

ABSTRACT
The assembly and maintenance of most cilia and eukaryotic flagella depends on intraflagellar transport (IFT), the bidirectional movement of multi-megadalton IFT trains along the axonemal microtubules. These IFT trains function as carriers, moving ciliary proteins between the cell body and the organelle. Whereas tubulin, the principal protein of cilia, binds directly to IFT particle proteins, the transport of other ciliary proteins and complexes requires adapters that link them to the trains. Large axonemal substructures, such as radial spokes, outer dynein arms and inner dynein arms, assemble in the cell body before attaching to IFT trains, using the adapters ARMC2, ODA16 and IDA3, respectively. Ciliary import of several membrane proteins involves the putative adapter tubby-like protein 3 (TULP3), whereas membrane protein export involves the BBSome, an octameric complex that co-migrates with IFT particles. Thus, cells employ a variety of adapters, each of which is substoichiometric to the core IFT machinery, to expand the cargo range of the IFT trains. This Review summarizes the individual and shared features of the known cargo adapters and discusses their possible role in regulating the transport capacity of the IFT pathway.
Keywords: BBSome, Cilia, Flagella, Intraflagellar transport, Dynein arms, Radial spokes
Summary: Intraflagellar transport trains use cargo adapters to bind certain ciliary proteins and building blocks, shuttling them from the cell body into growing cilia.
Introduction
Cilia are microtubule-based cell protrusions with motile and sensory functions. Ciliary malfunction leads to a plethora of diseases and developmental disorders, which has kindled an interest into how cilia are assembled and maintained (Brown and Witman, 2014; Fliegauf et al., 2007). Because cilia lack ribosomes, their building blocks need to be imported from the cell body (Rosenbaum and Child, 1967). On their way into the cilium, the proteins must pass through the transition zone, which functions as a barrier, rendering ciliary entry of larger proteins and complexes by diffusion inefficient (Garcia-Gonzalo and Reiter, 2012; Kee et al., 2012). Intraflagellar transport (IFT), the dedicated protein shuttle of cilia, moves many proteins and building blocks in and out of cilia, likely explaining the critical role of IFT in ciliary assembly (Kozminski et al., 1993; Lechtreck, 2015). In most cell types, IFT continues perpetually in full-length cilia, contributing to ciliary maintenance, signaling and adjustments of composition.
The IFT machinery consists of IFT-A and IFT-B complexes and the associated motors – heterotrimeric kinesin-2 for anterograde transport to the ciliary tip and IFT dynein for retrograde transport back to the cell body (Fig. 1A) (Cole et al., 1998; Pazour et al., 1999; Piperno and Mead, 1997). The IFT complexes transiently assemble into multi-megadalton IFT trains, which cycle through cilia (Wingfield et al., 2017). In Chlamydomonas, anterograde trains, which are key for protein import and ciliary assembly, form around a string of IFT-B particles (Hirano et al., 2017; Jordan et al., 2018; Wingfield et al., 2021). The IFT-B backbone is flanked by the shorter IFT-A particle layer and an even shorter layer of autoinhibited IFT dynein (Jordan et al., 2018; Toropova et al., 2019; van den Hoek et al., 2022). The interactions within and between the IFT complexes have been intensively studied, and preliminary structures of the IFT particles and anterograde trains are available (Katoh et al., 2015; Kobayashi et al., 2021; Lacey et al., 2022 preprint; McCafferty et al., 2022; Petriman et al., 2022; Taschner et al., 2014; Taschner et al., 2016). In addition to organizing the IFT motors, the IFT complexes also form a platform to which other ciliary proteins attach for transport (Fig. 1) (Lechtreck, 2015).
Fig. 1.

Overview of the IFT pathway and IFT interactions. (A) Schematic presentation of the IFT pathway. (1) IFT proteins and complexes (blue dots) are recruited to the cilia base, assembled into IFT trains (blue rectangles) and loaded with cargo. (2) Anterograde trains enter the cilium and move to the tip. (3) At the tip, the trains make a U-turn, (4) return by retrograde IFT to the ciliary base, and (5) are largely disassembled. (B) Organization of anterograde IFT trains. The trains form around an IFT-B backbone (dark blue) to which the IFT-A complex (light blue) and the IFT motors are attached. Whereas tubulin binds directly to IFT-B proteins, transport of several axonemal and membrane-bound cargoes involves cargo adapters. RSs, radial spokes.
Proteomic studies in a variety of species show that cilia consist of more than 1000 distinct proteins (Li et al., 2004; Pazour et al., 2005; Stolc et al., 2005). Although some proteins enter cilia by diffusion, the transport of many ciliary membrane, matrix and axonemal proteins involves the IFT pathway (Belzile et al., 2013; Craft Van De Weghe et al., 2020; Harris et al., 2016; Lechtreck, 2015). Extrapolation of the range of transported proteins confirmed by in vivo imaging suggests that IFT carries an abundance of distinct proteins as cargoes, raising the question of how the trains, which consist of just 22 proteins, interact with such a large and diverse set of proteins. For most cargoes, the IFT binding sites remain unknown, but a few such contacts have been characterized. In vitro studies have revealed that tubulin dimers bind to the N-terminal domains of the IFT-B proteins IFT74 and IFT81, and the removal of these tubulin-binding domains from both proteins by gene engineering impairs tubulin transport and ciliary assembly in Chlamydomonas (Bhogaraju et al., 2013a; Craft Van De Weghe et al., 2020; Kubo et al., 2016). Thus, tubulin, the main structural protein of cilia, appears to bind directly to the IFT-B backbone. Recent structural data of the IFT-B complex, however, indicate that the N-terminal domain of IFT81 is largely embedded in the IFT-B particles, raising the question of how it interacts with tubulin dimers in the assembled trains (Lacey et al., 2022; Petriman et al., 2022). Several other cargoes and axonemal substructures connect indirectly to the IFT trains using adapters, which are defined here as proteins or complexes necessary to link cargoes to IFT trains without being a steady or necessary part of either the trains or cargoes (Fig. 1B). This definition excludes certain IFT subunits – for example IFT25 (also known as HSPB11 in mammals), IFT27 (also known as RABL4) and IFT56 (also known as DYF13 in Chlamydomonas, TTC26 in mammals) – that are not essential for ciliary assembly or IFT itself but appear to function solely in cargo transport (Eguether et al., 2014; Huet et al., 2014, 2019; Ishikawa et al., 2014; Liew et al., 2014). However, because of cell type- and species-specific differences in the contribution of such proteins to ciliary assembly, they will not be discussed further. This Review focuses on three adapters for axonemal cargoes – ARMC2, IDA3 and ODA16 (also known as DAW1 or WDR69) – as well as tubby-like protein 3 (TULP3) and the BBSome complexes, which are involved in ciliary membrane protein traffic.
Adapters for axonemal proteins
In addition to α- and β-tubulin, subunits of the inner and outer dynein arms (IDAs and ODAs, respectively), the radial spokes, the central pair and the nexin–dynein regulatory complex (N-DRC) have been observed to move by IFT (Craft et al., 2015; Craft Van De Weghe et al., 2020; Dai et al., 2018; Hao et al., 2011; Lechtreck et al., 2018; Luo et al., 2017; Wren et al., 2013). ODAs (∼2 MDa), the double-headed IDA I1/f (∼1.5 MDa) and the ∼710 kDa radial spoke precursor, which lacks several subunits present in mature axoneme-bound radial spokes, are preassembled in the cell body before transport into the cilia by IFT. The transport of these three large axonemal substructures requires the adapter proteins ODA16, IDA3 and ARMC2, respectively (Fig. 1) (Ahmed et al., 2008; Hunter et al., 2018; Lechtreck et al., 2022). The adapters are not part of the substructures once assembled into the axoneme, and adapter mutants largely lack the corresponding substructure from the otherwise unaffected axonemes (Ahmed et al., 2008; Huang et al., 1981; Kamiya et al., 1991; Viswanadha et al., 2014). Thus, each of these adapters mediates transport of a single cargo by generating a cargo-specific binding site on the IFT trains. It is unknown whether other axonemal substructures, such as single-headed inner arm dyneins or central pair projections, also require adapters for transport into cilia.
ARMC2
The armadillo repeat protein ARMC2 is widely conserved in organisms with motile cilia (Merchant et al., 2007). Genome-wide association studies have identified ARMC2 as a locus associated with reduced lung function in humans (Soler Artigas et al., 2011). Whole-exome sequencing has shown that homozygous mutations in ARMC2 cause asthenozoospermia, male infertility caused by immotile spermatozoa (Coutton et al., 2019). In Chlamydomonas, ARMC2 is encoded by PF27, and pf27 mutant cilia are largely paralyzed due to the near absence of radial spokes, which regulate the activity of axonemal dyneins (Alford et al., 2013; Lechtreck et al., 2022). ARMC2 is not a component of the mature axoneme-bound radial spokes (Huang et al., 1981).
ARMC2 consists of an extended N-terminal intrinsically disordered region (IDR) and a C-terminal helical region encompassing several armadillo repeats forming an α-solenoid (Fig. 2A; Box 1). Fluorescent protein tagging of RSP3, an essential radial spoke protein present in the cell body precursor complex, has revealed co-migration of RSP3 with ARMC2 by IFT, and in the absence of ARMC2, IFT of RSP3 is abolished (Lechtreck et al., 2022). It has also been shown that ARMC2 dissociates from IFT trains at the ciliary tip, concurrently releasing its cargo, and returns to the cell body mostly by diffusion (Lechtreck et al., 2022). IFT of ARMC2 continues in mutants lacking intact radial spokes, indicating that ARMC2 binds to anterograde trains independently of its cargo (Lechtreck et al., 2022). Furthermore, anterograde IFT of fluorescently tagged ARMC2 (and of radial spokes, if present) is frequent during ciliary growth and is downregulated in full-length cilia, when demand for radial spokes in cilia is low (Lechtreck et al., 2022).
Fig. 2.
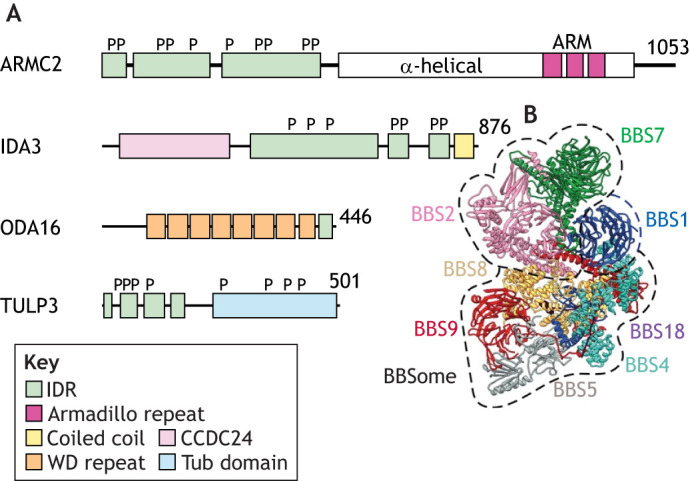
Domain structure of IFT cargo adapters. (A) Schematic representations of the adapter proteins ARMC2, IDA3 and ODA16 from Chlamydomonas and TULP3 from humans; Chlamydomonas TULPs have not yet been characterized. The number of residues and the domain structures are indicated. IDRs were predicted using Iupred3 (see Box 1) (Erdos and Dosztanyi, 2020). The positions of phosphorylation sites are marked (P) based on a previously published phosphoproteomic analysis for the Chlamydomonas proteins (Wang et al., 2014) and on the PhosphoSitePlus database record for human TULP3 (https://www.phosphosite.org/). TULP3 and ARMC2 each encompass two distinct domains: an IDR and either a Tub domain or an armadillo repeat domain (ARM) forming an α-solenoid, respectively. See Box 1 for a discussion of the possibility that the IDRs of these adapters bind to IFT particles, while their other domains form cargo-specific binding sites. (B) Atomic model of the mammalian BBSome. Image adapted from Singh et al. (2020), where it was published under a CC BY 4.0 license.
