P.X. Curcumin, a Multifaceted Hormetic Agent, Mediates an Intricate Crosstalk between Mitochondrial Turnover, Autophagy, and Apoptosis. Oxidative Med. Cell. Longev. 2020;2020:3656419. doi: 10.1155/2020/3656419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.De R., Kundu P., Swarnakar S., Ramamurthy T., Chowdhury A., Nair G.B., Mukhopadhyay A.K. Antimicrobial Activity of Curcumin against Helicobacter pylori Isolates from India and during Infections in Mice. Antimicrob. Agents Chemother. 2009;53:1592–1597. doi: 10.1128/AAC.01242-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Reddy P.J., Sinha S., Ray S., Sathe G.J., Chatterjee A., Prasad T.K., Dhali S., Srikanth R., Panda D., Srivastava S. Comprehensive analysis of temporal alterations in cellular proteome of Bacillus subtilis under curcumin treatment. PLoS ONE. 2015;10:e0120620. doi: 10.1371/journal.pone.0120620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Sharma O. Antioxidant activity of curcumin and related compounds. Biochem. Pharmacol. 1976;25:1811–1812. doi: 10.1016/0006-2952(76)90421-4. [DOI] [PubMed] [Google Scholar]
- 144.Ragunathan I., Panneerselvam N. Antimutagenic potential of curcumin on chromosomal aberrations in Allium cepa. J. Zhejiang Univ. Sci. B. 2007;8:470–475. doi: 10.1631/jzus.2007.B0470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Sun D., Zhuang X., Xiang X., Liu Y., Zhang S., Liu C., Barnes S., Grizzle W., Miller D., Zhang H.-G. A Novel Nanoparticle Drug Delivery System: The Anti-inflammatory Activity of Curcumin Is Enhanced When Encapsulated in Exosomes. Mol. Ther. 2010;18:1606–1614. doi: 10.1038/mt.2010.105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Hatcher H., Planalp R., Cho J., Torti F.M., Torti S.V. Curcumin: From ancient medicine to current clinical trials. Cell Mol. Life Sci. 2008;65:1631–1652. doi: 10.1007/s00018-008-7452-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Ronis M.J.J. Effects of soy containing diet and isoflavones on cytochrome P450 enzyme expression and activity. Drug Metab. Rev. 2016;48:331–341. doi: 10.1080/03602532.2016.1206562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Dixon R.A., Ferreira D. Genistein. Phytochemistry. 2002;60:205–211. doi: 10.1016/S0031-9422(02)00116-4. [DOI] [PubMed] [Google Scholar]
- 149.Sak K. Current epidemiological knowledge about the role of flavonoids in prostate carcinogenesis. Exp. Oncol. 2017;39:98–105. doi: 10.31768/2312-8852.2017.39(2):98-105. [DOI] [PubMed] [Google Scholar]
- 150.Kim I.-S. Current Perspectives on the Beneficial Effects of Soybean Isoflavones and Their Metabolites for Humans. Antioxidants. 2021;10:1064. doi: 10.3390/antiox10071064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Ganai A.A., Farooqi H. Bioactivity of genistein: A review of in vitro and in vivo studies. Biomed. Pharmacother. 2015;76:30–38. doi: 10.1016/j.biopha.2015.10.026. [DOI] [PubMed] [Google Scholar]
- 152.Uifălean A., Schneider S., Ionescu C., Lalk M., Iuga C.A. Soy Isoflavones and Breast Cancer Cell Lines: Molecular Mechanisms and Future Perspectives. Molecules. 2015;21:13. doi: 10.3390/molecules21010013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Kim S.-H., Kim C.-W., Jeon S.-Y., Go R.-E., Hwang K.-A., Choi K.-C. Chemopreventive and chemotherapeutic effects of genistein, a soy isoflavone, upon cancer development and progression in preclinical animal models. Lab. Anim. Res. 2014;30:143–150. doi: 10.5625/lar.2014.30.4.143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Malloy K.M., Wang J., Clark L.H., Fang Z., Sun W., Yin Y., Kong W., Zhou C., Bae-Jump V.L. Novasoy and genistein inhibit endometrial cancer cell proliferation through disruption of the AKT/mTOR and MAPK signaling pathways. Am. J. Transl. Res. 2018;10:784–795. [PMC free article] [PubMed] [Google Scholar]
- 155.Yu Y., Xing Y., Zhang Q., Zhang Q., Huang S., Li X., Gao C. Soy isoflavone genistein inhibits hsa_circ_0031250/miR -873-5p/FOXM1 axis to suppress non-small-cell lung cancer progression. IUBMB Life. 2020;73:92–107. doi: 10.1002/iub.2404. [DOI] [PubMed] [Google Scholar]
- 156.Alorda-Clara M., Torrens-Mas M., Morla-Barcelo P.M., Roca P., Sastre-Serra J., Pons D.G., Oliver J. High Concentrations of Genistein Decrease Cell Viability Depending on Oxidative Stress and Inflammation in Colon Cancer Cell Lines. Int. J. Mol. Sci. 2022;23:7526. doi: 10.3390/ijms23147526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Nie Z.-Y., Zhao M.-H., Cheng B.-Q., Pan R.-F., Wang T.-R., Qin Y., Zhang X.-J. Tanshinone IIA regulates human AML cell proliferation, cell cycle, and apoptosis through miR-497-5p/AKT3 axis. Cancer Cell Int. 2020;20:379. doi: 10.1186/s12935-020-01468-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Liu Z., Zhu W., Kong X., Chen X., Sun X., Zhang W., Zhang R. Tanshinone IIA inhibits glucose metabolism leading to apoptosis in cervical cancer. Oncol. Rep. 2019;42:1893–1903. doi: 10.3892/or.2019.7294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Curtis H., Noll U., Störmann J., Slusarenko A.J. Broad-spectrum activity of the volatile phytoanticipin allicin in extracts of garlic (Allium sativum L.) against plant pathogenic bacteria, fungi and Oomycetes. Physiol. Mol. Plant Pathol. 2004;65:79–89. doi: 10.1016/j.pmpp.2004.11.006. [DOI] [Google Scholar]
