

Abstract
The antibiotic formulary is threatened by high rates of antimicrobial resistance (AMR) among enteropathogens. Enteric bacteria are exposed to anaerobic conditions within the gastrointestinal tract, yet little is known about how oxygen exposure influences AMR. The facultative anaerobe Vibrio cholerae was chosen as a model to address this knowledge gap. We obtained V. cholerae isolates from 66 cholera patients, sequenced their genomes, and grew them under anaerobic and aerobic conditions with and without three clinically relevant antibiotics (ciprofloxacin, azithromycin, doxycycline). For ciprofloxacin and azithromycin, the minimum inhibitory concentration (MIC) increased under anaerobic conditions compared to aerobic conditions. Using standard resistance breakpoints, the odds of classifying isolates as resistant increased over 10 times for ciprofloxacin and 100 times for azithromycin under anaerobic conditions compared to aerobic conditions. For doxycycline, nearly all isolates were sensitive under both conditions. Using genome-wide association studies, we found associations between genetic elements and AMR phenotypes that varied by oxygen exposure and antibiotic concentrations. These AMR phenotypes were more heritable, and the AMR-associated genetic elements were more often discovered, under anaerobic conditions. These AMR-associated genetic elements are promising targets for future mechanistic research. Our findings provide a rationale to determine whether increased MICs under anaerobic conditions are associated with therapeutic failures and/or microbial escape in cholera patients. If so, there may be a need to determine new AMR breakpoints for anaerobic conditions.
Keywords: antimicrobial resistance, antibiotics, anaerobic, cholera, enteropathogens, Vibrio cholerae
Data Summary
All sequencing data generated in this study are available from the National Center for Biotechnology Information (NCBI) database under BioProject PRJNA818081.
Impact Statement.
Many bacterial pathogens experience anaerobic conditions in the gut, but antimicrobial resistance (AMR) phenotypes are generally tested under ambient aerobic conditions in the laboratory. To better understand AMR under conditions more similar to natural infections, we used Vibrio cholerae as a model enteric pathogen. We sequenced the genomes and assessed the growth of V. cholerae isolates with different concentrations of three antibiotics, under anaerobic and aerobic conditions. In support of the hypothesis that AMR varies according to oxygen exposure, V. cholerae was more resistant to antibiotics under anaerobic conditions. We found many previously known genes associated with resistance; however, some of these genes were only resistance-associated under anaerobic conditions. Resistance to azithromycin and doxycycline only had a detectable genetic component under anaerobic conditions. Together, our results point to distinct genetic mechanisms of resistance under anaerobic conditions and suggest several candidate genes for experimental follow-up.
Introduction
Clinically relevant laboratory methods are essential to gauge the extent to which the antibiotic formulary is threatened by antimicrobial resistance (AMR). Knowledge gaps remain on the degree to which in vitro AMR assays reflect in vivo AMR physiology. Facultative anaerobic pathogens experience hypoxia and anoxia within the gastrointestinal tract, yet AMR assays rely on aerobic conditions [1]. How oxygen exposure effects AMR is poorly understood. To investigate this question, we chose the facultative anaerobe Vibrio cholerae as a model system. In V. cholerae , classic mechanisms for AMR, and physiological pathways for anaerobic respiration and fermentation, are well characterized [2–9]. The disease cholera is also one of the few non-invasive diarrheal diseases for which antibiotics are indicated, albeit conditionally [10–12].
Rehydration is the definitive intervention for acute diarrheal disease [11]; antibiotics are supportive and indicated for only a few diarrheal diseases, including cholera. The World Health Organization (WHO) recommends ciprofloxacin, azithromycin or doxycycline for cholera patients with severe dehydration [10–12]; antibiotics shorten the frequency and duration of diarrhoea. In practice, guideline adherence in cholera-endemic regions may be low out of clinical concern that a patient ‘might’ have cholera and may develop severe dehydration, contributing to rates of inappropriate antibiotic usage that can rise above 90 % [13, 14]. Strong regional associations between antibiotic use and rise of AMR have been observed across enteric taxa [15, 16]. Given that AMR genes frequently co-localize on mobile elements [17], inappropriate single-agent therapy poses a risk of multidrug-resistance selection.
Associations between AMR phenotypes and genotypes are known for the three antibiotics recommended to treat cholera; the cognate AMR mechanisms share commonality across Gram-negative taxa. Ciprofloxacin (a fluoroquinolone) resistance mechanisms include mutations in genes encoding type II topoisomerases: heterotetrameric DNA gyrase (GyrA2,GyrB2) and DNA topoisomerase IV (ParC2,ParE2). Mutations in the quinolone-resistance-determining region (QRDR) of gyrA and parC can yield additive resistance phenotypes [18]. Fluoroquinolone resistance can also arise by efflux pump upregulation, by downregulation of outer membrane porins that permit quinolone entry, and by the expression of the quinolone-resistance protein (Qnr, a pentapeptide repeat protein) that protects the target gyrase protein [18]. Resistance can increase over 30-fold compared to wild-type when strains harbour qnr family genes.
In V. cholerae , diverse AMR genes, including qnr, often reside on an integrative and conjugative element (ICE; SXT in V. cholerae ) [17, 19, 20]. Azithromycin (a macrolide) resistance mechanisms are similarly diverse and include mutations in the 23S rRNA target genes and ribosomal protein genes. Macrolide resistance is conveyed by carriage of rRNA methyltransferase genes (erm) and associated induction mechanisms, cis-acting peptides, efflux systems (e.g. mef, msr), macrolide esterases (e.g. ere), and macrolide phosphotransferases such as mphA, which can reside on the V. cholerae SXT element [21]. Doxycycline (a tetracycline) resistance is conferred by mutations in the 16S rRNA component of the 30S ribosomal subunit [22]. Additional mechanisms include tetracycline-specific ribosomal protection proteins (RPPs), tetracycline-specific efflux pumps [e.g. tet(59)] that can reside on SXT element intrinsic efflux pumps, AraC-family transcriptional activators (e.g. MarA) and cytoplasmic ATP-dependent serine proteases [22].
Associations between AMR phenotypes and genotypes have been studied by random mutagenesis, phenotypic screening and network analyses [23–28], and applied in V. cholerae [29]. These approaches uncovered how the effect of an antibiotic is shaped by a large number of often more subtle physiological perturbations, including altered DNA synthesis/repair, central metabolism/growth and SOS response [30, 31]. AMR assays conducted under aerobic conditions alone may not reflect these physiological perturbations experienced in the host. Within bacteria, aerobic oxidative phosphorylation generates reactive oxygen species (ROS) that are lethal unless a sufficient defence is mounted by factors like superoxide dismutase, catalase and glutathione systems [32, 33]. Under anaerobic conditions, growth rate typically slows and proton motive force is reduced [34, 35], which can have both synergistic and antagonistic effects on antibiotics [31, 36]. In Escherichia coli , ROS are generated after fluroquinolone treatment under aerobic conditions [37], and fluoroquinolone resistance increases under anaerobic conditions [38, 39]. The extent to which tetracyclines and macrolides induce ROS and how anaerobiosis influences resistance and susceptibility is less known [30, 40].
The objective of this study was to compare AMR phenotypes, with underlying genotypes, under aerobic and anaerobic conditions among isolates obtained from cholera patients. The study rationale assumes that the lower gastrointestinal tract of cholera patients is hypoxic/anaerobic, despite animal experiments that suggest aerobic respiration in the upper gastrointestinal tract is important for infection [41, 42]. Using minimum inhibitory concentration (MIC) assays, we found that AMR increased under anaerobic conditions for select antibiotics, and novel genetic targets for AMR were discovered under anaerobic conditions.