Box 1. Several adapters have IDRs.
Several known adapters (ARMC2, IDA3, TULP3 and ODA16) are predicted to encompass structurally ambiguous domains referred to as intrinsically disordered regions (IDRs; Fig. 2). Intrinsically disordered proteins and protein domains are abundant in all organisms (Dyson and Wright, 2005; Wright and Dyson, 2015). Structural predictions using IUPred3 (see box figure, panel A; https://iupred3.elte.hu/) and AlphaFold2 (see box figure, panel B; https://alphafold.ebi.ac.uk/) indicate that human ARMC2 and TULP3 have a bipartite structure consisting of extended N-terminal IDRs and a C-terminal α-helical region or Tub domain, respectively. Since these adapters move different cargoes, IFT binding could be mediated by the common IDRs. Indeed, the short helical IFT-A-binding motif of mammalian TULP3 is embedded within the IDR (see box figure), and overexpression of this domain has a dominant-negative effect on GPCR transport into cilia (Mukhopadhyay et al., 2010). ODA16 possesses only a small C-terminal IDR, which is expendable for binding to IFT46 (Taschner et al., 2017). Furthermore, both the N-terminal domain and the ODA-binding β-propeller region of ODA16 are necessary to bind IFT46, arguing against a bipartite structure with separate IFT- and cargo-binding regions for this adapter (Taschner et al., 2017). However, the N-terminal region of IFT46, which is conserved in organisms with motile cilia and is critical for ODA16 binding and ODA transport, is predicted to be disordered (Bhogaraju et al., 2013b; Hou and Witman, 2017). Thus, in this case, the IDR of an IFT protein mediates IFT–adapter interaction. IDRs can undergo disorder-to-order transitions, allowing them to fold onto a heterologous binding partner to mediate protein–protein interaction. Disorder-to-order transitions of IDRs are often induced by posttranslational modifications, including phosphorylation (Bah and Forman-Kay, 2016; Iakoucheva et al., 2004). All known phosphorylated sites, as identified by whole-cell phosphoproteomics, of Chlamydomonas ARMC2 (10 sites), IDA3 (seven sites; Fig. 2), IFT46 (five sites), and the uncharacterized TULPs TLP1 (11 sites) and TLP2 (18 sites) are located in the predicted IDRs (Wang et al., 2014). Hypothetically, changes in the phosphorylation pattern of the IDRs could regulate adapter binding to IFT trains and thereby the transport frequency of the attached cargoes.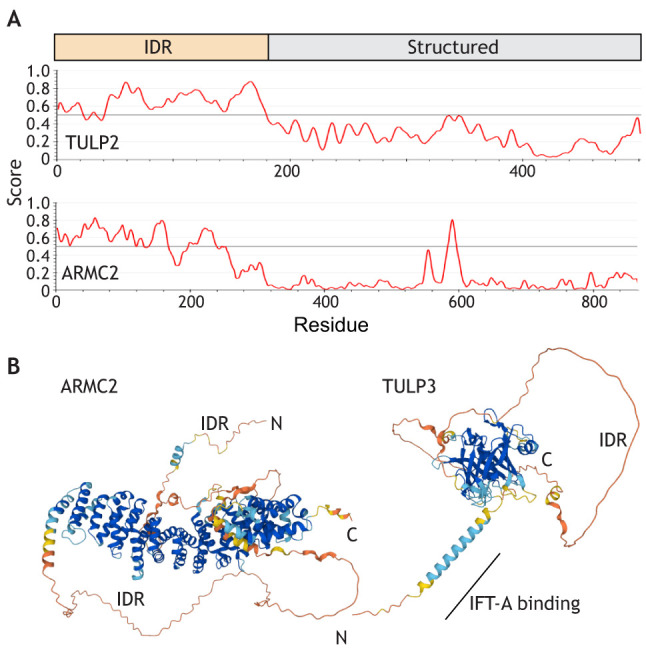
The radial spoke stalks continue to enter cilia and attach to axonemes in spoke-head-deficient mutants, suggesting that the stalk portion of the radial spoke precursor encompasses the ARMC2-binding site (Luck et al., 1977). Phylogenetic data suggest that the stalk protein RSP3 could be the binding partner of ARMC2 (Fig. 3) (Lechtreck et al., 2022). Interestingly, in axoneme-bound radial spokes, RSP3 interacts with the armadillo repeat proteins RSP8 and RSP14, which are not part of the precursor and are added to the spokes only after transport in the cilium (Gui et al., 2021; Gupta et al., 2012; Yang et al., 2006). The ARMC2 docking site on the IFT train remains unknown, but it is generally thought that structural proteins move via the IFT-B particle, as this complex is essential for ciliogenesis and is the known carrier of tubulin and the ODAs (Bhogaraju et al., 2013b). A few apparently normal radial spokes accumulate in the most proximal region of pf27 mutant cilia, indicating ARMC2-independent low-frequency entry into mutant cilia (Alford et al., 2013). In conclusion, ARMC2 is an adapter that mediates IFT of radial spokes to ensure assembly along the length of the cilia.
Fig. 3.

Overview of IFT adapter–cargo interactions. Diagram summarizing the interactions between IFT proteins, adapters and their cargoes. Tubulin dimers bind directly to IFT-B via IFT74 and IFT81. The BBSome associates with the IFT-B complex via IFT25–IFT27 (IFT25/27), and TULP3 associates with the IFT-A complex, apparently via IFT122. Both are multivalent adapters participating in the transport of diverse transmembrane and membrane-associated proteins, including GPCRs, ion channels and single-pass transmembrane (TM) proteins. The ciliary targeting sequences (CTSs; turquoise) of BBSome and TULP3 cargoes are also the adapter-binding sites. Ubiquitylation of some GPCRs (such as SMO and SSTR3) is required for BBSome-dependent export from cilia. The IFT complexes and the BBSome lack known ubiquitin-binding sites, and the interaction between the BBSome and ubiquitin sidechains may involve TOM1L2 as an additional adapter. The known adapters for axonemal substructures each have a single cargo specificity. For ODA16, the binding partner on the IFT trains is the N-terminal domain of IFT46, and IFT56 is reported to be the IFT binding partner for IDAs. RSP3 is the only radial spoke (RS) protein expressed in Drosophila auditory neurons, which form cilia by the IFT pathway, suggesting that it could be the binding partner of ARMC2.
IDA3
In addition to one or two types of ODA, most motile cilia contain several types of IDA, including IDA I1/f, which is the only IDA with two dynein heavy chains. IDA I1/f is almost completely missing from cilia of the Chlamydomonas ida3 mutant (Hunter et al., 2018; Viswanadha et al., 2014). IDA3, which is not conserved outside of green algae, is an 816-residue protein with extended IDRs and a coiled-coil domain-containing 24 motif (CCDC24; Fig. 2) (Hunter et al., 2018). Fluorescently tagged IDA3 moves by anterograde IFT to the tip, whereas it returns to the cell body mostly by diffusion; its anterograde transport frequency is highly upregulated during ciliary growth (Hunter et al., 2018). Levels of IDA I1/f and other IDAs are reduced in cilia of a Chlamydomonas mutant lacking the IFT-B protein IFT56, suggesting that IFT56 could be involved in IDA3 binding and IDA transport (Fig. 3) (Ishikawa et al., 2014). IDA3 and the IC140 subunit of I1/f (encoded by IDA7) can be co-immunoprecipitated from ciliary extracts (Hunter et al., 2018), and unpublished observations show that IDA3 and IC140 co-migrate by IFT, with the transport frequency of IDA3 exceeding that observed for IC140 (Emily Hunter, Juyeon Hwang, Win Sale and K.L., unpublished). IDA3 continues to move by IFT in the absence of IDA I1/f, revealing cargo-independent binding to IFT trains (Hunter et al., 2018). In conclusion, IDA3 is an adapter necessary IFT for of IDA I1/f.
ODA16
Outer dynein arm 16 (ODA16) is a ∼50 kDa WD-repeat protein that is conserved in many organisms with motile cilia (Fig. 2). In the Chlamydomonas oda16 mutant, IFT of ODAs is almost abolished and the number of ODAs in cilia is reduced by ∼75% (Ahmed et al., 2008; Dai et al., 2018). Knockdown of Oda16 in zebrafish impairs assembly of ODAs onto the axonemes (Gao et al., 2010). However, ODA16 is not part of mature axoneme-bound ODAs in Chlamydomonas (Ahmed and Mitchell, 2005). Taken together, these observations suggest a conserved role of ODA16 in ODA transport by IFT (Ahmed et al., 2008; Ahmed and Mitchell, 2005).
A yeast two-hybrid screen using human ODA16 as a bait against a mouse testis-derived cDNA library identified the IFT-B subunit IFT46 as a potential interactor (Fig. 3) (Ahmed et al., 2008). Indeed, Chlamydomonas strains expressing an N-terminally truncated IFT46 in an ift46 mutant background (ift46 IFT46ΔN) assemble cilia that lack most ODAs but are of otherwise normal ultrastructure (Hou et al., 2007; Hou and Witman, 2017). Chlamydomonas ODA16 forms an eight-bladed WD β-propeller, which interacts with the ODAs (Fig. 2A) (Taschner et al., 2017; Wang et al., 2020). The small N–terminal domain of Chlamydomonas ODA16 generates a cleft within the β-propeller to accommodate the unstructured N-terminal domain of IFT46. Thus, ODA16 interacts with the N-terminal domain of IFT46 to ensure efficient ODA transport (Fig. 3). Although structurally conserved, human ODA16 and IFT46 fail to form a complex in vitro, suggesting that additional proteins might be involved in mediating ODA–IFT interactions and ODA transport (Wang et al., 2020). Indeed, IFT of ODAs is occasionally observed in Chlamydomonas oda16 mutants, and curiously, residual ODAs cluster in a central region of oda16 mutant cilia (Dai et al., 2018). In contrast, ift46 IFT46ΔN mutants lack most ODAs, with only a few complexes clustered at the very proximal end of the cilia, as one would expect if some ODAs enter the cilia by diffusion and then dock to the nearest available axonemal sites (Dai et al., 2018; Hou and Witman, 2017). In the ift46 IFT46ΔN strain, but not the oda16 mutant, ODAs accumulate near the basal bodies, suggesting that ODA16 also participates in recruiting ODAs to the ciliary base for loading onto IFT trains (Dai et al., 2018). Once the ODA complexes are assembled in the cell body, putative maturation factors – ODA5, ODA8 and ODA10 – ensure that they become competent for transport by IFT and axonemal docking (Dai et al., 2018; Dean and Mitchell, 2015; Desai et al., 2015; Wirschell et al., 2004). In the ciliate Tetrahymena, the presence of ODAs in cilia is reduced following mutation of Shulin (an ortholog of human DNAAF9); this conserved protein functions as a packing factor, keeping ODAs inactive during transport (Mali et al., 2021). Thus, IFT of ODAs is complex and appears to involve other proteins in addition to ODA16 and IFT46.