- 160.Gruhlke M.C., Nwachwukwu I., Arbach M., Anwar A., Noll U., Slusarenko A.J. Allicin from garlic, effective in controlling several plant diseases, is a reactive sulfur species (RSS) that pushes cells into apoptosis; Proceedings of the Modern Fungicides and Antifungal Compounds VI, 16th International Reinhardsbrunn Symposium; Friedrichroda, Germany. 25–29 April 2010; pp. 325–330. [Google Scholar]
- 161.Ried K., Toben C., Fakler P. Effect of garlic on serum lipids: An updated meta-analysis. Nutr. Rev. 2013;71:282–299. doi: 10.1111/nure.12012. [DOI] [PubMed] [Google Scholar]
- 162.Warshafsky S., Kamer R.S., Sivak S.L. Effect of garlic on total serum cholesterol. A meta-analysis. Ann. Intern. Med. 1993;119:599–605. doi: 10.7326/0003-4819-119-7_Part_1-199310010-00009. [DOI] [PubMed] [Google Scholar]
- 163.Focke M., Feld A., Lichtenthaler H.K. Allicin, a naturally occurring antibiotic from garlic, specifically inhibits acetyl-CoA synthetase. FEBS Lett. 1990;261:106–108. doi: 10.1016/0014-5793(90)80647-2. [DOI] [PubMed] [Google Scholar]
- 164.Gupta N., Porter T.D. Garlic and garlic-derived compounds inhibit human squalene monooxygenase. J. Nutr. 2001;131:1662–1667. doi: 10.1093/jn/131.6.1662. [DOI] [PubMed] [Google Scholar]
- 165.Barceloux D.G. Medical Toxicology of Natural Substances: Foods, Fungi, Medicinal Herbs, Plants, and Venomous Animals. John Wiley & Sons, Inc.; Hoboken, NJ, USA: 2008. [Google Scholar]
- 166.Basch E., Gasparyan A., Giese N., Hashmi S., Miranda M., Sollars D., Seamon E., Tanguay-Colucci S., Ulbricht C., Varghese M., et al. Clove (Eugenia aromatica) and clove oil (eugenol). Natural standard monograph (www. naturalstandard. com) copyright© 2008. J. Diet Suppl. 2008;5:117–146. doi: 10.1080/19390210802335391. [DOI] [PubMed] [Google Scholar]
- 167.Leite A.M., Lima E.D.O., Souza E.L.D., Diniz M.D.F.F.M., Trajano V.N., de Medeiros I.A. Inhibitory effect of beta-pinene, alpha-pinene and eugenol on the growth of potential infectious endocarditis causing Gram-positive bacteria. Rev. Bras. Cienc. Farm. 2007;43:121–126. doi: 10.1590/S1516-93322007000100015. [DOI] [Google Scholar]
- 168.Ali S.M., A Khan A., Ahmed I., Musaddiq M., Ahmed K.S., Polasa H., Rao L.V., Habibullah C.M., A Sechi L., Ahmed N. Antimicrobial activities of Eugenol and Cinnamaldehyde against the human gastric pathogen Helicobacter pylori. Ann. Clin. Microbiol. Antimicrob. 2005;4:20. doi: 10.1186/1476-0711-4-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Leem H.-H., Kim E.-O., Seo M.-J., Choi S.-W. Antioxidant and Anti-Inflammatory Activities of Eugenol and Its Derivatives from Clove (Eugenia caryophyllata Thunb.) J. Korean Soc. Food Sci. Nutr. 2011;40:1361–1370. doi: 10.3746/jkfn.2011.40.10.1361. [DOI] [Google Scholar]
- 170.Kabuto H., Yamanushi T.T. Effects of zingerone [4-(4-hydroxy-3-methoxyphenyl)-2-butanone] and eugenol [2-methoxy-4-(2-propenyl) phenol] on the pathological progress in the 6-hydroxydopamine-induced parkinson’s disease mouse model. Neurochem. Res. 2011;36:2244–2249. doi: 10.1007/s11064-011-0548-5. [DOI] [PubMed] [Google Scholar]
- 171.Harikumar K.B., Kunnumakkara A.B., Sethi G., Diagaradjane P., Anand P., Pandey M.K., Gelovani J., Krishnan S., Guha S., Aggarwal B.B. Resveratrol, a multitargeted agent, can enhance antitumor activity of gemcitabine in vitro and in orthotopic mouse model of human pancreatic cancer. Int. J. Cancer. 2010;127:257–268. doi: 10.1002/ijc.25041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Hume W. Effect of Eugenol on Constrictor Responses in Blood Vessels of the Rabbit Ear. J. Dent. Res. 1983;62:1013–1015. doi: 10.1177/00220345830620090301. [DOI] [PubMed] [Google Scholar]
- 173.Markowitz K., Moynihan M., Liu M., Kim S. Biologic properties of eugenol and zinc oxide-eugenol: A clinically oriented review. Oral Surg. Oral Med. Oral Pathol. 1992;73:729–737. doi: 10.1016/0030-4220(92)90020-Q. [DOI] [PubMed] [Google Scholar]
- 174.Bhagwat S., Haytowitz D.B., Holden J.M. USDA Database for the Flavonoid Content of Selected Foods. J.U.D.o.A.B Release 3. Volume 159. [(accessed on 8 September 2022)];2011 Available online: https://www.ars.usda.gov/ARSUserFiles/80400525/data/flav/flav_r03.pdf.