Methods
Clinical sample collection
The two sample collections analysed were part of previously published ethical Institutional Review Board (IRB) approved studies [13, 43]. In the primary collection, stool samples were obtained during the spring cholera outbreak period of 2006 at the International Centre for Diarrhoeal Disease Research, Bangladesh (icddr,b) in Dhaka, Bangladesh. Samples were collected prior to hospital administration of antibiotics; patient histories were negative for known antibiotic exposure. The library consisted of 67 V . cholerae isolates (Table S1 available with the online version of this article); paired stool supernatant for MS was available for 50 isolates. In the secondary collection, samples were obtained in 2018 as part of a cholera surveillance study conducted across Bangladesh [13]. Samples were collected at hospital admission independent of reported antibiotic exposure; 277 out of 282 isolates cultured and were analysed to assess generalizability of the AMR profiles identified in the primary collection.
AMR testing
Growth kinetics and the MIC determinations for ciprofloxacin, azithromycin and doxycycline were performed on isolates from the primary collection in LB broth with twofold serial dilutions with concentrations spanning the Clinical and Laboratory Standards Institute (CLSI) MIC breakpoints [1] for V. cholerae (ciprofloxacin, 2 µg ml−1; azithromycin, 8 µg ml−1; doxycycline, 8 µg ml−1) [1]. Isolates were prepared and grown aerobically at 37 ˚C in 15 ml tubes containing 5 ml LB broth at 220 r.p.m. Bacteria were back-diluted to a final optical density at 600 nm of 0.01 (200 μl per well) in LB broth with or without the respective antibiotic dilution-series in black Corning CoStar clear-bottom 96-well plates. Plates were placed in a BioTek Synergy H1 reader pre-warmed to 37 ˚C with the lid on. Anaerobic conditions were generated using a continuous chamber flow (5 % CO2, 95 % N2) and a BioTek CO2/O2 gas controller; anaerobic growth plates were given a 10 min equilibration period. OD600 was measured every 2 min for 8 h at 37 ˚C with orbital shaking at 220 r.p.m. Growth was defined as a culture reaching an OD600 of 0.095 or greater (an increase of ~10-fold or greater). A standard logistic equation was fit to growth curve data using the R package growthcurver version 0.3.0 [44]. Outcome measures were intrinsic growth velocity (growth rate that would occur if there were no restrictions on total population size), carrying capacity (K; maximum possible population size) and area under the curve (AUC). The MIC was determined using a logistic fit for growth over the 12 twofold serial dilutions of the test antibiotic. Binary phenotypic sensitive/resistance categories were set in concordance with CLSI guidelines [45]. In general, CLSI breakpoints are set by clinical and bacteriological response data, pharmacokinetic and pharmacodynamic simulations, and expert working group experience. The breakpoints for V. cholerae are based on aerobic assays. Susceptible (sensitive) is defined by CLSI as the 'category that implies that isolates are inhibited by the usually achievable concentrations of antimicrobial agent when the dosage recommended to treat the site of infection is used’. Those not sensitive were scored as resistant (combines indeterminate and resistant). Three isolates and one reference strain (E7946) were used to assess media acidification during anaerobic and aerobic growth; the pH was measured using dipsticks (pH range 5.2–7.2). The assays were conducted with and without the addition of 20 mM fumarate as an alternative electron acceptor for anaerobic respiration.
Use of catalase to determine whether ROS contribute to antibiotic sensitivity
To test whether the reduction of ROS was associated with increased resistance to antibiotics under aerobic conditions, MICs were determined for two select strains (E7946, EN160) with and without catalase (10 U ml−1; final concentration) added to the media. Growth curves were performed with viable counts as endpoints to determine the minimum dose for lethality by H2O2 or protection by catalase.
Whole-genome sequencing
Genomic DNA was extracted from V. cholerae isolates from the primary collection using the Qiagen DNeasy blood and tissue kit. Library construction was completed using the Illumina Nextera XT v.2 DNA library preparation kit. Libraries were sequenced in three Illumina MiSeq runs. Two batches of 24 genomes and one batch of 19 were pooled and sequenced on a MiSeq for 500 cycles per run. Using CLC Genomics Workbench v20, raw reads were filtered by length, trimmed and mapped to the reference genome ( V. cholerae O1 El Tor E7946) to identify single-nucleotide variants. Of the 67 isolates, 66 yielded sufficient coverage (>50×) of the V. cholerae genome. We proceeded with these 66 genomes for further analysis. To identify genes not present in the reference genome, contigs were assembled de novo using CLC Genomics Workbench v20.
Genome-wide association studies (GWAS)
To extract genomic variants capturing all sources of variation in the genome (i.e. single nucleotide variants, indels and gene presence/absence) without a priori assumption about the underlying gene content of each sample (e.g. accessory genes or plasmids), unitigs were generated from the 66 genomes assembled using gatb [46]. Unitigs are sequences of variable length (unlike k-mers of fixed length k) that represent the variations in the population of genomes under study in high resolution. GWAS were performed using linear mixed models implemented in pyseer v.1.3.6 and adjusted for population stratification using the kinship matrix estimated from the phylogenetic tree [47].
To generate the phylogenetic tree, genome alignments consisting entirely of variable nucleotides were produced from whole-genome SNP data generated by CLC Genomics Workbench v20 using VCF-kit 0.1.6 [48]. The tree was then inferred by RaxML under the general time reversible (GTR) model with rate variation across sites following a gamma distribution [49]. We used the linear-mixed model approach to adjust for population stratification and linkage disequilibrium in microbial GWAS [50]. Heritability (h2 ), an estimate of the proportion of the phenotype variance that can be explained by total genomic variation represented in the unitigs, was also calculated using pyseer v.1.3.6. Likelihood-ratio test P values for the association tests were adjusted for multiple-testing by Bonferroni correction (at a genome-wide false discovery rate of 0.05) for the number of unique unitig patterns (i.e. only giving one count to a unitig with an identical presence/absence profile across genomes). We also removed unitigs tagged with the errors ‘bad-chisq’, ‘pre-filtering-failed’, ‘lrt-filtering-failed’, ‘firth-fail’ and ‘matrix-inversionerror’ after the analysis. To further remove false positive GWAS hits, we removed any considerable clusters of unitigs (>20) with identical P values, as these are likely to be lineage-specific markers or markers with strong linkage disequilibrium comprised of mostly non-causal variants linked on the same clonal frame. GWAS hits were annotated by mapping the unitigs to two reference genomes of V. cholerae , namely, E7946 [National Center for Biotechnology Information (NCBI) assembly accession number: GCA_002749635.1] and O1 biotype El Tor strain N16961 (NCBI assembly accession number: GCA_003063785.1) using bwa. Statistically significant GWAS hits were further annotated with the card resistance gene identifier (rgi) after filtering the ‘loose’ hits and hits with identity <0.90.
Antibiotic detection by liquid chromatography (LC) MS/MS
The approach was based on a prior study [51]. Stool supernatants from the primary collection were obtained by centrifugation and filtration (0.2 µM surfactant-free cellulose acetate, Nalgene; Thermo Fisher Scientific). Proteins were precipitated (1 : 7, v/v, water:methanol). Supernatants were diluted with methanol and water (1 : 1, v/v) in 0.1 % formic acid for LC, and 5 µl supernatant was injected for analysis. LC-MS/MS was performed on a 2.1×150 mm Hypersil Gold aQ column (particle size 3 µm) using a high-performance LC system (Thermo UltiMate 3000 series) with an LTQ XL ion trap mass spectrometer (Thermo Fisher Scientific). Mobile phases were 1 % formic acid in water (A) and 1 % formic acid in methanol (B), and held at a constant 5% B for 2 min before ramping to 95 % B at 15 min, where it was held for an additional minute before returning to starting conditions for a total run time of 25 min.
Eluent was ionized using electrospray ionization (ESI) in positive mode at a spray voltage of 5 kV, a nitrogen sheath gas flow rate of 8 l min−1, and capillary temperature of 300 °C. Two scan events were programmed to perform an initial scan from m/z 100 to 1000, which was followed by targeted collision induced dissociation based on a retention time and mass list. Retention time windows ranged from 0.35 to 6.50 min, depending on the elution range of the standards at high and low concentrations. Masses were targeted for the most abundant adduct or ion associated with each antibiotic (typically the [M+H]+ ion) with a m/z 1 window. Data analysis for amoxicillin, sulfamethoxazole/trimethoprim, azithromycin, tetracycline, doxycycline, metronidazole, nalidixic acid and ciprofloxacin was performed manually using extracted ion chromatograms and MS/MS matching with an in-house antibiotic MS/MS library using Xcalibur 2.2 SP 1.48 (Thermo Fisher Scientific).