Adapters for ciliary membrane proteins
Ciliary transmembrane proteins travel in post-Golgi vesicles or from the plasma membrane via the recycling endosome to the periciliary plasma membrane, from where they move laterally into the ciliary membrane (Dentler, 2013; Long and Huang, 2019; Monis et al., 2017; Quidwai et al., 2021). Several transmembrane proteins – including the TRPV channels OSM-9 and OCR-2 in Caenorhabditis elegans (Qin et al., 2005; van Krugten et al., 2022); the olfactory channel CNGA2, the odorant receptor Olfr78 (also known as OR51E2), adenylate cyclase III (also known as ADCY3; Williams et al., 2014) and G-protein-coupled receptors (GPCRs; Ye et al., 2018) in mammalian cells; and PKD2 in Chlamydomonas (Huang et al., 2007; Liu and Lechtreck, 2018) – have been observed to at least occasionally move by IFT. IFT has also been observed for proteins associated to the ciliary membrane by lipidation (Liu and Lechtreck, 2018; Yu et al., 2020). The small GTPase ARL13B, for example, undergoes IFT in murine olfactory cilia but mostly moves by diffusion in C. elegans and Chlamydomonas, indicating species-specific differences (Cevik et al., 2010; Dai et al., 2022; Roy et al., 2017; Williams et al., 2014). Certain membrane proteins enter and exit cilia by diffusion, either exclusively or in addition to IFT (Belzile et al., 2013; Calvert et al., 2006; Cao et al., 2015; Domire et al., 2011; Ye et al., 2018). Also, transmembrane proteins can exit cilia by vesicle shedding rather than by retrograde IFT (Ojeda Naharros and Nachury, 2022; Wang and Barr, 2018). To conclude, the role of IFT in the bulk delivery of membrane proteins during ciliary assembly and the tuning of membrane composition in development and signaling is less clear. IFT also functions in organizing ciliary membrane domains. The salt-sensing receptor-type guanylate cyclase GCY-22, for example, is concentrated in the ciliary tip region of C. elegans ASER neurons because anterograde IFT relentlessly returns escaped receptors diffusing along the cilium to the tip region (van der Burght et al., 2020). IFT-A and IFT-B participate in the transport of ciliary membrane proteins either directly (IFT-A) or, as discussed below, via the adapters TULP3 (attached to IFT-A) and the BBSome complex (attached to IFT-B) (Fu et al., 2016; Mukhopadhyay et al., 2010; Picariello et al., 2019). An adapter function has also been proposed for lebercilin (LCA5), which interacts with IFT-B proteins but is expendable for IFT and ciliary assembly (Boldt et al., 2011). LCA5 is required for proper translocation of opsin and arrestin proteins into the outer segment of photoreceptor cells (Boldt et al., 2011), but transport by IFT has not been shown.
The BBSome
The BBSome, which is occasionally referred to as the third IFT complex, is a conserved octameric complex that cycles through cilia via IFT (Fig. 2B) (Blacque et al., 2004; Lechtreck et al., 2009; Nachury et al., 2007; Singh et al., 2020). The BBSome is substoichiometric to the core IFT complexes (Box 2) and, in most cell types analyzed, is not required for IFT per se or the initial assembly of cilia (Lechtreck et al., 2009; Mykytyn et al., 2004). However, in various species, mutation of genes encoding Bardet–Biedl syndrome (BBS) protein subunits of the BBSome results in changes in the composition of the ciliary membrane, including loss or reduction and abnormal accumulation of proteins (Wingfield et al., 2018). The resulting biochemical defects interfere with ciliary maintenance and signaling and are thought cause the ciliopathy BBS in humans (Mykytyn et al., 2004; Nishimura et al., 2004; Sheffield, 2010). In C. elegans, the BBSome has an additional role in stabilizing IFT trains (Ou et al., 2005).
Box 2. Adapters and cargoes form an irregular coat on the IFT trains.
Here, I summarize data on the ratios between IFT particle proteins, adapters and cargoes within IFT trains. Quantitative mass spectrometry has revealed a ratio of 1:6 between BBS1 and the IFT-B protein IFT81 in Chlamydomonas cilia (Lechtreck et al., 2009). A similar 1:6.2 ratio for BBSomes and IFT-B has been estimated based on quantitative imaging of tagged proteins in primary cilia (Ye et al., 2018). Furthermore, peptides of IFT proteins were found to be abundant in a proteomic analysis of full-length Chlamydomonas cilia, whereas the axonemal cargo adapters ARMC2, IDA3 and ODA16, as well as most BBSome subunits, TLP1 and TLP2, were not detected (see the Chlamydomonas flagellar proteome database at http://chlamyfp.org/ChlamyFPv2/index.php; Pazour et al., 2005). The data indicate that adapters are substoichiometric to proteins of the IFT core machinery in full-length cilia.
In vivo imaging has shown that not all IFT trains carry a tagged BBSome and that not all BBSomes moving by IFT are engaged with a cargo (i.e. PLD; Liu and Lechtreck, 2018; Xue et al., 2020). The available in vivo imaging data for other IFT particle–adapter–cargo triads indicate similar ratios (i.e. IFT particle proteins>ARMC2>radial spoke proteins and IFT particle proteins>IDA3>IDA I1/f) (Hunter et al., 2018; Lechtreck et al., 2022). Thus, a given adapter is substoichiometric to the IFT complexes, and cargoes are substoichiometric to their adapters, suggesting that IFT trains carry an irregular coat of adapters and cargoes (see box figure). The IFT frequencies of several axonemal proteins and their adapters are increased during ciliary growth, suggesting that IFT trains entering growing cilia carry a higher load of adapters and cargoes.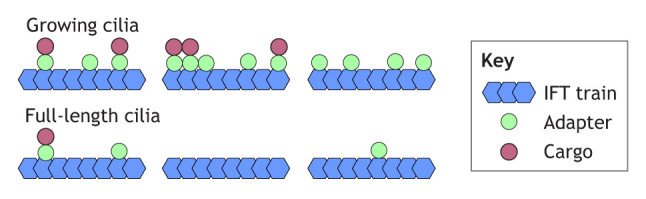
The main type of cargo of BBSome-dependent IFT in mammalian cilia are GPCRs (Wingfield et al., 2018). In vivo imaging studies suggest that the GPCR SSTR3 co-migrates with BBSomes on IFT trains, an association that ensures that ligand-bound SSTR3 is exported from cilia (Ye et al., 2018). However, several GPCRs, such as SSTR3, MCHR1 and NPY2R, are absent from cilia of murine cultured neuronal cells lacking subunits of the BBSome (Berbari et al., 2008b; Loktev and Jackson, 2013), and in Paramecium, ion channels are lost from cilia upon knockdown of BBS gene expression (Valentine et al., 2012), suggesting a contribution of the BBSome to the import of these proteins into cilia. The third intracellular loop and/or parts of the C-terminal tail of several ciliary GPCRs form a BBSome-binding motif (Berbari et al., 2008a; Klink et al., 2017; Nachury, 2018). It has been shown that ubiquitylation within the third intracellular loop earmarks GPCRs for ciliary export, which likely involves BBSome-dependent IFT (Fig. 3) (Desai et al., 2020; Lv et al., 2021; Shinde et al., 2022 preprint; Shinde et al., 2020). Recent data indicate that the conserved ESCRT protein target of Myb1-like 2 membrane trafficking protein (TOM1L2) mediates the interaction between ubiquitin chains on the GPCRs and the BBSome, but BBSome-dependent IFT of TOM1L2 has not yet been demonstrated (Fig. 3) (Shinde et al., 2022 preprint).
In Chlamydomonas, mutation of BBS genes results in accumulation of several lipidated membrane-associated proteins in cilia (Lechtreck et al., 2009). These include phospholipase D (PLD), which can enter cilia by diffusion but requires BBSome-dependent IFT to exit cilia (Liu and Lechtreck, 2018). Tagged PLD co-migrates with BBSomes on IFT trains, and IFT of PLD is abolished in the bbs1 mutant. The N-terminal 20 residues of PLD, which encompass the GC motif for myristoylation and palmitoylation, are sufficient for both ciliary accumulation in cells with BBS gene mutations and BBSome-dependent IFT in control cells (Liu and Lechtreck, 2018).
In mammalian cells and Chlamydomonas, BBSome docking to the IFT-B complex involves the IFT25–IFT27 heterodimer (Dong et al., 2017; Eguether et al., 2014; Liew et al., 2014). Murine Ift25−/− and Ift27−/− mutants and Chlamydomonas IFT25-knockdown cells assemble full-length cilia accumulating BBSomes and BBSome cargoes. Thus, the IFT25–IFT27 dimer appears to function specifically during retrograde IFT of BBSomes, suggesting the presence of a currently unknown, second BBSome docking site on the anterograde trains. However, the role of these IFT proteins in BBSome transport and IFT appears to differ between species and cell types, as IFT25 and IFT27 are absent in certain species with cilia and BBSomes – such as C. elegans and Drosophila melanogaster (van Dam et al., 2013), IFT25 depletion interferes with the assembly of new flagella in Trypanosoma (Huet et al., 2014, 2019), and murine mutants lacking IFT25 or IFT27 fail to assemble sperm flagella (Eguether et al., 2014; Liu et al., 2017).
The BBSome and its GPCR cargoes also accumulate in primary cilia following knockout or knockdown of leucine zipper transcription factor-like 1 (LZTFL1; Eguether et al., 2014; Seo et al., 2011). It has been suggested that this conserved coiled-coil protein mediates binding of the BBSome to IFT25–IFT27 dimers during retrograde IFT. However, Chlamydomonas LZTFL1 is of low abundance in cilia, and IFT of GFP-tagged LZTFL1 is not observed, arguing against a role as an adapter for BBSome anchoring onto the IFT trains (Sun et al., 2021). The regulation of BBSome–cargo and BBSome–IFT particle interactions by several small GTPases is discussed below. Taken together, data from several species indicate that the BBSome is an adapter mediating IFT of several ciliary membrane proteins.
TULP3
Tubby-like proteins (TULPs) are widely distributed in animals, plants and protists, and are characterized by the conserved C-terminal Tub domain (Pfam PF01167), which consists of a 12-strand β-barrel surrounding a central α-helix (Fig. 1; Box 1). TULPs bind to phosphatidylinositol 4,5-bisphosphate (PIP2) at the plasma membrane and function as transcriptional regulators in the nucleus (Mukhopadhyay and Jackson, 2011; Santagata et al., 2001). In mammals, mutations in several members of the family have been linked to obesity, kidney anomalies and retinal degeneration – a combination of features often linked to ciliary dysfunction (Kleyn et al., 1996). The abundance of numerous proteins – including the GPCRs SSTR3, GPR161 and MCHR1; the polycystin 1–polycystin 2 complex; and the single-pass cystogenic protein fibrocystin – are reduced in TULP3-deficient cilia, suggesting that TULP3 is a multivalent adapter (Badgandi et al., 2017; Legue and Liem, 2019). TULP3 is also involved in ciliary import of membrane-associated proteins, including the ciliary GTPase ARL13B and the inositol polyphosphate 5-phosphatase INPP5E, which both function in protein trafficking themselves (Gotthardt et al., 2015; Han et al., 2019; Hwang et al., 2019).
The current model proposes that TULP3 links membrane proteins to the IFT trains for transfer into cilia. In detail, TULP3 is recruited to the periciliary plasma membrane via binding of the Tub domain to PIP2. The activated Tub domain then binds the ciliary targeting sequences of its cargoes (Badgandi et al., 2017). The N-terminal region of mammalian TULP3, while predicted to be mostly disordered, encompasses an α-helix (residues 10–44) that binds the IFT-A core complex; recent data suggest that TULP3 binds to the IFT-A protein IFT122 (Figs 2 and 3; Box 1) (Hesketh et al., 2022 preprint; Mukhopadhyay et al., 2010). C. elegans mutants lacking TUB-1, the ortholog of mammalian TULP3, develop stunted olfactory cilia with an aberrant GPCR content (Brear et al., 2014; DiTirro et al., 2019). IFT of GFP-tagged TUB-1 was not observed in AWR neurons within the short proximal segment of cilia suitable for IFT imaging, generating some uncertainty about whether TUB-1 indeed functions as a co-migrating IFT adapter (DiTirro et al., 2019). In an alternative model, IFT-A, instead of TULP3, directly binds these transmembrane proteins (Nachury, 2018). IFT-A exposes an array of β-propeller and tetratricopeptide repeat (TPR) domains on its surface as potential cargo binding sites (Hesketh et al., 2022 preprint; McCafferty et al., 2022). In this scenario, TULP3, rather than being an adapter that co-migrates with trains in cilia, could function as a loading factor at the ciliary base and within cilia, priming IFT-A for cargo binding or recruiting the IFT-A complex to the membrane for cargo pick-up.