- 175.Jack C.R., Jr., Bennett D.A., Blennow K., Carrillo M.C., Dunn B., Haeberlein S.B., Holtzman D.M., Jagust W., Jessen F., Karlawish J.J., et al. NIA-AA research framework: Toward a biological definition of Alzheimer’s disease. Alzheimers Dement. 2018;14:535–562. doi: 10.1016/j.jalz.2018.02.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Takagaki N., Sowa Y., Oki T., Nakanishi R., Yogosawa S., Sakai T. Apigenin induces cell cycle arrest and p21/WAF1 expression in a p53-independent pathway. Int. J. Oncol. 2005;26:185–189. doi: 10.3892/ijo.26.1.185. [DOI] [PubMed] [Google Scholar]
- 177.Maggioni D., Garavello W., Rigolio R., Pignataro L., Gaini R., Nicolini G. Apigenin impairs oral squamous cell carcinoma growth in vitro inducing cell cycle arrest and apoptosis. Int. J. Oncol. 2013;43:1675–1682. doi: 10.3892/ijo.2013.2072. [DOI] [PubMed] [Google Scholar]
- 178.Hong S., Dia V.P., Baek S.J., Zhong Q. Nanoencapsulation of apigenin with whey protein isolate: Physicochemical properties, in vitro activity against colorectal cancer cells, and bioavailability. LWT. 2021;154:112751. doi: 10.1016/j.lwt.2021.112751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Seo H.-S., Jo J.K., Ku J.M., Choi H.-S., Choi Y.K., Woo J.-K., Kim H.I., Kang S.-Y., Lee K.M., Nam K.W., et al. Induction of caspase-dependent extrinsic apoptosis by apigenin through inhibition of signal transducer and activator of transcription 3 (STAT3) signalling in HER2-overexpressing BT-474 breast cancer cells. Biosci. Rep. 2015;35:e00276. doi: 10.1042/BSR20150165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Kashyap D., Sharma A., Tuli H.S., Sak K., Garg V.K., Buttar H.S., Setzer W.N., Sethi G. Apigenin: A natural bioactive flavone-type molecule with promising therapeutic function. J. Funct. Foods. 2018;48:457–471. doi: 10.1016/j.jff.2018.07.037. [DOI] [Google Scholar]
- 181.Seo H.-S., Ku J.M., Choi H.-S., Woo J.-K., Jang B.-H., Shin Y.C., Ko S.-G. Induction of caspase-dependent apoptosis by apigenin by inhibiting STAT3 signaling in HER2-overexpressing MDA-MB-453 breast cancer cells. Anticancer Res. 2014;34:2869–2882. [PubMed] [Google Scholar]
- 182.Choudhari S.M., Ananthanarayan L. Enzyme aided extraction of lycopene from tomato tissues. Food Chem. 2007;102:77–81. doi: 10.1016/j.foodchem.2006.04.031. [DOI] [Google Scholar]
- 183.Li N., Wu X., Zhuang W., Xia L., Chen Y., Wu C., Rao Z., Du L., Zhao R., Yi M., et al. Tomato and lycopene and multiple health outcomes: Umbrella review. Food Chem. 2020;343:128396. doi: 10.1016/j.foodchem.2020.128396. [DOI] [PubMed] [Google Scholar]
- 184.Durairajanayagam D., Agarwal A., Ong C., Prashast P. Lycopene and male infertility. Asian J. Androl. 2014;16:420–425. doi: 10.4103/1008-682x.126384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Agarwal S., Rao A.V. Tomato lycopene and low density lipoprotein oxidation: A human dietary intervention study. Lipids. 1998;33:981–984. doi: 10.1007/s11745-998-0295-6. [DOI] [PubMed] [Google Scholar]
- 186.Agarwal S., Rao A.V. Tomato lycopene and its role in human health and chronic diseases. Can. Med. Assoc. J. 2000;163:739–744. [PMC free article] [PubMed] [Google Scholar]
- 187.Zou J., Feng D., Ling W.-H., Duan R.-D. Lycopene suppresses proinflammatory response in lipopolysaccharide-stimulated macrophages by inhibiting ROS-induced trafficking of TLR4 to lipid raft-like domains. J. Nutr. Biochem. 2013;24:1117–1122. doi: 10.1016/j.jnutbio.2012.08.011. [DOI] [PubMed] [Google Scholar]
- 188.Göncü T., Oğuz E., Sezen H., Koçarslan S., Oğuz H., Akal A., Adıbelli F.M., Çakmak S., Aksoy N. Anti-inflammatory effect of lycopene on endotoxin-induced uveitis in rats. Arq. Bras. Oftalmol. 2016;79:357–362. doi: 10.5935/0004-2749.20160102. [DOI] [PubMed] [Google Scholar]
- 189.Moia V.M., Portilho F.L., Pádua T.A., Corrêa L.B., Ricci-Junior E., Rosas E.C., Alencar L.M.R., Sinfronio F.S.M., Sampson A., Iram S.H., et al. Lycopene used as Anti-inflammatory Nanodrug for the Treatment of Rheumathoid Arthritis: Animal assay, Pharmacokinetics, ABC Transporter and Tissue Deposition. Colloids Surf. B Biointerfaces. 2020;188:110814. doi: 10.1016/j.colsurfb.2020.110814. [DOI] [PubMed] [Google Scholar]
- 190.Thies F., Masson L.F., Rudd A., Vaughan N., Tsang C., Brittenden J., Simpson W.G., Duthie S., Horgan G.W., Duthie G. Effect of a tomato-rich diet on markers of cardiovascular disease risk in moderately overweight, disease-free, middle-aged adults: A randomized controlled trial. Am. J. Clin. Nutr. 2012;95:1013–1022. doi: 10.3945/ajcn.111.026286. [DOI] [PubMed] [Google Scholar]
- 191.Kelkel M., Schumacher M., Dicato M., Diederich M. Antioxidant and anti-proliferative properties of lycopene. Free. Radic. Res. 2011;45:925–940. doi: 10.3109/10715762.2011.564168. [DOI] [PubMed] [Google Scholar]
- 192.Riaz M., Zia-Ul-Haq M., Saad B. Anthocyanins and Human Health: Biomolecular and Therapeutic Aspects. Volume 10 Springer; Berlin/Heidelberg, Germany: 2016. [Google Scholar]
- 193.Mazza G., Miniati E. Anthocyanins in Fruits, Vegetables, and Grains. CRC Press; Boca Raton, FL, USA: 2018. [Google Scholar]
- 194.de Pascual-Teresa S., Moreno D.A., García-Viguera C. Flavanols and anthocyanins in cardiovascular health: A review of current evidence. Int. J. Mol. Sci. 2010;11:1679–1703. doi: 10.3390/ijms11041679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Wang L.S., Kuo C.T., Peiffer D., Seguin C., Stoner K., Huang Y.W., Huang T.H., Salzman N., Liu Z., Rosenberg D. Anthocyanins in Health and Disease. CRC Press; Boca Raton, FL, USA: 2013. Anthocyanins, anthocyanin derivatives, and colorectal cancer; pp. 225–243. [Google Scholar]