Statistical analysis
Bivariate analyses of categorical data were assessed using Fisher’s exact test, and continuous data were analysed using the Mann–Whitney U test (alpha=0.05). McNemar’s test was used to analyse paired data (alpha=0.05).
Results
Comparison of AMR under aerobic and anaerobic conditions
We measured baseline growth parameters for each isolate under anaerobic and aerobic conditions and found that carrying capacity, AUC and growth velocity were all significantly lower under anaerobic conditions (Table S2). We tested an assumption that under anaerobic conditions mixed fermentation and anaerobic respiration would occur in LB medium. Analysing a subset of three isolates and the reference strain E7946, we monitored for acidification as a sign of fermentation and assessed the impact of the addition of an alternative electron acceptor (20 mM fumarate) for anaerobic respiration. Under anaerobic conditions, one out of four strains acidified the LB medium to 6.0 (Table S3), suggesting fermentation in at least some isolates. The addition of fumarate as an alternative electron acceptor under anaerobic conditions resulted in a small increase in AUC of 25–27 % (Table S3), suggesting that in LB medium anaerobic respiration likely occurs but is limited for alternative electron acceptors. These results are consistent with mixed fermentation and anaerobic respiration in our experimental conditions.
In this physiological context and using standard antibiotic breakpoints established for aerobic conditions, AMR differed between anaerobic versus aerobic conditions (Fig. 1); distributions of single and multi-agent AMR phenotypes are shown (Fig. 2a). The MIC modes for ciprofloxacin were 8 µg ml−1 (min=0.016 µg ml−1; max=32 µg ml−1) and 2 µg ml−1 (min=0.004 µg ml−1; max=8 µg ml−1) under anaerobic and aerobic conditions, respectively (Table S4); the rates of resistance under anaerobic (93 %; N=62/67) and aerobic (54 %; N=36/67) conditions were significantly different (McNemar’s test P<0.001; Table S5). For azithromycin, the MIC modes were 32 µg ml−1 (min=8 µg ml−1; max=124 µg ml−1) and 4 µg ml−1 (min=1 µg ml−1; max=32 µg ml−1) under anaerobic and aerobic conditions, respectively (Table S4). The rates of resistance under anaerobic (n=67/67; 100 %) and aerobic (n=15/67; 22 %) conditions were significantly different (McNemar’s test P<0.001; Table S5). For doxycycline, the MIC modes were 1 µg ml−1 under both aerobic and anaerobic conditions (Table S4); one isolate was resistant under anaerobic conditions alone, and one isolate was resistant under both anaerobic and aerobic conditions. The odds of classifying isolates as resistant increased over 10 times for ciprofloxacin [odds ratio (OR)=10.5; 95 % confidence interval (CI)=3.61–37.7] and over 200 times for azithromycin (OR=213; 95 % CI=31.9–>5000) under anaerobic compared to aerobic conditions.
Fig. 1.
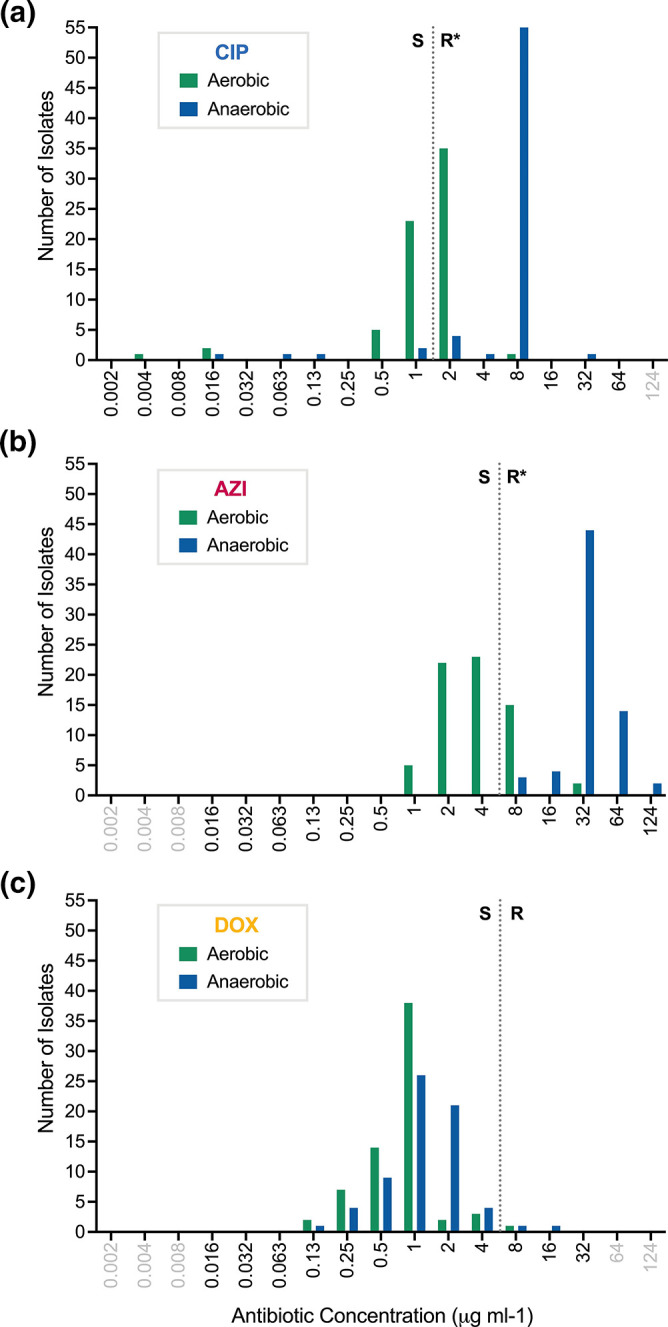
Distribution of MICs under aerobic and anaerobic conditions among clinical isolates from the primary collection. Ciprofloxacin (CIP) (a), azithromycin (AZI) (b) and doxycycline (DOX) (c). Data are from 67 human-shed V. cholerae isolates. The MICs for each isolate under aerobic (green) and anaerobic (blue) conditions were enumerated, and the number of isolates with a given MIC (µg ml−1) are represented as bars; values in grey text represent concentrations not tested (Table S4). Dotted lines are the breakpoints for resistance as per CLSI standards, which are based on assays under aerobic conditions (CIP=2 µg ml−1; AZI=8 µg ml−1; DOX=8 µg ml−1). S, Sensitive; R, resistant. * indicates a significant difference in the frequency of isolates identified as resistant to ciprofloxacin and azithromycin by McNemar’s test (both P<0.001).
Fig. 2.

AMR phenotypes and known AMR genetic elements in human-shed V. cholerae isolates from the primary collection. Pink, yellow and blue colour coding is used to indicate AMR phenotypes and genotypes to azithromycin (AZI), doxycycline (DOX) and ciprofloxacin (CIP), respectively; colours blend when overlapped. Isolates with no resistance (sensitive) are shown in white circles. (a) Proportional Venn diagram (Euler) of AMR phenotypes to AZI, DOX and/or CIP under aerobic (left) and anaerobic conditions (right). Counts indicate the number of isolates with the corresponding phenotype. (b) AMR genotypes from known resistance genes in whole-genome sequences. The distribution of known AMR genetic elements is shown by proportional Venn diagram (Euler; left) and bar chart (right). On the right, the x-axis of the bar chart depicts the presence (black points) of known AMR genes [mphA, gyrA, parC, tet(59), qnrVc ] in a given genome and the y-axis depicts the number of isolates that share the given combination of AMR genes. Coloured bars to the left indicate the number of isolate genomes encoding resistance genes to CIP (blue), AZI (pink) or DOX (yellow). AMR genetic elements to other antibiotics are not shown.