Shared and distinguishing attributes of adapters
A shared feature of all known adapters is that they are substoichiometric to the IFT particles (see Box 2), but distinguishing features of the adapters for axonemal and membrane proteins are also emerging. ARMC2, IDA3 and ODA16 each mediate IFT of a single complex, whereas the BBSome complex is a multivalent adapter involved in IFT of multiple proteins within a species and distinct types of proteins between species (Fig. 3) (Ahmed et al., 2008; Hunter et al., 2018; Lechtreck et al., 2022; Wingfield et al., 2018). TULP3 also participates in ciliary entry of several transmembrane proteins, although in vivo imaging data confirming transport by IFT are not yet available (Badgandi et al., 2017). The adapters for axonemal complexes are considerably more abundant in growing cilia than in full-length cilia, and ARMC2 and IDA3 show a marked preference for anterograde trains and diffuse back to the cell body, in agreement with a role in delivering building blocks to the cilium tip (Hunter et al., 2018; Lechtreck et al., 2022). In contrast, BBSomes move bidirectionally by IFT and are constitutively present in full-length cilia (Lechtreck et al., 2009). These differences could indicate principal differences in IFT of axonemal and ciliary membrane proteins, with the former transported predominantly during the assembly phase of cilia and the latter apparently transported constitutively to exchange membrane proteins as part of ciliary maintenance and in response to signals.
The function of IFT adapters
Is assisting in the transport of proteins by IFT the sole role of the adapters? If so, adapters would not be needed during IFT-independent cytoplasmic assembly of cilia, as occurs during gametogenesis in apicomplexan parasites and Drosophila testis. The Drosophila genome encodes orthologs of ARMC2 (CG32668), ODA16 (CG7568) and the BBSome subunits. However, expression of these proteins is not detected in testis but is observed in chordotonal neurons, which assemble auditory cilia, containing dynein arms and likely containing RSP3 (Rsph3), in an IFT-dependent manner (Zur Lage et al., 2019; Andrew Jarman, personal communication). Thus, adapters, like IFT proteins, are required during the assembly of projecting cilia but not during the formation of cytoplasmic cilia. Furthermore, Chlamydomonas ODAs isolated from oda16 mutant cell bodies will bind to axonemes from oda16 mutants in vitro, showing that both mutant ODAs and axonemes are docking competent (Ahmed et al., 2008). This suggests that ODA16 specifically serves to move ODAs from the cell body into the cilium.
Axonemal dyneins form in droplet organelles, which contain chaperones and other dynein-specific assembly factors (Horani et al., 2018; Lee et al., 2020). During cytoplasmic development of sperm flagella in Drosophila, ribonuclear particles, containing chaperones and dynein heavy chain mRNAs, localize near the tip of the elongating axoneme, likely supplying the axoneme with ODAs in an IFT-independent manner (Fingerhut and Yamashita, 2020). Thus, the assembly of large axonemal complexes could require an environment that is specific to the cell body and is absent from narrow, entry-limited, membrane-enclosed cilia.
The adapters for structural proteins identified so far assist in IFT of large preassembled axonemal complexes, raising the question of whether cargo size makes the use of adapters necessary for transport (Ahmed et al., 2008; Hunter et al., 2018; Lechtreck et al., 2022). In anterograde trains, the IFT-B particles are densely covered with IFT-A particles, BBSome, tubulin dimers, ODA16 and likely other adapters (Jordan et al., 2018). This limits the area available for other interactions, probably forcing many cargoes to cluster around exposed parts of the IFT-B complex (Lacey et al., 2022 preprint). In that case, attaching multiple large axonemal substructures to train-embedded IFT-B complexes could be sterically hindered, and the adapters could function as spacers between the IFT train and the actual cargoes, allowing for better use of the available train surface. It should be noted that the IFT-B layer is exposed where it protrudes from both ends of trains, potentially providing a large area for interactions (van den Hoek et al., 2022).
Are adapters involved in the regulation of ciliary protein transport?
Adapters for axonemal proteins
As linkers between the IFT trains and cargoes, adapter proteins are in a key position to regulate the amount of cargo entering a cilium. Anterograde IFT of many axonemal building blocks is upregulated during Chlamydomonas ciliary assembly and diminishes once cilia reach their full length (Craft et al., 2015; Wren et al., 2013). Similar to the axonemal building blocks, the transport frequency of ARMC2 and IDA3 is increased in short, growing cilia of Chlamydomonas (Hunter et al., 2018; Lechtreck et al., 2022). Relative to the IFT proteins, ARMC2, IDA3 and ODA16 are enriched in growing cilia, suggesting that IFT particle–adapter binding is regulated. Whereas cargoes require the adapters to move by IFT, IFT of ARMC2 and IDA3, as well as ciliary entry of ODA16, appear to be largely cargo independent (Box 2) (Ahmed and Mitchell, 2005; Hunter et al., 2018; Lechtreck et al., 2022). In cells possessing one growing and one full-length cilium, IFT of tubulin, other axonemal building blocks, and the ARMC2 and IDA3 adapters is increased specifically in the short cilium, indicating that the transport frequency is somehow regulated by ciliary length (Craft et al., 2015; Hunter et al., 2018; Lechtreck et al., 2022). The transport rates of ODAs, radial spokes and the IDA I1/f adapter IDA3 are not increased in full-length zygotic cilia lacking the respective structures (Dai et al., 2018; Hunter et al., 2018; Lechtreck et al., 2018; Wren et al., 2013). These findings indicate that the IFT system does not respond to ciliary paralysis and specific structural defects by increasing transport of the missing component. Thus, the IFT frequency of adapters for axonemal proteins is chiefly regulated in response to ciliary length. Taken together, the data suggest that the regulation of IFT particle–adapter binding critically controls the amount of ODAs, IDAs and radial spoke precursors entering cilia. How IFT particle–adapter binding is regulated remains unknown.
For tubulin, a mechanism regulating binding to IFT particles has been suggested. The N-terminal region of IFT-74, which is part of the putative IFT-74–IFT-81 tubulin-binding module, is phosphorylated by the C. elegans kinase DYF-5 in vitro, reducing its affinity for tubulin (Jiang et al., 2022). However, whether IFT of tubulin is indeed regulated by phosphorylation and altered in dyf-5 mutants it is not known yet.
Regulation of BBSome–IFT particle and BBSome–cargo interactions
Several RAB-like and ARL GTPases regulate the binding of the BBSome to IFT particles and cargoes (Fig. 4) (Yan and Shen, 2021). In Chlamydomonas, IFT22 (also known as RABL5, or as FAP9 in Chlamydomonas) and ARL6 (also known as BBS3), each in their GTP-bound form, recruit BBSomes to the ciliary base, passing them to IFT trains for ciliary entry. However, IFT of the BBSome in cilia is independent of ARL6 and IFT22 (Xue et al., 2020). In mammals and Chlamydomonas, retrograde IFT of BBSomes requires the IFT-B proteins IFT25 and the small GTPase IFT27 (Dong et al., 2017; Eguether et al., 2014; Liew et al., 2014). RABL2, which binds to IFT-B and functions as a pacemaker for train entry, also acts as a ciliary export factor for the BBSome by allowing cargo-carrying BBSomes to bind to IFT-B once RABL2 hydrolyzes its bound GTP and dissociates from the IFT train (Duan et al., 2021; Kanie et al., 2017; Nishijima et al., 2017).
Fig. 4.

Regulation of IFT particle–BBSome and BBSome–cargo interactions by small GTPases. (1) IFT22 and ARL6 stabilize each other in the cell body and recruit the BBSome (BBS) to the ciliary base for loading onto IFT trains (IFT). (2) The passage of the IFT trains through the transition zone (TZ) into the cilium is regulated by RABL2. (3) During retrograde IFT, IFT27 and IFT25 provide a docking site for the BBSome. (4) ARL6 recruits BBSomes to the ciliary membrane and activates BBSomes for cargo binding. In Chlamydomonas, BBSome-dependent export also depends on ARL13 and ARL3. (5) Exit of loaded BBSomes from cilia involves RABL2.
In addition to BBSome–IFT particle interactions, BBSome–cargo interactions are subject to a complex regulation, which is not yet fully understood. In the GTP-bound state, ARL6 recruits the BBSome to membranes via binding to the BBS1 subunit and stabilizes the BBSome in an open configuration for cargo pick-up (Fig. 4) (Jin et al., 2010; Mourao et al., 2014; Yang et al., 2020). Similar to subunits of the IFT-A complex, some BBSome subunits (such as BBS4 and BBS8) have similarities to coatomer proteins (van Dam et al., 2013), and in vitro, BBSomes are recruited to liposomes in a ARL6-GTP-dependent manner (Jin et al., 2010). It has been postulated that IFT27 exits the IFT trains to function as a guanine-nucleotide-exchange factor (GEF) for ARL6, which then recruits BBSomes to the membrane near the ciliary tip to form a planar BBSome coat, followed by cargo binding, retrograde IFT and ciliary exit (Nachury, 2018; Ye et al., 2018). BBSome coat formation can be triggered by signaling, such as ligand binding to an GPCR, initiating retrieval of activated receptors to the cell body, which is often followed by endocytosis (Pal et al., 2016; Ye et al., 2018).
BBSome-dependent export of PLD from Chlamydomonas cilia also requires ARL6, but it is unknown whether PLD transport is triggered by a signaling event or a constitutive process to minimize PLD presence in cilia (Liu and Lechtreck, 2018; Liu et al., 2021). Imaging data further suggest that individual BBSome complexes, rather than a coat-like cluster, transport PLD, suggesting that Chlamydomonas possesses a somewhat simpler BBSome system than that described above for mammalian cells (Liu and Lechtreck, 2018). BBSome-dependent IFT of PLD is abolished in Chlamydomonas arl13 mutants, and PLD accumulates in arl13 and arl3 mutant cilia (Dai et al., 2022; Liu et al., 2022), suggesting that these two small GTPases are required for cargo transport by BBSome-dependent IFT in Chlamydomonas. In primary cilia, ARL13B and ARL3 are widely known for their role in the import of lipidated proteins into cilia, but the contribution of IFT to this process remains unknown (Gotthardt et al., 2015). To summarize, BBSome-mediated IFT and BBSome–cargo interactions are regulated by several small ARL and RAB-like GTPases. Which cellular processes impinge on the BBSome pathway to regulate the presence of signaling proteins in cilia remains largely unknown.
Conclusions and future directions
In many biological processes, core machineries depend on a set of adapters to fulfill their function. Examples of this include cytoplasmic dynein, which in addition to the dynactin complex, employs numerous adapters that activate the motor and link it to a variety of cargoes, or F-box proteins that recruit substrates to the SCF ubiquitin ligase, increasing its target repertoire (Kipreos and Pagano, 2000; Reck-Peterson et al., 2018). Similarly, IFT employs adapters to move cargoes, which by themselves are unable to bind to IFT trains. IFT has a wide cargo spectrum that includes axonemal and membrane proteins, and the known adapters are structurally heterogenous, which likely reflects the heterogeneity of the cargoes. Besides extending the cargo range of the trains, the adapters likely contribute to regulating cargo quantity on the trains. Remaining questions to be addressed include elucidating where on IFT trains the cargoes and adapters are located, how adapters interact with IFT particles and the cargoes, how these interactions are regulated during loading and unloading, and whether other transports also involve adapters. The answers will pave the way for a deeper understanding how ciliary composition is established, maintained and adjusted by IFT.