- 196.Pour P.M., Fakhri S., Asgary S., Farzaei M.H., Echeverría J. The Signaling Pathways, and Therapeutic Targets of Antiviral Agents: Focusing on the Antiviral Approaches and Clinical Perspectives of Anthocyanins in the Management of Viral Diseases. Front. Pharmacol. 2019;10:1207. doi: 10.3389/fphar.2019.01207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Rice-Evans C., Miller N., Paganga G. Antioxidant properties of phenolic compounds. Trends Plant Sci. 1997;2:152–159. doi: 10.1016/S1360-1385(97)01018-2. [DOI] [Google Scholar]
- 198.Jomova K., Valko M. Advances in metal-induced oxidative stress and human disease. Toxicology. 2011;283:65–87. doi: 10.1016/j.tox.2011.03.001. [DOI] [PubMed] [Google Scholar]
- 199.Tsuda T., Horio F., Osawa T. Cyanidin 3-O-β-D-glucoside suppresses nitric oxide production during a zymosan treatment in rats. J. Nutr. Sci. Vitaminol. 2002;48:305–310. doi: 10.3177/jnsv.48.305. [DOI] [PubMed] [Google Scholar]
- 200.Polychronopoulos P., Magiatis P., Skaltsounis A., Myrianthopoulos V., Mikros E., Tarricone A., Musacchio A., Roe S., Pearl L., Leost M.J., et al. Anthocyanin-rich acai (Euterpe oleracea Mart.) fruit pulp fractions attenuate inflammatory stress signaling in mouse brain BV-2 microglial cells. J. Agric. Food Chem. 2004;47:1084–1093. doi: 10.1021/jf203989k. [DOI] [PubMed] [Google Scholar]
- 201.Medda R., Lyros O., Schmidt J.L., Jovanovic N., Nie L., Link B.J., Otterson M.F., Stoner G.D., Shaker R., Rafiee P. Anti inflammatory and anti angiogenic effect of black raspberry extract on human esophageal and intestinal microvascular endothelial cells. Microvasc. Res. 2015;97:167–180. doi: 10.1016/j.mvr.2014.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Park J.-S., Choi M.-A., Kim B.-S., Han I.-S., Kurata T., Yu R. Capsaicin protects against ethanol-induced oxidative injury in the gastric mucosa of rats. Life Sci. 2000;67:3087–3093. doi: 10.1016/S0024-3205(00)00890-0. [DOI] [PubMed] [Google Scholar]
- 203.Amna T., Hwang I., Shang K., Amina M., Al-Musayeib N.M., Al-Deyab S.S. Influence of capsaicin on inflammatory cytokines induced by lipopolysaccharide in myoblast cells under in vitro environment. Pharmacogn. Mag. 2017;13:S26–S32. doi: 10.4103/0973-1296.203984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Gram D.X., Hansen A.J., Wilken M., Elm T., Svendsen O., Carr R.D., Ahrén B., Brand C.L. Plasma calcitonin gene-related peptide is increased prior to obesity, and sensory nerve desensitization by capsaicin improves oral glucose tolerance in obese Zucker rats. Eur. J. Endocrinol. 2005;153:963–969. doi: 10.1530/eje.1.02046. [DOI] [PubMed] [Google Scholar]
- 205.Chen H.Y., Lee Y.H., Chen H.Y., Yeh C.A., Chueh P.J., Lin Y.-M.J. Capsaicin Inhibited Aggressive Phenotypes through Downregulation of Tumor-Associated NADH Oxidase (tNOX) by POU Domain Transcription Factor POU3F2. Molecules. 2016;21:733. doi: 10.3390/molecules21060733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Meral O., Alpay M., Kismali G., Kosova F., Cakir D.U., Pekcan M., Yigit S., Sel T. Capsaicin inhibits cell proliferation by cytochrome c release in gastric cancer cells. Tumor Biol. 2014;35:6485–6492. doi: 10.1007/s13277-014-1864-6. [DOI] [PubMed] [Google Scholar]
- 207.Amantini C., Mosca M., Nabissi M., Lucciarini R., Caprodossi S., Arcella A., Giangaspero F., Santoni G. Capsaicin-induced apoptosis of glioma cells is mediated by TRPV1 vanilloid receptor and requires p38 MAPK activation. J. Neurochem. 2007;102:977–990. doi: 10.1111/j.1471-4159.2007.04582.x. [DOI] [PubMed] [Google Scholar]
- 208.Backonja M.M., Malan T.P., Vanhove G.F., Tobias J. NGX-4010, a High-Concentration Capsaicin Patch, for the Treatment of Postherpetic Neuralgia: A Randomized, Double-Blind, Controlled Study with an Open-Label Extension. Pain Med. 2010;11:600–608. doi: 10.1111/j.1526-4637.2009.00793.x. [DOI] [PubMed] [Google Scholar]
- 209.Tesfaye S. Advances in the management of diabetic peripheral neuropathy. Curr. Opin. Support. Palliat. Care. 2009;3:136–143. doi: 10.1097/SPC.0b013e32832b7df5. [DOI] [PubMed] [Google Scholar]
- 210.Hoeks M., Bagguley T., van Marrewijk C., Smith A., Bowen D., Culligan D., Kolade S., Symeonidis A., Garelius H., Spanoudakis M., et al. Toxic iron species in lower-risk myelodysplastic syndrome patients: Course of disease and effects on outcome. Leukemia. 2020;35:1745–1750. doi: 10.1038/s41375-020-01022-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Kim E.-C., Min J.-K., Kim T.-Y., Lee S.-J., Yang H.-O., Han S., Kim Y.-M., Kwon Y.-G. [6]-Gingerol, a pungent ingredient of ginger, inhibits angiogenesis in vitro and in vivo. Biochem. Biophys. Res. Commun. 2005;335:300–308. doi: 10.1016/j.bbrc.2005.07.076. [DOI] [PubMed] [Google Scholar]
- 212.Schirle M., Bantscheff M., Kuster B. Mass Spectrometry-Based Proteomics in Preclinical Drug Discovery. Chem. Biol. 2012;19:72–84. doi: 10.1016/j.chembiol.2012.01.002. [DOI] [PubMed] [Google Scholar]
- 213.Thomford N.E., Senthebane D.A., Rowe A., Munro D., Seele P., Maroyi A., Dzobo K. Natural Products for Drug Discovery in the 21st Century: Innovations for Novel Drug Discovery. Int. J. Mol. Sci. 2018;19:1578. doi: 10.3390/ijms19061578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Rix U., Superti-Furga G. Target profiling of small molecules by chemical proteomics. Nat. Chem. Biol. 2009;5:616–624. doi: 10.1038/nchembio.216. [DOI] [PubMed] [Google Scholar]