To evaluate the generalizability of these findings from the primary sample collection, we also compared aerobic and anaerobic growth curves of 277 isolates from the secondary sample collection. For ciprofloxacin, the rates of resistance were significantly different under anaerobic conditions (21 %; n=58/277) compared to aerobic conditions (1.1 %; n=3/277; McNemar’s test P<0.001; Table S6). For azithromycin, the rates of resistance were significantly different under anaerobic conditions (100 %; N=277/277) compared to aerobic conditions (57 %; N=159/277; McNemar’s test P<0.001; Table S6). For doxycycline, only two isolates were resistant under anaerobic conditions alone and one under both anaerobic and aerobic conditions (Table S6). The odds of classifying isolates as resistant increased over 25 times for ciprofloxacin (OR=25.7; 95 % CI=18.8–34.6) and 119 times for azithromycin (OR=119; 95 % CI=20.95–4739) under anaerobic compared to aerobic conditions.
Addition of catalase to test whether ROS affect antibiotic resistance/sensitivity under aerobic conditions
In this experiment, catalase was added to the media to quench hydrogen peroxide with the objective of testing the hypothesis that susceptibility under aerobic conditions was associated with ROS (e.g. hydrogen peroxide). For ciprofloxacin, the MICs for the sensitive reference strain E7946 and the resistant clinical isolate EN160 remained unchanged when catalase was added to the media under aerobic conditions. The addition of catalase was not associated with differences in AUCs for both E7946 and EN160 in media containing ciprofloxacin, azithromycin or doxycycline at twofold below the MIC. The AUCs in LB medium with and without catalase for E7946 and EN160 were not statistically different (Table S7).
Molecular AMR correlates under aerobic and anaerobic conditions
The distribution of known AMR genetic elements is shown (Fig. 2b). AMR-associated point mutations (likely transmitted vertically, not on an established mobilizable element), and genes on known horizontally transferred mobilizable elements, are provided (Supplementary Material). The ICE SXT/R391 was found in 90 % (60/67) of isolates. The ICEs contained the pentapeptide repeat protein that confers fluoroquinolone resistance (qnrVc ), the macrolide-inactivating phosphotransferase (mphA) and the major facilitator superfamily (MFS) efflux pump conferring tetracycline resistance [tet(59)] [52–55]. The genes qnrVc , mphA and tet(59) were found in 79 % (52/66), 33 % (22/66) and 79 % (52/66) of isolates, respectively. Ciprofloxacin resistance under both anaerobic and aerobic conditions was significantly associated with qnrVC , gyrA and parC (Table S8). Identification of the known azithromycin AMR gene mphA was significantly associated with resistance under aerobic conditions alone (P<0.001). The gene tet(59) was not associated with doxycycline resistance under aerobic or aerobic conditions (both P=0.566).
We next used GWAS to comprehensively explore the genetic basis of AMR. This approach used the phenotype of AUC to represent growth with or without exposure to the three test antibiotics at five concentrations under either aerobic or anaerobic conditions. Growth phenotypes (analysed by AUCs) at similar antibiotic concentrations were positively correlated within aerobic and anaerobic conditions for all three antibiotics (Fig. 3). Growth phenotypes were also positively correlated between aerobic and anaerobic conditions for ciprofloxacin (Fig. 3a). However, phenotypes were weakly, or even negatively, correlated between aerobic and anaerobic conditions for azithromycin and doxycycline (Fig. 3b,c). These results support the hypothesis that anaerobic and aerobic growth under antibiotic pressure can differ and be distinct to antibiotic type.
Fig. 3.

Correlation analysis of growth phenotypes at different concentrations of antibiotics under aerobic and anaerobic conditions among V. cholerae clinical isolates from the primary collection. Antibiotic exposures were to ciprofloxacin (a), azithromycin (b) and doxycycline (c). AUC was analysed as the growth parameter. Aerobic/anaerobic conditions are labelled horizontally and vertically with the antibiotic concentration in µg ml−1 (e.g. anaerobic-0.06). Analyses are grouped: aerobic versus aerobic, red boxes; anaerobic versus aerobic, purple boxes; anaerobic versus anaerobic, blue boxes. Heatmaps show correlation coefficients (the keys are on the right) for similar (yellow) versus dissimilar (purple) growth at two given conditions.
The heritability of the AMR phenotypes (AUCs) was estimated prior to the GWAS. Heritability (h2 ) is defined as the proportion of phenotypic variation explained by genetic variation, measured as unique contiguous tracts of the assembled genomes (unitigs) that tag both single nucleotide variants, indels and gene content changes (see Methods). We found relatively high heritability (h2 in the range 0.60–0.99) of growth across concentrations of ciprofloxacin under both aerobic and anaerobic conditions, yielding statistically significant GWAS hits (Table 1; Data Files S1 and S2): 20 under aerobic conditions and 16 under anaerobic conditions. In contrast, heritability tended to be much lower under aerobic compared to anaerobic conditions for both azithromycin and doxycycline, yielding significant GWAS hits only under anaerobic conditions (Table 1; Data Files S3 and S4, respectively): 3 for azithromycin under anaerobic conditions alone and 57 for doxycycline under anaerobic conditions alone.
Table 1.
Identification of genetic elements by GWAS that associate with AMR
|
Condition |
Outcome* |
Antibiotic concn (µg ml−1) |
||||
|---|---|---|---|---|---|---|
|
Ciprofloxacin |
CIP 4 |
CIP 1 |
CIP 0.5 |
CIP 0.25 |
CIP 0.13 |
|
|
Aerobic |
Heritability (h2 ) |
0.99 |
0.73 |
0.60 |
0.74 |
0.92 |
|
Associated genes |
0 |
11 |
9 |
2 |
6 |
|
|
Anaerobic |
Heritability (h2 ) |
0.81 |
0.73 |
0.72 |
0.72 |
0.87 |
|
Associated genes |
8 |
8 |
8 |
10 |
4 |
|
|
Azithromycin |
AZI 16 |
AZI 8 |
AZI 4 |
AZI 2 |
AZI 1 |
|
|
Aerobic |
Heritability (h2 ) |
– |
0 |
0.02 |
0 |
0.11 |
|
Associated genes |
– |
– |
– |
– |
– |
|
|
Anaerobic |
Heritability (h2 ) |
0.30 |
0.77 |
0.66 |
0.38 |
0 |
|
Associated genes |
2 |
1 |
0 |
0 |
– |
|
Doxycycline |
DOX 1 |
DOX 0.5 |
DOX 0.25 |
DOX 0.13 |
DOX 0.06 |
|
|
Aerobic |
Heritability (h2 ) |
0 |
0.27 |
0.23 |
0.30 |
0.50 |
|
Associated genes |
– |
0 |
0 |
0 |
0 |
|
|
Anaerobic |
Heritability (h2 ) |
0.10 |
0.61 |
0.72 |
0.72 |
0.77 |
|
Associated genes |
0 |
23 |
55 |
54 |
17 |
AZI, Azithromycin; CIP, ciprofloxacin; DOX, doxycycline.
*Heritability is the proportion of phenotypic variation that is explained by genetic variation. Associated genes are all significant GWAS hits after correction for multiple hypothesis testing (P<0.05 after Bonferroni correction).
AMR genes identified by GWAS were diverse (Fig. 4; Data Files S1–S4). These candidates included known AMR genes, such as qnrVc and dfrA, which were associated with ciprofloxacin resistance under both aerobic and anaerobic conditions. We identified seven genes associated with ciprofloxacin resistance under anaerobic conditions alone (including the stress response gene barA and a radC homologue involved in DNA repair; Data File S2), and ten genes under aerobic conditions alone (including rtxB; Data File S1). Under anaerobic conditions, most genes were identified at ciprofloxacin concentrations at, or above, 0.25 µg ml−1; however, four genes, including barA, were identified under one of the lowest tested ciprofloxacin concentrations (0.13 µg ml−1; Data File S2). GWAS hits for azithromycin and doxycycline resistance were found only under anaerobic conditions. For azithromycin, two genetic elements were identified: mphA, and a region between ompT and dinG (ompT-dinG; Data File S3). For doxycycline, 23 genes were shared across concentrations; however, the gene discovery rate was higher at the lower concentrations (n=54 at 0.13 µg ml−1; n=27 at 0.06 µg ml−1; Data File S4). GWAS hits included the major facilitator superfamily antibiotic efflux pump tet(59) (Fig. 4, Data Files S1–S4). Most genetic elements identified have unknown function. The identification of known AMR genes by GWAS serves as positive control and suggests that the genes of unknown function may indeed play a role in AMR.