Acknowledgements
I thank Win Sale (Emory University, GA, USA), Esben Lorentzen (Aarhus University, Denmark) and Andrew Jarman (University of Edinburgh, UK) for discussion of the manuscript.
Footnotes
Funding
Research on ciliary protein transport in my laboratory is supported by a grant from the National Institutes of Health (R01GM110413). The content is solely the responsibility of the author and does not necessarily represent the official views of the National Institutes of Health. Deposited in PMC for release after 12 months.
References
- Ahmed, N. T. and Mitchell, D. R. (2005). ODA16p, a Chlamydomonas flagellar protein needed for dynein assembly. Mol. Biol. Cell 16, 5004-5012. 10.1091/mbc.e05-07-0627 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ahmed, N. T., Gao, C., Lucker, B. F., Cole, D. G. and Mitchell, D. R. (2008). ODA16 aids axonemal outer row dynein assembly through an interaction with the intraflagellar transport machinery. J. Cell Biol. 183, 313-322. 10.1083/jcb.200802025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alford, L. M., Mattheyses, A. L., Hunter, E. L., Lin, H., Dutcher, S. K. and Sale, W. S. (2013). The Chlamydomonas mutant pf27 reveals novel features of ciliary radial spoke assembly. Cytoskeleton (Hoboken) 70, 804-818. 10.1002/cm.21144 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Badgandi, H. B., Hwang, S. H., Shimada, I. S., Loriot, E. and Mukhopadhyay, S. (2017). Tubby family proteins are adapters for ciliary trafficking of integral membrane proteins. J. Cell Biol. 216, 743-760. 10.1083/jcb.201607095 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bah, A. and Forman-Kay, J. D. (2016). Modulation of intrinsically disordered protein function by post-translational modifications. J. Biol. Chem. 291, 6696-6705. 10.1074/jbc.R115.695056 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belzile, O., Hernandez-Lara, C. I., Wang, Q. and Snell, W. J. (2013). Regulated membrane protein entry into flagella is facilitated by cytoplasmic microtubules and does not require IFT. Curr. Biol. 23, 1460-1465. 10.1016/j.cub.2013.06.025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berbari, N. F., Johnson, A. D., Lewis, J. S., Askwith, C. C. and Mykytyn, K. (2008a). Identification of ciliary localization sequences within the third intracellular loop of G protein-coupled receptors. Mol. Biol. Cell 19, 1540-1547. 10.1091/mbc.E07-09-0942 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berbari, N. F., Lewis, J. S., Bishop, G. A., Askwith, C. C. and Mykytyn, K. (2008b). Bardet-Biedl syndrome proteins are required for the localization of G protein-coupled receptors to primary cilia. Proc. Natl. Acad. Sci. USA 105, 4242-4246. 10.1073/pnas.0711027105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhogaraju, S., Cajanek, L., Fort, C., Blisnick, T., Weber, K., Taschner, M., Mizuno, N., Lamla, S., Bastin, P., Nigg, E. A.et al. (2013a). Molecular basis of tubulin transport within the cilium by IFT74 and IFT81. Science 341, 1009-1012. 10.1126/science.1240985 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhogaraju, S., Engel, B. D. and Lorentzen, E. (2013b). Intraflagellar transport complex structure and cargo interactions. Cilia 2, 10. 10.1186/2046-2530-2-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blacque, O. E., Reardon, M. J., Li, C., McCarthy, J., Mahjoub, M. R., Ansley, S. J., Badano, J. L., Mah, A. K., Beales, P. L., Davidson, W. S.et al. (2004). Loss of C. elegans BBS-7 and BBS-8 protein function results in cilia defects and compromised intraflagellar transport. Genes Dev. 18, 1630-1642. 10.1101/gad.1194004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boldt, K., Mans, D. A., Won, J., van Reeuwijk, J., Vogt, A., Kinkl, N., Letteboer, S. J., Hicks, W. L., Hurd, R. E., Naggert, J. K.et al. (2011). Disruption of intraflagellar protein transport in photoreceptor cilia causes Leber congenital amaurosis in humans and mice. J. Clin. Invest. 121, 2169-2180. 10.1172/JCI45627 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brear, A. G., Yoon, J., Wojtyniak, M. and Sengupta, P. (2014). Diverse cell type-specific mechanisms localize G protein-coupled receptors to Caenorhabditis elegans sensory cilia. Genetics 197, 667-684. 10.1534/genetics.114.161349 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown, J. M. and Witman, G. B. (2014). Cilia and diseases. Bioscience 64, 1126-1137. 10.1093/biosci/biu174 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calvert, P. D., Strissel, K. J., Schiesser, W. E., Pugh, E. N., Jr. and Arshavsky, V. Y. (2006). Light-driven translocation of signaling proteins in vertebrate photoreceptors. Trends Cell Biol. 16, 560-568. 10.1016/j.tcb.2006.09.001 [DOI] [PubMed] [Google Scholar]
- Cao, M., Ning, J., Hernandez-Lara, C. I., Belzile, O., Wang, Q., Dutcher, S. K., Liu, Y. and Snell, W. J. (2015). Uni-directional ciliary membrane protein trafficking by a cytoplasmic retrograde IFT motor and ciliary ectosome shedding. Elife 4, e05242. 10.7554/eLife.05242 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cevik, S., Hori, Y., Kaplan, O. I., Kida, K., Toivenon, T., Foley-Fisher, C., Cottell, D., Katada, T., Kontani, K. and Blacque, O. E. (2010). Joubert syndrome Arl13b functions at ciliary membranes and stabilizes protein transport in Caenorhabditis elegans. J. Cell Biol. 188, 953-969. 10.1083/jcb.200908133 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cole, D. G., Diener, D. R., Himelblau, A. L., Beech, P. L., Fuster, J. C. and Rosenbaum, J. L. (1998). Chlamydomonas kinesin-II-dependent intraflagellar transport (IFT): IFT particles contain proteins required for ciliary assembly in Caenorhabditis elegans sensory neurons. J. Cell Biol. 141, 993-1008. 10.1083/jcb.141.4.993 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coutton, C., Martinez, G., Kherraf, Z. E., Amiri-Yekta, A., Boguenet, M., Saut, A., He, X., Zhang, F., Cristou-Kent, M., Escoffier, J.et al. (2019). Bi-allelic mutations in ARMC2 lead to severe astheno-teratozoospermia due to sperm flagellum malformations in humans and mice. Am. J. Hum. Genet. 104, 331-340. 10.1016/j.ajhg.2018.12.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Craft, J. M., Harris, J. A., Hyman, S., Kner, P. and Lechtreck, K. F. (2015). Tubulin transport by IFT is upregulated during ciliary growth by a cilium-autonomous mechanism. J. Cell Biol. 208, 223-237. 10.1083/jcb.201409036 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Craft Van De Weghe, J., Harris, J. A., Kubo, T., Witman, G. B. and Lechtreck, K. F. (2020). Diffusion rather than intraflagellar transport likely provides most of the tubulin required for axonemal assembly in Chlamydomonas. J. Cell Sci. 133, jcs249805. 10.1242/jcs.249805 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dai, J., Barbieri, F., Mitchell, D. R. and Lechtreck, K. F. (2018). In vivo analysis of outer arm dynein transport reveals cargo-specific intraflagellar transport properties. Mol. Biol. Cell 29, 2553-2565. 10.1091/mbc.E18-05-0291 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dai, J., Zhang, G., Alkhofash, R. A., Mekonnen, B., Saravanan, S., Xue, B., Fan, Z. C., Betleja, E., Cole, D. G., Liu, P.et al. (2022). Loss of ARL13 impedes BBSome-dependent cargo export from Chlamydomonas cilia. J. Cell Biol. 221, e202201050. 10.1083/jcb.202201050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dean, A. B. and Mitchell, D. R. (2015). Late steps in cytoplasmic maturation of assembly-competent axonemal outer arm dynein in Chlamydomonas require interaction of ODA5 and ODA10 in a complex. Mol. Biol. Cell 26, 3596-3605. 10.1091/mbc.E15-05-0317 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dentler, W. (2013). A role for the membrane in regulating Chlamydomonas flagellar length. PLoS One 8, e53366. 10.1371/journal.pone.0053366 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desai, P. B., Freshour, J. R. and Mitchell, D. R. (2015). Chlamydomonas axonemal dynein assembly locus ODA8 encodes a conserved flagellar protein needed for cytoplasmic maturation of outer dynein arm complexes. Cytoskeleton (Hoboken) 72, 16-28. 10.1002/cm.21206 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desai, P. B., Stuck, M. W., Lv, B. and Pazour, G. J. (2020). Ubiquitin links smoothened to intraflagellar transport to regulate Hedgehog signaling. J. Cell Biol. 219, e201912104. 10.1083/jcb.201912104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- DiTirro, D., Philbrook, A., Rubino, K. and Sengupta, P. (2019). The Caenorhabditis elegans Tubby homolog dynamically modulates olfactory cilia membrane morphogenesis and phospholipid composition. Elife 8, e48789. 10.7554/eLife.48789 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Domire, J. S., Green, J. A., Lee, K. G., Johnson, A. D., Askwith, C. C. and Mykytyn, K. (2011). Dopamine receptor 1 localizes to neuronal cilia in a dynamic process that requires the Bardet-Biedl syndrome proteins. Cell. Mol. Life Sci. 68, 2951-2960. 10.1007/s00018-010-0603-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dong, B., Wu, S., Wang, J., Liu, Y. X., Peng, Z., Meng, D. M., Huang, K., Wu, M. and Fan, Z. C. (2017). Chlamydomonas IFT25 is dispensable for flagellar assembly but required to export the BBSome from flagella. Biol. Open 6, 1680-1691. 10.1242/bio.026278 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duan, S., Li, H., Zhang, Y., Yang, S., Chen, Y., Qiu, B., Huang, C., Wang, J., Li, J., Zhu, X.et al. (2021). Rabl2 GTP hydrolysis licenses BBSome-mediated export to fine-tune ciliary signaling. EMBO J. 40, e105499. 10.15252/embj.2020105499 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dyson, H. J. and Wright, P. E. (2005). Intrinsically unstructured proteins and their functions. Nat. Rev. Mol. Cell Biol. 6, 197-208. 10.1038/nrm1589 [DOI] [PubMed] [Google Scholar]
- Eguether, T., San Agustin, J. T., Keady, B. T., Jonassen, J. A., Liang, Y., Francis, R., Tobita, K., Johnson, C. A., Abdelhamed, Z. A., Lo, C. W.et al. (2014). IFT27 links the BBSome to IFT for maintenance of the ciliary signaling compartment. Dev. Cell 31, 279-290. 10.1016/j.devcel.2014.09.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erdos, G. and Dosztanyi, Z. (2020). Analyzing protein disorder with IUPred2A. Curr. Protoc. Bioinformatics 70, e99. 10.1002/cpbi.99 [DOI] [PubMed] [Google Scholar]
- Fingerhut, J. M. and Yamashita, Y. M. (2020). mRNA localization mediates maturation of cytoplasmic cilia in Drosophila spermatogenesis. J. Cell Biol. 219, e202003084. 10.1083/jcb.202003084 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fliegauf, M., Benzing, T. and Omran, H. (2007). When cilia go bad: cilia defects and ciliopathies. Nat. Rev. Mol. Cell Biol. 8, 880-893. 10.1038/nrm2278 [DOI] [PubMed] [Google Scholar]