- 215.Aslam B., Basit M., Nisar M.A., Khurshid M., Rasool M.H. Proteomics: Technologies and Their Applications. J. Chromatogr. Sci. 2017;55:182–196. doi: 10.1093/chromsci/bmw167. [DOI] [PubMed] [Google Scholar]
- 216.de Souza L.P., Alseekh S., Scossa F., Fernie A.R. Ultra-high-performance liquid chromatography high-resolution mass spectrometry variants for metabolomics research. Nat. Methods. 2021;18:733–746. doi: 10.1038/s41592-021-01116-4. [DOI] [PubMed] [Google Scholar]
- 217.Chanukuppa V., Taware R., Chatterjee T., Sharma S., More T.H., Taunk K., Kumar S., Santra M.K., Rapole S. Current Understanding of the Potential of Proteomics and Metabolomics Approaches in Cancer Chemoresistance: A Focus on Multiple Myeloma. Curr. Top. Med. Chem. 2019;18:2584–2598. doi: 10.2174/1568026619666181130111202. [DOI] [PubMed] [Google Scholar]
- 218.Beck M., Claassen M., Aebersold R. Comprehensive proteomics. Curr. Opin. Biotechnol. 2011;22:3–8. doi: 10.1016/j.copbio.2010.09.002. [DOI] [PubMed] [Google Scholar]
- 219.Domon B., Aebersold R. Mass Spectrometry and Protein Analysis. Science. 2006;312:212–217. doi: 10.1126/science.1124619. [DOI] [PubMed] [Google Scholar]
- 220.Shlar I., Droby S., Rodov V. Modes of antibacterial action of curcumin under dark and light conditions: A toxicoproteomics approach. J. Proteom. 2017;160:8–20. doi: 10.1016/j.jprot.2017.03.008. [DOI] [PubMed] [Google Scholar]
- 221.Chang K., Cheng Y., Lai M., Hu A. Identification of carbonylated proteins in a bactericidal process induced by curcumin with blue light irradiation on imipenem-resistant Acinetobacter baumannii. Rapid Commun. Mass Spectrom. 2019;34:e8548. doi: 10.1002/rcm.8548. [DOI] [PubMed] [Google Scholar]
- 222.Chen J., Li L., Su J., Li B., Zhang X., Chen T. Proteomic analysis of G2/M arrest triggered by natural borneol/curcumin in HepG2 cells, the importance of the reactive oxygen species-p53 pathway. J. Agric. Food Chem. 2015;63:6440–6449. doi: 10.1021/acs.jafc.5b01773. [DOI] [PubMed] [Google Scholar]
- 223.Jain R., Kulkarni P., Dhali S., Rapole S., Srivastava S. Quantitative proteomic analysis of global effect of LLL12 on U87 cell’s proteome: An insight into the molecular mechanism of LLL12. J. Proteom. 2015;113:127–142. doi: 10.1016/j.jprot.2014.09.020. [DOI] [PubMed] [Google Scholar]
- 224.Wang J., Zhang J., Zhang C.-J., Wong Y.K., Lim T.K., Hua Z.-C., Liu B., Tannenbaum S.R., Shen H.-M., Lin Q. In situ Proteomic Profiling of Curcumin Targets in HCT116 Colon Cancer Cell Line. Sci. Rep. 2016;6:22146. doi: 10.1038/srep22146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Pisano M., Palomba A., Tanca A., Pagnozzi D., Uzzau S., Addis M.F., Dettori M.A., Fabbri D., Palmieri G., Rozzo C. Protein expression changes induced in a malignant melanoma cell line by the curcumin analogue compound D6. BMC Cancer. 2016;16:317. doi: 10.1186/s12885-016-2362-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Chen J., Cao X., Qin X., Liu H., Chen S., Zhong S., Li Y. Proteomic analysis of the molecular mechanism of curcumin/β-cyclodextrin polymer inclusion complex inhibiting HepG2 cells growth. J. Food Biochem. 2019;44:e13119. doi: 10.1111/jfbc.13119. [DOI] [PubMed] [Google Scholar]
- 227.Luo J., Yang J., Yu B.-Y., Liu W., Li M., Zhuang S.-M. Identification of Siah-interacting protein as a potential regulator of apoptosis and curcumin resistance. Oncogene. 2010;29:6357–6366. doi: 10.1038/onc.2010.358. [DOI] [PubMed] [Google Scholar]
- 228.Liu H., Liu Y.-Z., Zhang F., Wang H.-S., Zhang G., Zhou B.-H., Zuo Y.-L., Cai S.-H., Bu X.-Z., Du J. Identification of potential pathways involved in the induction of cell cycle arrest and apoptosis by a new 4-arylidene curcumin analogue T63 in lung cancer cells: A comparative proteomic analysis. Mol. BioSyst. 2014;10:1320–1331. doi: 10.1039/c3mb70553f. [DOI] [PubMed] [Google Scholar]
- 229.Zhu D.-J., Chen X.-W., Wang J.-Z., Ju Y.-L., Ou Yang M.-Z., Zhang W.-J. Proteomic analysis identifies proteins associated with curcumin-enhancing efficacy of irinotecan-induced apoptosis of colorectal cancer LOVO cell. Int. J. Clin. Exp. Pathol. 2014;7:1–15. [PMC free article] [PubMed] [Google Scholar]
- 230.Lee J.-G., McKinney K.Q., Pavlopoulos A.J., Park J.-H., Hwang S. Identification of anti-metastatic drug and natural compound targets in isogenic colorectal cancer cells. J. Proteom. 2015;113:326–336. doi: 10.1016/j.jprot.2014.10.009. [DOI] [PubMed] [Google Scholar]
- 231.Li H., Li L. Relationship of GSTP1 lower expression and multidrug resistance reversing of curcumin on human colon carcinoma cells. Zhonghua Yi Xue Za Zhi. 2015;95:2478–2482. [PubMed] [Google Scholar]
- 232.Sato T., Higuchi Y., Shibagaki Y., Hattori S. Phosphoproteomic Analysis Identifies Signaling Pathways Regulated by Curcumin in Human Colon Cancer Cells. Anticancer Res. 2017;37:4789–4798. doi: 10.21873/anticanres.11885. [DOI] [PubMed] [Google Scholar]
- 233.Cai X., Huang W., Qiao Y., Du S., Chen Y., Chen D., Yu S., Che R., Liu N., Jiang Y. Inhibitory effects of curcumin on gastric cancer cells: A proteomic study of molecular targets. Phytomedicine. 2013;20:495–505. doi: 10.1016/j.phymed.2012.12.007. [DOI] [PubMed] [Google Scholar]
- 234.Maioli E., Torricelli C., Valacchi G. Rottlerin and curcumin: A comparative analysis. Ann. N. Y. Acad. Sci. 2012;1259:65–76. doi: 10.1111/j.1749-6632.2012.06514.x. [DOI] [PubMed] [Google Scholar]