Fig. 4.

Distribution of AMR genes associated with AMR growth phenotypes at different concentrations of antibiotics under aerobic and anaerobic conditions among isolates from the primary collection. Venn diagrams show the overlap between genes associated with (a) ciprofloxacin resistance under aerobic versus anaerobic conditions, (b) ciprofloxacin at different concentrations (µg ml−1) under aerobic conditions, (c) ciprofloxacin at different concentrations (µg ml−1) under anaerobic conditions, and (d) doxycycline at different concentrations (µg ml−1) under anaerobic conditions. Genes shown in boxes had statistically significant associations.
Antibiotics detected in stool by LC-MS/MS
Finally, we sought to test the hypothesis that AMR genotypes and phenotypes would be associated with measured concentrations of antibiotics in stool. A combined total of 196 antibiotics were detected in the 51 stool supernatants tested by LC-MS/MS using a targeted technique for nine common antibiotics (Fig. 5). At least one antibiotic was detected in 98 % (n=50/51), at least two antibiotics were detected in 94 % (n=48/51), and three or more antibiotics were detected in 90 % (n=46/51) of stool supernatants (Fig. 5). Antibiotics detected were ciprofloxacin (n=48/51; 94 %), tetracycline/doxycycline (n=46/51; 90 %), nalidixic acid (n=41/51; 80 %), metronidazole (n=37/51; 73 %), sulfamethoxazole/trimethoprim (n=22/51; 43 %) and amoxicillin (n=2/51; 4 %); azithromycin was not detected. Detection of quinolone/fluoroquinolone and tetracycline/doxycycline in stool by LC-MS/MS was not associated with AMR genotypes nor phenotypes (Table S9). Associations for azithromycin could not be tested because azithromycin was not detected in any stool supernatant.
Fig. 5.

Antibiotic detection in stool supernatants by LC-MS/MS among cholera samples from the primary collection. Green with +, detected; white with −, not detected. CIP, Ciprofloxacin; TET/DOX, tetracycline and/or doxycycline; NAL, nalidixic acid; MET, metronidazole; BAC, sulfamethoxazole and/or trimethoprim; AMO, amoxicillin; ERY, erythromycin; CEF, ceftriaxone. Stool supernatants were not available for EN80, 86, 88, 92, 100, 103, 109, 116–120, 126, 124, 130 and 131.
Discussion
In this study, V. cholerae isolates from cholera patients were found to be more resistant to antibiotic exposure under anaerobic conditions compared to aerobic conditions (Fig. 1). This phenotype differed by antibiotic class. Novel genetic elements were found to associate with AMR under anaerobic conditions, which also differed by antibiotic class (Fig. 4). The approach of using a continuous variable (AUC) for the AMR phenotype within the framework of GWAS may provide a new approach to identify putative AMR genetic targets for future mechanistic research.
CLSI breakpoints for enteropathogens like V. cholerae were developed under aerobic conditions [45, 56]. CLSI and other clinical reference bodies set breakpoints to have clinical relevance despite limited data from clinical studies [56–58]. In this context, the odds of classifying isolates in the primary collection as resistant under anaerobic conditions compared to aerobic conditions increased over 10 times for ciprofloxacin and over 200 times for azithromycin. These results are likely generalisable across V. cholerae because in the secondary collection, which is separated by more than 10 years, we found that the odds of classifying isolates as resistant under anaerobic conditions compared to aerobic conditions increased over 20 times for ciprofloxacin and over 100 times for azithromycin.
There are several physiological explanations for increased antibiotic resistance under anaerobic conditions. ROS induce both intracellular and cell-wall stress, and are at higher concentrations under aerobic conditions [59]. ROS may have acted synergistically to potentiate antibiotic lethality [31, 60]. The assays that utilized catalase to quench hydrogen peroxide under aerobic conditions were conducted to evaluate this possibility. The MICs for all three antibiotics did not increase with the addition of catalase suggesting that the reduction of ROS alone cannot account for increased MICs observed under anaerobic conditions (Table S7). We hypothesize that reduced growth under anaerobic conditions might decrease the effectiveness of antimicrobial agents that directly or indirectly disrupt cell envelope integrity [61]. While reduced growth was observed under anaerobic conditions, the antibiotics tested are not known to directly disrupt the cell envelope. However, off-target effects (e.g. cell envelope stress) by ciprofloxacin, azithromycin and doxycycline may have occurred.
To further investigate AMR phenotypes under anaerobic conditions, future studies will benefit from the use of a defined medium (such as M9) where the carbon source and an alternative electron acceptor (e.g. fumarate, nitrate, DMSO or trimethylamine N-oxide) can be supplemented to strictly control anaerobic respiration versus fermentation. Buffering the medium (e.g. with PBS or bicarbonate) to a neutral pH is important because V. cholerae can acidify the medium over time, with toxic effects. In prior research, acidification occurred after 10 h [9]. In our shorter 8 h assay, acidification was observed under anaerobic conditions among one of four strains tested (Table S3). Furthermore, buffering to the alkaline pH found in cholera stool (pH 8.5–9) will provide important insight given that V. cholerae utilizes nitrate for anaerobic respiration only at alkaline pH [2, 9]. Here, we used an undefined medium (LB) without a defined, saturating concentration of an alternative electron acceptor. This approach likely resulted in mixed anaerobic respiration and fermentation. To address these limitations in future studies, robotic automation and 384-well formatted assays would enable a scalable system for multiple defined media across broad gradients of antibiotics. Ideally, we would simulate the conditions of the human gut, but in practice these conditions can only be approximated.
There are many knowledge gaps on the genetic basis of AMR phenotypes under diverse environmental conditions. This study prioritized the factor of oxygen exposure as a determinant of AMR phenotypes, because facultative anaerobic enteropathogens experience hypoxia and anoxia in the animal gut [9]. The first phase of the analysis focused on previously known AMR genotypes with known AMR phenotypes. For ciprofloxacin, mutations in parC and carriage of qnrVc were significantly associated with phenotypic resistance under aerobic and anaerobic conditions; mutations in gyrA were significantly associated with resistance under anaerobic conditions alone (Table S8). For azithromycin, mphA was identified and significantly associated with AMR under aerobic conditions alone. While tet(59) was identified, very few isolates were identified as resistant to doxycycline under aerobic (n=1) or anaerobic conditions (n=2). These associative data begin to reveal a difference between AMR genotypes and phenotypes under aerobic and anaerobic conditions.
The second phase of analysis sought to use GWAS to identify previously unknown genetic targets associated with AMR. The continuous variable of AUC, as opposed to the binary variable of growth/no growth, was used in assays with and without antibiotic exposure under aerobic and anaerobic conditions. Antibiotics at the breakpoint and sub-breakpoint concentrations were chosen based on a rationale that different genetic elements might contribute differently to AMR phenotypes at different antibiotic concentrations. As expected, GWAS identified qnrVc for ciprofloxacin exposure under both aerobic and anaerobic conditions (Fig. 4). GWAS identified mphA for azithromycin exposure under anaerobic conditions alone, and tet(59) for doxycycline exposure under anaerobic conditions alone. These results of known AMR genes served as positive controls for the GWAS. We also note that genes known to be important for anaerobic respiration (e.g. tatA1, tatC, ccmA–F, ccmH, moaA–D, moeA–B, napA–D, napF, fnr, narP–Q, nqrF, dsbA, dsbD, hemN) in V. cholerae were not identified as GWAS hits [9]. This suggests that our GWAS was specific to AMR phenotypes and was less liable to detect genes related to anaerobic conditions alone. Therefore, we expect novel GWAS hits to be likely candidates for involvement in AMR phenotypes.