- Fu, W., Wang, L., Kim, S., Li, J. and Dynlacht, B. D. (2016). Role for the IFT-A complex in selective transport to the primary cilium. Cell Rep. 17, 1505-1517. 10.1016/j.celrep.2016.10.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao, C., Wang, G., Amack, J. D. and Mitchell, D. R. (2010). Oda16/Wdr69 is essential for axonemal dynein assembly and ciliary motility during zebrafish embryogenesis. Dev. Dyn. 239, 2190-2197. 10.1002/dvdy.22355 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia-Gonzalo, F. R. and Reiter, J. F. (2012). Scoring a backstage pass: mechanisms of ciliogenesis and ciliary access. J. Cell Biol. 197, 697-709. 10.1083/jcb.201111146 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gotthardt, K., Lokaj, M., Koerner, C., Falk, N., Giessl, A. and Wittinghofer, A. (2015). A G-protein activation cascade from Arl13B to Arl3 and implications for ciliary targeting of lipidated proteins. Elife 4, e11859. 10.7554/eLife.11859 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gui, M., Ma, M., Sze-Tu, E., Wang, X., Koh, F., Zhong, E. D., Berger, B., Davis, J. H., Dutcher, S. K., Zhang, R.et al. (2021). Structures of radial spokes and associated complexes important for ciliary motility. Nat. Struct. Mol. Biol. 28, 29-37. 10.1038/s41594-020-00530-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gupta, A., Diener, D. R., Sivadas, P., Rosenbaum, J. L. and Yang, P. (2012). The versatile molecular complex component LC8 promotes several distinct steps of flagellar assembly. J. Cell Biol. 198, 115-126. 10.1083/jcb.201111041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han, S., Miyoshi, K., Shikada, S., Amano, G., Wang, Y., Yoshimura, T. and Katayama, T. (2019). TULP3 is required for localization of membrane-associated proteins ARL13B and INPP5E to primary cilia. Biochem. Biophys. Res. Commun. 509, 227-234. 10.1016/j.bbrc.2018.12.109 [DOI] [PubMed] [Google Scholar]
- Hao, L., Thein, M., Brust-Mascher, I., Civelekoglu-Scholey, G., Lu, Y., Acar, S., Prevo, B., Shaham, S. and Scholey, J. M. (2011). Intraflagellar transport delivers tubulin isotypes to sensory cilium middle and distal segments. Nat. Cell Biol. 13, 790-798. 10.1038/ncb2268 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harris, J. A., Liu, Y., Yang, P., Kner, P. and Lechtreck, K. F. (2016). Single-particle imaging reveals intraflagellar transport-independent transport and accumulation of EB1 in Chlamydomonas flagella. Mol. Biol. Cell 27, 295-307. 10.1091/mbc.E15-08-0608 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hesketh, S. J., Mukhopadhyay, A. G., Nakamura, D., Toropova, K. and Roberts, A. J. (2022). IFT-A structure reveals carriages for membrane protein transport into cilia. bioRxiv, 2022.08.09.503213. 10.1101/2022.08.09.503213 [DOI] [PubMed] [Google Scholar]
- Hirano, T., Katoh, Y. and Nakayama, K. (2017). Intraflagellar transport-A complex mediates ciliary entry and retrograde trafficking of ciliary G protein-coupled receptors. Mol. Biol. Cell 28, 429-439. 10.1091/mbc.e16-11-0813 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horani, A., Ustione, A., Huang, T., Firth, A. L., Pan, J., Gunsten, S. P., Haspel, J. A., Piston, D. W. and Brody, S. L. (2018). Establishment of the early cilia preassembly protein complex during motile ciliogenesis. Proc. Natl. Acad. Sci. USA 115, E1221-E1228. 10.1073/pnas.1715915115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hou, Y. and Witman, G. B. (2017). The N-terminus of IFT46 mediates intraflagellar transport of outer arm dynein and its cargo-adaptor ODA16. Mol. Biol. Cell 28, 2420-2433. 10.1091/mbc.e17-03-0172 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hou, Y., Qin, H., Follit, J. A., Pazour, G. J., Rosenbaum, J. L. and Witman, G. B. (2007). Functional analysis of an individual IFT protein: IFT46 is required for transport of outer dynein arms into flagella. J. Cell Biol. 176, 653-665. 10.1083/jcb.200608041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang, B., Piperno, G., Ramanis, Z. and Luck, D. J. (1981). Radial spokes of Chlamydomonas flagella: genetic analysis of assembly and function. J. Cell Biol. 88, 80-88. 10.1083/jcb.88.1.80 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang, K., Diener, D. R., Mitchell, A., Pazour, G. J., Witman, G. B. and Rosenbaum, J. L. (2007). Function and dynamics of PKD2 in Chlamydomonas reinhardtii flagella. J. Cell Biol. 179, 501-514. 10.1083/jcb.200704069 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huet, D., Blisnick, T., Perrot, S. and Bastin, P. (2014). The GTPase IFT27 is involved in both anterograde and retrograde intraflagellar transport. Elife 3, e02419. 10.7554/eLife.02419 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huet, D., Blisnick, T., Perrot, S. and Bastin, P. (2019). IFT25 is required for the construction of the trypanosome flagellum. J. Cell Sci. 132, jcs228296. 10.1242/jcs.228296 [DOI] [PubMed] [Google Scholar]
- Hunter, E. L., Lechtreck, K., Fu, G., Hwang, J., Lin, H., Gokhale, A., Alford, L. M., Lewis, B., Yamamoto, R., Kamiya, R.et al. (2018). The IDA3 adapter, required for intraflagellar transport of I1 dynein, is regulated by ciliary length. Mol. Biol. Cell 29, 886-896. 10.1091/mbc.E17-12-0729 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hwang, S. H., Somatilaka, B. N., Badgandi, H., Palicharla, V. R., Walker, R., Shelton, J. M., Qian, F. and Mukhopadhyay, S. (2019). Tulp3 regulates renal cystogenesis by trafficking of cystoproteins to cilia. Curr. Biol. 29, 790-802.e5. 10.1016/j.cub.2019.01.047 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iakoucheva, L. M., Radivojac, P., Brown, C. J., O'Connor, T. R., Sikes, J. G., Obradovic, Z. and Dunker, A. K. (2004). The importance of intrinsic disorder for protein phosphorylation. Nucleic Acids Res. 32, 1037-1049. 10.1093/nar/gkh253 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ishikawa, H., Ide, T., Yagi, T., Jiang, X., Hirono, M., Sasaki, H., Yanagisawa, H., Wemmer, K. A., Stainier, D. Y., Qin, H.et al. (2014). TTC26/DYF13 is an intraflagellar transport protein required for transport of motility-related proteins into flagella. Elife 3, e01566. 10.7554/eLife.01566 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang, X., Shao, W., Chai, Y., Huang, J., Mohamed, M. A. A., Okten, Z., Li, W., Zhu, Z. and Ou, G. (2022). DYF-5/MAK-dependent phosphorylation promotes ciliary tubulin unloading. Proc. Natl. Acad. Sci. USA 119, e2207134119. 10.1073/pnas.2207134119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jin, H., White, S. R., Shida, T., Schulz, S., Aguiar, M., Gygi, S. P., Bazan, J. F. and Nachury, M. V. (2010). The conserved Bardet-Biedl syndrome proteins assemble a coat that traffics membrane proteins to cilia. Cell 141, 1208-1219. 10.1016/j.cell.2010.05.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jordan, M. A., Diener, D. R., Stepanek, L. and Pigino, G. (2018). The cryo-EM structure of intraflagellar transport trains reveals how dynein is inactivated to ensure unidirectional anterograde movement in cilia. Nat. Cell Biol. 20, 1250-1255. 10.1038/s41556-018-0213-1 [DOI] [PubMed] [Google Scholar]
- Kamiya, R., Kurimoto, E. and Muto, E. (1991). Two types of Chlamydomonas flagellar mutants missing different components of inner-arm dynein. J. Cell Biol. 112, 441-447. 10.1083/jcb.112.3.441 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanie, T., Abbott, K. L., Mooney, N. A., Plowey, E. D., Demeter, J. and Jackson, P. K. (2017). The CEP19-RABL2 GTPase complex binds IFT-B to initiate intraflagellar transport at the ciliary base. Dev. Cell 42, 22-36.e12. 10.1016/j.devcel.2017.05.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katoh, Y., Nozaki, S., Hartanto, D., Miyano, R. and Nakayama, K. (2015). Architectures of multisubunit complexes revealed by a visible immunoprecipitation assay using fluorescent fusion proteins. J. Cell Sci. 128, 2351-2362. 10.1242/jcs.168740 [DOI] [PubMed] [Google Scholar]
- Kee, H. L., Dishinger, J. F., Blasius, T. L., Liu, C. J., Margolis, B. and Verhey, K. J. (2012). A size-exclusion permeability barrier and nucleoporins characterize a ciliary pore complex that regulates transport into cilia. Nat. Cell Biol. 14, 431-437. 10.1038/ncb2450 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kipreos, E. T. and Pagano, M. (2000). The F-box protein family. Genome Biol. 1, REVIEWS3002. 10.1186/gb-2000-1-5-reviews3002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kleyn, P. W., Fan, W., Kovats, S. G., Lee, J. J., Pulido, J. C., Wu, Y., Berkemeier, L. R., Misumi, D. J., Holmgren, L., Charlat, O.et al. (1996). Identification and characterization of the mouse obesity gene tubby: a member of a novel gene family. Cell 85, 281-290. 10.1016/S0092-8674(00)81104-6 [DOI] [PubMed] [Google Scholar]
- Klink, B. U., Zent, E., Juneja, P., Kuhlee, A., Raunser, S. and Wittinghofer, A. (2017). A recombinant BBSome core complex and how it interacts with ciliary cargo. Elife 6, e27434. 10.7554/eLife.27434 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kobayashi, T., Ishida, Y., Hirano, T., Katoh, Y. and Nakayama, K. (2021). Cooperation of the IFT-A complex with the IFT-B complex is required for ciliary retrograde protein trafficking and GPCR import. Mol. Biol. Cell 32, 45-56. 10.1091/mbc.E20-08-0556 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kozminski, K. G., Johnson, K. A., Forscher, P. and Rosenbaum, J. L. (1993). A motility in the eukaryotic flagellum unrelated to flagellar beating. Proc. Natl. Acad. Sci. USA 90, 5519-5523. 10.1073/pnas.90.12.5519 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kubo, T., Brown, J. M., Bellve, K., Craige, B., Craft, J. M., Fogarty, K., Lechtreck, K. F. and Witman, G. B. (2016). Together, the IFT81 and IFT74 N-termini together form the major module for intraflagellar transport of tubulin. J. Cell Sci. 129, 2019-2106. 10.1242/jcs.187120 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lacey, S. E., Foster, H. E. and Pigino, G. (2022). The Molecular Structure of Anterograde Intraflagellar transport trains. bioRxiv, 2022.08.01.502329. 10.1101/2022.08.01.502329 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lechtreck, K. F. (2015). IFT-Cargo Interactions and Protein Transport in Cilia. Trends Biochem. Sci. 40, 765-778. 10.1016/j.tibs.2015.09.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lechtreck, K. F., Johnson, E. C., Sakai, T., Cochran, D., Ballif, B. A., Rush, J., Pazour, G. J., Ikebe, M. and Witman, G. B. (2009). The Chlamydomonas reinhardtii BBSome is an IFT cargo required for export of specific signaling proteins from flagella. J. Cell Biol. 187, 1117-1132. 10.1083/jcb.200909183 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lechtreck, K. F., Liu, Y., Dai, J., Alkhofash, R. A., Butler, J., Alford, L. and Yang, P. (2022). Chlamydomonas ARMC2/PF27 is an obligate cargo adapter for intraflagellar transport of radial spokes. Elife 11, e74993. 10.7554/eLife.74993 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lechtreck, K. F., Mengoni, I., Okivie, B. and Hilderhoff, K. B. (2018). In vivo analyses of radial spoke transport, assembly, repair and maintenance. Cytoskeleton (Hoboken) 75, 352-362. 10.1002/cm.21457 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee, C., Cox, R. M., Papoulas, O., Horani, A., Drew, K., Devitt, C. C., Brody, S. L., Marcotte, E. M. and Wallingford, J. B. (2020). Functional partitioning of a liquid-like organelle during assembly of axonemal dyneins. Elife 9, e58662. 10.7554/eLife.58662 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Legué, E. and Liem, K. F., Jr. (2019). Tulp3 is a ciliary trafficking gene that regulates polycystic kidney disease. Curr. Biol. 29, 803-812.e5. 10.1016/j.cub.2019.01.054 [DOI] [PubMed] [Google Scholar]