- 235.Kreutz D., Sinthuvanich C., Bileck A., Janker L., Muqaku B., Slany A., Gerner C. Curcumin exerts its antitumor effects in a context dependent fashion. J. Proteom. 2018;182:65–72. doi: 10.1016/j.jprot.2018.05.007. [DOI] [PubMed] [Google Scholar]
- 236.Mittal L., Aryal U.K., Camarillo I.G., Raman V., Sundararajan R. Effective electrochemotherapy with curcumin in MDA-MB-231-human, triple negative breast cancer cells: A global proteomics study. Bioelectrochemistry. 2019;131:107350. doi: 10.1016/j.bioelechem.2019.107350. [DOI] [PubMed] [Google Scholar]
- 237.Sathe G., Pinto S.M., Syed N., Nanjappa V., Solanki H.S., Renuse S., Chavan S., Khan A.A., Patil A.H., Nirujogi R.S., et al. Phosphotyrosine profiling of curcumin-induced signaling. Clin. Proteom. 2016;13:13. doi: 10.1186/s12014-016-9114-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Taverna S., Fontana S., Monteleone F., Pucci M., Saieva L., De Caro V., Cardinale V.G., Giallombardo M., Vicario E., Rolfo C., et al. Curcumin modulates chronic myelogenous leukemia exosomes composition and affects angiogenic phenotype via exosomal miR-21. Oncotarget. 2016;7:30420. doi: 10.18632/oncotarget.8483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 239.Monteleone F., Taverna S., Alessandro R., Fontana S. SWATH-MS based quantitative proteomics analysis reveals that curcumin alters the metabolic enzyme profile of CML cells by affecting the activity of miR-22/IPO7/HIF-1α axis. J. Exp. Clin. Cancer Res. 2018;37:170. doi: 10.1186/s13046-018-0843-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.Lu D., Lin T., Wo X., Wang P., Zhou Y., Yang Z. Proteomic research on anti-atherosclerosis mechanism of curcumin in RAW264.7. China J. Chin. Mater. Medica. 2011;36:1207–1211. [PubMed] [Google Scholar]
- 241.Kim S.W., Choi J.H., Mukherjee R., Hwang K.-C., Yun J.W. Proteomic identification of fat-browning markers in cultured white adipocytes treated with curcumin. Mol. Cell. Biochem. 2016;415:51–66. doi: 10.1007/s11010-016-2676-3. [DOI] [PubMed] [Google Scholar]
- 242.Gouda M.M., Rex D.A.B., Es S.P., Modi P.K., Chanderasekaran J., Bhandary Y.P. Proteomics Analysis Revealed the Importance of Inflammation-Mediated Downstream Pathways and the Protective Role of Curcumin in Bleomycin-Induced Pulmonary Fibrosis in C57BL/6 Mice. J. Proteome Res. 2020;19:2950–2963. doi: 10.1021/acs.jproteome.9b00838. [DOI] [PubMed] [Google Scholar]
- 243.D’Aguanno S., D’Agnano I., De Canio M., Rossi C., Bernardini S., Federici G., Urbani A. Shotgun proteomics and network analysis of neuroblastoma cell lines treated with curcumin. Mol. Biosyst. 2012;8:1068–1077. doi: 10.1039/c2mb05498a. [DOI] [PubMed] [Google Scholar]
- 244.Hu Y.-H., Huang X.-R., Qi M.-X., Hou B.-Y. Curcumin inhibits proliferation of human lens epithelial cells: A proteomic analysis. J. Zhejiang Univ. B. 2012;13:402–407. doi: 10.1631/jzus.B1100278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Asher G.N., Fallon J.K., Smith P.C. UGT concentrations in human rectal tissue after multidose, oral curcumin. Pharmacol. Res. Perspect. 2016;4:e00222. doi: 10.1002/prp2.222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246.Lee D.E., Lee S.J., Kim S.J., Lee H.-S., Kwon O.-S. Curcumin ameliorates nonalcoholic fatty liver disease through inhibition of O-GlcNAcylation. Nutrients. 2019;11:2702. doi: 10.3390/nu11112702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Khoontawad J., Intuyod K., Rucksaken R., Hongsrichan N., Pairojkul C., Pinlaor P., Boonmars T., Wongkham C., Jones A., Plieskatt J., et al. Discovering proteins for chemoprevention and chemotherapy by curcumin in liver fluke infection-induced bile duct cancer. PLoS ONE. 2018;13:e0207405. doi: 10.1371/journal.pone.0207405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248.Gim S.-A., Lee S.-R., Shah F.-A., Koh P.-O. Curcumin attenuates the middle cerebral artery occlusion-induced reduction in γ-enolase expression in an animal model. Lab. Anim. Res. 2015;31:198–203. doi: 10.5625/lar.2015.31.4.198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Shah F.-A., Gim S.-A., Sung J.-H., Jeon S.-J., Kim M.-O., Koh P.-O. Identification of proteins regulated by curcumin in cerebral ischemia. J. Surg. Res. 2015;201:141–148. doi: 10.1016/j.jss.2015.10.025. [DOI] [PubMed] [Google Scholar]
- 250.Barnett M., Young W., Cooney J., Roy N. Metabolomics and Proteomics, and What to Do with All These ‘Omes’: Insights from Nutrigenomic Investigations in New Zealand. Lifestyle Genom. 2014;7:274–282. doi: 10.1159/000381349. [DOI] [PubMed] [Google Scholar]
- 251.Fang Y., Zhang Q., Wang X., Yang X., Wang X., Huang Z., Jiao Y., Wang J. Quantitative phosphoproteomics reveals genistein as a modulator of cell cycle and DNA damage response pathways in triple-negative breast cancer cells. Int. J. Oncol. 2016;48:1016–1028. doi: 10.3892/ijo.2016.3327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 252.Sotoca A.M., Gelpke M.D.S., Boeren S., Ström A., Gustafsson J., Murk A.J., Rietjens I., Vervoort J. Quantitative Proteomics and Transcriptomics Addressing the Estrogen Receptor Subtype-mediated Effects in T47D Breast Cancer Cells Exposed to the Phytoestrogen Genistein. Mol. Cell. Proteom. 2011;10 doi: 10.1074/mcp.M110.002170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Roh T., Kim S.W., Moon S.H., Nam M.J. Genistein induces apoptosis by down-regulating thioredoxin-1 in human hepatocellular carcinoma SNU-449 cells. Food Chem. Toxicol. 2016;97:127–134. doi: 10.1016/j.fct.2016.09.003. [DOI] [PubMed] [Google Scholar]