For ciprofloxacin, seven genes were associated with AMR in anaerobic conditions alone; these included a gene involved in DNA repair (radC), a two-component histidine kinase involved in stress response (barA) and an ATP-dependent zinc protease. The gene dfrA31 encodes a trimethoprim-resistant dihydrofolate reductase and APH(6)-Id encodes a streptomycin phosphotransferase enzyme; both genes were identified for aerobic and anaerobic conditions. These two genes are located on the SXT element along with qnrVc and, therefore, may be associated due to genetic linkage rather than due to causal roles in ciprofloxacin resistance. For azithromycin, one additional genetic element under anaerobic conditions was discovered to associate with AMR: an intergenic region between ompT (porin; known to be associated with AMR) [62, 63] and dinG (ATP-dependent DNA helicase). For doxycycline, a diverse set of 57 genetic elements under anaerobic conditions alone were discovered to associate with AMR. These include vexK (efflux RND transporter permease associated with AMR) [64–66] and zorA (anti-phage defence system ZorAB subunit A; a putative proton channel that may respond to membrane perturbation by depolarization) [67].
In addition to the SXT element, genes associated with an AMR phenotype were also discovered on the Vibrio Pathogenicity Island II (VSPII; N16961 VC0506–VC0512/E7946 loci RS02705–RS02745); these loci are genetically diverse [68]. The GWAS hits in VSPII encode both biofilm/auto-aggregation associated factors, as well as an aerotaxis protein (AerB; VC0512) [69]; findings consistent with roles in AMR and aerobic/anaerobic conditions. These GWAS analyses were of limited power due to the modest sample size, and could be sensitive to false positives at AMR ‘hot-spots’ like SXT. Despite these limitations, GWAS enabled the discovery of an intriguing list of genetic targets that were associated with AMR and require future mechanistic molecular analysis to test for causal relationships.
LC-MS/MS analysis on the stools from the primary collection, stools from which the isolates were obtained, was conducted to test the hypothesis that the rates of AMR genotypes and phenotypes were higher when the stool samples contained the cognate antibiotic. Nearly all patients shed at least one antibiotic, making it difficult to identify AMR correlates to exposure (Fig. 5). This finding is important because studies that leverage natural infection to set clinically meaningful AMR breakpoints under aerobic conditions, and now anaerobic conditions, cannot readily be performed because of the degree of antibiotic exposure among diarrheal patients. Therefore, future interventional clinical studies with known antibiotic exposure determined a priori may be required. Given that the primary collection is from patients that self-reported not taking antibiotics, the detection of a combined total of 196 antibiotics further highlights the ubiquity of antibiotics and the limited value of self-reported antibiotic exposure.
Conclusions
Facultative enteropathogens are exposed to antibiotics under aerobic and anaerobic conditions in both the human gut and in the environment. We used the facultative anaerobic enteropathogen V. cholerae as a model to test for differences in AMR phenotypes under aerobic and anaerobic conditions. Increased resistance was found under anaerobic conditions compared to aerobic conditions. Using AMR breakpoints established for aerobic conditions, the odds of classifying isolates as resistant under anaerobic compared to aerobic conditions increased over 10 times for two of the three antibiotics tested. While several known resistance genes were associated with AMR under both conditions, many genes were only associated with AMR under one condition. Heritability tended to be higher, and more genes associated with resistance, under anaerobic conditions. This suggests that key genetic determinants of resistance may be missed when experiments are only performed aerobically. Our findings provide a rationale to determine whether increased MICs under anaerobic conditions are associated with therapeutic failures and/or microbial escape in cholera patients and, if true, there may be a need to determine AMR breakpoints for anaerobic conditions.
Supplementary Data
Funding information
This work was supported by National Institutes of Health grants to E.J.N. [DP5OD019893, R21TW010182] and K.B.B. [S10 OD021758-01A1], and internal support from the Emerging Pathogens Institute, and the Department of Pediatrics and the Department of Environmental and Global Health at the University of Florida. B.J.S. and M.M.S. were supported by a Genome Canada and Genome Quebec Bioinformatics and Computational Biology grant. A.C.-M. was supported in part by a grant from the Children’s Miracle Network (Florida). The funders had no role in study design, data collection and analysis, decision to publish nor preparation of the manuscript.
Acknowledgements
We thank the patients for participating in this study, and the icddr,b clinical and laboratory teams that collected the samples. We are grateful to Randy Autrey and Krista Berquist for their administrative expertise, as well as Glenn Morris at the Emerging Pathogens Institute and Desmond Schatz in the Department of Pediatrics at the University of Florida for their ongoing support. Stephen Calderwood, Jason Harris and Regina LaRocque were the principal investigators (PIs) of the parent study/IRB protocol (Massachusetts General Hospital, Harvard University School of Medicine, USA) under which the samples were collected by E.J.N. when he was a National Institutes of Health Fogarty Fellow. Andrew Camilli provided additional reagents and insights for this manuscript; F.Q. was the PI in Bangladesh, and PI of the ethical review committee (ERC)/research review committee (RRC) approvals at the icddr,b. This generous research infrastructure and support was invaluable to the success of this study.
Conflicts of interest
The authors declare that there are no conflicts of interest
Footnotes
Abbreviations: AMR, antimicrobial resistance; AUC, area under the curve; CI, confidence interval; CLSI, Clinical and Laboratory Standards Institute; GWAS, genome-wide association studies; icddr,b, International Centre for Diarrhoeal Disease Research, Bangladesh; ICE, integrative and conjugative element; LC, liquid chromatography; MIC, minimum inhibitory concentration; OR, odds ratio; ROS, reactive oxygen species.
All supporting data, code and protocols have been provided within the article or through supplementary data files. Nine supplementary tables are available with the online version of this article.
References
- 1.CLSI Performance Standards for Antimicrobial Susceptibility Testing, 27th edn. Wayne, PA: Clinical and Laboratory Standards Institute; 2017. [Google Scholar]
- 2.Bueno E, Sit B, Waldor MK, Cava F. Genetic dissection of the fermentative and respiratory contributions supporting Vibrio cholerae hypoxic growth. J Bacteriol. 2020;202:e00243-20. doi: 10.1128/JB.00243-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bueno E, Pinedo V, Cava F. Adaptation of Vibrio cholerae to hypoxic environments. Front Microbiol. 2020;11:739. doi: 10.3389/fmicb.2020.00739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Xu Q, Dziejman M, Mekalanos JJ. Determination of the transcriptome of Vibrio cholerae during intraintestinal growth and midexponential phase in vitro . Proc Natl Acad Sci USA. 2003;100:1286–1291. doi: 10.1073/pnas.0337479100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Mandlik A, Livny J, Robins WP, Ritchie JM, Mekalanos JJ, et al. RNA-Seq-based monitoring of infection-linked changes in Vibrio cholerae gene expression. Cell Host Microbe. 2011;10:165–174. doi: 10.1016/j.chom.2011.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Beaber JW, Hochhut B, Waldor MK. SOS response promotes horizontal dissemination of antibiotic resistance genes. Nature. 2004;427:72–74. doi: 10.1038/nature02241. [DOI] [PubMed] [Google Scholar]