- Li, J. B., Gerdes, J. M., Haycraft, C. J., Fan, Y., Teslovich, T. M., May-Simera, H., Li, H., Blacque, O. E., Li, L., Leitch, C. C.et al. (2004). Comparative genomics identifies a flagellar and basal body proteome that includes the BBS5 human disease gene. Cell 117, 541-552. 10.1016/S0092-8674(04)00450-7 [DOI] [PubMed] [Google Scholar]
- Liew, G. M., Ye, F., Nager, A. R., Murphy, J. P., Lee, J. S., Aguiar, M., Breslow, D. K., Gygi, S. P. and Nachury, M. V. (2014). The intraflagellar transport protein IFT27 promotes BBSome exit from cilia through the GTPase ARL6/BBS3. Dev. Cell 31, 265-278. 10.1016/j.devcel.2014.09.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu, P. and Lechtreck, K. F. (2018). The Bardet-Biedl syndrome protein complex is an adapter expanding the cargo range of intraflagellar transport trains for ciliary export. Proc. Natl. Acad. Sci. USA 115, E934-E943. 10.1073/pnas.1713226115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu, H., Li, W., Zhang, Y., Zhang, Z., Shang, X., Zhang, L., Zhang, S., Li, Y., Somoza, A. V., Delpi, B.et al. (2017). IFT25, an intraflagellar transporter protein dispensable for ciliogenesis in somatic cells, is essential for sperm flagella formation. Biol. Reprod. 96, 993-1006. 10.1093/biolre/iox029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu, Y. X., Xue, B., Sun, W. Y., Wingfield, J. L., Sun, J., Wu, M., Lechtreck, K. F., Wu, Z. and Fan, Z. C. (2021). Bardet-Biedl Syndrome 3 protein promotes ciliary exit of the signaling protein phospholipase D via the BBSome. Elife 10, e59119. 10.7554/eLife.59119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu, Y. X., Sun, W. Y., Xue, B., Zhang, R. K., Li, W. J., Xie, X. and Fan, Z. C. (2022). ARL3 mediates BBSome ciliary turnover by promoting its outward movement across the transition zone. J. Cell Biol. 221, e202111076. 10.1083/jcb.202111076 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loktev, A. V. and Jackson, P. K. (2013). Neuropeptide Y family receptors traffic via the Bardet-Biedl syndrome pathway to signal in neuronal primary cilia. Cell Rep 5, 1316-1329. 10.1016/j.celrep.2013.11.011 [DOI] [PubMed] [Google Scholar]
- Long, H. and Huang, K. (2019). Transport of ciliary membrane proteins. Front. Cell Dev. Biol. 7, 381. 10.3389/fcell.2019.00381 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luck, D., Piperno, G., Ramanis, Z. and Huang, B. (1977). Flagellar mutants of Chlamydomonas: studies of radial spoke-defective strains by dikaryon and revertant analysis. Proc. Natl. Acad. Sci. USA 74, 3456-3460. 10.1073/pnas.74.8.3456 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo, W., Ruba, A., Takao, D., Zweifel, L. P., Lim, R. Y. H., Verhey, K. J. and Yang, W. (2017). Axonemal lumen dominates cytosolic protein diffusion inside the primary cilium. Sci. Rep. 7, 15793. 10.1038/s41598-017-16103-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lv, B., Stuck, M. W., Desai, P. B., Cabrera, O. A. and Pazour, G. J. (2021). E3 ubiquitin ligase Wwp1 regulates ciliary dynamics of the Hedgehog receptor Smoothened. J. Cell Biol. 220, e202010177. 10.1083/jcb.202010177 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mali, G. R., Ali, F. A., Lau, C. K., Begum, F., Boulanger, J., Howe, J. D., Chen, Z. A., Rappsilber, J., Skehel, M. and Carter, A. P. (2021). Shulin packages axonemal outer dynein arms for ciliary targeting. Science 371, 910-916. 10.1126/science.abe0526 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCafferty, C. L., Papoulas, O., Jordan, M. A., Hoogerbrugge, G., Nichols, C., Pigino, G., Taylor, D. W., Wallingford, J. B. and Marcotte, E. M. (2022). Integrative modeling reveals the molecular architecture of the intraflagellar transport A (IFT-A) complex. Elife, 11, e81977. 10.7554/eLife.81977 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Merchant, S. S., Prochnik, S. E., Vallon, O., Harris, E. H., Karpowicz, S. J., Witman, G. B., Terry, A., Salamov, A., Fritz-Laylin, L. K., Marechal-Drouard, L.et al. (2007). The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science 318, 245-250. 10.1126/science.1143609 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monis, W. J., Faundez, V. and Pazour, G. J. (2017). BLOC-1 is required for selective membrane protein trafficking from endosomes to primary cilia. J. Cell Biol. 216, 2131-2150. 10.1083/jcb.201611138 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mourao, A., Nager, A. R., Nachury, M. V. and Lorentzen, E. (2014). Structural basis for membrane targeting of the BBSome by ARL6. Nat. Struct. Mol. Biol. 21, 1035-1041. 10.1038/nsmb.2920 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mukhopadhyay, S. and Jackson, P. K. (2011). The tubby family proteins. Genome Biol. 12, 225. 10.1186/gb-2011-12-6-225 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mukhopadhyay, S., Wen, X., Chih, B., Nelson, C. D., Lane, W. S., Scales, S. J. and Jackson, P. K. (2010). TULP3 bridges the IFT-A complex and membrane phosphoinositides to promote trafficking of G protein-coupled receptors into primary cilia. Genes Dev. 24, 2180-2193. 10.1101/gad.1966210 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mykytyn, K., Mullins, R. F., Andrews, M., Chiang, A. P., Swiderski, R. E., Yang, B., Braun, T., Casavant, T., Stone, E. M. and Sheffield, V. C. (2004). Bardet-Biedl syndrome type 4 (BBS4)-null mice implicate Bbs4 in flagella formation but not global cilia assembly. Proc. Natl. Acad. Sci. USA 101, 8664-8669. 10.1073/pnas.0402354101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nachury, M. V. (2018). The molecular machines that traffic signaling receptors into and out of cilia. Curr. Opin. Cell Biol. 51, 124-131. 10.1016/j.ceb.2018.03.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nachury, M. V., Loktev, A. V., Zhang, Q., Westlake, C. J., Peranen, J., Merdes, A., Slusarski, D. C., Scheller, R. H., Bazan, J. F., Sheffield, V. C.et al. (2007). A core complex of BBS proteins cooperates with the GTPase Rab8 to promote ciliary membrane biogenesis. Cell 129, 1201-1213. 10.1016/j.cell.2007.03.053 [DOI] [PubMed] [Google Scholar]
- Nishijima, Y., Hagiya, Y., Kubo, T., Takei, R., Katoh, Y. and Nakayama, K. (2017). RABL2 interacts with the intraflagellar transport-B complex and CEP19 and participates in ciliary assembly. Mol. Biol. Cell 28, 1652-1666. 10.1091/mbc.e17-01-0017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishimura, D. Y., Fath, M., Mullins, R. F., Searby, C., Andrews, M., Davis, R., Andorf, J. L., Mykytyn, K., Swiderski, R. E., Yang, B.et al. (2004). Bbs2-null mice have neurosensory deficits, a defect in social dominance, and retinopathy associated with mislocalization of rhodopsin. Proc. Natl. Acad. Sci. USA 101, 16588-16593. 10.1073/pnas.0405496101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ojeda Naharros, I. and Nachury, M. V. (2022). Shedding of ciliary vesicles at a glance. J. Cell Sci. 135, jcs246553. 10.1242/jcs.246553 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ou, G., Blacque, O. E., Snow, J. J., Leroux, M. R. and Scholey, J. M. (2005). Functional coordination of intraflagellar transport motors. Nature 436, 583-587. 10.1038/nature03818 [DOI] [PubMed] [Google Scholar]
- Pal, K., Hwang, S. H., Somatilaka, B., Badgandi, H., Jackson, P. K., DeFea, K. and Mukhopadhyay, S. (2016). Smoothened determines beta-arrestin-mediated removal of the G protein-coupled receptor Gpr161 from the primary cilium. J. Cell Biol. 212, 861-875. 10.1083/jcb.201506132 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pazour, G. J., Dickert, B. L. and Witman, G. B. (1999). The DHC1b (DHC2) isoform of cytoplasmic dynein is required for flagellar assembly. J. Cell Biol. 144, 473-481. 10.1083/jcb.144.3.473 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pazour, G. J., Agrin, N., Leszyk, J. and Witman, G. B. (2005). Proteomic analysis of a eukaryotic cilium. J. Cell Biol. 170, 103-113. 10.1083/jcb.200504008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petriman, N. A., Loureiro-López, M., Taschner, M., Zacharia, N. K., Georgieva, M. M., Boegholm, N., Wang, J., Mourão, A., Russell, R. B., Andersen, J. S.et al. (2022). Biochemically validated structural model of the 15-subunit intraflagellar transport complex IFT-B. EMBO J. e112440. 10.15252/embj.2022112440 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Picariello, T., Brown, J. M., Hou, Y., Swank, G., Cochran, D. A., King, O. D., Lechtreck, K., Pazour, G. J. and Witman, G. B. (2019). A global analysis of IFT-A function reveals specialization for transport of membrane-associated proteins into cilia. J. Cell Sci. 132, jcs220749. 10.1242/jcs.220749 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piperno, G. and Mead, K. (1997). Transport of a novel complex in the cytoplasmic matrix of Chlamydomonas flagella. Proc. Natl. Acad. Sci. USA 94, 4457-4462. 10.1073/pnas.94.9.4457 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin, H., Burnette, D. T., Bae, Y. K., Forscher, P., Barr, M. M. and Rosenbaum, J. L. (2005). Intraflagellar transport is required for the vectorial movement of TRPV channels in the ciliary membrane. Curr. Biol. 15, 1695-1699. 10.1016/j.cub.2005.08.047 [DOI] [PubMed] [Google Scholar]
- Quidwai, T., Wang, J., Hall, E. A., Petriman, N. A., Leng, W., Kiesel, P., Wells, J. N., Murphy, L. C., Keighren, M. A., Marsh, J. A.et al. (2021). A WDR35-dependent coat protein complex transports ciliary membrane cargo vesicles to cilia. Elife 10, e69786. 10.7554/eLife.69786 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reck-Peterson, S. L., Redwine, W. B., Vale, R. D. and Carter, A. P. (2018). The cytoplasmic dynein transport machinery and its many cargoes. Nat. Rev. Mol. Cell Biol. 19, 382-398. 10.1038/s41580-018-0004-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenbaum, J. L. and Child, F. M. (1967). Flagellar regeneration in protozoan flagellates. J. Cell Biol. 34, 345-364. 10.1083/jcb.34.1.345 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roy, K., Jerman, S., Jozsef, L., McNamara, T., Onyekaba, G., Sun, Z. and Marin, E. P. (2017). Palmitoylation of the ciliary GTPase ARL13b is necessary for its stability and its role in cilia formation. J. Biol. Chem. 292, 17703-17717. 10.1074/jbc.M117.792937 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santagata, S., Boggon, T. J., Baird, C. L., Gomez, C. A., Zhao, J., Shan, W. S., Myszka, D. G. and Shapiro, L. (2001). G-protein signaling through tubby proteins. Science 292, 2041-2050. 10.1126/science.1061233 [DOI] [PubMed] [Google Scholar]