- 254.Yan G.R., Zou F.Y., Dang B.L., Zhang Y., Yu G., Liu X., He Q.Y. Genistein-induced mitotic arrest of gastric cancer cells by downregulating KIF 20 A, a proteomics study. Proteomics. 2012;12:2391–2399. doi: 10.1002/pmic.201100652. [DOI] [PubMed] [Google Scholar]
- 255.Narasimhan K., Lee Y.M., Lim T.K., A Port S., Han J.-H., Chen C.-S., Lin Q. Genistein exerts anti-leukemic effects on genetically different acute myeloid leukemia cell lines by inhibiting protein synthesis and cell proliferation while inducing apoptosis–molecular insights from an iTRAQ™ quantitative proteomics study. Oncoscience. 2015;2:111–124. doi: 10.18632/oncoscience.120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 256.Sun Z., Biela L.M., Hamilton K.L., Reardon K.F. Concentration-dependent effects of the soy phytoestrogen genistein on the proteome of cultured cardiomyocytes. J. Proteom. 2012;75:3592–3604. doi: 10.1016/j.jprot.2012.04.001. [DOI] [PubMed] [Google Scholar]
- 257.Fuchs D., de Pascual-Teresa S., Rimbach G., Virgili F., Ambra R., Turner R., Daniel H., Wenzel U. Proteome analysis for identification of target proteins of genistein in primary human endothelial cells stressed with oxidized LDL or homocysteine. Eur. J. Nutr. 2004;44:95–104. doi: 10.1007/s00394-004-0499-6. [DOI] [PubMed] [Google Scholar]
- 258.Li Y., Zhu M., Huo Y., Zhang X., Liao M. Anti-fibrosis activity of combination therapy with epigallocatechin gallate, taurine and genistein by regulating glycolysis, gluconeogenesis, and ribosomal and lysosomal signaling pathways in HSC-T6 cells. Exp. Ther. Med. 2018;16:4329–4338. doi: 10.3892/etm.2018.6743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 259.Wang J., Betancourt A.M., Mobley J.A., Lamartiniere C.A. Proteomic Discovery of Genistein Action in the Rat Mammary Gland. J. Proteome Res. 2011;10:1621–1631. doi: 10.1021/pr100974w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 260.Rowell C., Carpenter D.M., Lamartiniere C.A. Chemoprevention of Breast Cancer, Proteomic Discovery of Genistein Action in the Rat Mammary Gland. J. Nutr. 2005;135:2953S–2959S. doi: 10.1093/jn/135.12.2953S. [DOI] [PubMed] [Google Scholar]
- 261.Wang J., Betancourt A., Jenkins S., Biro F., Pinney S.M., Chen D., Russo J., Lamartiniere C.A. Altered blood proteome in girls with high urine concentrations of bisphenol A, genistein, mono-ethyl hexylphthalate and mono-benzyl phthalate. MOJ Proteom. Bioinform. 2015;2:44. doi: 10.15406/mojpb.2015.02.00040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262.Betancourt A., Mobley J.A., Wang J., Jenkins S., Chen N., Kojima K., Russo J., Lamartiniere C.A. Alterations in the Rat Serum Proteome Induced by Prepubertal Exposure to Bisphenol A and Genistein. J. Proteome Res. 2014;13:1502–1514. doi: 10.1021/pr401027q. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263.Schwab K., Stein R., Scheler C., Theuring F. Dietary genistein enhances phosphorylation of regulatory myosin light chain in the myocardium of ovariectomized mice. Electrophoresis. 2012;33:1795–1803. doi: 10.1002/elps.201100607. [DOI] [PubMed] [Google Scholar]
- 264.Schwab K., Neumann B., Vignon-Zellweger N., Fischer A., Stein R., Jungblut P.R., Scheler C., Theuring F. Dietary phytoestrogen supplementation induces sex differences in the myocardial protein pattern of mice: A comparative proteomics study. Proteomics. 2011;11:3887–3904. doi: 10.1002/pmic.201000785. [DOI] [PubMed] [Google Scholar]
- 265.Cao W., Zhou Y., Li Y., Zhang X., He M., Zang N., Zhou Y., Liao M. iTRAQ-based proteomic analysis of combination therapy with taurine, epigallocatechin gallate, and genistein on carbon tetrachloride-induced liver fibrosis in rats. Toxicol. Lett. 2015;232:233–245. doi: 10.1016/j.toxlet.2014.11.009. [DOI] [PubMed] [Google Scholar]
- 266.Cao W., Li Y., Li M., Zhang X., Liao M. Txn1, Ctsd and Cdk4 are key proteins of combination therapy with taurine, epigallocatechin gallate and genistein against liver fibrosis in rats. Biomed. Pharmacother. 2017;85:611–619. doi: 10.1016/j.biopha.2016.11.071. [DOI] [PubMed] [Google Scholar]
- 267.Bagheri M., Rezakhani A., Nyström S., Turkina M.V., Roghani M., Hammarström P., Mohseni S. Amyloid Beta1-40-Induced Astrogliosis and the Effect of Genistein Treatment in Rat: A Three-Dimensional Confocal Morphometric and Proteomic Study. PLoS ONE. 2013;8:e76526. doi: 10.1371/journal.pone.0076526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 268.Pan T.-L., Hung Y.-C., Wang P.-W., Chen S.-T., Hsu T.-K., Sintupisut N., Cheng C.-S., Lyu P.-C. Functional proteomic and structural insights into molecular targets related to the growth inhibitory effect of tanshinone IIA on HeLa cells. Proteomics. 2010;10:914–929. doi: 10.1002/pmic.200900178. [DOI] [PubMed] [Google Scholar]
- 269.Long X., Zhang J., Zhang Y., Yao J., Cai Z., Yang P. Nano-LC-MS/MS based proteomics of hepatocellular carcinoma cells compared to Chang liver cells and tanshinone IIA induction. Mol. BioSyst. 2011;7:1728–1741. doi: 10.1039/c0mb00343c. [DOI] [PubMed] [Google Scholar]