- 7.Narendrakumar L, Gupta SS, Johnson JB, Ramamurthy T, Thomas S. Molecular adaptations and antibiotic resistance in Vibrio cholerae: a communal challenge. Microb Drug Resist. 2019;25:1012–1022. doi: 10.1089/mdr.2018.0354. [DOI] [PubMed] [Google Scholar]
- 8.Das B, Verma J, Kumar P, Ghosh A, Ramamurthy T. Antibiotic resistance in Vibrio cholerae: understanding the ecology of resistance genes and mechanisms. Vaccine. 2020;38 (Suppl. 1):A83–A92. doi: 10.1016/j.vaccine.2019.06.031. [DOI] [PubMed] [Google Scholar]
- 9.Bueno E, Sit B, Waldor MK, Cava F. Anaerobic nitrate reduction divergently governs population expansion of the enteropathogen Vibrio cholerae . Nat Microbiol. 2018;3:1346–1353. doi: 10.1038/s41564-018-0253-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Nelson EJ, Nelson DS, Salam MA, Sack DA. Antibiotics for both moderate and severe cholera. N Engl J Med. 2011;364:5–7. doi: 10.1056/NEJMp1013771. [DOI] [PubMed] [Google Scholar]
- 11.WHO The Treatment of Diarrhoea – a Manual for Physicians and Other Senior Health Workers, 4th rev. Geneva: World Health Organization; 2005. [Google Scholar]
- 12.Leibovici-Weissman Y, Neuberger A, Bitterman R, Sinclair D, Salam MA, et al. Antimicrobial drugs for treating cholera. Cochrane Database Syst Rev. 2014;2014:CD008625. doi: 10.1002/14651858.CD008625.pub2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Khan AI, Mack JA, Salimuzzaman M, Zion MI, Sujon H, et al. Electronic decision support and diarrhoeal disease guideline adherence (mHDM): a cluster randomized controlled trial. Lancet Digit Health. 2020;2:e250–258. doi: 10.1016/S2589-7500(20)30062-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Biswas D, Hossin R, Rahman M, Bardosh KL, Watt MH, et al. An ethnographic exploration of diarrheal disease management in public hospitals in Bangladesh: from problems to solutions. Soc Sci Med. 2020;260:113185. doi: 10.1016/j.socscimed.2020.113185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ingle DJ, Levine MM, Kotloff KL, Holt KE, Robins-Browne RM. Dynamics of antimicrobial resistance in intestinal Escherichia coli from children in community settings in South Asia and sub-Saharan Africa. Nat Microbiol. 2018;3:1063–1073. doi: 10.1038/s41564-018-0217-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Towner KJ, Pearson NJ, Mhalu FS, O’Grady F. Resistance to antimicrobial agents of Vibrio cholerae E1 Tor strains isolated during the fourth cholera epidemic in the United Republic of Tanzania. Bull World Health Organ. 1980;58:747–751. [PMC free article] [PubMed] [Google Scholar]
- 17.Burrus V, Marrero J, Waldor MK. The current ICE age: biology and evolution of SXT-related integrating conjugative elements. Plasmid. 2006;55:173–183. doi: 10.1016/j.plasmid.2006.01.001. [DOI] [PubMed] [Google Scholar]
- 18.Hooper DC, Jacoby GA. Topoisomerase inhibitors: fluoroquinolone mechanisms of action and resistance. Cold Spring Harb Perspect Med. 2016;6:a025320. doi: 10.1101/cshperspect.a025320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Garriss G, Waldor MK, Burrus V. Mobile antibiotic resistance encoding elements promote their own diversity. PLoS Genet. 2009;5:e1000775. doi: 10.1371/journal.pgen.1000775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Fonseca EL, Dos Santos Freitas F, Vieira VV, Vicente ACP. New qnr gene cassettes associated with superintegron repeats in Vibrio cholerae O1. Emerg Infect Dis. 2008;14:1129–1131. doi: 10.3201/eid1407.080132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Fyfe C, Grossman TH, Kerstein K, Sutcliffe J. Resistance to macrolide antibiotics in public health pathogens. Cold Spring Harb Perspect Med. 2016;6:a025395. doi: 10.1101/cshperspect.a025395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Grossman TH. Tetracycline antibiotics and resistance. Cold Spring Harb Perspect Med. 2016;6:a025387. doi: 10.1101/cshperspect.a025387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zhu Z, Surujon D, Ortiz-Marquez JC, Huo W, Isberg RR, et al. Entropy of a bacterial stress response is a generalizable predictor for fitness and antibiotic sensitivity. Nat Commun. 2020;11:4365. doi: 10.1038/s41467-020-18134-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wood S, Zhu K, Surujon D, Rosconi F, Ortiz-Marquez JC, et al. In: The Pangenome: Diversity, Dynamics and Evolution of Genomes. Tettelin H, Medini D, editors. Cham: Springer; 2020. A pangenomic perspective on the emergence, maintenance, and predictability of antibiotic resistance; pp. 169–202. [PubMed] [Google Scholar]
- 25.Warrier I, Ram-Mohan N, Zhu Z, Hazery A, Echlin H, et al. The transcriptional landscape of Streptococcus pneumoniae TIGR4 reveals a complex operon architecture and abundant riboregulation critical for growth and virulence. PLoS Pathog. 2018;14:e1007461. doi: 10.1371/journal.ppat.1007461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Jensen PA, Zhu Z, van Opijnen T. Antibiotics disrupt coordination between transcriptional and phenotypic stress responses in pathogenic bacteria. Cell Rep. 2017;20:1705–1716. doi: 10.1016/j.celrep.2017.07.062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.van Opijnen T, Dedrick S, Bento J. Strain dependent genetic networks for antibiotic-sensitivity in a bacterial pathogen with a large pan-genome. PLoS Pathog. 2016;12:e1005869. doi: 10.1371/journal.ppat.1005869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Cain AK, Barquist L, Goodman AL, Paulsen IT, Parkhill J, et al. A decade of advances in transposon-insertion sequencing. Nat Rev Genet. 2020;21:526–540. doi: 10.1038/s41576-020-0244-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Dörr T, Delgado F, Umans BD, Gerding MA, Davis BM, et al. A transposon screen identifies genetic determinants of Vibrio cholerae resistance to high-molecular-weight antibiotics. Antimicrob Agents Chemother. 2016;60:4757–4763. doi: 10.1128/AAC.00576-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kohanski MA, Dwyer DJ, Hayete B, Lawrence CA, Collins JJ. A common mechanism of cellular death induced by bactericidal antibiotics. Cell. 2007;130:797–810. doi: 10.1016/j.cell.2007.06.049. [DOI] [PubMed] [Google Scholar]
- 31.Dwyer DJ, Belenky PA, Yang JH, MacDonald IC, Martell JD, et al. Antibiotics induce redox-related physiological alterations as part of their lethality. Proc Natl Acad Sci USA. 2014;111:E2100–E2109. doi: 10.1073/pnas.1401876111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Staerck C, Gastebois A, Vandeputte P, Calenda A, Larcher G, et al. Microbial antioxidant defense enzymes. Microb Pathog. 2017;110:56–65. doi: 10.1016/j.micpath.2017.06.015. [DOI] [PubMed] [Google Scholar]
- 33.Smirnova G, Muzyka N, Oktyabrsky O. Transmembrane glutathione cycling in growing Escherichia coli cells. Microbiol Res. 2012;167:166–172. doi: 10.1016/j.micres.2011.05.005. [DOI] [PubMed] [Google Scholar]
- 34.Bryan LE, Kwan S. Mechanisms of aminoglycoside resistance of anaerobic bacteria and facultative bacteria grown anaerobically. J Antimicrob Chemother. 1981;8 (Suppl. D):1–8. doi: 10.1093/jac/8.suppl_d.1. [DOI] [PubMed] [Google Scholar]
- 35.Bryan LE. General mechanisms of resistance to antibiotics. J Antimicrob Chemother. 1988;22 (Suppl. A):1–15. doi: 10.1093/jac/22.supplement_a.1. [DOI] [PubMed] [Google Scholar]
- 36.Bryant RE, Fox K, Oh G, Morthland VH. Beta-lactam enhancement of aminoglycoside activity under conditions of reduced pH and oxygen tension that may exist in infected tissues. J Infect Dis. 1992;165:676–682. doi: 10.1093/infdis/165.4.676. [DOI] [PubMed] [Google Scholar]
- 37.Dwyer DJ, Kohanski MA, Hayete B, Collins JJ. Gyrase inhibitors induce an oxidative damage cellular death pathway in Escherichia coli . Mol Syst Biol. 2007;3:91. doi: 10.1038/msb4100135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hong Y, Li Q, Gao Q, Xie J, Huang H, et al. Reactive oxygen species play a dominant role in all pathways of rapid quinolone-mediated killing. J Antimicrob Chemother. 2020;75:576–585. doi: 10.1093/jac/dkz485. [DOI] [PubMed] [Google Scholar]