- Seo, S., Zhang, Q., Bugge, K., Breslow, D. K., Searby, C. C., Nachury, M. V. and Sheffield, V. C. (2011). A novel protein LZTFL1 regulates ciliary trafficking of the BBSome and Smoothened. PLoS Genet. 7, e1002358. 10.1371/journal.pgen.1002358 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheffield, V. C. (2010). The blind leading the obese: the molecular pathophysiology of a human obesity syndrome. Trans. Am. Clin. Climatol. Assoc. 121, 172-181; discussion 181-2. [PMC free article] [PubMed] [Google Scholar]
- Shinde, S. R., Mick, D. U., Aoki, E., Rodrigues, R. B., Gygi, S. P. and Nachury, M. V. (2022). The ancestral ESCRT protein TOM1L2 selects ubiquitinated cargoes for retrieval from cilia. bioRxiv, 2022.09.23.509287. 10.1101/2022.09.23.509287 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shinde, S. R., Nager, A. R. and Nachury, M. V. (2020). Ubiquitin chains earmark GPCRs for BBSome-mediated removal from cilia. J. Cell Biol. 219, e202003020. 10.1083/jcb.202003020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh, S. K., Gui, M., Koh, F., Yip, M. C. and Brown, A. (2020). Structure and activation mechanism of the BBSome membrane protein trafficking complex. Elife 9. e53322. 10.7554/eLife.53322 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soler Artigas, M., Loth, M., Wain, D. W., Gharib, L. V., Obeidat, S. A., Tang, M., Zhai, W., Zhao, G., Smith, J. H., Huffman, A. V., E., J.et al. (2011). Genome-wide association and large-scale follow up identifies 16 new loci influencing lung function. Nat. Genet. 43, 1082-1090. 10.1038/ng.941 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stolc, V., Samanta, M. P., Tongprasit, W. and Marshall, W. F. (2005). Genome-wide transcriptional analysis of flagellar regeneration in Chlamydomonas reinhardtii identifies orthologs of ciliary disease genes. Proc. Natl. Acad. Sci. USA 102, 3703-3707. 10.1073/pnas.0408358102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun, W. Y., Xue, B., Liu, Y. X., Zhang, R. K., Li, R. C., Xin, W., Wu, M. and Fan, Z. C. (2021). Chlamydomonas LZTFL1 mediates phototaxis via controlling BBSome recruitment to the basal body and its reassembly at the ciliary tip. Proc. Natl. Acad. Sci. USA 118, e2101590118. 10.1073/pnas.2101590118 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taschner, M., Kotsis, F., Braeuer, P., Kuehn, E. W. and Lorentzen, E. (2014). Crystal structures of IFT70/52 and IFT52/46 provide insight into intraflagellar transport B core complex assembly. J. Cell Biol. 207, 269-282. 10.1083/jcb.201408002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taschner, M., Mourao, A., Awasthi, M., Basquin, J. and Lorentzen, E. (2017). Structural basis of outer dynein arm intraflagellar transport by the transport adaptor protein ODA16 and the intraflagellar transport protein IFT46. J. Biol. Chem. 292, 7462-7473. 10.1074/jbc.M117.780155 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taschner, M., Weber, K., Mourao, A., Vetter, M., Awasthi, M., Stiegler, M., Bhogaraju, S. and Lorentzen, E. (2016). Intraflagellar transport proteins 172, 80, 57, 54, 38, and 20 form a stable tubulin-binding IFT-B2 complex. EMBO J. 35, 773-790. 10.15252/embj.201593164 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toropova, K., Zalyte, R., Mukhopadhyay, A. G., Mladenov, M., Carter, A. P. and Roberts, A. J. (2019). Structure of the dynein-2 complex and its assembly with intraflagellar transport trains. Nat. Struct. Mol. Biol. 26, 823-829. 10.1038/s41594-019-0286-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valentine, M. S., Rajendran, A., Yano, J., Weeraratne, S. D., Beisson, J., Cohen, J., Koll, F. and Van Houten, J. (2012). Paramecium BBS genes are key to presence of channels in Cilia. Cilia 1, 16. 10.1186/2046-2530-1-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Dam, T. J., Townsend, M. J., Turk, M., Schlessinger, A., Sali, A., Field, M. C. and Huynen, M. A. (2013). Evolution of modular intraflagellar transport from a coatomer-like progenitor. Proc. Natl. Acad. Sci. USA 110, 6943-6948. 10.1073/pnas.1221011110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van den Hoek, H., Klena, N., Jordan, M. A., Alvarez Viar, G., Righetto, R. D., Schaffer, M., Erdmann, P. S., Wan, W., Geimer, S., Plitzko, J. M.et al. (2022). In situ architecture of the ciliary base reveals the stepwise assembly of intraflagellar transport trains. Science 377, 543-548. 10.1126/science.abm6704 [DOI] [PubMed] [Google Scholar]
- van der Burght, S. N., Rademakers, S., Johnson, J. L., Li, C., Kremers, G. J., Houtsmuller, A. B., Leroux, M. R. and Jansen, G. (2020). Ciliary Tip Signaling Compartment Is Formed and Maintained by Intraflagellar Transport. Curr. Biol. 30, 4299-4306.e5. 10.1016/j.cub.2020.08.032 [DOI] [PubMed] [Google Scholar]
- van Krugten, J., Danne, N. and Peterman, E. J. G. (2022). A local interplay between diffusion and intraflagellar transport distributes TRPV-channel OCR-2 along C. elegans chemosensory cilia. Commun Biol 5, 720. 10.1038/s42003-022-03683-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Viswanadha, R., Hunter, E. L., Yamamoto, R., Wirschell, M., Alford, L. M., Dutcher, S. K. and Sale, W. S. (2014). The ciliary inner dynein arm, I1 dynein, is assembled in the cytoplasm and transported by IFT before axonemal docking. Cytoskeleton (Hoboken) 71, 573-586. 10.1002/cm.21192 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang, J. and Barr, M. M. (2018). Cell-cell communication via ciliary extracellular vesicles: clues from model systems. Essays Biochem. 62, 205-213. 10.1042/EBC20170085 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang, H., Gau, B., Slade, W. O., Juergens, M., Li, P. and Hicks, L. M. (2014). The global phosphoproteome of Chlamydomonas reinhardtii reveals complex organellar phosphorylation in the flagella and thylakoid membrane. Mol. Cell. Proteomics 13, 2337-2353. 10.1074/mcp.M114.038281 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang, J., Taschner, M., Petriman, N. A., Andersen, M. B., Basquin, J., Bhogaraju, S., Vetter, M., Wachter, S., Lorentzen, A. and Lorentzen, E. (2020). Purification and crystal structure of human ODA16: Implications for ciliary import of outer dynein arms by the intraflagellar transport machinery. Protein Sci. 29, 1502-1510. 10.1002/pro.3864 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams, C. L., McIntyre, J. C., Norris, S. R., Jenkins, P. M., Zhang, L., Pei, Q., Verhey, K. and Martens, J. R. (2014). Direct evidence for BBSome-associated intraflagellar transport reveals distinct properties of native mammalian cilia. Nat. Commun. 5, 5813. 10.1038/ncomms6813 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wingfield, J. L., Mengoni, I., Bomberger, H., Jiang, Y. Y., Walsh, J. D., Brown, J. M., Picariello, T., Cochran, D. A., Zhu, B., Pan, J.et al. (2017). IFT trains in different stages of assembly queue at the ciliary base for consecutive release into the cilium. Elife 6, e26609. 10.7554/eLife.26609 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wingfield, J. L., Lechtreck, K. F. and Lorentzen, E. (2018). Trafficking of ciliary membrane proteins by the intraflagellar transport/BBSome machinery. Essays Biochem. 62, 753-763. 10.1042/EBC20180030 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wingfield, J. L., Mekonnen, B., Mengoni, I., Liu, P., Jordan, M., Diener, D., Pigino, G. and Lechtreck, K. (2021). In vivo imaging shows continued association of several IFT-A, IFT-B and dynein complexes while IFT trains U-turn at the tip. J. Cell Sci. 134, jcs259010. 10.1242/jcs.259010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wirschell, M., Pazour, G., Yoda, A., Hirono, M., Kamiya, R. and Witman, G. B. (2004). Oda5p, a novel axonemal protein required for assembly of the outer dynein arm and an associated adenylate kinase. Mol. Biol. Cell 15, 2729-2741. 10.1091/mbc.e03-11-0820 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wren, K. N., Craft, J. M., Tritschler, D., Schauer, A., Patel, D. K., Smith, E. F., Porter, M. E., Kner, P. and Lechtreck, K. F. (2013). A differential cargo-loading model of ciliary length regulation by IFT. Curr. Biol. 23, 2463-2471. 10.1016/j.cub.2013.10.044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wright, P. E. and Dyson, H. J. (2015). Intrinsically disordered proteins in cellular signalling and regulation. Nat. Rev. Mol. Cell Biol. 16, 18-29. 10.1038/nrm3920 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xue, B., Liu, Y. X., Dong, B., Wingfield, J. L., Wu, M., Sun, J., Lechtreck, K. F. and Fan, Z. C. (2020). Intraflagellar transport protein RABL5/IFT22 recruits the BBSome to the basal body through the GTPase ARL6/BBS3. Proc. Natl. Acad. Sci. USA 117, 2496-2505. 10.1073/pnas.1901665117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan, X. and Shen, Y. (2021). Rab-like small GTPases in the regulation of ciliary Bardet-Biedl syndrome (BBS) complex transport. FEBS J. e16232. 10.1111/febs.16232 [DOI] [PubMed] [Google Scholar]
- Yang, P., Diener, D. R., Yang, C., Kohno, T., Pazour, G. J., Dienes, J. M., Agrin, N. S., King, S. M., Sale, W. S., Kamiya, R.et al. (2006). Radial spoke proteins of Chlamydomonas flagella. J. Cell Sci. 119, 1165-1174. 10.1242/jcs.02811 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang, S., Bahl, K., Chou, H. T., Woodsmith, J., Stelzl, U., Walz, T. and Nachury, M. V. (2020). Near-atomic structures of the BBSome reveal the basis for BBSome activation and binding to GPCR cargoes. Elife 9, e55954. 10.7554/eLife.55954 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ye, F., Nager, A. R. and Nachury, M. V. (2018). BBSome trains remove activated GPCRs from cilia by enabling passage through the transition zone. J. Cell Biol. 217, 1847-1868. 10.1083/jcb.201709041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu, K., Liu, P., Venkatachalam, D., Hopkinson, B. M. and Lechtreck, K. F. (2020). The BBSome restricts entry of tagged carbonic anhydrase 6 into the cis-flagellum of Chlamydomonas reinhardtii. PLoS One 15, e0240887. 10.1371/journal.pone.0240887 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zur Lage, P., Newton, F. G. and Jarman, A. P. (2019). Survey of the ciliary motility machinery of Drosophila sperm and ciliated mechanosensory neurons reveals unexpected cell-type specific variations: a model for motile ciliopathies. Front Genet 10, 24. 10.3389/fgene.2019.00024 [DOI] [PMC free article] [PubMed] [Google Scholar]