- 270.Lin L.-L., Hsia C.-R., Hsu C.-L., Huang H.-C., Juan H.-F. Integrating transcriptomics and proteomics to show that tanshinone IIA suppresses cell growth by blocking glucose metabolism in gastric cancer cells. BMC Genom. 2015;16:41. doi: 10.1186/s12864-015-1230-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 271.Pan T.-L., Wang P.-W. Explore the Molecular Mechanism of Apoptosis Induced by Tanshinone IIA on Activated Rat Hepatic Stellate Cells. Evid. -Based Complement. Altern. Med. 2012;2012:734987. doi: 10.1155/2012/734987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 272.Shi S.L., Li Q.F., Liu Q.R., Xu D.H., Tang J., Liang Y., Zhao Z.L., Yang L.M. Nuclear matrix protein, prohibitin, was down-regulated and translocated from nucleus to cytoplasm during the differentiation of osteosarcoma MG-63 cells induced by ginsenoside Rg1, cinnamic acid, and tanshinone IIA (RCT) J. Cell Biochem. 2009;108:926–934. doi: 10.1002/jcb.22324. [DOI] [PubMed] [Google Scholar]
- 273.Yin C.-F., Kao S.-C., Hsu C.-L., Chang Y.-W., Cheung C.H.Y., Huang H.-C., Juan H.-F. Phosphoproteome Analysis Reveals Dynamic Heat Shock Protein 27 Phosphorylation in Tanshinone IIA-Induced Cell Death. J. Proteome Res. 2020;19:1620–1634. doi: 10.1021/acs.jproteome.9b00836. [DOI] [PubMed] [Google Scholar]
- 274.Pan T.L., Wang P.W., Hung Y.C., Huang C.H., Rau K.M. Proteomic analysis reveals tanshinone IIA enhances apoptosis of advanced cervix carcinoma CaSki cells through mitochondria intrinsic and endoplasmic reticulum stress pathways. Proteomics. 2013;13:3411–3423. doi: 10.1002/pmic.201300274. [DOI] [PubMed] [Google Scholar]
- 275.Cabrera-Pérez R., Monguió-Tortajada M., Gámez-Valero A., Rojas-Márquez R., Borràs F.E., Roura S., Vives J. Osteogenic commitment of Wharton’s jelly mesenchymal stromal cells: Mechanisms and implications for bioprocess development and clinical application. Stem Cell Res. Ther. 2019;10:356. doi: 10.1186/s13287-019-1450-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 276.Yan S.-H., Zhao N.-W., Geng Z.-R., Shen J.-Y., Liu F.-M., Yan D., Zhou J., Nie C., Huang C.-C., Fang Z.-Y. Modulations of Keap1-Nrf2 signaling axis by TIIA ameliorated the oxidative stress-induced myocardial apoptosis. Free Radic. Biol. Med. 2018;115:191–201. doi: 10.1016/j.freeradbiomed.2017.12.001. [DOI] [PubMed] [Google Scholar]
- 277.Yan Y., Su W., Zeng S., Qian L., Chen X., Wei J., Chen N., Gong Z., Xu Z. Effect and Mechanism of Tanshinone I on the Radiosensitivity of Lung Cancer Cells. Mol. Pharm. 2018;15:4843–4853. doi: 10.1021/acs.molpharmaceut.8b00489. [DOI] [PubMed] [Google Scholar]
- 278.Liu X., Wang Y., Ma C., Zhang L., Wu W., Guan S., Yang M., Wang J., Jiang B., Guo D.-A. Proteomic Assessment of Tanshinone II A Sodium Sulfonate on Doxorubicin Induced Nephropathy. Am. J. Chin. Med. 2011;39:395–409. doi: 10.1142/S0192415X11008907. [DOI] [PubMed] [Google Scholar]
- 279.Weng W., Goel A. Curcumin and colorectal cancer: An update and current perspective on this natural medicine. Semin. Cancer Biol. 2020;80:73–86. doi: 10.1016/j.semcancer.2020.02.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 280.Anand P., Kunnumakkara A.B., Newman R.A., Aggarwal B.B. Bioavailability of curcumin: Problems and promises. Mol. Pharm. 2007;4:807–818. doi: 10.1021/mp700113r. [DOI] [PubMed] [Google Scholar]
- 281.Huang Y.-F., Zhu D.-J., Chen X.-W., Chen Q.-K., Luo Z.-T., Liu C.-C., Wang G.-X., Zhang W.-J., Liao N.-Z. Curcumin enhances the effects of irinotecan on colorectal cancer cells through the generation of reactive oxygen species and activation of the endoplasmic reticulum stress pathway. Oncotarget. 2017;8:40264–40275. doi: 10.18632/oncotarget.16828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 282.Singh L., Sharma S., Xu S., Tewari D., Fang J. Curcumin as a Natural Remedy for Atherosclerosis: A Pharmacological Review. Molecules. 2021;26:4036. doi: 10.3390/molecules26134036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 283.Silvestro S., Sindona C., Bramanti P., Mazzon E. A State of the Art of Antioxidant Properties of Curcuminoids in Neurodegenerative Diseases. Int. J. Mol. Sci. 2021;22:3168. doi: 10.3390/ijms22063168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 284.Gregory J., Vengalasetti Y., Bredesen D., Rao R. Neuroprotective Herbs for the Management of Alzheimer’s Disease. Biomolecules. 2021;11:543. doi: 10.3390/biom11040543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 285.Simeonova R., Zheleva D., Valkova I., Stavrakov G., Philipova I., Atanasova M., Doytchinova I. A Novel Galantamine-Curcumin Hybrid as a Potential Multi-Target Agent against Neurodegenerative Disorders. Molecules. 2021;26:1865. doi: 10.3390/molecules26071865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 286.Goedert M., Spillantini M.G., Del Tredici K., Braak H. 100 years of Lewy pathology. Nat. Rev. Neurol. 2012;9:13–24. doi: 10.1038/nrneurol.2012.242. [DOI] [PubMed] [Google Scholar]
- 287.Vallés S.L., Borrás C., Gambini J., Furriol J., Ortega A., Sastre J., Pallardó F.V., Viña J. Oestradiol or genistein rescues neurons from amyloid beta-induced cell death by inhibiting activation of p38. Aging Cell. 2008;7:112–118. doi: 10.1111/j.1474-9726.2007.00356.x. [DOI] [PubMed] [Google Scholar]
- 288.Cai Q., Wei H. Effect of dietary genistein on antioxidant enzyme activities in SENCAR mice. Nutr. Cancer. 1996;25:1–7. doi: 10.1080/01635589609514423. [DOI] [PubMed] [Google Scholar]
- 289.Hao H., Wang G., Cui N., Li J., Xie L., Ding Z. Pharmacokinetics, Absorption and Tissue Distribution of Tanshinone IIA Solid Dispersion. Planta Medica. 2006;72:1311–1317. doi: 10.1055/s-2006-951698. [DOI] [PubMed] [Google Scholar]
- 290.Zhou L., Zuo Z., Chow M.S.S. Danshen: An Overview of Its Chemistry, Pharmacology, Pharmacokinetics, and Clinical Use. J. Clin. Pharmacol. 2005;45:1345–1359. doi: 10.1177/0091270005282630. [DOI] [PubMed] [Google Scholar]