- 39.Luan G, Hong Y, Drlica K, Zhao X. Suppression of reactive oxygen species accumulation accounts for paradoxical bacterial survival at high quinolone concentration. Antimicrob Agents Chemother. 2018;62:e01622-17. doi: 10.1128/AAC.01622-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zhao X, Drlica K. Reactive oxygen species and the bacterial response to lethal stress. Curr Opin Microbiol. 2014;21:1–6. doi: 10.1016/j.mib.2014.06.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Van Alst AJ, Demey LM, DiRita VJ. Vibrio cholerae requires oxidative respiration through the bd-I and cbb(3) oxidases for intestinal proliferation. PLoS Pathog. 2022;18:e1010102. doi: 10.1371/journal.ppat.1010102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Van Alst AJ, DiRita VJ. Aerobic metabolism in Vibrio cholerae is required for population expansion during infection. mBio. 2020;11:e01989-20. doi: 10.1128/mBio.01989-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Nelson EJ, Chowdhury A, Harris JB, Begum YA, Chowdhury F, et al. Complexity of rice-water stool from patients with Vibrio cholerae plays a role in the transmission of infectious diarrhea. Proc Natl Acad Sci USA. 2007;104:19091–19096. doi: 10.1073/pnas.0706352104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Sprouffske K, Wagner A. Growthcurver: an R package for obtaining interpretable metrics from microbial growth curves. BMC Bioinformatics. 2016;17:172. doi: 10.1186/s12859-016-1016-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.CLSI Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria, M45, 3rd edn. Wayne, PA: Clinical and Laboratory Standards Institute; 2017. [Google Scholar]
- 46.Drezen E, Rizk G, Chikhi R, Deltel C, Lemaitre C, et al. GATB: genome assembly & analysis tool box. Bioinformatics. 2014;30:2959–2961. doi: 10.1093/bioinformatics/btu406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Lees JA, Galardini M, Bentley SD, Weiser JN, Corander J. pyseer: a comprehensive tool for microbial pangenome-wide association studies. Bioinformatics. 2018;34:4310–4312. doi: 10.1093/bioinformatics/bty539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Cook DE, Andersen EC. VCF-kit: assorted utilities for the variant call format. Bioinformatics. 2017;33:1581–1582. doi: 10.1093/bioinformatics/btx011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Stamatakis A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 2014;30:1312–1313. doi: 10.1093/bioinformatics/btu033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Saber MM, Shapiro BJ. Benchmarking bacterial genome-wide association study methods using simulated genomes and phenotypes. Microb Genom. 2020;6:000337. doi: 10.1099/mgen.0.000337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Alexandrova L, Haque F, Rodriguez P, Marrazzo AC, Grembi JA, et al. Identification of widespread antibiotic exposure in patients with cholera correlates with clinically relevant microbiota changes. J Infect Dis. 2019;220:1655–1666. doi: 10.1093/infdis/jiz299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Leclercq SO, Wang C, Zhu Y, Wu H, Du X, et al. Diversity of the tetracycline mobilome within a Chinese pig manure sample. Appl Environ Microbiol. 2016;82:6454–6462. doi: 10.1128/AEM.01754-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Grossman TH, Starosta AL, Fyfe C, O’Brien W, Rothstein DM, et al. Target- and resistance-based mechanistic studies with TP-434, a novel fluorocycline antibiotic. Antimicrob Agents Chemother. 2012;56:2559–2564. doi: 10.1128/AAC.06187-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Noguchi N, Emura A, Matsuyama H, O’Hara K, Sasatsu M, et al. Nucleotide sequence and characterization of erythromycin resistance determinant that encodes macrolide 2’-phosphotransferase I in Escherichia coli . Antimicrob Agents Chemother. 1995;39:2359–2363. doi: 10.1128/AAC.39.10.2359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Chesneau O, Tsvetkova K, Courvalin P. Resistance phenotypes conferred by macrolide phosphotransferases. FEMS Microbiol Lett. 2007;269:317–322. doi: 10.1111/j.1574-6968.2007.00643.x. [DOI] [PubMed] [Google Scholar]
- 56.Humphries RM, Ambler J, Mitchell SL, Castanheira M, Dingle T, et al. CLSI methods development and standardization working group best practices for evaluation of antimicrobial susceptibility tests. J Clin Microbiol. 2018;56:e01934-17. doi: 10.1128/JCM.01934-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Humphries RM, Abbott AN, Hindler JA. Understanding and addressing CLSI breakpoint revisions: a primer for clinical laboratories. J Clin Microbiol. 2019;57:e00203-19. doi: 10.1128/JCM.00203-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ellington MJ, Ekelund O, Aarestrup FM, Canton R, Doumith M, et al. The role of whole genome sequencing in antimicrobial susceptibility testing of bacteria: report from the EUCAST subcommittee. Clin Microbiol Infect. 2017;23:2–22. doi: 10.1016/j.cmi.2016.11.012. [DOI] [PubMed] [Google Scholar]
- 59.Stokes JM, Lopatkin AJ, Lobritz MA, Collins JJ. Bacterial metabolism and antibiotic efficacy. Cell Metab. 2019;30:251–259. doi: 10.1016/j.cmet.2019.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Wang X, Zhao X, Malik M, Drlica K. Contribution of reactive oxygen species to pathways of quinolone-mediated bacterial cell death. J Antimicrob Chemother. 2010;65:520–524. doi: 10.1093/jac/dkp486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kan B, Habibi H, Schmid M, Liang W, Wang R, et al. Proteome comparison of Vibrio cholerae cultured in aerobic and anaerobic conditions. Proteomics. 2004;4:3061–3067. doi: 10.1002/pmic.200400944. [DOI] [PubMed] [Google Scholar]
- 62.Mathur J, Waldor MK. The Vibrio cholerae ToxR-regulated porin OmpU confers resistance to antimicrobial peptides. Infect Immun. 2004;72:3577–3583. doi: 10.1128/IAI.72.6.3577-3583.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Li CC, Crawford JA, DiRita VJ, Kaper JB. Molecular cloning and transcriptional regulation of ompT, a ToxR-repressed gene in Vibrio cholerae . Mol Microbiol. 2000;35:189–203. doi: 10.1046/j.1365-2958.2000.01699.x. [DOI] [PubMed] [Google Scholar]
- 64.Buckley AM, Webber MA, Cooles S, Randall LP, La Ragione RM, et al. The AcrAB-TolC efflux system of Salmonella enterica serovar Typhimurium plays a role in pathogenesis. Cell Microbiol. 2006;8:847–856. doi: 10.1111/j.1462-5822.2005.00671.x. [DOI] [PubMed] [Google Scholar]
- 65.Taylor DL, Bina XR, Bina JE. Vibrio cholerae VexH encodes a multiple drug efflux pump that contributes to the production of cholera toxin and the toxin co-regulated pilus. PLoS One. 2012;7:e38208. doi: 10.1371/journal.pone.0038208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Bina XR, Philippart JA, Bina JE. Effect of the efflux inhibitors 1-(1-naphthylmethyl)-piperazine and phenyl-arginine-beta-naphthylamide on antimicrobial susceptibility and virulence factor production in Vibrio cholerae . J Antimicrob Chemother. 2009;63:103–108. doi: 10.1093/jac/dkn466. [DOI] [PubMed] [Google Scholar]
- 67.Doron S, Melamed S, Ofir G, Leavitt A, Lopatina A, et al. Systematic discovery of antiphage defense systems in the microbial pangenome. Science. 2018;359:eaar4120. doi: 10.1126/science.aar4120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Baddam R, Sarker N, Ahmed D, Mazumder R, Abdullah A, et al. Genome dynamics of Vibrio cholerae isolates linked to seasonal outbreaks of cholera in Dhaka, Bangladesh. mBio. 2020;11:e03339-19. doi: 10.1128/mBio.03339-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Murphy SG, Johnson BA, Ledoux CM, Dörr T. Vibrio cholerae’s mysterious seventh pandemic island (VSP-II) encodes novel Zur-regulated zinc starvation genes involved in chemotaxis and autoaggregation. PLoS Genet. 2021;17:e1009624. doi: 10.1371/journal.pgen.1009624. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.